
Submitted:
30 July 2024
Posted:
02 August 2024
You are already at the latest version
Abstract
Fall armyworm, Spodoptera frugiperda, is one of the major migratory polyphagous insect pests of various crops. The essential nutrient and mineral profile of host plants determines the feeding fitness of herbivorous insects. As a result, growth and development of insects is affected. To determine the effect of nutrient and mineral profile of different host plants (maize, castor bean, cotton, cabbage, okra and sugarcane) on the growth and development of S. frugiperda, biological parameters like larval weight, pupal weight (male/female), feeding and growth indices were calculated. The proximate compositions and mineral contents of the tested host plants showed a significant difference (p<0.5). Better nutrients were observed in castor bean and maize host plants. The growth indices on these host plants also showed the statistical difference (p<0.05) except efficiency of ingested food (p>0.05). The maximum relative growth rate (RGR), relative consumption rate (RCR) and consumption index (CI) were recorded in S. frugiperda larvae that fed on maize and castor bean leaves. Larval, male and female pupal weights were the maximum in the larvae feeding on castor bean host plant. In case of fitness indices, the highest larval growth, pupal growth, standard growth and fitness indices were observed in S. frugiperda larvae that fed on maize. The findings of the present study highlight the dynamic growth and development of S. frugiperda larvae on maize plants and provide novel information based on nutritional ecology to develop sustainable integrated pest management strategies using selective crop rotation and limiting alternative host plants.
Keywords:
Fitness indices
; growth indices
; host plants
; nutritional compositions
; plant minerals
; Spodoptera frugiperda
1. Introduction
Insect herbivores confront with innumerable constraints when they consume their host plants [1]. Plants have evolved certain physical barriers to cope with insect herbivore attacks, such as thick leaves, glandular trichomes, and a wide range of chemical defenses [2,3,4]. A diverse range of natural compounds and constituents, that are unique to a certain plant, act as repellents, deterrents or toxins and impede the digestion of food. However, herbivores have evolved mechanisms to counteract these defenses, by escaping the detection or suppressing plant defenses of host plants [5]. Polyphagous insect herbivores compete with a wide range of plant defenses from various taxa, but their metabolic system is less optimized than mono and oligophagous herbivores [6,7,8].
Therefore, polyphagous insect herbivores have a benefit of getting a better nutritional balance, increased resource availability, and ability to dilute the particular host plant defenses by consuming a variety of plant species; however, their growth and survival fitness and population dynamics may vary on different plant species having varying nutritional and mineral contents [9,10,11]. The nutrients and minerals are also essential for the insects to grow, develop, maintain tissues, reproduce and to get energy. They accomplish these necessities; they feed and try to utilize the host plants in their best interest. Plants with high mineral content are more resistant to insect pests because they are stronger and healthier than nutrient-deficient plants and these nutrients may alter the acceptability of the host plants in herbivore insect pests [12].
A high plant species richness is helpful in reducing insect herbivory risks because specific host plant resistance plays significant role in minimizing the crop losses [13]. The nutrients and minerals composition in the host plants have a direct impact on the physiological characteristics of insects, including their growth, development, and general health [14,15]. Insects require balanced nutrients for their growth, survival, and reproduction. Plants with a higher protein content may promote growth and development of herbivorous insects by increasing their body size, shortening life cycles, and increasing fecundity [16].
Mineral contents in plants, such as phosphorus, potassium, and calcium, may also alter the insect physiology and morphology [17,18]. Imbalanced nutrients can result in suboptimal growth and development in insects [16,19]. The nutritional physiology and development of insect pests are altered by consuming and utilizing the different host plants with varied nutrients and minerals content. The impact of host plants on insect nutritional physiology and development are critical responses. In addition to host plant defense mechanisms, the nutritional and mineral profiles of the host plants can also be helpful in devising the pest management strategies by altering the host plant preference, lifecycle and biology of the insect herbivores [20,21].
The host plants can be managed under good crop selection and rotation plans. While plant nutrient and mineral constituents may be managed with good breeding and crop policies and agricultural systems can be improved for host plant defenses against insect herbivores. This approach involves selecting the crop variations with higher nutrients use efficiency with enhanced natural pest resistance [22,23]. This can also be attained through balanced fertilization based on soil tests, and using nutrient and mineral modifications to reinforce the structural host plant defenses. Additionally, crop rotation and inter-cropping practices are also critical for disrupting the insect pest life cycle and maintaining the soil health [24].
Recently, the fall armyworm (FAW) Spodoptera frugiperda J.E. Smith (Lepidoptera: Noctuidae) has posed a significant threat to maize production worldwide. Maize is a very important cereal crop in the world and stands at 3rd place after wheat and rice based on its production and consumption in different fields [25]. It is used for human, poultry and livestock both as food and feed. Maize plays an important role in combating the global food crisis due to world's rapid population growth. FAW population is a great threat in Pakistan also, and its destructive nature may also harm the rice crop in the coming season after maize, resulting in massive crop protection losses [26].
FAW is a polyphagous pest that can rapaciously diminish production of several agricultural crops [27]. It is a major pest of many field crops because of its preference to feed on all parts of plants, higher fecundity, increased mobility, potential for migration, and capacity to develop resistance against a variety of insecticides [27,28]. It feeds on more than 350 commercial and non-commercial plant species from 76 families, such as wheat, barley, sorghum, maize, and soybean [29]. FAW larvae are voracious feeders; early first and second instars feed on one side of the leaves, and after skeletonization of mandibles, final instars feed on all parts of their host plants [30,31]. In maize, it usually attacks maize during stages 3 to 6, when the whorls' protective zones are still intact. The early feeding kills the growing points, resulting in no more leaf or cob development [28]. Usually, one to two larvae feed in a whorl, as lager larvae can feed on younger larvae to reduce competition. FAW attacks at cobs development stage can devoid cobs of the grains [31,32].
The integration of information regarding the host plant nutrient and mineral components into existing agricultural practices with special reference to use of resistant crop cultivars, intercropping, fertilizers and micronutrients would improve the practical cropping policies. By doing this, IPM will be more resilient towards target insect pest herbivores. This holistic approach will not only boost the plant defense system to cope insects’ herbivory but also encourages sustainable and eco-friendly agricultural practices by maintaining soil fertility and plant health. Moreover, comprehensive farmer education along with research provision for host plant’s nutrients based IPM would further reinforce crop resilience with reduced herbivore damages and overall agricultural sustainability. Focusing the above facts, the current study was conducted, with the goal of determining the effect of mineral and nutritional constituents of different host plants including castor bean (Ricinus communis L.), cotton (Gossypium hirsutum L.), maize (Zea mays L.), cabbage (Brassica oleracea), okra (Abelmoschus esculentus L.) and sugarcane (Saccharum officinarum L.) on the growth, development and feeding physiological fitness of S. frugiperda.
2. Materials and Methods
2.1. Study Area
The present study was carried out at the Department of Entomology, Faculty of Agriculture and Environment, The Islamia University of Bahawalpur, Pakistan. The city is in plain condition with 214m altitude from sea level and has longitude of 29° 25' 5.0448'' N and 71° 40' 14.4660'' E.
2.2. Insect Culture
S. frugiperda larvae were collected from the insecticide free maize fields from university research field area. The larvae were shifted into cylindrical plastic screened cages (30 cm h, 18 cm d) and allowed to feed on natural diet (caster bean leaves) under laboratory conditions (25±2 ˚C, R.H 65±5 % and 16L:8D photoperiod) [33]. After pupation, male and female pupae were identified, paired and placed in Petri dishes and shifted into fresh cylindrical plastic screened cages for mating and egg production after adult emergence. Egg laying sheets were stretched from the top into the rearing cages as oviposition substrate and were fixed with strong rubber bands. Newly emerged FAW adults were provided with a 20% honey solution using the cotton plugs. Egg sheets were removed and shifted into new screened plastic cages until hatching. The larvae were provided with natural diet (caster bean leaves) under the same laboratory conditions until the next generation. Third instar S. frugiperda larvae from 6th generation were used for the experiments.
2.3. Host Plants
Different host plants including castor bean (R. communis L.), cotton (G. hirsutum L.), maize (Z. mays L.), okra (A. esculentus L.) cabbage (B. oleracea) and sugarcane (S. officinarum L.) were maintained as insecticide free source of feed for S. frugiperda larvae at field research area of the Islamia University of Bahawalpur. Fresh leaves of these host plants were used for experiments.
2.4. Analyses of Nutritional and Mineral Profiles of Host Plants
Proximate composition and mineral contents analyses of the host plants were performed at the Department of Animal Nutrition and Central Laboratory Complex (CLC), University of Veterinary and Animal Sciences, Lahore, respectively. Proximate nutritional compositions for crude ash, crude protein, crude fat, dry matter, and crude fiber of the host plants were estimated using the standard protocols (AOAC, 2003). Plant leaves were dried to a constant weight in a hot air oven at 70 °C for 24 h to determine dry matter and then grinded into a fine size with a blinder. The crude protein was determined by Kjeldahl analyses. Crude fat was determined by exhaustive Soxhlet Extraction using petroleum ether (40–60 °C, BP) and crude fiber was estimated by using a fiber analyzer. Atomic absorption spectrophotometer (STA-4800 Spectrophotometer, Stalwart Analytics) was used to determine the mineral contents of the host plants including calcium (Ca), magnesium (Mg), potassium (K), zinc (Zn) and phosphorus (P) from dried leaves of the host plants.
2.5. Experimentation
Three hundred newly hatched larvae were randomly divided into six groups and were allowed to feed on six different host plants i.e., castor bean, cabbage, cotton, maize, sugarcane, okra and sorghum in screened plastic containers (32×15×15 cm). After 3rd instar, they were shifted individually in plastic Petri dishes (8 cm in diameter). Their survival was checked on daily basis. Quantified fresh host plant leaves were offered to the larvae until their pupation on daily basis. Pupae were separated on gender basis. The newly emerged moths were paired (one female and one male) in screened plastic containers (32×15×15 cm) and fed with a 20% honey solution supplied through a small cotton wick. The egg sheets (baby liner cloth) were used as oviposition substrates hanged from the top of the containers fixed with strong rubber bands. Eggs were collected daily and deposited in plastic Petri dishes (8 cm diameter) until the emergence of the neonate larvae to calculate hatching rate. Each treatment was replicated five times. The effects of different diets on biological parameters in terms of larval weight, pupal weight, feeding indices and fitness indices were calculated during the whole experiment. The larvae on the first day of each instar and the pupae on the second day were weighed with a high precision electronic weighing balance (Shimadzu Scientific Instruments, ATX/ATY Unibloc Analytical Balance, Japan).
2.6. Feeding Indices
The larval weight (mg), diet weight (mg), and faeces weight (mg) were recorded before and after the feeding at an interval of 24 h to calculate the feeding indices parameters by using high precision balance (Shimadzu Scientific Instruments, ATX/ATY Unibloc Analytical Balance, Japan) with a range of 200g/0.0001g. Equations to calculate growth indices were derived from suggestions of Waldbauer (1968) [34] as follows:
Where, A is the mean weight (mg) of the insects on last day, B is the original mean weight of insects (mg), D is food biomass ingested (mg) per insect, F is faeces biomass produced (mg) per insect and T is the feeding period (days).
2.7. Fitness Indices
The fitness indices in terms of larval growth index, immature growth index, pupal growth index, standardized growth index, and fitness index of S. frugiperda larvae were calculated with the formulae suggested by Ali et al. (2019) [35] as below
2.8. Statistical Data Analyses
Data regarding larval survival, larval weight and length, pupal weight and length, feeding indices -RGR, RCR, ECI and CI- and growth fitness indices -LGI, IGI, PGI, SGI, and FI- were subjected to factorial ANOVA and means were compared using Duncan Multiple Range (DMR) test at 5% significance level. Nutritional values in terms of crude ash, crude protein, crude fat, dry matter, and crude fiber and minerals Ca, Mg, K, Zn, and P content values from the host plants were subjected to One-way ANOVA and means were compared using DMR test at 5% significance level. Growth and feeding indices were correlated to the proximate composition and mineral contents analyses using Pearson Correlation. All statistical analyses were performed using the SPSS software, version 21 (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Proximate Composition of Different Host Plants Used to Feed S. frugiperda Larvae
The tested host plants showed a statistically significant difference (P<0.001) among their proximate composition in terms of their ash, crude protein, crude fiber, dry matter, and crude fat. The maximum ash content (12.13 %) was found in maize plant sample whiles the minimum ash content (9.17%) was detected in cotton plant sample. The crude protein content was the highest (23.48%) in castor bean plant and sugarcane contained the lowest (4.57%) crude protein value. The maximum (33.20%) crude fiber value was recorded in case of sugarcane plant while its minimum (9.36%) value was observed in maize sample. Similarly, the highest dry matter (43.65%) was recorded in castor bean plant sample and the lowest (18.27%) dry matter content was found in okra plant sample. The maximum crude fat (9.50%) was found in cotton plant sample while its minimum (0.87%) content was found in cabbage plant sample (Figure 1).
3.2. Mineral Composition Analysis of Different Host Plants Used to Feed S. frugiperda Larvae
These host plants also showed a significant difference (P<0.001) among their tested mineral contents including calcium (Ca), magnesium (Mg), potassium (K) and zinc (Zn), and phosphorus (P) (P>0.05). The maximum Ca content (28.33mg) was found in okra plant sample while, minimum Ca (2.49mg) was found in cotton plant. The highest Mg contents (38.92mg) were detected in cabbage plant sample while the lowest (1.36mg) Mg contents were present in sugarcane plant. The maximum (26.31 mg) K content was found in cabbage plant while its minimum (15.93 mg) percent value was seen in cotton plant. Additionally, the highest Zn contents (0.2mg) were recorded in case of okra plant sample while its lowest (0.002 mg) contents were fund in castor bean plant sample (Figure 2).
3.3. Feeding Indices Parameters for S. frugiperda Larvae Feeding on Different Host Plants
The feeding indices calculations revealed that tested host plants including maize, castor bean, cotton, cabbage, okra and sugarcane had significantly affected the relative growth rate (RGR) (F=63.4, P<0.001), relative consumption rate (RCR) (F= 588, P<0.001) and consumption index (CI) (F= 3.14, P<0.05) of S. frugiperda larvae. However, larvae of S. frugiperda did not show significant difference in their efficiency of conversion of ingested food (ECI) (F=3.45, P>0.05) while feeding one these different host plants (Table 1). The highest RGR (1.13 mg/mg/day) value for S. frugiperda larvae was observed when they were feeding on maize plant while the same was found to be the lowest (0.59 mg/mg/day) when S. frugiperda larvae fed on sugarcane leaves. The S. frugiperda larvae also exhibited a similar trend in case of RCR and CI calculations where the maximum RCR (50.47 mg/mg/day) and CI (68.9%) values were calculated when S. frugiperda larvae fed on maize. S. frugiperda larvae showed the minimum RCR and CI (19.2 mg/mg/day and 65.69 %, respectively) values when they fed on the sugarcane host plant (Table 1).
3.4. Growth Parameters for S. frugiperda Larvae Feeding on Different Host plants
3.4.1. Larval Length and Weight
From the data, it was depicted that larval length was significantly differed (F= 1860, P<0.001) among the S. frugiperda larvae when they fed on the different host plants. The larval length was found to be the largest (32.97 mm) when S. frugiperda larvae fed on castor bean plant while the smallest (26.65 mm) larval length was found in larvae which fed on sugarcane plant. It was also depicted that larval weights were significantly (F= 446, P<0.001) differed among S. frugiperda larvae when they fed on the different host plants. The larval weight was found to be the maximum (205 mg) when S. frugiperda larvae fed on castor bean plant material while the minimum larval weight (140 mg) was recorded when S. frugiperda larvae fed on sugarcane plant material (Figure 3).
3.4.2. Pupal Length and Weight
There was a significant difference in female and male pupal length of S. frugiperda (F=4366 and 553 respectively, P<0.001) when fed on six different host plants (Figure 4). Female and male pupae were significantly larger (17.97 and 17.67 mm, respectively) when S. frugiperda larvae fed on castor bean plant while the smallest pupal lengths (14.79 mm and 14.39 mm) were recorded on sugarcane plant. S. frugiperda female and male pupae also showed significant differences (F=705 and 505 respectively, P<0.001) in their weights when S. frugiperda larvae fed on six different host plants (Figure 5). Female and male pupae were significantly heavier (198.6 and 191.6 mg, respectively) when S. frugiperda larvae fed on castor bean plant material while the lowest pupal weights (male 131.8 mg and female 136.2 mg) were recorded when they fed on sugarcane plant material. Female pupae were relatively heavier than male pupae when fed on the provided different host plant materials (P<0.05).
3.4.3. Total Larval Survival Rate
The larval survival of S. frugiperda showed significant difference (F=951, P<0.001) when S. frugiperda fed on six different host plants (Figure 6). The highest and statistically similar survival rate was revealed on maize, castor bean and cotton (96.64%, 96.43% and 96.12%, respectively) while the minimum survival rate (62.10%) was seen in S. frugiperda larvae feeding on sugarcane plant material.
3.5. Growth Fitness Indices Parameters for S. frugiperda Larvae Feeding on Different Host Plants
The data revealed that utilization of different host plants significantly affected (P < 0.001) the larval, pupal, immature and standard growth indices of S. frugiperda larvae (Figure 5-9and). The maximum larval growth index (13.13) was observed in S. frugiperda larvae feeding on maize plant material, while the minimum larval growth index (0.72) was observed when larvae fed on sugarcane. Similarly, the highest pupal growth indices (23.38 and 20.69) of S. frugiperda were recorded on maize and castor bean plants, respectively, while the lowest pupal growth index (12.9) was observed when larvae were fed with okra plant material. Similarly, the S. frugiperda larvae showed the maximum immature growth index (7.55) on castor bean plant and the minimum (4.82) on cabbage followed by maize (5.33) and okra (4.96), respectively. In the same line, the highest standard growth index (26.69) and fitness index (606.79) for S. frugiperda larvae were also recorded on maize plant material as their feed while the lowest standard growth index (8.56) and fitness index (177.81) were observed on sugarcane and cotton, respectively (Table 2).
3.6. Matrix of Pearson’s Correlation Coefficient among Proximate Analysis with Growth and Feeding Indices of S. frugiperda
The results of Pearson’s correlation among fitness, growth indices of FAW larvae and nutritional contents of the host plants are given in table 3 and 4. The crude protein (CP) positively correlated with ash and dry matter (DM). It means that higher protein material will definitely produce higher level of ash and dry matter. Crude fiber (CF) is negatively correlated with CP, ether extract (EE) and ash. A food material rich in crude fiber will be a weaker source of CP, EE and ash. Statistically, highly significant relationship exists between CF and EE. Larval growth Index (LGI), pupal growth Index (PGI), immature growth index (IGI) and standard growth index (SGI) are positively co related with CP of the sample, indicating better larval and pupal growth. There is no linear relationship in case of fitness index (FI) and CP (0.00). In case of crude fiber, it had a significantly negative relationship with fat, ash, LGI, PGI, IGI, SGI and FI, while crude fat had a significant positive correlation with LGI, PGI, IGI, SGI and FI of S. frugiperda. It means that higher fat content does not warrant better larval growth. Moreover, Ash contents of the insect diet have a positive impact on insect growth as indicated in the table.3. More ash contents positively correlated with growth indices (LGI, PGI, IGI and SGI). Because generally, ash is a source of minerals and is relative measure of protein contents of the diet. However, it did not show a pronounce impact on fitness index (FI). Pupal growth Index (PGI), immature growth index (IGI), standard growth index (SGI) and fitness index (FI) showed a weaker co relation with dry matter (Table 3).
The results of Pearson’s correlation showed that growth at various stages (larval and pupal etc.) significantly impact the subsequent stages of the FAW development and resulted into fitness index (FI) as given in Table.3. There are stronger association among these growth indices as they are interplay of each other. For example, LGI have stronger correlation with other growth indices (PGI, IGI and SGI). Likewise, a very strong correlation exists among PGI, IGI and SGI. IGI significantly showed a linear correlation with SGI and similarly SGI also have significant association with overall fitness index (FI). The above given scenario resultantly improved the fitness index of the diet. As we know that healthy larval growth is a key indicator of further better development of all other immature stage of the FAW like pupal etc. Therefore, good larval, pupal growth translates into better standard growth index (SGI) and overall fitness. Consequently, all these parameters determined that a strong fitness index (FI) has been observed due to better LGI PGI, IGI and SGI. In case of feeding indices, the RGR had a statistically positive relationship with RCR, DM and ash contents. The RCR had a significant positive relationship with DM and ash content. DM had a significant positive relation with crude protein. CP had a significant relation with ash while negative relation with crude fiber (Table 4).
4. Discussion
The quality of host plants affects the biological and physiological characteristics of herbivores by changing their food preference, consumption and utilization [36]. Host plants exert selection pressure on insect herbivores, leading to differential physiological and morphological changes [37]. S. frugiperda is a highly polyphagous and invasive pest that attacks numerous economically important crops, including maize, rice, sorghum, soybeans, and cotton [38]. Their survival, growth, development, and reproductive potential are predisposed wide-ranging nutritional, mineral, and secondary metabolic substances of host plants [39]. In the present study, experimental host plants exerted substantial effects on the survival, feeding and growth indices of S. frugiperda larvae. Plant and insect growth are interdependent in several ways. Plants, with different nutrients availability and assimilation ability, differentially affects the herbivore insects [40]. In our study, nutritional and mineral composition of the food plants differed significantly. These variations might affect the plant- insect interactions which ultimately affect their spread and population dynamics on different host crop plants [41].
The nutritional and chemical elements of host plants are influential in host plant resistance and tolerance against insect pests [42]. Feeding indices for instance RGR, RCR, and CI are vital indicators for recognizing resistance in selected crops and executing pest management tactics. In this study, S. frugiperda exhibited the highest feeding indices on maize and castor bean plants. RCR is linked with food bioavailability, nutrients, minerals and allelochemicals and affect the growth and development of S. frugiperda through nutrient assimilation and conversion [43]. The crude protein had a positive relationship with growth indices. Maize and castor bean were identified as suitable hosts for S. frugiperda, supporting higher rates of survival and growth due to their nutritional profiles, particularly higher protein content in maize [44,45] which makes the maize a highly preferred crop plant for S. frugiperda. Our findings align with previous studies by Cock, et al. [46], Ganiger, et al. [47] and Sharanabasappa, et al. [48] which reported maize as a preferred host for S. frugiperda. The higher protein and fat content and lower fiber content in maize and castor bean may enhance S. frugiperda growth and development. The presence of favorable nutritional profiles in the plant makes it ideal for the pest's development, highlighting the need for targeted pest management strategies in maize crop.
Mineral contents in host plants, like K and P, can improve resistance against insect pests by promoting secondary metabolic compound production and reducing carbohydrate accumulation [12,41,49]. This beneficial effect by minerals is largely predominated in many host plants against the plant hoppers, beetles, lepidopterans and mites [12]. Our findings indicated that maize had relatively lower mineral content, making it the most suitable host plant for S. frugiperda. Incorporating minerals into crops could improve their resistance to pests, as the abridged larval survival and body weight of rice leaf folder and sugarcane borer was evidenced due to higher K levels [12,49]. This climaxes the importance of understanding the mineral contents of host plants in devising effective pest management policies.
Insect body growth is influenced by the consumption, utilization and assimilation of plant food. Protein, nitrogen, carbohydrates and water contents are the essential primary nutrients for insect growth and development [39,50]. Our findings showed that the LGI, PGI, SGI and FI were found maximum on maize, while IGI was the maximum on castor bean. Altaf, et al. [51] reported better growth of S. frugiperda on maize compared to sorghum and wheat, likely because of differences in nutritional and mineral content, and defensive compounds in these plants. These factors impact host preference, insect survival, and development. Our results align with Cowie, et al. [52], who also found higher larval survival and development on maize plants as compared to rice. Host plant suitability is indicated by factors like larval development, growth, fecundity, and overall generation time, with higher feeding and growth indices indicating greater suitability [53].
The results showed that S. frugiperda had the highest larval and pupal lengths, weights, and survival rates when fed on maize and castor bean leaves. Female pupae were heavier than male pupae. Differences in nutritional quality, chemical stress, and secondary metabolites in these host plants affected the larval and pupal growth, development and survival [54]. Previously, Awmack and Leather [39] and Kumar, et al. [55] reported that the larval and pupal weights of S. frugiperda and Lymantria dispar (gypsy moth) increased when fed on high protein diets. Our results are aligned with Xie, et al. [56], Liu, et al. [57] and Chi and Su [58] who depicted that female pupae were heavier than male, it might be due to one to two days earlier emergence of female pupae than male. The rate of development and survival is directly associated with the quantity and quality of ingested food; as the quantity of ingested food decreases, the insect becomes smaller and lighter in weight with delayed development [59]. Larval and pupal weights and survival of S. frugiperda are greatly affected by host plants, with significantly weightier larvae and pupae were recorded on maize. This indicates good feeding fitness on maize. Wang, He, Zhang, Liu, Jing and Zhang [53] found no significant variation in male and female pupal weights on the same host plant while, Chen, et al. [60] and He, et al. [61] found heavier male pupae. These disparities are due to different host plants with varied nutritional and mineral values which influence larval fitness, development and ultimate survival [62].
Despite contradictory studies on the impact of plant nutrition and mineral constituents on insect pests, it is evident that these constituents influence the growth and feeding fitness of herbivorous insects. The current findings suggest implementing sustainable mixed cropping systems with non-preferred host plants to disrupt S. frugiperda infestations. Nutrient management strategies can be part of effective cultural control actions in IPM to reduce infestations. Further studies are required to develop effective plant protection strategies focusing on insect nutrition, potentially offering new tools for plant protection. By focusing on the nutritional and mineral needs of both crops and pests, we can develop integrated approaches that not only improve crop yields but also reduce the impact of pests like S. frugiperda. This holistic approach to pest management is essential for ensuring food security and sustainable agricultural practices in the future.
5. Conclusions
Role of host plants in modulating the growth and development of S. frugiperda is crucial for assessing the impact of nutrients and minerals on insect growth and development as these factors play a key role in activating plant defense mechanisms against pest stress. This study intended to determine the effect of nutrients and mineral contents in different host plants on survival, feeding and growth indices of S. frugiperda. Overall, survival rates, larval and pupal performance, and growth and feeding indices were the highest on maize and castor bean plants. This research highlights the threats to maize-growing areas under monocropping systems. These outcomes have weighty implications to understand FAW outbreaks from a nutritional ecology viewpoint and assist in devising new pest control approaches based on nutrient and mineral administration in crops. Additionally, further studies are warranted to evaluate the menaces to other crops that might be susceptible to future incursions.
Author Contributions
Conceptualization, M.S.A. and S.A.; methodology, M.S.A. and S.A.; software, M.S.A.; validation, S.A.; formal analysis, M.S.A. and S.A.; investigation, M.S.A.; resources, S.A.; data curation, S.A., A.J., and M.F.S.; writing—original draft preparation, M.S.A.; writing—review and editing, S.A., E.R., A.J., M.F.S., and S.C.; visualization, M.S.A. and S.A.; supervision, S.A.; project administration, M.F.S..; funding acquisition, S.C., and A.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the first corresponding author.
Acknowledgments
The authors are thankful for technical support of lab and field staff during the experimentation and data collection.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sarmento, R.A.; Lemos, F.; Bleeker, P.M.; Schuurink, R.C.; Pallini, A.; Oliveira, M.G.A.; Lima, E.R.; Kant, M.; Sabelis, M.W.; Janssen, A. A herbivore that manipulates plant defence. Ecol. Lett. 2011, 14, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.J.; Rausher, M.D. Evolution of resistance to a multiple-herbivore community: genetic correlations, diffuse coevolution, and constraints on the plant's response to selection. Evolution 2013, 67, 1767–1779. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.; Villarroel, C.; Ataide, L.; Dermauw, W.; Glas, J. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Intraspecific variation in a generalist herbivore accounts for differential induction and impact of host plant defences. Proc. R. Soc. Ser. B Biol. Sci. 2008, 275, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Pentzold, S.; Zagrobelny, M.; Roelsgaard, P.S.; Møller, B.L.; Bak, S. The multiple strategies of an insect herbivore to overcome plant cyanogenic glucoside defence. PloS One 2014, 9, e91337. [Google Scholar] [CrossRef] [PubMed]
- Halon, E.; Eakteiman, G.; Moshitzky, P.; Elbaz, M.; Alon, M.; Pavlidi, N.; Vontas, J.; Morin, S. Only a minority of broad-range detoxification genes respond to a variety of phytotoxins in generalist Bemisia tabaci species. Sci. Rep. 2015, 5, 17975. [Google Scholar] [CrossRef] [PubMed]
- Katsanis, A.; Rasmann, S.; Mooney, K.A. Herbivore diet breadth and host plant defense mediate the tri-trophic effects of plant toxins on multiple coccinellid predators. PLoS One 2016, 11, e0155716. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.J.; Ray, S.; Davidson-Lowe, E.; Ali, J.; Luthe, D.S.; Felton, G. Plant nutrition influences resistant maize defense responses to the fall armyworm (Spodoptera frugiperda). Front. Ecol. Evol. 2022, 10, 844274. [Google Scholar] [CrossRef]
- Pentzold, S.; Zagrobelny, M.; Rook, F.; Bak, S. How insects overcome two-component plant chemical defence: plant β-glucosidases as the main target for herbivore adaptation. Biol. Rev. 2014, 89, 531–551. [Google Scholar] [CrossRef]
- Beran, F.; Petschenka, G. Sequestration of plant defense compounds by insects: from mechanisms to insect–plant coevolution. Annu. Rev. Entomol. 2022, 67, 163–180. [Google Scholar] [CrossRef]
- Bala, K.; Sood, A.; Pathania, V.S.; Thakur, S. Effect of plant nutrition in insect pest management: A review. J. Pharmacognosy Phytochem. 2018, 7, 2737–2742. [Google Scholar]
- Castagneyrol, B.; Jactel, H.; Vacher, C.; Brockerhoff, E.G.; Koricheva, J. Effects of plant phylogenetic diversity on herbivory depend on herbivore specialization. J. Appl. Ecol. 2014, 51, 134–141. [Google Scholar] [CrossRef]
- Bernays, E.A.; Chapman, R.F. Host-plant selection by phytophagous insects; Springer Science & Business Media: 2007.
- Wang, D.; Zhou, L.; Wang, Q.; Ding, J. Plant chemistry determines host preference and performance of an invasive insect. Front. Plant Sci. 2020, 11, 594663. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.; Simpson, C. The mechanisms of nutritional compensation by phytophagous insects. Insect-Plant Interactions 2017, 111–160. [Google Scholar]
- Singh, V.; Sood, A. Plant Nutrition: A tool for the management of hemipteran insect-pests-A review. Agri. Rev. 2017, 38, 260–270. [Google Scholar] [CrossRef]
- Queiroz, R.B.; Lopes, M.C.; Costa, T.L.; da Silva, R.S.; Galdino, T.V.; Gontijo, P.d.C.; Martinez, H.E.P.; Picanço, M.C. Influence of tomato plants nutritional status on the fitness and damage of Tuta absoluta (Lepidoptera: Gelechiidae). Agric. For. Entomol. 2022, 24, 260–266. [Google Scholar] [CrossRef]
- Cease, A.J.; Harrison, J.F.; Hao, S.; Niren, D.C.; Zhang, G.; Kang, L.; Elser, J.J. Nutritional imbalance suppresses migratory phenotypes of the Mongolian locust (Oedaleus asiaticus). R. Soc. Open Sci. 2017, 4, 161039. [Google Scholar] [CrossRef]
- Sarwar, M.; Ahmad, N.; Toufiq, M. Identification of susceptible and tolerant gram (Cicer arietinum L.) genotypes against gram pod borer (Helicoverpa armigera)(Hubner). Pak. J. Bot.
- Kassi, A.K.; Javed, H.; Mukhtar, T. Relationship of physico-morphic characters of okra cultivars with their resistance to Helicoverpa armigera. Pak. J. Zool. 2019, 51, 835. [Google Scholar] [CrossRef]
- Snyder, L.D.; Gómez, M.I.; Power, A.G. Crop varietal mixtures as a strategy to support insect pest control, yield, economic, and nutritional services. Front. Sustainable Food Syst. 2020, 4, 60. [Google Scholar] [CrossRef]
- Jaworski, C.C.; Thomine, E.; Rusch, A.; Lavoir, A.V.; Wang, S.; Desneux, N. Crop diversification to promote arthropod pest management: A review. Agri. Comm. 2023, 1, 100004. [Google Scholar] [CrossRef]
- Ratnadass, A.; Fernandes, P.; Avelino, J.; Habib, R. Plant species diversity for sustainable management of crop pests and diseases in agroecosystems: a review. Agron. Sustainable Dev. 2012, 32, 273–303. [Google Scholar] [CrossRef]
- Khan, A.G.; Imran, M.; Khan, A.-u.-H.; Fares, A.; Šimůnek, J.; Ul-Haq, T.; Alsahli, A.A.; Alyemeni, M.N.; Ali, S. Performance of spring and summer-sown maize under different irrigation strategies in Pakistan. Sustainability 2021, 13, 2757. [Google Scholar] [CrossRef]
- Yousaf, S.; Rehman, A.; Masood, M.; Ali, K.; Suleman, N. Occurrence and molecular identification of an invasive rice strain of fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) from Sindh, Pakistan, using mitochondrial cytochrome c oxidase I gene sequences. J. Plant Dis. Prot. 2022, 129, 71–78. [Google Scholar] [CrossRef]
- Montezano, D.G.; Sosa-Gómez, D.; Specht, A.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.d.; Peterson, J.A.; Hunt, T. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Malo, M.; Hore, J. The emerging menace of fall armyworm (Spodoptera frugiperda JE Smith) in maize: A call for attention and action. J. Entomol. Zool. Stud. 2020, 8, 455–465. [Google Scholar]
- Yang, X.; Sun, X.; Zhao, S.; Li, J.; Chi, X.; Jiang, Y.; Wu, K. Population occurrence, spatial distribution and sampling technique of fall armyworm Spodoptera frugiperda in wheat fields. Plant Prot. 2020, 46, 23. [Google Scholar]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J. Fall armyworm: impacts and implications for Africa. Outlooks Pest Manage. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Gilal, A.A.; Bashir, L.; Faheem, M.; Rajput, A.; Soomro, J.A.; Kunbhar, S.; Mirwani, A.S.; Mastoi, G.S.; Sahito, J.G.M. First record of invasive fall armyworm (Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae)) in corn fields of Sindh, Pakistan. Pak. J. Agri. Res. 2020, 33(2), 47. [Google Scholar] [CrossRef]
- FAO. Integrated management of the fall armyworm on maize: a guide for farmer field schools in Africa. 2018.
- Gergs, A.; Baden, C.U. A dynamic energy budget approach for the prediction of development times and variability in Spodoptera frugiperda rearing. Insects 2021, 12, 300. [Google Scholar] [CrossRef]
- Waldbauer, G. The consumption and utilization of food by insects. In Adv. Insect Physiol., Elsevier, 1968, 5, 229–288. [Google Scholar]
- Ali, S.; Ullah, M.I.; Saeed, M.F.; Khalid, S.; Saqib, M.; Arshad, M.; Afzal, M.; Damalas, C.A. Heavy metal exposure through artificial diet reduces growth and survival of Spodoptera litura (Lepidoptera: Noctuidae). Environ. Sci. Poll. Res. 2019, 26, 14426–14434. [Google Scholar] [CrossRef]
- Cabezas, M.; Nava, D.; Geissler, L.; Melo, M.; Garcia, M.; Krüger, R. Development and leaf consumption by Spodoptera cosmioides (Walker) (Lepidoptera: Noctuidae) reared on leaves of agroenergy crops. Neotrop. Entomol. 2013, 42, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Barros, E.M.; Torres, J.B.; Ruberson, J.R.; Oliveira, M.D. Development of Spodoptera frugiperda on different hosts and damage to reproductive structures in cotton. Entomol. Exp. Appl. 2010, 137, 237–245. [Google Scholar] [CrossRef]
- Neelima, C.; Nameirakam, L.; Devi, Y.K.; Scholar, M. The invasive fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) in Maize, status and infestation taken control under sustainable management: A Review. J. Emerg. Technol. Innov. Res 2020, 7, 1459–1471. [Google Scholar]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Haldhar, S.M.; Bhargava, R.; Choudhary, B.; Pal, G.; Kumar, S. Allelochemical resistance traits of muskmelon (Cucumis melo) against the fruit fly (Bactrocera cucurbitae) in a hot arid region of India. Phytoparasitica 2013, 41, 473–481. [Google Scholar] [CrossRef]
- Shah, J.; Walling, L. Advances in plant-hemipteran interactions. Front. Plant Sci. 2017, 8, 1652. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Prakash, A.; Johri, B. Induced systemic resistance (ISR) in plants: mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef]
- De Sousa Ramalho, F.; Azeredo, T.L.; de Nascimento, A.R.B.; Fernandes, F.S.; Nascimento Júnior, J.L.; Malaquias, J.B.; da Silva, C.A.D.; Zanuncio, J.C. Feeding of fall armyworm, Spodoptera frugiperda, on Bt transgenic cotton and its isoline. Entomol. Exp. Appl. 2011, 139, 207–214. [Google Scholar] [CrossRef]
- Hutasoit, R.T.; Kalqutny, S.H.; Widiarta, I.N. Spatial distribution pattern, bionomic, and demographic parameters of a new invasive species of armyworm Spodoptera frugiperda (Lepidoptera; Noctuidae) in maize of South Sumatra, Indonesia. Biodivers. J. Biol. Divers. 2020, 21. [Google Scholar] [CrossRef]
- Maharani, Y.; Puspitaningrum, D.; Istifadah, N.; Hidayat, S.; Ismail, A. Biology and life table of fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) on maize and rice. Serangga 2021, 26, 161–174. [Google Scholar]
- Cock, M.J.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef] [PubMed]
- Ganiger, P.; Yeshwanth, H.; Muralimohan, K.; Vinay, N.; Kumar, A.; Chandrashekara, K. Occurrence of the new invasive pest, fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), in the maize fields of Karnataka, India. Curr. Sci. 2018, 115, 621–623. [Google Scholar] [CrossRef]
- Sharanabasappa, S.; Kalleshwaraswamy, C.; Maruthi, M.; Pavithra, H. Biology of invasive fall army worm Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) on maize. Indian J. Entomol. 2018, 80, 540–543. [Google Scholar] [CrossRef]
- Rashid, M.M.; Jahan, M.; Islam, K.S. Impact of nitrogen, phosphorus and potassium on brown planthopper and tolerance of its host rice plants. Rice Sci. 2016, 23, 119–131. [Google Scholar] [CrossRef]
- Hosseininejad, A.; Naseri, B.; Razmjou, J. Comparative feeding performance and digestive physiology of Helicoverpa armigera (Lepidoptera: Noctuidae) larvae-fed 11 corn hybrids. J. Insect Sci. 2015, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Altaf, N.; Idrees, A.; Ullah, M.I.; Arshad, M.; Afzal, A.; Afzal, M.; Rizwan, M.; Li, J. Biotic potential induced by different host plants in the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 921. [Google Scholar] [CrossRef] [PubMed]
- Cowie, B.W.; Strathie, L.W.; Goodall, J.M.; Venter, N.; Witkowski, E.T.; Byrne, M.J. Does host plant quality constrain the performance of the Parthenium beetle Zygogramma bicolorata? Biol. Cont. 2019, 139, 104078. [Google Scholar] [CrossRef]
- Wang, W.; He, P.; Zhang, Y.; Liu, T.; Jing, X.; Zhang, S. The population growth of Spodoptera frugiperda on six cash crop species and implications for its occurrence and damage potential in China. Insects 2020, 11, 639. [Google Scholar] [CrossRef]
- Wouters, F.C.; Blanchette, B.; Gershenzon, J.; Vassão, D.G. Plant defense and herbivore counter-defense: benzoxazinoids and insect herbivores. Phytochem. Rev. 2016, 15, 1127–1151. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.M.; Gadratagi, B.G.; Paramesh, V.; Kumar, P.; Madivalar, Y.; Narayanappa, N.; Ullah, F. Sustainable management of invasive fall armyworm, Spodoptera frugiperda. Agronomy 2022, 12, 2150. [Google Scholar] [CrossRef]
- Xie, W.; Zhi, J.; Ye, J.; Zhou, Y.; Li, C.; Liang, Y.; Yue, W.; Li, D.; Zeng, G.; Hu, C. Age-stage, two-sex life table analysis of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) reared on maize and kidney bean. Chem. Biol. Technol. Agric. 2021, 8, 1–8. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, Y.; Wang, Q.; Liu, X.; Fu, Y.; Zhang, Y.; Chen, J. Push–pull plants in wheat intercropping system to manage Spodoptera frugiperda. J. Pest Sci. 2023, 96, 1579–1593. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.-Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Khedr, M.A.; Al-Shannaf, H.M.; Mead, H.M.; Shaker, S.A. Comparative study to determine food consumption of cotton leafworm, Spodoptera littoralis, on some cotton genotypes. J. Plant Prot. Res. 2015, 55. [Google Scholar] [CrossRef]
- Chen, W.H.; Itza, B.; Kafle, L.; Chang, T.Y. Life table study of fall armyworm (Spodoptera frugiperda) (Lepidoptera: Noctuidae) on three host plants under laboratory conditions. Insects 2023, 14, 329. [Google Scholar] [CrossRef]
- He, L.M.; Wang, T.L.; Chen, Y.C.; Ge, S.S.; Wyckhuys, K.A.; Wu, K.M. Larval diet affects development and reproduction of East Asian strain of the fall armyworm, Spodoptera frugiperda. J. Integr. Agric. 20, 736–744. [CrossRef]
- He, L.M.; Wu, Q.L.; Gao, X.W.; Wu, K.M. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J. Integr. Agric. 20, 745–754. [CrossRef]
Figure 1.
Proximate composition analysis (%±SE) of different host plants used as feed for S. frugiperda larvae.
Figure 1.
Proximate composition analysis (%±SE) of different host plants used as feed for S. frugiperda larvae.
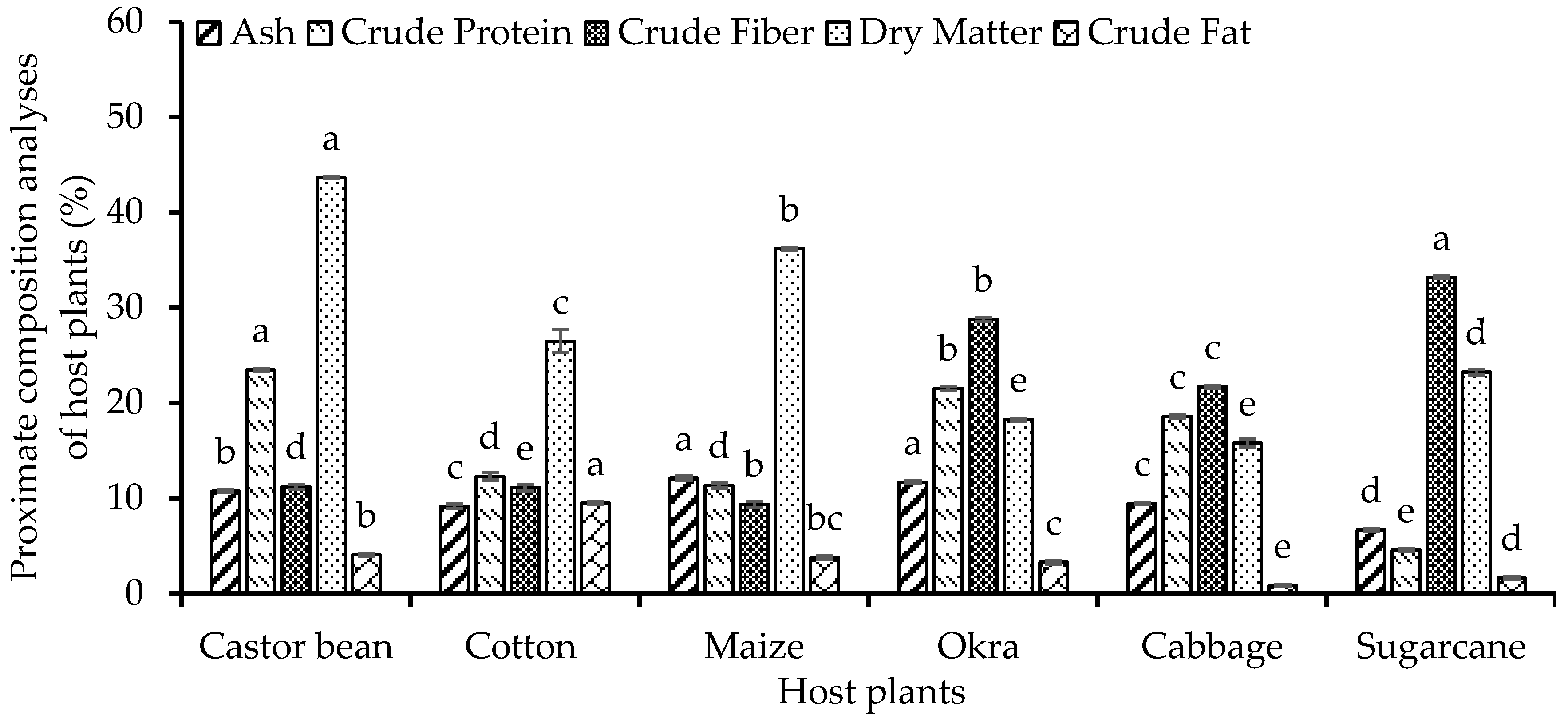
Figure 2.
Mineral composition (mg/g±SE) in different host plants used as feed for S. frugiperda larvae.
Figure 2.
Mineral composition (mg/g±SE) in different host plants used as feed for S. frugiperda larvae.

Figure 3.
Larval weight (mg±SE) and larval length (mm±SE) of S. frugiperda in response to their feeding on different host plants.
Figure 3.
Larval weight (mg±SE) and larval length (mm±SE) of S. frugiperda in response to their feeding on different host plants.

Figure 4.
Pupal length (mm±SE) of S. frugiperda in response to their feeding on different host plants.
Figure 4.
Pupal length (mm±SE) of S. frugiperda in response to their feeding on different host plants.
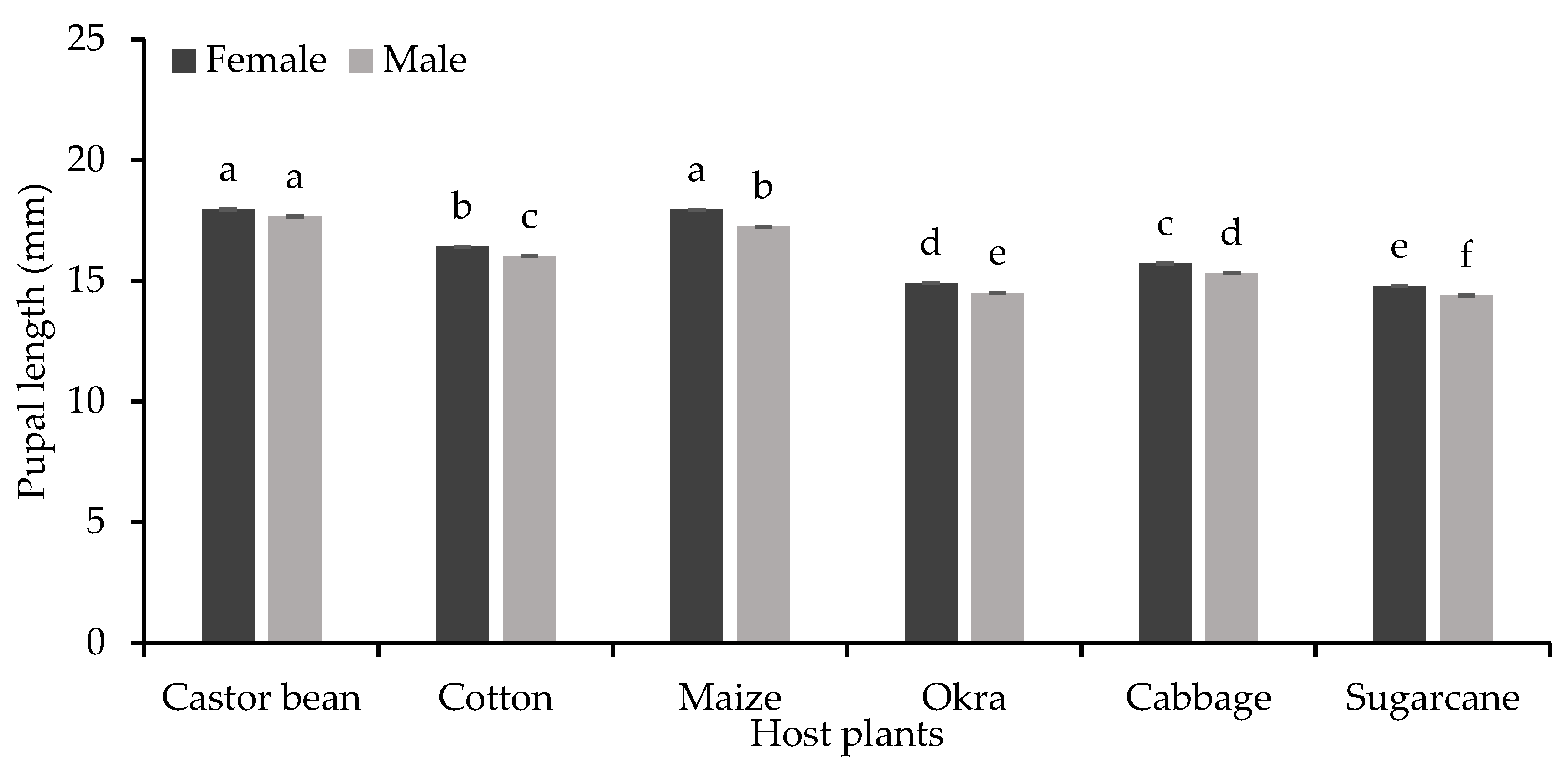
Figure 5.
Pupal weight (mg±SE) of S. frugiperda in response to their feeding on different host plants.
Figure 5.
Pupal weight (mg±SE) of S. frugiperda in response to their feeding on different host plants.
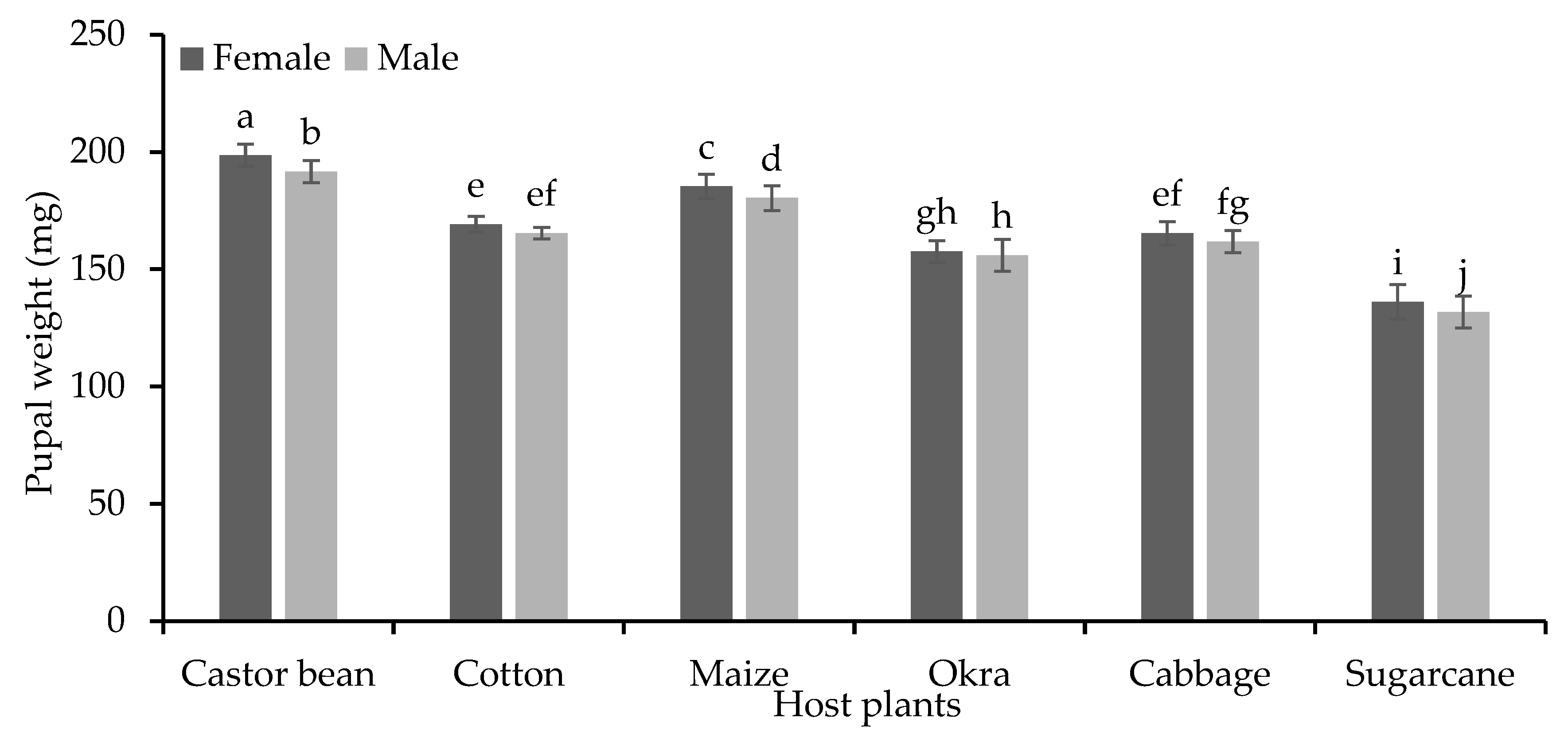
Figure 6.
Survival (%±SE) of S. frugiperda in response to their feeding on different host plants.
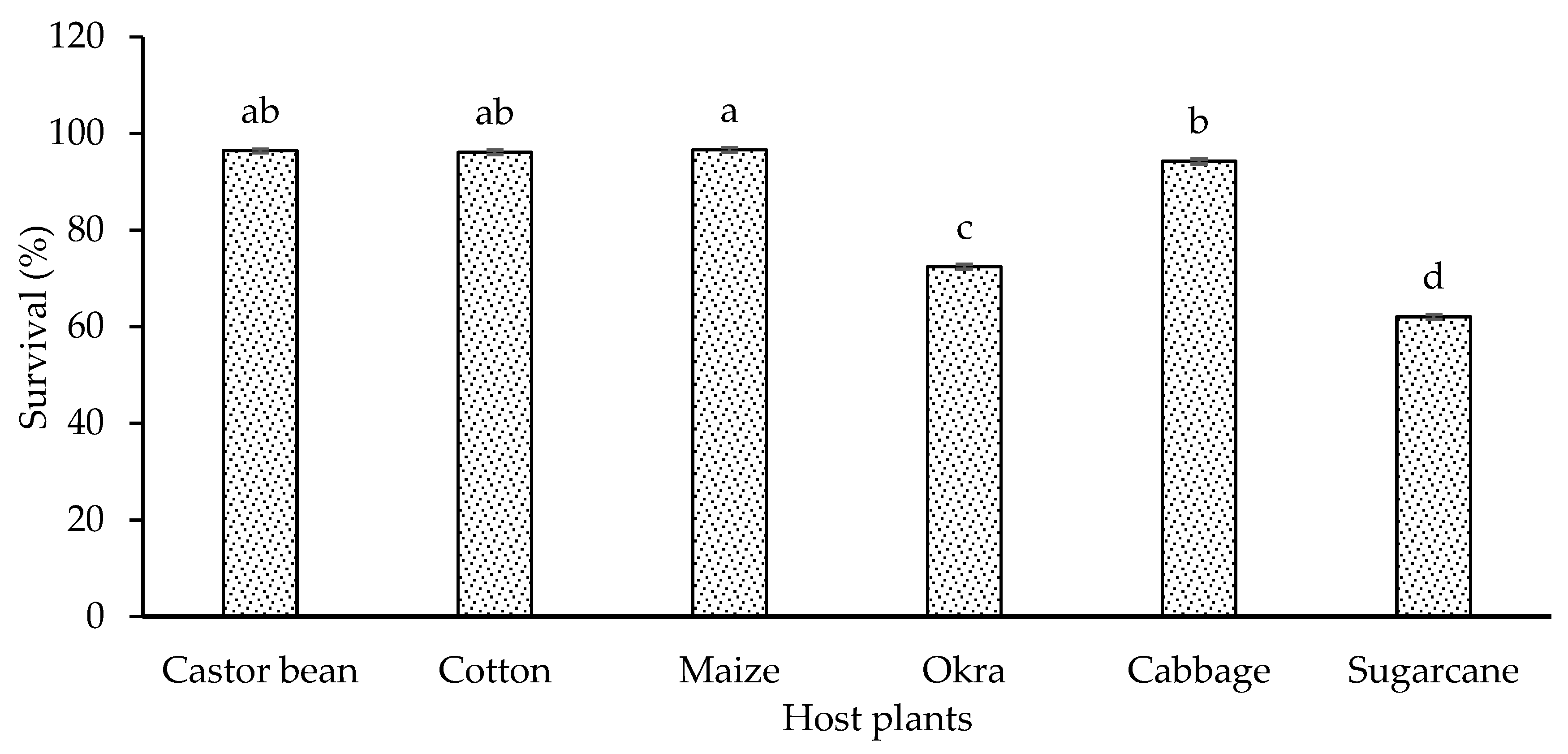

Table 1.
Feeding indices of S. frugiperda larvae in response to their feeding on different host plants.
Table 1.
Feeding indices of S. frugiperda larvae in response to their feeding on different host plants.
| Host plants | RGR | RCR | ECI | CI |
|---|---|---|---|---|
| Castor bean | 0.97±0.03b | 47.13±0.45b | 71.80±1.25a | 67.68ab |
| Cotton | 0.85±0.01c | 33.49±0.47c | 71.11±1.32a | 67.84ab |
| Maize | 1.13±0.03a | 50.47±0.61a | 70.77±1.45a | 68.39a |
| Okra | 0.71±0.03d | 19.66±0.46d | 69.22±2.03a | 67.08ab |
| Cabbage | 0.77±0.01cd | 33.22±0.50c | 69.87±1.40a | 67.47ab |
| Sugarcane | 0.59±0.01e | 19.2±0.40d | 70.02±1.69a | 65.69b |
| df | 5 | 5 | 5 | 5 |
| F-value | 63.4 | 588 | 3.45 | 3.14 |
| p-value | <0.001 | <0.001 | 0.08 | 0.03 |
Means in columns sharing similar letters are not significantly different according to Duncan Multiple Range (DMR) test at P>0.05.
Table 2.
Growth fitness indices of S. frugiperda larvae in response to their feeding on different host plants.
Table 2.
Growth fitness indices of S. frugiperda larvae in response to their feeding on different host plants.
| Host plants | Larval growth Index | Pupal Growth Index | Immature Growth Index | Standard Growth Index | Fitness Index |
|---|---|---|---|---|---|
| Castor bean | 10.21±0.85b | 20.69±0.52a | 7.55±0.15a | 21.53±0.17b | 408.21±41.04b |
| Cotton | 5.48±0.34d | 15.47±1.50b | 6.05±0.82ab | 15.62±0.51d | 177.81±10.52c |
| Maize | 13.13±0.66a | 23.38±0.41a | 5.33±0.36a | 26.59±1.22a | 606.79±27.95a |
| Okra | 8.57±0.71bc | 12.9±0.58b | 4.96±0.95b | 17.87±0.36c | 233.08±19.96c |
| Cabbage | 7.18±0.56cd | 15.95±0.82b | 4.82±0.88b | 16.52±0.46cd | 192.15.75c |
| Sugarcane | 0.72±0.10e | 0±0.00c | -- | 8.56±1.88e | -- |
| df | 5 | 5 | 4 | 5 | 4 |
| F-value | 51.4 | 108 | 2.5 | 74.7 | 52 |
| p-value | <0.001 | <0.001 | 0.04 | <0.001 | <0.001 |
Means in columns sharing similar letters are not significantly different according to Duncan Multiple Range (DMR) test at P>0.05.
Table 3.
Matrix of Pearson’s correlation coefficient among proximate analysis and growth indices of S. frugiperda.
Table 3.
Matrix of Pearson’s correlation coefficient among proximate analysis and growth indices of S. frugiperda.
| DM | CP | CF | EE | Ash | LGI | PGI | IGI | SGI | |
|---|---|---|---|---|---|---|---|---|---|
| CP | 0.43* | ||||||||
| CF | -0.30 | -0.73** | |||||||
| EE | 0.13 | -0.04 | -0.59** | ||||||
| Ash | 0.26 | 0.62** | -0.38* | 0.10 | |||||
| LGI | -0.15 | 0.19 | -0.45* | 0.64** | 0.47** | ||||
| PGI | 0.14 | 0.39* | -0.60** | 0.68** | 0.69** | 0.87** | |||
| IGI | 0.06 | 0.40* | -0.58** | 0.57** | 0.58** | 0.82** | 0.88** | ||
| SGI | -0.05 | 0.21 | -0.50** | 0.70** | 0.48** | 0.93** | 0.90** | 0.80** | |
| FI | -0.09 | 0.00 | -0.41* | 0.80** | 0.34 | 0.95** | 0.85** | 0.76** | 0.92** |
DM (dry matter), CP (crude protein), CF (crude fiber), EE (ether extract/ crude fat), LGI (larval growth index), PGI (pupal growth index), IGI (immature growth index), SGI (standard growth index), FI (fitness index)
Table 4.
Matrix of Pearson’s correlation coefficient among proximate analysis and feeding indices of S. frugiperda.
Table 4.
Matrix of Pearson’s correlation coefficient among proximate analysis and feeding indices of S. frugiperda.
| RGR | RCR | DM | CP | CF | EE | Ash | |
|---|---|---|---|---|---|---|---|
| RCR | 0.91** | ||||||
| DM | 0.50** | 0.64** | |||||
| CP | 0.21 | 0.23 | 0.43* | ||||
| CF | -0.10 | -0.05 | -0.30 | -0.73** | |||
| EE | 0.29 | 0.21 | 0.13 | -0.04 | -0.59 | ||
| ASH | 0.69** | 0.53** | 0.26 | 0.62** | -0.38 | 0.10 | |
| ECI | -0.30 | 0.43 | 0.01 | 0.13 | 0.64 | 0.52 | 0.26 |
RGR (relative growth rate), RCR (relative consumption rate), ECI (efficiency of conversion of ingested food), DM (dry matter), CP (crude protein), CF (crude fiber), EE (ether extract/ crude fat)
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



