
Submitted:
01 August 2024
Posted:
02 August 2024
You are already at the latest version
Abstract
A truss network was applied to analyze morphometric differentiation among three wild populations of Hoplias microlepis, captured in three different sites from two habitats of Guayas Hydrographic Basin: Daule-Peripa dam, Quevedo and Babahoyo rivers. This study can help to deepen to the knowledge of the species adaptation to different environments and highlights some factors which may be helpful for future management and conservation of the freshwater resources of Ecuador. A sample of 225 adult fish were collected, 49 morphometric characteristics and 6 meristic counts were analyzed, and a statistical model was built through multifactorial discriminant analysis. The standard length of Hoplias microlepis was 32.14 cm, total length 37.82 cm and sexual maturity index 4.39. The Hoplias microlepis with the greatest weight, length and sexual maturity index were in the dam where the food availability is better, and the fishing pressure is lower. Finally, the cluster analysis grouped the fish from the rivers in cluster 1 and those from the dam in cluster 2, being samples of dam clearly distinct from rivers, probably due to differences in the habitat condition. Fish body condition and morphological variation at different sites may be useful indicators for both, habitat disturbance and consequently its conservation, and the sustainability of fishery management plans.
Keywords:
Guayas River Basin
; native resources
; morphological differentiation
; discriminant analysis
; conservation
; fishing pressure
1. Introduction
Latin American countries are home to a large variety of fish species. With 951 native freshwater species, Ecuador may be considered as a biodiversity reserve [1]. The fluvial network of Ecuador is complex and diverse, and the Guayas River Hydro-graphic Basin (CHG), covering 53,299 km2 (Figure 1), is the largest Pacific Ocean basin in South America [2]. The CHG is made up of 7 sub-basins with high geomorphological, climatic, and biological diversity. This basin contains 123 species be-longing to 14 orders [3], many of which are locally endemic and inhabiting different freshwater and estuarial habitats [1].
According to Olaya Carbó et al. [4], the CHG is made up of several fragile river ecosystems, which are permanently subject to risk factors, such as modification, fragmentation and destruction of habitats, introduction of non-native species (Oreochromis spp), overfishing, environmental contamination (herbicides, heavy metals, etc.), development of large-scale intensive forestry practices, loss of modification of natural hydro-logical regime including within basin river connectivity and finally climate change [2,5,6,7]. The anthropogenic disturbances over the freshwater ecosystems had a strong impact, were characterized by the destruction of habitats, environmental pollution, climate change, the introduction of invasive species [8], and the overexploitation of natural resources [9,10]. All these combined have led to a rapid loss of biodiversity. On the contrary, an adequate regulation of human activity, which contributes to the risk factors mentioned above, would be essential for conservation of these eco-systems [11,12].
The changes in environmental factors could induce changes in behavior, morphology, and physiology of wild populations of native fishes [13,14,15,16,17] between environmental variations and the morphological diversification of a population. Therefore, habitat modifications will influence changes in the composition and population structure of the species [5,18,19]. In this sense, diversity of species could be considered as an ecosystem restoration indicator [20].
The evaluation of morphological variations in a native freshwater species living under different conditions within the same habitat could help to identify the factors responsible for these differences [21]. Ndiwas et al. [15] reported that morphological variations that are strongly correlated to the environmental conditions. Kurtul et al. [22] reported how fish populations in Lake Bafa (Western Anatolia) respond to ecological shifts. So, long-term biodiversity monitoring is crucial for freshwater ecosystems as it enables the detection of even subtle changes and biodiversity trends, guiding conservation efforts and ensuring the sustainability of these vital habitats. Despite some recent works investigating functional changes in aquatic communities and their consequences on ecosystem functioning [23,24], studies investigating drivers of such changes have remained insufficient. This lack of linking environmental changes to effects on an ecosystem makes conservation or management interventions difficult, exacerbating a possible deterioration of native genetic resources and the fishery production [20,22,25].
The biodiversity conservation programs and fisheries policies of each country seek food sovereignty, the maintenance of genetic resources and biodiversity through economic incentives and regulatory measures [26]. The construction of simple habitat conservation indicators in Latin America would be a useful although complex tool due to the lack of data on the characterization of wild fish populations and studies of productive behavior [27]. The development of conservation programs in genetic resources implies a morphology and productive characterization [11]. Morphological characterization may be helpful to establish a conservation program. Regarding the conservation of native fish genetic resources, the evaluation of the morphostructural variations shown by a species in different conditions within the same hydrographic zone could help to identify the causes responsible for these differences [21]. Previous studies using geometric morphometrics to investigate variation in fishes have found differences between sexes [28], diets [29,30], geographic location [25,31], and habitat characteristics [32,33].
Therefore, knowledge of the morphological variations of native fish in certain rivers could help, on the one hand, to understand native resources, the association between morphological variations and ecosystem changes, and to the development of public fishing and restoration policies of ecosystems [34,35].
The guanchiche, Hoplias microlepis (Günther, 1864), is a native fish of Ecuador widely distributed in most Pacific River basins from Costa Rica to Peru [36]. It constitutes a protein source of high nutritional value for indigenous and local communities [37] and, it is generally caught with artisanal nets. Guanchiche is a benthic fish and has a slightly compressed and elongated cylindrical body [38]. This fish has dark brown coloration, brown fins, and black patches band-shaped. It has a strong jaw with cone-shaped teeth. Benthic fishes have the capacity to withstand temperature and salinity fluctuations, and an optional use of anaerobic metabolism in periods with low oxygen in the environment [38]. Considering all these characteristics, guanchiche is a quite tough fish that could remain out of the water for a long time. It shows preference for slow-moving river and is commonly found in swamps and shallow wells, where it stands motionless within vegetation to capture their prey (fish, amphibians, insects, rodents, birds, etc.) [37]. It was considered a common species, however, the latest data collected indicate a decline in number and length of fish caught [39]. Intensive overfishing of guanchiche resulted in current stage, where the fish is considered as endangered species [40,41].
Our objective was to investigate the morphological variation of guanchiche traits in three wild populations of the Guayas Basin in the face of environmental changes and the impact of fishing policies. Under the hypothesis that the morphological differences be-tween populations of native freshwater species could be used as bioindicators. In this research meristic and morphometric traits were measured to examine morphological divergence of Hoplias microlepis in different parts of the drainage and in contrasting habitat types, river sites and artificial impoundments. Knowledge of the phenotypic variations in relation to environmental modifications could be used to identify key factors for policy makers in terms of the development of both diversity conservation programs and sustainable fishing practices.
Ferrito et al. [42] and Mir et al. [43] conducted similar studies on other fresh-water species, and Dasgupta et al. [44] stated that morphological discrimination in various populations is strongly influenced by habitat differences. Growth variations also occur in response to different habitats [45].
The general hypothesis of this study was that fishing policies, resource management, and environmental conditions influence the differentiation of populations due to the phenotypic plasticity of Hoplias microlepis. Subsequently, three hypotheses were formulated, as displayed below:
Hypothesis 1.
Fishing and resource management practices (such as fishing methods and pressure, respect of the closure periods, land use, endogeneity level, and competition with native and non-native species) influence the morphological variation of Hoplias microlepis in the CHG. The null hypothesis (H0) was that there would be an equality of means between the populations of Hoplias microlepis in Daule-Peripa Dam and Babahoyo river (H0: μ1 = μ3). In contrast, the alternative hypothesis was that significant differences would exist due to fishing management practices (H1: μ1 ≠ μ3).
Hypothesis 2.
The physical environmental conditions influence the morphological variation of Hoplias microlepis. The null hypothesis (H0) was that there would be an equality of means between the populations of Hoplias microlepis in the Quevedo and Babahoyo rivers (H0: μ2 = μ3). In contrast, the alternative hypothesis was that significant differences would exist due to a change in environmental conditions (H1: μ2 ≠ μ3).
Hypothesis 3.
The fishing management and environmental conditions influence the morphological variation of Hoplias microlepis in the CHG. The relationships among three populations were analyzed through discriminant analysis, where three groups were identified: Population 1 (Daule-Peripa Dam) with low fishing pressure and low flow velocity, Population 2 (Quevedo river) with a high fishing pressure and white water or water containing a high concentration of oxygen, and Population 3 (Babahoyo river) with a high fishing pressure and a low flow velocity.
This study aims to investigate the adaptation of fish morphology to human impacts on the habitats as previous step to establish a conservation plan in Hoplias microlepis. Therefore, how fishing policies, resource management, and environmental conditions influence the differentiation of populations due to the phenotypic plasticity of Hoplias microlepis was evaluated.
2. Materials and Methods
2.1. Study Area
The hydrographic basin of Guayas fluvial system (Figure 1) comprises three geo-morphic reliefs (upper-middle basin, lower-middle basin, and lower basin) which provide different ecosystems used in the rearing of native species in Ecuador [4]. Three samples of native fish were selected, two belonging to riparian habitats (Quevedo River and Babahoyo River) and the other is from lentic habitat (dam) (Figure 1).
The Daule-Peripa dam (sampling 1), collects the fluvial network from the Chongón-Colonche Mountain range west and corresponds to the upper-middle basin, amongst the provinces of Los Ríos, Manabí, Santo Domingo de Los Tsáchilas and Guayas. It comprises an area of 295 km2 and a maximum depth of 85 m allowing the rearing of many freshwater species [3]. The physical characteristics of reservoirs drive changes in the morphological attributes of native fish populations [14]. It has become a controlled place in the face of indiscriminate fishing for native freshwater species. However, the introduction of species such as cachama (Colossoma macropomum) and paiche (Arapaima gigas) is causing a decrease in the native species [46].
In the middle basin of the Guayas is the Quevedo river (sampling 2), originated in the foothills of the Cordilleras of Los Andes at a height of 625 m, it has a route from north to south and a length of 163 km. This river is in the province of Los Ríos, ending at 79 m, in the Babahoyo river in the province of Guayas. The maximum depth ranges from 2 m in the dry season (between June and November) to 5 m in the rainy season (between December and May). Different native species inhabit this river, such as vieja azul (Andinoacara ri-vulatus), vieja colorada (Cichlasoma festae), raton (Leporinus ecuadorensis), dama (Brycon dentex), bocachico (Prochilodus magdalenae), etc.
In the lower basin of the Guayas delta, the Babahoyo river is located (sampling 3), being slow-flowing due to its reduced slope. This type of flow is suitable for benthic fish such as guanchiche, which coexist with introduced species like Oreochromis spp [6]. This river is under the highest pressure due to the fishing cooperatives authorized by the National Directorate of Fisheries (DNP for its acronym in Spanish) and to the high preference for the consumption of freshwater species by the locals.
The overall CHG area has a tropical climate with an average temperature of 25 °C, annual rainfall of 2,400 mm and relative humidity of 82% [28]. Further details regarding the physiological characteristics of our sample sites can be found in Table 1.
The availability and diversity of fishes are indicators of the degree of human intervention and determine the growth and development of different fish species [12]. Food availability and contamination are conditioned by different abiotic parameters, such as current flow, turbidity, and dissolved oxygen concentration, among others [47]. The Quevedo river is rich in phytoplankton, while the Babahoyo river is rich in zooplankton and fishes as Astyanax sp. Furthermore, in the Babahoyo river, the presence of Polymyxus coronalis has been related to eutrophic waters. In the case of the Quevedo river, Jiménez-Prado et al. [3] determined a high diversity of phyto-plankton and low diversity of zooplankton. In the Babahoyo and Quevedo rivers there is less diversity of species (Table 1).
The most commonly used fishing gear are hand-held line with hooks (10% of catch), gill net (20%), with a length of 100-150 m, 4 to 5 m in height and ¾ to 1 inch in mesh light diameter; the trammel (30%), with a mesh size between 2 and 3½ inches; casts (35%) with a mesh size between ½ and 1 inch and others such as harpoon hooks, etc. (5%). Balsas and canoes were ancestrally used by fishermen, while the bongo (15-70 CV), the boat (25-50 CV) and lancha (30-180 CV) use motors providing greater strength and autonomy. Previous research by MAGAP [48], Ochoa Ubilla et al. [40] and Pacheco-Bedoya [41], widely analyzed the diversity of traditional fishing tackle utilized in CHG [25].
2.2. Water Quality
Jiménez-Prado et al. [3] propose different water quality parameters for freshwater fish farming. Table 2 shows the values reported for the three study areas where the physical, chemical, and biological characteristics are included, with values within the range of those recommended for freshwater fish farming [29]. Furthermore, Alvarez-Mieles et al. [5] and Aguilar et al. [18] established critical values and identified maximum contamination situations that would serve to prevent any alteration in fish production due to human induced environmental change. Moreover, hydrological and geological variability stand out among the physical factors, which significantly influence the structure and function of communities in river ecosystems [49,50]. The natural physical attributes (pH, temperature, color, conductivity, turbidity, dissolved oxygen, total dissolved solids), chemical (chlorides, alkalinity, nitrates, ammonium) and biological indicators (phytoplankton) of different waters are a result of their origin and uses [12].
2.3. Data Collection
Local fishermen were informed about the study objectives and their importance to conserve the guanchiche and their livelihoods. They in turn, told us about the poor situation of guanchiche and the rearing area. The questionnaires included questions regarding the reasons for river area preference, sites of capture, causes of the fish population decrease and actions for its conservation. The fishermen caught 250 adult specimens of guanchiche during 2020-2021 in three sampling sites with weight and length greater than 170 g and 20 cm, and phenotypic characteristics of the species. They were fishery both in rainy and dry season, although most of them were captured, after spawning, at the beginning of the dry season, between March and April. The fishes were caught using a traditional fishing system called “wing deployment” [33], which sweeps the net in the opposite direction to the flow of water. Specimens selected had a state of sexual maturity greater than 1. The weigh and observation of the gonads was carried out according to the methodology described by Less and López [55] and Revelo [56]. The non-representative specimens of the species (10% of the sample), with small size, fish of other species, mutilated, as well as young individuals were im-mediately returned to the river.
Therefore, a sample formed by 225 mature fishes from Hoplias microlepis (132 females and 93 males) were used in this research from three different sites; 75 the Daule-Peripa dam, 75 from the Quevedo river and 75 from the Babahoyo river. These captured fishes were deposited in a cage attached to the net, in the same river. Subse-quently, they were placed in a transition pond under natural conditions before being transported alive following the recommendations of Gonzalez-Martinez et al. [30]. In the laboratory, all specimens were kept in two tanks of 500 l each (dissolved oxygen = 6.20 ± 0.0 mg/l, temperature = 20.5 ± 0.2 °C and pH = 5.6 ± 0.1). Fishes rested for 48 h before the experiment, with fasting time of 24 h before their death. On the experience day, the water was reduced by half in the tank, the fishes were quickly caught with a net and transferred into a plastic box (100 l). To stun the fish, 40 l of ice and 40 l of water (0.8 °C) were added to the plastic box. After approximately 20 minutes, each fish was identified with a number and weight, and morphometric measurements and the corresponding meristic counts were recorded. Sex was determined through external phenotypic characteristics. The procedure was carried out according to the Canadian Council on Animal Care guide for investigative fish management [57]. We have tried to take all the necessary steps to assure the welfare of native fishes in this research, during the handling, transport, and sacrifice phases. Nevertheless, the suggested method of euthanasia by the referenced authority (CCAC) was not used. According to the Guide to Good Practice for Slaughter [58], they were sacrificed. The animal welfare in each research step was supervised by a veterinary practitioner.
2.4. Body Measurements
Morphometric measurements were taken by the same person, on the left side of the fish, following the conventional method described by Diodatti et al. [59]. Animals were measured using an ichthyometer, graduated digital calipers and tapes with 0.01 mm of precision. The meristic characters were examined following the indications of Mattox et al. [36]. A total of twenty-six morphometric measurements based on nineteen landmarks on the left side of the fish and six meristic counts were obtained (Table 3 and Figure 2). Sexual identification was carried out by direct observation of external sexual characteristics. To determine the states of sexual maturity, the modified scale of Less and López [55] was used; from 1 to 5 (1 immature, 2 recovery, 3 sexual maturity, 4 to spawn, 5 spawned). The selection of variables and the methodology followed Gonzalez et al. [29], Caez et al. [28] and Gonzalez-Martinez et al. [25,30,33] approaches for further comparisons.
2.3. Fulton Condition Coefficient (K)
Fulton’s Condition factor (K), which is defined as the wellbeing of the fish, was calculated using the following equation K = 100 BW/SL3, where BW is total weight (g), and SL is the standard length in cm. K is widely used in fisheries and general fish biology studies for quantifying variations in fish populations [29,60].
2.6. Statistical Analysis
Morphometric variables used were standardized according to Gonzalez Martinez et al. [25,33] and Elliot et al. [61]. Kolmogorov–Smirnov test and Bartlett test were per-formed to verify the normality and equality of the data variance (homoscedasticity). The KMO sampling adequacy test showed a value greater or 0.7, while the Bartlett test showed a satisfactory probability value (p < 0.001), thus indicating that the sampling was adequate [25,30,33].
Firstly, morphometric characters (original and adjusted) were compared by Student t-test and the meristic variables were compared by Kruskal-Wallis test with river or dam included as a fixed effect (Hypotheses 1 and 2). Sex was not considered as a fixed effect, since no significant differences between males and females had been found previously in an exploratory study.
Secondly, discriminant analysis was used to test Hypothesis 3 of this research. DISCRIM procedure was used to perform a canonical discriminant analysis of size-adjusted geometric morphometric data, using the three wild fish population (sampling of Daule-Peripa Dam, Quevedo River and Babahoyo River) as the grouping variable. The probabilities to enter and to stay in the model were both set at P < 0.05. The selection of the most discriminant variables was conducted by applying the F-Snedecor, Wilks’ lambda, and 1-Tolerance methods. The correct assignment percentage was considered and the Mahalanobis distances were represented graphically by cluster. Also, the spatial distribution of fish was represented by canonical scatterplot. Statistica 12.0 for Windows software was used to perform the statistical analyses (StatSoft, Tulsa, OK, USA).
3. Results
The measured guanchiche individuals had an average weight of 442.66 ± 29.52 g (CV = 68.97%), a standard length of 32.14 ± 0.60 cm (CV = 19.28%) and a total length of 37.82 ± 0.69 cm (CV = 18.96%) (Table 4). The fishes with the largest size were found in Daule-Peripa dam and Quevedo river, whilst fish morphometric measures in Babahoyo river had lower values. The coefficient of variation obtained in morphometric measures was high for the overall dataset, and low when considering each sampling population separately. The condition factor (K) averaged 1.47 with a high coefficient of variation (69.70%), and significant differences found according to sampling populations (P < 0.001): Daule-Peripa dam showing the highest values of weight (BW) and total length (TL), and Babahoyo river showing the lowest values. The growth of fishes was not isometric, with an allometric coefficient of 0.186. However, all the correlations were significant (P < 0.001) (data no presented).
In relation to Hypothesis 1, the results of the morphometric comparisons between two wild populations from the Daule-Peripa dam and Babahoyo river with different fishing management practices were presented in Table 2 (A × C). There were significant differ-ences in body weight or the condition factor between the two populations (P < 0.001). Fourteen of the 25 morphometric measures showed significant differences for the populations from the Daule-Peripa dam and Babahoyo river (BW, K, HL, Pre-PvL, DFL, PcFL, PVFL, AFL, AFRL, AC1, AC2, AC3, P2 and LC2).
Regarding Hypothesis 2, Table 2 (B × C) shows the influence of environmental conditions on morphological differentiation in populations of Guanchiche. Significant dif-ferences between the Quevedo and Babahoyo rivers were found for 13 morphometric variables: BW, K, Pre-OL, Pre-PvL, DFL, PVFL, AFL, AFRL, AC1, AC2, AC3, P1 and P2.
Guanchiche’s sexual maturity index obtained was 4.39 ± 0.12 g (28.20%). Significant differences in the sexual maturity index among three populations were found (Figure 3). Daule-Peripa dam and Quevedo river showed the highest values and Babahoyo river the lowest values. Furthermore, the Babahoyo river had a temperature higher by 8 °C than in the other sites (Table 2).
Regarding meristic characters, the following values were found for the average number of fin rays: dorsal (12.96 ± 0.11), pectoral (10.71 ± 0.20), pelvic, (7.79 ± 0.05), anal (9.45 ± 0.12), caudal (16.96 ± 0.08) and branchial (3.65 ± 0.05). In most of the variables no significant differences were found between native population (A x C) and (B x C) (Table 5).
The coefficients of variation value ranged between 5 and 23%, in response to the differences in the number of fin rays recorded in this species. The number of rays of the dorsal fin ranged between 10 and 15 (66% of the sample had 13-14) and of the pectoral fin was within the range 4 and 14 (75% of individuals had 10-13). The results for pelvic fin ranged between 6 and 9 (76% of the fish had 8 rays). With significantly higher values in Babahoyo river compared to Daule-Peripa dam (P < 0.05). The anal fin ranged between 6 and 11 (67% of the fish had 9-10).
The caudal fin showed values between 14 and 18 rays (52% of the specimens had 17). Daule-Peripe dam and Babahoyo river showed the lowest values, while the highest values were obtained in the Quevedo river (P < 0.05). The number of gill arches ranged between 2 and 4 (68% of the fish had 4). The number of rays of the different fins and gill arches was similar among sampled population. However, it was found that: in Daule-Peripa dam, no specimen showed 15 rays on the dorsal fin, specimens with 14 rays on pectoral fin or 6 rays on the pelvic fin; in Quevedo river, no individuals were found with 10 rays on the dorsal fin, 6 and 9 rays on the pelvic fin, 14 rays on the caudal fin; in Babahoyo river, no specimen showed 14 rays on its pectoral end.
Before performing the discriminant analysis, all data of body, Fulton´s factor and morphometric measures were standardized by Elliot et al. [61]. The adjusted variables showed significant differences in weight, factor de condition and 14 morphometric measures (P < 0.05) (Table S1). Fish specimens in sampling Daule-Peripa dam showed the highest weight and size values, and those in Babahoyo river the lowest values. In general, the fish in Quevedo river had a greater length (HL, ED, Pre-AL, DFL, PcFL, PvFL) with a higher mean value in 16 morphometric measurements. In contrast, those specimens from Babahoyo river showed a lower mean value in majority of morphometric measurements.
There were no significant correlations (P > 0.05) between standardized morphometric measurements and standard length, and therefore all were used in the discriminant analysis. As predictors, fifteen morphometric variables were selected, six of them were significant (P < 0.05), five of those relative to length (AFRL, Pre-PvL, PvFL, UJL y DFRL) and one to body depth (AC3) (Table 6). According to the results of the F-Snedecor, Wilks’ lambda, and 1-Toler methods, there were three major discriminant variables in the model (25%): anal fin ray length (AFRL), pre-pelvic fin length (Pre-PvL) and pelvic fin length (PvFL).
The classification matrix based on body shape showed that 87.74% of the fishes were correctly classified to their original group, with the highest correct adscription for the specimens of Babahoyo river (90.91%). The errors in sampling Daule-Peripa dam y Quevedo river were mostly due to misclassification with specimens from Babahoyo river. All this can be better observed in graphic representation of overall specimens from canonical scores (Figure 4). Two groups were identified by the cluster analysis from Mahalanobis distances performed on adjusted morphometric measurements, by clearly separating all specimens of Quevedo river from Daule-Peripa dam and Babahoyo river (Figure 4).
4. Discussion
The morphostructure is crucial in biological studies and organisms form detection, which have been attributed to the ecological condition where they live [62]. Variations in the shape of freshwater fish are modulated by different habitat conditions and other anthropogenic activities [63]. Moreover, environmental disturbances greatly influence the developmental stability aspect that leads to the species inability to grow naturally [28,29,30].
Populations of Hoplias microlepis from river and dam of the Guayas Basin differ significantly in morphology due to phenotypic plasticity or contemporary evolution [32]. Phenotypic plasticity, non-genetic change in phenotypes, may well be the cause of the differences founded in Hoplias microlepis [32]. However, the contemporary evolution, change in allele frequencies, need a few hundred years to be measurable but dams of Guayas Basin are relatively news [32].
The fishes caught in our study were of comparable size, average 32.14 cm SL and 37.82 cm TL, and sexual maturity, to those recorded in previous studies [64,65]. Bussing [64] observed a maximum size in Costa Rica about 40 cm TL; Laaz and Torres [65] reported an average size of 32 cm SL in Ecuador specimens; and Revelo [56] observed about 48% of the specimens with a sexual maturity index less than 3. The present study is showing higher values of both length and sexual maturity in each population, but lower body size when compared to Bussing [64]. Accordingly, the high coefficient of variation obtained reinforces the heterogeneity between populations. However, the low standard error and the low variability within each site indicate homogeneity within the group due to a potential shared morphological plasticity and a parallel adaptation to similar habitat types [32]. The strong variability on the traits of guanchiche found in the Guayas Basin was one of the causes that motivated this research. According to Matox et al. [36] Hoplias spp group represents one of the most complex taxonomical problems in the systematics of Neotropical fishes with many nominal species, some of which without known type material, renders the problem more complicated. In addition, the variability is greater in native wild fish than in cultured fish [28,29,30]. When we confronted the morphometric results obtained with the three proposed research hypotheses, these were whole or in part accepted:
In hypothesis 1 the differences between Hoplias microlepis population from Daule-Peripe Dam and Babahoyo river were influenced by the fishery policies, high fishing pressure, the overfishing of native species in the Babahoyo river, and the introduction of non-native species [1,56,66,67]. In this sense, hypothesis 1, that fishing policies and resource management influence the morphological differentiation of Guanchiche populations between the Daule Peripe dam and Babahoyo river, was accepted for 14 of the 25 morphometric variables, for 2 meristics characters and the sexual maturity index.
Hoplias microlepis from the Daule-Peripa dam were bigger than the fish from Ba-bahoyo river. With specimens from the dam showing the highest values in most of mor-phometric measurements relationship with weight, body depth variables (AC1, AC2 and AC3), maturity sexual and two meristic characters. In Babahoyo river has been main-taining significant lowest values mean in all morphometric variables of the captured specimens.
The morphological differences in BW, K and HL between the Daule-Peripe dam and Babahoyo river were significant. For some traits linked to the body development of both populations (perimeter, P2 and depth of body, LC2), significant differences were found too. However, contrary to expectations, there were no differences in most of the variables, aligned to total length size and structure of the fish among these populations (Hypothesis 1), most likely because the local fishing regimes are closer than initially considered [61,67].
Several factors help to understand these morphometric variations among populations, such as the habitat diversity of Ecuador’s rivers [5,19,47,50]. In the dam, the fishing pressure was low and controlled, as this area acts as a reservoir of native freshwater species and encourages the use of environmentally friendly agricultural practices. Furthermore, the fishing gear used and the fishing pressure to which each river is subjected also determine captures frequency and size [40,41]. According to MAGAP [48], Ochoa Ubilla et al. [40] and Pacheco-Bedoya [41], the native species fishing pressure in Babahoyo river is very high due to the high number of fishing cooperatives, fishing off the fishing season. The use of illegal fishing gear has also in-creased, contributing to the Babahoyo River deterioration. Instead, the largest specimens were found in Daule-Peripa dam. Therefore, the guanchiche captures quality could be used as an indicator of the habitat conservation and the endogeneity degree [6,16,21]. So, it also could be used to assess fishing pressure and the effectiveness of sustainability policies [25,68].
In the hypothesis 2 the differences in the guanchiche morphology between Quevedo and Babahoyo rivers could be explained by fragmentation, deterioration of ecosystem. hypothesis 2, was accepted for 13 of the 25 morphometric variables, for 1 meristics characters and the sexual maturity index. Specimens from the Quevedo River presented greater weight, width, and sexual maturity index; meanwhile, those from the Babahoyo River showed greater body depth (AC1 and AC3). The different morphology found in specimens of the Babahoyo river might be due to the fact that Babahoyo river was slower, shallower, presented greater turbidity, and also a higher level of photosynthesis and oxi-dative reactions. Nevertheless, Melvin et al. [69] and Ujjania and Kohli [70], pointed out the importance of genetic and environmental interactions to complement the mor-phology approach. The condition factor (K), indicative of the nutritional status of fishes, was lower in the Quevedo river (Quevedo river). This variability interpretation responds to multifactorial causes [71], such as food availability [68] or seasonality [72], including their interactions [73]. The current characterization carried out is of a preliminary nature since it only considers the morphological aspects. To determine the relative contribution of genetic and plasticity (i.e., environmentally induced phenotypic differences), genetic analyses and common garden rearing experiments would be a necessary next step.
Hypothesis 3 was partially accepted. Although the discriminant model was accept-ed-ed, Mahalanobis distances only differentiated two groups: Guanchiche’s populations from Quevedo river versus those from Daule-Peripa dam and Babahoyo river. Six mor-phological variables were selected. Six variables were significant in the model (P<0.05) and four variables were highly significant (P>0.01) and showed high discriminant power.
The discriminant and cluster analysis determined the existence of two groups: Cluster 1, which included Daule-Peripe dam and Babahoyo river population characterized by deepest water and watertight and Cluster 2, circumscribed to the Quevedo river with fast waters.
These results were not agreed to those obtained by Aguirre et al. [32] and Cocha-Alulema et al. [21], and could be due to the high mix in the morphology of the fished from the three sampling sites according to the classification matrix and representation of canonical scores. Also, we have found three major discriminant variables in the model, two relationships with fin length (anal fin ray length and pelvic fin length) and other one with body length (pre-pelvic fin length). Changes in fin lengths may be a morphological response to more hydrodynamic bodies [7].
Wild populations of guanchiche from rivers and dam differed morphologically by discriminant analysis according to Aguirre et al. [32]. Surprisingly, the Babahoyo river specimens showed the higher rate adscription in classification matrix (Babahoyo river); on the contrary, the rate adscription in the dam was low [32]. A marginal error of 11.54% was observed between the cases of Babahoyo river and Daule-Peripa dam. This could be because both (Babahoyo River and Dam) present slow flow water, little oxygen and a high degree of oxidative reactions. The relationship between flow water and body shape is not sufficient clear (Meyers and Belk, 2014) but some studies have tested it for intraspecific variation of body shape in benthic fishes across multiple flow [14,74,75].
The manifestation of a certain shape is a marker of fish mobility and habitat selection [76]. According to the results obtained in this research, guanchiche specimens occurring in fast-flowing rivers have a different shape than stationary water fish for increasing steady swimming efficiency [70]. The body shape is also conditioned by the nature, ecology, and innate characteristics of the organism [76], and by factors such as diet adaptation [29,30]. Such is the case of the head, the bigger head, the bigger mouth, and the faster suction speed [77], or such as the deep bodies that increase their ability to maneuver when feeding [78]. According to Libay et al. [76], native fish species adapt their morphology and are resilient to the environment. Therefore, morphometry is a very useful tool to associate morphometric variations with habitat type and to build conservation indicators [28,29,30]. Besides, morphometric models can be used to evaluate the effectiveness of plans for the sustainable use of fishery resources [40,41,48].
The conservation aquaculture has among its aims the conservation of fish popula-tions along with their locally adapted gene pools and characteristic phenotypes and be-haviors [79]. Likewise, the characterization of a native resource must be known before the development of a conservation program FAO [27]. Thus, it is important to control the overfishing, to follow the guidelines on Ecuadorian polices aquaculture, and to work on the maintenance of fish habitats and to avoid any introduction of non-native species, problem that has already been described above (with Oreochromis sp.).
5. Conclusions
The guanchiche morphology showed high variability, mostly affected by the site. However, standard length (32.14 ± 0.60 cm), total length (37.82 ± 0.69 cm) and sexual maturity index (4.39 ± 0.12) showed lower variability. The guanchiche showed morpho-logical variations according to the site where it develops its activity. As descends from the highlands to the lowlands of the Guayas hydrographic basin, three variables of length were decreasing. The specimens with the greatest weight, height and sexual maturity were found in Daule-Peripa dam. However, the lower size and sexual mature animals were found in the Babahoyo river.
Morphometric differences were found between the three guanchiche populations of the Guayas Basins influenced by fishing management and environmental conditions. In-depth knowledge of both factors is necessary for diversity maintenance. The different fishing policies and resource management practices present in the studied rivers have generated morphological differences that are associated with body development. Under the different environmental conditions differences in traits related to the weight, K factor, structure of fish and sexual maturity index have primarily arisen.
Discriminant analysis was a useful way to differentiate populations by easy morphometric traits. Six morphometric measurements were found to have the greatest discriminant power and were deemed to be appropriate for population discrimination. These variables were: anal fin ray length, body depth, pre-pelvic length, pelvic fin length, upper jaw length, and dorsal fin ray length. With a very small number of easily obtained morphometric traits in each population could be used to determine the biodiversity status.
The guanchiche morphometric model can be a useful to work in its conservation and thus be an indicator of habitat status by quickly detecting changes in fish shape. Furthermore, the size, weight and sexual maturity of the captured specimens, have been of great help for years to evaluate fishery management policies, considering the fishing pressure, the respect of the closure periods, the traditional fishing tackle utilized, and the native species captured in CHG of Ecuador.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Descriptive statistics of body measurements (Mean±ES) (ajusted data) of Hoplias microlepis, Table S2: Classification matrix (%) of Hoplias microlepis.
Author Contributions
Conceptualization and methodology, all authors; Formal analysis, software, data curation, data processing, A.G-M., A.G. and C.B.; Statistical analysis, A.G-M and A.G.; Validation and investigation, A.G., A. G-M and C.B.; Supervision, project administration, A.G., and M.G.; Data acquisition, M.G. and C.R.D-V.; All authors have been involved in developing, writing, commenting, editing and reviewing the manuscript. All authors read and approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The authors thank Research Project by competitive scientific and technological research funds of FOCICYT of Technical State University of Quevedo (Ecuador).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board of Quevedo State Technical University for research and the code of practices for the housing and care of animals used in scientific procedures.
Data Availability Statement
This is not applicable as the data are not in any data repository of public access, however if editorial committee needs access, we will happily provide them, please use this email: pa1gamaa@uco.es.
Acknowledgments
We want to thank the RED-RIIDPA, the open research and innovation network for the development of alternative aquaculture productions between University Técnica Estatal of Quevedo (Ecuador), University of Cordoba (Spain), and Agricultural Polytechnic of Manabi (Ecuador).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barriga, R. Lista de peces de agua dulce e intermareales del Ecuador. Rev. Politec. 2012, 30, 83-119.
- Rodríguez, J.; González, A.; Angón, E.; Vivas, R.; Barba, C.; González, M.A.; Peña, F.; García, A. Efecto del tamaño de las reproductoras en la producción de alevines de Cichlasoma festae en condiciones semicontroladas en Ecuador. ITEA 2020, 116 (2), 93-105. [CrossRef]
- Jiménez-Prado, P.; Aguirre, W.; Laaz-Moncayo, E.; Navarrete-Amaya, R.; Nugra-Salazar, F.; Rebolledo-Monsalve, E.; Zárate-Hugo, E.; Torres-Noboa, A.; Valdiviezo-Rivera, J. Guía de peces para aguas continentales en la vertiente occidental del Ecuador. Pontificia Universidad Católica del Ecuador Sede Esmeraldas (PUCESE); Universidad del Azuay (UDA) y Museo Ecuatoriano de Ciencias Naturales (MECN) del Instituto Nacional de Biodiversidad. Esmeraldas, Ecuador. 2015, 416 pp.
- Olaya Carbó P. Estado ecológico del sistema estuarino del Río Guayas, Cantón Durán, Ecuador: Simulación numérica de su dinámica fluvial y principios ecológicos para el diseño de actuaciones de restauración y/o recuperación. MSc. Universidad de Alcalá, Universidad Complutense de Madrid, Universidad Rey Juan Carlos, Universidad Politécnica de Madrid. 2016.
- Alvarez-Mieles, G.; Irvine, K.; Griensven, A.V.; Arias-Hidalgo, M.; Torres, A. Mynett, A.E. Relationships between aquatic biotic communities and water quality in a tropical river–wetland system (Ecuador). Environ. Sci. Policy. 2013, 34, 115-127.
- Ansah, Y.B.; Frimpong, E.A.; Hallerman, E.M. Genetically-Improved Tilapia Strains in Africa: Potential Benefits and Negative Impacts. Sustainability 2014, 6, 3697-3721. [CrossRef]
- Aguirre, W. E., Alvarez-Mieles, G., Anaguano-Yancha, F., Burgos Morán, R., Cucalón, R. V.; Escobar-Camacho, D.; ... & Zárate Hugo, E. Conservation threats and future prospects for the freshwater fishes of Ecuador: A hotspot of Neotropical fish diversity. J. Fish Biol. 2021, 99(4), 1158-1189.
- Soto, I.; Balzani, P.; Carneiro, L.; Cuthbert, R. N.; Macêdo, R.; Serhan Tarkan, A.; ... & Haubrock, P. J. Taming the terminological tempest in invasion science. Biol. Rev. 2024.
- Corlett, R. T. The Anthropocene concept in ecology and conservation. Trends Ecol. Evol. 2015, 30(1), 36-41. [CrossRef] [PubMed]
- Pievani, T. (2014). The sixth mass extinction: Anthropocene and the human impact on biodiversity. Rend. Fis. Acc. Lincei. 2014, 25, 85-93.
- Food and Agriculture Organization of the United Nations. Report on strategic priorities for action. For the sustainable use, development and conservation of animal genetic resources for food and agriculture. Second draft. Roma. Italia. 2005. Available online: (accessed on 25 July 2024).
- Choi, J-Y.; Kim, S-K. Effects of Aquatic Macrophytes on Spatial Distribution and Feeding Habits of Exotic Fish Species Lepomis macrochirus and Micropterus salmoides in Shallow Reservoirs in South Korea. Sustainability 2020, 12, 1447; doi:10.3390/su12041447. [CrossRef]
- Schluter, D. The Ecology of Adaptative Radiation, Oxford, UK: Oxford University Press. 2000.
- Travis, C.H.; Blum, M.J.; Heins, D.C. Morphological responses of a stream fish to water impoundment. Biol. Lett. 2010, 6, 803-806.
- Ndiwas, T.C.; Nyingi, D.W.; Claude, J.; Agnése, J.F. Morphological variation of wild populations of Nile tilapia (Oreochromis niloticus) living in extreme environmental conditions in the Kenyan Rift-Valley. Environ. Biol. Fish. 2016, 99, 473-485. [CrossRef]
- Escanta-Molina, R.; Jiménez-Prado, P. Uso de la morfometría geométrica para establecer contrastes biológicos y ambientales en poblaciones de peces del río Teaone. Revista Científica Hallazgos 2019, 4 (1), 55-69.
- Estrada Guagua, EP. Diferencias en la población de Eretmobrycon ecuadorensis del río Sálima, Cantón Atacames, a diferentes niveles altitudinales, como un bioindicador de la calidad ambiental. BSc. Pontificia Universidad Católica de Ecuador. 2019.
- Aguilar.; C.; González-Sansón, G.; Cabrera, Y.; Ruiz, A.; Allen Curry, R. Inter-habitat variation in density and size composition of reef fishes from the Cuban Northwestern shelf. Rev. Biol. Trop. 2014, 62 (2), 589-602.
- Kerezsy, A.; Arthington, A.H.; Balcombe, S.R. Fish Distribution in Far Western Queensland, Australia: The Importance of Habitat, Connectivity and Natural Flows. Diversity 2014, 6, 380-395. [CrossRef]
- Arceo-Carranza, Y.; Arceo-Carranzaa, D.; Gamboab, E.; Teutli-Hernández, C.; Badillo-Alemánay, M.; Herrera-Silveira, J.A. Conservación. Los peces como indicador de restauración de áreas de manglar en la costa norte de Yucatán. Rev. Mex. Biodiv. 2016, 87, 489-496.
- Cocha-Alulema, A.P. Análisis de la variación morfológica de Hoplias malabaricus (Bloch 1794), asociada al tipo de hábitat utilizando morfometría geométrica. Facultad de Ciencias Biológicas, Universidad Central del Ecuador, Quito, Ecuador. 2018.
- Kurtul, I.; Kaya, C.; Kaykaç, H.; Ilhan, A.; Düzbastilar, F. O.; Tosunoğlu, Z.; ... & Haubrock, P. J. How fish populations in Lake Bafa (Western Anatolia) respond to ecological shifts. Aquat. Conserv.: Mar. Freshw. Ecosyst. 2024, 34(5), e4154.
- Jeppesen, E.; Meerhoff, M.; Holmgren, K.; González-Bergonzoni, I.; Teixeira-de Mello, F.; Declerck, S. A.; ... Lazzaro, X. Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia, 2010, 646, 73-90.
- Le Hen, G.; Balzani, P.; Haase, P.; Kouba, A.; Liu, C.; Nagelkerke, L. A.; ... Haubrock, P. J. Alien species and climate change drive shifts in a riverine fish community and trait compositions over 35 years. Sci. Total Environ. 2023, 867, 161486. [CrossRef]
- Gonzalez-Martinez, A.; De-Pablos-Heredero, C.; González, M.; Rodriguez, J.; Barba, C.; García, A. Morphological Variations of Wild Populations of Brycon dentex (Characidae, Teleostei) in the Guayas Hydrographic Basin (Ecuador). The Impact of Fishing Policies and Environmental Conditions. Animals 2021, 11(7), 1901. [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. El estado de la biodiversidad para la alimentación y la agricultura; comisión de recursos genéticos para la alimentación y la agricultura. Evaluaciones: Rome, Italy. 2019. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/1f51259a-7584-4cfc-bab7-d9109361199c/content (accessed on 25 July 2024).
- Food and Agriculture Organization of the United Nations. Phenotypic characterization of animal genetic resources. FAO Animal Production and Health Guidelines. No.11. 2012. Available online: http://14.139.252.116/agrportal/phenochar.htm (accessed on 25 July 2024).
- Caez, J.; González, A.; González, M.A.; Angón, E.; Rodríguez, J.M.; Peña, F.; Barca, C. García, A. Application of multifactorial discriminant analysis in the morphostructural differentiation of wild and cultured populations of Vieja Azul (Andinoacara rivulatus). Turk. J. Zool. 2019, 43, 516-530.
- González, M.A.; Rodríguez, J.M.; Angón, E.; Martínez, A.; García, A.; Peña, F. Characterization of morphological and meristic traits and their variations between two different populations (wild and cultured) of Cichlasoma festae, a species native to tropical Ecuadorian rivers. Archiv. Tierz. 2016, 59, 435-444. [CrossRef]
- Gonzalez-Martinez, A.; Lopez, M.; Molero, H.M.; Rodriguez, J.; Gonzalez, M.; Barba, C.; García, A. Morphometric and meristic characterization of native chame fish (Dormitator latifrons) in Ecuador using multivariate analysis. Animals, 2020, 10, 1805. [CrossRef] [PubMed]
- Teimori, A.; Schulz-Mirbach, T.; Esmaeli, H.R.; Reichenbacher, B. Geographical differentiation of Aphanius dispar (Teleostei: Cyprinodontae) from Southern Iran. J. Zool. Syst. Res. 2012, 50 (4), 289-304. [CrossRef]
- Aguirre, W.W.; Shervette, V.R.; Navarrete, R.; Calle, P.; Agorastos, S. Morphological and Genetic Divergence of Hoplias microlepis (Characiformes: Erythrinidae) in Rivers and Artificial Impoundments of Western Ecuador. Copeia, 2013, 2, 312-323.
- Gonzalez-Martinez, A.; De-Pablos-Heredero, C.; González, M.; Rodriguez, J.; Barba, C.; García, A. Usefulness of discriminant analysis in the morphometric differentiation of six native freshwater species from Ecuador. Animals, 2021, 11, 111,.
- Siddik, M. A. B.; Hanif, M. A.; Chaklader, M. R.; Nahar, A.; Mahmud, S. Fishery biology of gangetic whiting Sillaginopsis panijus (Hamilton, 1822) endemic to Ganges delta, Bangladesh. Egypt. J. Aquat. Res. 2015, 41(4), 307-313. [CrossRef]
- Dauda, A.; Abbaya, H. Y.; Ebegbulem, V. N. Application of multifactorial discriminant analysis of morphostructural differentiation of sheep. Journal of Genetics and Genetic Engineering, 2018, 2(2), 11-16.
- Mattox, G.M.T.; Bifi, A.G.; Oyakawa, O.T. Taxonomic study of Hoplias microlepis (Günther: 1864): a trans-Andean species of trahiras (Ostariophysi: Characiformes: Erythrinidae). Neotrop. Ichthyol. 2014, 12 (2), 343-352. [CrossRef]
- Rodriguez Vizcaino, N.E. Análisis proximal de pescados continentales de mayor consumo humano en Ecuador. BSc. Pontificia Universidad Católica del Ecuador, Quito-Ecuador. 2017. Available online: https://repositorio.puce.edu.ec/items/6ad10381-fd92-4668-9d18-9b8cc91d5892 (accessed on 25 July 2024).
- Werlinger, C. Biologia Marina y Oceanográfica: Conceptos y procesos. Concepción, Chile. Consejo Nacional del Libro y la Lectura. Universidad de Concepción. 2004.
- Lucas Pozo, A. R. Análisis interanual de cinco especies capturadas en el embalse parque lago Chongón, provincia del Guayas, Ecuador, 2008-2018 (Bachelor’s thesis) La Libertad: Universidad Estatal Península de Santa Elena, Ecuador. 2021. Available online: https://repositorio.upse.edu.ec/handle/46000/6634 (accessed on 25 July 2024).
- Ochoa Ubilla, B.Y.; Mendoza Nieto, K.X.; Vivas Moreira, R.; Zambrano, J.U.; Ferrer-Sánchez, Y. Ecuador Structure of catch sizes and length-weight ratio of native fish in the Abras de Mantequilla wetland, Ecuador (in Spanish). Cien. y Tecnol UTEQ 2016, 9 (2), 19-27.
- Pacheco Bedoya, J.L. Aspectos biológicos y pesqueros de las principales especies capturadas en el río Babahoyo y afluentes en el cantón Samborondón de la provincia del Guayas. Available on line: http://www.institutopesca.gob.ec/wp-content/uploads/2018/01/Aspectos.-Biol%C3%B3gicos-y-Pesqueros-R%C3%ADo-Babahoyo-y-Afluentes-Cant%C3%B3n-Samborond%C3%B3n-2015-2.pdf (accessed on 25 July 2024).
- Ferrito, V.; Mannino, M.C.; Pappalardo, A.M.; Tigano, C. Morphological variation among populations of Aphanius fasciatus Nardo, 1827 (Teleostei, Cyprinodontidae) from the Mediterranean. J. Fish Biol. 2007, 70, 1–20. [CrossRef]
- Mir, F.A.; Mir, J.I.; Chandra, S. Phenotypic variation in the Snowtrout Schizothorax richardsonii (Gray, 1832) (Actinopterygii: Cypriniformes: Cyprinidae) from the Indian Himalayas. Contrib. Zool. 2013, 82, 115-122. [CrossRef]
- Dasgupta, S.; Moqbul Hossain, M.D.; Huq, M.; Wheeler, D. Facing the hungry tide: Climate change, livelihood threats, and household responses in coastal Bangladesh. Clim. Change Econ. 2016, 7, 1650007. [CrossRef]
- Hossain, M.; Nahiduzzaman, M.; Saha, D.; Khanam, M.; Alam, M. Landmark-based morphometric and meristic variations of the endangered carp, Kalibaus labeocalbasu, from stocks of two isolated Rivers, the Jamuna and Halda and a hatchery. Zool. Stud. 2010, 49, 556–563.
- Rodriguez, J. Caracterización de la Cichlasoma festae (Vieja Colorada) en la cuenca hidrográfica del Guayas. Ecuador. Bachelor’s thesis. University of Cordoba, Spain, 2017. Available online: https://helvia.uco.es/xmlui/handle/10396/14921 (accessed on 25 July 2024).
- Prado, M.; Bucheli, R.; Calderón, G. Composición, distribución y abundancia del plancton en sistemas fluviales de la provincia de los ríos-ecuador. Boletín Científico y Técnico 2010, 20 (6), 1-52.
- MAGAP. Aspectos pesqueros de las principales especies capturadas en el embalse parque lago Chongón. Instituto Nacional de Pesca 2008-2013.
- Pazmiño-Rodríguez, J.C.; Zambrano-Ganchozo, G.L.; Coello-Burgos, H.A. Modelización de la calidad del agua del estero Aguas Claras, cantón Quevedo, Ecuador. DYNA 2018, 85, 204-214. [CrossRef]
- Zhang, H.; Kang, M.; Wu, J.; Wang, C.; Li, J.; Du, H.; Yang, H.; Wei, O.V. Increasing River Temperature Shifts Impact the Yangtze Ecosystem: Evidence from the Endangered Chinese Sturgeon. Animals 2019, 9, 583. [CrossRef] [PubMed]
- Huayamave, J. Estudio de las Aguas y Sedimentos del Río Daule, en la provincia del Guayas desde el punto de vista Físico químico, orgánico, bacteriológico y toxicológico. Tesis Doctoral. Universidad de las Palmas de Gran Canaria, Departamento de Ingeniería de Procesos. Gran Canaria – España. 2013. Available online: http://hdl.handle.net/10553/11262 (accessed on 25 July, 2024).
- Bermeo, M.; Villa, F.; Toro, P.; Valdiviezo, C. Metales Pesados represa Daule, Peripa. Editorial Grupo Compas S.A. ISBN 978-9942-760-59-3. Guayaquil – Ecuador. 2017.
- Cuero Ordoñez, L.E. (2017). Evaluación de la incidencia de las descargas contaminantes en la calidad del agua del río Quevedo, cantón Quevedo, provincia de los Ríos, año 2016. BSc, Quevedo-Los Rios, Ecuador. Available online: https://repositorio.uteq.edu.ec/items/80926713-fd24-4a8c-9da6-0059335eec0e (accessed on 25 July, 2024).
- Sandal, M. (2019). Efecto de las descargas de aguas residuales de la empresa EMSABA EP sobre la calidad del agua del rio Babahoyo, año 2018. Tesis de maestría. Que-vedo - Ecuador, 2019. Avaiable online: https://repositorio.uteq.edu.ec/server/api/core/bitstreams/8eddc4fa-7955-436c-ac91-d7cfc70f8ea7/content (accessed on 25 July, 2024).
- Less, B.; López, E. Estudios de la Biología de los Peces del Río Vinces. Boletín Científico y Técnico. Instituto Nacional de Pesca 1974, 3 (1), 1- 40.
- Revelo, W. Aspectos biológicos y pesqueros de los principales peces del sistema hídrico de la provincia de los Ríos, durante 2009. Boletín Científico y Técnico 2010, 20 (6), 53-84.
- Canadian Council on Animal Care. Guidelines on the care and use of fish research, teaching and testing. 2005. Available online: https://www.ccac.ca/Documents/Standards/Guidelines/Fish.pdf (accessed on 25 July 2024).
- Norma Española UNE 173300 – “Piscicultura: Guía de prácticas correctas para el sacrificio”. Available online: https://www.mapa.gob.es/es/pesca/temas/calidad-seguridad-alimentaria/pisciculturaguiadepracticascorrectasparaelsacrificio_tcm30-291479.pdf (accessed on 25 July 2024).
- Diodatti, F.C.; Fonseca de Freitas, R.T.; Freato, T.A.; Pérez Ribeiro, P.A.; Solis Murgas, L.D. Parámetros morfométricos en el rendimiento de los componentes corporales de tilapia del Nilo (Oreochromis Niloticus). Anales de Veterinaria (Murcia) 2008, 24, 45–55.
- Nash, R.R.M.; Valencia, A.H.; Geffen, A.J. The Origin of Fulton´s Condition Factor-Setting the Record Straight. Fisheries 2006, 31 (5), 236-238.
- Elliott, N.G.; Haskard, K.; Koslow, J.A. Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of southern Australia. J. Fish Biol. 1995, 46, 202-220. [CrossRef]
- Requiron, E.A.; Torres, M.A.J.; Demayo, C.G. 2012. Applications of relative warp analysis in describing of scale morphology between sexes of the snakehead fish Channa striata. Int. J. Biol.Ecol. Envir. Sci. 2012, 1 (6), 205-209.
- Portillo, JN.; Patulili, R.R.; Lucas, M.M.A.; Alaijos, O.; Demayo, C.G. Body Shape Variation in the Goby.; Glossogobius giuris Collected in Selected Areas in the River of Norzagaray Bulacan Using Landmark-Based Geometric Morphometrics. J. Inform. Math. Sci. 2017, 9 (4), 1109-1116.
- Bussing, W. Peces de las Aguas Continentales de Costa Rica. Editorial de la Universidad de Costa Rica, San Jose, Costa Rica. 1998.
- Laaz Moncayo, E.; Torres Noboa, A. Lista de Peces continentales de la Cuenca del Río Guayas. 2014.
- Gonzáles, A.; Acosta, J.; Andrade, S. (2008). Evaluación de las inundaciones de la cuenca baja del Guayas, datos y manejo. CLIRSEN. In Proceedings of the XI Congreso Ecuatoriano de la Ciencia del Suelo, Quito, Ecuador, 29–31 October.
- Barriga, R. Peces de los Afluentes de la Costa de Ecuador. In Cuencas Pericon-tinentales de Colombia, Ecuador, Perú y Venezuela. Serie Editorial Recursos Hidro-biológicos y Pesqueros Continentales de Colombia; Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt, Bogotá-Colombia. 2015.
- Revelo, W.; Laaz, E. Catálogo de peces de aguas continentales de la provincia de los Ríos – Ecuador. Boletín Especial 2012, 03 (5), 1-56.
- Melvin, G.D.; Dadswell, M.J.; McKenzie, A. Usefulness of meristic and morphometric characters in discriminating populations of American shad (Alosa sapidissima) (Ostreichthys: Clupeidae) inhabiting a marine environment. Can. J. Fish. Aquat. Sci. 1992, 49, 266-280. [CrossRef]
- Ujjania, N.C.; Kohli, M.P.S. Landmark-based morphometric analysis for selected species of indian major carp (catla catla.; ham. 1822). Int. J. Food Agri. Vet. Sci. 2011, 1, 64-74.
- Froese, R. Cube law, condition factor and weight–length relationships: history, meta-analysis and recommendations. J. Appl. Ichth. 2006, 22, 241-253. [CrossRef]
- Trudel, M.; Tucker, S.; Morris, J.; Higgs, D.; Welch, D. Indicators of energetic status in juvenile coho and chinook salmon. N. Am. J. Fish. Manag. 2005, 25, 374-390. [CrossRef]
- Rennie, M.D.; Verdon, R. Evaluation of condition indices for the lake whitefish, Coregonus clupeaformis. N. Am. J. Fish. Manag. 2008, 28, 1270-1293. [CrossRef]
- Cureton, J.C.; Broughton, R.E. Rapid morphological divergence of a stream fish in response to changes in water flow. Biol. Lett. 2014, 10, 20140352. [CrossRef]
- Radojković, N.; Marinoić, Z.; Milosković, A.; Radenković, M.; Duretanović, S.; Luji, J.; Simi, V. Effects of Stream Damming on Morphological Variability of Fish: Case Study on Large Spot Barbell Barbus balcanicus. Turk. J. Fish. & Aquat. Sci. 2018, 19 (3), 231-239.
- Libay, CP.; Ratunil, V.B. Jr.; Ebarsabal, G.A.; Gamboa, G.Z. Jr.; Borja, E.A.; Ga, J.A.A.; Eclipse, M.G.E.; Mahomoc, D.Q.; Cabuga, C.C. Jr. Geometric morphometric analysis in determining phenotypic variability of Bugwan, (Hypseleotris agilis, Herre) in Lake Mainit, Philippines. Int. J. Biosci. 2019, 14 (6), 61-70.
- Caldecutt, WC.; Adams, DC. Morphometrics of trophic osteology in the three spine stickleback, Gasterosteus aculeatus. Copeia 1998, 4, 827-838. [CrossRef]
- Webb, PW. Locomotor patterns in the evolution of actinopterygian fish. Am. Zool. 1982, 22, 329-342. [CrossRef]
- Anders, P.J. Conservation aquaculture: An adaptive approach to prevent extinction of an endangered white sturgeon population. Fisheries 1998, 23 (11), 28-31.
Figure 1.
Guayas Hydrographic Basin (CHG); Daule-Peripa dam; Quevedo river and Babahoyoriver (n = sampling size).
Figure 1.
Guayas Hydrographic Basin (CHG); Daule-Peripa dam; Quevedo river and Babahoyoriver (n = sampling size).
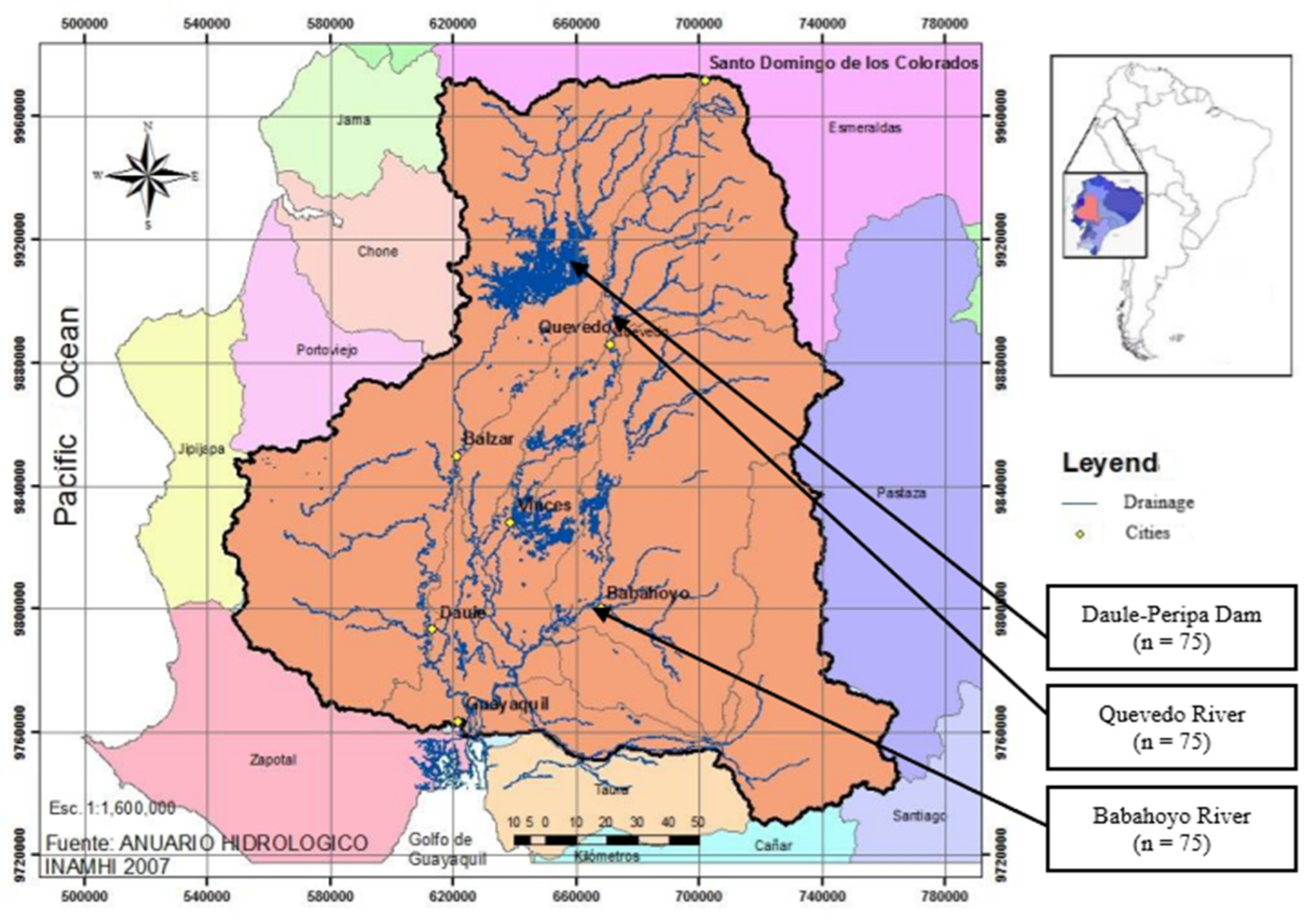
Figure 2.
Location of 19 anatomical landmark points1 designed on the left view of the Hoplias microlepis. 1 Landmark points: 1: most cranial point of the upper pre-maxilla; 2: most cranial point of the lower pre-maxilla; 3: commissure of the mouth; 4: anterior edge of the eye; 5: posterior edge of the eye; 6: end of operculum; 7: down of operculum; 8: origin of pectoral fin; 9: origin of dorsal fin; 10: ending of dorsal fin; 11: dorsal origin of caudal fin; 12: ventral origin of caudal fin; 13: most cranial point of caudal peduncle; 14: most caudal point of caudal peduncle; 15: ending of anal fin; 16: origin of anal fin; 17: anal opening; 18: origin of pelvic fin; 19: ending of the pelvic fin radius.
Figure 2.
Location of 19 anatomical landmark points1 designed on the left view of the Hoplias microlepis. 1 Landmark points: 1: most cranial point of the upper pre-maxilla; 2: most cranial point of the lower pre-maxilla; 3: commissure of the mouth; 4: anterior edge of the eye; 5: posterior edge of the eye; 6: end of operculum; 7: down of operculum; 8: origin of pectoral fin; 9: origin of dorsal fin; 10: ending of dorsal fin; 11: dorsal origin of caudal fin; 12: ventral origin of caudal fin; 13: most cranial point of caudal peduncle; 14: most caudal point of caudal peduncle; 15: ending of anal fin; 16: origin of anal fin; 17: anal opening; 18: origin of pelvic fin; 19: ending of the pelvic fin radius.

Figure 3.
Sexual maturity index by sampled population of Guanchiche. a, b, superscript letters indicate significative differences among populations (P < 0.01).
Figure 3.
Sexual maturity index by sampled population of Guanchiche. a, b, superscript letters indicate significative differences among populations (P < 0.01).
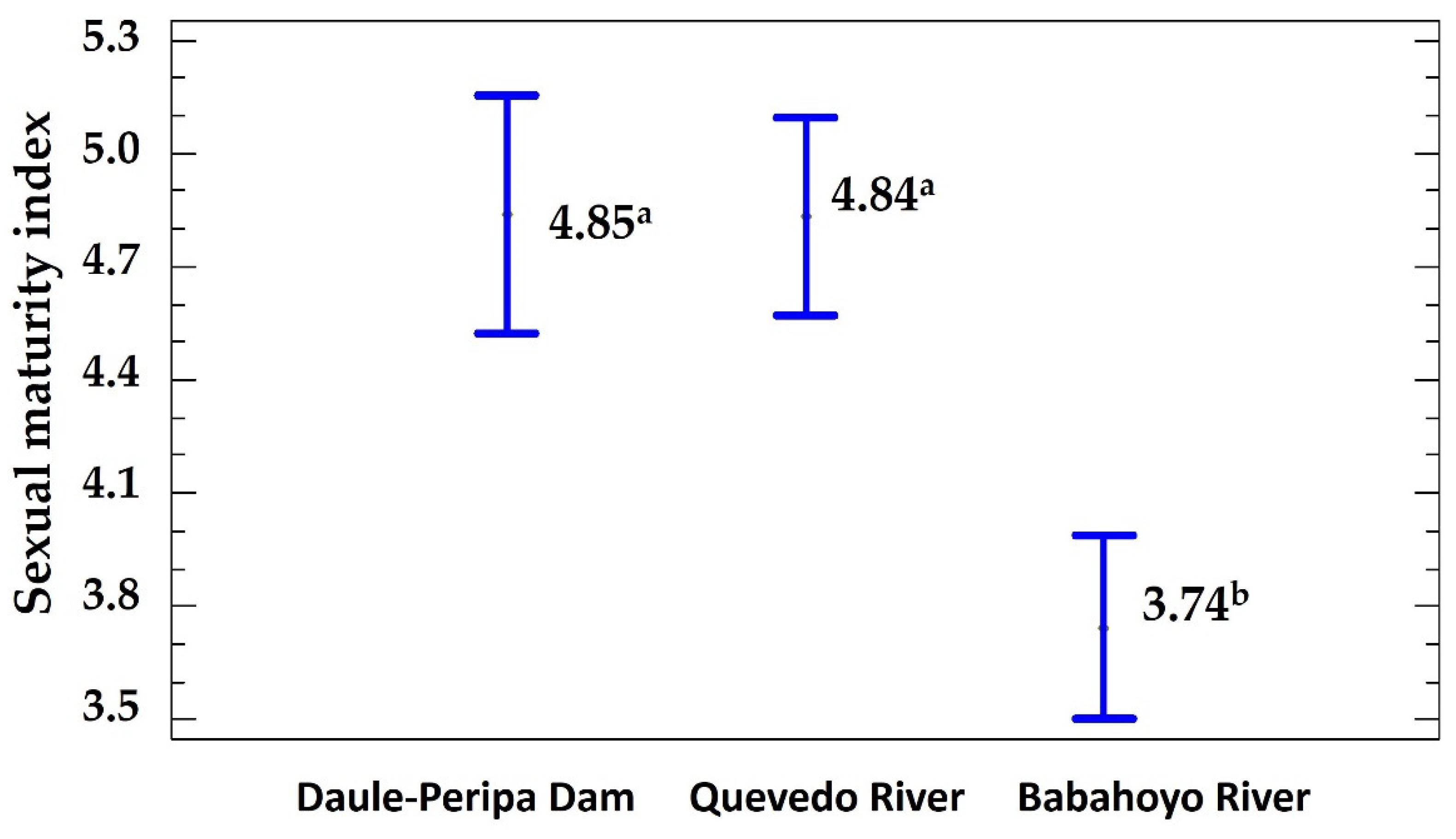
Figure 4.
Plot of the individual observation discriminant scores obtained with the canonical discri-minant function (a) and Cluster from Mahalanobis distances (b) for three populations of of Hoplias microlepis.
Figure 4.
Plot of the individual observation discriminant scores obtained with the canonical discri-minant function (a) and Cluster from Mahalanobis distances (b) for three populations of of Hoplias microlepis.
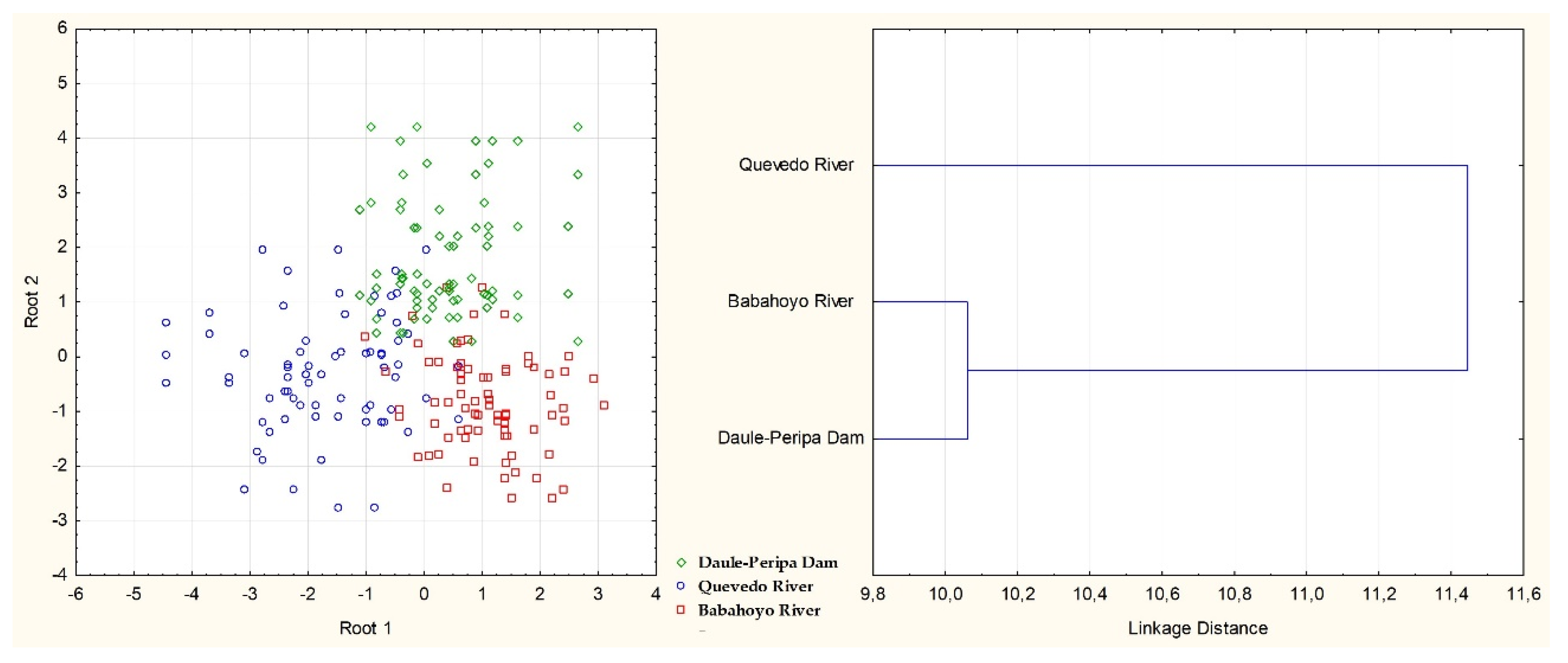

Table 1.
Guayas Hydrographic Basin sampling sites characterization (CHG).
| Habitat | Daule-Peripa dam | Quevedo river | Babahoyo river |
|---|---|---|---|
| Locality description | Reservoir of species. All species | White-water with native species: dama, bocachico, ratón, vieja azul, vieja colorada, among others | Slow-flowing waters which favour benthic species rearing such as guanchiche. |
| Depth | The maximun depth is 85 m with body water of 295 km | The average depth is 5 m in the rainy season, while its depth decreases to 1.92 m in the dry season, with a route of 163 km | It has an average of 5m of depth and 40 km of route |
| Endogeneity and competition | Carnivorous species such as cachama (Colossoma macropomum) and paiche (Arapaima gigas) | Non-native species like tilapia (Oreochromis niloticus) displace native species | Guanchiche coexists with tilapia (Oreochromis niloticus) |
| Plankton-insects | Maximum concentration in October and minimum in August | Rich in phytoplankton | Rich in zooplankton and larvae of different species |
| Fishing vessels | Bongo1, bote1, lancha1 | Balsa, canoe, bongo1, bote1 | Balsa, canoe, bongo1, bote1 |
| Fishing methods | Riverbank manual thrownets, trammel nets, fishing spear | Trawl line, riverbank manual thrownets, trammel nets, thrownets, fishhooks | Trammel nets, thrownets |
| Fishing pressure | Low fishing pressure. The largest reservoir of native freshwater species in Ecuador. | Traditional fishing. Medium fishing pressure of native species | Traditional fishing. Strong fishing pressure of native species |
115-180 hp motor boats.
Table 2.
Physicochemical1 characteristics water in three sampling sites of the Guayas River Basin-Ecuador.
Table 2.
Physicochemical1 characteristics water in three sampling sites of the Guayas River Basin-Ecuador.
| Indicators | Daule-Peripa dam | Quevedo river | Babahoyo river |
|---|---|---|---|
| Physical / chemical parameters | |||
| pH | 7.42 | 8.23 | 7.44 |
| Electric conductivity (μS/cm) | 104.9 | 95.3 | 114.3 |
| Temperature (°C) | 19.3 | 19.2 | 27.25 |
| Turbidity (NTU) | 4.9 | 7.4 | 14.1 |
| Colour (UC Pt-Co) | 16 | 5 | 63 |
| Total dissolved solids (mg/l) | 112 | 96 | <150 |
| Alkalinity (mg/l) | 51.80 | 47.73 | 40.2 |
| Total hardness (mg/l) | 38.25 | 41.43 | 29.3 |
| Calcium hardness (mg/l) | 24.22 | 27.97 | 25.9 |
| CO2 (mg/l) | 3.93 | 0.55 | 1.97 |
| SIO2 (mg/l) | 35 | 20.1 | 28 |
| Cations | |||
| Ca2+ (mg/l) | 9.7 | 11.2 | 9.25-12.1 |
| Mg2+ (mg/l) | 3.38 | 3.28 | 3.93 |
| Metals | |||
| Na+(mg/l) | 7.88 | 5.48 | 7.43 |
| Mn (mg/l) | <0.016 | <0.016 | X |
| Cu (mg/l) | <0.024 | <0.024 | <0.03 |
| Cr (mg/l) | < 1 | < 1 | <0.082 |
| Fe (mg/l) | 0.70 | 0.84 | 1.89 |
| K+ (mg/L) | 2.815 | 2.065 | 1.9-1.7 |
| Pb (mg/l) | <1 | <1 | <0.050 |
| Hg (ug/l) | 0 | <0.001 | 0 |
| Cd (mg/l) | <0.010 | <0.001 | <0.010 |
| Anions | |||
| (HCO3)- mg/l | 51.80 | 42.92 | 32.83 |
| (CO3)2- mg/l | 0 | 4.81 | 0 |
| (SO4)2- mg/l | 7.84 | 12.32 | 12 |
| Cl- (mg/l) | <5.30 | <5.3 | <5.0 |
| NH4+ (mg/l9) | 0.33 | 0.03 | <0.0014 |
| NO3- (mg/l) | <0.21 | 0.92 | <0.84 |
| NO2- (mg/l | <0.05 | <0.05 | 0.072 |
| (PO4)3- (mg/l | <0.50 | <0.50 | 2 |
| F-(mg/l) | 0.12 | 0.5 | 0.17 |
| Organic constituents | |||
| Biochemical oxygen demand (BOD5, ml/g) | 5.77 | 3.95 | 2.47 |
| Chemical oxygen demand (DQO, mg/l) | 17.38 | 12.98 | 4.6 |
| Dissolved oxygen (OD, mg/l) | 1.80 | 7.05 | 6.53 |
| OD (% sat) | 48 | 87.6 | 82.7 |
Table 3.
Morphometric measurements and meristic counts of Hoplias microlepis used in this study.
| Measurement | Description | Acronym |
|---|---|---|
| Weight | Total weight including the gut and gonads | BW |
| Total length | Tip of the upper jaw to the top of the caudal superior end of the caudal fin | TL |
| Standard length | Tip of the upper jaw to the tail base | SL |
| Head length | From the front of the upper lip to the posterior end of the opercula membrane | HL |
| Eye diameter | The greatest bony diameter of the orbit | ED |
| Pre-orbital length | Front of the upper lip to the cranial eye edge | Pre-OL |
| Pre-dorsal fin length | Front of the upper lip to the origin of the dorsal fin | Pre-DL |
| Pre-pectoral fin length | Front of the upper lip to the origin of the pectoral fin | Pre-PcL |
| Pre-pelvic fin length | Front of the upper lip to the origin of the pelvic fin | Pre-PvL |
| Pre-anal fin length | Front of the upper lip to the origin of the anal fin | Pre-AL |
| Dorsal fin length | From the base of the first dorsal spine to the base of the last dorsal ray | DFL |
| Dorsal fin ray length | From the base to the tip of the fifth dorsal ray | DFRL |
| Pectoral fin length | From the base to the tip of the pectoral fin | PcFL |
| Pelvic fin length | From the base to the tip of the pelvic fin | PvFL |
| Anal fin length | From the base of the first anal spine to the base of the last anal ray | AFL |
| Anal fin ray length | From the base to the tip of the last anal ray | AFRL |
| Upper jaw length | Straight line measurement between the snout tip and posterior edge of maxilla | UJL |
| Body perimeter 1 | Body perimeter at the level of the first ray of the dorsal fin | P1 |
| Body perimeter 2 | Body perimeter at the level of the first ray of the anal fin | P2 |
| Body perimeter 3 | Body perimeter at the level of the last ray of the dorsal fin | P3 |
| Body perimeter 4 | Body perimeter at the level of the last ray of the anal fin | P4 |
| Body width 1 | Straight line measurement from side to side at the level of the base of the first dorsal spine | LC1 |
| Body width 2 | Straight line measurement from side to side at the level of the base of the first anal spine | LC2 |
| Body width 3 | Straight line measurement from side to side at the level of the base of the last dorsal ray | LC3 |
| Body width 3 | Straight line measurement from side to side at the level of the base of the last anal ray | LC4 |
| Body depth 1 | Body depth at the level of the first ray of the dorsal fin | AC1 |
| Body depth 2 | Body depth at the level of the first ray of the anal fin | AC2 |
| Body depth 3 | Body depth at the level of the first radius of the caudal fin | AC3 |
| Dorsal fin rays | Number of thorns in the dorsal fin | DFR |
| Pectoral fin rays | Number of thorns in the pectoral fin | PcFR |
| Pelvic fin rays | Number of thorns in the pelvic fin | PvFR |
| Anal fin rays | Number of thorns in the anal fin | AFR |
| Caudal fin rays | Number of thorns in the caudal fin | CFR |
Table 4.
Descriptive statistics of body measurements (original data) of Hoplias microlepis from three different sampling sites (Mean ± SE (CV, %)).
Table 4.
Descriptive statistics of body measurements (original data) of Hoplias microlepis from three different sampling sites (Mean ± SE (CV, %)).
| Parameter1 | All | Daule-Peripa Dam (A) |
Quevedo River (B) |
Babahoyo River (C) |
p-Value2 | |
|---|---|---|---|---|---|---|
| A x C | B x C | |||||
| BW | 442.66±29.52 (68.97) | 778.65±74.39 (48.71) | 398.59±31.68 (48.34) | 281.18±20.49 (48.33) | *** | * |
| K | 1.47±0.1 (69.7) | 1.84±0.12 (34.15) a | 1.06±0.1 (60.36) | 1.59±0.2 (83.48) | *** | *** |
| TL | 37.82±0.69 (18.97) | 41.55±1.18 (14.52) | 40.34±0.92 (13.85) | 33.49±1.03 (20.38) | n.s. | n.s. |
| SL | 32.14±0.6 (19.28) | 34.88±1.12 (16.35) | 34.18±0.81 (14.38) | 28.79±0.9 (20.8) | n.s. | n.s. |
| HL | 8.31±0.15 (18.77) | 8.45±0.36 (21.49) | 9.03±0.23 (15.28) | 7.63±0.19 (16.33) | * | n.s. |
| ED | 1.34±0.03 (23.71) | 1.33±0.06 (22.19) | 1.43±0.05 (22.96) | 1.27±0.05 (24.42) | n.s. | n.s. |
| Pre-OL | 1.34±0.04 (31.79) | 1.48±0.06 (22.33) | 1.47±0.08 (32.15) | 1.14±0.05 (30.86) | n.s. | * |
| Pre-DL | 16.16±0.3 (19.25) | 17.59±0.57 (16.41) | 17.31±0.43 (15.11) | 14.35±0.42 (19.2) | n.s. | n.s. |
| Pre-PcL | 8.33±0.17 (20.79) | 8.25±0.4 (24.67) | 8.97±0.22 (14.71) | 7.83±0.26 (21.8) | n.s. | n.s. |
| Pre-PvL | 11.08±0.22 (20.63) | 12.95±0.46 (18.22) | 11.19±0.42 (22.87) | 9.89±0.12 (7.83) | *** | *** |
| Pre-AL | 25.29±0.55 (22.44) | 28.26±1.04 (18.8) | 27.86±0.66 (14.34) | 21.37±0.72 (22.37) | n.s. | n.s. |
| DFL | 3.58±0.08 (22.28) | 3.94±0.19 (24.64) | 3.81±0.13 (21.3) | 3.16±0.06 (12.87) | *** | *** |
| DFRL | 3.99±0.08 (20.37) | 3.59±0.13 (18.92) | 4.14±0.15 (22.43) | 4.10±0.11 (17.44) | n.s. | n.s. |
| PcFL | 2.80±0.08 (28.85) | 2.80±0.18 (33.54) | 3.22±0.13 (23.75) | 2.46±0.09 (23.86) | ** | n.s. |
| PvFL | 3.63±0.09 (25.49) | 4.03±0.16 (20.01) | 4.26±0.12 (17.66) | 2.86±0.07 (16.34) | ** | ** |
| AFL | 2.99±0.08 (28.73) | 3.13±0.18 (29.24) | 3.37±0.15 (27.26) | 2.58±0.08 (21.67) | ** | ** |
| AFRL | 2.95±0.07 (24.3) | 3.04±0.12 (20.54) | 3.56±0.09 (15.25) | 2.39±0.06 (15.95) | ** | * |
| UJL | 2.18±0.05 (24.51) | 2.14±0.11 (26.75) | 1.98±0.07 (22.55) | 2.37±0.08 (21.96) | n.s. | n.s. |
| AC1 | 3.56±0.05 (14.14) | 3.95±0.06 (7.25) | 3.50±0.06 (10.3) | 3.39±0.09 (17.32) | *** | ** |
| AC2 | 4.77±0.06 (12.22) | 5.19±0.08 (8.11) | 4.59±0.07 (9.89) | 4.68±0.1 (13.83) | * | * |
| AC3 | 5.44±0.06 (11.69) | 5.98±0.09 (7.85) | 5.22±0.08 (9.11) | 5.31±0.1 (12.52) | * | * |
| P1 | 13.66±0.19 (14.23) | 13.26±0.49 (18.91) | 14.03±0.17 (7.28) | 13.6±0.32 (15.73) | n.s. | *** |
| P2 | 14.19±0.24 (17.69) | 14.23±0.17 (6.09) | 14.13±0.55 (23.89) | 14.21±0.35 (16.56) | *** | * |
| P3 | 12.36±0.13 (10.91) | 12.00±0.24 (10.35) | 12.59±0.23 (10.93) | 12.38±0.21 (11.06) | n.s. | n.s. |
| P4 | 8.75±0.09 (11.17) | 8.60±0.18 (10.8) | 8.85±0.16 (11.33) | 8.75±0.15 (11.36) | n.s. | n.s. |
| LC1 | 3.11±0.04 (12.55) | 3.10±0.07 (11.39) | 3.13±0.07 (13.07) | 3.11±0.06 (13.01) | n.s. | n.s. |
| LC2 | 3.13±0.04 (13.52) | 3.22±0.06 (9) | 3.06±0.07 (14.86) | 3.15±0.07 (14.6) | * | n.s. |
| LC3 | 2.54±0.04 (17.36) | 2.40±0.07 (14.86) | 2.57±0.08 (18.4) | 2.60±0.07 (17.31) | n.s. | n.s. |
| LC4 | 1.23±0.04 (31.49) | 1.09±0.07 (34.31) | 1.33±0.06 (28.57) | 1.24±0.06 (31.25) | n.s. | n.s. |
1 BW = body weight; K = Fulton´s factor; TL = total length; SL = standard length; HL = head length; ED = eye diameter; Pre-OL = pre-orbital length; Pre-DL = pre-dorsal fin length; Pre-PcL = pre-pectoral fin length; Pre-PvL = pre-pelvic fin length; Pre-AL = pre-anal fin length; DFL = dorsal fin length; DFRL = dorsal fin ray length; PcFL = pectoral fin length; PvFL = pelvic fin length; AFL = anal fin length; AFRL = anal fin ray length; UJL = upper jaw length; AC1 = body depth 1; AC2 = body depth 2; AC3 = body depth 3; P1 = body perimeter 1; P2 = body perimeter 2; P3 = body perimeter 3; LC1 = body width 1; LC2 = body width 2; LC3 = body width 3; LC4 = body width 4. 2 p-Value: *p < 0.05; ** p < 0.01; *** p < 0.001; n.s. not significant differences between columns.
Table 5.
Descriptive statistics of the meristic characters (original data) of Hoplias microlepis from three different sites (Mean ± SE (CV)).
Table 5.
Descriptive statistics of the meristic characters (original data) of Hoplias microlepis from three different sites (Mean ± SE (CV)).
| Parameter 1 | All | Daule-Peripa Dam (A) |
Quevedo River (B) |
Babahoyo River (C) |
p-Value2 | |
|---|---|---|---|---|---|---|
| A × C | B × C | |||||
| DFR | 12.96±0.11 (8.73) | 12.85±0.25 (10.02) | 13.00±0.15 (7.25) | 13.00±0.18 (9.24) | n.s. | n.s. |
| PcFR | 10.71±0.20 (19.79) | 10.50±0.47 (22.74) | 10.84±0.34 (19.15) | 10.73±0.31 (18.90) | n.s. | n.s. |
| PvFR | 7.79±0.05 (6.37) | 7.77±0.10 (6.62) | 7.86±0.06 (4.41) | 7.73±0.09 (7.57) | * | n.s. |
| AFR | 9.45±0.12 (13.58) | 9.38±0.25 (13.50) | 9.43±0.20 (12.87) | 9.50±0.21 (14.45) | n.s. | n.s. |
| CFR | 16.96±0.08 (4.99) | 16.92±0.19 (5.77) | 17.00±0.12 (4.38) | 16.95±0.13 (5.08) | * | * |
| G | 3.65±0.05 (14.61) | 3.62±0.11 (15.80) | 3.65±0.09 (14.75) | 3.68±0.08 (14.07) | n.s. | n.s. |
1 DFR: Dorsal fin rays; PcFR: Pectoral fin rays; PvFR: Pelvic fin rays; AFR: Anal fin rays; CFR: Caudal fin rays; G: Gill arches. 2 p-Value: *p < 0.05; n.s. not significant differences between columns.
Table 6.
Discriminant functions for truss measurements of Hoplias microlepis from three populations.
Table 6.
Discriminant functions for truss measurements of Hoplias microlepis from three populations.
| Parameter1 | Wilks’ Lambda | Partial Lambda | F-Remove | p-Level2 | Toler | 1-Toler |
|---|---|---|---|---|---|---|
| AFRL | 0.22 | 0.85 | 7.60 | ** | 0.69 | 0.31 |
| AC3 | 0.22 | 0.85 | 7.76 | ** | 0.90 | 0.10 |
| Pre-PvL | 0.24 | 0.78 | 12.37 | *** | 0.72 | 0.28 |
| PvFL | 0.22 | 0.85 | 7.65 | ** | 0.68 | 0.32 |
| UJL | 0.20 | 0.92 | 4.09 | * | 0.88 | 0.12 |
| DFRL | 0.21 | 0.90 | 4.79 | * | 0.82 | 0.18 |
| LC4 | 0.20 | 0.94 | 3.05 | n.s. | 0.51 | 0.49 |
| P1 | 0.20 | 0.94 | 2.77 | n.s. | 0.94 | 0.06 |
| HL | 0.19 | 0.96 | 1.85 | n.s. | 0.59 | 0.41 |
| TL | 0.19 | 0.96 | 1.85 | n.s. | 0.68 | 0.32 |
| ED | 0.20 | 0.94 | 2.68 | n.s. | 0.90 | 0.10 |
| LC3 | 0.19 | 0.97 | 1.52 | n.s. | 0.50 | 0.50 |
| Pre-PcL | 0.19 | 0.97 | 1.33 | n.s. | 0.59 | 0.41 |
| Pre-DL | 0.19 | 0.96 | 1.73 | n.s. | 0.64 | 0.36 |
| AFL | 0.19 | 0.97 | 1.21 | n.s. | 0.60 | 0.40 |
1 TL = total length; HL = head length; ED = eye diameter; Pre-DL = pre-dorsal fin length; Pre-PcL = pre-pectoral fin length; Pre-PvL = pre-pelvic fin length; DFRL = dorsal fin ray length; PvFL = pelvic fin length; AFL = anal fin length; AFRL = anal fin ray length; UJL = upper jaw length; AC3 = body depth 3; P1 = body perimeter 1; LC3 = body width 3; LC4 = body width 4. 2 p-Value: *p < 0.05; ** p < 0.01; *** p < 0.001; n.s. not significant differences between columns.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



