
Submitted:
02 August 2024
Posted:
02 August 2024
You are already at the latest version
Abstract
The composition of the gut microbiota varies among end-stage renal disease (ESRD) patients on the basis of their mode of renal replacement therapy (RRT), with notably more pronounced dysbiosis in those undergoing hemodialysis (HD). Interventions such as dialysis catheters, unstable hemodynamics, strict dietary restrictions, and pharmacotherapy significantly alter the intestinal microenvironment, thus disrupting the gut microbiota composition in HD patients. The gut microbiota may influence HD-related complications, including cardiovascular disease (CVD), infections, anemia, and malnutrition, through mechanisms such as bacterial translocation, immune regulation, and the production of gut bacterial metabolites, thereby affecting both the quality of life and the prognosis of patients. This review focuses on alterations in the gut microbiota and its metabolites in HD patients. Additionally, understanding the impact of the gut microbiota on the complications of HD could provide insights into the development of novel treatment strategies to prevent or alleviate complications in HD patients.
Keywords:
gut microbiota
; hemodialysis
; complications
; uremic toxins
Introduction
The number of patients with chronic kidney disease (CKD) continues to increase, with approximately 10% of the world’s adult population suffering from CKD and causing 1.2 million deaths each year [1]. Some CKD patients experience end-stage renal disease (ESRD), and these patients often need renal replacement therapy (RRT) to maintain life. By 2030, the global use of RRT is expected to reach 5.439 billion people [2]. Hemodialysis (HD) remains the most commonly used RRT [3]. HD patients face a series of challenges, such as cardiovascular disease (CVD), infections, anemia, and protein-caloric malnutrition [4]. As these complications are often not effectively solved, the mortality of HD patients remains high [5].
The human microbiome includes bacteria, fungi, and viruses, which form a large and complex ecosystem [6]. The gut microbiota directly or indirectly affects human health, and a complex bidirectional relationship between the gut microbiota and the heart, brain, and kidney has been confirmed [7,8,9]. The gut microbiota and kidney can interact through metabolic and immune mechanisms to form a bidirectional regulatory axis known as the gut‒kidney axis [10]. On the one hand, uremia can change the composition of the gut microbiota and function (known as gut microbiota dysbiosis) [11]. On the other hand, gut microbiota dysbiosis can disrupt the intestinal barrier, resulting in increased translocation of endotoxins and gut-derived uremic toxins and activation of inflammatory responses, leading to various complications [12,13].
This review discusses the changes in the gut microbiota and its metabolites in HD patients, as well as the effects of the gut microbiota on HD complications, to provide insight into treatment and diagnosis from the perspective of the gut microbiota.
The Gut Microbiota and HD
The gut microbiota plays a crucial role in helping the host resist exogenous bacteria and maintaining the integrity of the intestinal barrier and immune homeostasis [14,15,16]. In healthy people, Firmicutes and Bacteroidetes account for approximately 90% of the microbiome, followed by Actinobacteria, Proteobacteria, Fusobacteria, and Verrucomicrobia [17].
The gut microbiota of ESRD patients (predialysis and postdialysis) undergoes profound changes[18]. This is related to the rapid decline in renal function in ESRD patients, the accumulation of toxins in the body, the large amount of urea circulating into the gut lumen, and the stimulation of the overproduction of urease-containing bacteria [19,20]. Special dietary restrictions (reduced intake of fruits, vegetables, and dairy products), comorbidities (diabetes, hypertension, etc.), and medications (phosphate binders, iron, and antibiotics) can also lead to gut microbiota dysbiosis [11,21,22,23]. In HD, some waste products in the blood are removed through the semipermeable membrane, which means that there are vascular access interventions (arteriovenous fistula/graft or venous catheter), inadequate dialysis, and low immunity, which may further aggravate gut microbiota disorders [24,25]. In addition, during the process of dialysis, ultrafiltration or the administration of anticoagulants may induce hemodynamic instability in the gastrointestinal tract, which can cause intestinal barrier disturbances [26,27]. Damage to the intestinal barrier can result in increased translocation of the gut microbiota and toxic metabolites, leading to microinflammation in HD patients [25].
Proteobacteria and Firmicutes were increased in HD patients compared with healthy controls [11]. This observation is consistent with recent findings reported by Wu et al. [28]. However, some investigators have shown that the abundance of Firmicutes is significantly reduced in HD patients [25,28]. The abundance of Bacteroidetes shows different trends in adult and pediatric HD patients [28,29]. The differences in these research results may be due to variations in the subjects’ age, genetic history, diet, lifestyle, and dialysis adequacy. Owing to the lack of comparative studies before and after dialysis in patients with ESRD, we cannot accurately determine the specific causes of the microbiome changes mentioned above. These changes may be caused by dialysis treatment itself, by ESRD itself, or even by the combined effects of both. To gain a deeper understanding of these changes, Lou et al. specifically selected ESRD patients who had not yet started dialysis, peritoneal dialysis (PD) patients, and HD treatment patients for a detailed comparative analysis, and their findings indicated that HD treatment had a more significant effect on the gut microbiota of ESRD patients. Their study revealed that HD significantly increased the proportion of beneficial bacteria in the microbial community but also stimulated the growth and reproduction of certain potentially pathogenic bacteria, which may introduce new risks to the stability of ecosystems [30]. At the phylum level, HD patients presented the lowest levels of Bacteroidetes[30]. At the genus level, there was a decrease in the abundance of Prevotella and Paraprevotella and an increase in the abundance of Akkermansia, Coprococcus, Acinetobacter, Proteus, and Pseudomonas [30]. Prevotella and Paraprevotella, two anaerobic gram-negative rods associated with infections of the gastrointestinal, respiratory and urinary tracts, are rich in peptidases[31], which degrade proteins and produce large amounts of ammonia [32]. A high concentration of ammonia destroys intestinal epithelial tight junctions, leading to intestinal mucosal injury [33]. Akkermansia and Coprococcus are associated with the production of short-chain fatty acids (SCFAs). Akkermansia is a promising probiotic resident in the mucus layer that reduces inflammation and improves host metabolism [34]. Studies have shown that increasing the abundance of Akkermansia may reduce the chronic inflammatory state of CKD patients [35]. Acinetobacter is an important opportunistic pathogen in hospitalized patients [36]. Proteus, a family of Enterobacteriaceae, is a common commensal bacterium in the gastrointestinal tract that secretes virulence factors such as the protease ZapA and is therefore potentially pathogenic [37]. The abundance of Pseudomonas was positively correlated with plasma tryptophan levels [38]. Pseudomonas is found in the blood of ESRD patients and is associated with increased levels of inflammatory markers (high-sensitivity C-reactive protein (CRP) and IL-6) [39]. He et al. reported that HD increased the abundance of the beneficial bacteria Bifidobacterium and Lactobacillus acidophilus and reduced the abundance of the pathogenic bacteria Escherichia coli and Enterococcus faecalis compared with those in ESRD patients who did not undergo dialysis[40].
The immune system, including adaptive and innate immunity, plays a crucial role in the gut‒kidney axis. HD can induce T-cell-mediated immunodeficiency, thereby impairing the host immune system [41]. HD-related factors (such as the dialysis process, frequency, ultrafiltration, and intradialytic exercise) and non-HD-related factors (uremia and medication) can both alter the intestinal barrier [42]. Additionally, gut microbiota dysbiosis also plays a role in disrupting the intestinal barrier, further increasing the translocation of bacterial components and toxic metabolites, which may activate the innate immune system and systemic inflammation [43]. After intestinal barrier integrity is compromised, there is a substantial influx of lipopolysaccharide (LPS) from the intestines into the circulatory system through the compromised intestinal wall [44]. LPS is a component of the cell wall of Escherichia coli and Enterococcus faecalis, which can bind to Toll-like receptors (TLRs), activate the immune system, increase the production of inflammatory cytokines, and thus exacerbate inflammatory responses [45,46].
Gut microbiota dysbiosis can also result in alterations in microbial metabolites. On the one hand, the production of beneficial SCFAs in HD patients is reduced, which may be related to the reduction in SCFA-producing bacteria such as Lactobacillus, Prevotellaceae and Ruminococcus [47,48]. The plasma SCFA levels are linked to the host’s energy homeostasis and immune system [49,50]. SCFAs, including acetate, propionate and butyrate, serve as major sources of nutrients for colon cells[49] and can be absorbed into the bloodstream through the intestinal lumen for transport to distant organs, such as the heart, kidney, muscle and adipose tissue, where they provide a source of energy for host metabolism[51,52,53]. In addition, SCFAs also have anti-inflammatory effects [54,55]. Among them, butyrate has potent anti-inflammatory effects and reduces the levels of the inflammatory factors tumor necrosis factor-α (TNF-α) and IL-6 by inhibiting the activation of the nuclear factor kappa B (NF-κB) cell signaling pathway[50]. On the other hand, the levels of uremic toxins such as p-cresyl sulfate (pCS) and indoxyl sulfate (IS) in the plasma of HD patients are elevated [56], which is linked to the proliferation of bacteria that produce urease, uricase, p-cresol, and indoles (specifically, Clostriadiaceae and Enterobacteriaceae) [47]. p-Cresol is a byproduct of the putrefactive metabolism of tyrosine. Indole is a compound produced by the fermentation of tryptophan. When p-cresol and indole are absorbed by the body, most of them are further metabolized by the liver. p-Cresol is oxidized into pCS, while indole is oxidized into IS. In normal renal function, these two kinds of metabolites are excreted from the urine [57]. The protein binding rate of pCS and IS is as high as 95%, which leads to limited clearance of HD and is significantly associated with poor prognosis [58]. Furthermore, elevated levels of the gut-derived microbial metabolite trimethylamine N-oxide (TMAO) are closely associated with cardiovascular and all-cause mortality [59,60]. TMAO is a small-molecule, water-soluble poison metabolized by quaternary amines (choline and L-carnitine). Quaternary amine metabolism produces trimethylamine (TMA), which is converted to TMAO by flavin-containing monooxygenase 3 (FMO3) [61]. The plasma TMAO concentration in HD patients is more than 20 times greater than that in patients with normal kidney function [62,63]. In conclusion, higher levels of uremic toxins can have harmful effects on HD patients [64,65].
The Gut Microbiota and CVD
CVD has a high mortality rate among HD patients [66]. Research has revealed that the relative risk of CVD mortality in HD patients is 20 times greater than that in the general population [67]. This is mainly because HD patients are usually accompanied by a series of metabolic disorders, including hypertension, hyperglycemia, and hyperlipidemia. In addition, the chronic inflammatory state, oxidative stress, and uremic toxins, among other nontraditional risk factors, play key roles in the development of CVD in HD patients [68]. After HD treatment, the increase in intestinal permeability leads to the migration of the gut microbiota and metabolites into the bloodstream, resulting in an increase in inflammatory cytokines. Systemic inflammation can promote the development and progression of CVD [69,70] (Figure 1).
Several studies have revealed a significant correlation between the gut microbiota composition and CVD incidence. Among them, Sumida et al. studied the composition of circulating microorganisms in HD patients who died from cardiovascular events and reported that the abundance of Actinobacteria increased while that of Proteobacteria decreased [71]. Similar findings have also been documented in comparative studies involving cohorts of healthy individuals and those diagnosed with CVD [72]. The proportions of the Actinobacteria and Proteobacteria phyla are significantly correlated with the levels of nuclear factor erythroid 2-related factor 2 (Nrf2) in the blood [71]. Nrf2 is a key regulator of the antioxidant response and plays a crucial role in immune regulation. Dysregulation of Nrf2 activation is associated with the occurrence and progression of CVD [73]. In ESRD patients, analysis of CVD mortality data revealed a decrease in Bacteroides and Phascolarctobacterium, suggesting that these two bacterial communities may have a protective effect on CVD [30]. It has been demonstrated that Bacteroides can prevent atherosclerosis through a reduction in LPS production [74]. Phascolarctobacterium, a genus of SCFA-producing bacteria, inhibits the growth of Clostridium difficile and holds promise as a therapeutic option for patients afflicted with this pathogen [75]. However, in psoriatic patients, Phascolarctobacterium is considered a risk factor for CVD [76]. Further investigations are warranted to explore the impact of diverse microbial communities on the prognosis of HD patients.
The uremic toxins (TMAO, pCS, and IS) are associated with increased CVD risk. Several studies have demonstrated that elevated TMAO levels represent an independent and significant risk factor for cardiovascular events in HD patients [60,77,78]. TMAO is associated with the progression of atherosclerosis, and its mechanisms include affecting cholesterol metabolism, promoting thrombosis, activating inflammation, and damaging endothelial cells [79,80]. TMAO increases the expression of receptor cluster of differentiation (CD) 36 and scavenger receptor A on macrophages, thereby inhibiting reverse cholesterol transport (RCT) and promoting the formation of foam cells [81]. TMAO induces mitogen-activated protein kinase and NF-κB signaling, thereby promoting vascular inflammation [82,83]. TMAO induces inflammation and endothelial dysfunction by activating the ROS-TXNIP-NLRP3 inflammasome pathway [84]. Moreover, elevated TMAO levels can upregulate the expression of thrombin, ADP, and collagen and trigger intracellular calcium release and platelet hyperreactivity, thereby contributing to the modulation of platelet function and thrombosis in vivo [85]. Recent studies also suggest that TMAO is closely associated with increased arterial stiffness and vascular calcification in HD patients [86,87]. An increase in arterial stiffness may lead to increased blood pressure and increased heart burden. Vascular calcification may cause the vessel wall to become fragile, increasing the risk of CVD. Cardiovascular calcification (CVC) is a well-known cardiovascular risk factor in HD patients[88].
Unlike TMAO, the protein-bound molecules pCS and IS are challenging to remove through conventional dialysis. The pathophysiological mechanisms associated with cardiovascular injury in pCS and IS include the induction of endothelial dysfunction, inflammatory responses, and oxidative stress[89]. Clinical studies have revealed a close association between these metabolites and the overall mortality and cardiovascular event mortality of HD patients [90,91,92,93,94]. These studies provide strong evidence that abnormal changes in metabolites may have a significant impact on the prognosis of HD patients. However, not all researchers have observed this significant correlation[95,96,97]. Therefore, further research is needed to determine the relationship between these metabolites and the prognosis of HD patients and whether changes in these metabolites can serve as biomarkers or targets for HD treatment.
The Gut Microbiota and Infection
Infection is the second leading cause of death among ESRD patients, and sepsis accounts for more than 75% of all infection-related deaths, posing a serious threat to their lives and health [98]. Owing to impaired immune system function, these patients are more susceptible to infection by various pathogens, leading to infection and sepsis. In a 7-year follow-up of 4005 HD patients, 11.7% of HD patients had at least one episode of sepsis [99]. The mortality rate for sepsis in HD patients is significantly greater, ranging from 100–300 times greater than that of the general population [4]. The presence of an arteriovenous fistula/graft or a dialysis catheter, older age, malnutrition, diabetes, and the frequency of dialysis are predisposing factors for infections [100,101,102,103]. Immune dysfunction and dysbiosis of the gut microbiota also play critical roles in the occurrence and development of infections [104,105]. Elevated concentrations of free pCS in HD patients have been shown to increase the risk of infection-related hospitalizations (IH) [106,107].
Infections related to vascular access represent a common source of infectious complications [108]. Although only approximately 19% of HD patients utilize central venous catheters (CVCs), catheter-related bloodstream infections (CRBSIs) account for 70% of vascular access infections [109]. Staphylococcus aureus, enterococci and coagulase-negative staphylococci are the most common causative microorganisms implicated in CRBSIs [110]. Moreover, the increasing resistance of HD patients to antibiotics has resulted in a heightened prevalence of infections caused by multidrug-resistant microorganisms (MDROs) [111]. Vancomycin-resistant enterococci (VREs) and methicillin-resistant Staphylococcus aureus (MRSA) are common pathogens associated with MDROs [112,113]. An increase in the genes encoding antibiotic resistance in HD patients may result in bacterial insusceptibility to conventional antibiotic therapies, consequently leading to a substantial reduction in the efficacy of antibiotics [114]. Microbial virulence factors are a class of molecules produced by pathogenic microorganisms that have a toxic effect on the host. They play crucial roles in promoting the proliferation, spread, and worsening of infection within the host organism [115]. In HD patients, 12 virulence factors exhibited significant changes. The levels and activities of these factors differ between patients and healthy individuals, which may have a negative impact on the health of patients. Among them, pyridoxamine 5-phosphate oxidase and streptothricin-acetyl-transferase are relatively highly abundant in HD patients and may affect the course of the disease and the treatment effect to some extent [114]. Further research into these two enzymes may provide new ideas and methods for the treatment of HD patients.
The Gut Microbiota and Anemia
Anemia is a common complication in HD patients. According to data from the China Dialysis Outcomes and Practice Patterns Study (DOPPS), 21% of patients receiving HD treatment in China had hemoglobin levels less than 9 g/dl [116]. These data highlight the widespread prevalence of anemia among dialysis patients in China, which significantly impacts their quality of life and prognosis. Notably, the corresponding figures in Japan and North America are 10% and 3%, respectively [116]. This may be related to differences in the level of development of dialysis treatment, medical resource allocation, and patient education in various countries.
There are many causes of anemia in HD patients, including altered iron homeostasis, erythropoietin (EPO) deficiency, hyperparathyroidism, and chronic inflammation [117]. Several in vitro experiments have shown that IS can inhibit the generation of EPO through a hypoxia-inducible factor (HIF)-dependent oxygen-sensing mechanism [118,119,120]. HIF is a transcription factor that is activated in low-oxygen environments and can bind to the promoter of the EPO gene, thereby regulating EPO generation [121]. Additionally, IS can induce suicidal erythrocyte death or eryptosis, both of which are associated with a shortened lifespan of erythrocytes [122]. However, no significant correlation between IS or pCS and anemia has been reported in clinical studies of HD patients [117,123]. The significant difference between the in vivo and in vitro experiments may be due to the significant differences in the experimental environments. The concentration of uremic toxins in in vitro experiments may be relatively high. Therefore, it is essential to fully consider this possibility when designing experiments to ensure the accuracy and reliability of the experimental results [117].
Iron supplementation and EPO-stimulating agents (ESAs) are commonly used in HD patients. However, EPO hyporesponsiveness (EH) occurs in 10% of patients treated with ESAs [124,125], which may be related to iron metabolism disorders and EPO receptor dysfunction [126]. In a study on the responsiveness to EPO treatment, researchers reported that 9 bacteria in HD patients had predictive value for EH, with Neisseria (area under the curve (AUC) = 0.9535 (95% CI, 90.2–100, P < 0.0001) showing the highest predictive value [127]. The author also reported that the majority of enzymes related to butyrate synthesis were significantly enriched in HD patients with a good EH response, which may contribute to improving anemia [127]. A recent study has shown that supplementing dietary fiber (DF) can significantly improve anemia in HD patients. There are likely multiple mechanisms by which supplementing DF can improve renal anemia, one of which may be the increased production of butyrate and butyrate-producing bacteria (such as Bifidobacterium, Lactobacillus, and Lactobacillaceae) [126]. However, this discovery provides only a preliminary indication of this possibility. To ensure that this discovery can truly benefit HD patients, it is necessary to conduct multicenter, large-sample, and long-term clinical studies to comprehensively evaluate its clinical value.
The Gut Microbiota and Malnutrition
HD patients are at high risk of malnutrition due to impaired kidney function, which makes them prone to sarcopenia and protein-energy wasting (PEW). Under both of these conditions, the loss of muscle mass plays a crucial role in pathogenesis [128]. However, the concept of sarcopenia is no longer limited to a decrease in muscle mass but also includes the loss of muscle strength [129]. Protein loss during dialysis, reduced physical activity, chronic inflammation, etc., can all contribute to the development of sarcopenia [130]. The gut‒muscle axis suggests that the gut microbiota plays a crucial role in maintaining skeletal muscle homeostasis [131]. A recent study demonstrated a decrease in gut microbiota diversity and alterations in microbial structure in HD patients with sarcopenia[132]. However, there have been few studies on HD patients, and most of them were observational studies with small sample sizes, which cannot prove a causal relationship between the gut microbiota and sarcopenia. Tang et al. reported significant reductions in muscle function and mass in mice colonized with gut microbiota from HD patients with sarcopenia, accompanied by a decrease in the abundance of Akkermansia, a producer of SCFAs [133]. Notably, SCFAs have been proven by multiple studies to have a positive effect on skeletal muscle mass [134,135]. Additionally, a study revealed that IS may cause metabolic disorders, further leading to impaired mitochondrial function [136]. Mitochondria are energy factories in cells and are crucial for the growth and maintenance of muscles. The impairment of mitochondrial function may ultimately result in a reduction in muscle mass [136]. Additionally, IS can induce myotube atrophy by activating the ROS-ERK and JNK-MAFbx signaling pathways, thereby affecting muscle generation and maintenance [137]. However, many aspects of the pathogenesis of sarcopenia remain unknown, and future research will continue to explore these areas to provide more effective treatment options for HD patients.
Similarly, PEW is a disease related to malnutrition that primarily occurs in people who are unable to obtain sufficient food or nutrients, leading to severe deficiencies in body protein and energy [138]. The prevalence of PEW in HD patients is 30% to 75% [139]. Decreased nutrient intake, systemic inflammation, and inadequate dialysis are associated with the development of PEW [140,141]. The abundance of the butyric acid-producing bacteria Faecalibacterium prausnitzii and Roseburia was reportedly reduced in HD patients with PEW [140,142]. The gut microbiota may also be useful for predicting PEW in HD patients [143]. Another researcher reported a positive correlation between the levels of Actinobacteria and Bifidobacteriaceae and PEW indicators such as serum albumin levels, lean tissue mass (LTM), and the lean tissue index (LTI) [143]. The level of TMAO was significantly greater in HD patients with PEW [144]. These findings suggest that TMAO may play an important role in this disease process. Further research revealed that circulating TMAO levels are significantly associated with the incidence of PEW in HD patients [144], which means that an increase in TMAO levels may increase the risk of PEW. To better understand this phenomenon, it is necessary to conduct in-depth research on the causal relationship between TMAO and PEW.
Strategies to Attenuate Gut Microbiota Dysbiosis in HD
Research on probiotics, prebiotics, and synbiotics has received widespread attention from scholars at home and abroad in recent years. These three substances are believed to have significant effects on the balance of the gut microbiota and human health. In evaluating the effects of these supplements on diseases, most research currently uses indicators such as endotoxins, uremic toxins, inflammation, and metabolic markers. Probiotics, as active microorganisms, can not only reduce inflammatory responses [145] but also positively impact glucose homeostasis, oxidative stress [146,147], kidney function [147], nutritional status[148], and quality-of-life indicators[149,150]. Moreover, probiotics have been shown to significantly reduce the production of uremic toxin precursors (such as phenol and p-cresol) [151] but have no significant positive effect on IS or pCS in HD patients [152,153].
Prebiotics are nondigestible food ingredients [154]. In fourteen HD patients who were supplemented with curcumin for three months, curcumin significantly lowered plasma pCS levels [155]. Resistant starch (RS) is a prebiotic compound that promotes the proliferation of SCFA-producing groups (such as Roseburia and Ruminococcus gauvreauii) [156], increases the production of SCFAs, reduces the levels of pCS and IS [157], and alleviates inflammation and oxidative stress markers [158,159]. However, the efficacy of RS has been confirmed in many but not all studies. Esgalhado et al. reported that RS only reduced plasma IS levels, with no significant effect on pCS levels [158]. High-amylose resistant starch (HAM-RS2) can reduce serum creatinine and p-cresol levels but has no significant effect on IS levels [160]. Moreover, RS does not seem to play a role in altering plasma TMAO levels, as it does not contribute to any significant changes in the concentration of this metabolite in the bloodstream[161].
Synbiotics are mixtures of prebiotics and probiotics. A trial investigating synbiotic ingestion for four weeks in HD patients revealed that such therapy altered the fecal microbiota (Bifidobacterium enrichment) and significantly increased the levels of acetic acid and butyric acid [162]. In 58 HD subjects, a 7-week synbiotic treatment decreased the serum IS, pCS and urea concentrations [163]. Probiotic intervention has demonstrated potential benefits in reducing the levels of serum high-sensitivity C-reactive protein (hs-CRP), IL6, and endotoxins [164]. Furthermore, the combined administration of synbiotics and probiotics for 12 weeks has been shown to ameliorate anemia in HD patients [165].
Conclusions
In general, HD restored the abundance of beneficial microbes but induced some potentially pathogenic bacteria. The gut microbiota can play a crucial role in post-HD complications, such as cardiovascular events, infections, anemia, and nutritional complications, through metabolites and immune regulation. Studies aimed at restoring the appropriate gut microbiota composition and alleviating uremic toxins have been conducted. It seems that dietary interventions consisting of probiotics, prebiotics, and synbiotics are promising strategies. These dietary supplements have been shown to restore the gut microbiota composition, reduce IS and pCS levels, and reduce inflammatory marker levels. However, there is a lack of clear guidelines to inform HD patients when to take these dietary supplements, as well as the types and doses that must be taken, which can confuse clinicians when prescribing them for patients. All of these measures have limited effects. They do not fundamentally solve the problem of gut microbiota imbalance.
In addition, there are many limitations in the study of the gut microbiota and metabolites in HD patients. First, research on the gut microbiota and metabolites in HD patients is mostly based on single-center, small-sample studies at present. These studies largely ignore the influence of diet, dialysis duration, and drug intake on the gut microbiota and metabolites, leading to highly heterogeneous conclusions and difficulty in forming a unified scientific understanding. Second, research on the relationship between the gut microbiota and phenotype in HD patients is relatively limited. Most of the existing studies are observational studies and have not fully demonstrated the causal relationship between the gut microbiota and phenotypes. In addition, the molecular mechanisms by which metabolites regulate HD complications remain unclear, which limits our understanding and means of preventing and treating HD complications. Finally, reliable evidence from the gut microbiota perspective to guide clinical management strategies is challenging to obtain because of the complex relationships between hosts and microorganisms, individual differences, and susceptibility to multiple factors. This requires a deeper understanding of the diversity and dynamics of the gut microbiota, as well as further research on the relationship between the gut microbiota and HD patient phenotypes to reveal the underlying biological mechanisms. In the future, we look forward to gaining a deeper understanding of the gut microbiota and metabolites of HD patients through large-scale clinical trials, considering the effects of diet, dialysis duration, and drug intake, to provide a more comprehensive and scientific basis for clinical management strategies.
Author Contributions
Conceptualization: J.D., X.Z., and H.Z., methodology, validation, and formal analysis: Q.R., H.W., X.W., C.S, and Y.D., writing—original draft preparation: J.D., and X.Z.; writing—review and editing: X.D., Q.H., and D.Z.; supervision: H.Z.; funding acquisition: D.Z., and H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The National Natural Science Foundation of China (No. 62271506, No. 61971441), the National Key R&D Program of China (Nos. 2021YFC1005300), sJinzhongzi project of Beijing Chao-yang Hospital (No. CYJZ202203).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kalantar-Zadeh, K.; et al. Chronic kidney disease. Lancet 2021, 398, 786–802. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, T.; et al. Worldwide access to treatment for end-stage kidney disease: a systematic review. Lancet, 2015, 385, 1975–82. [Google Scholar] [CrossRef] [PubMed]
- Thiery, A.; et al. Survival advantage of planned haemodialysis over peritoneal dialysis: a cohort study. Nephrology Dialysis Transplantation, 2018, 33, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Himmelfarb, J. Hemodialysis complications. Am J Kidney Dis, 2005, 45, 1122–31. [Google Scholar] [CrossRef] [PubMed]
- Saran, R.; et al. US Renal Data System 2016 Annual Data Report: Epidemiology of Kidney Disease in the United States. Am J Kidney Dis, 2017, 69, A7–a8. [Google Scholar] [CrossRef] [PubMed]
- Structure, function and diversity of the healthy human microbiome. Nature, 2012, 486, 207–14. [CrossRef] [PubMed]
- Montemurno, E.; et al. What would you like to eat, Mr CKD Microbiota? A Mediterranean Diet, please! Kidney Blood Press Res, 2014, 39, 114–23. [Google Scholar] [CrossRef] [PubMed]
- Galla, S.; et al. Microbiotal-Host Interactions and Hypertension. Physiology (Bethesda), 2017, 32, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, A.; et al. Role of the Gut Microbiome in Uremia: A Potential Therapeutic Target. Am J Kidney Dis, 2016, 67, 483–98. [Google Scholar] [CrossRef]
- Stavropoulou, E.; et al. Focus on the Gut-Kidney Axis in Health and Disease. Front Med (Lausanne), 2020, 7, 620102. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; et al. Chronic kidney disease alters intestinal microbial flora. Kidney Int, 2013, 83, 308–15. [Google Scholar] [CrossRef] [PubMed]
- Evenepoel, P. , Poesen, R. ; Meijers, B., The gut–kidney axis. Pediatric Nephrology, 2016, 32, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Cosola, C.; et al. Microbiota metabolites: Pivotal players of cardiovascular damage in chronic kidney disease. Pharmacol Res, 2018, 130, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; et al. Gut microbiota as a key regulator of intestinal mucosal immunity. Life Sci, 2024, 345, 122612. [Google Scholar] [CrossRef] [PubMed]
- Mörbe, U.M.; et al. Human gut-associated lymphoid tissues (GALT); diversity, structure, and function. Mucosal Immunology, 2021, 14, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Bourlioux, P.; et al. he intestine and its microflora are partners for the protection of the host: report on the Danone Symposium “The Intelligent Intestine,” held in Paris, June 14, 2002. Am J Clin Nutr 2003, 78, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; et al. Enterotypes of the human gut microbiome. Nature, 2011, 473, 174–80. [Google Scholar] [CrossRef] [PubMed]
- Tourountzis, T.; et al. Microbiome in Chronic Kidney Disease. Life (Basel), 2022, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.L.; Vaziri, N.D. The Leaky Gut and Altered Microbiome in Chronic Kidney Disease. J Ren Nutr, 2017, 27, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; et al. Expansion of urease- and uricase-containing, indole- and p-cresol-forming and contraction of short-chain fatty acid-producing intestinal microbiota in ESRD. Am J Nephrol, 2014, 39, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Khoury, T.; et al. The gut-kidney axis in chronic renal failure: A new potential target for therapy. Hemodial Int, 2017, 21, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Zhao, Y-Y.; Pahl, M.V. Altered intestinal microbial flora and impaired epithelial barrier structure and function in CKD: the nature, mechanisms, consequences and potential treatment. Nephrology Dialysis Transplantation, 2016, 31, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Simões-Silva, L.; et al. The microbiome in chronic kidney disease patients undergoing hemodialysis and peritoneal dialysis. Pharmacological Research, 2018, 130, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Berg, R.D. , Wommack, E. ; Deitch, E.A., Immunosuppression and intestinal bacterial overgrowth synergistically promote bacterial translocation. Arch Surg, 1988, 123, 1359–64. [Google Scholar] [PubMed]
- Shi, K.; et al. Gut bacterial translocation may aggravate microinflammation in hemodialysis patients. Dig Dis Sci, 2014, 59, 2109–17. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, C.W.; et al. Circulating endotoxemia: a novel factor in systemic inflammation and cardiovascular disease in chronic kidney disease. Clin J Am Soc Nephrol, 2011, 6, 133–41. [Google Scholar] [CrossRef] [PubMed]
- Blikslager, A.T.; et al. Restoration of barrier function in injured intestinal mucosa. Physiol Rev, 2007, 87, 545–64. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; et al. Microbiota analysis in the hemodialysis population—Focusing on Enterobacteriaceae. J Microbiol Immunol Infect, 2023, 56, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Salgado, J.; et al. Intestinal microbiota in pediatric patients with end stage renal disease: a Midwest Pediatric Nephrology Consortium study. Microbiome, 2016, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; et al. The Effects of Hemodialysis and Peritoneal Dialysis on the Gut Microbiota of End-Stage Renal Disease Patients, and the Relationship Between Gut Microbiota and Patient Prognoses. Front Cell Infect Microbiol, 2021, 11, 579386. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Yu, Z. , Genomic Insights into the Distribution of Peptidases and Proteolytic Capacity among Prevotella and Paraprevotella Species. Microbiol Spectr, 2022, 10, e0218521. [Google Scholar] [CrossRef] [PubMed]
- Morotomi, M.; et al. Paraprevotella clara gen. nov., sp. nov. and Paraprevotella xylaniphila sp. nov., members of the family ‘Prevotellaceae’ isolated from human faeces. Int J Syst Evol Microbiol 2009, 59, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A. The determination of urea, ammonia, and urease. Methods Biochem Anal, 1969, 17, 311–24. [Google Scholar] [PubMed]
- Zhang, T.; et al. Akkermansia muciniphila is a promising probiotic. Microb Biotechnol, 2019, 12, 1109–1125. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; et al. Alterations to the Gut Microbiota and Their Correlation With Inflammatory Factors in Chronic Kidney Disease. Front Cell Infect Microbiol, 2019, 9, 206. [Google Scholar] [CrossRef] [PubMed]
- Visca, P. , Seifert, H. ; Towner, K.J., Acinetobacter infection--an emerging threat to human health. IUBMB Life, 2011, 63, 1048–54. [Google Scholar] [PubMed]
- Hamilton, A.L.; et al. Proteus spp. as Putative Gastrointestinal Pathogens. Clin Microbiol Rev, 2018, 31, 3. [Google Scholar]
- Chen, L.; et al. Unilateral ureteral obstruction causes gut microbial dysbiosis and metabolome disorders contributing to tubulointerstitial fibrosis. Exp Mol Med, 2019, 51, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; et al. Gut bacterial translocation is associated with microinflammation in end-stage renal disease patients. Nephrology (Carlton), 2012, 17, 733–8. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Xie, Y. , Effect of Different Hemodialysis Methods on Microbiota in Uremic Patients. Biomed Res Int, 2020, 2020, 6739762. [Google Scholar] [CrossRef]
- Ghassabi, A.; et al. Potential contribution of the immune system to the emergence of renal diseases. Immunology Letters, 2022, 248, 1–6. [Google Scholar] [CrossRef] [PubMed]
- March, D.S.; et al. Intestinal Barrier Disturbances in Haemodialysis Patients: Mechanisms, Consequences, and Therapeutic Options. Biomed Res Int, 2017, 2017, 5765417. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Zhao, Y.Y.; Pahl, M.V. , Altered intestinal microbial flora and impaired epithelial barrier structure and function in CKD: the nature, mechanisms, consequences and potential treatment. Nephrol Dial Transplant, 2016, 31, 737–46. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; et al. Immunosenescence, gut dysbiosis, and chronic kidney disease: Interplay and implications for clinical management. Biomed J, 2023, 47, 100638. [Google Scholar] [CrossRef] [PubMed]
- Dickson, K.; Lehmann, C. , Inflammatory Response to Different Toxins in Experimental Sepsis Models. Int J Mol Sci, 2019, 20, 18. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.J.; Andersen, K.; Stecher, B. , The intestinal microbiota, a leaky gut, and abnormal immunity in kidney disease. Kidney Int, 2013, 83, 1010–6. [Google Scholar] [CrossRef]
- Wong, J.; et al. Expansion of Urease- and Uricase-Containing, Indole- and p-Cresol-Forming and Contraction of Short-Chain Fatty Acid-Producing Intestinal Microbiota in ESRD. American Journal of Nephrology, 2014, 39, 230–237. [Google Scholar] [CrossRef]
- Bao, W.H.; et al. Relationship between gut microbiota and vascular calcification in hemodialysis patients. Ren Fail, 2023, 45, 2148538. [Google Scholar] [CrossRef] [PubMed]
- Roediger, W.E. , Utilization of nutrients by isolated epithelial cells of the rat colon. Gastroenterology, 1982, 83, 424–9. [Google Scholar] [CrossRef] [PubMed]
- Segain, J.P.; et al. Butyrate inhibits inflammatory responses through NFkappaB inhibition: implications for Crohn’s disease. Gut, 2000, 47, 397–403. [Google Scholar] [CrossRef] [PubMed]
- He, J.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int J Mol Sci, 2020, 21, 17. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; et al. Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am J Physiol Gastrointest Liver Physiol, 2013, 305, G900–G910. [Google Scholar] [CrossRef] [PubMed]
- Knowles, S.E.; et al. Production and utilization of acetate in mammals. Biochem J, 1974, 142, 401–11. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; et al. The role of short-chain fatty acids in health and disease. Adv Immunol, 2014, 121, 91–119. [Google Scholar] [PubMed]
- Brown, A.J.; et al. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J Biol Chem, 2003, 278, 11312–9. [Google Scholar] [CrossRef] [PubMed]
- Sirich, T.L.; et al. Prominent accumulation in hemodialysis patients of solutes normally cleared by tubular secretion. J Am Soc Nephrol, 2014, 25, 615–22. [Google Scholar] [CrossRef] [PubMed]
- Evenepoel, P.; Poesen, R.; Meijers, B. The gut-kidney axis. Pediatr Nephrol, 2017, 32, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.C.; Sirich, T.L. , Indoxyl Sulfate-Review of Toxicity and Therapeutic Strategies. Toxins (Basel), 2016, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.B.; et al. Advanced chronic kidney disease populations have elevated trimethylamine N-oxide levels associated with increased cardiovascular events. Kidney Int, 2016, 89, 1144–1152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; et al. Association of trimethylamine N-Oxide with cardiovascular and all-cause mortality in hemodialysis patients. Ren Fail, 2020, 42, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Agus, A.; Clément, K.; Sokol, H. , Gut microbiota-derived metabolites as central regulators in metabolic disorders. Gut, 2021, 70, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Hai, X.; et al. Mechanism of Prominent Trimethylamine Oxide (TMAO) Accumulation in Hemodialysis Patients. PLoS One, 2015, 10, e0143731. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, J.R.; et al. Serum Trimethylamine-N-Oxide is Elevated in CKD and Correlates with Coronary Atherosclerosis Burden. J Am Soc Nephrol, 2016, 27, 305–13. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; et al. Association of Circulating Trimethylamine-N Oxide With Malnutrition and the Risk of Coronary Artery Disease in Patients With Maintenance Hemodialysis. J Ren Nutr, 2023, 33, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Bammens, B.; et al. Free serum concentrations of the protein-bound retention solute p-cresol predict mortality in hemodialysis patients. Kidney Int, 2006, 69, 1081–7. [Google Scholar] [CrossRef]
- Ahmadmehrabi, S.; Tang, W.H.W. Hemodialysis-induced cardiovascular disease. Semin Dial, 2018, 31, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Echefu, G.; et al. Pathophysiological concepts and screening of cardiovascular disease in dialysis patients. Front Nephrol, 2023, 3, 1198560. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, M.; et al. Cardiovascular disease in dialysis patients. Nephrology Dialysis Transplantation 2018, 33, iii28–iii34. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.; et al. Gut Bacterial Translocation May Aggravate Microinflammation in Hemodialysis Patients. Digestive Diseases and Sciences, 2014, 59, 2109–2117. [Google Scholar] [CrossRef] [PubMed]
- Donath, M.Y.; Meier, D.T.; Böni-Schnetzler, M. Inflammation in the Pathophysiology and Therapy of Cardiometabolic Disease. Endocr Rev, 2019, 40, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Sumida, K.; et al. Circulating Microbial Signatures and Cardiovascular Death in Patients With ESRD. Kidney Int Rep, 2021, 6, 2617–2628. [Google Scholar] [CrossRef] [PubMed]
- Dinakaran, V.; et al. Elevated levels of circulating DNA in cardiovascular disease patients: metagenomic profiling of microbiome in the circulation. PLoS One, 2014, 9, e105221. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Gupta, D. , Crosstalk of toll-like receptors signaling and Nrf2 pathway for regulation of inflammation. Biomed Pharmacother, 2018, 108, 1866–1878. [Google Scholar] [CrossRef]
- Yoshida, N.; et al. Bacteroides vulgatus and Bacteroides dorei Reduce Gut Microbial Lipopolysaccharide Production and Inhibit Atherosclerosis. Circulation, 2018, 138, 2486–2498. [Google Scholar] [CrossRef]
- Nagao-Kitamoto, H.; et al. Interleukin-22-mediated host glycosylation prevents Clostridioides difficile infection by modulating the metabolic activity of the gut microbiota. Nat Med, 2020, 26, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Valentini, V.; et al. A Possible Link between Gut Microbiome Composition and Cardiovascular Comorbidities in Psoriatic Patients. J Pers Med, 2022, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; et al. Trimethylamine-N-oxide is an independent risk factor for hospitalization events in patients receiving maintenance hemodialysis. Ren Fail, 2020, 42, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Shafi, T.; et al. Trimethylamine N-Oxide and Cardiovascular Events in Hemodialysis Patients. J Am Soc Nephrol, 2017, 28, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Gui, T.; et al. Diverse roles of macrophages in atherosclerosis: from inflammatory biology to biomarker discovery. Mediators Inflamm, 2012, 2012, 693083. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; et al. Implication of Gut Microbiota in Cardiovascular Diseases. Oxid Med Cell Longev, 2020, 2020, 5394096. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med, 2013, 19, 576–85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gérard, P. Diet-gut microbiota interactions on cardiovascular disease. Comput Struct Biotechnol J, 2022, 20, 1528–1540. [Google Scholar] [CrossRef] [PubMed]
- Seldin, M.M.; et al. Trimethylamine N-Oxide Promotes Vascular Inflammation Through Signaling of Mitogen-Activated Protein Kinase and Nuclear Factor-κB. J Am Heart Assoc, 2016, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; et al. Trimethylamine N-oxide induces inflammation and endothelial dysfunction in human umbilical vein endothelial cells via activating ROS-TXNIP-NLRP3 inflammasome. Biochem Biophys Res Commun, 2016, 481, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Zhen, J.; et al. The gut microbial metabolite trimethylamine N-oxide and cardiovascular diseases. Front Endocrinol (Lausanne), 2023, 14, 1085041. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.Y.; et al. Serum Trimethylamine N-Oxide Level Is Positively Associated with Aortic Stiffness Measured by Carotid-Femoral Pulse Wave Velocity in Patients Undergoing Maintenance Hemodialysis. Toxins (Basel), 2023, 15, 9. [Google Scholar] [CrossRef] [PubMed]
- He, L.; et al. Higher serum trimethylamine-N-oxide levels are associated with increased abdominal aortic calcification in hemodialysis patients. Ren Fail, 2022, 44, 2019–2027. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, M.; et al. Cardiovascular disease in dialysis patients. Nephrol Dial Transplant 2018, 33, iii28–iii34. [Google Scholar] [CrossRef] [PubMed]
- Vanholder, R.; et al. The uremic toxicity of indoxyl sulfate and p-cresyl sulfate: a systematic review. J Am Soc Nephrol, 2014, 25, 1897–907. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; et al. Serum total indoxyl sulfate and clinical outcomes in hemodialysis patients: results from the Japan Dialysis Outcomes and Practice Patterns Study. Clin Kidney J, 2021, 14, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; et al. Serum total indoxyl sulfate levels and all-cause and cardiovascular mortality in maintenance hemodialysis patients: a prospective cohort study. BMC Nephrol, 2022, 23, 231. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.J.; et al. Serum p-cresyl sulfate predicts cardiovascular disease and mortality in elderly hemodialysis patients. Arch Med Sci, 2013, 9, 662–8. [Google Scholar] [CrossRef] [PubMed]
- Wu, I.-W.; et al. Serum free p -cresyl sulfate levels predict cardiovascular and all-cause mortality in elderly hemodialysis patients—a prospective cohort study. Nephrology Dialysis Transplantation, 2011, 27, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.S.; et al. Association of indoxyl sulfate with heart failure among patients on hemodialysis. Clin J Am Soc Nephrol, 2015, 10, 111–9. [Google Scholar] [CrossRef] [PubMed]
- Shafi, T.; et al. Results of the HEMO Study suggest that p-cresol sulfate and indoxyl sulfate are not associated with cardiovascular outcomes. Kidney Int, 2017, 92, 1484–1492. [Google Scholar] [CrossRef]
- Lin, T.Y.; et al. Indoxyl Sulfate and Incident Peripheral Artery Disease in Hemodialysis Patients. Toxins (Basel) 2020, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Melamed, M.L.; et al. Retained organic solutes, patient characteristics and all-cause and cardiovascular mortality in hemodialysis: results from the retained organic solutes and clinical outcomes (ROSCO) investigators. BMC Nephrol, 2013, 14, 134. [Google Scholar] [CrossRef] [PubMed]
- Sarnak, M.J.; Jaber, B.L. , Mortality caused by sepsis in patients with end-stage renal disease compared with the general population. Kidney Int, 2000, 58, 1758–64. [Google Scholar] [CrossRef] [PubMed]
- Powe, N.R.; et al. Septicemia in dialysis patients: incidence, risk factors, and prognosis. Kidney Int, 1999, 55, 1081–90. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; et al. Risk factors for catheter-associated bloodstream infection in hemodialysis patients: A meta-analysis. PLoS One, 2024, 19, e0299715. [Google Scholar] [CrossRef] [PubMed]
- Descamps-Latscha, B. The immune system in end-stage renal disease. Curr Opin Nephrol Hypertens, 1993, 2, 883–91. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.; Hörl, W.H. Immune dysfunction in uremia—an update. Toxins (Basel), 2012, 4, 962–90. [Google Scholar] [PubMed]
- Shorr, A.F.; et al. Prediction of infection due to antibiotic-resistant bacteria by select risk factors for health care-associated pneumonia. Arch Intern Med, 2008, 168, 2205–10. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; et al. Aspects of immune dysfunction in end-stage renal disease. Clin J Am Soc Nephrol, 2008, 3, 1526–33. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, M.T.; et al. Protein-Bound Uremic Toxins and Immunity. Methods Mol Biol, 2021, 2325, 215–227. [Google Scholar] [PubMed]
- Banerjee, T.; et al. Free and total p-cresol sulfate levels and infectious hospitalizations in hemodialysis patients in CHOICE and HEMO. Medicine (Baltimore), 2017, 96, e5799. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.J.; et al. Serum protein-bound uraemic toxins and clinical outcomes in haemodialysis patients. Nephrol Dial Transplant, 2010, 25, 3693–700. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Yassin, H.M. Infection and Hemodialysis Access: An Updated Review. Infectious Disorders—Drug Targets, 2013, 13, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.B.; et al. National Healthcare Safety Network (NHSN) Dialysis Event Surveillance Report for 2014, Clin J Am Soc Nephrol, 2017, 12, 1139–1146.
- Gupta, V.; Yassin, M.H. Infection and hemodialysis access: an updated review. Infect Disord Drug Targets, 2013, 13, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; et al. Association of Kidney Function with Infections by Multidrug-Resistant Organisms: An Electronic Medical Record Analysis. Sci Rep, 2018, 8, 13372. [Google Scholar] [CrossRef] [PubMed]
- Zacharioudakis, I.M.; et al. Vancomycin-resistant enterococci colonization among dialysis patients: a meta-analysis of prevalence, risk factors, and significance. Am J Kidney Dis, 2015, 65, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.B.; et al. CDC National Health Report: leading causes of morbidity and mortality and associated behavioral risk and protective factors--United States, 2005-2013. MMWR Suppl, 2014, 63, 3–27. [Google Scholar] [PubMed]
- Shi, X.; et al. Alterations of gut microbial pathways and virulence factors in hemodialysis patients. Front Cell Infect Microbiol, 2022, 12, 904284. [Google Scholar] [CrossRef] [PubMed]
- Leitão, J.H. Microbial Virulence Factors. Int J Mol Sci, 2020, 21, 15. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; et al. Anemia Management in the China Dialysis Outcomes and Practice Patterns Study. Blood Purif, 2016, 42, 33–43. [Google Scholar] [CrossRef]
- Bataille, S.; et al. Indole 3-acetic acid, indoxyl sulfate and paracresyl-sulfate do not influence anemia parameters in hemodialysis patients. BMC Nephrol, 2017, 18, 251. [Google Scholar] [CrossRef] [PubMed]
- Madai, S.; et al. Activation of the hypoxia-inducible factor pathway protects against acute ischemic stroke by reprogramming central carbon metabolism. Theranostics, 2024, 14, 2856–2880. [Google Scholar] [CrossRef]
- Chiang, C.K.; et al. Indoxyl sulfate, a representative uremic toxin, suppresses erythropoietin production in a HIF-dependent manner. Lab Invest, 2011, 91, 1564–71. [Google Scholar] [CrossRef]
- Tanaka, T.; et al. Indoxyl sulfate signals for rapid mRNA stabilization of Cbp/p300-interacting transactivator with Glu/Asp-rich carboxy-terminal domain 2 (CITED2) and suppresses the expression of hypoxia-inducible genes in experimental CKD and uremia. Faseb j, 2013, 27, 4059–75. [Google Scholar] [CrossRef]
- Nakai, T.; et al. The drug-specific properties of hypoxia-inducible factor-prolyl hydroxylase inhibitors in mice reveal a significant contribution of the kidney compared to the liver to erythropoietin induction. Life Sci, 2024, 346, 122641. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.S.; et al. Triggering of suicidal erythrocyte death by uremic toxin indoxyl sulfate. BMC Nephrol, 2013, 14, 244. [Google Scholar] [CrossRef] [PubMed]
- Capo-Chichi, J.C.C.; et al. Is there an association between the plasma levels of uremic toxins from gut microbiota and anemia in patients on hemodialysis? Int Urol Nephrol, 2022, 54, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
- Marcelli, D.; et al. Dynamics of the erythropoiesis stimulating agent resistance index in incident hemodiafiltration and high-flux hemodialysis patients. Kidney Int, 2016, 90, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Macdougall, I.C.; Cooper, A.C. Erythropoietin resistance: the role of inflammation and pro-inflammatory cytokines. Nephrol Dial Transplant, 2002, 17, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; et al. The prebiotic effects of soluble dietary fiber mixture on renal anemia and the gut microbiota in end-stage renal disease patients on maintenance hemodialysis: a prospective, randomized, placebo-controlled study. J Transl Med, 2022, 20, 599. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; et al. Gut Microbiota Correlates With Clinical Responsiveness to Erythropoietin in Hemodialysis Patients With Anemia. Front Cell Infect Microbiol, 2022, 12, 919352. [Google Scholar] [CrossRef] [PubMed]
- Mori, K. , Maintenance of Skeletal Muscle to Counteract Sarcopenia in Patients with Advanced Chronic Kidney Disease and Especially Those Undergoing Hemodialysis. Nutrients, 2021, 13, 5. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing, 2010, 39, 412–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-L.; et al. Impact of sarcopenia and its diagnostic criteria on hospitalization and mortality in chronic hemodialysis patients: A 3-year longitudinal study. Journal of the Formosan Medical Association, 2020, 119, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.; et al. The gut microbiota influences skeletal muscle mass and function in mice. Sci Transl Med, 2019, 11, 502. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; et al. Characterization of the gut microbiota in hemodialysis patients with sarcopenia. Int Urol Nephrol, 2022, 54, 1899–1906. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; et al. The gut microbiota from maintenance hemodialysis patients with sarcopenia influences muscle function in mice. Front Cell Infect Microbiol, 2023, 13, 1225991. [Google Scholar] [CrossRef] [PubMed]
- Han, D.S.; et al. Differences in the gut microbiome and reduced fecal butyrate in elders with low skeletal muscle mass. Clin Nutr, 2022, 41, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.Q.; et al. Human gut microbiome impacts skeletal muscle mass via gut microbial synthesis of the short-chain fatty acid butyrate among healthy menopausal women. J Cachexia Sarcopenia Muscle, 2021, 12, 1860–1870. [Google Scholar] [CrossRef] [PubMed]
- Sato, E.; et al. Metabolic alterations by indoxyl sulfate in skeletal muscle induce uremic sarcopenia in chronic kidney disease. Sci Rep, 2016, 6, 36618. [Google Scholar] [CrossRef] [PubMed]
- Changchien, C.Y.; et al. Indoxyl sulfate induces myotube atrophy by ROS-ERK and JNK-MAFbx cascades. Chem Biol Interact, 2019, 304, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Fouque, D.; et al. A proposed nomenclature and diagnostic criteria for protein-energy wasting in acute and chronic kidney disease. Kidney Int, 2008, 73, 391–8. [Google Scholar] [CrossRef] [PubMed]
- Herselman, M.; et al. Protein-energy malnutrition as a risk factor for increased morbidity in long-term hemodialysis patients. J Ren Nutr, 2000, 10, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Hung, S.C. , Association of subjective global assessment of nutritional status with gut microbiota in hemodialysis patients: a case-control study. Nephrol Dial Transplant, 2021, 36, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Mitch, W.E. Mechanisms of muscle wasting in chronic kidney disease. Nat Rev Nephrol, 2014, 10, 504–16. [Google Scholar] [CrossRef]
- Hu, J.; et al. Correlation between intestinal flora disruption and protein-energy wasting in patients with end-stage renal disease. BMC Nephrol, 2022, 23, 130. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; et al. Gut microbiota dysbiosis and protein energy wasting in patients on hemodialysis: an observational longitudinal study. Front Nutr, 2023, 10, 1270690. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; et al. Correlation between serum trimethylamine-N-oxide concentration and protein energy wasting in patients on maintenance hemodialysis. Ren Fail, 2022, 44, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.; et al. The effect of probiotic supplementation on systemic inflammation in dialysis patients. Kidney Res Clin Pract, 2022, 41, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, A.; et al. Probiotic supplementation in diabetic hemodialysis patients has beneficial metabolic effects. Kidney Int, 2017, 91, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; et al. Effects of Probiotic Supplementation on Nutrient Intake, Ghrelin, and Adiponectin Concentrations in Diabetic Hemodialysis Patients. Altern Ther Health Med, 2023, 29, 36–42. [Google Scholar] [PubMed]
- Yamamoto, M.; et al. Clostridium butyricum affects nutrition and immunology by modulating gut microbiota. Biosci Microbiota Food Health, 2022, 41, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Belova, I.V.; et al. Colon Microbiocenosis and Its Correction in Patients Receiving Programmed Hemodialysis. Sovrem Tekhnologii Med, 2021, 12, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Shamanadze, A.; et al. IMPACT OF MICROBIOME COMPOSITION ON QUALITY OF LIFE IN HEMODIALYSIS PATIENTS. Georgian Med News 2022, 101–106. [Google Scholar]
- Eidi, F.; et al. Effect of Lactobacillus Rhamnosus on serum uremic toxins (phenol and P-Cresol) in hemodialysis patients: A double blind randomized clinical trial. Clin Nutr ESPEN, 2018, 28, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Borges, N.A.; et al. Probiotic Supplementation in Chronic Kidney Disease: A Double-blind, Randomized, Placebo-controlled Trial. J Ren Nutr, 2018, 28, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Hyun, H.S.; Paik, K.H.; Cho, H.Y. p-Cresyl sulfate and indoxyl sulfate in pediatric patients on chronic dialysis. Korean J Pediatr, 2013, 56, 159–64. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J. , Fiber and prebiotics: mechanisms and health benefits. Nutrients, 2013, 5, 1417–35. [Google Scholar] [CrossRef] [PubMed]
- Salarolli, R.T.; et al. Can curcumin supplementation reduce plasma levels of gut-derived uremic toxins in hemodialysis patients? A pilot randomized, double-blind, controlled study. Int Urol Nephrol, 2021, 53, 1231–1238. [Google Scholar] [PubMed]
- Kemp, J.A.; et al. The Impact of Enriched Resistant Starch Type-2 Cookies on the Gut Microbiome in Hemodialysis Patients: A Randomized Controlled Trial. Mol Nutr Food Res, 2021, 65, e2100374. [Google Scholar] [CrossRef] [PubMed]
- Sirich, T.L.; et al. Effect of increasing dietary fiber on plasma levels of colon-derived solutes in hemodialysis patients. Clin J Am Soc Nephrol, 2014, 9, 1603–10. [Google Scholar] [CrossRef] [PubMed]
- Esgalhado, M.; et al. Could resistant starch supplementation improve inflammatory and oxidative stress biomarkers and uremic toxins levels in hemodialysis patients? A pilot randomized controlled trial. Food Funct, 2018, 9, 6508–6516. [Google Scholar]
- Tayebi Khosroshahi, H.; et al. Effect of high amylose resistant starch (HAM-RS2) supplementation on biomarkers of inflammation and oxidative stress in hemodialysis patients: a randomized clinical trial. Hemodial Int, 2018, 22, 492–500. [Google Scholar] [CrossRef]
- Khosroshahi, H.T.; et al. Effects of fermentable high fiber diet supplementation on gut derived and conventional nitrogenous product in patients on maintenance hemodialysis: a randomized controlled trial. Nutr Metab (Lond), 2019, 16, 18. [Google Scholar] [CrossRef] [PubMed]
- Kemp, J.A.; et al. Resistant Starch Type-2 Supplementation Does Not Decrease Trimethylamine N-Oxide (TMAO) Plasma Level in Hemodialysis Patients. J Am Nutr Assoc, 2022, 41, 788–795. [Google Scholar] [CrossRef]
- Miyoshi, M.; et al. Synbiotics Improved Stool Form via Changes in the Microbiota and Short-Chain Fatty Acids in Hemodialysis Patients. Kobe J Med Sci, 2021, 67, E112–e118. [Google Scholar] [PubMed]
- Lopes, R.; et al. Synbiotic meal decreases uremic toxins in hemodialysis individuals: A placebo-controlled trial. Food Res Int, 2019, 116, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Haghighat, N.; et al. Effects of Synbiotics and Probiotics Supplementation on Serum Levels of Endotoxin, Heat Shock Protein 70 Antibodies and Inflammatory Markers in Hemodialysis Patients: a Randomized Double-Blinded Controlled Trial. Probiotics Antimicrob Proteins, 2020, 12, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Haghighat, N.; et al. The Effect of Synbiotic and Probiotic Supplementation on Mental Health Parameters in Patients Undergoing Hemodialysis: A Double-blind, Randomized, Placebo-controlled Trial. Indian J Nephrol, 2021, 31, 149–156. [Google Scholar] [PubMed]
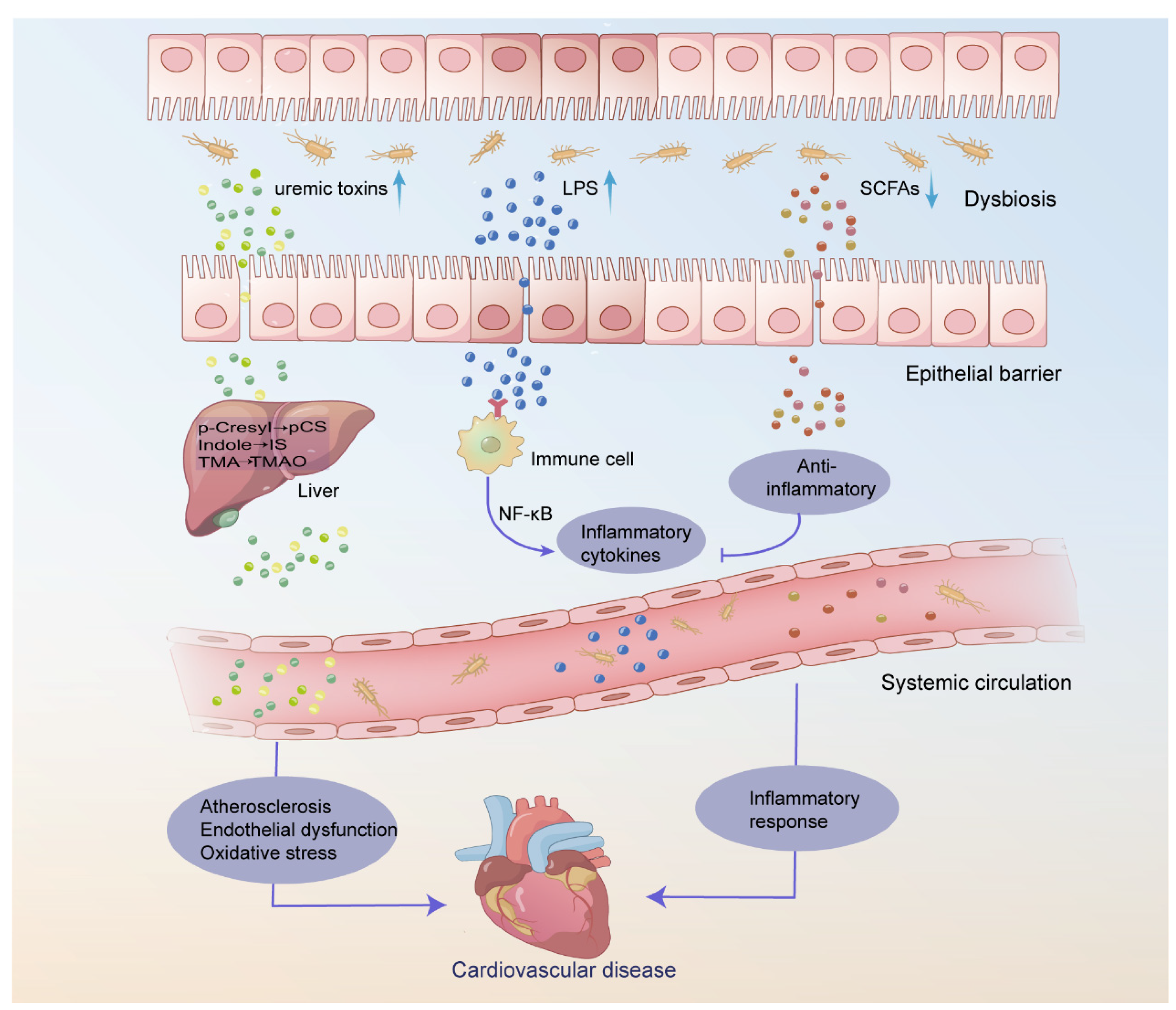
Figure 1.
Roles of gut microbiota dysbiosis in the development of cardiovascular disease (CVD) in hemodialysis (HD) patients. Disruption of the gut microbiota in HD patients leads to increased levels of uremic toxins produced in the liver, such as p-cresol sulfate (pCS), indole sulfate (IS), and trimethylamine N-oxide (TMAO), while beneficial SCFA metabolites are reduced. Additionally, gut microbiota dysbiosis causes damage to the integrity of the intestinal barrier, increasing bacterial translocation. This allows lipopolysaccharide (LPS) and uremic toxins to accumulate in the systemic circulation, leading to an increased release of inflammatory factors and triggering a series of pathophysiological changes. These changes include the development of atherosclerosis, endothelial dysfunction, oxidative stress, and chronic inflammation. Ultimately, this increases the risk of CVD.
Figure 1.
Roles of gut microbiota dysbiosis in the development of cardiovascular disease (CVD) in hemodialysis (HD) patients. Disruption of the gut microbiota in HD patients leads to increased levels of uremic toxins produced in the liver, such as p-cresol sulfate (pCS), indole sulfate (IS), and trimethylamine N-oxide (TMAO), while beneficial SCFA metabolites are reduced. Additionally, gut microbiota dysbiosis causes damage to the integrity of the intestinal barrier, increasing bacterial translocation. This allows lipopolysaccharide (LPS) and uremic toxins to accumulate in the systemic circulation, leading to an increased release of inflammatory factors and triggering a series of pathophysiological changes. These changes include the development of atherosclerosis, endothelial dysfunction, oxidative stress, and chronic inflammation. Ultimately, this increases the risk of CVD.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



