
Submitted:
30 July 2024
Posted:
02 August 2024
You are already at the latest version
Abstract
Infection with the Chikungunya virus (CHIKV) manifests as a febrile illness known as chikungunya fever (CHIKF). CHIKF is characterized by sudden-onset high fever, rash, polyarthritis, and polyarthralgia. Alt-hough the infection typically resolves within two weeks, many patients experience recurrent joint pain and inflammation that can outlast the other symptoms for many months. CHIKF has been previously linked to rheumatoid arthritis (RA) due to similar inflammatory processes. This study aimed to identify molecular markers in the blood of CHIKV-infected individuals associated with joint pain and chronicity of CHIKF. Sequencing of B and T cell receptors (BCR and TCR) revealed that CHIKV infection reduces CDR3 diver-sity. Lower BCR diversity was linked to increased expression of genes involved in osteoclast differentiation and activation through RANK/RANKL signaling. If osteoclast involvement in CHIKF pathogenesis is con-firmed, existing therapeutic approaches could be used as alternative treatments for CHIKF patients.
Keywords:
Chikungunya fever
; Rheumatoid arthritis
; RANK signaling
; CDR3
; B cells
; T cells
1. Introduction
The Chikungunya virus (CHIKV), transmitted to humans by Aedes mosquitoes, causes a febrile illness known as chikungunya fever (CHIKF). Originally from East Africa, CHIKF is now present in more than 60 countries in four continents. The infection commonly resolves in two weeks. The acute phase of CHIKF typically lasts around one week and is characterized by sudden onset of fever, polyarthralgia, skin rash, and myalgia [1]. Mortality rates are no larger than 0.8%, but CHIKF can progress to a chronic state characterized by persistent joint pain and inflammation that can last for months, leading to mobility problems and decreased quality of life [2]. These chronic symptoms are associated with large economic impacts, especially in third world countries [1].
CHIKF has been previously linked to rheumatoid arthritis (RA) due to similar inflammatory processes. CHIKV infection of fibroblast-like synoviocytes induces the expression of chemotactic agents, including RANKL, IL-6, IL-8, and MCP-1, which recruit and differentiate phagocytes into osteoclast-like cells. These cells produce TNF-α and IL-6, the primary mediators of arthritis [3]. Osteoclasts play a crucial role in bone erosion in RA, and there is evidence of increased osteoclast precursor abundance and activity in peripheral blood cells of RA patients [4]. The balance between osteoclast and osteoblast activity determines the degree of bone erosion in RA [5]. CHIKV infection of osteoblasts increases RANKL and IL-6 production while decreasing osteoprotegerin (OPG) production, promoting osteoclastogenesis [5].
Given the immune response links between CHIKF and RA, this study aimed to analyze publicly available RNAseq data in order to identify immune-related blood markers in CHIKV-infected individuals that are shared with chronic RA patients and could potentially explain the underlying causes of joint pain and chronicity in CHIKF. The discovery of such common biological processes could open the possibility for new and improved treatments for chronic joint pain in CHIKF patients.
2. Materials and Methods
2.1. RNA-Seq Data
Two publicly available RNA-Seq datasets were utilized for the analyses. The Chikungunya dataset, as detailed in reference [6], includes 24 CHIKV-infected patients, all in the acute phase of the disease, with symptom onset occurring 0 to 4 days before sample collection, and 13 non-infected patients. Real-time reverse transcription polymerase chain reaction (RT-PCR) was performed to test for CHIKV. Peripheral blood RNA extraction and sequencing using HiSeq Illumina 1500 are described in the original article [6].
The Dengue dataset, described in reference [7] and used as a comparative control, consists of 18 DENV-infected individuals and 16 non-infected individuals. Acute dengue was confirmed using IgG ELISA, virus isolation, and RT-PCR. Peripheral blood RNA extraction and sequencing using Illumina NovaSeq 6000 are detailed in the original article [7].
Both datasets include individuals aged 15 to 72 years, and sample quality control was performed for all samples. Outliers were excluded during the estimation of BCR and TCR CDR3 diversity, as detailed below. A summary of the datasets and samples is presented in Figure 1A, Tables S2 and S3.
2.2. Differential Gene Expression Analysis
To conduct the analysis of differential gene expression, initial normalization of gene expression values was performed using the DESeq2 R package (version 1.38.2) [8], wherein counts per million reads mapped (CPM) were utilized. For the Chikungunya dataset, CPM values were directly provided by the authors. In the case of the Dengue dataset, values were computed subsequent to mapping the sequences to the human genome (GRCh38.p13 release 104) employing STAR software (version 2.7.10a) [9]. Following alignment, the reads were summarized into count matrices utilizing the R package Rsubread (version 2.0.1) [10]. Differential expression analysis was then conducted using the R package DESeq2 (version 1.38.2) to identify genes exhibiting differential expression. The analysis is summarized in Figure 1B.
2.3. Cell Abundance, CDR3 Diversity and Gene Set Enrichment Analysis
The Immune Cell Abundance Identifier (ImmuCellAI) tool [11] was employed to infer the abundance of immune system cells from counts per million (CPM) values. Sequences of the heavy chain complementary-determining region 3 (CDR3), representing the entire functional Ig heavy chain (IgH) repertoire, were acquired for T cell receptors (TCR) and B cell receptors (BCR) using MiXCR software (version 3.0.13) [12]. TCR encompass TCRα (TRA) and TCRβ (TRB), while BCR include heavy (IGH), kappa (IGK), and lambda (IGL) chains.
Diversity analysis of CDR3 sequences for each chain was conducted utilizing the immunarch R package (version 0.9.0) [13], with Chao1 as the diversity estimator. Pearson correlation coefficients were computed to evaluate the association between variance stabilizing transformation count data (VST) and CDR3 diversity values, using by the Hmisc R package (version 4.5-0) [14]. These coefficients were subsequently utilized in Gene Set Enrichment Analysis (GSEA) using the fgsea R package (version 1.24.0) [15], with gene modules sourced from the Molecular Signatures Database [16]. The normalized enrichment score (NES) from GSEA is calculated by normalizing the enrichment score (ES) for each gene set to account for differences in gene set size. This normalization allows for comparison across gene sets of different sizes. Specifically, NES is determined by dividing the actual ES by the mean of the ESs obtained from all permutations of the dataset, as described by Subramanian et al. (2005)[17].
3. Results
3.1. CDR3 Diversity and Cell Abundance
During the acute phase of CHIKV infection, individuals showed reduced diversity of BCR and TCR CDR3 compared to healthy individuals (Figure 1C and Figure S1). In contrast, individuals infected with DENV exhibited decreased diversity of TCR only (Figure 1C and Figure S2). Similarly, the abundance of T cells in the blood was lower in both CHIKV and DENV-infected patients, while the abundance of B cells was lower only in CHIKV-infected patients (Figure 1D and Figure S3).
3.2. Gene Set Enrichment Analysis
Gene Set Enrichment Analysis (GSEA) analysis identified gene sets that were correlated with CDR3 diversity. Some of these pathways were related to osteoclast differentiation and activation, negatively correlated with CDR3 diversity, and enriched primarily in the Chikungunya dataset, as shown in Figure 2.
3.3. Transcriptomic Changes in Peripheral Blood during CHIKV Infection
Genes associated with osteoclast differentiation and activation were up-regulated in CHIKV-infected individuals compared to non-infected individuals, as shown in Figure 3A,B. The expression of these genes is consistently increased in CHIKV-infected individuals, but not in Dengue-infected individuals (see Table S1).
4. Discussion
During the acute phase of CHIKV infection, patients showed lower CDR3 diversity (Figure 1C, Figure S1). This is a common observation in infections due to the expansion of lymphocytes after antigen contact [18]. Lymphopenia, a characteristic of the acute phase of CHIKF [19], may explain the lower diversity observed in CHIKV-infected. Lower frequencies of B and T cells (Figure 1D, Figure S3) were also observed in CHIKV-infected, which could lead to a lower number of BCR and TCR reads and underestimate the diversity [20].
GSEA analysis revealed a negative correlation between CDR3 diversity and the expression of genes associated with osteoclast differentiation and activation pathways in CHIKV-infected individuals. This suggests that these genes are more highly expressed during the acute phase of infection (Figure 2).
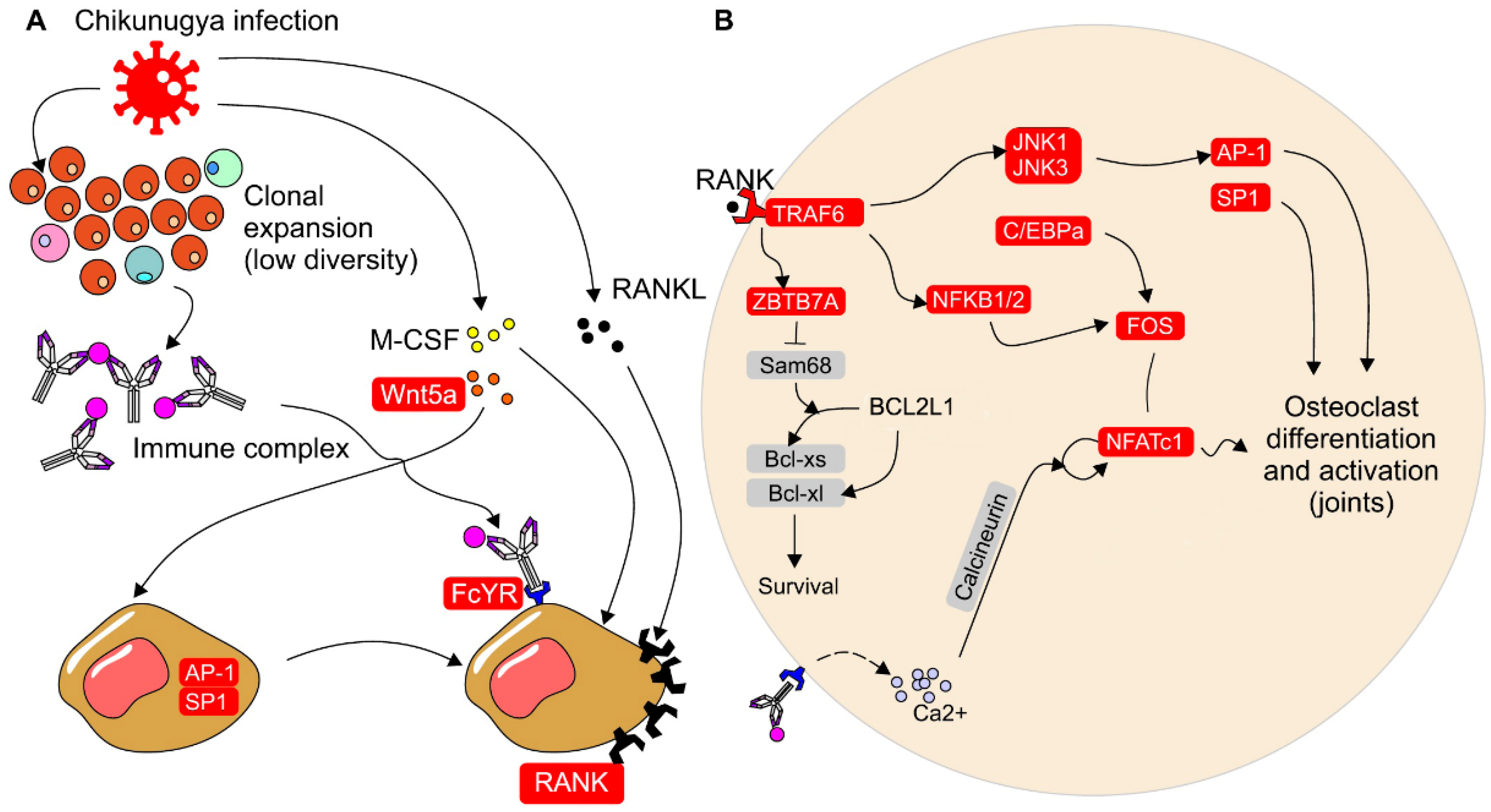
Individuals infected with CHIKV show up-regulation of receptor activator of nuclear factor-κB (RANK), macrophage-colony stimulating factor (M-CSF), and FcyR, which are involved in the induction of osteoclast differentiation and activation [21] (Figure 3, Supplementary Table S1). Additionally, WNT5A expression is up-regulated, and has been demonstrated to stimulate the overexpression of RANK in osteoclast precursor cells through JNK, SP-1, and AP-1 signaling [22]. The activation of RANK and its ligand RANKL is crucial for osteoclast differentiation, as it leads to the expression of Nuclear Factor of Activated T Cells 1 (NFATC1) in precursor cells [23]. Upon RANK stimulation, intracellular signaling is initiated, involving the recruitment of TRAF6 and activation of the MAP kinases (MAPKs) and NF-κB pathways. The MAPKs pathway induces the expression of Ap-1, while the NF-κB pathway induces the expression of FOS [24]. Signaling through RANK also promotes the expression of C/EBPα (CCAAT/enhancer binding protein α) in osteoclast precursors [25]. Collectively, Ap-1, FOS, and C/EBPα facilitate the expression of NFATc1, which in turn stimulates the expression of genes responsible for osteoclast differentiation [21]. Activation of FCyR can also increase intracellular calcium concentration, further promoting NFATc1 expression [21]. In CHIKV-infected individuals, not only are the expression levels of RANK and NFATC1 up-regulated, but also the other genes involved in these pathways (Figure 3, Supplementary Table S1).
The literature on RA combined with the results produced by our own analyses leads to the hypothesis that the chronicity of CHIKV infection may be linked to a higher number of circulating osteoclast precursors during the acute phase of infection, which are more resistant to apoptosis. These are observed in RA [26,27], and could be driven by Ig and T cells through a persistent ligand [28]. These resilient osteoclasts can migrate to the joints and remain activated, leading to chronic joint inflammation. RANK signaling induces the expression of ZBTB7A in osteoclast precursors, which in turn promotes the overexpression of NFATC1 and favors the alternative splicing of the BCL2L1 gene into Bcl-xl, an anti-apoptotic protein [29]. Xu et al. (2022) demonstrated that ZBTB7A overexpression leads to the degradation of Sam68, which favors alternative Bcl-xl splicing and longer osteoclast survival. Moreover, ZBTB7A expression is up-regulated by cytokines in inflammatory conditions. ZBTB7A expression is also increased in CHIKV-infected individuals during the acute phase of infection (Figure 3B, Supplementary Table S1).
Because there is little to no bone erosion associated with CHIKF [30], the mechanism that generates chronic joint pain must be distinct from RA. Osteoclasts activity is also seen in viral infections not associated with erosive arthritic symptoms [31,32]. Additionally, osteoclasts perform activities not related to bone erosion [33]. Lower CDR3 diversity and higher expression of genes involved in osteoclastogenesis may be linked to persistent presence of antigen. Osteoclasts are activated by FcγR through IgG antibodies bound to their antigens in the form of immunocomplexes, which promotes their differentiation and activation [34,35]. In acute CHIKF, lymphocytes may undergo clonal expansion after contact with CHIKV antigens, followed by antibody production and larger pools of monoclonal T cells. Antibodies activate FcγR signaling in osteoclast precursors, enhancing osteoclastogenesis. Due to the nature of CHIKV infecting several cell types sequentially, including macrophages and synovial cells, the inflammatory response can become sustained. This is also furthered by T cell cytokine mediated recruitment of host cells to the synovial tissues, sustaining ligands and promoting sustained B cell antibody production at high circulating levels [36]. Ultimately these factors can contribute to joint pain and the development of the chronic phase of CHIKF.
The findings presented in this study stem from analyzing the transcriptome of peripheral blood mononuclear cells (PMBCs) from patients infected with CHIKV. Consequently, discriminating the specific cell types responsible for the observed changes in gene expression poses a challenge, particularly for genes expressed across various cell types. To delve into this further, investigating the presence and prevalence of osteoclast precursors, along with their expression patterns during CHIKF, requires both in vivo and in vitro investigations. These investigations would include techniques such as flow cytometry, cell sorting, single-cell sequencing, and assays focused on osteoclastogenesis. Subsequent studies will facilitate a comprehensive understanding of the molecular disruptions induced by CHIKF.
If the hypothesis suggesting osteoclast involvement in CHIKF proves correct, existing interventions utilized for other conditions might offer potential benefits for both the acute and chronic phases of CHIKF. One such intervention is the monoclonal antibody denosumab. It works by binding to RANKL, thereby preventing its interaction with RANK, which halts the process of osteoclast differentiation and activation [37]. Furthermore, if immune complex formation plays a role in osteoclast activation during CHIKF, a possible treatment avenue involves reducing FcγR signaling by promoting IgG sialylation [8]. Lastly, inducing osteoclast apoptosis, a strategy already successful in treating arthritis, could also serve as a viable option for managing chronic joint pain associated with CHIKF [38].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: CDR3 diversity is lower in CHIKV-infected samples; Figure S2: T cell CDR3 diversity is lower in DENV-infected samples; Table S1: Differentially expressed genes in CHIKV- and DENV-infected samples.
Author Contributions
Conceptualization, H.N., A.U., A.N.; methodology, A.U.; validation, A.U.; formal analysis, A.U., V.M. and A.C-M.; investigation, A.U.; resources, H.N.; data curation, A.U.; writing—original draft preparation, A.U.; writing—review and editing, F.M., A.N. and H.N.; visualization, A.U., H.N. and F.M.; supervision, H.N.; project administration, A.U.; funding acquisition, H.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Finance Code 001).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used here can be downloaded from the following links: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA507472 and https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8578928/
Acknowledgments
The authors would like to thank Andreas Suhrbier and two anonymous reviewers for helpful comments on the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Manzoor, K.N.; Javed, F.; Ejaz, M.; Ali, M.; Mujaddadi, N.; Khan, A.A.; Khattak, A.A.; Zaib, A.; Ahmad, I.; Saeed, W.K.; et al. The global emergence of Chikungunya infection: An integrated view. Rev. Med Virol. 2021, 32, e2287. [Google Scholar] [CrossRef]
- Hoarau, J.-J.; Bandjee, M.-C.J.; Trotot, P.K.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T.; et al. Persistent Chronic Inflammation and Infection by Chikungunya Arthritogenic Alphavirus in Spite of a Robust Host Immune Response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef] [PubMed]
- Phuklia, W.; Kasisith, J.; Modhiran, N.; Rodpai, E.; Thannagith, M.; Thongsakulprasert, T.; Smith, D.R.; Ubol, S. Osteoclastogenesis induced by CHIKV-infected fibroblast-like synoviocytes: A possible interplay between synoviocytes and monocytes/macrophages in CHIKV-induced arthralgia/arthritis. Virus Res. 2013, 177, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Boire, G.; Komarova, S.; Dixon, S.; Sims, S.; Harrison, R.; Nabavi, N.; Maria, O.; Manolson, M.; Mizianty, M.; et al. The increased in vitro osteoclastogenesis in patients with rheumatoid arthritis is due to increased percentage of precursors and decreased apoptosis — The In Vitro Osteoclast Differentiation in Arthritis (IODA) study. Bone 2011, 48, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Noret, M.; Herrero, L.; Rulli, N.; Rolph, M.; Smith, P.N.; Li, R.W.; Roques, P.; Gras, G.; Mahalingam, S. Interleukin 6, RANKL, and Osteoprotegerin Expression by Chikungunya Virus-Infected Human Osteoblasts. J. Infect. Dis. 2012, 206, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Soares-Schanoski, A.; Cruz, N.B.; de Castro-Jorge, L.A.; de Carvalho, R.V.H.; da Rós, N.; Oliveira. ; Costa, D.D.; dos Santos, C.L.S.; Cunha, M.d.P.; Oliveira, M.L.S.; et al. Systems analysis of subjects acutely infected with the Chikungunya virus. PLOS Pathog. 2019, 15, e1007880. [Google Scholar] [CrossRef] [PubMed]
- Tsalik, E.L.; Fiorino, C.; Aqeel, A.; Liu, Y.; Henao, R.; Ko, E.R.; Burke, T.W.; Reller, M.E.; Bodinayake, C.K.; Nagahawatte, A.; et al. The Host Response to Viral Infections Reveals Common and Virus-Specific Signatures in the Peripheral Blood. Front. Immunol. 2021, 12, 741837. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 2014; 15(12),550.
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef]
- Miao, Y.R.; Zhang, Q.; Lei, Q.; Luo, M.; Xie, G.Y.; Wang, H.; Guo, A.Y. ImmuCellAI: A Unique Method for Comprehensive T-Cell Subsets Abundance Prediction and its Application in Cancer Immunotherapy. Adv. Sci. 2020, 7, 1902880. [Google Scholar] [CrossRef]
- A Bolotin, D.; Poslavsky, S.; Davydov, A.N.; E Frenkel, F.; Fanchi, L.; I Zolotareva, O.; Hemmers, S.; Putintseva, E.V.; Obraztsova, A.S.; Shugay, M.; et al. Antigen receptor repertoire profiling from RNA-seq data. Nat. Biotechnol. 2017, 35, 908–911. [Google Scholar] [CrossRef] [PubMed]
- Nazarov, V.; Tsvetkov, V.; Fiadziushchanka, S.; et al. immunarch: Bioinformatics Analysis of T-Cell and B-Cell Immune Repertoires. https://immunarch.com/. https://github.com/immunomind/immunarch. 2023.
- Harrell, F.E., Jr.; HMISC: Harrell Miscellaneous. R Package Version 4.1-1. 2018. Available online: http://CRAN.R-project.org/package=Hmisc (accessed on 21 March 2022).
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.N. Fast gene set enrichment analysis. bioRxiv. 2016. Available from: https://www.biorxiv.org/content/10. 1101. [Google Scholar]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular signatures database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Nikolich-Žugich, J.; Slifka, M.K.; Messaoudi, I. The many important facets of T-cell repertoire diversity. Nat. Rev. Immunol. 2004, 4, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Thiberville, S.-D.; Boisson, V.; Gaudart, J.; Simon, F.; Flahault, A.; de Lamballerie, X. Chikungunya Fever: A Clinical and Virological Investigation of Outpatients on Reunion Island, South-West Indian Ocean. PLOS Neglected Trop. Dis. 2013, 7, e2004. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.S.; Botero, S.; Simon, S.M. Sequencing the peripheral blood B and T cell repertoire – Quantifying robustness and limitations. J. Immunol. Methods 2018, 463, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Takahashi N, Kobayashi Y, Udagawa N. Osteoclasts. Principles of Bone Biology [Internet]. Elsevier; 2020. p. 111–131. Available from: https://linkinghub.elsevier. 9780.
- Maeda, K.; Kobayashi, Y.; Udagawa, N.; Uehara, S.; Ishihara, A.; Mizoguchi, T.; Kikuchi, Y.; Takada, I.; Kato, S.; Kani, S.; et al. Wnt5a-Ror2 signaling between osteoblast-lineage cells and osteoclast precursors enhances osteoclastogenesis. Nat. Med. 2012, 18, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.-I.; et al. Induction and Activation of the Transcription Factor NFATc1 (NFAT2) Integrate RANKL Signaling in Terminal Differentiation of Osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef]
- Ramos-Junior, E.S.; Taira, T.M.; Fukada, S.Y. Bone—From macrophage to osteoclast and osteolytic diseases. In Macrophages in the Human Body. Camara, N.O.S., Braga, T.T. Eds.; Academic Press, 2022; p. 161–180.
- Chen, W.; Zhu, G.; Hao, L.; Wu, M.; Ci, H.; Li, Y.-P. C/EBPα regulates osteoclast lineage commitment. Proc. Natl. Acad. Sci. 2013, 110, 7294–7299. [Google Scholar] [CrossRef]
- Allard-Chamard, H.; Carrier, N.; Dufort, P.; Durand, M.; de Brum-Fernandes, A.; Boire, G.; Komarova, S.; Dixon, S.; Harrison, R.; Manolson, M.; et al. Osteoclasts and their circulating precursors in rheumatoid arthritis: Relationships with disease activity and bone erosions. Bone Rep. 2020, 12, 100282. [Google Scholar] [CrossRef]
- Durand, M.; Komarova, S.V.; Bhargava, A.; Trebec-Reynolds, D.P.; Li, K.; Fiorino, C.; Maria, O.; Nabavi, N.; Manolson, M.F.; Harrison, R.E.; et al. Monocytes from patients with osteoarthritis display increased osteoclastogenesis and bone resorption: The In Vitro Osteoclast Differentiation in Arthritis study. Arthritis Rheum. 2012, 65, 148–158. [Google Scholar] [CrossRef]
- Yap, H.-Y.; Tee, S.Z.-Y.; Wong, M.M.-T.; Chow, S.-K.; Peh, S.-C.; Teow, S.-Y. Pathogenic Role of Immune Cells in Rheumatoid Arthritis: Implications in Clinical Treatment and Biomarker Development. Cells 2018, 7, 161. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Shobuike, T.; Shiraki, M.; Kamohara, A.; Hirata, H.; Murayama, M.; Mawatari, D.; Ueno, M.; Morimoto, T.; Kukita, T.; et al. Leukemia/lymphoma-related factor (LRF) or osteoclast zinc finger protein (OCZF) overexpression promotes osteoclast survival by increasing Bcl-xl mRNA: A novel regulatory mechanism mediated by the RNA binding protein SAM68. Mod. Pathol. 2022, 102, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Suhrbier, A. Rheumatic manifestations of chikungunya: emerging concepts and interventions. Nat. Rev. Rheumatol. 2019, 15, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.; Creecy, A.; Awosanya, O.D.; et al. SARS-CoV-2 and its Multifaceted Impact on Bone Health: Mechanisms and Clinical Evidence. Current Osteoporosis Reports 2024, 22: 135-145.
- Caetano, C.C.S.; Azamor, T.; Meyer, N.M.; Onwubueke, C.; Calabrese, C.M.; Calabrese, L.H.; Visperas, A.; Piuzzi, N.S.; Husni, M.E.; Foo, S.-S.; et al. Mechanistic insights into bone remodelling dysregulation by human viral pathogens. Nat. Microbiol. 2024, 9, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Drissi, H.; Sanjay, A. The Multifaceted Osteoclast; Far and Beyond Bone Resorption. J. Cell. Biochem. 2016, 117, 1753–1756. [Google Scholar] [CrossRef] [PubMed]
- Negishi-Koga, T.; Gober, H.-J.; Sumiya, E.; Komatsu, N.; Okamoto, K.; Sawa, S.; Suematsu, A.; Suda, T.; Sato, K.; Takai, T.; et al. Immune complexes regulate bone metabolism through FcRγ signalling. Nat. Commun. 2015, 6, 6637–6637. [Google Scholar] [CrossRef] [PubMed]
- Harre, U.; Lang, S.C.; Pfeifle, R.; Rombouts, Y.; Frühbeißer, S.; Amara, K.; Bang, H.; Lux, A.; Koeleman, C.A.; Baum, W.; et al. Glycosylation of immunoglobulin G determines osteoclast differentiation and bone loss. Nat. Commun. 2015, 6, 6651. [Google Scholar] [CrossRef] [PubMed]
- Bartholomeeusen, K.; Daniel, M.; LaBeaud, D.A.; et al. Chikungunya fever. Nat Rev Dis Primers 2023, 9(1), 17. [Google Scholar] [CrossRef]
- Lacey, D.L.; Boyle, W.J.; Simonet, W.S.; Kostenuik, P.J.; Dougall, W.C.; Sullivan, J.K.; Martin, J.S.; Dansey, R. Bench to bedside: elucidation of the OPG–RANK–RANKL pathway and the development of denosumab. Nat. Rev. Drug Discov. 2012, 11, 401–419. [Google Scholar] [CrossRef]
- Deng, C.; Zhang, Q.; He, P.; Zhou, B.; He, K.; Sun, X.; Lei, G.; Gong, T.; Zhang, Z. Targeted apoptosis of macrophages and osteoclasts in arthritic joints is effective against advanced inflammatory arthritis. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Study design, CDR3 diversity and cell abundance. Panel A: number of samples and group division in each dataset. B: workflow for integrating immune repertoire, deconvolution, and transcriptomic analysis from RNA-seq. C: CDR3 Chao1 diversity for BCR (IGH, IGK, and IGL) and TCR (TRA and TRB), normalized by z-score for each chain within the same dataset. D: summary of the estimated cells in each sample.
Figure 1.
Study design, CDR3 diversity and cell abundance. Panel A: number of samples and group division in each dataset. B: workflow for integrating immune repertoire, deconvolution, and transcriptomic analysis from RNA-seq. C: CDR3 Chao1 diversity for BCR (IGH, IGK, and IGL) and TCR (TRA and TRB), normalized by z-score for each chain within the same dataset. D: summary of the estimated cells in each sample.
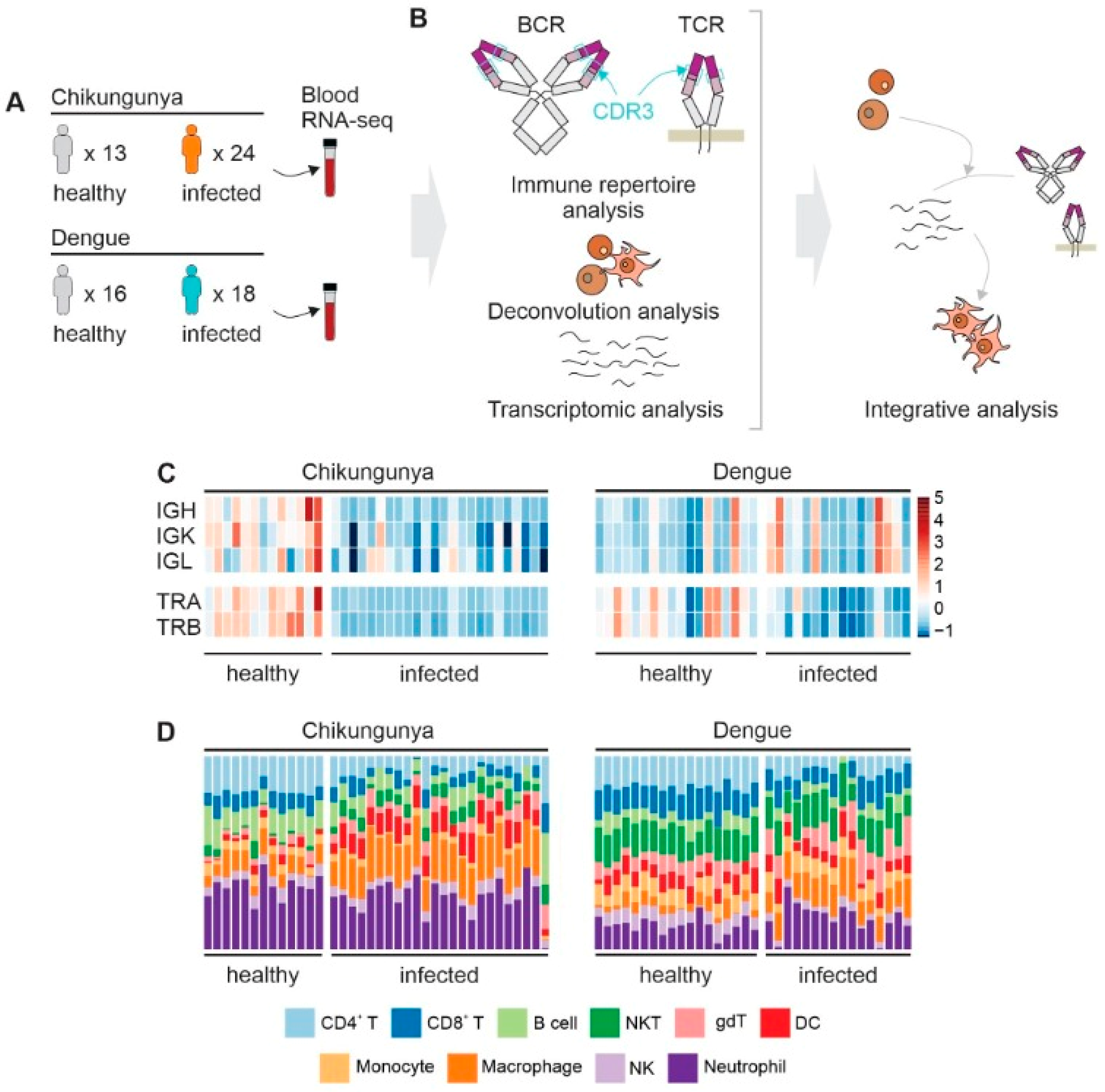
Figure 2.
Gene set enrichment analysis of the correlation between gene expression and CDR3 diversity. The dot plot shows the normalized enrichment score (NES) for each gene set, which reflects the correlation coefficients between gene expression and CDR3 diversity. T cell receptor complex and Immunoglobulin complex were included as positive controls for TCR and BCR diversity, respectively. Circle size reflects number of genes that contribute to the enrichment. Circles with black borders indicate an adjusted p-value < 0.1.
Figure 2.
Gene set enrichment analysis of the correlation between gene expression and CDR3 diversity. The dot plot shows the normalized enrichment score (NES) for each gene set, which reflects the correlation coefficients between gene expression and CDR3 diversity. T cell receptor complex and Immunoglobulin complex were included as positive controls for TCR and BCR diversity, respectively. Circle size reflects number of genes that contribute to the enrichment. Circles with black borders indicate an adjusted p-value < 0.1.
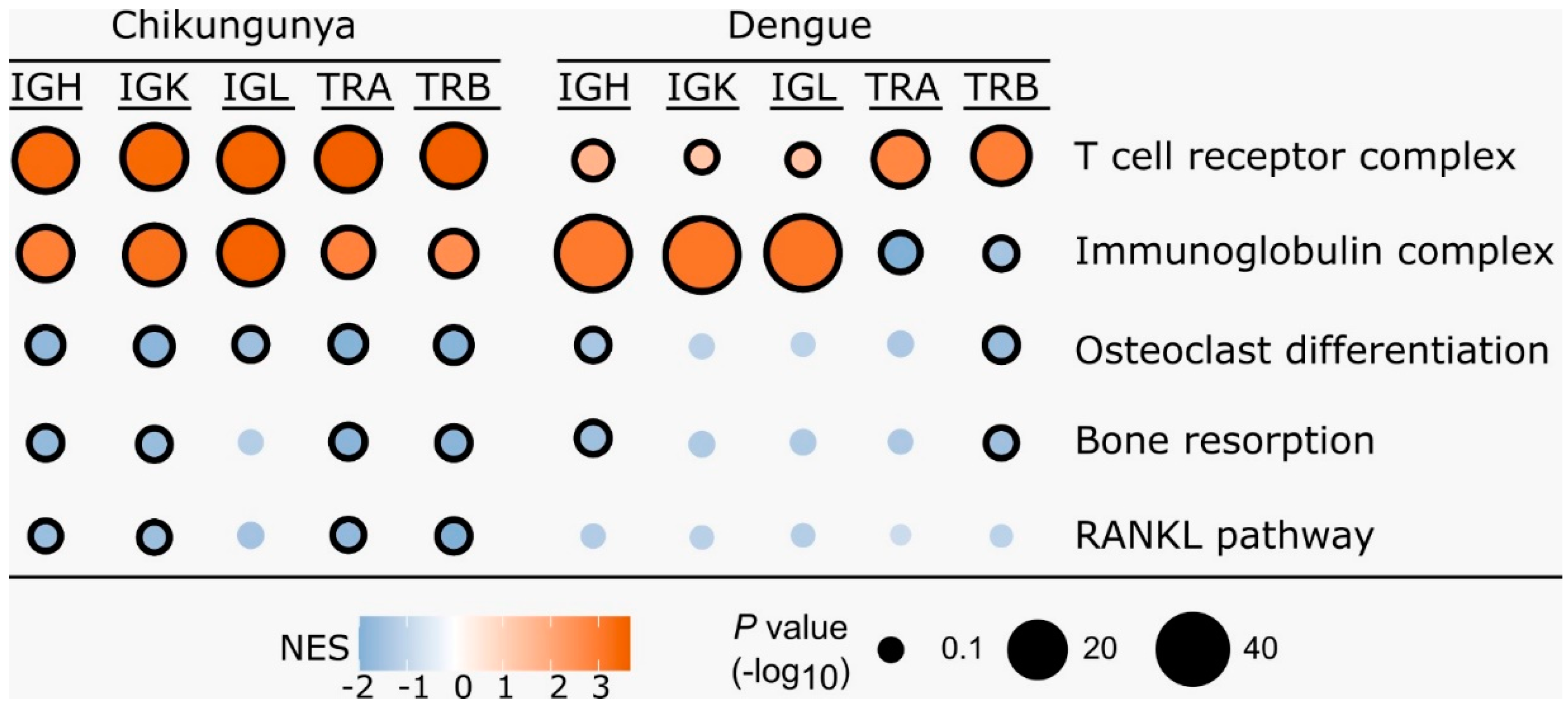
Figure 3.
CHIKV infection increases expression of genes associated to osteoclast differentiation. Proposed model of how CHIKV infection increases expression of genes associated to osteoclast differentiation and activation. Panel A shows that WNT5A expression is elevated during CHIKV infection, leading to increased RANK expression in osteoclast precursors through AP-1 and SP1 signaling. CHIKV infection also reduces BCR and TCR CDR3 diversity and may result in the formation of immunocomplexes that bind to FCyR. Osteoclast precursors that overexpress RANK and are stimulated through FCyR, RANK, and M-CSF receptors undergo differentiation and activation. Panel B illustrates that RANK signaling in osteoclast precursors ultimately leads to NFATC1 expression and osteoclastogenesis. Additionally, ZBTB7A expression promotes cell survival by favoring the alternative splicing of Bcl-xl, an antiapoptotic protein. Signaling through FCyR can increase intracellular calcium levels, which enhances NFATC1 expression. Genes upregulated during CHIKV infection are highlighted in red.
Figure 3.
CHIKV infection increases expression of genes associated to osteoclast differentiation. Proposed model of how CHIKV infection increases expression of genes associated to osteoclast differentiation and activation. Panel A shows that WNT5A expression is elevated during CHIKV infection, leading to increased RANK expression in osteoclast precursors through AP-1 and SP1 signaling. CHIKV infection also reduces BCR and TCR CDR3 diversity and may result in the formation of immunocomplexes that bind to FCyR. Osteoclast precursors that overexpress RANK and are stimulated through FCyR, RANK, and M-CSF receptors undergo differentiation and activation. Panel B illustrates that RANK signaling in osteoclast precursors ultimately leads to NFATC1 expression and osteoclastogenesis. Additionally, ZBTB7A expression promotes cell survival by favoring the alternative splicing of Bcl-xl, an antiapoptotic protein. Signaling through FCyR can increase intracellular calcium levels, which enhances NFATC1 expression. Genes upregulated during CHIKV infection are highlighted in red.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



