
Submitted:
30 July 2024
Posted:
31 July 2024
You are already at the latest version
Abstract
Additives for anaerobic digestion (AD) can play a significant role in optimising the process by increasing biogas production, stabilising the system and improving digestate quality. The role of additives largely boils down to: (i) enhancing direct interspecies electron transfer (DIET) between microbial communities, resulting in improved syntrophic interactions and methane production rates (e.g. biochar, magnetite and carbon nanotubes), (ii) adsorption of toxic substances that may inhibit microbial activity (e.g. activated carbon, zeolites), (iii) improving microbial activity and increasing process stability (e.g. cobalt, nickel, iron, selenium), (iv) maintaining optimal pH levels for microbial activity (e.g. magnesium oxide), (v) reducing inhibition (the aforementioned adsorbents and conductive substances), (vi) accelerating the decomposition of complex organic materials into simpler compounds that are more easily digested by microorganisms, thereby increasing the rate of hydrolysis (enzymes, including cellulases, proteases and lipases). Through the aforementioned action, additives can significantly affect AD performance. The function of these materials varies, from enhancing microbial activity to maintaining optimal conditions and protecting the system from inhibitors. The choice of additives should be carefully tailored to the specific needs and conditions of the digester to maximise benefits and ensure sustainability. In light of these considerations, this paper characterizes the most commonly used additives and their combinations based on a comprehensive review of recent scientific publications, including a report on the results of conducted studies. The publication features chapters that describe: carbon-based conductive materials, metal oxide nanomaterials, trace metal and biological additives, including enzymes and microorganisms. It concludes with a chapter summarising reports on various additives and discussing their indications for functional systems with determined properties. A notable advantage of this work is the updated literature data, clear summaries, and a substantive description of the performance of the additives discussed.
Keywords:
anaerobic digestion
; additives
; material systems
; enhanced efficiency
; DIET
1. Introduction
Anaerobic digestion (AD), as an environmentally friendly method of waste management and a source of renewable energy in the form of biogas, is a highly recommended solution in times of ongoing geopolitical changes and energy crises [1]. AD is a complex, multistep biotechnological process, and its efficiency is highly dependent on sensitive microorganisms [2,3]. The performance of these microorganisms is influenced by factors such as substrate type, process environment conditions (e.g., pH and temperature), bioreactor design, operational modes, and access to the medium. Various challenges can adversely affect the process, including inefficient biodegradation, acidification, lag phases, foam formation, complex rheology, and high apparent viscosity [4].
Several methods are employed to enhance AD, such as substrate pretreatment, co-digestion, and modifications to reactor configurations and operating conditions. One promising approach to improving AD kinetics and effectiveness is the use of additives in anaerobic reactors. These additives can be broadly categorised into inorganic additives (e.g. carbon-based materials, macro- and micro-elements, nanoparticles, and nanomaterials) and biological additives (e.g. microorganisms and enzymes) [5]. While existing scientific research on AD additives is both encouraging and revealing, many issues remain unresolved. The published results offer a solid foundation and inspiration for further research into new materials, additive systems with defined effects, and advanced analyses to clarify key phenomena and the relationships between additive properties and their functions [6].
Studies on the supplementation of the AD process with inorganic, organic, and biological additives have shown positive results in enhancing the efficiency of digesters. Among the notable additives are carbon-based conductive materials, such as Granular Activated Carbon (GAC), Powdered Activated Carbon (PAC), and multi-wall or single-wall Carbon Nanotubes (CNTs) [7,8]. These materials are characterised by high electron conductivity, a large specific surface area that facilitates microorganism adhesion and the adsorption of potentially toxic chemicals, and high corrosion resistance [9]. Additives with high conductivity have been found to stimulate Direct Interspecies Electron Transfer (DIET), a concept initially proposed by Kato et al. [10]. The authors demonstrated that DIET could be a new form of syntrophic association, where electrons are transferred directly from electron donors to electron acceptors, involving fewer biological enzymatic reactions [11,12]. It was confirmed that DIET had great potential for shortening the start-up period, promoting organic degradation and enhancing methane production in the AD process. In addition to carbon-based conductive additives, iron-based conductive materials have also been tested for stimulating DIET and methane formation. These include magnetite (Fe3O4), hematite (Fe2O3) and stainless steel [13,14]. Although these oxides differ in their chemical composition and electrical conductivity (magnetite has 106 times higher conductivity than hematite [15]), their DIET stimulation, in terms of lag time and methane generation rate, is similar. It is also worth noting that inorganic carbon- or iron-based conductive materials can be replaced by conductive polymers, e.g. polyaniline (PANI) nanorods [16]. In addition to the previously mentioned chemical additive applications, research also focuses on the impact of supplementing with enzymes and microorganisms. These biological additives are often considered as a form of pre-treatment for difficult-to-biodegrade substrates, typically performed outside the bioreactor [17,18]. The direct application of biological additives into the digester has been studied and utilised less frequently due to the challenging environmental conditions. The goal of inoculating with biological additives is to increase the diversity of microorganisms to enhance hydrolytic and/or methanogenic activity.
Given the growing trend of using additives in anaerobic digestion and the development and study of functional additive systems with various origins and properties, this work provides a comprehensive characterisation of these additives based on the latest research reports. Combining functional additives in AD requires a strategic approach to maximise their benefits and ensure the stability and efficiency of the digestion process. Thus, creating up-to-date literature reviews is valuable as it provides scientists and practitioners with inspiration for designing effective solutions and technologies.
2. Carbon–Based Conductive Materials
Enhancing the efficiency of anaerobic digestion (AD) through the use of conductive materials enables increased profitability of operations and optimisation of feedstock management to maximise its utilisation. Carbon-based conductive materials are becoming increasingly important. Notable examples include Granular Activated Carbon (GAC), Powdered Activated Carbon (PAC), biochar, Carbon Nanotubes (CNTs), as well as graphite and graphene. These materials vary in their chemical and physical properties, which lead to different process outcomes. Graphite is a natural mineral with conductive properties and a high adsorption surface area. It has several beneficial properties for the AD process [19] and is also used as a feedstock for producing graphene. In a study [20] observed that the addition of graphite to AD processes, where the feedstock were cattle manure by-products and food waste, resulted in increased methane production under mesophilic conditions. Graphene, on the other hand, is a material consisting of a single layer of carbon atoms. A comparative study [21] involving graphite, graphene, and graphite oxide found that the best fermentation results for wastewater sludge and food waste were achieved with the addition of graphite to the reaction mixture. This resulted in a 36.09% improvement in methane production and a 23.07% increase in organic degradation compared to the control sample.
2.1. GAC and PAC
Granular Activated Carbon (GAC) is a conductive material derived from the thermal processing of carbon-rich feedstocks (such as coconut shells and coal). During thermal processing, the material’s activity is increased by reducing the oxygen level in the furnace (higher oxygen levels reduce GAC activity) [22]. GAC features a porous structure and a large adsorption surface area (see Figure 1), which is crucial for the AD process. GAC performs several functions, including enhancing DIET, promoting the growth of Firmicutes microorganisms in AD, and reducing lag phase in the process.
In the study by Zhang et al. [23], GAC improved the fermentation of wastewater at 20°C and also induced an increase in Geobacter bacteria from 9% to 12%. Additionally, there was a 300% increase in biomass in the system. It was found that DIET and physiological changes in organisms led to favourable changes in the Chemical Oxygen Demand (COD) growth rate to 2000 mg COD/L/day, compared to 500 mg COD/L/day without GAC. It was observed that this change resulted from interspecies interactions. Another study [9] noted that GAC can also be beneficial under low temperature conditions. A method for wastewater treatment using anaerobic digestion at temperatures of 15°C, 25°C, and 35°C was proposed. It was found that the addition of GAC (6 g/L) reduced the lag phase from 15.1 days to 10.6 days (a 29.8% reduction) and increased methane production from 6.4 mL/day to 7.9 mL/day. Thus, GAC supplementation can significantly contribute to reducing the sludge decomposition time and enhancing methane production. Furthermore, modifying GAC with magnetite has been shown to reduce propionate. A experience [8] found that doping GAC with magnetite increased the syntrophic degradation of propionate, which is a compound that is difficult for microorganisms to break down. The highest methane production (220 mL/g) and COD of 70% were achieved in the bioreactor containing the modified GAC. In another study [24], thermally treated sludge and untreated hydrolysate were examined. It was observed that under three temperature conditions: psychrophilic (16°C–24°C), mesophilic (35°C), and thermophilic (55°C), the addition of GAC (10 g/L) resulted in a decrease in CH4 yield (ranging from 6.5% to 36.9%), as well as a reduction in the length of the methanogenic lag phase (19.3% to 30.6%), which accelerated the process and improved efficiency. It was noted that the acceleration of the process was associated with a reduction in phenolic compounds, which contributed to the overall enhancement of the process. Ultimately, the methane yield decreased (by 5.9%–8.1%) after the addition of GAC. It has been suggested to combine pre-treatment of the sludge with thermal processing to counteract the reduced methane yield in the AD process.
Powdered Activated Carbon (PAC) is another form of activated carbon. Both PAC and GAC can promote the syntrophic metabolism of alcohols and volatile fatty acids (VFAs) from brewery wastewater. However, PAC generally enhances process efficiency more effectively. The more abundant structure of mesopores and micropores in PAC provides better accessibility for microorganisms of the genus Methanosarcina in the system [25]. PAC is also more effective in accelerating the start of methanogenesis compared to GAC [26]. Literature data suggest that researchers have focused more on the granular form of activated carbon. The effects of GAC and PAC are summarised in Table 1, demonstrating that both materials can enhance AD efficiency.
2.2. Biochar
Biochar is a material obtained from a process waste feedstock, unlike activated carbon. Biochar is produced through the thermochemical conversion of biomass in the absence of air. The production of biochar is environmentally sustainable. Feedstock for biochar production can include vegetable waste, garden waste, and agricultural residues. Additionally, animal and household waste can also serve as feedstock [34]. The main differences between biochar and activated carbon, besides the source of feedstock, is its lower porosity. According to researchers [28], the conductivity of GAC is approximately 1000 times greater than that of biochar, which is significant for its impact on DIET. Furthermore, the production temperature of biochar is lower: activated carbon is produced at temperatures between 600°C and 900°C, while biochar is produced at temperatures below 600°C. The lower production temperature and use of waste feedstock make biochar a more environmentally friendly option. Figure 2 shows an SEM microscopic image of biochar.
Biochar is an additive that supports the growth of microorganisms. According to researchers [36], biochar can enrich the microbial community in AD processes in oil. In mesophilic systems, methane production increased by 32.5% with granular biochar. In thermophilic systems, a 13.3% increase was noted in digesters enriched with powdered biochar. Additionally, biochar was found to alleviate the accumulation of propionic acid, which positively impacts the AD process. There are also modifications of biochar that affect ammonia reduction. Research [37] on animal manure treated with biochar activated by acid reduced NH3 emissions by 37% to 51% over a month of storage. In contrast, non-activated biochar did not reduce emissions. Another study [38] compared biochar obtained through pyrolysis and hydrothermal carbonisation. It was found that biochar accelerates methane fermentation by mitigating mild ammonia concentrations (2.1 TAN kg-1; TAN – total ammonium nitrogen) for pyrolysed biochar. Biochar produced via hydrothermal carbonisation increased methane yield by 32%. Additionally, biochar addition helped alleviate the effects of ammonia at concentrations of 2450 mg/mL, stimulating process efficiency and reactor stability, particularly at a high OLR (6 g/VS L-d; VS – volatile solids, OLR – organic loading rate) [39]. Table 2 shows selected system parameters and effects of biochar.
2.3. SWNTs and MWNTs
Conductive nanomaterials are the subject of numerous studies aimed at enhancing the performance of the AD process. One such material is carbon nanotubes, which form spatial structures resembling cylindrical networks of hexagonal carbon rings. They represent an allotrope of carbon, similar to diamond and fullerene. Carbon nanotubes are categorised into single-walled nanotubes (SWNTs) and multiwalled nanotubes (MWNTs). SWNT consist of a single layer of carbon atoms, while MWNT are composed of multiple layers. These structures exhibit various properties that can be useful for AD, including promoting DIET, accelerating both abiotic and biotic reductions of electrophilic compounds, though they can also be toxic to microorganisms [44]. They are known for their high mechanical and thermal strength, as well as their large specific surface area [45]. It has been indicated [46] that carbon nanotubes improve organic loading rate and process stability, reduce ammonia levels, and support DIET. Research [47] on the effect of single-walled carbon nanotubes in wastewater sludge showed that adding 1000 mg/L did not impact the maximum methane yield but accelerated the process and increased electrical conductivity in the sludge, potentially enhancing DIET. Researchers Shen et al. [48], in studies conducted on a UASB (upflow anaerobic sludge blanket) reactor under thermophilic conditions with high acetate concentrations, found that the addition of 1000 mg/L SWNT had little impact on methanogenesis and acetate concentration but induced syntrophic acetate oxidation (SAO). This led to the growth of Coprothermobacter and Thermoacetogenium, which played key roles in the SAO pathway. Thermoacetogenium bacteria contributed to a potential DIET pathway, ultimately accelerating the process.
Research [49] on MWNTs indicates that they increase methane production by influencing specific trophic levels, such as methanogens and syntrophic bacteria. An experiment with anaerobic sludge and river sediment demonstrated increased methane yields for acetate, butyrate, ethanol, and hydrogen by factors of 2.6, 2.1, 1.2, and 1.1, respectively. Additionally, MWNTs doped with iron achieved significantly higher methane yields compared to those without iron. Another study [50] investigated various concentrations of nanotubes used in methane production from poultry manure under mesophilic conditions, with concentrations ranging from 0.5 to 6.5 g/L. The study found that the addition of 5 g/L of nanotubes had the most significant impact on methane production. Furthermore, nanotube structures influenced the biodegradation of short-chain fatty acids (SCFAs) such as butyrate, propionate, and acetate, resulting in a 15–16% increase in methane production with nanotube concentrations of 5–6.5 g/L. The conclusion drawn was that nanotube addition improves both the kinetics and productivity of the AD process in anaerobic reactors.
Figure 3 shows a diagram related to the most important functions of the above-described carbon-based conductive materials.
2.4. DIET Mechanism
Studies have shown that conductive carbon materials significantly accelerate and improve the efficiency of anaerobic digestion by intensifying direct interspecies electron transfer (DIET) [51]. DIET promotes the acetogenic methanogens metabolic pathway, one indicator of which is an increased concentration of acetic acid and a decreased concentration of propionic acid, with CO2 and CH4 being the final products, as described by the following equation [52]:
DIET pathway: CO2 + 8H+ + 8e− → CH4 + 2H2O
In DIET communities, electrons released from exoelectrogenic microorganisms are directly transferred to electron-capturing microorganisms via membrane-associated cytochromes and conductive pili that form electrical connections between syntrophic partners. The mechanism can also be based on the abiotic conductivity of materials within methanogens. Below (see Figure 4) is a diagram illustrating the DIET mechanism [53].
The DIET mechanism through conductive materials varies depending on the type of conductive material used. Conductive materials typically have a large specific surface area, to which microorganisms can attach, grow, and transfer electrons on their surface.
Regarding the action of carbon materials [11], found that the reason active carbon promotes DIET is due to its higher conductivity compared to methanogenic aggregates. However, the promotional effect of biochar is nearly the same as that of active carbon, even though biochar's conductivity is only one-thousandth that of activated carbon [54,55]. This suggests that any conductivity above a certain threshold is sufficient to trigger the DIET methanogenic pathway [56].
Currently, researchers are striving to discover various materials that mediate electron transfer involved in methanogenesis. Modern spectroscopy techniques provide crucial information on the composition and structural organisation of biofilms, which can be useful in elucidating the additional functions of this unique layer of microbial cells. However, there are many challenges such as the unclear biological mechanisms, influences of non-DIET mechanisms, limitations of organic matters syntrophically oxidised by way of DIET, and problems with practical application of DIET mediated by conductive materials [57]. When discussing the ambiguities surrounding the mechanism, the electron transfer biological pathway between microorganisms is considered not fully explained. It has not been discovered how electron-donating bacteria transfer electrons outside the cell. The most commonly used microbial model in studies of the mechanism is Geobacter because of its fully sequenced genome and advanced gene engineering methods.
3. Metal Oxide Nanomaterial
Analyses by Baniamerian et al. [58] show that metal oxide nanoparticles (NPs) are the second most commonly used type of nanomaterial in methane fermentation processes (metallic nanoparticles – 38%, metal oxide nanomaterials – 35%, carbon-based nanomaterials – 18%, nanocomposite materials – 9%). The relatively broad application of these metal oxides not only in the energy sector but also in many other industries is due to their unique physical, chemical, mechanical, and biological properties [59,60]. The advantages of metal oxide nanoparticles include small particle size, large surface area and density, stability, high tensile strength, and resistance to degradation by chemicals or microorganisms. They also exhibit high-temperature superconductivity, low cost, ease of use, and recovery post-fermentation. Additionally, these nanoparticles are non-toxic and available in various forms [58,61,62].
Years of research have shown that metal oxide nanoparticles used as a biological additive in methane fermentation increase the efficiency and effectiveness of the process, leading to the production of more biogas and biomethane (see Table 3). Nanoparticles stimulate energy production by shortening the lag phase [63,64], which is the adaptation period of microorganisms to new environmental conditions. According to Arya et al. [65] and Chen et al. [66], metal oxide nanoparticles can promote the synthesis of enzymes and cellular components essential for the proper growth and development of microorganisms, improving their ability to acclimate in the bioreactor and serving as a source of essential nutrients. Additionally, nano-TiO2 has demonstrated the ability to reduce hydrogen sulphide content in the fermentation environment by forming irreversible bonds with sulphur [67]. Metal oxide nanoparticles enrich the microbial community in the digester, leading to the growth and development of methanogenic bacteria (e.g. Methanobacterium sp., Methanobrevibacter sp., Methanothrix sp.) and hydrolytic microorganisms capable of degrading specific feedstock components (e.g. Chloroflexi sp., Bacteroidetes sp. – polysaccharide decomposition, Lysinibacillus sp., Ruminofilibacter sp. – cellulose decomposition). Nano-Al2O3 and nano-Fe2O3 also promote the action of coenzyme F420, which is indicative of methanogenic bacterial activity and is a key element in methane formation [64]. Furthermore, nanoparticles increase biogas and biomethane production by stimulating methanogen activity through inherent indirect electron transfer (IET) and previously discussed DIET between methanogens and fermentative bacteria [13,63,68], evidenced by the high efficiency of organic matter degradation [69]. Wang et al. [70] found that nanoparticles promote DIET in methanogenesis pathways using acetate and H2/CO2, and reduce the risk of reactor acidification. Li et al. [71] reported that nano-Fe3O4 (magnetite) stimulated DIET with increased acetic acid concentration and decreased propionic acid concentration. The stimulating effect of iron-based materials differs from that of conductive carbon materials, likely due to their size and structural differences. Magnetite nanoparticles are generally 20–50 nm in size, which is smaller than microbial cells, while conductive carbon materials are significantly larger and provide a substantial surface area for microbial attachment.
The proper selection of a metal oxide nanoparticle is crucial for the success of the methane fermentation process [59], as some metal ions exhibit strong toxic properties (see Table 3). Nanoparticles such as ZnO and CuO increase the amount of reactive oxygen species (ROS), which heightens the risk of oxidative stress [72]. This stress can damage cell membranes, cause leakage of cytoplasmic contents, inactivate enzymes, and disrupt DNA structure, thereby limiting the growth of methanogenic bacteria (e.g. Methanothrix sp.). This can lead to apoptosis of microorganisms and inhibition of bioenergy production [64,67]. Analyses conducted by Chen et al. [66] indicated that some metal oxides negatively affect the methane fermentation process during the hydrolysis stage of the feedstock. This is evidenced by a decrease in the degradation rate of soluble proteins and polysaccharides compared to the control sample. The authors found that zinc and copper cations inhibit the activity of protease and cellulase, enzymes responsible for the breakdown of proteins and cellulose present in the substrates. A low amount of soluble proteins and polysaccharides can slow the conversion of butyric acid to acetic acid, and subsequently acetic acid to methane. Furthermore, high concentrations of volatile fatty acids negatively impact the functioning of methanogenic bacteria [73,74].

Table 3.
Yield of biogas and biomethane in anaerobic digestion using metal oxide nanomaterials.
| Type of additive | Type of substrate | Yield of biogas/biomethane | References |
|---|---|---|---|
| Al2O3 | animal fat | increase in biogas production by 285% | [64] |
| Al2O3 | sewage sludge | increase in biogas production by 23.4% | [66] |
| Al2O3 | waste-activated sludge | increase in methane production by 14.8% | [69] |
| Fe2O3 | animal fat | increase in biogas production by 45.87% | [64] |
| Fe2O3 | granular sludge | increase in methane production by 38% | [7] |
| Fe2O3 | waste activated sludge | increase in methane production by 117% | [58] |
| Fe3O4 | corn straw and sewage sludge | increase in methane production by 60.47% | [71] |
| Fe3O4 | waste sludge | increase in methane yield by 58.7% | [75] |
| Fe3O4 | wastewater sludge | increase in biogas production by 96%increase in methane production by 144% | [68] |
| Fe3O4 | wastewater sludge | increase in biogas production by 107%increase in methane production by 153% | [68] |
| Fe3O4 | municipal solid waste | increase in methane yield by 72.09% | [76] |
| TiO2 | fresh dairy cattle manure | increase in methane yield by 121% | [67] |
| TiO2 | anaerobic sludge | increase in methane yield by 14.9% | [69] |
| CeO2 | waste-activated sludge | increase in methane production by 9.2% | [77] |
| MnO2 | seed sludge | decrease in methane production by 93% | [72] |
| MgO | waste activated sludge | decrease in methane production by 99% | [58] |
| CoO | sewage sludge | decrease in biogas production by 60% | [69] |
| CeO2 | sludge | decrease in biogas production by 35% | [58] |
| CeO2 | cellulose | decrease in biogas production by 100% | [58] |
| CuO | cattle manure | decrease in biogas production by 96% | [58] |
| CuO | sewage sludge | decrease in biogas production by 17.3% | [66] |
| ZnO | animal fat | decrease in biogas production by 17% | [64] |
| ZnO | municipal solid waste | decrease in biogas production by 15% | [78] |
| ZnO | waste-activated sludge | decrease in methane production by 50% | [73] |
| ZnO | sewage sludge | decrease in biogas production by 90.2% | [66] |
4. Trace Elements
Trace elements (TEs) are crucial additives in the process of anaerobic digestion to enhance the efficiency and stability of biogas production. Trace elements, also known as micronutrients, are required in small amounts but play significant roles in various metabolic pathways of the microorganisms involved in anaerobic digestion. The primary functions of TEs include: (i) supporting the metabolic activity of microorganisms, leading to increased biogas production and higher methane content, (ii) ensuring the proper functioning of enzymes and metabolic pathways, which helps maintain a stable biodegradation process, (iii) reducing inhibitory compounds, preventing the accumulation of inhibitors such as volatile fatty acids (VFAs) and ammonia, and (iv) preventing toxicity caused by excessive accumulation of certain metals, such as copper or zinc [79,80].
Trace metals such as cobalt (Co), nickel (Ni), selenium (Se), iron (Fe), molybdenum (Mo), and tungsten (W) are important cofactors necessary for the enzymes that catalyse methane production. Their addition to AD has been shown to stimulate and stabilise the biogas production process [81,82]. Combinations of trace elements can exhibit synergistic effects [83]. For example, the combination of elevated concentrations of Ni and Co can accelerate the initial exponential rate of the process, increase methanogen density, and thereby improve AD efficiency [83,84]. The supplementation of iron also plays a significant role in the AD process. A study on the feasibility of using iron-rich activated sludge as a stabilising agent for the AD of food waste confirmed the positive impact of Fe addition on AD stability [81]. In another study, Ketheesan et al. [85] illustrated the importance of Fe2+ bioavailability in controlling VFA concentration in batch reactors and organic shock loads in submerged anaerobic membrane bioreactor (SAMBR). The authors noted an increase in methane production from acetate and propionate compared to reactors without iron addition.
It is also important to note that different metals have different bio-uptake processes due to different kinetic and equilibrium processes. The presence of one metal can also influence the speciation and thus the bioavailability of another metal. For example, high concentrations of iron in the AD digester can promote co-precipitation, adsorption, and ion substitution of Co and Ni on FeS [86]. Therefore, to optimise the concentration of TEs (and maximise methanogenic activity), it is essential to understand how the speciation of TEs affects their bioavailability [87]. Due to the significance and effects of TE bioavailability in the microenvironment, Zhang et al. [88] developed an innovative approach to encapsulate Ni in dry water.
Trace elements play various roles in biochemical transformations and the AD process. Below are the key functions of trace elements in the process:
- Iron (Fe) plays many roles in anaerobic processes, mainly due to its exceptionally high reduction potential. Because of its properties, it plays a special role in energy metabolism. Iron is crucial for the activity of various enzymes, including hydrogenases and ferredoxins, which are involved in electron transfer and hydrogen metabolism [82,89]. In summary, this metal is utilised in the transport of methanogenic bacteria for the conversion of CO2 to CH4 and serves as both an electron acceptor and donor [5]. Adequate iron levels can enhance biogas production and stabilise the AD process.
- Nickel (Ni) is essential for the function of several enzymes, such as methyl-coenzyme M reductase, which is key in the final step of methanogenesis. Anaerobic bacteria are heavily dependent on nickel when carbon dioxide and hydrogen are the sole sources of energy. The nickel tetrapyrrole, coenzyme F430, is known to bind to methyl-S-CoM reductase, which catalyses methane formation from methyl-S-CoM in both acetoclastic and hydrogenotrophic methanogens [90]. This coenzyme is part of the methyl-coenzyme M reductase enzyme, which reduces methyl-coenzyme M to methane [84,91]. Nickel supplementation can improve methane production, especially in nickel-deficient substrates.
- Cobalt (Co) is a critical component of vitamin B12, which activates carboxypeptidase and is required for the metabolism of certain methanogens. Corrinoids, such as vitamin B12, containing a cobalt ion, bind to methyl-coenzyme M (CoM) reductase, which catalyses methane formation in both acetoclastic methanogens and hydrogenotrophic bacteria [84]. The enzyme carbon monoxide dehydrogenase (CODH) also utilises cobalt [92]. Therefore, cobalt can increase methane yield and improve the metabolic activities of acetoclastic methanogens.
- Molybdenum (Mo) is a cofactor for enzymes like formate dehydrogenase, which participates in the conversion of formate to carbon dioxide. The Mo enzyme is synthesised only when Mo is present in the growth medium [82]. This metal can inhibit sulfate-reducing bacteria, limiting the formation of sulphides. Molybdenum can also stimulate methane production from corn silage and municipal waste substrates [84,93]. In summary, molybdenum enhances formate decomposition, thereby supporting the entire AD process.
- Selenium and Tungsten (Se and W) are parts of several selenoproteins that play roles in protecting cells from oxidative damage and participating in redox reactions [82]. Selenium, like tungsten, is a component of the enzyme formate dehydrogenase (FDH), which catalyses formate production by propionate oxidizers. Certain methanogenic bacteria contain W and Mo enzymes for the same purpose [94]. Few studies have been conducted on the effects of Se and W on methanogenesis. One study conducted on a laboratory scale, with food industry waste, showed evidence of increased methane production under the influence of Se and W, and additionally in combination with Co [95].
- Zinc (Zn) is involved in enzyme function, stabilising protein structures, and regulating gene expression. Zinc is a part of enzymes such as formate dehydrogenase (FDH), superoxide dismutase (SODM), and hydrogenase [82,83]. Zn has been found in remarkably high concentrations (50–630 ppm) in 10 methanogenic bacteria [96]. This metal is necessary for maintaining microbial activity and diversity in AD.
- Copper (Cu) functions in redox reactions and electron transport. In general, the role of copper in methanogenesis is contradictorily perceived. It has rarely been studied, making it difficult to fully understand the role of Cu in biogas production. However, it has not been found to have a noticeable stimulating effect on biogas production [82]. It is important to note that while Cu is essential in small amounts, its excess can be toxic to microorganisms [86].
To ensure the effectiveness of trace elements, it is essential to follow principles of monitoring, supplementation, and balancing. Regular monitoring of trace element concentrations is crucial for maintaining optimal levels. The required concentration of trace metals during anaerobic digestion varies significantly depending on the type of substrate, process temperature (mesophilic or thermophilic), the mode of AD implementation (mono or co-digestion), and the type of methanogens [86,97]. These factors significantly differentiate the biochemical processes related to the dynamics of metals. Different substrates are characterised by varying metal contents, and the temperature of anaerobic systems can also determine different nutritional requirements. As identified in one study [93], the minimum requirements for Ni, Co, Zn, and Fe in thermophilic glucose fermentation are 0.40, 0.45, 2.0, and 3.5 mg/L, respectively. However, these values differ for mesophilic acetate fermentation or mesophilic AD of organic solid waste [93,97].
Proper supplementation is also crucial, aiming to correct deficiencies and improve performance. The form of supplementation (e.g. soluble salts, chelated forms) and dosing strategies are important factors to consider. While supplementation with trace elements enhances AD, as has been repeatedly emphasised, trace elements themselves do not biodegrade and accumulate in biomass, potentially causing inhibition of AD. Therefore, it is important to monitor and balance trace element levels.
5. Biological Additives
In recent years, increasing attention has been given to finding new and innovative solutions that not only enhance the efficiency of biogas and biomethane production but also increase the ecological value of the entire anaerobic digestion process. A promising solution to this problem is the use of biological additives in the form of microorganism cells and/or enzymes they produce (see Table 4), which do not pose a threat to the natural environment or human health [5,98,99]. Biological additives can be introduced directly into the bioreactor where anaerobic digestion is conducted, or they can be used as a pre-treatment method for the substrate preceding the discussed process [100,101].
5.1. Enzyme Supplementation
The role of commercial enzyme preparations and solutions containing extracellular enzymes isolated from microbial cultures, used as biological additives in anaerobic digestion, is to increase the efficiency of the first stage of this process. Hydrolytic metabolites are responsible for converting hard-to-degrade substrate components into soluble micro-molecules of simpler structure [102], thereby accelerating feedstock hydrolysis [103,104]. Enzymes create gaps in the substrate structure through which successive groups of hydrolytic metabolites migrate, thus increasing the surface area available for enzyme and microorganism activity [105,106,107]. This consequently allows more efficient use of the feedstock, accelerates the entire anaerobic digestion process, and increases the amount of produced biogas and biomethane.
Literature reports indicate that enzymatic supplementation is particularly important for lignocellulosic substrates, such as plant biomass, which is primarily composed of cellulose, hemicellulose, and lignin [100,108]. This highly complex lignocellulosic structure protects the plant cell wall from degradation, consequently slowing down feedstock decomposition and reducing energy generation efficiency [101,109]. Therefore, the hydrolytic enzymes most commonly used as biological additives in anaerobic digestion include lipases and peroxidases, which modify and hydrolyse lignin [110], as well as cellulases, which break β-1,4-glycosidic bonds between glucose molecules in cellulose [18,111,112]. What is more, amylases responsible for breaking down complex sugars and proteases that hydrolyse bonds in proteins, thereby releasing nitrogen molecules that are subsequently converted to ammonia, are often used [103,104].
The activity of enzymatic preparations is directly dependent on the reaction environment. Hence, a key challenge in supplementation is the proper preparation of the bioreactor, particularly maintaining the appropriate pH level and adding the necessary cofactors to achieve the desired enzymatic activity. Moreover, it is important to remember that anaerobic digestion occurs under anaerobic conditions, so metabolites used as biological additives must function effectively in an oxygen-free environment [104]. Another critical aspect is the analysis of the structure and composition of the substrates used [101,113], which will determine the selection of the appropriate enzyme [110,112,114], as well as its concentration, as evidenced by studies conducted by Bhatnagar et al. [115]. Their research indicates that both excessively high and low enzyme concentrations inhibit the anaerobic digestion process, resulting in the production of less biogas and biomethane compared to the control sample. Due to the highly complex structure of feedstock, Liu et al. [104], Speda et al. [111], and Weide et al. [114] suggest that an effective solution is to use preparations composed of various enzyme groups, which will simultaneously break down multiple substrate components.
Enzyme supplementation is a promising method for accelerating substrate hydrolysis because enzymes have a relatively broad tolerance to varying environmental conditions and high resistance to stress factors that may occur in the digester [98]. Compared to microbial cells, these metabolites are smaller, more soluble, and more mobile, which allows enzymes easier access to the substrate structure and more efficient hydrolysis of its components [5,99].
5.2. Bioaugmentation
Bioaugmentation involves introducing additional monocultures of microorganisms or consortia with specific properties into the digester [99,104], thereby modifying the bioreactor's microflora and restoring balance that may have been disrupted by stress conditions [98], particularly in the early stages of the process [114]. The main goal of this method is to enhance and select specific groups of microorganisms to stimulate their hydrolytic and methanogenic activity [5], which ultimately leads to increased biogas and biomethane production.
The groups of microorganisms most commonly used for bioaugmentation include hydrolytic bacteria and fungi [116,117,118]. These microorganisms modify the structure of lignin and hemicellulose through the production of enzymes and secondary metabolites, reduce the crystallinity of cellulose, thereby increasing substrate porosity and breaking down its components [101,119,120]. Another significant group comprises typical methanogenic bacteria (methanogens) responsible for carrying out anaerobic digestion and producing biogas and biomethane [121,122]. Recent literature indicates that an effective method is the use of microbial consortia consisting of different fungal species [123,124] or bacterial species [125,126], as well as consortia containing both bacteria and fungi [99,127,128]. Vinzelj et al. [129] suggest that an innovative solution involves utilising symbiotic relationships between anaerobic fungi, which produce hydrogen and formate, and methanobacteria, which convert these compounds into methane [130,131]. Furthermore, Swift et al. [132] report that methanogenic bacteria positively interact with hydrolytic enzymes produced by fungi, thereby promoting their activity.
A study by Li et al. [121] indicates that bioaugmentation using a consortium of methanogens stimulated signalling mechanisms and the activity of genes responsible for cell motility, stabilised the pH level in the bioreactor, and reduced the concentration of volatile fatty acids [123]. Tian et al. [133] demonstrated that bioaugmentation led to a reduction in instant hydrogen partial pressure, thereby creating favourable conditions for acetate oxidation and fatty acid degradation. Lebiocka et al. [125] showed that bioaugmentation using a consortium of various bacterial species increased the efficiency of volatile solids removal despite a reduced hydraulic retention time. Furthermore, the aforementioned authors found that the addition of new groups of microorganisms accelerated the metabolic rate and stimulated microbial growth and activity, thus leading to an increase in the efficiency of the methane fermentation process.
Just like with enzymatic preparations, the functioning of microorganisms and their ability to exhibit desired properties are directly dependent on the environment in which they reside. Therefore, a key task is to select suitable bacterial and/or fungal species that can withstand stress conditions (such as excessive ammonia concentrations [133,134], inappropriate temperature and pH), ensuring that they properly grow and develop in the fermentation bioreactor [5]. Another important aspect is the correct concentration of the introduced inoculum [135], as well as the type, composition, and moisture level of the feedstock used [101,119]. Furthermore, Romero-Güiza et al. [5] and Paritosh et al. [98] reported that innovative bioaugmentation approaches suggest that microorganisms should be introduced into the digester in the form of cultures immobilised on suitable carriers (e.g. microcapsules), indicating that the properties of the auxiliary material also play a role in the selection of biological additives.
Table 4.
Yield of biogas and biomethane in anaerobic digestion using biological additives.
| Type of additive | Type of substrate | Yield of biogas/biomethane | References |
|---|---|---|---|
| cellulases, xylanases, β-glucosidases |
ensiled forage ley | increase in methane production by 19% | [111] |
| lipase from Aspergillus | animal fat | increase in methane production by 80.8% | [110] |
| arachis oil | increase in methane production by 26.9% | [110] | |
| floatable grease | increase in methane production by 37% | [110] | |
| lipase from Candida | animal fat | increase in methane production by 157.7% | [110] |
| arachis oil | increase in methane production by 53.8% | [110] | |
| floatable grease | increase in methane production by 40.7% | [110] | |
| bio-additive Digest P3 (carbohydrases, pectinase, xylanase) |
poultry litter | increase in biogas production by 59.7% increase in methane production by 91.4% |
[136] |
| bio-additive APD (Aerobacter, Pseudomonas, Alcaligenes, cellulase, lipase) |
igniscum silage | increase in biogas production by 6% decrease in methane production by 7% |
[18] |
| maize silage | increase in biogas production by 53% increase in methane production by 74% |
[18] | |
| bio-additive PPT (Pseudomonas, Flavobacterium, Lactobacillus, cellulase, lipase) |
igniscum silage | increase in biogas production by 16% increase in methane production by 26% |
[18] |
| maize silage | increase in biogas production by 62% increase in methane production by 79% |
[18] | |
| bio-additive HAP (Clostridium, Micrococcus, cellulase, lipase) |
igniscum silage | increase in biogas production by 12% increase in methane production by 30% |
[18] |
| maize silage | increase in biogas production by 32% increase in methane production by 46% |
[18] | |
| bio-additive JENOR (Pichia, Trichoderma, cellulase, lipase) |
igniscum silage | increase in biogas production by 13% increase in methane production by 16% |
[18] |
| maize silage | increase in biogas production by 17% increase in methane production by 26% |
[18] | |
| Orpinomyces sp. | barley, triticale, rye, wheat, cow manure |
increase in methane production by 33% | [117] |
| Ochrobactrum sp. | sewage sludge | increase in biogas production by 22.06% | [137] |
| Caldicellulosiruptor bescii | birch wood chips | increase in methane production by 44% | [120] |
| Clostridium thermocellum | wheat straw, cow manure |
increase in methane production by 39% | [116] |
|
Clostridium cellulolyticumClostridium cellulovorans Clostridium aceticum Mesotoga infera Methanosarcina barkeri Methanosaeta concilii |
Axonopus compressus | increase in methane production by 20.7% | [126] |
|
Neocallimastix sp. Orpinomyces sp. fermentative bacteria |
wheat straw | increase in methane production of 290% | [127] |
| mushroom spent straw | increase in methane production by 330% | [127] | |
|
Aspergillus sp. Trichoderma viride |
maize straw | increase in methane production by 31.7% | [128] |
| Trichoderma atroviride | water hyacinth | increase in biogas production by 65% increase in methane production by 117% |
[118] |
| Trichoderma reesei | rice straw and soybean straw |
increase in biogas production by 318% increase in methane production by 807% |
[138] |
|
Orpinomyces sp. Piromyces sp. Anaeromyces sp. Neocallimastix frontalis |
algal biomass | increase in methane production by 40.6% | [123] |
| cow manure | increase in methane production by 60% | [124] |
6. AD Additives Research – Summary
This chapter presents a summary of the most commonly used additives. The discussion will start with Granulated Activated Carbon (GAC). This substance has often been used as a conductive material for DIET stimulation so far, as its preparation and application in the reactor are not very problematic, while its properties are typical of carbon-based materials. It is interesting to note that the conductivity of GAC is approximately 1,000 times higher than that of biocarbon [13]. In their study on the AD process of wastewater, Park et al. [9], proved that thanks to GAC supplementation it was possible to shorten the delay time by 29.8% and increase the methane production rate by 23.4% at a low temperature of 25°C. So far, additive systems introduced into anaerobic bioreactors have been extremely rarely analysed. Barua et al. [8], were the first to study GAC doped with magnetite, through the use of which the scientist achieved the highest methane production and COD removal (70%) in fed-batch tests. Powdered Activated Carbon (PAC), similarly to GAC, facilitates electron transfer between microorganisms by acting as the electrical bridge and enhances both hydrogenotrophic and aceticlastic pathways. However, in the case of that compound, a more favourable effect is noticed at the hydrolysis stage compared to GAC: effective acceleration of volatile fatty acids (VFAs) consumption and thereby alleviation of acidification and lag phase. The stimulation of hydrolysis by PAC was also demonstrated by Yan et al. [31]. The obtained results revealed that genes coding for hydrolytic enzymes and xenobiotics metabolism were highly expressed in the presence of PAC.
Another additive selected for this study was Carbon Nanotubes (CNTs), recognisable as cylindrical carbon nanostructures consisting of a hexagonal mesh. They are characterised by high electrical conductivity and high mechanical strength and thermal conductivity. They also stimulate DIET to form methane, although they are toxic to microorganisms. Ambuchi et al. [7] investigated the role of multi-wall carbon nanotubes (MWCNTs) in enhancing the AD performance of granular activated sludge. It was demonstrated that MWCNTs improved the kinetics and effectiveness of biogas generation. The carbon-based conductive materials described above can be replaced by conductive polymers.
Literature confirm also that supplementation with Polyaniline Nanorods (PANI), from the semi-flexible rod polymer family, increases methane formation in an anaerobic reactor. Zhou et al. [16] used conductive polyaniline hydrogel (three-dimensional polyaniline) in their experiment. The porous structure and hydrophilic surface of the PANI hydrogel promoted the adhesion of the anaerobes. Accelerated production of CH4 in the anaerobic degradation of organics in wastewater was reported, as the result of the DIET effect. Nanostructured materials, a product of nanotechnology, also have a significant position as effective additives in the AD process [139]. In that category, additional materials for methane fermentation are metal oxide NPs, such as ZnO, CuO, TiO2, MgO, NiO, Fe2O3 etc., while those selected for this project, due to their high conductivity, are: CuO, MgO and Fe3O4. Hamed et al. [140] presented the functions and effects of individual metal oxide NPs in AD. Thus, CuO brings good results in the fermentation of cattle manure and municipal waste activated sludge, MgO – waste activated sludge and Fe3O4 – raw manure and municipal solid waste. The authors of the research carried out so far have obtained various results, not always confirming an increase in CH4 production under the influence of oxides but giving good results in the shortening of the lag phase [64].
Trace Elements (TEs), are another type of additive with a significant impact on kinetics and efficiency. Trace elements are key cofactors in many enzymatic reactions of the metabolic methane production pathway. Fe, Ni and Co are the most often studied NS, since they are essential cofactors of carbon monoxide dehydrogenase, acetyl-CoA decarbonylase, metreductase and other enzymes involved in the acetoclastic methanogenesis pathway [5]. Furthermore, those metals have also proved to be essential for the acetotrophic pathway of methanogenesis, which is currently gaining importance. For example, Pobeheim et al. [84], noted in their work that Ni and Co deficiency has a negative impact on the stability of the process (i.e. VFAs accumulation).
Supplementation of enzymes and microorganisms, known as biological additives, is also widely studied as an alternative to the physicochemical pretreatment of substrates before AD, while their direct introduction into fermentation chambers has received less attention. In this project, a strain of Trichoderma sp. fungi and an enzyme preparation were selected as biological additives. The main characteristics of fungi of the Trichoderma genus that influence their key use in the anaerobic digestion process are their very strong cellulolytic and hemicellulolytic properties, which allow them to accelerate the hydrolysis of the polysaccharides that make up the cell wall of plant waste biomass. Despite the abundant supply of lignocellulosic waste biomass on the Earth, it is used in AD relatively rarely [17]. When it comes to the use of an enzyme preparation, Fugol et al. [18], in their study with the use of different lignocellulosic substrates, introduced four commercial vaccines containing different bacteria species or a yeast and mould mixture into the reactor along with the inoculum and reported improved biogas production. In turn, Noyola and Tinajero [141] after using the lyophilized bacilli system with a solution of micronutrients managed to increase the production of methane dramatically, which confirms the validity of using appropriate combinations of additives. The presented summary constitutes the basis for formulating reasons why research in the field of additives for anaerobic digestion should be continued in a multi-faceted manner using advanced techniques.
7. Feasibility and Perspectives
The incorporation of an increasingly diverse range of waste materials with varying chemical compositions and origins into the anaerobic digestion process often leads to technical challenges, negatively affecting the kinetics and efficiency of the process [1,142]. These challenges primarily include poor biodegradation of the fermented material due to inhibition, low microbial activity, high activation energy, and other factors. As a result, the optimal extraction of chemical (primary) energy from the substrates is hindered [143]. These difficulties are often addressed by using effective yet largely unrecognised additives in the AD process. Literature on such additives is frequently speculative, underscoring the need for continued research and expansion of knowledge.
The previously cited reports suggest that attempts to combine additives have yielded very promising results, confirming that this approach is promising for improving the kinetics and effectiveness of AD. Successfully integrating functional additives into AD requires a comprehensive strategy that takes into account the interactions between additives, optimises their application through controlled dosing and real-time monitoring, and ensures both economic and regulatory compliance. Understanding synergistic and antagonistic effects is crucial for developing additive systems. Some additives may enhance each other's effects, leading to improved efficiency. For instance, adding both trace elements and specific enzymes can result in more effective breakdown and conversion of organic matter. Conversely, other additives may interfere with each other, reducing effectiveness or even inhibiting the process. To avoid such conflicts, careful selection and dosing are essential. Thus, by strategically integrating additives, AD systems can achieve higher efficiency, stability, and biogas production, contributing to more sustainable and effective waste management and energy generation.
To sum up, additives have the potential to significantly enhance anaerobic digestion by improving biogas yield, process stability, and the quality of digestate. However, their use must be carefully managed to balance performance benefits with economic and environmental considerations. Additives must also comply with regulatory standards for environmental safety and public health. It is crucial to ensure that additives do not introduce harmful contaminants into the digestate, which could affect its suitability as a fertiliser. Continued research and technological advancements are expected to expand the range of effective and safe additives available for AD.
8. Conclusions
The supplementation of various additives, including those discussed in this paper — conductive carbon materials, metal oxide nanomaterials, trace metals, and biological additives — in anaerobic digesters is currently a topic of significant interest. The primary objective of research and attempts to implement additive-based technologies is to increase biogas production, while secondary goals include reducing process inhibition and foaming, and improving the rheology of the digestate. Most studies have been conducted in laboratory-scale batch reactors, with less emphasis on continuous reactor operations, which partly limits the ability to model and extrapolate results to industrial scales.
The results presented largely confirm the positive impact of the tested additives, particularly conductive carbon and iron-based materials, as well as trace metals and selected metal oxide nanomaterials. Inorganic additives are the subject of numerous research studies. Trace elements, as cofactors in various enzymatic reactions, are often studied individually and in combinations, yielding promising results. Notably, iron has been frequently reported as a promising additive due to its low cost. However, not all types of nanomaterial oxides have produced the expected results — some, such as CuO and ZnO, are toxic to microorganisms and thus unsuitable for use. Bioaugmentation, on the other hand, has been the focus of more research than enzymatic supplementation with hydrolytic properties. The results clearly indicate that bioaugmentation can be a useful technique for improving AD performance, though its industrial application remains limited, mainly due to economic factors and the relatively small number of studies conducted so far.
Overall, research on additives focuses on understanding their mechanisms of action, optimising dosages, and application methods. Advanced analytical techniques, such as metagenomics and proteomics, are employed to study microbial communities and their interactions with additives. The development of new additives is driven by environmental and economic considerations, sustainability, and cost-effectiveness. These trends reflect a holistic approach to improving AD processes by enhancing microbial activity, substrate availability, and process stability, ultimately leading to higher biogas yields and more efficient waste processing.
Author Contributions: Conceptualization, A.A.P.; software, T.K. and A.K.; formal analysis, A.A.P., A.W.-M. and J.D.; resources, A.A.P., K.P., T.K. and A.K.; writing—A.A.P., K.P., T.K. and A.K. ; writing—review and editing, A.A.P., A.W-M., and A.N.; visualization, T.K.; supervision, A.A.P., A.W.-M., A.N. and J.D. ; project administration, A.A.P.; funding acquisition, A.A.P. and K.P. All authors have read and agreed to the published version of the manuscript.
Informed Consent Statement: Not applicable.
Data Availability Statement: No new data were created or analysed in this study. Data sharing is not applicable to this article.
Conflicts of Interest: The authors declare no conflicts of interest.
References
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
- Pilarska, A.A.; Wolna-Maruwka, A.; Pilarski, K.; Janczak, D.; Przybył, K.; Gawrysiak-Witulska, M. The Use of Lignin as a Microbial Carrier in the Co-Digestion of Cheese and Wafer Waste. Polymers 2019, 11, 2073. [Google Scholar] [CrossRef] [PubMed]
- Amani, T.; Nosrati, M.; Sreekrishnan, T.R. Anaerobic digestion from the viewpoint of microbiological, chemical, and operational aspects - A review. Environ. Rev. 2010, 18, 255–278. [Google Scholar] [CrossRef]
- Gadirli, G.; Pilarska, A.A.; Dach, J.; Pilarski, K.; Kolasa-Więcek, A.; Borowiak, K. Fundamentals, Operation and Global Prospects for the Development of Biogas Plants - A Review. Energies 2024, 17, 568. [Google Scholar] [CrossRef]
- Pramanik, S.K.; Suja, F.B.; Zain, S.M.; Pramanik, B.K. The anaerobic digestion process of biogas production from food waste: Prospects and constraints. Bioresour. Technol. Rep. 2019, 8, 100310. [Google Scholar] [CrossRef]
- Romero-Güiza, M.S.; Vila, J.; Mata-Alvarez, J.; Chimenos, J.M.; Astals, S. The role of additives on anaerobic digestion: A review. Renew. Sustain. Energy Rev. 2016, 58, 1486–1499. [Google Scholar] [CrossRef]
- Leca, E.; Zennaro, B.; Hamelin, J.; Carrère, H.; Sambusiti, C. Use of additives to improve collective biogas plant performances: A comprehensive review. Biotechnol. Adv. 2023, 65, 108129. [Google Scholar] [CrossRef] [PubMed]
- Ambuchi, J.J.; Zhang, Z.; Feng, Y. Biogas Enhancement Using Iron Oxide Nanoparticles and Multi-Wall Carbon Nanotubes. Int. J. Chem. Biomol. Eng. 2016, 10, 1305–1311. [Google Scholar]
- Barua, S.; Zakaria, B.S.; Lin, L.; Dhar, B.R. Magnetite Doped Granular Activated Carbon as an Additive for High-Performance Anaerobic Digestion. Mater. Sci. Energy Technol. 2019, 2, 377–384. [Google Scholar] [CrossRef]
- Park, J.H.; Park, J.H.; Lee, S.H.; Jung, S.P.; Kim, S.H. Enhancing anaerobic digestion for rural wastewater treatment with granular activated carbon (GAC) supplementation. Bioresour. Technol. 2020, 315, 123890. [Google Scholar] [CrossRef]
- Kato, S.; Hashimoto, K.; Watanabe, K. Methanogenesis facilitated by electric syntrophy via (semi) conductive iron-oxide minerals. Environ. Microbiol. 2012, 14, 1646–1654. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Q.; Sun, D.; Wang, Y.; Dong, H.; Dang, Y.; Holmes, D.E. Applying potentials to conductive materials impairs High-loading anaerobic digestion performance by affecting direct interspecies electron transfer. Bioresour. Technol. 2020, 297, 122422. [Google Scholar] [CrossRef]
- Chen, L.; Fang, W.; Chang, J.; Liang, J.; Zhang, P.; Zhang, G. Improvement of Direct Interspecies Electron Transfer via Adding Conductive Materials in Anaerobic Digestion: Mechanisms, Performances, and Challenges. Front. Microbiol. 2022, 13, 860749. [Google Scholar] [CrossRef]
- Park, J.H.; Kang, H.J.; Park, K.H.; Park, H.D. Direct interspecies electron transfer via conductive materials: A perspective for anaerobic digestion applications. Bioresour. Technol. 2018, 254, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Pilarska, A.A.; Wolna-Maruwka, A.; Niewiadomska, A.; Grządziel, J.; Gałązka, A.; Paluch, E.; Borowiak, K.; Pilarski, K. Quantitative and Qualitative Changes in the Genetic Diversity of Bacterial Communities in Anaerobic Bioreactors with the Diatomaceous Earth/Peat Cell Carrier. Cells 2022, 11, 2571. [Google Scholar] [CrossRef] [PubMed]
- Guskos, N.; Papadopoulos, G.J.; Likodimos, V.; Patapis, S.; Yarmis, D.; Przepiera, A.; Przepiera, K.; Majszczyk, J.; Typek, J.; Wabia, M.; Aidinis, K.; Drazek, Z. Photoacoustic, EPR and electrical conductivity investigations of three synthetic mineral pigments: Hematite, goethite and magnetite. Mater. Res. Bull. 2002, 37, 1051–1061. [Google Scholar] [CrossRef]
- Zhou, N.; Wang, T.; Chen, S.; Hu, Q.; Cheng, X.; Sun, D.; Vupputuri, S.; Qiu, B.; Liu, H.; Guo, Z. Conductive Polyaniline Hydrogel Enhanced Methane Production from Anaerobic Wastewater Treatment. J. Colloid Interface Sci. 2021, 581, 314–322. [Google Scholar] [CrossRef]
- Mutschlechner, M.; Illmer, P.; Wagner, A.O. Biological pre-treatment: Enhancing biogas production using the highly cellulolytic fungus Trichoderma viride. Waste Manag. 2015, 43, 98–107. [Google Scholar] [CrossRef]
- Fugol, M.; Prask, H.; Szlachta, J.; Dyjakon, A.; Pasławska, M.; Szufa, S. Improving the Energetic Efficiency of Biogas Plants Using Enzymatic Additives to Anaerobic Digestion. Energies 2023, 16, 1845. [Google Scholar] [CrossRef]
- Hou, D.; Li, K.; Ma, R.; Liu, Q. Influence of order degree of coaly graphite on its structure change during preparation of graphene oxide. Journal of Materiomics, 2020, 6, 628–641. [Google Scholar] [CrossRef]
- Muratçobanoğlu, H.; Gökçek, Ö.B.; Mert, R.A.; Zan, R.; Demirel, S. Simultaneous Synergistic Effects of Graphite Addition and Co-Digestion of Food Waste and Cow Manure: Biogas Production and Microbial Community. Bioresour. Technol. 2020, 309, 123365. [Google Scholar] [CrossRef]
- Wang, P.; Zheng, Y.; Lin, P.; Li, J.; Dong, H.; Yu, H.; Qi, L.; Ren, L. Effects of graphite, graphene, and graphene oxide on the anaerobic co-digestion of sewage sludge and food waste: Attention to methane production and the fate of antibiotic resistance genes. Bioresour. Technol. 2021, 339, 125585. [Google Scholar] [CrossRef] [PubMed]
- Burachevskaya, M.; Mandzhieva, S.; Bauer, T.; Minkina, T.; Rajput, V.; Chaplygin, V.; Fedorenko, A.; Chernikova, N.; Zamulina, I.; Kolesnikov, S.; Sushkova, S.; Perelomov, L. The Effect of Granular Activated Carbon and Biochar on the Availability of Cu and Zn to Hordeum sativum Distichum in Contaminated Soil. Plants 2021, 10, 841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, L.; Guo, B.; Zhou, Y.; Gao, M.; Sharaf, A.; Liu, Y. Granular activated carbon stimulated microbial physiological changes for enhanced anaerobic digestion of municipal sewage. Chem. Eng. J. 2020, 400, 125838. [Google Scholar] [CrossRef]
- Jiang, Q.; Liu, H.; Zhang, Y.; Cui, M.H.; Fu, B.; Liu, H.B. Insight into sludge anaerobic digestion with granular activated carbon addition: Methanogenic acceleration and methane reduction relief. Bioresour. Technol. 2021, 319, 124131. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; He, C.; Luo, L.; Lü, F.; He, P.; Cui, L. Comparing activated carbon of different particle sizes on enhancing methane generation in upflow anaerobic digester. Bioresour. Technol. 2015, 196, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wei, H.; Su, Y.; Gu, W.; Wang, B.; Xie, B. Powdered activated carbon facilitates methane productivity of anaerobic co-digestion via acidification alleviating: Microbial and metabolic insights. Bioresour. Technol. 2020, 313, 123706. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Holmes, D.E.; Zhao, Z.; Woodard, T.L.; Zhang, Y.; Sun, D.; Wang, L.Y.; Nevin, K.P.; Lovley, D.R. Enhancing anaerobic digestion of complex organic waste with carbon-based conductive materials. Bioresour. Technol. 2016, 220, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Park, J.H.; Seong, H.; Sul, W.J.; Jin, K.H.; Park, H.D. Metagenomic insight into methanogenic reactors promoting direct interspecies electron transfer via granular activated carbon. Bioresour. Technol. 2018, 259, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, R.; Wang, H.; Yang, K. Direct interspecies electron transfer stimulated by granular activated carbon enhances anaerobic methanation efficiency from typical kitchen waste lipid-rapeseed oil. Sci. Total Environ. 2020, 704, 135282. [Google Scholar] [CrossRef]
- Patrícya Florentino, A.; Sharaf, A.; Zhang, L.; Liu, Y. Overcoming Ammonia Inhibition in Anaerobic Blackwater Treatment with Granular Activated Carbon: The Role of Electroactive Microorganisms. Environ. Sci. Water Res. Technol. 2019, 5, 383–396. [Google Scholar] [CrossRef]
- Yan, W.; Zhang, L.; Wijaya, S.M.; Zhou, Y. Unveiling the role of activated carbon on hydrolysis process in anaerobic digestion. Bioresour. Technol. 2020, 296, 122366. [Google Scholar] [CrossRef] [PubMed]
- Cuetos, M.J.; Martinez, E.J.; Moreno, R.; Gonzalez, R.; Otero, M.; Gomez, X. Enhancing anaerobic digestion of poultry blood using activated carbon. J. Adv. Res. 2017, 8, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Dastyar, W.; Mirsoleimani Azizi, S.M.; Meshref, M.N.A.; Dhar, B.R. Powdered activated carbon amendment in percolate tank enhances high-solids anaerobic digestion of organic fraction of municipal solid waste. Process. Saf. Environ. Prot. 2021, 151, 63–70. [Google Scholar] [CrossRef]
- Poluszyńska, J.; Ślęzak, E.; Wieczorek, P.P. Biowęgiel jako środek polepszający właściwości gleby. Przemysł Chemiczny, 2019, 1, 100–107. [Google Scholar] [CrossRef]
- Hassan, S.; Ngo, T.; Khudur, L.S.; Krohn, C.; Dike, C.C.; Hakeem, I.G.; Shah, K.; Surapaneni, A.; Ball, A.S. Biosolids-Derived Biochar Improves Biomethane Production in the Anaerobic Digestion of Chicken Manure. Resources 2023, 12, 123. [Google Scholar] [CrossRef]
- Lü, F.; Liu, Y.; Shao, L.; He, P. Powdered biochar doubled microbial growth in anaerobic digestion of oil. Appl. Energy 2019, 247, 605–614. [Google Scholar] [CrossRef]
- Baral, K.R.; McIlroy, J.; Lyons, G.; Johnston, C. The Effect of Biochar and Acid Activated Biochar on Ammonia Emissions during Manure Storage. Environ. Pollut. 2023, 317, 120815. [Google Scholar] [CrossRef]
- Mumme, J.; Srocke, F.; Heeg, K.; Werner, M. Use of biochars in anaerobic digestion. Bioresour. Technol. 2014, 164, 189–197. [Google Scholar] [CrossRef]
- Giwa, A.S.; Xu, H.; Chang, F.; Wu, J.; Li, Y.; Ali, N.; Ding, S.; Wang, K. Effect of biochar on reactor performance and methane generation during the anaerobic digestion of food waste treatment at long-run operations. J. Environ. Chem. Eng. 2019, 7, 103067. [Google Scholar] [CrossRef]
- Luo, C.; Lü, F.; Shao, L.; He, P. Application of eco-compatible biochar in anaerobic digestion to relieve acid stress and promote the selective colonization of functional microbes. Water Res. 2015, 68, 710–718. [Google Scholar] [CrossRef]
- Lim, E.Y.; Tian, H.; Chen, Y.; Ni, K.; Zhang, J.; Tong, Y.W. Methanogenic pathway and microbial succession during start-up and stabilization of thermophilic food waste anaerobic digestion with biochar. Bioresour. Technol. 2020, 314, 123751. [Google Scholar] [CrossRef] [PubMed]
- Gómez, X.; Meredith, W.; Fernández, C.; Sánchez-García, M.; Díez-Antolínez, R.; Garzón-Santos, J.; Snape, C.E. Evaluating the effect of biochar addition on the anaerobic digestion of swine manure: Application of Py-GC/MS. Environ. Sci. Pollut. Res. 2018, 25, 25600–25611. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, Q.; Yuwen, C.; Gong, K.; Sheng, L.; Li, Y.; Xing, Y.; Chen, R. Biochar Triggers Methanogenesis Recovery of a Severely Acidified Anaerobic Digestion System via Hydrogen-Based Syntrophic Pathway Inhibition. Int. J. Hydrogen Energy 2021, 46, 9666–9677. [Google Scholar] [CrossRef]
- Salvador, A.F.; Martins, G.; Melle-Franco, M.; Serpa, R.; Stams, A.J.M.; Cavaleiro, A.J.; Pereira, M.A.; Alves, M.M. Carbon nanotubes accelerate methane production in pure cultures of methanogens and in a syntrophic coculture. Environ. Microbiol. 2017, 19, 2727–2739. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Rahimian Koloor, S.S.; Alshehri, A.H.; Arockiarajan, A. Carbon Nanotube Characteristics and Enhancement Effects on the Mechanical Features of Polymer-Based Materials and Structures - A Review. J. Mater. Res. Technol. 2023, 24, 6495–6521. [Google Scholar] [CrossRef]
- Jeyakumar, R.B.; Vincent, G.S. Recent Advances and Perspectives of Nanotechnology in Anaerobic Digestion: A New Paradigm towards Sludge Biodegradability. Sustainability 2022, 14, 7191. [Google Scholar] [CrossRef]
- Li, L.L.; Tong, Z.H.; Fang, C.Y.; Chu, J.; Yu, H.Q. Response of anaerobic granular sludge to single-wall carbon nanotube exposure. Water Res. 2015, 70, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Liang, Z.; Chen, Y.; Song, H.; Wan, J. Enhancement of syntrophic acetate oxidation pathway via single walled carbon nanotubes addition under high acetate concentration and thermophilic condition. Bioresour. Technol. 2020, 306, 123182. [Google Scholar] [CrossRef] [PubMed]
- Cavaleiro, A.J.; Salvador, A.F.; Martins, G.; Oliveira, C.C.; Liu, Y.; Martins, V.R.; Castro, A.R.; Soares, O.S.G.P.; Pereira, M.F.R.; Pereira, L.; Langenhoff, A.A.M.; Pereira, M.A.; Alves, M.M. Multi-Walled Carbon Nanotubes Enhance Methanogenesis from Diverse Organic Compounds in Anaerobic Sludge and River Sediments. Appl. Sci. 2020, 10, 8184. [Google Scholar] [CrossRef]
- Ziganshina, E.E.; Bulynina, S.S.; Ziganshin, A.M. Anaerobic Digestion of Chicken Manure Assisted by Carbon Nanotubes: Promotion of Volatile Fatty Acids Consumption and Methane Production. Fermentation 2022, 8, 641. [Google Scholar] [CrossRef]
- Li, L.; Xu, Y.; Dai, X.; Dai, L. Principles and advancements in improving anaerobic digestion of organic waste via direct interspecies electron transfer. Renew. Sustain. Energy Rev 2021, 148, 111367. [Google Scholar] [CrossRef]
- Madondo, N.I.; Rathilal, S.; Bakare, B.F.; Tetteh, E.K. Application of Bioelectrochemical Systems and Anaerobic Additives in Wastewater Treatment: A Conceptual Review. Int. J. Mol. Sci. 2023, 24, 4753. [Google Scholar] [CrossRef] [PubMed]
- Kong, T.; Zhang, W. Enhanced Anaerobic Digestion Using Conductive Materials through Mediation of Direct Microbial Interspecies Electron Transfer: A Review. Fermentation 2023, 9, 884. [Google Scholar] [CrossRef]
- Chen, S.; Rotaru, A.E.; Liu, F.; Philips, J.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Carbon cloth stimulates direct interspecies electron transfer in syntrophic co-cultures. Bioresour. Technol. 2014, 173, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Li, S.; Cai, J.; He, P.; Lü, F. Ability of biochar to facilitate anaerobic digestion is restricted to stressed surroundings. J. Clean. Prod. 2019, 238, 117959. [Google Scholar] [CrossRef]
- Barua, S.; Zakaria, B.S.; Dhar, B.R. Enhanced methanogenic co-degradation of propionate and butyrate by anaerobic microbiome enriched on conductive carbon fibers. Bioresour. Technol. 2018, 266, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Storck, T.; Virdis, B.; Batstone, D.J. Modelling extracellular limitations for mediated versus direct interspecies electron transfer. The ISME Journal. 2016, 10, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Baniamerian, H.; Isfahani, P.G.; Tsapekos, P.; Alvarado-Morales, M.; Shahrokhi, M.; Vossoughi, M.; Angelidaki, I. Application of Nano-Structured Materials in Anaerobic Digestion: Current Status and Perspectives. Chemosphere 2019, 229, 188–199. [Google Scholar] [CrossRef]
- Mu, H.; Chen, Y.; Xiao, N. Effects of metal oxide nanoparticles (TiO2, Al2O3, SiO2 and ZnO) on waste activated sludge anaerobic digestion. Bioresour. Technol. 2011, 102, 10305–10311. [Google Scholar] [CrossRef]
- Kökdemir Ünşar, E.; Perendeci, N.A. What Kind of Effects Do Fe2O3 and Al2O3 Nanoparticles Have on Anaerobic Digestion, Inhibition or Enhancement? Chemosphere 2018, 211, 726–735. [Google Scholar] [CrossRef]
- Stankic, S.; Suman, S.; Haque, F.; Vidic, J. Pure and Multi Metal Oxide Nanoparticles: Synthesis, Antibacterial and Cytotoxic Properties. J. Nanobiotechnology 2016, 14, 73. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.E.; Mohammad, A.; Ali, W.; Sharwani, A.A. Recent Developments in Properties and Applications of Metal Oxides. In Inorganic Anticorrosive Materials; Verma, E., Aslam, J., Hussain, C.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 95–111. [Google Scholar]
- Ren, S.; Usman, M.; Tsang, D.C.W.; O-Thong, S.; Angelidaki, I.; Zhu, X.; Zhang, S.; Luo, G. Hydrochar-facilitated anaerobic digestion: evidence for direct interspecies electron transfer mediated through surface oxygen-containing functional groups. Environ. Sci. Technol. 2020, 54, 5755–5766. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Pu, H.; Yang, Z. Study on the effect of different additives on the anaerobic digestion of hybrid Pennisetum: Comparison of nano-ZnO, nano-Fe2O3 and nano-Al2O3. Heliyon 2023, 9, e16313. [Google Scholar] [CrossRef] [PubMed]
- Arya, I.; Poona, A.; Dikshit, P.K.; Pandit, S.; Kumar, J.; Singh, H.N.; Jha, N.K.; Rudayni, H.A.; Chaudhary, A.A.; Kumar, S. Current Trends and Future Prospects of Nanotechnology in Biofuel Production. Catalysts 2021, 11, 1308. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, Z.; Zhang, Y.; Xiang, Y.; Xu, R.; Jia, M.; Cao, J.; Xiong, W. Effects of different conductive nanomaterials on anaerobic digestion process and microbial community of sludge. Bioresour. Technol. 2020, 304, 123016. [Google Scholar] [CrossRef]
- Farghali, M.; Andriamanohiarisoamanana, F.J.; Ahmed, M.M.; Kotb, S.; Yamashiro, T.; Iwasaki, M.; Umetsu, K. Impacts of iron oxide and titanium dioxide nanoparticles on biogas production: Hydrogen sulfide mitigation, process stability, and prospective challenges. J. Environ. Manag. 2019, 240, 160–167. [Google Scholar] [CrossRef]
- Heikal, G.; Shakroum, M.; Vranayova, Z.; Abdo, A. Impact of Nanoparticles on Biogas Production from Anaerobic Digestion of Sewage Sludge. J. Ecol. Eng. 2022, 23, 222–240. [Google Scholar] [CrossRef]
- Ajay, C.M.; Mohan, S.; Dinesha, P.; Rosen, M.A. Review of impact of nanoparticle additives on anaerobic digestion and methane generation. Fuel 2020, 277, 118234. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, D.; Dai, L.; Dong, B.; Dai, X. Magnetite triggering enhanced direct interspecies electron transfer: a scavenger for the blockage of electron transfer in anaerobic digestion of high-solids sewage sludge. Environ. Sci. Technol. 2018, 52, 7160–7169. [Google Scholar] [CrossRef]
- Li, P.; Wang, Q.; He, X.; Yu, R.; He, C.; Shen, D.; Jiao, Y. Investigation on the effect of different additives on anaerobic co-digestion of corn straw and sewage sludge: Comparison of biochar, Fe3O4, and magnetic biochar. Bioresour. Technol. 2022, 345, 126532. [Google Scholar] [CrossRef]
- Tian, T.; Qiao, S.; Yu, C.; Zhou, J. Effects of nano-sized MnO2 on methanogenic propionate and butyrate degradation in anaerobic digestion. J. Hazard. Mater. 2019, 364, 11–18. [Google Scholar] [CrossRef]
- Zhang, L.; He, X.; Zhang, Z.; Cang, D.; Nwe, K.A.; Zheng, L.; Li, Z.; Cheng, S. Evaluating the Influences of ZnO Engineering Nanomaterials on VFA Accumulation in Sludge Anaerobic Digestion. Biochem. Eng. J. 2017, 125, 206–211. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, Z.; Tian, L.; Zhang, L.; Cheng, S.; Li, Z.; Cang, D. Mechanistic investigation of toxicological change in ZnO and TiO2 multi-nanomaterial systems during anaerobic digestion and the microorganism response. Biochem. Eng. J. 2019, 147, 62–71. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, L.; Wang, Y.; Zhao, Y.; She, Z.; Gao, M.; Guo, Y. Application of iron oxide (Fe3O4) nanoparticles during the two-stage anaerobic digestion with waste sludge: Impact on the biogas production and the substrate metabolism. Renewable Energy 2020, 146, 2724–2735. [Google Scholar] [CrossRef]
- Ali, A.; Mahar, R.B.; Soomro, R.A.; Sherazi, S.T.H. Fe3O4 nanoparticles facilitated anaerobic digestion of organic fraction of municipal solid waste for enhancement of methane production. Energy Source Part A 2017, 39, 1815–1822. [Google Scholar] [CrossRef]
- Ünşar, E.K.; Çığgın, A.S.; Erdem, A.; Perendeci, N.A. Long- and Short-Term Impacts of CuO, Ag and CeO2 Nanoparticles on Anaerobic Digestion of Municipal Waste Activated Sludge. Environ. Sci.: Processes Impacts 2016, 18, 277–288. [Google Scholar]
- Temizel, I.; Emadian, S.M.; Di Addario, M.; Onay, T.T.; Demirel, B.; Copty, N.K.; Karanfil, T. Effect of nano-ZnO on biogas generation from simulated landfills. Waste Manage 2017, 63, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, B.; Li, A.; Zhang, L.; Li, R.; Yang, T.; Xing, W. Mechanism of process imbalance of long-term anaerobic digestion of food waste and role of trace elements in maintaining anaerobic process stability. Bioresour. Technol. 2019, 275, 172–182. [Google Scholar] [CrossRef]
- Molaey, R.; Bayrakdar, A.; Sürmeli, R.Ö.; Çalli, B. Influence of trace element supplementation on anaerobic digestion of chicken manure: Linking process stability to methanogenic population dynamics. J. Clean. Prod. 2018, 181, 794–800. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, L.; Li, A. Enhanced anaerobic digestion of food waste by trace metal elements supplementation and reduced metals dosage by green chelating agent [S, S]-EDDS via improving metals bioavailability. Water Res. 2015, 2015 84, 266–277. [Google Scholar] [CrossRef]
- Nakhate, S.P.; Gulhane, M.; Singh, A.K.; Purohit, H.J.; Shah, M.P.; Khardenavis, A.A. Trace metals as key controlling switches regulating the efficiencies of aerobic and anaerobic bioprocesses. Biochem. Eng. J. 2023, 198, 108999. [Google Scholar] [CrossRef]
- Linville, J.L.; Shen, Y.; Schoene, R.P.; Nguyen, M.; Urgun-Demirtas, M.; Snyder, S.W. Impact of trace element additives on anaerobic digestion of sewage sludge with in-situ carbon dioxide sequestration. Process Biochem. 2016, 51, 1283–1289. [Google Scholar] [CrossRef]
- Pobeheim, H.; Munk, B.; Lindorfer, H.; Guebitz, G.M. Impact of nickel and cobalt on biogas production and process stability during semi-continuous anaerobic fermentation of a model substrate for maize silage. Water Res. 2011, 45, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Ketheesan, B.; Thanh, P.M.; Stuckey, D.C. Iron deficiency and bioavailability in anaerobic batch and submerged membrane bioreactors (SAMBR) during organic shock loads. Bioresour. Technol. 2016, 211, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Myszograj, S.; Stadnik, A.; Płuciennik-Koropczuk, E. The influence of trace elements on anaerobic digestion process. Civ. Environ. Eng. Rep. 2018, 28, 105–115. [Google Scholar] [CrossRef]
- Gustavsson, J.; Shakeri Yekta, S.; Sundberg, C.; Karlsson, A.; Ejlertsson, J.; Skyllberg, U.; Svensson, B.H. Bioavailability of cobalt and nickel during anaerobic digestion of sulfur-rich stillage for biogas formation. Appl. Energy 2013, 112, 473–477. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, J.; Loh, K.C. Enhanced food waste anaerobic digestion: An encapsulated metal additive for shear stress-based controlled release. J. Clean. Prod. 2019, 235, 85–95. [Google Scholar] [CrossRef]
- Vintiloiu, A.; Boxriker, M.; Lemmer, A.; Oechsner, H.; Jungbluth, T.; Mathies, E.; Ramhold, D. Effect of ethylenediaminetetraacetic acid (EDTA) on the bioavailability of trace elements during anaerobic digestion. Chem. Eng. J. 2013, 223, 436–441. [Google Scholar] [CrossRef]
- Friedmann, H.C.; Klein, A.; Thauer, R.K. Structure and function of the nickel porphinoid, coenzyme F430, and of its enzyme, methyl coenzyme M reductase. FEMS Microbiol. Lett. 1990, 87, 339–348. [Google Scholar] [CrossRef]
- Gustavsson, J.; Yekta, S.S.; Karlsson, A.; Skyllberg, U.; Bo, H.S. Potential bioavailability and chemical forms of Co and Ni in the biogas process - An evaluation based on sequential and acid volatile sulfide extractions. Eng. Life Sci. 2013, 13, 572–579. [Google Scholar] [CrossRef]
- Murakami, E.; Ragsdale, S.W. Evidence for Intersubunit Communication during Acetyl-Coa Cleavage by the Multienzyme Co Dehydrogenase/Acetyl-Coa Synthase Complex from Methanosarcina thermophila Evidence That the Beta Subunit Catalyzes C-C and C-S Bond Cleavage. J. Biol. Chem. 2000, 275, 4699–4707. [Google Scholar] [CrossRef]
- Uemura, S. Mineral requirements for mesophilic and thermophilic anaerobic digestion of organic solid waste. Int. J. Environ. Res. 2010, 4, 33–40. [Google Scholar]
- Yu, B.; Zhang, D.; Shan, A.; Lou, Z.; Yuan, H.; Huang, X.; Yuan, W.; Dai, X.; Zhu, N. Methane-rich biogas production from waste-activated sludge with the addition of ferric chloride under a thermophilic anaerobic digestion system. RSC Adv. 2015, 5, 38538–38546. [Google Scholar] [CrossRef]
- Feng, X.M.; Karlsson, A.; Svensson, B.H.; Bertilsson, S. Impact of trace element addition on biogas production from food industrial waste ^ linking process to microbial communities. FEMS Microbiol. Ecol. 2010, 74, 226–240. [Google Scholar] [CrossRef]
- Scherer, P.; Lippert, H.; Wolff, G. Composition of the Major Elements and Trace Elements of 10 Methanogenic Bacteria Determined by Inductively Coupled Plasma Emission Spectrometry. Biol. Trace Elem. Res. 1983, 5, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Takashima, M.; Shimada, K.; Speece, R.E. Minimum Requirements for Trace Metals (Iron, Nickel, Cobalt, and Zinc) in Thermophilic and Mesophilic Methane Fermentation from Glucose. Water Environ. Res. 2011, 83, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Paritosh, K.; Yadav, M.; Chawade, A.; Sahoo, D.; Kesharwani, N.; Pareek, N.; Vivekanand, V. Additives as a Support Structure for Specific Biochemical Activity Boosts in Anaerobic Digestion: A Review. Front. Energy Res. 2020, 8, 88. [Google Scholar] [CrossRef]
- Li, X.; Wang, Z.; He, Y.; Wang, Y.; Wang, S.; Zheng, Z.; Wang, S.; Xu, J.; Cai, Y.; Ying, H. A Comprehensive Review of the Strategies to Improve Anaerobic Digestion: Their Mechanism and Digestion Performance. Methane 2024, 3, 227–256. [Google Scholar] [CrossRef]
- Ferdeș, M.; Dincă, M.N.; Moiceanu, G.; Zăbavă, B.Ș.; Paraschiv, G. Microorganisms and Enzymes Used in the Biological Pretreatment of the Substrate to Enhance Biogas Production: A Review. Sustainability 2020, 12, 7205. [Google Scholar] [CrossRef]
- Kubiak, A.; Pilarska, A.A.; Wolna-Maruwka, A.; Niewiadomska, A.; Panasiewicz, K. The Use of Fungi of the Trichoderma Genus in Anaerobic Digestion: A Review. Int. J. Mol. Sci. 2023, 24, 17576. [Google Scholar] [CrossRef]
- Kim, M.; Li, D.; Choi, O.; Sang, B.I.; Chiang, P.C.; Kim, H. Effects of supplement additives on anaerobic biogas production. Korean J. Chem. Eng. 2017, 34, 2678–2685. [Google Scholar] [CrossRef]
- Ye, M.; Liu, J.; Ma, C.; Li, Y.Y.; Zou, L.; Qian, G.; Xu, Z.P. Improving the stability and efficiency of anaerobic digestion of food waste using additives: A critical review. J. Clean. Prod. 2018, 192, 316–326. [Google Scholar] [CrossRef]
- Liu, M.; Wei, Y.; Leng, X. Improving Biogas Production Using Additives in Anaerobic Digestion: A Review. J. Clean. Prod. 2021, 297, 126666. [Google Scholar] [CrossRef]
- Rajendran, K.; Drielak, E.; Varma, V.S.; Muthusamy, S.; Kumar, G. Updates on the Pretreatment of Lignocellulosic Feedstocks for Bioenergy Production—A Review. Biomass Conv. Bioref. 2018, 8, 471–483. [Google Scholar] [CrossRef]
- Kainthola, J.; Podder, A.; Fechner, M.; Goel, R. An Overview of Fungal Pretreatment Processes for Anaerobic Digestion: Applications, Bottlenecks and Future Needs. Bioresour. Technol. 2021, 321, 124397. [Google Scholar] [CrossRef] [PubMed]
- Kovács, E.; Szűcs, C.; Farkas, A.; Szuhaj, M.; Maróti, G.; Bagi, Z.; Rákhely, G.; Kovács, K.L. Pretreatment of Lignocellulosic Biogas Substrates by Filamentous Fungi. J. Biotechnol. 2022, 360, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.M.; Wright, M.M. Anaerobic digestion fundamentals, challenges, and technological advances. Phys. Sci. Rev. 2023, 8, 2819–2837. [Google Scholar] [CrossRef]
- Manyi-Loh, C.E.; Lues, R. Anaerobic Digestion of Lignocellulosic Biomass: Substrate Characteristics (Challenge) and Innovation. Fermentation 2023, 9, 755. [Google Scholar] [CrossRef]
- Meng, Y.; Luan, F.; Yuan, H.; Chen, X.; Li, X. Enhancing anaerobic digestion performance of crude lipid in food waste by enzymatic pretreatment. Bioresour. Technol. 2017, 224, 48–55. [Google Scholar] [CrossRef]
- Speda, J.; Johansson, M.A.; Odnell, A.; Karlsson, M. Enhanced Biomethane Production Rate and Yield from Lignocellulosic Ensiled Forage Ley by in Situ Anaerobic Digestion Treatment with Endogenous Cellulolytic Enzymes. Biotechnol. Biofuels 2017, 10, 129. [Google Scholar] [CrossRef]
- Wang, S.; Li, F.; Wu, D.; Zhang, P.; Wang, H.; Tao, X.; Ye, J.; Nabi, M. Enzyme pretreatment enhancing biogas yield from corn stover: Feasibility, optimization, and mechanism analysis. J. Agric. Food Chem. 2018, 66, 10026–10032. [Google Scholar] [CrossRef] [PubMed]
- Herrero Garcia, N.; Benedetti, M.; Bolzonella, D. Effects of enzymes addition on biogas production from anaerobic digestion of agricultural biomasses. Waste Biomass Valor. 2019, 10, 3711–3722. [Google Scholar] [CrossRef]
- Weide, T.; Baquero, C.D.; Schomaker, M.; Brügging, E.; Wetter, C. Effects of enzyme addition on biogas and methane yields in the batch anaerobic digestion of agricultural waste (silage, straw, and animal manure). Biomass Bioenergy 2020, 132, 105442. [Google Scholar] [CrossRef]
- Bhatnagar, R.; Ryan, D.; Murphy, R.; Enright, A.M. Effect of Co-digestion Ratio and Enzyme Treatment on Biogas Production from Grass Silage and Chicken Litter. Waste Biomass Valor 2019, 10, 3271–3277. [Google Scholar] [CrossRef]
- Öner, B.E.; Akyol, Ç.; Bozan, M.; Ince, O.; Aydin, S.; Ince, B. Bioaugmentation with Clostridium thermocellum to enhance the anaerobic biodegradation of lignocellulosic agricultural residues. Bioresour. Technol. 2018, 249, 620–625. [Google Scholar] [CrossRef]
- Akyol, Ç.; Ince, O.; Bozana, M.; Ozbayramb, E.G.; Ince, B. Fungal bioaugmentation of anaerobic digesters fed with lignocellulosic biomass: What to expect from anaerobic fungus Orpinomyces sp. Bioresour. Technol. 2019, 277, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ilo, O.P.; Nkomo, S.L.; Mkhize, N.M.; Mutanga, O.; Simatele, M.D. The effects of Trichoderma atroviride pretreatment on the biogas production from anaerobic digestion of water hyacinth. Energy Environ. 2024, 35, 1339–1358. [Google Scholar] [CrossRef]
- Ali, S.S.; Mustafa, A.M.; Kornaros, M.; Manni, A.; Sun, J.; Khalil, M.A. Construction of novel microbial consortia CS-5 and BC-4 valued for the degradation of catalpa sawdust and chlorophenols simultaneously with enhancing methane production. Bioresour. Technol. 2020, 301, 122720. [Google Scholar] [CrossRef]
- Mulat, D.G.; Huerta, S.G.; Kalyani, D.; Horn, S.J. Enhancing methane production from lignocellulosic biomass by combined steam-explosion pretreatment and bioaugmentation with cellulolytic bacterium Caldicellulosiruptor bescii. Biotechnol. Biofuels 2018, 11, 19. [Google Scholar] [CrossRef]
- Li, Y.; Yang, G.; Li, L.; Sun, Y. Bioaugmentation for overloaded anaerobic digestion recovery with acid-tolerant methanogenic enrichment. Waste Manag. 2018, 79, 744–751. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Sun, Y.; Yuan, Z. Bioaugmentation Strategy for Enhancing Anaerobic Digestion of High C/N Ratio Feedstock with Methanogenic Enrichment Culture. Bioresour. Technol. 2018, 261, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Yildirim, E.; Ince, O.; Ince, B. Rumen anaerobic Fungi create new opportunities for enhanced methane production from microalgae biomass. Algal Res. 2017, 23, 150–160. [Google Scholar] [CrossRef]
- Yildirim, E.; Ince, O.; Aydin, S.; Ince, B. Improvement of biogas potential of anaerobic digesters using rumen fungi. Renew. Energy 2017, 109, 346–353. [Google Scholar] [CrossRef]
- Lebiocka, M.; Montusiewicz, A.; Cydzik-Kwiatkowska, A. Effect of Bioaugmentation on Biogas Yields and Kinetics in Anaerobic Digestion of Sewage Sludge. Int. J. Environ. Res. Public Health 2018, 15, 1717. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.E.; Wang, Q.; Lim, E.Y.; Liu, Z.; He, J.; Tong, Y.W. Optimization of Bioaugmentation of the Anaerobic Digestion of Axonopus Compressus Cowgrass for the Production of Biomethane. J. Clean. Prod. 2020, 258, 120932. [Google Scholar] [CrossRef]
- Ferraro, A.; Dottorini, G.; Massini, G.; Mazzurco Miritana, V.; Signorini, A.; Lembo, G.; Fabbricino, M. Combined bioaugmentation with anaerobic ruminal fungi and fermentative bacteria to enhance biogas production from wheat straw and mushroom spent straw. Bioresour. Technol. 2018, 260, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zheng, Z.; Cai, Y.; Zhao, Y.; Zhang, Y.; Gao, Y.; Cui, Z.; Wang, X. Accelerated biomethane production from lignocellulosic biomass: Pretreated by mixed enzymes secreted by Trichoderma viride and Aspergillus sp. Bioresour. Technol. 2020, 309, 123378. [Google Scholar] [CrossRef]
- Vinzelj, J.; Joshi, A.; Insam, H.; Podmirseg, S.M. Employing anaerobic fungi in biogas production: Challenges & opportunities. Bioresour. Technol. 2020, 300, 122687. [Google Scholar]
- Wilken, S.E.; Saxena, M.; Petzold, L.R.; O’Malley, M.A. In Silico Identification of Microbial Partners to Form Consortia with Anaerobic Fungi. Processes 2018, 6, 7. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Jin, W.; Sharpton, T.J.; Mackie, R.I.; Cann, I.; Cheng, Y.; Zhu, W. Combined genomic, transcriptomic, proteomic, and physiological characterization of the growth of Pecoramyces sp. F1 in monoculture and co-culture with a syntrophic methanogen. Front. Microbiol. 2019, 10, 435. [Google Scholar] [CrossRef]
- Swift, C.L.; Brown, J.L.; Seppälä, S.; O’Malley, M.A. Co-cultivation of the anaerobic fungus Anaeromyces robustus with Methanobacterium bryantii enhances transcription of carbohydrate active enzymes. J. Ind. Microbiol. Biotechnol. 2019, 46, 1427–1433. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Yan, M.; Treu, L.; Angelidaki, I.; Fotidis, I.A. Hydrogenotrophic methanogens are the key for a successful bioaugmentation to alleviate ammonia inhibition in thermophilic anaerobic digesters. Bioresour. Technol. 2019, 293, 122070. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, W.; Liu, C.; Zhang, R.; Liu, G. Mitigation of ammonia inhibition through bioaugmentation with different microorganisms during anaerobic digestion: Selection of strains and reactor performance evaluation. Water Res. 2019, 155, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Undiandeye, J.; Gallegos, D.; Lenz, J.; Nelles, M.; Stinner, W. Effect of Novel Aspergillus and Neurospora Species-Based Additive on Ensiling Parameters and Biomethane Potential of Sugar Beet Leaves. Appl. Sci. 2022, 12, 2684. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Ryan, D.; Murphy, R.; Enright, A.M. Trace Element Supplementation and Enzyme Addition to Enhance Biogas Production by Anaerobic Digestion of Chicken Litter. Energies 2020, 13, 3477. [Google Scholar] [CrossRef]
- Poszytek, K.; Karczewska-Golec, J.; Ciok, A.; Decewicz, P.; Dziurzynski, M.; Gorecki, A.; Jakusz, G.; Krucon, T.; Lomza, P.; Romaniuk, K.; Styczynski, M.; Yang, Z.; Drewniak, L.; Dziewit, L. Genome-Guided Characterization of Ochrobactrum sp. POC9 Enhancing Sewage Sludge Utilization - Biotechnological Potential and Biosafety Considerations. Int. J. Environ. Res. Public Health 2018, 15, 1501. [Google Scholar] [CrossRef]
- Deng, Y.; Dai, B.; Xu, J.; Liu, X.; Xu, J. Anaerobic co-digestion of rice straw and soybean straw to increase biogas production by pretreatment with Trichoderma reesei RUT C30. Environ. Prog. Sustain. Energy 2018, 37, 1050–1057. [Google Scholar] [CrossRef]
- Fernández-García, M.; Rodriguez, J.A. Metal Oxide Nanoparticles. In Encyclopedia of Inorganic Chemistry; John Wiley & Sons.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Hamed, S.M.; Hassan, S.H.; Selim, S.; Kumar, A.; Khalaf, S.M.H.; Wadaan, M.A.M.; Hozzein, W.N.; AbdElgawad, H. Physiological and biochemical responses to aluminum-induced oxidative stress in two cyanobacterial species. Environ. Pollut. 2019, 251, 961–969. [Google Scholar] [CrossRef]
- Noyola, A.; Tinajero, A. Effect of biological additives and micronutrients on the anaerobic digestion of physicochemical sludge. Water Sci Technol, 2005, 52, 275–281. [Google Scholar] [CrossRef]
- Pilarska, A.A.; Kulupa, T.; Kubiak, A.; Wolna-Maruwka, A.; Pilarski, K.; Niewiadomska, A. Anaerobic Digestion of Food Waste - A Short Review. Energies 2023, 16, 5742. [Google Scholar] [CrossRef]
- Pilarski, K.; Pilarska, A.A.; Kolasa-Więcek, A.; Suszanowicz, D. An Agricultural Biogas Plant as a Thermodynamic System: A Study of Efficiency in the Transformation from Primary to Secondary Energy. Energies 2023, 16, 7398. [Google Scholar] [CrossRef]
Figure 1.
SEM microscopic image of GAC microstructure [22].
Figure 1.
SEM microscopic image of GAC microstructure [22].

Figure 2.
SEM microscopic image of biochar microstructure [35].
Figure 2.
SEM microscopic image of biochar microstructure [35].

Figure 3.
Functions of selected carbon-based materials in AD process (authors’ own scheme).
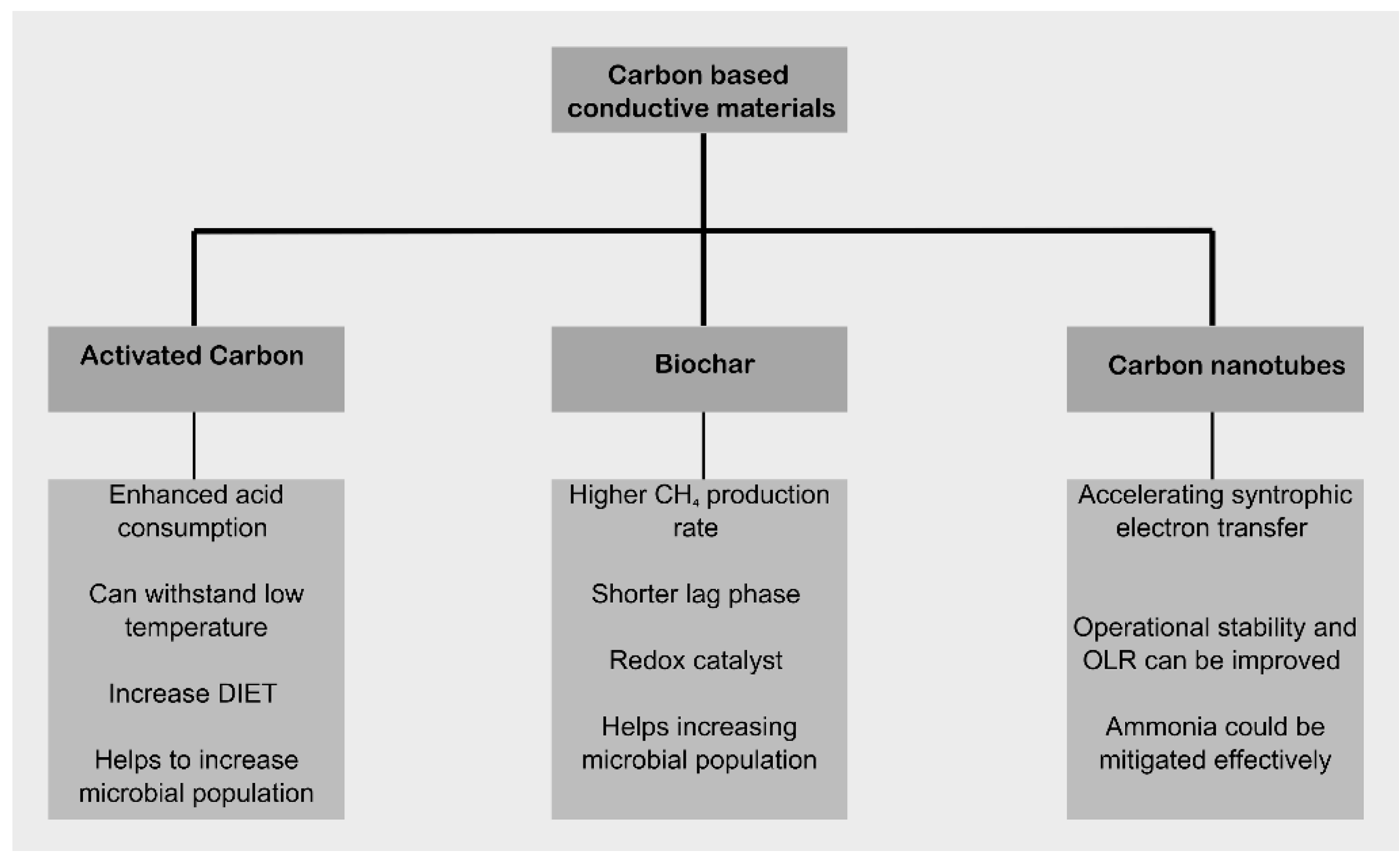
Figure 4.
DIET via conductive materials, based on [53].
Figure 4.
DIET via conductive materials, based on [53].
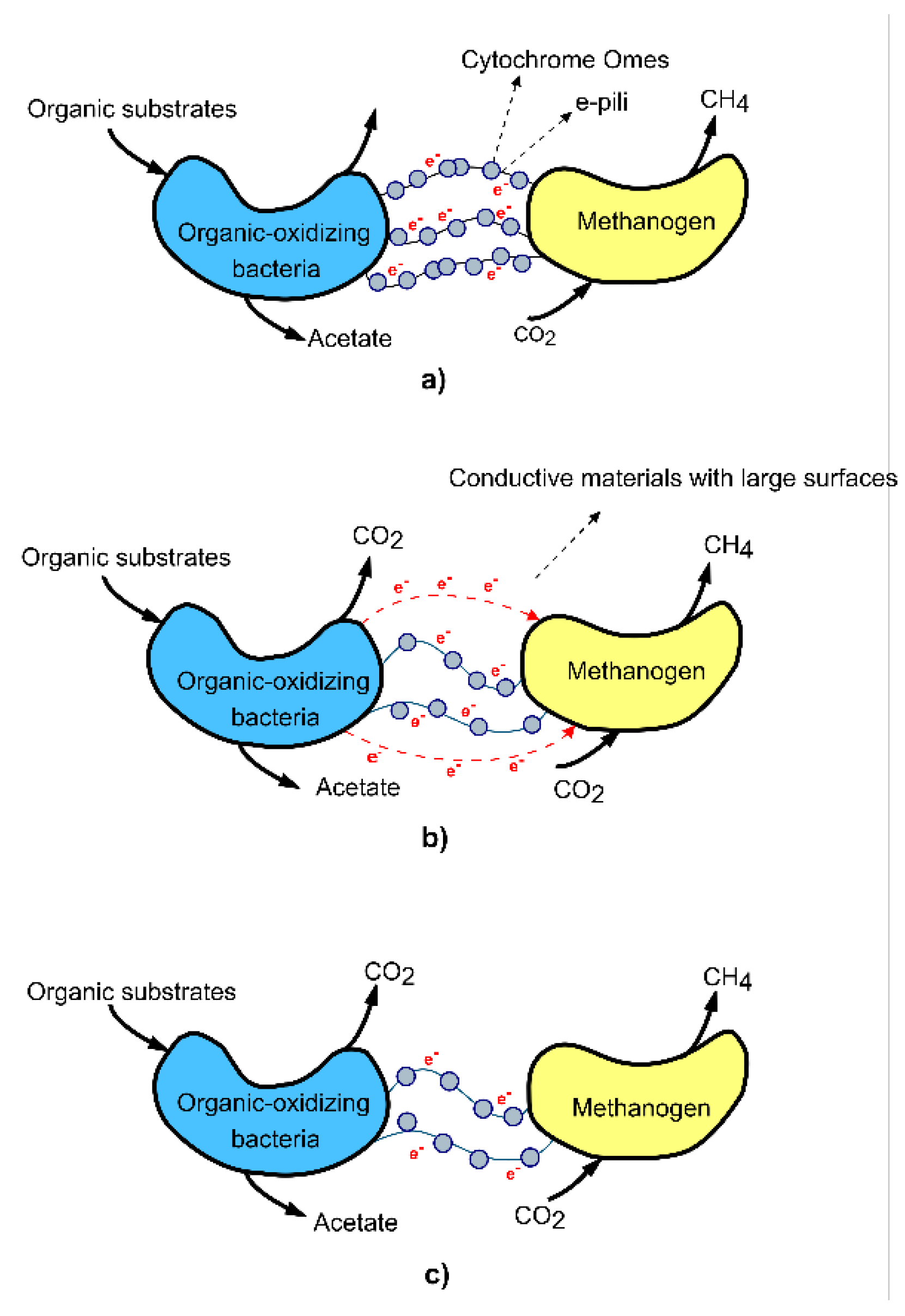
Table 1.
Dosage and effects of GAC and PAC in the AD process.
| Type of additive | Dosage (g/L) | Type of substrate | Effect | References |
|---|---|---|---|---|
| GAC | 50.0 | dog food | enables the process to proceed at high OLR values and improves process efficiency | [27] |
| GAC | 6.0 | acetic acid and ethanol |
increase in methane yield by 31% | [28] |
| GAC | – | rapeseed oil | increase in methane yield | [29] |
| GAC | 33.3 | black water (urine and feces) |
increase in methane yield by 57% | [30] |
| PAC | 5.0 10.0 |
food waste, vegetable waste |
higher yield provided by PAC than GAC, PAC degrades VFA more efficiently than GAC |
[26] |
| PAC | 0.125–1 | pre-treated activated sludge subjected to thermal hydrolysis |
stimulates hydrolysis activity, increases methanogenic activity, and speeds up VS removal |
[31] |
| PAC | 15.0 | poultry blood | enhances syntrophic metabolism | [32] |
| PAC | 15.0 | organic fraction of municipal solid waste |
reduction of inhibitor content (FAN, VFA), 17% higher methane yield |
[33] |
Table 2.
Dosage and effects of biochar in the AD process.
| Dose of biochar (g/L) |
Type of substrate | Effect | References |
|---|---|---|---|
| 10 | glucose | reduced downtime and faster fermentation start | [40] |
| 10 | food waste | increased average methane yield by 14% | [41] |
| – | waste water | improved methane yield and enhanced degradation of protein substances |
[42] |
| 20 | volatile fatty acids | reduced downtime and faster methane production start | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



