
Submitted:
30 July 2024
Posted:
31 July 2024
You are already at the latest version
Abstract
Candida albicans is present in oral health and has been linked with dental caries. So far, the behavior of the yeast in the presence of Streptococcus sanguinis has not been tested. The aim was to determine the effect of excess sugar as a cariogenic environment on the biological and physico-chemical parameters of single-species (C. albicans) and dual-species biofilms of C. albicans and S. sanguinis. An in vitro caries model was used. The biofilms were exposed to 10% sucrose or 0.9% NaCl for 5 min 3x/day/5 days. Viable cells, hyphal quantification, SEM, biofilm pH, H2O2 quantification and surface microhardness of enamel slabs were determined. Microbial viability and the number of hyphae were higher for sucrose exposure, in the dual-species biofilms induced by excess sugar and coexistence between the two microorganisms. Sucrose-exposed biofilms showed a progressive pH drop over the time course. H2O2 production was about 6 times higher for the dual-species biofilms. Enamel demineralization was higher in the single-species, suggesting an inhibitory effect due to S. sanguinis. Excess sugar increases the pathogenic potential of C. albicans biofilm. In dual-species biofilms, S. sanguinis prevents pronounced drops in pH, decreasing demineralization and stimulates the development of hyphae in the yeast.
Keywords:
Excess sugar
; Candida albicans
; Streptococcus sanguinis
; single-species biofilm
; dual-species biofilm
1. Introduction
Abundant communities of hundreds of species comprising complex biofilms can be found in the oral cavity of humans [1,2]. To date, the expanded Human Oral Microbiome database (eHOMD; www.homd.org) informs 774 oral bacterial species, of which only 58% are officially named. Still scarcely understood, species within these biofilms may act as commensal or as competitive microorganisms, thus showing antagonistic or synergistic interactions. Given the complexity of the oral microbiome, control mechanisms should take place among the component microorganisms to preserve an equilibrated condition of eubiosis [3]. Until now, it has been very difficult to study the variety of interactions between the different microorganisms that form part of the dental biofilm in health and disease oral, i.e., dental caries. Species of the genus Streptococcus colonize the oral cavity (about 20%) at an early stage and are the pioneers in the formation of dental biofilm, a favorable oral health condition [3,4]. S. sanguinis is an early colonizer in dental biofilm formation. It is associated with the surface of caries-free teeth and is a native inhabitant microorganism of the mouth [4,5]. This bacterium produces and secretes hydrogen peroxide (H2O2) as an antimicrobial compound to compete with other oral streptococci [5,6]. It has been described that H2O2 production by commensal streptococci (as S. sanguinis) is the principal determinant, bacteriostatic at low concentrations and capable of preventing the growth of bacteria such as S. mutans, a cariogenic microorganism [7], reviewed in [8]. Hence, H2O2 is proposed as a key component for the maintenance of oral ecology associated with healthy conditions [9]. On the other hand, Candida spp. are also common commensal microorganisms in the oral cavity, with approximately 50-60% prevalence in healthy people [10,11]. Under conditions of disturbance of the oral microenvironment or dysbiosis (as dental caries or periodontitis), Candida spp. become opportunistic and pathogenic, condition known as pathobiont, where resident and commensal microorganisms may turn pathogenic and trigger the onset of oral diseases [12]. Even though Candida albicans is the most frequently isolated fungi from the oral cavity, its interaction with commensal microorganisms remains poorly understood. Candida spp. have been linked with dental caries, being isolated with a high frequency (mainly C. albicans) from children with caries lesions compared to caries-free children [13,14]. However, it is still uncertain whether the increase in C. albicans counts involves it with dental caries or wether it is due to factors directly implicated in the beginning of the caries process or the yeast plays an active role in promoting this disease. To date, the biological interaction between S. sanguinis and C. albicans has not been tested, although both are part of the dental biofilm. In this study, we will use S. sanguinis, which is directly correlated with oral health and presents some features of a model microorganism, to study molecular commensalism [5].
On the other hand, the emerging and scarce knowledge on the oral fungal microbiota, including Candida spp., has limited the studies on the interactions between bacteria-fungus and the role they play in health and disease [15,16]. Recently, studies using “omics” technologies have characterized the oral mycobiome in oral health and disease (as dental caries). A marked prevalence in the presence and proportions of Candida spp. yeasts was reported [17,18]. Candida spp. present several virulence traits that could influence caries development, such as adhesion to tooth surfaces, degradation of extracellular matrix and proteins [19], colonization of the oral mucosa and fermentation of carbohydrates [20] contributing to the acidic environment existing in caries. In this context, the hydroxyapatite, the major component of dental enamel formed by a phosphate and calcium apatite complex, is severely affected [21]. Under this acidic environment (pH<5.5), the enamel, the outermost mineral layer of the tooth, begins to demineralize [22]. Moreover, C. albicans changes from yeast-like (ovoid cell) to hypha, a frequently analyzed virulence marker whose morphology prevails when it is part of biofilms [23].
C. albicans co-aggregates with S. sanguinis in in vivo and in vitro conditions [24,25]. On the other hand, in the oral cavity, Candida spp. is constantly exposed to reactive oxidative species, such as H2O2, produced by some bacteria (such as S. sanguinis) and immune cells present in the oral cavity [26]. H2O2 induces genotoxic and oxidative stress in yeast, promoting hyphal formation [27,28]. However, this yeast is protected through several detoxification systems [29].
Current concepts accept that C. albicans is a commensal microorganism under oral health conditions but may transition to a pathobiont when the ecological balance is lost. The specific factors (intrinsic conditions of the oral microenvironment or of the oral microbiota present) that trigger the transition are mostly unknown. This article evaluates the effect of S. sanguinis on C. albicans in dual-species biofilms regarding biological and physico-chemical parameters, in an in vitro model of caries.
The results of this study may contribute important information to further develop novel strategies for caries control, including fungi, along with bacteria, as a potential therapeutic target.
2. Materials and Methods
Microorganisms and growth conditions: Oral clinical isolate of Candida albicans from caries-free preschool children (ICDAS II code = 0), (part of the strain collection of our laboratory) and S. sanguinis SK36 (from human dental plaque) were used in this study. The Todd Hewitt Broth medium (THB; BD, MD, USA) was used for individual growth of bacteria pre-cultures with 5% CO2 for 18 h at 37ºC. Sabouraud-dextrose (SD; BD, MD, USA) broth was used in C. albicans aerobic pre-cultures overnight at 37ºC. Yeast Nitrogen Base (YNB; BD, MD, USA) broth supplemented with 0.1 mM glucose or 10% sucrose according to experimental design was used in single- and dual-species biofilms. The study protocol and informed consent (because of the use of human saliva for biofilm formation) were approved by the Ethics Committee of the Faculty of Dentistry, University of Chile (certificate Nº5/2022).
Single-species biofilm (C. albicans). This model was performed according to [30] with some modifications. That is, the biofilms were formed on saliva-coated enamel slabs (prepared from bovine incisors as described in [31]) as substrates to grow biofilms. Human non-stimulated saliva was obtained and filtered using filters with a pore size of 0.22 μM (Corning, NY, USA), obtained from healthy volunteers fasting for 10 h mixed with 1X AB buffer (50 mM KCl, 1 mM KPO4 [0.35 mM K2HPO4 and 0.65 mM KH2PO4], 1 mM CaCl2·2H2O, 0.1 mM MgCl2·6H2O, pH 6.5) and 0.1 M PMSF protease inhibitor (Sigma-Aldrich, MD, USA) was used to form the acquired salivary pellicle on the enamel slabs (sES). The sES were suspended vertically in a 24-well culture plate (Costar®, Corning, NY, USA) by orthodontic wire. sES were inoculated with C. albicans cultures (OD600nm of 0.38, equivalent to 107 CFU/mL) in YNB broth supplemented with 10% sucrose solution and incubated for 1.5 h at 37ºC with gentle agitation (to allow the adherence phase of biofilms). Then, 10% fetal bovine serum (FBS; Gibco, NY, USA) (for hyphal formation) was added and incubated for 10 h at 37ºC in microaerophilic conditions. The biofilm was allowed to mature in YNB broth supplemented with 0.1 mM glucose for 18 h in the same conditions as above, with a fresh medium change at 12 h. Subsequently, the experimental design consisted of the biofilms being exposed 5 min, 3 times/day for 5 days to 10% sucrose solution (excess sugar) or 0.9% NaCl (control condition). The treatment was carried out in 6 biological replicates per condition from two independent experiments (n=12).
C. albicans and S. sanguinis dual-species biofilm. This model is similar to the single-species biofilms with modifications [32]. After the C. albicans biofilms attachment phase, 10% FBS, S. sanguinis culture (OD600nm of 0.5, equivalent to 107 CFU/mL), THB medium and YNB supplemented with 10% sucrose solution were added and incubated for 10 h at 37°C under microaerophilic conditions. The biofilms were allowed to mature in YNB broth supplemented with 0.1 mM glucose and THB medium for 18 h under the same conditions as above, with a fresh medium change at 12 h. Subsequently, the biofilms were exposed to the same experimental treatment as the single-species biofilms, in 6 biological replicates in 2 independent experiments (n=12).
Biological and physico-chemical parameters (described below) were measured from single- and dual-species biofilms. Additionally, observations of the biofilms were made using Scanning Electron Microscopy (SEM) under the conditions previously indicated.
Biological parameters: A) Viable cells: After 5 days of experimental treatment of the single- and dual-species biofilms, enamel slabs were removed from the wire and transferred into sterile tubes with 1 mL 1X PBS pH 7.4. The biofilms were dispersed with a sonicator with 3 pulses of 30 s at 7 W with 1 min interval on ice. This “master solution” was serially diluted in PBS and seeded in duplicate on THB agar and SD agar (supplemented with chloramphenicol at a final concentration of 20 μg/mL) to obtain CFU/mL of bacteria and yeast, respectively. The plates were incubated aerobically at 37ºC for 48 h for the development of yeast colonies, while quantification of bacterial colonies was performed by incubating the plates in 5% CO2 at 37ºC for 24 h. Colony quantification was done in 6 biological replicates per condition from 2 independent experiments (n=12). B) Hyphal quantification: In single- and dual-species biofilms, this quantification was performed as previously described by [25]. Briefly, 50 μL of master solution of each biofilm were spread on a glass slide and viewed under a light microscope (Standard 20, Carl Zeiss, Jena, Germany) at 40x magnification. Ten microscopic fields per slide were analyzed in duplicate in the 3 biological replicates per condition.
Physico-chemical parameters: A) pH measure in biofilms: pH was determined according to [32] introducing a microelectrode (35811-98 OAKTON, IL, USA) coupled to a pH-meter (ION 700 OAKTON, IL, USA) into each well. Readings were taken twice per day in the spent medium, after overnight incubation and before the last fresh medium change of the day, and during the entire course of the experiment (approx. 142 h). B) Surface microhardness (enamel demineralization): It was estimated through the loss of Knoop’s surface hardness (SH) in enamel slabs from single- and dual-species biofilms, described in [31]. Briefly, initial surface hardness (SH)i was evaluated by 3 linear indentations in a Knoop microindenter with a microhardness tester (402 MVD, Wolpert Wilson Instruments, USA), at 50 g/5 s. After the experimental treatment, 3 indentations were made to the slabs, adjacent to the initial indentations, considered as final SH (SH)f (kg/mm2). The average (SH)i and (SH)f were used to calculate the percentage loss of SH (%SHL), interpreted as demineralization: (average (SH)i − average (SH)f) × 100/(SH)i. C) H2O2 production: At the end of the assays, the filtered supernatant of the culture medium of each well in single- and dual-species biofilms and the AmplexTM Red kit were used (ThermoFisher Scientific, MA, USA). Briefly, in the presence of peroxidase, Amplex™ Red reagent reacts with H2O2 (released from biological samples) in a 1:1 stoichiometry to create a red fluorescent oxidation product, resorufin. Subsequently, H2O2 concentration in the biofilm supernatant was calculated according to the supplier’s instructions. The quantification was done in 3 biological replicates per condition from 2 independent experiments.
Observation of single- and dual-species biofilms by microscopy: After experimental treatment, single-species (C. albicans) biofilms on circular glass discs (1 cm diameter) and dual-species biofilms on the enamel slabs were examined by SEM (JEOL JSM IT300LV; Tokyo, Japan) operated at 20 kV and a 10-mm working distance. Glass discs and enamel slabs with the biofilms were fixed with 2.5% glutaraldehyde and then treated with alcohol in ascending concentration from 50% to 100% which dehydrated the samples. Subsequently, they were subjected to critical point drying and metallized with carbon. The images were the results of general observations of the samples.
Statistical analysis. Data were analyzed using STATA SE software v14. All data were expressed as mean and SD respective or median with ranges, as appropriate. T-test, ANOVA and Tukey’s post hoc test were used as well as Wilcoxon test for non-parametric samples. Differences were considered significant if p<0.05.
3. Results
3.1. Effect of Sucrose on Biological Parameters in Single- and Dual-Species Biofilms
3.1.1. Cell Viability of C. albicans and S. sanguinis
When sucrose exposed and unexposed biofilms were assessed, non-significant differences were detected on yeast viability of single-species biofilms with C. albicans, with a trend for higher viability under sucrose condition compared to the control (NaCl) (p>0.05). Median values were 1.2x106 CFU/mL (range 2.4x105–5.3x106) and 8.8x105 CFU/mL (range 3.6x105–2.6x106), respectively (Table 1). For the dual-species biofilms, a significant median difference (p<0.001) between the control condition (1.1x106 CFU/mL, range: 3.0x105–4.5x106) and the sucrose condition (1.4x106 CFU/mL, range: 2.7x105–3.9x106) was observed. The median viability of S. sanguinis in the control condition was 8.0x107 (range: 4.0x106–4.1x108), and in the sucrose condition 1.0x108 (range: 7.0x106–3.7x108) (p<0.001) (Table 1).
3.1.2. Filamentation of C. albicans
In the single-species C. albicans biofilms, the control condition showed slightly higher hyphal counts than the sucrose condition, but the differences were not significant (p>0.05), with a mean of 5.31 ± 2.56 and 4.42 ± 1.47, respectively (Figure 1). In contrast, the number of hyphae in the dual-species biofilms for both control condition (7.6 ± 2.3) and the sucrose condition (14.9 ± 4.6) showed a significant difference (p<0.001). The highest hyphal count was observed in the dual-species biofilms for both experimental conditions (p<0.001).
3.1.3. Biofilms Formation
Biofilm formation for both experimental conditions is shown in Figure 2. In the NaCl condition (Figure 2A) of single-species biofilms (C. albicans), hyphal structures and their blastospores, as well as some groups of cells forming small aggregates, were observed. Figure 2B depicts sucrose condition with more hyphal and blastospore structures and aggregates are observed, forming a typical biofilm structure.
In the dual-species biofilms, a greater number of oval forms and yeast hyphae were observed in the sucrose condition (Figure 2D), compared to the control condition (Figure 2C). In addition, proximity (physical interaction) between C. albicans and S. sanguinis was observed in both conditions.
Noticeably, for both conditions the biofilm extracellular matrix cannot be observed due to the treatment needed for microscopic observation.
3.2. Effect of Sucrose on Physico-Chemical Parameters in Single- and Dual-Species Biofilms
3.2.1. Acidogenicity Measurement in the Biofilms
Figure 3a shows the mean pH values (representing acidogenicity from the biofilms) of the C. albicans single-species biofilms for both experimental conditions. Biofilms exposed to sucrose showed higher acidogenicity compared to biofilms exposed to NaCl. The mean pH value was similar in both conditions until approximately 22 h after the beginning of the experiment (p>0.05), and from 30 h to the end of the experiment, a significant decrease in pH was observed in the sucrose condition versus the NaCl condition (p<0.05). The highest pH for sucrose-exposed biofilms was 4.18 ± 0.42 at 102 h, while the lowest pH for the control was 5.25 ± 0.33 at 126 h. Figure 3b shows the variation in pH for the dual-species biofilms during the entire course of the experiment (142 h). From 46 h to 142 h, there were significant differences in the pH values between both experimental conditions (p<0.001). The lowest pH obtained in the control condition was 6.29 ± 0.14, while the lowest pH obtained in the sucrose condition was 4.98 ± 0.20, both at 126 h.
3.2.2. Surface Microhardness on Enamel Slabs
Figure 4 shows surface microhardness loss data, which represents enamel demineralization. Enamel slabs exposed to sucrose showed 71.6% ± 9.71, which is slightly higher than the NaCl condition, with 65.96% ± 16.69 (p>0.05), in the single-species biofilms. For the dual-species biofilms, sucrose condition showed higher numbers than NaCl, 64.94% ± 13.24 and 53.95% ± 13.60, respectively (p<0.05). The lowest demineralization percentages were in the dual-species biofilms for both experimental conditions.
3.2.3. H2O2 Production in Biofilms
Table 2 shows H2O2 quantification for both types of biofilms under the described experimental conditions. For dual-species biofilms, H2O2 levels were 3.05 ± 0.99 μM for sucrose and 2.51 ± 0.77 μM for NaCl (p>0.05), while in the single-species biofilms they were lower than 0.5 μM. For both experimental conditions, H2O2 levels were higher for dual-species than single-species biofilms (p<0.05).
4. Discussion
Effect of Sucrose on Biological Parameters Evaluated in Single- and Dual-Species Biofilms
Cell Viability of C. albicans and S. sanguinis
This study confirmed the ability of C. albicans to metabolize sucrose, which favors increased cell viability of dual biofilms. Bezerra et al. [33] observed an increased production of soluble and insoluble extracellular polysaccharides (EPS) in yeast monospecies biofilms subjected to high concentrations of glucose, favoring biofilm formation and increased cell viability. High Candida yeast counts have been reported in microenvironments with high sugar concentration and acidic salivary pH. For example, studies have been carried out with oral clinical isolates of C. albicans obtained from saliva of patients with type II diabetes mellitus. It was determined that when these isolates were exposed to different concentrations of glucose, they had greater cell viability than those from saliva of non-diabetic subjects [34,35]. High counts of this yeast genus have also been observed in similar microenvironments, such as in the saliva of preschool children with severe caries lesions or ICDAS II code 6 [13]. In the review by Lok et al. [36], it was described that C. albicans has the ability to double its viable count only in the presence of sugars such as glucose, fructose and galactose.
In the same article, it was mentioned that the effect of different carbon sources and the biofilm formation capacity of this yeast were tested, and it was determined that under exposure to sucrose the EPS content, roughness, height and thickness of its biofilm were higher compared to glucose, arabinose and lactate. Interestingly, the article by Xiang et al. [37] determined that clinical oral isolates of C. albicans from dental plaque of toddlers with severe childhood caries showed high proteinase activity and acidogenic and aciduric properties. In addition, it was observed that these isolates showed variations in hyphal growth, i.e., some of them had defective hyphal formation and others were hyperfilamentous. Finally, it was observed that mixed biofilms of these yeast isolates with S. mutans increased biomass and EPS accumulation versus using reference yeast strains.
In the study by Ev et al. [38], using RNA-seq analysis, it was established that C. albicans isolates from active root caries lesions showed overexpression of genes related to metabolic activity, sugar transport, stress tolerance, invasion and pH regulation, compared to isolates from healthy root surfaces, where only genes related to biofilm formation were overexpressed.
Huffines and Scoffield [8] observed that dual-species biofilms of C. albicans with S. parasanguinis (one of the most abundant commensals) were less robust than those of yeast with S. mutans. In addition, S. parasanguinis reduced the synergy between them in the biofilm. In the same study, no significant differences were found in the biomass of S. parasanguinis in single-species versus dual (with C. albicans) and three-species (with C. albicans + S. mutans) biofilms. In our study, the increase of viable S. sanguinis cells in the dual-species biofilm with excess sugar could be explained by the presence of yeast, for its role as a scaffold shaping the structure of polymicrobial biofilms [39], and, by the intrinsic ability of the bacterium to metabolize sugar for cell proliferation [40].
Taken together, both yeast and bacterial viability are promoted induced by excess sugar and when they co-exist in dual-species biofilm.
Hyphal Formation in C. albicans
C. albicans is characterized by the fact that it typically grows as a yeast (oval shaped and usually in budding) but can transition to a pseudofilamentous and filamentous form (hyphae) under the influence of environmental cues, such as alterations in temperature, pH and available nutrients. This way of changing from a yeast-like to a hyphal state and vice versa is considered the most pathogenic form of yeast, facilitating the formation of biofilms [41]. It has been determined that in the presence of sugars such as fructose and glucose, the 3 types of cell morphology of this yeast coexist with other sugars (reviewed in [36]).
In their study, do Rosario Palma et al. [25], observed the absence of inhibition of C. albicans filamentation. Furthermore, there was transcriptional overexpression (5-fold) of the gene encoding the hyphal wall protein (HWP1), involved in yeast adhesion and biofilm formation in the presence of S. sanguinis versus monospecies biofilms. Similarly, reviewed in Lohse et al. [42] and [28], pointed to a synergistic interaction between yeast and oral streptococci, such as S. sanguinis, promoting bacterial colonization and biofilm formation and, on the other hand, favoring yeast virulence and tissue invasion.
In the study by Morse et al. [43], they observed that hyphal formation increased when C. albicans was cultured together with S. sanguinis, S. gordonii, Actinomyces viscosus and A. odontolyticus. And when adding Porphyromonas gingivalis to this microbial consortium, this filamentation capacity was reduced. With this background, we can partly outline that one of the roles of C. albicans within these microbial societies is to have a structural function that allows bacterial co-aggregation and biofilm proliferation.
It is worth noting that yeast filamentation was only observed in the dual-species biofilm, where the only modified variable was the presence of S. sanguinis, so we can speculate that the presence of the bacterium alone could be inducing hyphal formation. The study by Xu et al. [44] demonstrated that the commensal bacterium S. oralis activates the filamentation pathway in C. albicans, promoting the interaction between kingdoms (prokaryote—eukaryote) and thus the formation of biofilms. This shows that the morphogenetic trait of the yeast is promoted by the presence of a bacterium alone.
It has been described that an alkaline pH (pH above 6) induces filamentation in yeast (reviewed in [45]). However, a study by Merino et al. [46] described that this morphological transition also occurs in a pH range from 4.4 to 7.4, which would be promoting the formation of biofilms. Thus, the ability to form hyphae could be a unique trait of the yeast that influences the virulence of biofilms [38].
In vitro studies have reported that H2O2 (commercial) induces the morphological transition of C. albicans from the yeast form to the hyphal stage [29,47]. Nevertheless, the effect of H2O2 biologically produced by S. sanguinis on filamentation is unknown. Preliminary studies in our laboratory showed an increase in filamentation of this yeast when exposed to supernatant (filtrate) from S. sanguinis cultures in late exponential phase, compared to commercial H2O2 and fetal bovine serum (unpublished data). Together with the few existing studies of interactions between C. albicans and S. sanguinis, the results of this study and unpublished data speculate that the bacteria modulate the pathogenic traits of the yeast.
Effect of Sucrose on Physico-Chemical Parameters Evaluated in Single- and Dual-Species Biofilms
Acidogenicity of the Biofilms
In dual-species biofilms of C. albicans with S. mutans, Willems et al. [48] found that the pH of the medium was higher than the pH of the single-species biofilm of the bacteria, so that C. albicans would be raising the pH in acidogenic microenvironments, as confirmed by Eidt et al. [49]. Despite the above, it should be noted that the articles mentioned are not comparable, because the C. albicans strains, the culture medium and the substrate used to form the biofilms are different. However, in our experiments pH values in the C. albicans biofilm and in excess sugar were acidic (close to 4.2), whereas for dual-species biofilm these values reached minimum of 5.0, indicating the inhibitory effect of S. sanguinis on pH in slightly acidic environments. That is, the bacterium induces the arginine deiminase system (ADS) [50], which generates ammonium from arginine as a substrate present in the proteins of the medium, raising the pH. C. albicans cannot produce lactic acid because it lacks the lactate dehydrogenase enzyme. However, it produces other acids, such as malic, pyruvic and acetic acid, which may contribute to the predominant acidic pool, such as a cariogenic condition [20]. This was also confirmed in the study by Bezerra et al. [33], where the pH drop showed a dose-dependent effect with respect to the glucose concentration in the culture medium in monospecies C. albicans biofilms. Conversely, monospecies S. sanguinis biofilms under similar experimental conditions reached pH values close to 6.0 under excess sugar [51]. Therefore, it is possible to conclude that the presence of S. sanguinis in conditions of excess sugar hampers pH drops in dual-species biofilms.
Enamel Slabs Demineralization
One of the pathogenic traits of Candida spp. is the production and secretion of extracellular enzymes, such as aspartyl proteases, phospholipases and lipases that are activated at low pH and degrade proteins and extracellular matrix components [19]. It has also been described that Candida spp. are able to dissolve dentin and consequently release calcium into the medium [52], but this has not been tested on tooth enamel, which has a higher mineral content than dentin. de Abreu et al. [20] observed that C. albicans biofilms formed on bovine enamel slabs coated with a pool of human saliva from preschool children with and without caries showed a higher percentage of demineralization compared to the control, which corresponded to enamel slabs coated with saliva but without the addition of yeast. Also, in the study by Charone et al. [53] using oral clinical isolates of Candida spp. from supragingival plaque of HIV-positive children, observed a gradual reduction in the surface microhardness of human tooth enamel from day 1 to day 5 after biofilm formation; it was also observed that C. albicans produced calcium release during the experiment. Our study, yeast monospecies biofilms yielded slightly more enamel demineralization, although not significant, in the excess sugar condition than in the control. However, the demineralization percentages were lower in the dual biofilm. This would mean that in excess sugar the presence of S. sanguinis inhibits the enamel demineralization preventing a pronounced decrease in pH values.
Furthermore, in the study by Willems et al. [48], it is shown that biofilms of yeast and S. mutans in a glucose medium supplemented with 0.2% sucrose had a more alkaline pH and consequently less calcium release compared to the single-species biofilm of the bacterium. Under these experimental conditions, we can speculate that C. albicans has also a role in decreasing cariogenicity in sugar-rich microenvironments and in the presence of an acidogenic bacterium. However, that study did not use enamel slabs for biofilm growth, but hydroxyapatite discs. With the results of our study, we can propose that excess sugar has a greater impact on enamel demineralization in yeast monospecies biofilms than in dual-species biofilms in the presence of the commensal bacterium S. sanguinis.
H2O2 Production in Biofilms
S. sanguinis produced H2O2 in the dual-species biofilm under the conditions of the study and tended to be slightly higher in excess sugar (around 3 μM). In the article by Díaz-Garrido et al. [51], H2O2 production was determined to be around 15 μM and 25 μM in a dual-species biofilm of S. sanguinis with S. mutans and in a single-species S. sanguinis biofilm, respectively, using the same in vitro caries model. Competition between these two bacteria has been widely described in the literature [7,32,54]. Thus, this difference in H2O2 production levels compared to our results represents that S. sanguinis does not need to compete for microenvironment with yeast or it is sufficient to maintain the attenuated virulent traits of C. albicans. As for the chemical or metabolic interaction of S. sanguinis with C. albicans, evidence is scarce. Nevertheless, it is worth mentioning the work by Zheng et al. [55], who found that the carbon catabolite control protein A (CcpA) of S. sanguinis regulates the expression of the spxB gene, encoding the pyruvate oxidase enzyme (involved in H2O2 production). However, this regulation is not influenced by glucose concentration. This relationship may explain why we did not observe significant differences in H2O2 production in the control versus sucrose condition.
On the other hand, oral streptococci (S. oralis, S. mitis, S. sanguinis and S. gordonii) have been reported to release H2O2 into the surrounding environment, which can be inhibitory or toxic to fungal cells, limiting the proliferation of, for example, Candida spp. in the host [47]. This fungus has different mechanisms to neutralize ROS and cope with oxidative stress, through the induction of the catalase enzyme gene (CAT1) that degrades H2O2; the synthesis of antioxidants such as glutathione can also repair oxidative damage to DNA, proteins and lipids (Reviewed in [56]). Additionally, C. albicans synthesizes farnesol, a molecule derived from farnesoic acid, which acts in cell-cell communication (quorum sensing), attenuates oxidative stress, and plays a key role in inhibiting bud-to-hyphal transition at high cell densities, without preventing elongation of pre-existing hyphae or affecting population growth [57,58]. Therefore, it can be suggested that H2O2 has an inhibitory effect on the viability, filamentation and ability of C. albicans to form biofilms with commensal microorganisms, which was not observed in our results. In addition, studies indicate that a neutral or alkaline pH of the medium promotes the formation of filamentous forms of C. albicans, while a low pH (present in medium with excess sugar) should inhibit the formation of filamentous forms of C. albicans (reviewed in [41]).
Consequently, we can state that excess sugar in a dual-species biofilm of C. albicans and S. sanguinis has no effect on H2O2 production. It remains to be studied whether this chemical produces oxidative stress in yeast.
Candida albicans Role in the Oral Biofilms Structure
The effect of a high-sugar diet on multispecies biofilms has been described in models involving S. mutans and C. albicans. Bowen et al. [59] indicated that this relationship between kingdoms is due to EPS formation, coadhesion and the role of exoenzymes, such as bacterial Gtfs, in the production of an extracellular matrix (ECM) that allows synergistic interaction between these species. Once these biofilms are formed, both species can exchange metabolites, substrates and growth factors, while continuing to produce exoenzymes and increasing the ECM volume of the biofilm. Thus, it can be deduced that both microorganisms play a structural role within the biofilm. Under oral health conditions, C. albicans may interact synergistically with S. sanguinis (physically, chemically or metabolically), whereby hyphal formation serves as a surface for the adhesion of these bacteria and leads to the establishment of biofilms [39]. In our results, it was observed that the number of yeast cells and hyphae increased considerably under a microenvironment with excess sugar and in the dual-species biofilm. Thus, the only difference with the yeast monospecies biofilm regarding these parameters was the presence of S. sanguinis. Unfortunately, SEM images do not show the volume and biomass of the biofilms. Still, in the case of the dual-species biofilm, they show the physical proximity between the two microorganisms. Therefore, using inverted microscopy or CLSM would give us more information about the biovolumes of the biofilms.
While synergism between S. mutans and C. albicans has been described, there is little evidence regarding oral commensal streptococci and their interaction with yeast. An example of this is the article by Huffines and Scoffield [8], who observed that in multispecies biofilms, S. parasanguinis inhibits C. albicans adherence to glucans and glucan formation by S. mutans by blocking the activity of its Gtfs enzymes, indicating the potential ability of S. parasanguinis to attenuate the synergy between these microorganisms and prevent oral dysbiosis. On the other hand, it has been reported that S. gordonii cannot co-aggregate with hyphal-defective C. albicans isolates. However, the yeast develops robust biofilms with S. mutans, independent of the filamentation status [37].
Our results suggest that S. sanguinis modulates the virulence of C. albicans by affecting hyphal formation and demineralizing capacity. Further studies are needed to understand these interactions and to develop new therapeutic strategies. It is also recommended to investigate how oral microenvironmental factors influence these interactions.
The study and understanding of the biological interactions occurring in oral biofilms should not only be based on bacterial targets. Further research is needed to elucidate how different microorganisms interact and organize in dental biofilm during health or in caries.
5. Conclusions
Excess sugar increases the pathogenic potential of mono-species C. albicans biofilms, in cell viability, filamentation and demineralizing capacity. Under conditions of excess sugar, the presence of S. sanguinis in dual-species biofilms with an oral clinical isolate of C. albicans attenuates abrupt pH drops, decreasing demineralization induced by the yeast. In addition, under this condition, there is increased viability of the bacteria, without affecting H2O2 production.
Further studies should deepen these findings, exploring the biochemical and molecular basis of the interaction between C. albicans and other commensal oral bacteria, such as S. sanguinis. The results suggest potential new strategies for preventing dental caries by harnessing the natural inhibitory effects of S. sanguinis on pathogenic yeasts and bacteria. This could lead to the development of probiotic treatments or other microbiome-based therapies.
Author Contributions
CPL designed and performed all the experiments, processed, and analyzed the data and drafted the first manuscript. EMC performed dual-species biofilms, processed, and analyzed the data. NGM carried out surface microhardness measurements of the enamel slabs. RAG and CPL wrote the final manuscript. CL and XL critically revised and contributed with new ideas to the paper. All authors revised and approved the final version of the article.
Funding
Funding of the study was contributed by the Faculty of Dentistry, University of Chile Grant FIOUCH 2022/02 to CPL.
Data Availability Statement
All data generated or analyzed during this study are included in this article. Further enquiries for specific data can be directed at the corresponding author.
Acknowledgments
Thanks to Mr. Juan Fernández, from the language and Translation Services of the Faculty of Dentistry, University of Chile, for kindly proofreading and checking the spelling and grammar of this manuscript.
Conflicts of Interest
There are no conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
- Takahashi, N.; Nyvad, B. The role of bacteria in the caries process: ecological perspectives. J. Dent. Res. 2011, 90, 294–303. [Google Scholar] [CrossRef]
- Simón-Soro, A.; Ren, Z.; Krom, B.P.; Hoogenkamp, M.A.; Cabello-Yeves, P.J.; Daniel, S.G.; Bittinger, K.; Tomas, I.; Koo, H.; Mira, A. Polymicrobial aggregates in human saliva build the oral biofilm. mBio. 2022, 13, e0013122. [Google Scholar] [CrossRef]
- Marsh, P.D.; Zaura, E. Dental biofilm: ecological interactions in health and disease. J. Clin. Periodontol. 2017, 44, S12–S22. [Google Scholar] [CrossRef]
- Marsh, P.; Martin, M. Williams, D. In Oral Microbiology, 5th ed.; Lewis, MA., Ed.; Edinburgh London New York Oxford Philadelphia St Louis Sydney Toronto, Churchill Livingstone Elsevier, 2009.
- Kreth, J.; Giacaman, R.A.; Raghavan, R.; Merritt, J. The road less traveled - defining molecular commensalism with Streptococcus sanguinis. Mol. Oral Microbiol. 2017, 32, 181–196. [Google Scholar] [CrossRef]
- Zhu, L.; Kreth, J. The role of hydrogen peroxide in environmental adaptation of oral microbial communities. Oxid. Med. Cell. Longev. 2012, 2012, 717843. [Google Scholar] [CrossRef] [PubMed]
- Kreth, J.; Merrit, J.; Shi, W.; Qi, F. Competition and coexistence between Streptococcus mutans and Streptococcus sanguinis in the dental biofilm. J. Bacteriol. 2005, 187, 7193–7203. [Google Scholar] [CrossRef]
- Huffines, J.T.; Scoffield, J.A. Disruption of Streptococcus mutans and Candida albicans synergy by a commensal streptococcus. Sci. Rep. 2020, 10, 19661. [Google Scholar] [CrossRef] [PubMed]
- Redanz, S.; Cheng, X.; Giacaman, R.A.; Pfeifer, C.S.; Merritt, J.; Kreth, J. Live and let die: hydrogen peroxide production by the commensal flora and its role in maintaining a symbiotic microbiome. Mol. Oral Microbiol. 2018, 33, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Thein, Z.M.; Samaranayake, Y.H.; Samaranayake, L.P. Effect of oral bacteria on growth and survival of Candida albicans biofilms. Arch. Oral Biol. 2006, 51, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Romo, J.A.; Kumamoto, C.A. On commensalism of Candida. J Fungi 2020, 6, 16. [Google Scholar] [CrossRef]
- Simón-Soro, A.; Guillen-Navarro, M.; Mira, A. Metatranscriptomics reveals overall active bacterial composition in caries lesions. J. Oral Microbiol. 2014, 6, 25443. [Google Scholar] [CrossRef]
- Lozano, C.; Rodríguez, G.; Lefimil, C.; Morales-Bozo, I.; Urzúa-Orellana, B. Prevalence of Candida albicans and carriage of Candida non-albicans in the saliva of preschool children, according to their caries status. Acta Odontol. Scand. 2017, 75, 30–35. [Google Scholar] [CrossRef]
- Du, Q.; Ren, B.; He, J.; Peng, X.; Guo, Q.; Zheng, L.; Li, J.; Dai, H.; Chen, V.; Zhang, L. ; Zhou, X; Xu, X. Candida albicans promotes tooth decay by inducing oral microbial dysbiosis. ISME J. 2021, 15, 894–908. [Google Scholar] [CrossRef]
- Díaz, P.I.; Strausbaugh, L.D.; Dongari-Bagtzoglou, A. Fungal-bacterial interactions and their relevance to oral health: linking the clinic and the bench. Front. Cell. Infect. Microbiol. 2014, 4, 101–106. [Google Scholar] [CrossRef]
- Janus, M.M.; Willems, H.M.E.; Krom, B.P. Candida albicans in multispecies oral communities; A keystone commensal? Adv. Exp. Med. Biol. 2016, 931, 13–20. [Google Scholar] [CrossRef]
- Baraniya, D.; Chen, T.; Nahar, A.; Alakwaa, F.; Hill, J.; Tellez, M.; Ismail, A.; Puri, S.; Al-Hebshi, N.N. Supragingival mycobiome and inter-kingdom interactions in dental caries. J. Oral Microbiol. 2020, 12, 1729305–1729313. [Google Scholar] [CrossRef]
- Hong, B.Y.; Hoare, A.; Cardenas, A.; Dupuy, A.K.; Choquette, L.; Salner, A.L.; Schauer, P.K.; Hegde, U.; Petersen, D.E.; Dongari-Bagtzoglou, A.; Strausbauhg, L.D.; Díaz, P.I. The salivary mycobiome contains 2 ecologically distinct mycotypes. J. Dent. Res. 2020, 99, 730–738. [Google Scholar] [CrossRef]
- Li, W.; Yu, D.; Gao, S.; Lin, J.; Chen, Z.; Zhao, W. Role of Candida albicans-secreted aspartyl proteinases (Saps) in severe early childhood caries. Int. J. Mol. Sci. 2014, 15, 10766–10779. [Google Scholar] [CrossRef]
- de Abreu, T.C.; Barbosa, M.; Moraes, P.; Barbosa, G.F.; Cople, L.; Fonseca-Goncalves, A. Demineralizing potential of dental biofilm added with Candida albicans and Candida parapsilosis isolated from preschool children with and without caries. Microb. Pathog. 2016, 100, 51–55. [Google Scholar] [CrossRef]
- Lamont, R.J.; Jenkinson, H.F. Microbiology of Caries. In Oral Microbiology at a Glance, 1st ed.; Wiley-Blackwell publication, United Kingdom, 2010; Volume 1, pp. 34.
- Akküç, S.; Duruk, G.; Keles, A. Remineralization effect of three different agents on initial caries and erosive lesions: a micro-computed tomography and scanning electron microscopy analysis. BMC Oral Health 2023, 23, 106–117. [Google Scholar] [CrossRef]
- Soon-Hwan, O.; Martin-Yken, H.; Coleman, D.A.; Dague, E.; Hoyer, L.L. Development and use of a monoclonal antibody specific for the Candida albicans cell-surface protein Hwp1. Front. Cell. Infect. Microbiol. 2022, 12, 907453. [Google Scholar] [CrossRef]
- Ten Cate, J.M.; Klis, F.M.; Pereira-Cenci, T.; Crielaard, W.; de Groot, P.W.J. Molecular and cellular mechanisms that lead to Candida biofilm formation. J. Dent. Res. 2009, 88, 105–115. [Google Scholar] [CrossRef]
- do Rosário Palma, A.; Domingues, N.; de Barros, P.; Brito, G.; Cardoso, A. Influence of Streptococcus mitis and Streptococcus sanguinis on virulence of Candida albicans: in vitro and in vivo studies. Folia Microbiol. 2019, 64, 215–222. [Google Scholar] [CrossRef]
- Brown, G. Innate antifungal immunity: the key role of phagocytes. Ann. Rev. Immunol. 2011, 29, 1–21. [Google Scholar] [CrossRef]
- Nasution, O.; Srinivasa, K.; Kim, M.; Kim, Y.J.; Kim, W.; Jeong, W.; Choi, W. Hydrogen peroxide induces hyphal differentiation in Candida albicans. Eukaryot. Cell. 2008, 7, 2008–2011. [Google Scholar] [CrossRef]
- Koo, H.; Andes, D.R.; Krysan, D.J. Candida-streptococcal interactions in biofilm-associated oral diseases. PLoS Pathog. 2018, 14, e1007342. [Google Scholar] [CrossRef]
- Komalapriya, C.; Kaloriti, D.; Tillmann, A.T.; Yin, Z.; Herrero-de-Dios, C.; Jacobsen, M.D.; Belmonte, R.C.; Cameron, G.; Haynes, K.; Grebogi, C.; de Moura, A.P.; Gow, N.A.; Thiel, M.; Quinn, J.; Brown, A.J.; Romano, M.C. Integrative Model of Oxidative Stress Adaptation in the Fungal Pathogen Candida albicans. PloS One 2015, 10, e0137750. [Google Scholar] [CrossRef]
- Díaz-Garrido, N.; Lozano, C.; Giacaman, R. Frequency of sucrose exposure on the cariogenicity of a biofilm-caries model. Eur. J. Dent. 2016, 10, 345–350. [Google Scholar] [CrossRef]
- Giacaman, R.A.; Umaña, R.; Nuñez, M.J.; Díaz-Garrido, N.; Echeverría, C.; García-Manríquez, N.; Mira, A.; Fernández, C.E.; Gambetta-Tessini, K.; Lozano, C.P. Saliva decreases sucrose-induced cariogenicity in an experimental biological caries model. Microorganisms 2023, 11, 1426. [Google Scholar] [CrossRef]
- Lozano, C.; Díaz-Garrido, N.; Kreth, J.; Giacaman, R. Streptococcus mutans and Streptococcus sanguinis Expression of Competition-Related Genes, Under Sucrose. Caries Res. 2019, 53, 194–203. [Google Scholar] [CrossRef]
- Bezerra, N.V.F.; Brito, A.C.M.; de Medeiros, M.M.D.; de França Leite, K.L.; Bezerra, I.M.; de Almeida, L.F.D.; Aires, C.P.; Cavalcanti, Y.W. Glucose supplementation effect on the acidogenicity, viability, and extracellular matrix of Candida single- and dual-species biofilms. J. Investig. Clin. Dent. 2019, 10, e12412. [Google Scholar] [CrossRef]
- Aitken-Saavedra, J.; Lund, R.G.; González, J.; Huenchunao, R.; Perez-Vallespir, I.; Morales-Bozo, I.; Urzúa, B.; Tarquinio, S.C.; Maturana-Ramírez, A.; Martos, J.; Fernandez-Ramires, R.; Molina-Berríos, A. Diversity, frequency and antifungal resistance of Candida species in patients with type 2 diabetes mellitus. Acta Odontol. Scand. 2018, 76, 580–586. [Google Scholar] [CrossRef]
- Domingues, P.C.A.; Oliveira, V.C.; Bim, F.L.; Aires, C.P.; Santos, A.P.D.; Castro, D.T.; Silva-Lovato, C.H.; Andrade, D.; Watanabe, E. Influence of glucose supplementation on biofilm formation of Candida albicans and Candida glabrata isolated from diabetic and non-diabetic individuals. Arch. Oral Biol. 2022, 134, 105339. [Google Scholar] [CrossRef]
- Lok, B.; Ahmad, M.A.; Mohd, L.Z.; Chukwudi, N.A.; Sandai, R.; Sandai, D. The assimilation of different carbon sources in Candida albicans: Fitness and pathogenicity. Med. Mycol. 2021, 59, 115–125. [Google Scholar] [CrossRef]
- Xiang, Z.; Wakade, R.S.; Ribeiro, A.A.; Hu, W.; Bittinger, K.; Simon-Soro, A.; Kim, D.; Li, J.; Krysan, D. J.; Liu, Y.; Koo, H. Human Tooth as a Fungal Niche: Candida albicans Traits in Dental Plaque Isolates. mBio. 2023, 14, e0276922. [Google Scholar] [CrossRef]
- Ev, L.D.; Damé-Teixeira, N.; DO, T.; Maltz, M.; Cavalcanti, C. The role of Candida albicans in root caries biofilms: an RNA-seq analysis. J. Appl. Oral Sci. 2020, 28, e20190578. [Google Scholar] [CrossRef]
- Ren, Z.; Jeckel, H.; Simon-Soro, A.; Xiang, Z.; Liu, Y.; Cavalcanti, I.M.; Xiao, J.; Tin, N.-N.; Hara, A.; Drescher, K.; Koo, H. Interkingdom assemblages in human saliva display group-level surface mobility and disease-promoting emergent functions. PNAS 2022, 119, e2209699119. [Google Scholar] [CrossRef]
- Díaz-Garrido, N.; Lozano, C.P.; Kreth, J.; Giacaman, R.A. Extended biofilm formation time by Streptococcus sanguinis modifies its non-cariogenic behavior, in vitro. Braz. Oral Res. 2022, 36, e107–10. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, X.; Ren, B.; Cheng, L. The regulation of hyphae growth in Candida albicans. Virulence 2020, 11, 337–348. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single- and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef]
- Morse, D.J.; Wilson, M.J.; Wei, X.; Bradshaw, D.J.; Lewis, M.A.O.; Williams, D.W. Modulation of Candida albicans virulence in in vitro biofilms by oral bacteria. Lett. Appl. Microbiol. 2019, 68, 337–343. [Google Scholar] [CrossRef]
- Xu, H.; Sobue, T.; Bertolini, M.; Thompson, A.; Vickerman, M.; Nobile, C.J.; Dongari-Bagtzoglou, A.D. S. oralis activates the Efg1 filamentation pathway in C. albicans to promote cross-kingdom interactions and mucosal biofilms. Virulence 2017, 8, 1602–1617. [Google Scholar] [CrossRef]
- Basso, V.; d’Enfert, C.; Znaidi, S.; Bachellier-Bassi, S. From Genes to Networks: The Regulatory Circuitry Controlling Candida albicans Morphogenesis. Curr. Top. Microbiol. Immunol. 2019, 422, 61–99. [Google Scholar] [CrossRef]
- Merino Guzmán, G.; Cedillo Ramírez, L.; Silva Andrade, F.; Muñoz García, A.A.; Castañeda Roldán, E.I. Análisis morfológico de biopelículas de Candida albicans producidas en diferentes condiciones de pH y temperatura analizadas por microscopía óptica y de fuerza atómica. [In English: Morphological analysis of Candida albicans biofilms produced at different pH and temperature conditions analyzed by optic and atomic force microscopy]. Rev. Mex. Mic. 2011, 33, 1–8. [Google Scholar]
- Westwater, C.; Balish, E.; Schofield, D.A. Candida albicans-conditioned medium protects yeast cells from oxidative stress: a possible link between quorum sensing and oxidative stress resistance. Eukaryot. Cell. 2005, 4, 1654–1661. [Google Scholar] [CrossRef]
- Willems, H.M.; Kos, K.; Jabra-Rizk, M. A.; Krom, B.P. Candida albicans in oral biofilms could prevent caries. Pathog. Dis. 2016, 74, ftw039. [Google Scholar] [CrossRef]
- Eidt, G.; Gomes de Andrade, C.; de Cássia, T.; Arthur, R.A. Role of Candida albicans on enamel demineralization and on acidogenic potential of Streptococcus mutans in vitro biofilms. J. Appl. Oral Sci. 2019, 27, e20180593. [Google Scholar] [CrossRef]
- Huang, X.; Schulte, R.M.; Burne, R.A.; Nascimento, M.M. Characterization of the arginolytic microflora provides insights into pH homeostasis in human oral biofilms. Caries Res. 2015, 49, 165–176. [Google Scholar] [CrossRef]
- Díaz-Garrido, N.; Lozano, C.P.; Kreth, J.; Giacaman, R.A. Competition and caries on enamel of a dual-species biofilm model with Streptococcus mutans and Streptococcus sanguinis. Appl. Environ. Microbiol. 2020, 86, e01262–20. [Google Scholar] [CrossRef]
- Szabó, B.; Majoros, L.; Papp-Falusi, E.; Szabó, Z.; Szabó, J.; Márton, I.; Kelentey, B. Studies on the possible aetiological role of different Candida species in pathogenesis of dentine caries by monitoring the calcium release from tooth particles. Acta Microbiol. Immunol. Hung. 2014, 61, 11–17. [Google Scholar] [CrossRef]
- Charone, S.; Portela, M.B.; Martins, K.O.; Soares, R.M.; Castro, G.F. Role of Candida species from HIV infected children in enamel caries lesions: an in vitro study. J. Appl. Oral Sci. 2017, 25, 53–60. [Google Scholar] [CrossRef]
- Kreth, J.; Zhang, Y.; Herzberg, M. Streptococcal antagonism in oral biofilms: Streptococcus sanguinis and Streptococcus gordonii interference with Streptococcus mutans. J. Bacteriol. 2008, 190, 4632–4640. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, Z.; Itzek, A.; Herzberg, M.C.; Kreth, J. CcpA regulates biofilm formation and competence in Streptococcus gordonii. Mol. Oral Microbiol. 2012, 27, 83–94. [Google Scholar] [CrossRef]
- d’Enfert, C.; Kaune, AK.; Alaban, LR.; Chakraborty, S.; Cole, N.; Delavy, M.; et al. The impact of the Fungus-Host-Microbiota interplay upon Candida albicans infections: current knowledge and new perspectives. FEMS Microbiol. Rev. 2021, 45, fuaa060. [Google Scholar] [CrossRef]
- Vila, T.; Kong, E.F.; Ibrahim, A.; Piepenbrink, K.; Shetty, A.C.; McCracken, C.; Bruno, V.; Jabra-Rizk, M.A. Candida albicans quorum-sensing molecule farnesol modulates staphyloxanthin production and activates the thiol-based oxidative-stress response in Staphylococcus aureus. Virulence 2019, 10, 625–642. [Google Scholar] [CrossRef]
- Sebaa, S.; Boucherit-Otmani, Z.; Courtois, P. Effects of tyrosol and farnesol on Candida albicans biofilm. Mol. Med. Rep. 2019, 19, 3201–3209. [Google Scholar] [CrossRef]
- Bowen, W.; Burne, R.; Wu, H.; Koo, H. Oral Biofilms: Pathogens, Matrix, and Polymicrobial Interactions in Microenvironments. Trends Microbiol. 2018, 26, 229–242. [Google Scholar] [CrossRef]
Figure 1.
Quantification of C. albicans hyphae in single- and dual-species biofilms in both experimental conditions. Bars indicate the average of two independent experiments (n=6). Error lines show standard deviation (SD). ANOVA, p<0.05.
Figure 1.
Quantification of C. albicans hyphae in single- and dual-species biofilms in both experimental conditions. Bars indicate the average of two independent experiments (n=6). Error lines show standard deviation (SD). ANOVA, p<0.05.
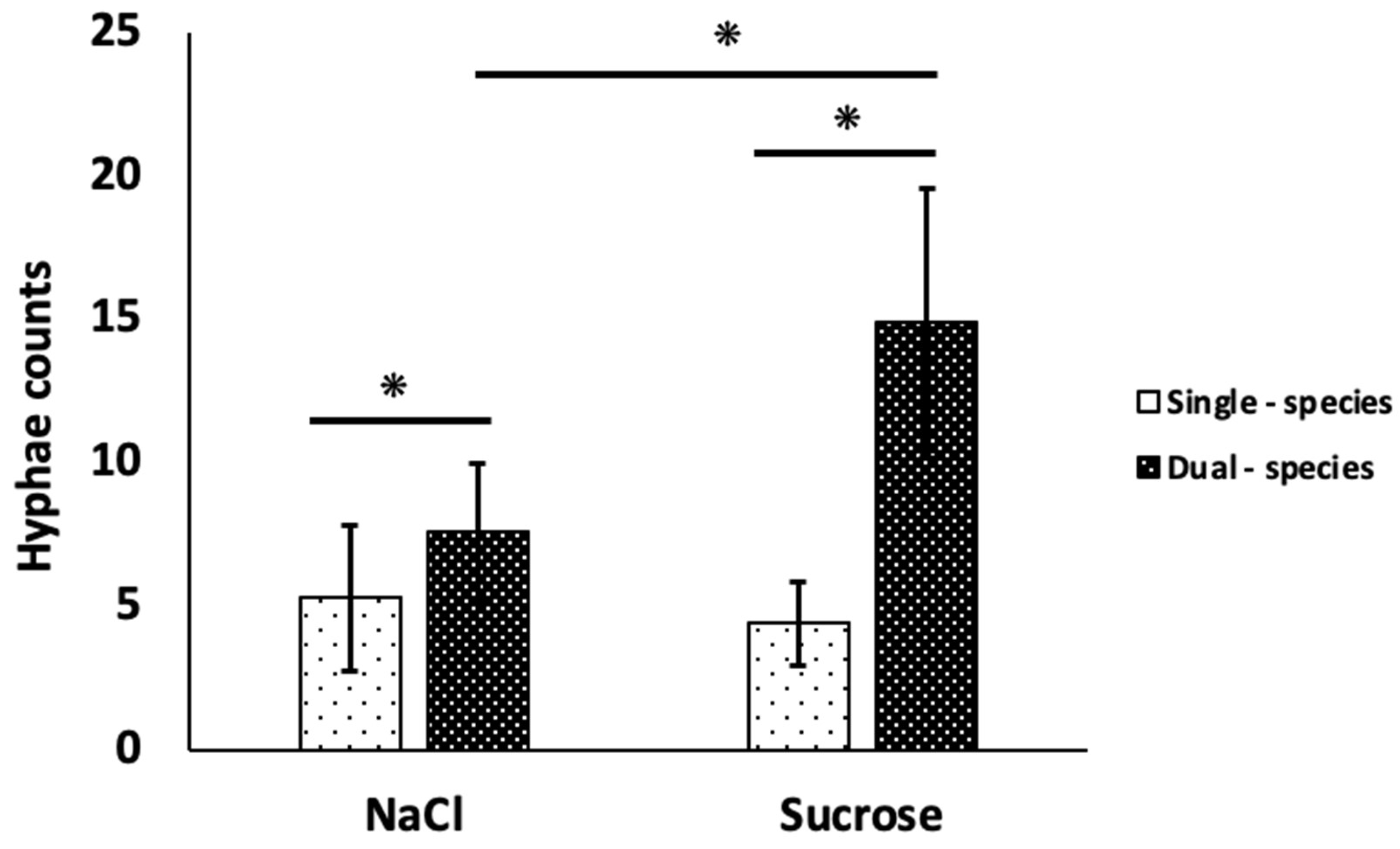
Figure 2.
Representative scanning electron microphotographs obtained from the single- and dual-species biofilms in both experimental conditions. A), C) single- (800x) and dual-species (1300x) biofilms in 0.9% NaCl, respectively, and B), D) single- (500x) and dual-species (4500x) biofilms in 10% sucrose, respectively.
Figure 2.
Representative scanning electron microphotographs obtained from the single- and dual-species biofilms in both experimental conditions. A), C) single- (800x) and dual-species (1300x) biofilms in 0.9% NaCl, respectively, and B), D) single- (500x) and dual-species (4500x) biofilms in 10% sucrose, respectively.

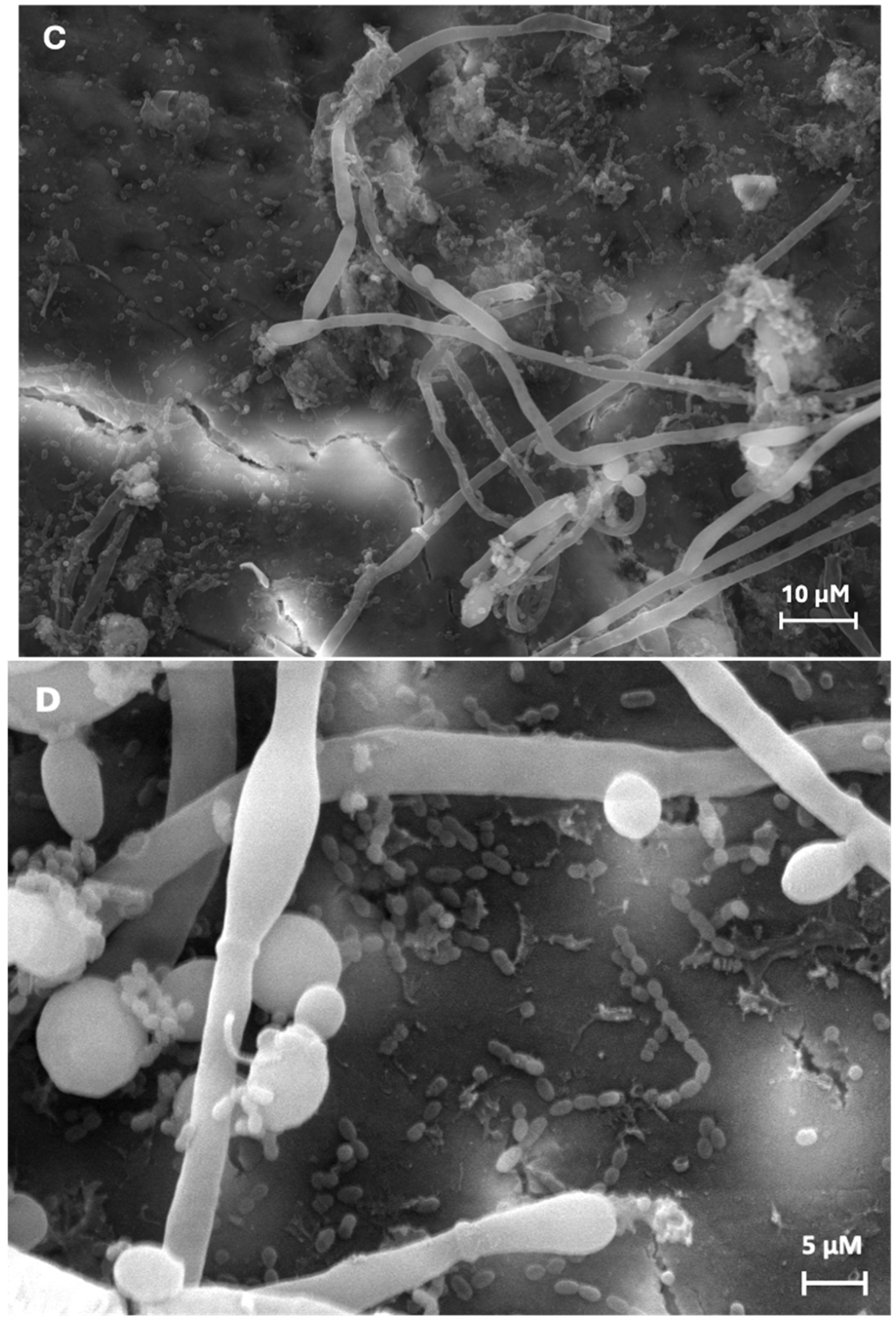
Figure 3.
pH variations during the experiment in single-and dual-species biofilms. The blue line represents the control condition (NaCl) and the red line the sucrose condition from two independent experiments (n=12) in C. albicans biofilm (a); and C. albicans—S. sanguinis biofilm (b), respectively. Asterisk represents statistically significant differences among conditions. Error lines show standard deviation (SD). ANOVA and Tukey’s post hoc test, p<0.05.
Figure 3.
pH variations during the experiment in single-and dual-species biofilms. The blue line represents the control condition (NaCl) and the red line the sucrose condition from two independent experiments (n=12) in C. albicans biofilm (a); and C. albicans—S. sanguinis biofilm (b), respectively. Asterisk represents statistically significant differences among conditions. Error lines show standard deviation (SD). ANOVA and Tukey’s post hoc test, p<0.05.
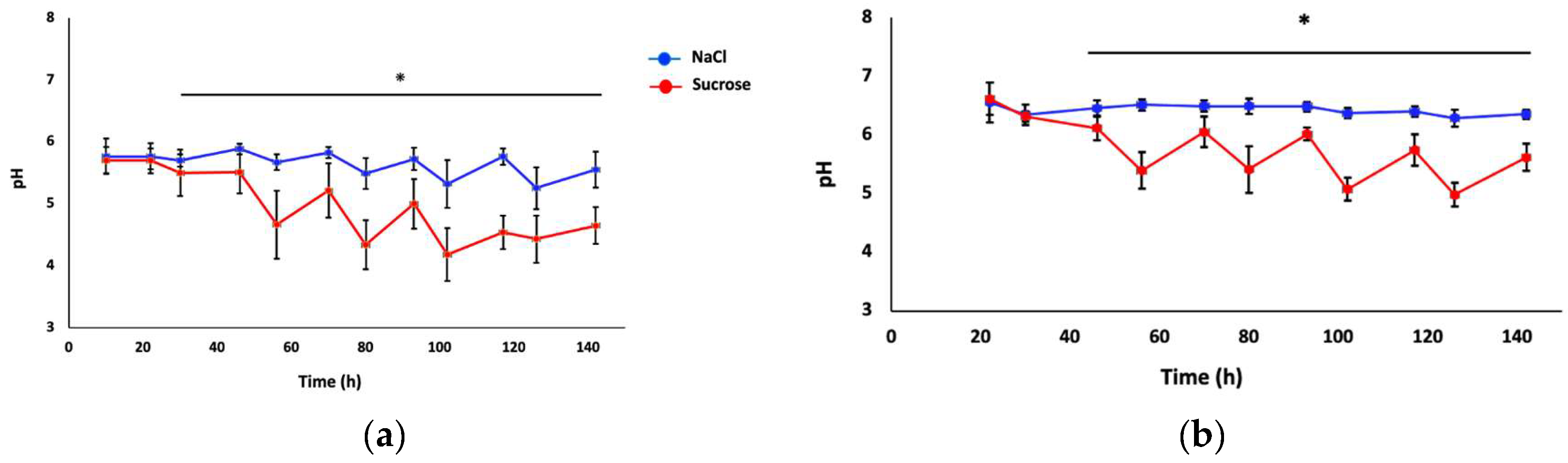
Figure 4.
Enamel slabs demineralization of single- and dual-species biofilms after exposure to sucrose and NaCl condition. The surface microhardness of the enamel slabs was measured before and after the experimental treatment and interpreted as demineralization percentage. Each bar indicates the average of two independent experiments conducted six times (n=12). Error lines show the standard deviation (SD). Asterisks represent statistically significant differences among conditions ANOVA, p<0.05.
Figure 4.
Enamel slabs demineralization of single- and dual-species biofilms after exposure to sucrose and NaCl condition. The surface microhardness of the enamel slabs was measured before and after the experimental treatment and interpreted as demineralization percentage. Each bar indicates the average of two independent experiments conducted six times (n=12). Error lines show the standard deviation (SD). Asterisks represent statistically significant differences among conditions ANOVA, p<0.05.
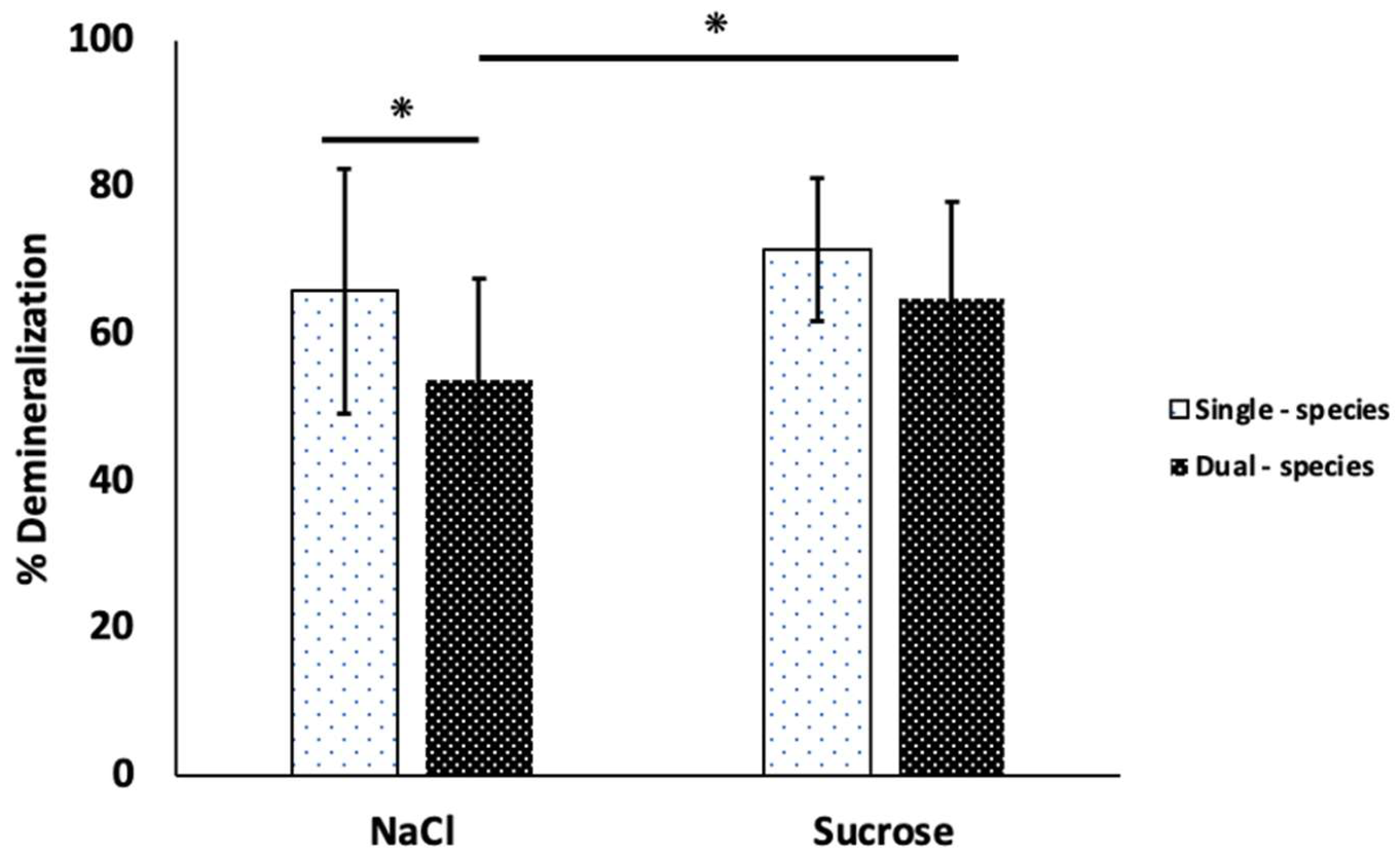

Table 1.
Viable cell counts of C. albicans and S. sanguinis in single- and dual-species biofilms in NaCl and sucrose condition.
Table 1.
Viable cell counts of C. albicans and S. sanguinis in single- and dual-species biofilms in NaCl and sucrose condition.
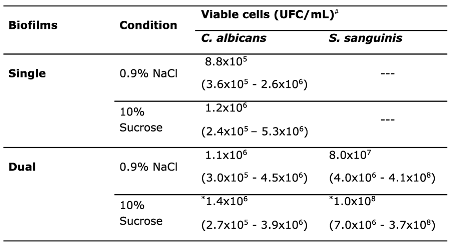 |
#Median (ranges) of two independent experiments conducted six times (n=12). Asterisks represent statistically significant differences among experimental conditions. Wilcoxon test, p<0.05.
Table 2.
H2O2 quantification in single- and dual-species biofilms under NaCl and sucrose conditions.
Table 2.
H2O2 quantification in single- and dual-species biofilms under NaCl and sucrose conditions.
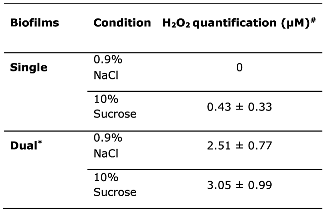 |
#Mean ± SD of two independent experiments conducted three times (n=6). Asterisk represent statistically significant differences among biofilms. T test (p<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



