
Submitted:
29 July 2024
Posted:
01 August 2024
You are already at the latest version
Abstract
Abstract: Epidemiological, experimental, and ecological data have indicated the controversial effect of in utero chronic low dose rate (100 mGy) dose radiation exposure. Our main goal of this study was to examine if different low dose rates of chronic pre- or/and post-natal radiation exposure with accumulative high doses could induce hippocampal cellular, mRNA, and miRNA changes leading to neuropsychiatric disorders. The comprehensive mouse phenotypic traits, organ weight pathological, blood mRNA and miRNA changes were also studied. Using different approaches including SmithKline, Harwell, Imperial College, Royal Hospital, Phenotype Assessment (SHIRPA), neurobehavioral tests, pathological examination, immunohistochemistry, mRNA and miRNA sequencing, and real-time quantitative polymerase chain reaction ( qRT-PCR) validation, we found that in prenatally irradiated (100 mGy/d for 18 days with an accumulative dose of 1.8 Gy) 1-year-old mice, no cellular changes including neurogenesis, mature neurons and glial cells in the hilus of the dentate gyrus and development of cognitive impairment, neuropsychiatric disorders occurred. However, a significant reduction in body weight and mass index (BMI) was indicated by the SHIRPA test. A reduced exploratory behavior was shown by an open field test. Organ weight measurement showed a significant reduction of the testes, kidneys, heart, liver, and epididymides although no abnormal changes from pathological examination. mRNA and miRNA sequencing and qRT-PCR validation revealed the upregulation of Rubcnl, Abhd14b and downregulation of Hspa1b, P4ha1, and Banp genes in both hippocampus and blood of mice prenatally irradiated with 100 mGy/d. Meanwhile, downregulation of miR-448-3p, miR1298-5p in the hippocampus, miR-320-3p, miR-423-5p, miR-486b-5p, miR-486b-3p, miR-423-3p, miR-652-3p, miR-324-3p, miR-181b-5p, miR-let-7b and miR-6904-5p in the blood was induced. The target scan revealed that Rubcnl is one of the miR-181b-5p targets in the blood. We, therefore, concluded that prenatal chronic irradiation with a low dose rate of 100 mGy/d and accumulative dose of 1.8Gy or below may not induce significant adverse health effects on the offspring. Further study of different low dose rate radiation exposure with accumulative high doses may provide threshold doses for authorities or regulators to set new radiation safety guidelines to replace those extrapolated from acute high dose/dose rate irradiation to reduce unnecessary emergency evacuation or expanding once a nuclear accident or leakage occurs.
Keywords:
Prenatal
; low dose rate
; chronic radiation exposure
; brain effect
1. Introduction
Acute high doses/dose-rates prenatal irradiation-induced adverse health effects, such as neural tube defects, microcephaly, severe mental retardation, cancers, and unprovoked seizures have been due to radiotherapy [1,2,3,4] and atomic bomb exposure in Hiroshima and Nagasaki [5,6,7]. Acute exposure to low doses at a high dose rate, as seen in prenatal diagnostic X-ray exposures, may not significantly increase the risk of cancer development [8] except for leukemia as reported by Schulze-Rath [9] and Wakeford [10]. While Down’s syndrome [11], trisomy 21 [12,13,14], neural tube defects, and microcephaly have been reported after Chernobyl chronic radiation exposure in humans [15,16,17], and microcephaly in the birds exposed to Chernobyl chronic radiation [18] and in monkeys exposed to Fukushima nuclear accident [19], no individual dose/dose rate has been reported for mother or embryo at different developmental stages of the prenatal period. These reports are not in agreement with the International Commission on Radiological Protection (ICRP) conclusion that irradiation throughout pregnancy with exposure rates of less than 0.05 Gy/day (or 2.08 mGy/h) are not measurably deleterious to the surviving offspring (ICRP report 90, Item 424). So far, limited lifetime or long-term chronic low dose rate ionizing radiation exposures in animal models have been done, and inconsistent radiation effects including life span, carcinogenesis, and neoplastic death have been reported [20,21,22,23,24,25,26,27]. Our previous review shows the variations in the effects of acute prenatal radiation exposures on the progeny[4], and a recent literature review concludes that both epidemiological and experimental studies do not provide convincing evidence to support radiation-induced developmental defects in the progeny after exposure in utero [28]. Furthermore, there are no epidemiological or experimental studies on cellular changes in the brain after prenatal chronic low (<100 mGy) or high dose with low dose rate (<6 mGy/h) radiation exposure. In the present study, the neurological, neurocognitive, and neuropsychiatric changes were examined in prenatal chronic continuous low dose rate irradiated progeny with SmithKline, Harwell, Imperial College, Royal Hospital, Phenotype Assessment (SHIRPA), novel object recognition, open field, forced swim and tail suspension tests respectively. Using different brain cell markers including newly generated immature neuronal marker doublecortin (DCX), mature neuronal marker neuronal nuclear protein (NeuN), astrocyte marker glial fibrillary acidic protein (GFAP), microglial marker ionized calcium-binding adapter molecule 1 (IBA1), and oligodendrocyte precursor cell marker platelet-derived growth factor receptor alpha (PDGFRα), we aimed to investigate prenatal low dose rate radiation effect on the progeny’s cellular changes in the dentate gyrus of the hippocampus. We focused on the dentate gyrus of the hippocampus as previous studies have shown obvious pathophysiological changes including neuronal loss, impairment of neurogenesis, and glial activation after different patterns of radiation exposure (4, 28). By immunohistochemical staining of DNA damage and repair marker H2A histone family member X (γH2AX), we examined if prenatal irradiation induced persistent DNA damage foci in the progeny’s brain. miRNA and mRNA sequencing and real-time quantitative reverse transcription PCR (qRT-PCR) analysis were done to reveal irradiation-induced gene changes. Furthermore, a pathological study was also done for different body organs. The results of this study may provide evidence for a better understanding of the effect of nuclear accidents on the offspring of pregnant women staying near the accident site or exposed to nuclear fallout.
2. Materials & Methods
2.1. Animals and Irradiation
Six-week-old mice (SPF C57BL/6JJcl females and C3H/HeNJcl males) purchased from CLEA Japan Inc., (Tokyo, Japan) were used as parent stocks and bred as described by Gulay et al. [27]. Pregnant dams were continuously exposed to whole body irradiation using 137Cs gamma rays for 22 h/day(d), at daily doses of 1 or 20 mGy/d, from gestation day (GD) 0 (when vaginal plugs are detected) up to post-natal day 30 (approximately 48 consecutive days), to total accumulated doses of 48 and 960 mGy, respectively. A third irradiation group was exposed to 100 mGy/d from GD 0-18 to a total dose of 1.8 Gy. Age-matched non-irradiated pregnant dams were also included.
The absorbed doses by the pregnant dam are based on measurements made using thermoluminescence dosimeters (TLDs) inserted into the abdomen of mice as described by Shiragai et al. [29]. Pups were carefully counted (as soon as possible after birth) and were allowed to stay with their dams until weaning at 21 days (3 weeks) of age, at which time they were individually identified with ear notches, weighed, separated by sex and group-caged (5 mice/cage).
A total of 64 male mice (n=16/group) including age-matched non-irradiated controls used in the study were housed at the Low-Dose Radiation Effects Research Facility (LERF) of the Institute for Environmental Sciences (IES), Japan.
The entire study was conducted under similar SPF environmental conditions and husbandry practices; which includes12 h light-dark cycle, weekly cage change, ad libitum feed and water supply, daily health monitoring or clinical inspection, and monthly monitoring of SPF status; as described previously [24]. All experiments were conducted according to legal regulations in Japan and following the Guidelines for Animal Experiments of the Institute for Environmental Sciences.
2.2. Behavioral Studies
A total of 32 B6C3F1 males (n = 16 non-irradiated and n=16 exposed in utero to 100 mGy/d for 18 days), were tested using the SmithKline, Harwell, Imperial College, Royal Hospital, Phenotype Assessment (SHIRPA), Open Field (Locomotor), Novel Object Recognition, Forced Swim and Tail Suspension Tests.
2.2.1. The SmithKline, Harwell, Imperial College, Royal Hospital, Phenotype Assessment (SHIRPA) test
SHIRPA is a rapid battery of tests of motor activity, coordination, postural control, muscle tone, autonomic functions, and emotional reactivity, as well as reflexes dependent on visual, auditory, and tactile modalities. Individual scores in SHIRPA are sensitive in detecting phenotypes of several experimental models of neural disease, especially cerebellar degeneration and Alzheimer’s disease, and combined sub-scores have been useful in estimating the impact of vascular anomalies and exposure to infectious agents [30]. The RIKEN Modified SHIRPA (Version 4*) was used in the present study. It included 68 test items related to the evaluation of morphology, behavior, sensory response, and athletic ability. The test was executed as a series of six categories comprising “In the viewing jar” to allow the mouse to get used to the environment and observe it; “In the Arena” to observe the behavior in the arena; “Above the Arena 1,” to observe the behavior above the grid put on the arena; “On the Arena,” to observe the morphology and behavior on the grid put on the arena; “Above the Arena 2” to observe the morphology and locomotion with the arena; and “Additional Comments” to evaluate the behavior and somatotype. The test took approximately 15 minutes to complete for each mouse (Please refer to https://ja.brc.riken.jp/lab/jmc/shirpa/ for the detailed protocol) [31].
2.2.2. Open Field (Locomotor) Test
The open field was done in an empty and opaque box with the dimensions 50 cm x 50 cm. The arena is divided into 3 areas; center, corners and outer in the software to track the distance travelled and time spent in each area. The mouse was placed in the center area at the start of the test and was allowed to explore for 30 minutes to observe behavior. The ANY-maze software (ANY-maze, Wood Dale, IL, US) detects the center of the mouse’s body, thus only detecting entry and tracking time and distance once half of the mouse’s body is in the area.
2.2.3. Novel Object Recognition Test
This test was done over 4 days: 2 habituation days, a training day, and a long-term test day. Mice were placed in an empty box with the dimensions 50 cm x 25 cm for 10 minutes. Habituation and training were done with 2 identical simple objects placed on opposite sides of the box. The long-term test day was conducted 24 hours after the training day where one of the simple objects from training was replaced with a novel object. Using the ANY-maze investigation zone feature (ANY-maze, Wood Dale, IL, US), we set a zone around the object with a radius similar to the mouse body length (2–3 cm). The software will detect and measure the amount of time the mouse spent in the 2 different object zones.
2.2.4. Forced Swim Test
A cylindrical tank of 20 cm diameter was used for the forced swim test with a water temperature of 24–26oC. Mice were placed in water and movement was recorded for 8 minutes. Immobility was detected by the ANY-maze (ANY-maze, Wood Dale, IL, US) when there were no limb movements for 3 seconds or more. Immobilization is regarded as an indication of depression-like behavior [32].
2.2.5. Tail Suspension Test
This test requires the mice to be suspended by their tails that were taped to a hook. Their movements were recorded with ANY-maze (ANY-maze, Wood Dale, IL, US) for 6 minutes with immobility detected when there were no limb movements for 3 seconds or more. The time spent as immobile was recorded for consideration for mice’s depressive-like behavior.
2.3. Pathological Examination
All 64 B6C3F1 male mice were sacrificed at approximately 1 year of age by carbon monoxide asphyxiation after which blood samples were collected via cardiac puncture and then subjected to necropsy (gross examination). Whole blood, 0.5 mL was transferred into 2 mL tubes pre-loaded with RNAlater solution and stored frozen at -80℃ until analyzed. Organs were collected, examined, weighed, and fixed in 10% neutral buffered formalin for histopathological examination based on a standard protocol [25]. When deemed necessary, additional tissue samples were collected from neoplasms and organs or tissues with gross abnormalities and special histochemical procedures were performed for diagnostic purposes. Histopathological examination was performed blindly and neoplasms were classified based on the proposed nomenclatures of WHO/IARC [33] and the NTP [34] as described previously [25]. Multiple primary neoplasms and pathologies were treated as in the previous lifespan study [25] wherein multiple (including multiple or metastatic foci) neoplasms of the same type were counted only once. All neoplasms were counted into the overall incidence.
The whole brain was dissected and separated sagittally into the left and right hemispheres. The right hemisphere was fixed in 4% paraformaldehyde for 24 hours, then transferred to 30% sucrose in 0.1M phosphate buffer (pH 7.4) for immunohistochemistry. The hippocampus was dissected from the left hemisphere and stored frozen at -80ºC until processed for RNA extraction.
2.4. Immunohistochemical Staining of the Hippocampus
Seven to nine serial sagittal sections of the left hemisphere of the brain (40μm thick) from 8 non-irradiated, 7 from the 100 mGy/d group, and 5 each from the 1 and 20 mGy/d groups were placed in 24-well-plate with PBS and immunostained for DCX, NeuN, PDGFRα, GFAP, IBA1, and γH2AX according to our previous study [35]. After treatment with 3% H2O2 (Sigma-Aldrich Pte Ltd., Singapore) and blocking with serum (Vector Laboratories Inc., Burlingame, CA, USA), free-floating sections were incubated with goat primary antibody against DCX (1: 500; Santa Cruz Biotechnology Inc., CA, USA, Catalog No: SC-8066), rabbit primary antibodies against NeuN (1: 1000; Invitrogen, MA, USA, Catalog No: PA5-37407), and PDGFRα (1: 200) (Catalog No: 3174S), GFAP (1: 200) (Catalog No: 12389S), IBA1 (1: 200) (Catalog No: 17189S) and γH2AX (1: 200) (Catalog No: 9718S) (Cell Signaling Technology, MA, USA) overnight. The sections were then washed and incubated with respective donkey anti-goat (Catalog No: ab6884) (Abcam Inc. MA, USA) and goat anti-rabbit (Catalog No: 14789S) (Cell Signaling Technology, MA, USA) secondary antibodies (1: 200) followed by avidin-biotin complex (ABC) reagent (Vector Laboratories Inc., Burlingame, CA, USA). After reaction in 3, 3′-diaminobenzidine (DAB) Peroxidase Substrate (Vector Laboratories Inc., Burlingame, CA, USA), the sections were then washed, mounted, counterstained, and covered.
The immunostained sections were examined and photographed (Leica Microsystems GmbH, Wetzlar, Germany) and the Stereologer System (Stereology Resource Center, Biosciences Inc. Tampa, FL, USA) was used to unbiasedly analyze the number of NeuN, PDGFRα, and GFAP immunopositive cells in the hilus; and, IBA1 immunopositive cells in the hilus and stratum granulosum, and indicated as the number/volume (mm3). DCX immunopositive cells in the subgranular zone were counted and indicated as a number/per subgranular length (mm).
2.5. RNA Extraction from the Hippocampus and Whole Blood
RNA extraction from the hippocampus was performed in 6 non-irradiated control and 6 prenatally irradiated mice from the 100 mGy/d group using the miRNeasy Mini Kit (Qiagen, Hilden, Germany). The hippocampus was homogenized in 700µl QIAzol lysis reagent, placed at room temperature for 5 min, added with 140 µl chloroform, and shaken vigorously for 15s. The tube was centrifuged for 15 min at 12000 x g at 4 0C. After centrifugation, the samples were separated into 3 phases. The upper colorless aqueous phase containing RNA was collected into a new tube, and mixed with 1.5 volumes of 100% ethanol. The above mixture was loaded into an RNeasy Mini spin column, and centrifuged at ≥8000 x g for 15s. The column was washed and centrifuged. RNA from the column membrane was finally eluded with 40 µl RNase-free water.
RNA from whole blood was isolated using Mouse RiboPure™-Blood RNA Isolation Kit (Life Technologies Holdings Pte Ltd. Singapore). Mouse blood was collected in a tube with pre-loaded RNAlater solution, and centrifuged for 3min at 15000 x g. The supernatant was removed. The cell pellet was reconstituted by adding a lysis solution, and vortexed vigorously, followed by 200 µl 3M sodium acetate and 1.5mL acid phenol: chloroform added. The tube was centrifuged for 10 min at 2000 x g. The aqueous upper phase was recovered and mixed with 1.2 volumes of 100% ethanol. The sample was then vacuum-filtered through a Filter Cartridge, washed, and eluted with 150 µl nuclease-free water. RNA concentration and integrity were checked using the Nanodrop and Bioanalyzer system (Agilent Technologies, Santa Clara, CA, USA) before being subjected to miRNA sequencing and qPCR analysis.
2.6. Systematic miR Sequencing (miRSeq) and mRNA Sequencing Analysis
miRSeq and mRNA sequencing carried out using the DNB SEQ platform (BGI, Beijing, P.R. China), detected 1976 miRs, and 19,039 mRNAs were further analyzed by DESeq2 method.
2.7. Real-Time Quantitative Reverse Transcription PCR (qRT-PCR) Analysis of miR
RNA was first reversely transcribed using the miScript II RT kit (Qiagen, Hilden, Germany). The 20 µl reaction mixture, including 4 µL 5x HiSpec buffer, 2 µL 10x nucleotide mix, 2 µL reverse transcripts mix, 5 µL template RNA, and 7 µL nuclease-free water, was incubated at 37 °C for 1 hour followed by 95 °C for 5 minutes. The resulting cDNA was then diluted by adding 80 µl of nuclease-free water and stored at -80 °C until analysis.
20 µL of master mix, for real-time PCR, was prepared as follows: 2 µL diluted cDNA, 10 µL 2x miScript SYBR green PCR master mix, 2 µL 10x miScript universal primer and 2 µL primer for target miR (Table 1), and 4 µL nuclease-free water. Samples were denatured at 95 °C for 15 minutes, followed by 40 cycles of denaturation at 94 °C for 15 seconds, annealing at 55 °C for 30 seconds, and extension at 70 °C for 30 seconds. PCR amplification was carried out in QuantStudio 6 Real-Time PCR Systems (Thermo Fisher Scientific, Waltham, MA, USA). Fluorescence data were then collected. The expression of miR-68 was used as an internal control.
2.8. Real-Time RT-PCR Analysis for mRNA
For real-time RT-PCR analysis of mRNA, RNA was reverse transcribed using Maxima first-strand cDNA synthesis kits (Thermo Fisher Scientific, Waltham, MA, USA). 1 µg RNA was added to 4 µL 5X Reaction Mix, 2 µL Maxima Enzyme Mix, and topped up to 20 µL with nuclease-free water. The tubes were thereafter incubated at 25 ˚C for 10 minutes, followed by 50 ˚C for 45 minutes and 85 ˚C for 5 minutes. The resulting cDNA was then diluted by adding 100 µl of nuclease-free water and stored at -20 ˚C.
A master mix of 20 µL for real-time PCR was prepared as follows: 2 µL diluted cDNA, 10 µL 2x Maxima SYBR Green qPCR Master Mix, 2 µL 10x forward and reverse primers for target genes (Table 2), and 4 µL nuclease-free water.
PCR amplification was carried out in QuantStudio 6 Real-Time PCR Systems (Thermo Fisher Scientific, Waltham, MA, USA). The samples were initially denatured at 95 ˚C for 10 minutes, followed by 40 cycles of: denaturation at 95 ˚C for 15 seconds, annealing at 60 ˚C for 30 seconds, and extension at 72 ˚C for 30 seconds. The fluorescence data was collected after the extension step. The expression of GAPDH was used as internal control.
2.9. Statistical Analyses
Student’s t-test was used to compare behavioral changes between the non-irradiated control and irradiated mice. Fischer’s exact tests were used to analyze the non-neoplastic lesions and neoplasm incidence. Levels of significance for incidence rates of non-neoplastic lesions and neoplasms were chosen as P < 0.05. For detected miRs and mRNAs analysis, the parameters for calculating the significantly differential expression of DEseq2 are |log2FC| >0.585 and P<0.05. To compare the immunostained DCX, NeuN, PDGFRα, GFAP, and IBA1 cells among the control, pre- and/or post-natally irradiated mice, One-Way ANOVA followed by a Post Hoc test was done. Brown-Forsythe test and Bartlett’s test were used to check data normality and homogeneity respectively. Turkey test was done as a post hoc test to compare the mean of each group of mice with the mean of every other group.
3. Results
3.1. Behavioral Studies
3.1.1. SHIRPA Test
The results of the SHIRPA test (68 parameters) in male B6C3F1 at 1 year after prenatal irradiation showed significant differences in body weight and body mass index (BMI) between irradiated mice (100 mGy/d) and the non-irradiated controls (Figure 1). Here, BMI is defined as
.
ere W is body weight in grams and L is body length in mm. There was no significant change in other parameters examined (for details, please refer to the website https://ja.brc.riken.jp/lab/jmc/mouse_clinic/SOPs/Classification_by_Platform/Other/RIKENMPP_001_004_02_modified_SHIRPA_v4.xml). The results suggest that prenatal irradiation with 100 mGy/d for 18 days does not induce any impairment of sensorimotor function nor emotional reactivity 1 year after radiation exposure.
3.1.2. Open Field Test
Open field test showed that the average distance the irradiated mice travelled to different areas of the box was reduced significantly (P<0.05) (Figure 2A) compared to the non-irradiated controls, although the time spent in each area was not significantly different (Figure 2B). Results suggest that irradiated mice had decreased locomotor activity or reduced exploratory behavior than the non-irradiated control mice.
3.1.3. Novel Object Recognition, Forced Swim (Figure 3B), and Tail Suspension Tests
Results of the novel object recognition test showed that both the non-irradiated control and the irradiated groups spent the same amount of time with the novel object, suggesting that prenatal irradiation did not impair novel object recognition (Figure 3A). Forced swim (Figure 3B) and tail suspension (Figure 3C) tests did not show significant differences between groups in immobile time, suggesting that these irradiated mice did not develop depression.
3.2. Pathology
As the SHIRPA test indicated a significant reduction of body weight in mice prenatally irradiated with 100 mGy/d, adipose tissue deposits and different organs including left and right testis, kidney, heart, liver, lung, epididymides, spleen, thymus were collected, and weighed, the results did not show a significant reduction in the weight of adipose tissue deposits between the irradiated and control mice (Figure 4A). However, there was a significant reduction of the weight of testis (control, right 130±7.11, Exp right 92±2.08; control left 119±2.07), kidney (Control right 293.63±6.89, Exp right 266.19±3.71), heart (Control 192.44, Exp 166.14±10.35), liver (control 2269.44±92.63, Exp 2019.50±60.02) and epididymides (control 105.63± 6.55, Exp 80± 1.28) (Figure 4B).
Histopathological examination showed that the most common neoplastic lesions in males were hepatocellular adenoma, hepatocellular carcinoma, and lung adenoma. However, there was no significant difference in the incidence of these lesions between the non-irradiated and irradiated groups (Table I). Non-neoplastic lesions observed included subcapsular cell hyperplasia of the adrenal gland, valvular endocardiosis in the heart, cytoplasmic vacuolation and granular degeneration in the liver, and inflammation in the lung but incidence rates were not significantly different between the non-irradiated and irradiated groups. The study indicates that continuous low-dose-rate irradiation during the gestation period (gestation days 0-18) and the juvenile period (1-30 days) does not lead to the development of neoplasms or non-neoplastic lesions at the examined endpoint.

Table 1.
Histopathological findings in male B6C3F1 mice.
| Lesion | Organ | Lesion | Control (0 mGy) n=16 (%) |
1 mGy/d (48 mGy) n=16 (%) |
20 mGy/d (960 mGy) n=16 (%) |
100 mGy/d (1800 mGy) n=16 (%) |
|---|---|---|---|---|---|---|
| Neoplastic lesion | Liver | Adenoma, Hepatocellular | 1 (6.3) | 3(18.8) | 1 (6.3) | |
| Carcinoma, Hepatocellular |
1 (6.3) | 2 (12.5) | ||||
| Lung |
Adenoma, Bronchiolo-Alveolar | 4 (25.0) | 3 (18.8) | 1 (6.3) | ||
| Non-neoplastic lesion | Adrenal gland | Accessory cortical tissue | 1(6.3) | 1 (6.3) | 1 (6.3) | 1 (6.3) |
| Hyperplasia, subcapsular cell | 6 (37.5) | 5 (31.3) | 6 (37.5) | 10 (62.5) | ||
| Heart | Valvular endocardiosis | 6 (37.5) | 5 (31.3) | 3 (18.8) | 4 (25.0) | |
| Kidney | Cysts | 1 (6.3) | ||||
| Hyperplasia, renal tubular | 1 (6.3) | |||||
| Liver | Cellular alteration, foci | 1 (6.3) | 1 (6.3) | |||
| Cytoplasmic vacuolization | 14 (87.5) | 11 (68.8) | 12 (75.0) | 13 (81.3) | ||
| Granular degeneration | 9 (56.3) | 9 (56.3) | 12 (75.0) | 9 (56.3) | ||
| Inflammation | 1 (6.3) | 1 (6.3) | ||||
| Lung | Congestion | 1 (6.3) | 1 (6.3) | 1 (6.3) | 2 (12.5) | |
| Metaplasia, osseous | 1 (6.3) | |||||
| MNC inf, perivascular | 2 (12.5) | |||||
| Pneumonia, interstitial | 1 (6.3) | 2 (12.5) |
3.3. Immunohistochemistry
Immunohistochemical study did not show significant difference in the number of mature neurons (NeuN, Figure 5A-E) in the hilus, immature neurons (DCX, Figure 5A1-E1) in the subgranular zone, microglia (IBA1, Figure 5A2-E2) in both hilus and the stratum granulosum, astrocytes (GFAP, Figure 5A3-E3) and oligodendrocyte precursor cells (PDGFRα, Figure 5A4-E4) in the hilus of the dentate gyrus of the hippocampus between the non-irradiated control and irradiated groups.
γ -H2AX immunostaining did not detect persistent DNA damage foci 1 year after pre-and/or post-natally irradiated male mice (Figure 5A5-D5).
3.4. Systematic miR Sequencing (miRSeq) and mRNA Sequencing Analysis
mRNA sequencing analysis showed 1549 differentially expressed mRNA in whole blood and 228 differentially expressed mRNA in the hippocampus. The Venn analysis indicated that 27 mRNAs were altered in both whole blood and hippocampus (Figure 6A). Of the differentially expressed mRNA, 20 or 25 highly and significantly expressed mRNA with the same up- or down-expression trend from the hippocampus or whole blood respectively were validated with qRT-PCR. Some differentially expressed mRNAs from mRNA sequencing results were summarized in heatmap in the hippocampus and blood (Figure 6B & 6C). The PCR and mRNA sequencing results showed that RUN and cysteine-rich domains containing beclin 1 interacting protein like (Rubcnl) and Alpha/beta hydrolase domain-containing protein 14B (Abhd14b) genes were up-regulated, whereas heat shock protein family A member 1B (Hspa1b), prolyl 4-hydroxylase subunit alpha 1 (P4ha1) and BTG3 associated nuclear protein (Banp) genes were down-regulated in both the hippocampus (Figure 6B & 7A) and whole blood (Figure 6C & 7B). In the hippocampus, significant down-regulation of Dnajb1, Arc, Pdia4, Fos, Tm6sf2, Tent5, Cdkl5, H2bc23, Arhgef5, Crybb3 (Figure 6B & 7C) were observed. In whole blood, CD74 was up-regulated while F5, Stip1, Ahsa2, Dap, Sh3bgrl2, Nptn, Hsp90aa1and Hsp90ab1genes were down-regulated (Figure 6C & 7D). There was no change in the expressions of F5, Dusp6, Egr2, Hspb1, CD74 in the hippocampus (Fig. Suppl 1A), nor of Vmn1r58, Scd1, Cd59b, Slc5a3, Ccdc117, Cish, Fosb, Gem, H2bc24, Fam122a, Sesn3 in whole blood (Fig. Suppl 1B) demonstrated. Of 4 differentially expressed miRNA (miR-202-5p, miR-448-3p, miR1298-5p, and miR-212-5p) in the hippocampus from sequencing analysis (Figure 8A), qRT-PCR validated the down-regulation of miR-448-3p and miR1298-5p (Figure 8B). Of the 75 differentially expressed miRNA in the whole blood from sequencing analysis, 16 highly or significantly expressed miRNAs were tested by qRT-PCR. miR-320-3p, miR-423-5p, miR-486b-5p, miR-486b-3p, miR-423-3p, miR-652-3p, miR-324-3p, miR-181b-5p, miR-let-7b and miR-6904-5p were down-regulated both in miRNA sequencing and qRT-PCR (Figure 8C&D). However, miR-122-5p, miR-744-5p, miR-let-7d-3p, miR-328-3p, miR-151-3p, miR-296-5p were not changed (Fig. Suppl 1C). The target scan indicated that among whole blood miRNA and mRNA changes, Rubcnl is one of the miR-181b-5p targets.
4. Discussion
4.1. Main Findings
Prenatal chronic low dose rate irradiation with 100mGy/d significantly reduced body weights and BMI, but no other change was observed when the SHIRPA test was done on the offsprings at the age of 1-year-old male B6C3F1 mice These mice also showed reduction in exploratory behavior, but no memory loss and neuropsychiatric impairment as tested by the novel object recognition, forced swim, and tail suspension tests respectively. Histopathological examinations showed neoplastic and non-neoplastic lesions in various organs such as the liver, lung, heart, adrenal glands, and kidney, there was no significant difference among the non-irradiated control and irradiated groups exposed to 1, 20, and 100 mGy/d although some organ weight was reduced. The chronic low dose rate irradiation did not induce any significant cellular changes in the dentate gyrus including newly generated neurons in the sub-granular zone, mature polymorphic neurons, microglia, astrocytes, oligodendrocyte precursor cells, in the hilus. Prenatal irradiation with 100 mGy/d (accumulated dose of 1.8Gy) also did not induce persistent DNA damage foci stained by γ H2AX in the granule cells of the dentate gyrus, suggesting that prenatal irradiation-induced DNA damage may be repaired by the age of 1-year-old. mRNA and miRNA sequencing and qRT-PCR validation revealed the changes of some mRNAs and miRNAs, in particular, up-regulation of Rubcnl and Abhd14b, and down-regulation of Hspa1b, P4ha1, Banp genes in both hippocampus and whole blood of mice irradiated with 100 mGy/d. In the whole blood, Rubcnl may be one of miR-181b-5p targets as indicated by the Target scan.
4.2. Pre- and Post-Natal (First 31 Days) Irradiation with a Low Dose Rate of 1mGy/d and 20 mGy/d Did Not Induce Obvious Pathological Changes in the Brain and Other Organs
A previous report on in-utero low dose-rate gamma-ray exposure of 20 mGy/d for the entire gestation period (accumulated dose: 360 mGy) in B6C3F1 mice did not cause any significant effect in pups when compared to the nonirradiated controls up to 10 weeks of age [27]. Further study indicated that in utero low dose-rate gamma ray exposures to 0.05, 1.0 and 20 mGy/d did not affect the reproductive parameters such as litter size and weaning rates among the 3 groups. Mean life spans and tumor spectra were not significantly different among the groups irradiated with 0.05, 1.0, and 20 mGy/d compared to the non-irradiated controls [26]. Results of these two studies suggest that chronic in-utero exposure to gamma rays at dose rates below 20 mGy/d for the entire gestation period does not induce obvious harmful health effects, at both early and late adult life stages of B6C3F1 mice. The negative results or no adverse effect from combined pre- and post-natal chronic irradiation with 1mGy/d (accumulated dose of 48 mGy) or 20 mGy/d (accumulated dose of 960 mGy) in 1-year-old (middle age) mice further support the conclusion that prenatal irradiation with 20 mGy/d does not induce harmful health effects from the early adulthood, middle age until natural death. This conclusion was also consistent with previous studies showing that in utero r-ray irradiation for the entire gestation period (days 1 to 18) in mice [36,37] and rats [38] with dose rates ranging from 25 mGy/d to 124 mGy/d produce no harmful effect. However, the induced histopathological changes at different life stages of the animal after prenatal medium dose rates of 200 mGy/d and 400 mGy/d to total accumulated doses of 3600 mGy and 7200 mGy [26], respectively, suggest that a threshold dose/dose rate may exist from 20 to 200mGy/d when one of the parameters is fixed.
4.3. Prenatal Irradiation with Dose Rate of 100 mGy/d (4.55 mGy/h, Total Accumulated Dose= 1.8 Gy) Did Not Induce Obvious Pathophysiological Changes in Middle-Aged Mice
While the present study showed that the prenatal chronic low dose rate irradiation with 100 mGy/d significantly reduced body weights, BMI, and some organ weight, the pathological study did not indicate any abnormal changes. Further study with a large sample size may confirm if this dose rate truly induces animal body physical changes. No detectable changes in the offspring’s motor activity, coordination, postural control, muscle tone, autonomic functions, emotional reactivity, reflexes (dependent on visual, auditory, and tactile modalities) as well as neurobehavior including cognition and neuropsychiatry after the prenatal chronic low dose-rate irradiation with 100 mGy/d suggest that animal may have a normal life at least during the first half of their lifespan. At the molecular level, the significance of the up-regulation of Rubcnl, Abhd14b, and down-regulation of Hspa1b, P4ha1, and Banp genes in both the hippocampus and whole blood, up- or down-regulation of other mRNA and miRNA in either the hippocampus or whole blood of mice prenatally irradiated with 100 mGy/d remain unknown due to limited functional study of the roles of these mRNA and miRNA.
Rubcnl, a recently identified novel accessory protein of PtdIns3K complexes, positively regulates autophagosome maturation and is primarily localized in the endoplasmic reticulum (ER) and autophagic structures. It antagonizes RUBCN to stimulate PIK3C3/Vps34 kinase activity and to recruit PtdIns3K and HOPS complexes to the autophagosome for their site-specific activation by anchoring to the autophagosomal SNARE STX17. Hepatocyte-specific Rubcnl ablation in mice results in impaired autophagy flux, glycogen and lipid accumulation, and liver fibrosis, whereas overexpression of Rubcnl in mouse livers to alleviate non-alcoholic fatty liver disease [39]. Alpha/beta hydrolase domain-containing protein 14B (Abhd14b) belongs to the α/β hydrolase superfamily of enzymes, which are known for their diverse roles in lipid metabolism, signal transduction, and cellular homeostasis, it is found in various tissues throughout the body, including the brain, liver, and adipose tissue, suggesting its potential roles in lipid metabolism, neuronal function, and possibly other cellular processes since enzymes in the α/β hydrolase family often act as lipases, esterases, or thioesterases, cleaving ester bonds in various lipid molecules. From a speculative point of view, the upregulation of Rubcnl and Abhd14b in the blood may play a beneficial role in alleviating radiation-induced decrease of different organ weights and potential abnormal lipid metabolism due to their anti-fibrosis and involvement in lipid metabolism. The upregulation of Rubcnl and Abhd14b in the hippocampus remains to be further investigated.
Hspa1b gene and its protein HSP70-2 are involved in oxidative stress response and an indel polymorphism in Hspa1b may be associated with the risk of sudden cardiac death [40]. Maternal separation (MS) enhanced the expression of Hspa1b mRNA in the blood and medial prefrontal cortex (mPFC) of juvenile and pre-adolescent rats [41] and was accompanied by an increase in the Hspa1a/1b protein levels in the mPFC and hippocampus of juvenile rats that persisted in the mPFC until adulthood. These changes suggest that Hspa1b may be a potential candidate peripheral and brain biomarkers of early-life stress (ELS)-induced changes in brain functioning [34], hepatocellular carcinoma [42], and in the early prediction and progression of Type 2 diabetes mellitus (T2DM) [43]. The level of Hspa1b in whole blood is also a sensitive marker for distinguishing tophi patients from healthy people [44]. Prolyl 4-hydroxylase subunit alpha 1 (P4ha1) has been identified as a valuable transcriptional genetic marker in the blood, liver, and adrenal tissues of female Sprague–Dawley rats under heat stress conditions [45]. P4ha1 is one of the metabolic-related genes for predicting poor clinical prognosis and immune microenvironment in primary melanomas [46], lung adenocarcinoma [47], and squamous cell carcinoma of the head and neck [48]. P4ha1 protects nasopharyngeal carcinoma (NPC) cells from erastin-induced ferroptosis by activating cytoplasmic 3-hydroxy-3-methylglutaryl-CoA synthase (HMGCS1), suggesting it may be a novel molecular marker of NPC ferroptosis resistance that results in poor prognosis, and that the P4ha1/HMGCS1 axis may be a new target for the treatment of NPC progression [49]. P4ha1 may also be a potential biomarker in human atrial fibrillation [49] since hypoxia-induced endothelial P4ha1 overexpression has been reported to enhance angiogenesis by promoting glycolytic metabolism reprogramming through the P4ha1/α-ketoglutarate (α-KG)/ten-eleven translocation 2 (Tet2)/fructose-1,6-biphosphatase (Fbp1) promoter pathway, leading to the decreased Fbp1, enhanced glycolytic metabolism, suggesting its therapeutic potential for post-ischemic tissue repair [50]. Banp gene has been identified as a new prognostic biomarker of high-risk acute myeloid leukemia (AML) [51]. Whether the down-regulation of Hspa1b, P4ha1, and Banp genes in whole blood and in the hippocampus in the present study is involved in the body’s self-protective mechanism in preventing low dose rate irradiation-induced early-life stress and cancers remains unknown. Given that limited studies have been done to investigate the functional changes of miRNAs validated in this study, the functional significance of the down-regulation of miR-448-3p, miR1298-5p in the hippocampus and miR-320-3p, miR-423-5p, miR-486b-5p, miR-486b-3p, miR-423-3p, miR-652-3p, miR-324-3p, miR-181b-5p, miR-let-7b and miR-6904-5p in whole blood needs to be further studied. Interestingly, among miRNA and mRNA changes in whole blood, Rubcnl is one of the miR-181b-5p targets by Target Scan. It remains to be studied whether the interaction of blood miR-181b-5p with Rubcnl is involved in animal weight changes induced by prenatal low dose rate radiation exposure.
4.4. Summary
The present study indicates that the prenatal and postnatal (first 30 days) continuous low dose rates of irradiation with 1 or 20 mGy/d, prenatal continuous low dose rates of irradiation with 100 mGy/d may not induce pathological changes in different organs, in particular, brain cellular distribution in the dentate gyrus of the hippocampus. SHIRPA and different neurobehavioural tests suggest that these animals may have a normal life, at least during the first half of their lifespan. The significant changes of miRNA and mRNA in the blood and hippocampus of animals with prenatal irradiation with 100 mGy/d remain unknown. Our results suggest that the chronic low dose rate irradiation with accumulated high dose may not be as harmful as previously expected from the linear no-threshold (LNT) model. The investigation of the health effect of chronic low dose rate of radiation with a cumulative threshold dose may be a promising area to explore in radiobiology research which may provide solid evidence for understanding the health effect of chronic low dose rate radiation exposure, for environmental radioprotection and safety policy preparation. The traditional LNT model can not be used for the evaluation of the health effects of chronic low dose rate radiation exposure.
Author Contributions
Conceptualization, F.R.T., I.B.T; methodology, F.R.T., I.B.T., H.W., S.T., S.L., investigation, H.W., S.T., S.L., A.T., D.T. and A.A. data curation, F.R.T.; I.B.T.; writing: original draft preparation, F.R.T.; writing review and editing, F.R.T., I.B.T.; H.W., S.L., A.T., supervision, F.R.T.; I.B.T.; funding acquisition, F.R.T., I.B.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research Foundation of Singapore and the Aomori Prefectural Government, Japan
Institutional Review Board Statement
All experiments were conducted according to legal regulations in Japan and following the Guidelines for Animal Experiments of the Institute for Environmental Sciences (Animal Research Ethics Committee Processing number: 22)
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
This study was supported by grants from the National Research Foundation of Singapore to the Singapore Nuclear Research and Safety Initiative (F.R.T.), the Aomori Prefectural Government, Japan (I.B.T), and the animal behavioral experiments were carried out at the Low-Dose Radiation Effects Research Facility (LERF) of the Institute for Environmental Sciences (IES), Japan. The technical support from Kael Lee Koon Lam is greatly appreciated.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dekaban, A.S. , Abnormalities in children exposed to x-radiation during various stages of gestation: tentative timetable of radiation injury to the human fetus. I. J. Nucl. Med. 1968, 9, 471–477. [Google Scholar]
- Goldstein, L. , Murphy, D.P., Etiology of ill health in children born after maternal pelvic irradiation. II. Defective children born after post conceptional maternal irradiation. Am. J. Roentgenol. 1929, 22, 322–331. [Google Scholar]
- Michalet, M. , et al., Radiotherapy and pregnancy. Cancer Radiother. 2021, 26, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Yang, B., B. X. Ren, and F.R. Tang, Prenatal irradiation-induced brain neuropathology and cognitive impairment. Brain Dev. 2017, 39, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Otake, M. and W.J. Schull, Radiation-related brain damage and growth retardation among the prenatally exposed atomic bomb survivors. Int J Radiat Biol 1998, 74, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Otake, M., W. J. Schull, and H. Yoshimaru, A review of forty-five years study of Hiroshima and Nagasaki atomic bomb survivors. Brain damage among the prenatally exposed. J Radiat Res 1991, 32, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, J.N., S. W. Wright, and P.M. Wright, A study of the outcome of pregnancy in women exposed to the atomic bomb blast in Nagasaki. J. Cell. Comp. Physiol. 1954, 43, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Wit, F., C. Vroonland, and H. Bijwaard, Prenatal X-ray Exposure and the Risk of Developing Pediatric Cancer-A Systematic Review of Risk Markers and a Comparison of International Guidelines. Health Phys. 2021, 121, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Rath, R., G. P. Hammer, and M. Blettner, Are pre- or postnatal diagnostic X-rays a risk factor for childhood cancer? A systematic review. Radiat. Environ. Biophys. 2008, 47, 301–312. [Google Scholar] [CrossRef]
- Wakeford, R. , Childhood leukaemia following medical diagnostic exposure to ionizing radiation in utero or after birth. Radiat. Prot. Dosim. 2008, 132, 166–174. [Google Scholar] [CrossRef]
- Zatsepin, I. , et al., Down syndrome time-clustering in January 1987 in Belarus: link with the Chernobyl accident? Reprod Toxicol 2007, 24, 289–295. [Google Scholar] [CrossRef]
- Sperling, K., H. Neitzel, and H. Scherb, Evidence for an increase in trisomy 21 (Down syndrome) in Europe after the Chernobyl reactor accident. Genet Epidemiol 2012, 36, 48–55. [Google Scholar] [CrossRef]
- Sperling, K. , et al., Significant increase in trisomy 21 in Berlin nine months after the Chernobyl reactor accident: temporal correlation or causal relation? Bmj 1994, 309, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Sperling, K. , et al., Frequency of trisomy 21 in Germany before and after the Chernobyl accident. Biomed Pharmacother 1991, 45, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Wertelecki, W. , Chornobyl radiation-congenital anomalies: A persisting dilemma. Congenit Anom (Kyoto) 2021, 61, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Wertelecki, W. , et al., Chornobyl 30 years later: Radiation, pregnancies, and developmental anomalies in Rivne, Ukraine. Eur J Med Genet 2017, 60, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Wertelecki, W. , et al., Chornobyl, radiation, neural tube defects, and microcephaly. Eur J Med Genet 2018, 61, 556–563. [Google Scholar] [CrossRef]
- Møller, A.P. , et al., Chernobyl birds have smaller brains. PLoS One 2011, 6, e16862. [Google Scholar] [CrossRef]
- Hayama, S.I. , et al., Small head size and delayed body weight growth in wild Japanese monkey fetuses after the Fukushima Daiichi nuclear disaster. Sci Rep 2017, 7, 3528. [Google Scholar] [CrossRef]
- Lorenz, E. , Gorgdon, C., Deringer, M., Hollcraft, J. Long-term effects of acute and chronic irradiation in mice: Survival and tumor incidence following chronic irradiation of 0.11 r per day. J. Natl. Cancer Inst. 1049. [Google Scholar]
- Spalding, J. , Thomas, R., Tietjen, G. Life-span of c57 mice ad influenced by radiation dose: dose rate at exposure. Rep No Vc 48-LA 9528. Los Alamos, Los Alamos National Laboratory, 1982.
- Carnes, B.A. , Fritz, T.E. Rsponses of the beagle to protracted irradiation. I. Effect of total dose and dose rate. Radiat Res. 1991, 128, 125–132. [Google Scholar] [CrossRef]
- Carnes, B.A. , Fritz, T.E. Continuous irradiation of beagles with gamma rays. Radiat Res. 1993, 136, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S. , et al., No lengthening of life span in mice continuously exposed to gamma rays at very low dose rates. Radiat. Res 2003, 160, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, I.B. , 3rd, et al., Cause of death and neoplasia in mice continuously exposed to very low dose rates of gamma rays. Radiat.Res. 2007, 167, 417–437. [Google Scholar] [CrossRef]
- Tanaka, I.B. , et al., Life Span, Cause of Death and Neoplasia in B6C3F1 Mice Exposed In Utero to Low- and Medium-Dose-Rate Gamma Rays. Radiat. Res. 2022, 198, 553–572. [Google Scholar] [CrossRef] [PubMed]
- . Gulay, K.C.M., et al., Effects of continuous gamma-ray exposure in utero in B6c3f1 mice on gestation day 18 and at 10 weeks of age. Radiat Res 2018, 189, 425–440. [CrossRef]
- Benotmane, M.A. and K.R. Trott, Epidemiological and experimental evidence for radiation-induced health effects in the progeny after exposure in utero. Int J Radiat Biol 2023, 1–12. [CrossRef] [PubMed]
- Shiragai, A., et al., Estimation of the Absorbed Dose to Mice in Prolonged Irradiation by Low-Dose Rate γ-Rays from 137Cs Sources. Radioisotopes 1997, 46, 904–911. [CrossRef]
- Lalonde, R., M. Filali, and C. Strazielle, SHIRPA as a Neurological Screening Battery in Mice. Curr Protoc 2021, 1, e135. [CrossRef]
- Masuya. H,; Inoue, M.; Wada, Y.; Shimizu, A.; Nagano, J.; Kawai, A.; Inoue, A.; Kagami, T.; Hirayama, T.; Yamaga, A.; et al. Implementation of the modified-SHIRPA protocol for screening of dominant phenotypes in a large-scale ENU mutagenesis program. Mamm Genome. 2005, 16, 829–837. [CrossRef]
- Tang, F.R., Loke, W. K., Wong, P., & Khoo, B. C.Radioprotective effect of ursolic acid in radiation-induced impairment of neurogenesis, learning and memory in adolescent BALB/c mouse. Physiology & Behavior, 2017. 175, 37-46. 33.
- Mohr, U., International Classification of Rodent Tumors: The Mouse. 2001: Springer-Verlag, Berlin, Heidelberg.
- Maronpot, R.R., Pathology of the Mouse: Reference and Atlas.. 1999: Cache River Press, Vienna, Illinois, USA.
- Wang, H.; Ma, Z.; Shen, H.; Wu, Z.; Liu, L.; Ren, B.; Wong, P.; Sethi, G.; Tang, F.R. Early Life Irradiation-Induced Hypoplasia and Impairment of Neurogenesis in the Dentate Gyrus and Adult Depression Are Mediated by MicroRNA- 34a-5p/T-Cell Intracytoplasmic Antigen-1 Pathway. Cells 2021, 10, 2476.
- Konermann, G., [Mouse germ development following continuous Co60 gamma irradiation during blastogenesis, organogenesis and fetal period]. Strahlentherapie 1969, 137, 451–466.
- 37. Russell L.B., B.S.K., Saylors C.L., Comparison of the effects of acute, continuous and fractionated irradiation during embryonic development. In: Buzzati-Traverso A.A. (ed.). Immediate and Low-level Effects of Ionizing Radiation Conference Held in Venice. 1960, London.: Taylor & Francis. 343-359.
- Vorisek, P., [EFFECT OF CONTINUOUS INTRAUTERINE IRRADIATION ON PERINATAL MORTALITY OF THE FETUS]. Strahlentherapie 1965, 127, 112–120.
- Cheng, X. and Q. Sun, RUBCNL/Pacer and RUBCN/Rubicon in regulation of autolysosome formation and lipid metabolism. Autophagy 2019, 15, 1120–1121. [CrossRef] [PubMed]
- Yang, Z., et al., Genetic association study of a novel indel polymorphism in HSPA1B with the risk of sudden cardiac death in the Chinese populations. Forensic Sci Int 2021, 318, 110637. [CrossRef] [PubMed]
- Solarz, A., et al., A Search for Biomarkers of Early-life Stress-related Psychopathology: Focus on 70-kDa Heat Shock Proteins. Neuroscience 2021, 463, 238–253. [CrossRef]
- He, Y., et al., Single-cell profiling of human CD127(+) innate lymphoid cells reveals diverse immune phenotypes in hepatocellular carcinoma. Hepatology 2022, 76, 1013–1029. [CrossRef]
- Ali, H.S., et al., Identification of a Multi-Messenger RNA Signature as Type 2 Diabetes Mellitus Candidate Genes Involved in Crosstalk between Inflammation and Insulin Resistance. Biomolecules, 2022. 12(9).
- Li, Y., et al., Up-regulation of HSPA1A and HSPA1B in the blood of tophi patients and its clinical significance. Acta Biochim Pol 2022, 69, 781–785.
- Dou, J., et al., Comprehensive RNA-Seq Profiling Reveals Temporal and Tissue-Specific Changes in Gene Expression in Sprague-Dawley Rats as Response to Heat Stress Challenges. Front Genet 2021, 12, 651979. [CrossRef] [PubMed]
- Eriksson, J., et al., Prolyl 4-hydroxylase subunit alpha 1 (P4HA1) is a biomarker of poor prognosis in primary melanomas, and its depletion inhibits melanoma cell invasion and disrupts tumor blood vessel walls. Mol Oncol 2020, 14, 742–762. [CrossRef]
- Zhou, H., et al., Overexpression of P4HA1 Is Correlated with Poor Survival and Immune Infiltrates in Lung Adenocarcinoma. Biomed Res Int 2020, 2020, 8024138.
- Cao, Y., et al., A novel metabolic-related gene signature for predicting clinical prognosis and immune microenvironment in head and neck squamous cell carcinoma. Exp Cell Res 2023, 428, 113628. [CrossRef]
- Zhou, R., et al., P4HA1 activates HMGCS1 to promote nasopharyngeal carcinoma ferroptosis resistance and progression. Cell Signal 2023, 105, 110609. [CrossRef]
- Xu, Y., et al., Hypoxia-induced P4HA1 overexpression promotes post-ischemic angiogenesis by enhancing endothelial glycolysis through downregulating FBP1. J Transl Med 2024, 22, 74. [CrossRef] [PubMed]
- Villar, S., et al., The transcriptomic landscape of elderly acute myeloid leukemia identifies B7H3 and BANP as a favorable signature in high-risk patients. Front Oncol 2022, 12, 1054458. [CrossRef] [PubMed]
Figure 1.
Animal body changes: The SHIRPA test indicates a significant reduction of body weight (A) and body mass index (BMI) (B) in mice prenatally irradiated with a dose rate of 100 mGy/d for the entire gestation period. No difference in body length is observed (C).
Figure 1.
Animal body changes: The SHIRPA test indicates a significant reduction of body weight (A) and body mass index (BMI) (B) in mice prenatally irradiated with a dose rate of 100 mGy/d for the entire gestation period. No difference in body length is observed (C).
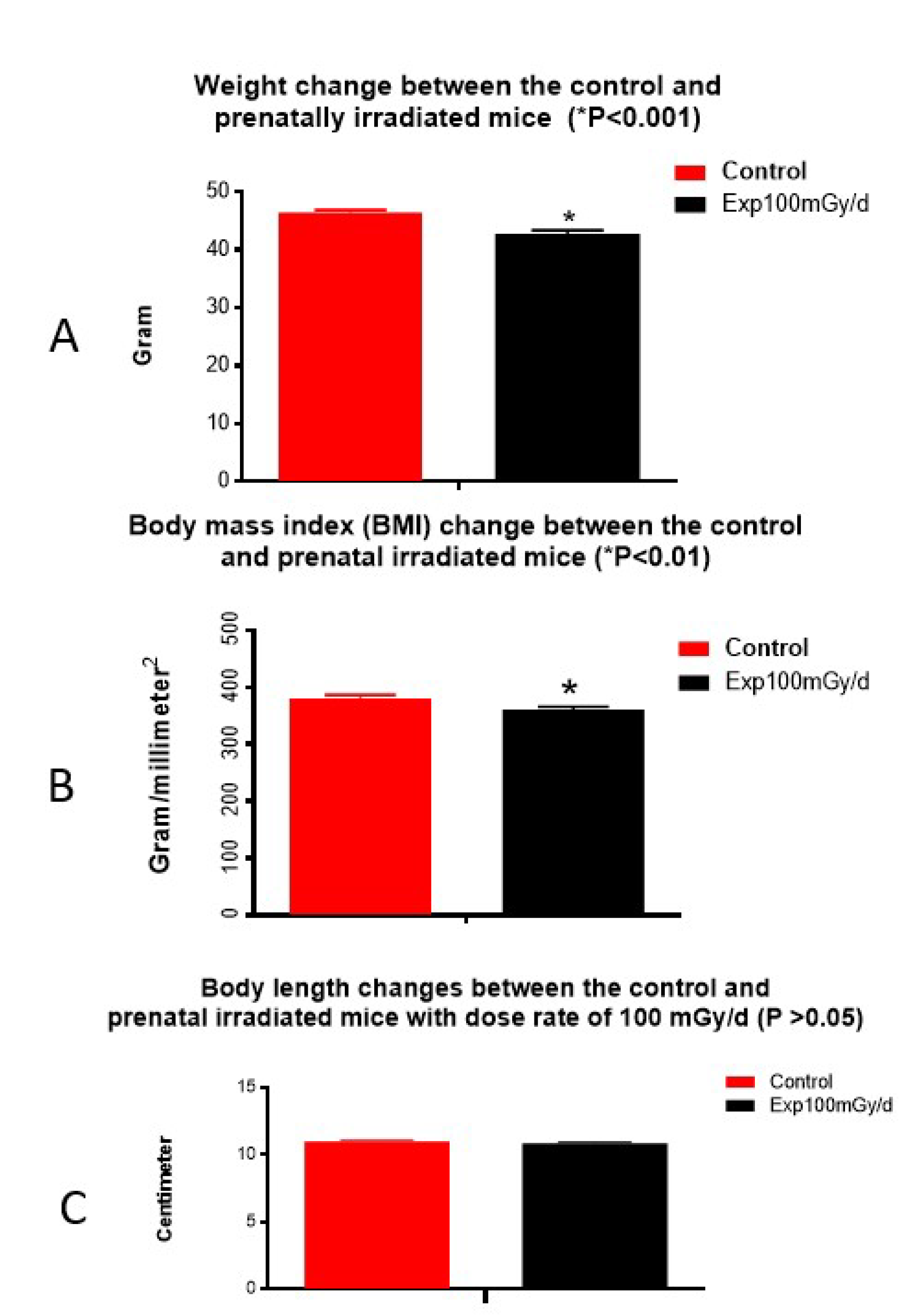
Figure 2.
Open field test: The distance irradiated animals traveled in each center, peripheral, and corner area is reduced (A) (P<0.01), but the time spent is similar (B) between the control and the prenatally irradiated mice. Center-C: the distance control animal traveled in the center area, Center-E: the distance experimental or irradiated (100mGy/d) animal traveled in the center area, Corner-C: the distance control animal traveled in the corner area, Corner-E: the distance irradiated animal traveled in the corner area, Peripheral-C: the distance control animal traveled in the peripheral area, Peripheral-E: the distance irradiated animal traveled in the peripheral area,.
Figure 2.
Open field test: The distance irradiated animals traveled in each center, peripheral, and corner area is reduced (A) (P<0.01), but the time spent is similar (B) between the control and the prenatally irradiated mice. Center-C: the distance control animal traveled in the center area, Center-E: the distance experimental or irradiated (100mGy/d) animal traveled in the center area, Corner-C: the distance control animal traveled in the corner area, Corner-E: the distance irradiated animal traveled in the corner area, Peripheral-C: the distance control animal traveled in the peripheral area, Peripheral-E: the distance irradiated animal traveled in the peripheral area,.
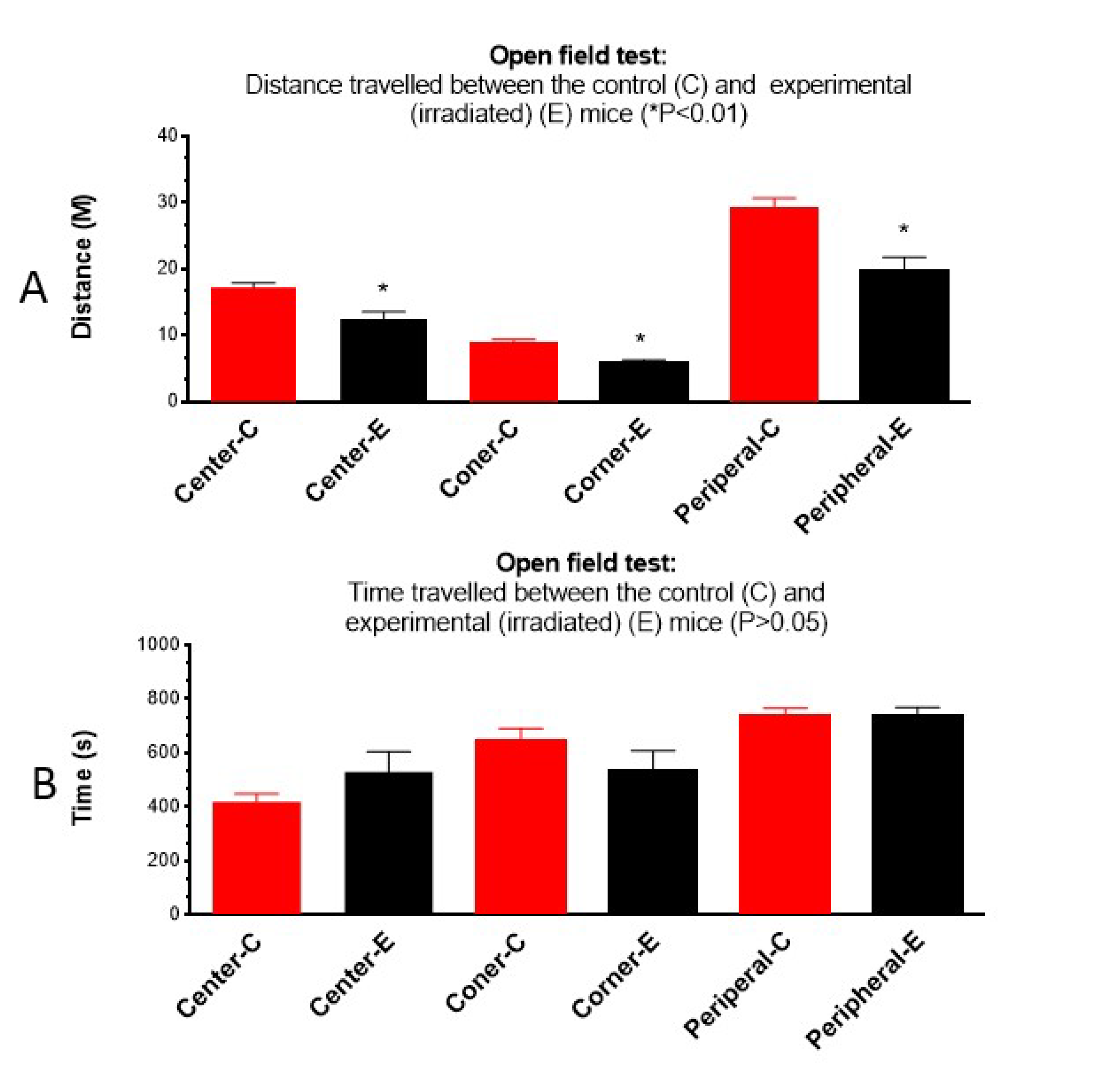
Figure 3.
Novel object recognition (A), forced swim (B) and tail suspension (C)tests: In A, both control and prenatally irradiated mice spend more time with novel object. In B and C, there is no difference in immobile time, and in B, no difference in swimming time is observed between the control and prenatally irradiated mice.
Figure 3.
Novel object recognition (A), forced swim (B) and tail suspension (C)tests: In A, both control and prenatally irradiated mice spend more time with novel object. In B and C, there is no difference in immobile time, and in B, no difference in swimming time is observed between the control and prenatally irradiated mice.
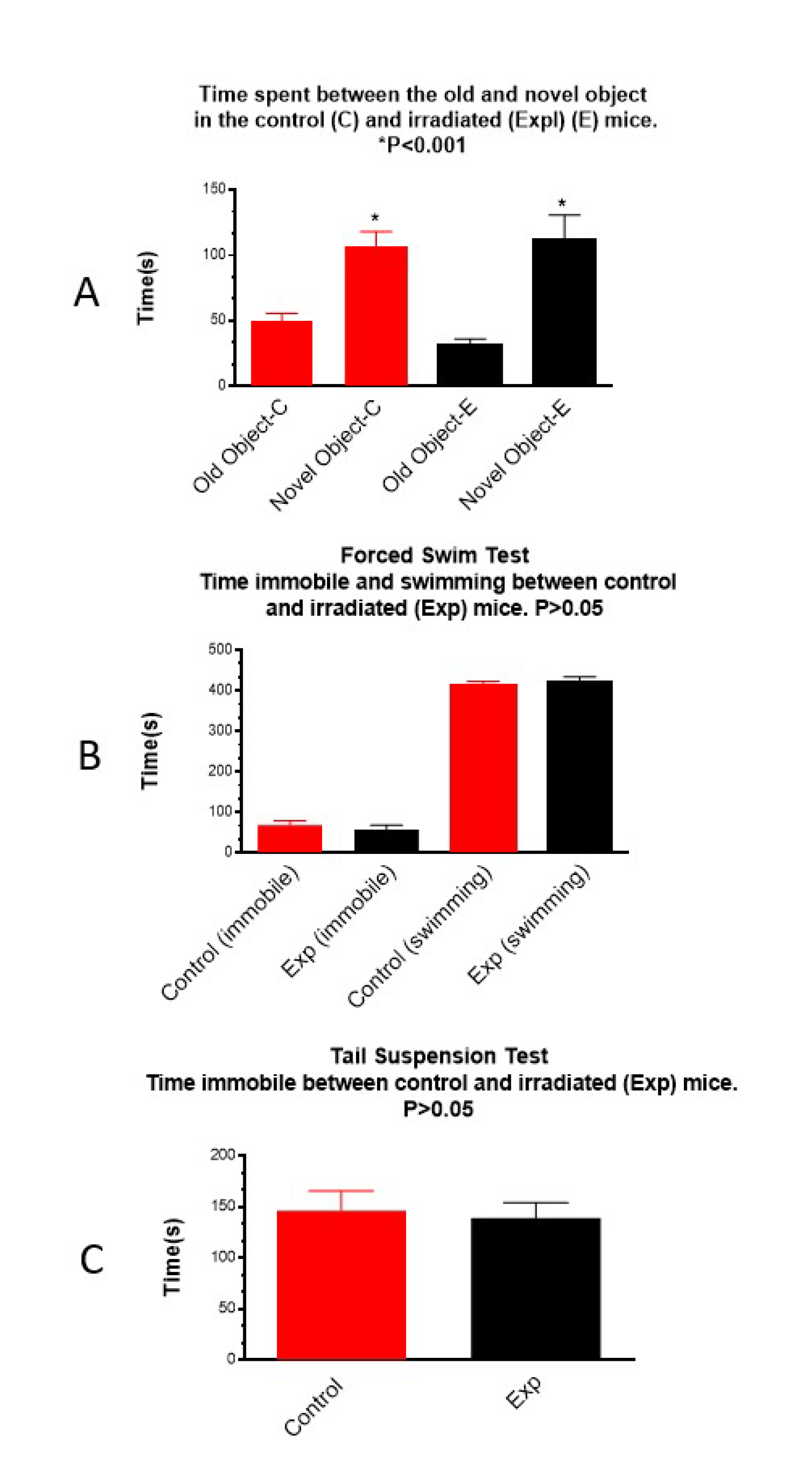
Figure 4.
Body fat (A) and organ (B) weight changes: Irradiation does not affect fat deposit (A), but organ weight reduction occurs in the testis, kidney, heart, epididymides (P<0.01), and liver (P<0.05)(B).
Figure 4.
Body fat (A) and organ (B) weight changes: Irradiation does not affect fat deposit (A), but organ weight reduction occurs in the testis, kidney, heart, epididymides (P<0.01), and liver (P<0.05)(B).
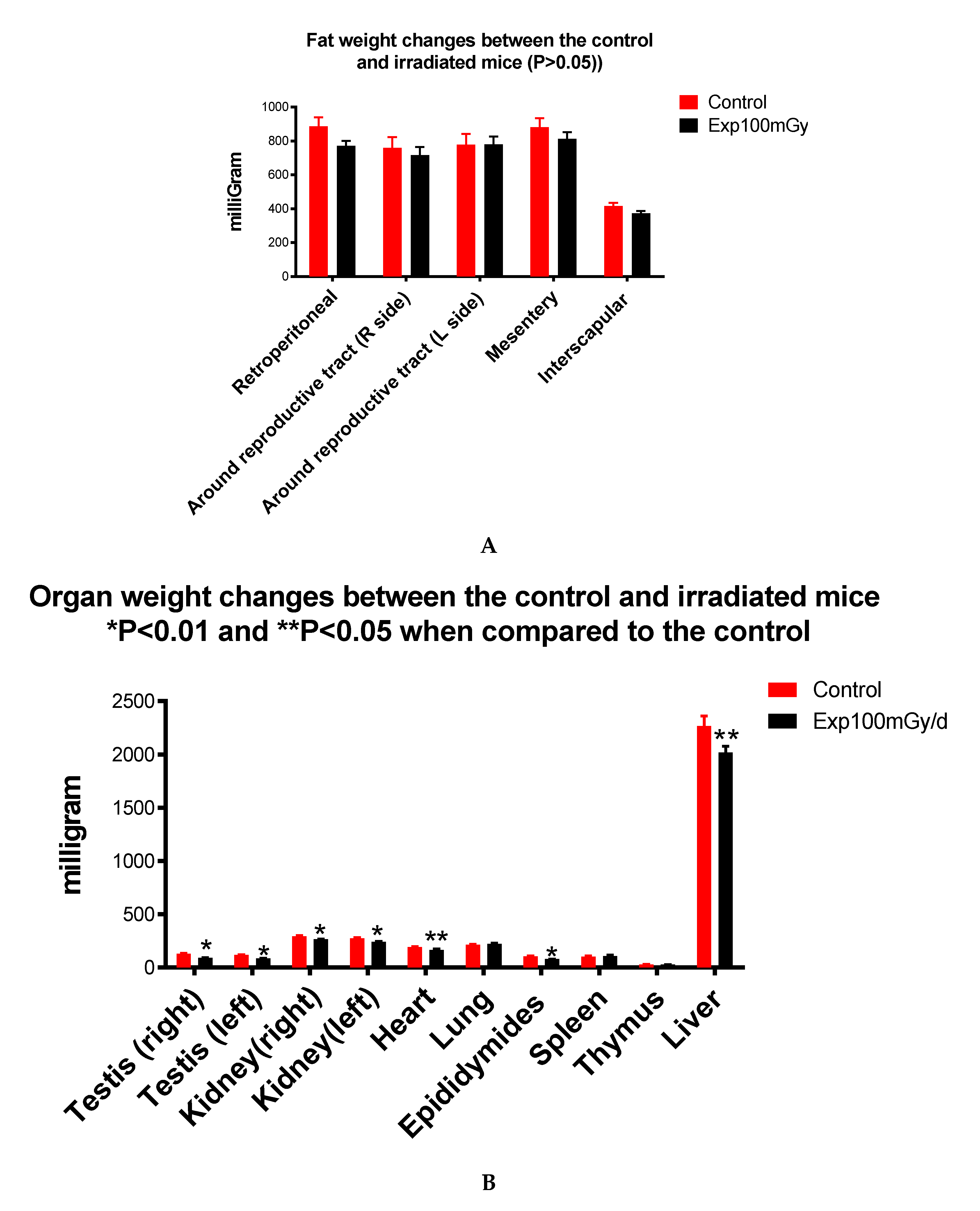
Figure 5.
Immunohistochemical staining: There is no obvious change in the number of NeuN immunopositive mature neurons (arrow) in the hilus (A, B, C, D, E), DCX immunopositive immature neurons (arrow) (A1, B1, C1, D1, E1) in the subgranular zone, IBA1 immunopositive microglia (arrow) in the hilus and the granule cell layer (GC) (A2, B2, C2, D2, E2), GFAP immunopositive astrocytes (arrow) (A3, B3, C3, D3, E3) and PDGFRα immunopositive oligodendrocyte precursor cells (arrow) (A4, B4, C4, D4, E4) among the control and experiment mice irradiated with 1, 20 and 100 mGy/d respectively. γ -H2AX immunostaining shows DNA damage foci occasionally in the granule cells (insert in C5). GC: granule cell layer, ML: molecular layer. Scale bar =100µm in A applies to A1-A4, B-B4, C-C4, D-D4, Scale bar =50µm in A5 applies to B5-D5.
Figure 5.
Immunohistochemical staining: There is no obvious change in the number of NeuN immunopositive mature neurons (arrow) in the hilus (A, B, C, D, E), DCX immunopositive immature neurons (arrow) (A1, B1, C1, D1, E1) in the subgranular zone, IBA1 immunopositive microglia (arrow) in the hilus and the granule cell layer (GC) (A2, B2, C2, D2, E2), GFAP immunopositive astrocytes (arrow) (A3, B3, C3, D3, E3) and PDGFRα immunopositive oligodendrocyte precursor cells (arrow) (A4, B4, C4, D4, E4) among the control and experiment mice irradiated with 1, 20 and 100 mGy/d respectively. γ -H2AX immunostaining shows DNA damage foci occasionally in the granule cells (insert in C5). GC: granule cell layer, ML: molecular layer. Scale bar =100µm in A applies to A1-A4, B-B4, C-C4, D-D4, Scale bar =50µm in A5 applies to B5-D5.
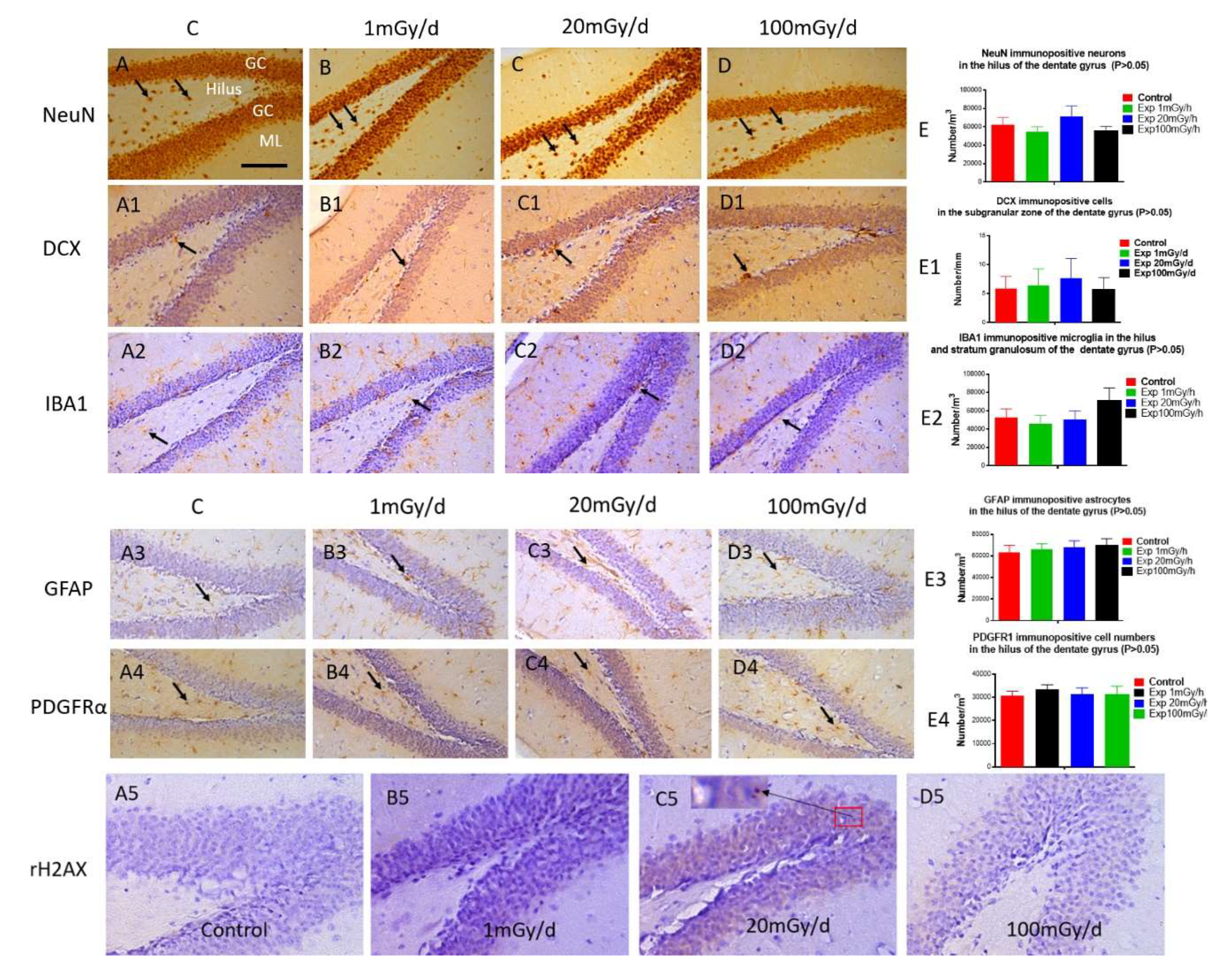
Figure 6.
Venn diagram and heatmap of mRNA sequencing results in the hippocampus and blood samples. A. Venn diagram of differentially expressed mRNA between hippocampus and blood; B and C. Heatmap of selected mRNAs which are differentially expressed in hippocampus and blood.
Figure 6.
Venn diagram and heatmap of mRNA sequencing results in the hippocampus and blood samples. A. Venn diagram of differentially expressed mRNA between hippocampus and blood; B and C. Heatmap of selected mRNAs which are differentially expressed in hippocampus and blood.
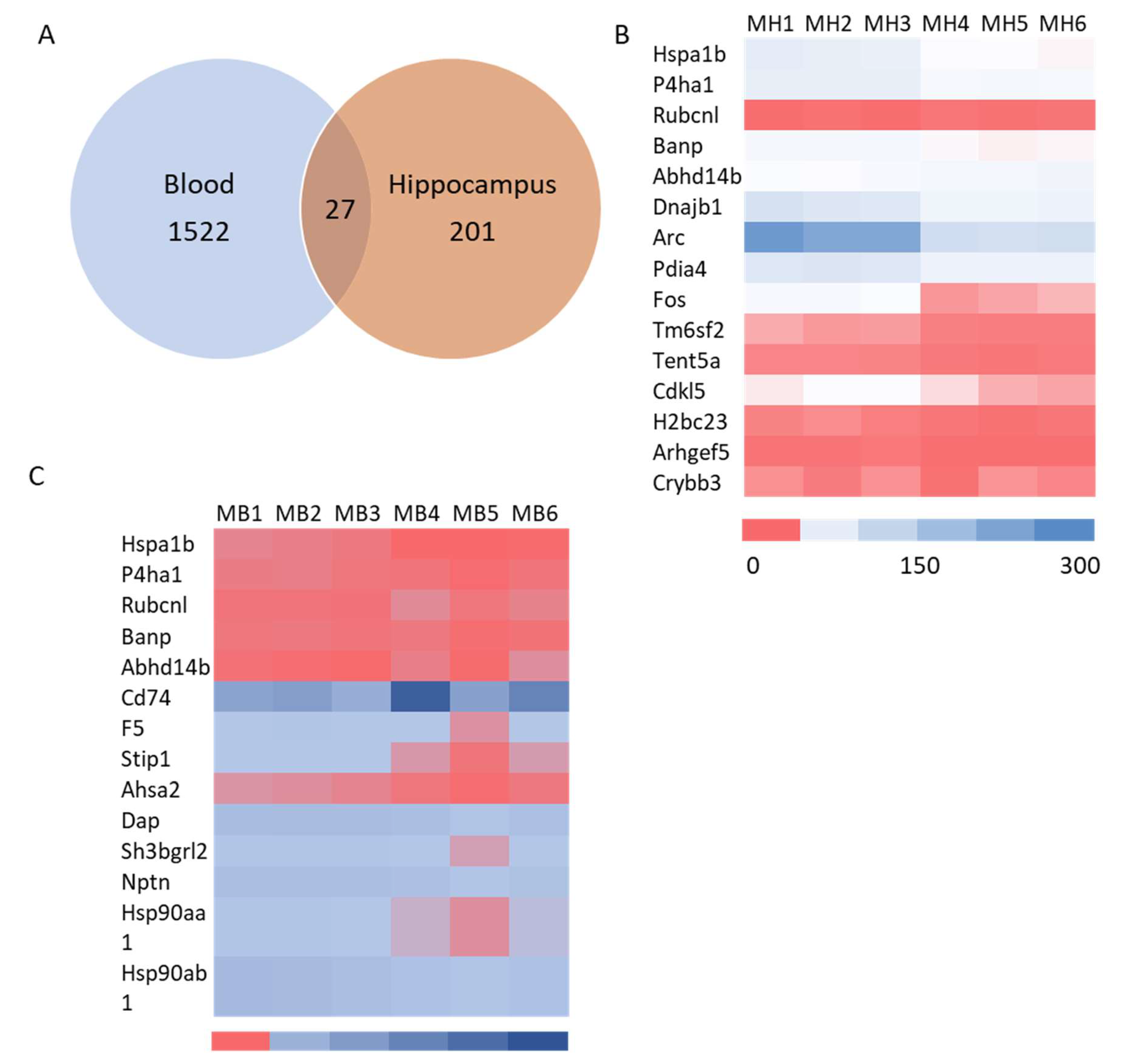
Figure 7.
qRT-PCR indicates a down-regulation of Hspa1b, P4ha1, Banp, and an up-regulation of Rubanl and Abhd14b in both hippocampus (A) and whole blood (B) (P<0.05). In the hippocampus, there is a significant down-regulation of Dnajb1, Arc, Pdia4, Fos, Tm6sf2, Tent5, Cdkl5, H2bc23, Arhgef5, Crybb3 (C), whereas CD74 is up-regulated, F5, Stip1, Ahsa2, Dap, Sh3bgrl2, Nptn, Hsp90aa1and Hsp90ab1 genes are down-regulated in whole blood (D). * p < 0.05.
Figure 7.
qRT-PCR indicates a down-regulation of Hspa1b, P4ha1, Banp, and an up-regulation of Rubanl and Abhd14b in both hippocampus (A) and whole blood (B) (P<0.05). In the hippocampus, there is a significant down-regulation of Dnajb1, Arc, Pdia4, Fos, Tm6sf2, Tent5, Cdkl5, H2bc23, Arhgef5, Crybb3 (C), whereas CD74 is up-regulated, F5, Stip1, Ahsa2, Dap, Sh3bgrl2, Nptn, Hsp90aa1and Hsp90ab1 genes are down-regulated in whole blood (D). * p < 0.05.
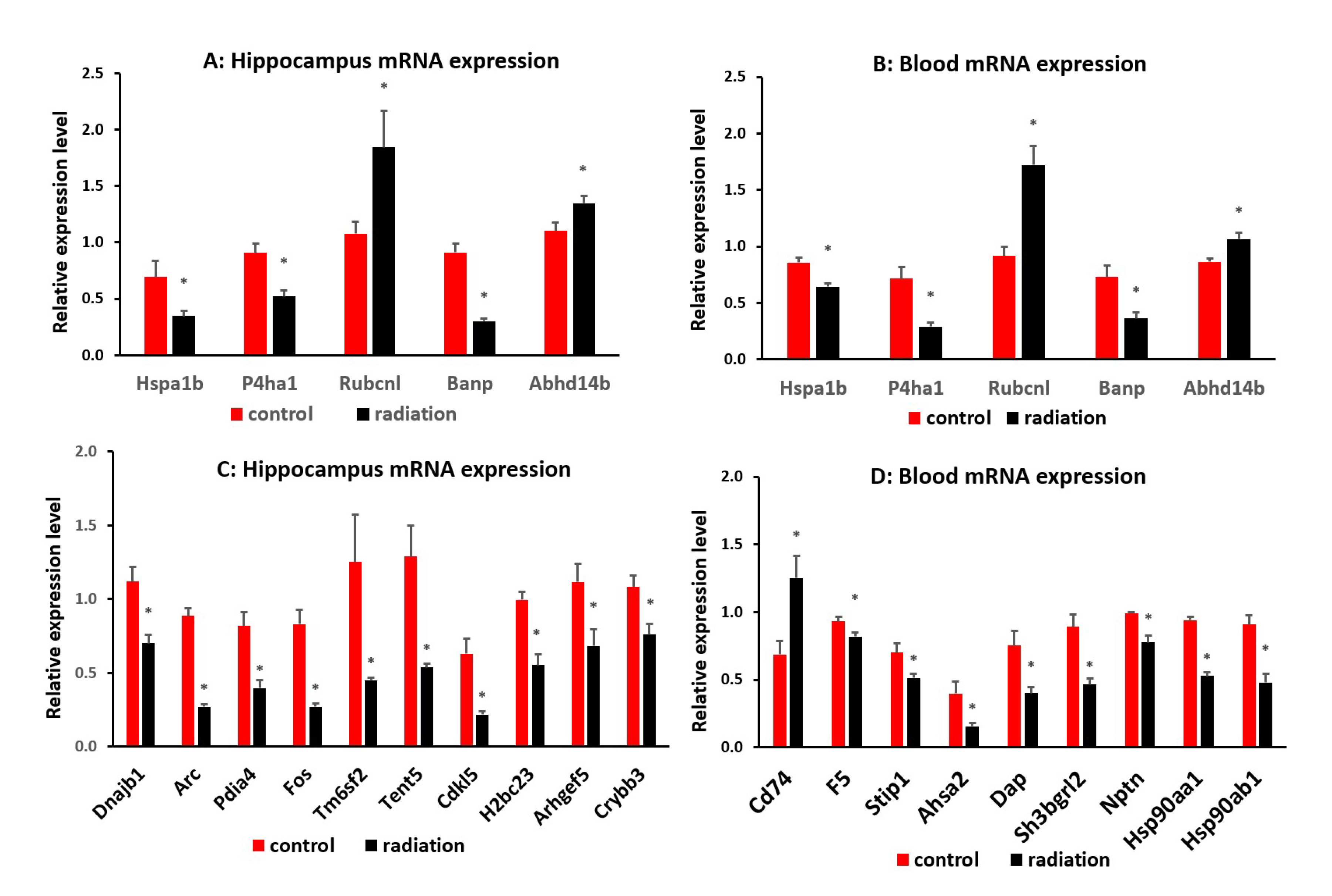
Figure 8.
Heatmap of miRNA sequencing results and qRT-PCR indicates a down-regulation of miR-448-3p and miR-1298-5p in the hippocampus (A&B) and miR-320-3p, miR-423-5p, miR-486b-5p, miR-486b-3p, miR-423-3p, miR-652-3p, miR-324-3p, miR-181b-5p, miR-let-7b and miR-6904-5p in whole blood (C&D). * p < 0.05.
Figure 8.
Heatmap of miRNA sequencing results and qRT-PCR indicates a down-regulation of miR-448-3p and miR-1298-5p in the hippocampus (A&B) and miR-320-3p, miR-423-5p, miR-486b-5p, miR-486b-3p, miR-423-3p, miR-652-3p, miR-324-3p, miR-181b-5p, miR-let-7b and miR-6904-5p in whole blood (C&D). * p < 0.05.
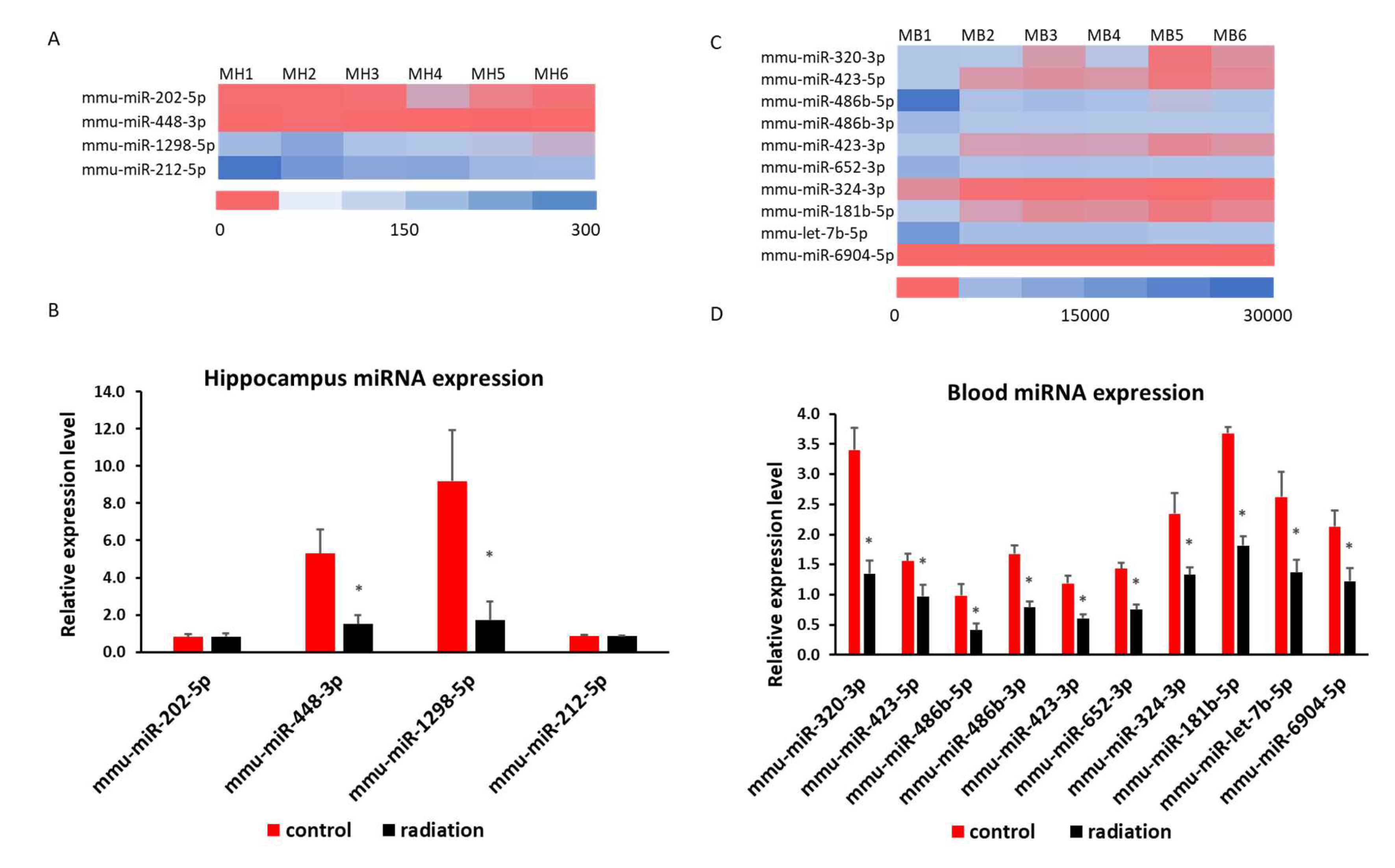
Table 1.
miRNA sequences for qRT-PCR.
| miRNA primers | Sequence |
|---|---|
| mmu-miR-68 | GCTGTACTGACTTGATGAAAGTAC |
| mmu-miR-202-5p | GCTTCCTATGCATATACTTCTTT |
| mmu-miR-448-3p | TTGCATATGTAGGATGTCCCAT |
| mmu-miR-1298-5p | TTCATTCGGCTGTCCAGATGTA |
| mmu-miR-212-5p | ACCTTGGCTCTAGACTGCTTACT |
| mmu-miR-320-3p | AAAAGCTGGGTTGAGAGGGCGA |
| mmu-miR-423-5p | TGAGGGGCAGAGAGCGAGACTTT |
| mmu-miR-486b-5p | TCCTGTACTGAGCTGCCCCGAG |
| mmu-miR-486b-3p | CGGGGCAGCTCAGTACAGGA |
| mmu-miR-423-3p | AGCTCGGTCTGAGGCCCCTCAGT |
| mmu-miR-652-3p | AATGGCGCCACTAGGGTTGTG |
| mmu-miR-324-3p | ATT CCA CTG CCC CAG GTG CTG CT |
| mmu-miR-122-5p | TGGAGTGTGACAATGGTGTTTG |
| mmu-miR-744-5p | TGCGGGGCTAGGGCTAACAGCA |
| mmu-miR-181b-5p | AACATTCATTGCTGTCGGTGGGTT |
| mmu-let-7b-5p | TGAGGTAGTAGGTTGTGTGGTT |
| mmu-miR-6904-5p | TCCTGGGGTTAGAGTTGAGTGG |
Table 2.
mRNA sequences for qRT-PCR.
| mRNA primers | Sequence | Direction |
|---|---|---|
| Dnajb1 F | CCAATGGGTATGGGTGGCTT | forward |
| Dnajb1 R | GCCTTCTCCAGGGACTTTCC | reverse |
| Arc F | GGAGGGAGGTCTTCTACCGT | forward |
| Arc R | TCCTCCTCAGCGTCCACATA | reverse |
| Pdia4 F | GGCCTCTTGGATGTGAATGC | forward |
| Pdia4 R | CAGGGCTGAAAGTGTGGTGA | reverse |
| Fos F | AGTCAAGGCCTGGTCTGTGT | forward |
| Fos R | TGGAACACGCTATTGCCAGG | reverse |
| Tm6sf2 F | TTCTCACACATGGGTGCCTC | forward |
| Tm6sf2 R | CTTGGTCCTGTGGCGAAGAT | reverse |
| Tent5a F | CTCCAGGACTGACCAAGGC | forward |
| Tent5a R | CGGACACCTATGCCCTTCTC | reverse |
| Cdkl5 F | AACGGCGAGAATCCAAGCAT | forward |
| Cdkl5 R | AAGGCGTTTGTTGGTCACTGT | reverse |
| H2bc23 F | TACAACAAGCGCTCGACCAT | forward |
| H2bc23 R | TGTCACTGAACACGTGCCTT | reverse |
| Hspb1 F | ATAGAGACCTGAAGCACCGC | forward |
| Hspb1 R | CGGTCATGTTCTTGGCTGGT | reverse |
| Crybb3 F | AAGCAGGTCTCTGCCTCCT | forward |
| Crybb3 R | TACGATCTCCATCTTGCGCC | reverse |
| Vmn1r58 F | GGTCAAAACACGGCCAAACC | forward |
| Vmn1r58 R | AGGAGAAACAGCCTTCTCTCAA | reverse |
| Scd1 F | GAGTAGCTGAGCTTTGGGCT | forward |
| Scd1 R | ACTTCATCAGCGGGGACTTG | reverse |
| Cd59b F | CTGTTGCCTTGGATCAGCCT | forward |
| Cd59b R | TGATACACTTGCCTTCCGGC | reverse |
| Stip1 F | GTGTTCAACCAGTGAGCAGG | forward |
| Stip1 R | CAGGTCTGACGGCTTGTTCT | reverse |
| Ahsa2 F | GACCAACGTGAACAACTGGC | forward |
| Ahsa2 R | CGTCTTGAGTGCCTTCAGGT | reverse |
| Dnaja1 F | GGCTCGGCTACAAAAGAGGT | forward |
| Dnaja1 R | ATGCGTTCTCCATGACCCTG | reverse |
| Dap F | TCCCTAAAGGGTCGTTGAACC | forward |
| Dap R | AGGAGCCCATCCCTCCTTAG | reverse |
| F5 F | GCTTGCCTTCTCAAGCGTTC | forward |
| F5 R | CCCAAGTGACTTTGCGTGTG | reverse |
| Hspa1b F | GGCACCGATTACTGTCAAGG | forward |
| Hspa1b R | ACAGTGCCAAGACGTTTGTT | reverse |
| P4ha1 F | AAGACTGTTCTGCCGCTACC | forward |
| P4ha1 R | TTCGTAGCCAGACAGCCAAG | reverse |
| Rubcnl F | AGGTGATCCGAACCTGTCG | forward |
| Rubcnl R | TCCGAGCATCACCTACGCC | reverse |
| Banp F | AACACCACGAGAATTCCGCA | forward |
| Banp R | GCACTTTGTTGCAGGTCTGG | reverse |
| Abhd14b F | TAGCACACGCCATTCTCCTG | forward |
| Abhd14b R | AAGGTATCCACCACAGCAGC | reverse |
| GAPDH F | ACC ACA GTC CAT GCC ATC AC | forward |
| GAPDH R | TCC ACC ACC CTG TTG CTG TA | reverse |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



