
Submitted:
02 August 2024
Posted:
06 August 2024
You are already at the latest version
Abstract
The cyclamen mite, Phytonemus pallidus, is one of the important pests in strawberry cultivation in Canada. This insect pest hides on young strawberry leaves that have not yet opened, which limits their control. In addition, the fight against the latter involves the use of synthetic chemicals that are not without effect on the environment, human health and beneficial organisms. We hypothesized that vectorization by bees could reduce the number of applications, product runoff and provide targeted control of these pathogens. To do this, field trials were conducted to evaluate the effectiveness of bumblebees Bombus terrestris in propagating Beauveria bassiana to control this pest. Our results showed that bumblebees were able to disseminate the inoculum on strawberry plants. Also, all the products used (B. bassiana and neem oil) were effective on P. pallidus. It should be noted that high mortality has been noted on this pest thanks to the application of this entomopathogen by spraying. In addition, neither of the two products tested had a significant effect on lady beetles C. septempunctata. These results therefore suggest the use of bee vectoring and neem essential oils in the sustainable management of strawberry mites.
Keywords:
bee vectoring
; beneficial organisms
; pests
; strawberries
; Bombus terrestris
; Phytonemus pallidus
; C. septempunctata
; environment
1. Introduction
The strawberry, Fragaria x pineapple Duch., is one of the most significant crops in Canada and around the world. In 2020, 24,158 metric tonnes of strawberries were produced in all provinces and territories in Canada [1]. The province of Ontario, on the other hand, is one of the largest producers of strawberries in North America. It was in second place in terms of area dedicated to strawberry production, with 2,737 acres, just behind Quebec, which had 5,281 acres in 2021 [2]. Unfortunately, this production is highly threatened by crop diseases and pests, which justifies the reduction in production area by almost half compared to the 2000s. One of the pests responsible for this decrease is the cyclamen mite (P. pallidus Banks; Acari: Tarsonemidae) [3]. It is a pest that abounds in newly developed strawberry leaves that are still folded [4]. According to [5], the juvenile stage is very dependent on leaf moisture. Due to its small body size (≤ 0.25 mm) and my way of survival, it is very difficult to control it with a chemical [6]. In addition, the use of these products is increasingly criticized due to their impact on human health and the environment [7]. Endosulfan (Thiodan, Thionex), a miticide originally used in Canada to control this pest was banned and removed from the marketplace as of December 31, 2016 [8]. Given the stringent regulations and consumer demands, iIt is becoming crucial to explore alternative approaches to the sustainable management of this pathogen. One solution would be to use the entomovectorization system or apivectorization. It is a system that integrates pollination and crop protection by mobilizing pollinators, such as bees, bumblebees, to transport biological control agents against plant pathogens [9]. It requires the use of a biological control agent such as a entomopathogenic and a fungal/bacterial inhibitor [10]. Thanks to a good formulation of this entomopathogen, bees are able to ensure better transport of this product for the reduction of insect pest infestations. This innovative approach leads to increased yields by promoting improved seed production while providing protection against plant diseases. Several trials of this technology have been carried out in North America, Central and South America, Europe, Australia, New Zealand and Kenya [11]. [12] have demonstrated that Bombus impatiens is capable of transporting the fungus Clonostachys rosea for the control of Botrytis cinerea in tomato and pepper plants in Canada. [13] evaluated the use of entomovectorization with Prestop-Mix and the Bombus terrestris vector to control Botrytis cinerea on strawberry plants in greenhouses. Their studies have shown that significant strawberry yields can be achieved, 2 to 2.5 times higher than normal without the use of these biocontrol agents. Other biological control agents combining the use of pollinators have been evaluated against various other pests and diseases. However, few studies have been conducted on this innovative approach in Northern Ontario. TheThe objective of this study was to determine the extent to which the conidia of the entomopathogen B. bassiana could be spread by bumblebees to control the cyclamen mite population (P. pallidus). Different factors were considered: (1) the susceptibility of the mite to a formulation B. bassiana and neem oil, (2) the method of application of B. bassiana and bioinsecticide, (2) entomovectorization on mites, and (3) the main risks of products on natural enemies.
2. Materials and Methods
2.1. Experimental Site
This real-world study took place from May to September 2023, at Leisure Farms, a farm located in the farming community of West Nipissing (46°20′N, 79°57′W) and approximately 100km from Collège Boréal, Sudbury, Ontario (Figure 1).
West Nipissing has a humid continental climate, with no dry season, and is classified as a temperate category (Dfb) according to the Köppen-Geiger classification. Precipitation in West Nipissing is significantly higher during the summer than in the winter. On average annually, its temperature reaches 4.3°C, accompanied by an average rainfall of 1030.1mm.
The laboratory of Collège Boréal’s Centre for Applied Research in Biodiversity (CRAB) served as a framework for the preparation of the various treatments to be applied to the field, followed by the analysis of the insect samples collected.
2.2. Inoculum, Vectors and Distributors
An end-use product of BotaniGard 22WP Biological Fungicide® (strain GHA; PlantProducts) containing a concentration of 4.4 X conidia per kg of product was used in all trials. Plantonix’s commercial neem oil has also been tested on insect pests. It is an oil extracted through cold pressing of the fruits and seeds of the Neem tree (Azadirachta Indica).
The Multi hives for bumblebees (B. terrestris) for outdoor use were obtained from Plant Products Inc. (member of the Biobest Group Belgium) and installed in a box of size (0.30m x 0.23m). They each contained a colony with a queen and 40 to 50 workers. All hives were equipped with two-way dispensers. In a two-way dispenser, the bumblebees pass through the tray of the dispenser containing the biopesticide to exit, but the inward movement is done through an empty tunnel to avoid contamination of the internal parts of the hives by the product (Figure 2).
2.3. Strawberry Varieties Used
The strawberry varieties F. x ananassa cultivar “Cabot” resulting from the cross between (Elsanta zx K79-5) and (ArKing x K7-40) were used for the experiments. The field that served as an experimental plot was established in 2022 and covers 0.5 ha. The strawberries were planted in the field in single rows (4 rows) with 10 plants per metre. No synthetic chemicals other than the experimental products designated above were used on the experimental plot. Strawberry varieties were placed on either side of the experimental variety to avoid the effects of contamination.
2.4. Experimental Methodology
Weekly surveys were carried out on strawberry plots between May and July 2023. The aim was to identify the presence of the cyclamen mite based on the characteristic symptoms of mite attacks using tools such as a magnifying glass and a manual counter. Mite counts were conducted on infested plants at the research site. Two surveys were carried out per week; ten and fifteen unfolded young leaves as well as flower buds were randomly collected and placed individually in Ziploc® bags for laboratory checks. These were immediately stored in a refrigerated cooler at 4°C for later counting in the laboratory. Each sample collected was carefully identified with information such as the sampling location, the name of the plant, the date of sampling, and the GPS coordinates of the sampling area.
Once in the laboratory, both sides of the leaflets were examined using a microscope and held wide open to clearly identify P. pallidus on the veins and folds of the leaves. In addition, the number of eggs, larvae, females and males of the pest was visually counted.
The economic threshold for deciding whether to set up trials was established by evaluating the number of mites, set at between 12 and 15 mites per plant. Following the determination of the economic threshold, four bumblebee hives were positioned inside the plots that received treatments 2, 3, 4 and 5 before the start of flowering. The entrances to the bumblebee nests remained closed until the beginning of flowering, i.e., an average of 1 to 2 flowers per plant were open.
The viability of the commercial formulation of B. bassiana, GHA strain at a concentration of 4.4 X conidia per kg of product was evaluated with a view to adjusting the amount of product used. To determine this viability, six 0.01g samples of Botanigard 22WP® were mixed with 100 ml of distilled water and Tween 80 (0.1%). Then, 200 μL of the conidia suspension was added to a 1 mL of PDA culture medium and incubated at 24 ± 1 C for 24 hours. After this period, four groups of 200 conidia were analyzed for their germination rate using a hemocytometer and a compound microscope. This viability was between 95% and 99% for all the trials conducted.
Seven experimental plots were randomly tagged on the rows. Each plot consisted of 12 strawberry plants and was covered with Hoolerry brand 43.2 cm greenhouse hoops with white insect netting (0.6mm/1” mesh). The netting provided adequate ventilation, but it isolated the flowers from both pollinating insects and bumblebee vectors. The effect of the entomopathogen on bumblebees was evaluated by adding 8 g of Botanigard to the dispenser before the first opening of the hive entrances. The distributor was examined daily to determine the quantity and condition of the B. bassiana inoculum. Every five days, the remaining powder in the dispenser was removed, and the dispenser was cleaned and filled with fresh powder. The procedure was repeated when the powder was agglomerated due to high air humidity. Seven (7) treatments were applied to evaluate the effect of B. bassiana, carried by bumblebees and neem oil, on the establishment of the cyclamen mite P. pallidus in a completely random block: untreated control without the presence of bumblebees; corn flour and use of bumblebees; combination of botanigard, corn flour and bumblebees at three different concentrations; direct spraying of neem oil; and botanigard. The treatments using Botanigard’s inoculum are as follows: T2: Control (Bumblebees + 25g maize flour); T3: Bumblebees + 3.3 x 1013 conidia/g + 25g corn flour; Size 4: Bumblebees + 2.2 x 1013 conidia/g + 50g corn flour; T5: Bumblebees + 1.1 x 1013 conidia/g + 75g corn flour). It should be noted that autoclaved maize flour is considered to be the most effective medium for the spread of B. bassiana. As for the T6 and T7 treatments respectively (14.19 Neem + 1 litre of water; 3.3 x 1013 conidia/g + 1 litre of water), they were applied to all parts of the plant using a manual sprayer with back pressure model H-7986 with 4 gallons, and size 15 x 21 in.
2.5. Statistical Analysis
The data collected were analyzed with the XLSTAT software version 10.0. The analysis of variances (ANOVA) was carried out in connection with the test of comparing variables according to classification: block, treatment, application (watering of products) and harvest. The Student-Newman-keuls (SNK) test at the 5% threshold made it possible to analyze and compare the overall average rates of leaf attack by the mite and the associated adult mortality rates.
3. Results
3.1. Distribution of B. bassiana by Bumblebees
The monitoring carried out for 10 minutes at the distributors of the B. bassiana inoculum reported the exit and entry of bumblebees from the hive. The bumblebee samples all bore viable spores on different parts of the plant (leaf, flowers). The mean number of B. bassiana spores on dead insects of P. pallidus was 3.4 x CFU.
3.2. Effect of Treatments on the Cyclamen Mite and Damage Recorded
Table 1 shows the effect of the suspension of B. bassiana spores on the cyclamen mite and the damage recorded on the leaf as a function of time. A significant difference was noted between the number of dead mites on the organs of the strawberry plant as a function of time (F = 8.122; df = 8; p < 0.0001). Between 7 and 12 days after application of the products, the average dead mite population count was 1.119 ± 0.38ab, respectively; 2,524 ± 0.81a. This mortality of dust mites decreased after 12 days to 2,048 ± 0.57ab at 17 days.
As for the damage associated with the mode of action of the cyclamen mite on the organs of the plant, especially the leaf, a significant reduction was noted as a function of time (F = 22.501; df = 8; p < 0.0001). A reduction in the rate of damage was observed between 7 and 12 days after application of the treatments, from 1.762 ± 0.22ab to 1.190 ± 0.142a. A resumption of damage was noted on the 17 days at 2,238 ± 0.20ab. This variation testifies to the effectiveness of the treatments on mites and on the reduction of the damage caused by them.
3.3. Efficacy of the Spraying Mode of B. bassiana and Neem oil on P. pallidus
Mite mortality was significantly affected by B. bassiana application methods (F = 9.348; df = 6; p < 0.0001) (Figure 3). The mean number of dead mites due to the B. bassiana contact spray method (T7) was 5.833 ± 0.226ab compared to the control (T1) which obtained 0.556 ± 0.280. In addition, the use of neem oil also induced the death of insect mites (T6) with 1.50 ± 0.121 compared to control. However, B. bassiana demonstrated effective control against P. pallidus with direct contact spraying.
3.4. Effectiveness of Bumblebee Biocontrol of P. pallidus and B. bassiana
The use of the entomovector B. terrestris for the transmission of B. bassiana at any concentration induced cyclamen mite mortality. Treatments T3, T4 and T5 caused dust mite mortality at 2,389 ± 0.229ab, respectively; 0.611 ± 0.118b; and 1,667 ± 0.114b compared to controls (T1) and (T2) on which a low mortality rate of 0.556 ± 0.280a and 0.722 ± 0.256a was noted. Effective control of the cyclamen mite was achieved with the T3 concentration.
3.5. Effect of B. bassiana Transmitted by Bumblebees and Spray Treatments on Beneficial Organisms
The ladybird Coccinella septempunctata was identified during our prospecting. The results of the analysis of variance showed that there were no significant effects on the mode of application of B. bassiana (transmission through bees and use of a sprayer) on the one hand, and on the use of neem oil on the other hand on the presence of C. septempunctata (F = 0.909; df = 6; Pr>F = 0.491). Although no significant effect of the treatments was observed on the presence of ladybugs, it is important to note that treatment 7 promoted an increase in the ladybug population from 0.222 ± 0.101a for treatment 1 to 0.333± 0.101a (Figure 4).
4. Discussion
Among the four orders of insects involved in plant pollination, insects belonging to the order Hymenoptera, especially bees, play a very essential role because of their morphology, diet, asphalting behaviour and mode of action with flowers [14,15]. Compared to honeybees, bumblebees have the great advantage of ensuring both the pollination of several flowers in a few minutes and the biological control of insect pests and diseases through the principle of entomovectorization [16]. This study explored the potential for biological control of B. bassiana (strain GHA) against the mite P. pallidus by conventional spraying and bumblebee vectoring. Our results show that, regardless of the mode of application of B. bassiana on strawberries, it had a significant effect on the cyclamen mite as well as on the damage caused by this pest compared to the control treatments (T1 and T2). These results corroborate the study of [17] He noted that entomopathogenic fungi have a good capacity to manage insects and crop pests, such as mites. Because of their specificity, these fungi can induce epizootics in pests and thus regulate their population. [18] had demonstrated the efficacy of two strains of B. bassiana on the mortality of two-spotted spider mite (TSM) populations up to 70%. The B. bassiana strain GHA used in this work is a fungal strain registered in Proposed Registration Decision 1925-0908PRD2009-03 [19]. This is a strain that, once in contact with the insect, germinates and enters the body of the latter. It produces a filament called a hyphae that induces white muscardine disease to its host and eventually kills it. It is indeed this mode of action that was identified in the dead mites at the end of the collection of samples whose spore concentration was noted at 3.4 x CFU. As for the spraying of B. bassiana, it induced a high mortality of P. pallidus compared to entomovectorization by bumblebees (T7, 5.833 ± 0.226ab; T3, 2,389 ± 0,229ab; T4, 0.611 ± 0.118b; and T5, 1.667 ± 0.114b). These results are in line with those of Macharia Kanyi et al. (2020) who have shown that spraying T. harzianum had a significant effect on the control of grey mould in strawberries than treatment with the use of entomovecteurs. Since mites hide in the hollows of young leaves, spraying has promoted contact between spores and pests; and thus created the conditions favourable to their development to the detriment of these pests. In addition, through the process of entomovectorization, bees need to carry a sufficient amount of colony-forming units on their bodies in order to ensure effective mite control, which would require bees to pass in several turns over the different plant parts to ensure a good deposition of inoculum capable of controlling this population. Macharia Kanyi et al. (2020) have demonstrated the effectiveness of bees in transmitting T. harzianum on the parts of the plant, which corroborates our results on the mortality of mites in the face of the use of bumblebees and the fungus B. bassiana. It should be noted that few studies have been conducted on the pathogenicity of GHA B. bassiana on pathogenic insects. [12] have shown that the use of B. bassiana of the active strain Balsamo was able to suppress 49% of T. vaporariorum on tomatoes, on flowers and leaves, respectively. As for the INRS IP and INRS-CFL isolates from B. bassiana Used as an insecticide, they have been shown to be 80% effective in the laboratory against tarnished plant bug adults and adults of strawberry flower anthonoma and strawberry root weevil at day seven after inoculation [21]. These results are in line with our results in the sense that either the damage caused by the P. pallidus are reduced from the seventh day after spraying. In terms of beneficial organisms, the case of the ladybird considered useful for the biological control of aphids and certain mites, no significant effect was noted with regard to the use of B. bassiana on the latter. These results are in line with those of [22] who have shown that the use of B. bassiana vectored by bees had no impact on certain beneficial insects such as the parasitoid species and A. swirskii. In addition [23] also demonstrated that B. bassiana has no effect on predators Coccinella undecimpunctataandHippodamia variegata survival, duration, longevity of adults and fertility.
5. Conclusion
This study confirmed the role of bumblebees in the transmission of entomopathogens for the control of cyclamen mites and the potential of neem-based essential oils in the management of this pest. In addition, the various products used had no impact on the natural enemies, in particular the ladybirds identified in this research. The application of these different biological control methods would thus contribute to strengthening strawberry production in Canada, reducing the use of pesticides and protecting the environment.
Acknowledgments
This study was funded through the Natural Sciences and Engineering Research Council of Canada’s Applied Research and Technology Partnership Grant to support agriculture and agri-food businesses in Northern Ontario. Our thanks go to Leisure farms for the collaboration in setting up our experiments in a real environment. We also thank the Health Sciences North Research Institute (HSNRI) for allowing us to analyze our samples in their laboratory.
References
- Agriculture et Agroalimentaire Canada, A. et A. Produire des fraises tout aussi savoureuses, mais plus résistantes aux maladies grâce aux rayons ultraviolets. http://agriculture.canada.ca/fr/science/science-racontee/realisations-scientifiques-agriculture/produire-fraises-savoureuses-plus-resistantes-aux-maladies-grace-aux-rayons-ultraviolets (accessed 2024-02-11).
- Gouvernement du Canada, S. C. Envie de vous bourrer la fraise? https://www.statcan.gc.ca/o1/fr/plus/1300-envie-de-vous-bourrer-la-fraise (accessed 2024-02-11).
- Patenaude, S.; Tellier, S.; Fournier, V. Cyclamen Mite (Acari: Tarsonemidae) Monitoring in Eastern Canada Strawberry (Rosaceae) Fields and Its Potential Control by the Predatory Mite Neoseiulus Cucumeris (Acari: Phytoseiidae). Can. Entomol. 2020, 152 (2), 249–260. [CrossRef]
- Renkema, J. M.; Pate, E.; Olivier, C. The Temporal Distribution of Cyclamen Mite, Phytonemus Pallidus (Acari: Tarsonemidae), in Strawberry and Comparison of Sampling Methods. Can. Entomol. 2022, 154 (1), e33. [CrossRef]
- Wiesmann, R.; Wiesmann, R. Investigations on the Biology and Control of the Strawberry Mite, T. Pallidus. Landw Jb Schweiz Bern 1941, 55, pts. 3, 259–329.
- Johansen, N. S.; Trandem, N.; Le, V. H.; Stensvand, A. The Potential for Using Aerated Steam to Eradicate Strawberry Mite and Two-Spotted Spider Mite on Strawberry Transplants. Exp. Appl. Acarol. 2022, 88 (3–4), 243–262. [CrossRef]
- Tamm, L.; Speiser, B.; Niggli, U. Réduction des produits phytosanitaires en Suisse: la contribution de l’agriculture biologique. 2018, 9 (2), 8.
- Réseau d’avertissement phytosanitaire. Bulletin d’informations Général; Message important de Santé Canada aux utilisateurs d’insecticides à base d’endosulfan. 3; 2017; p 2. https://www.agrireseau.net/documents/Document_95140.pdf.
- Temmermans, J.; Smagghe, G. Different Bees as Vectors for Entomovectoring with Enhanced Pollination and Crop Protection Control: Current Practices, Use Cases and Critical View on Transport. Rev. Sci. Tech. Int. Off. Epizoot. 2022, 41, 107–116. [CrossRef]
- Hazlegreaves, S. How is apivectoring innovating agricultural systems?. Open Access Government. https://www.openaccessgovernment.org/how-is-apivectoring-innovating-agricultural-systems/107527/ (accessed 2024-02-11).
- Smagghe, G., Boecking, O., Maccagnani, B., Mänd, M., Kevan, P. G. Entomovectoring for Precision Biocontrol and Enhanced Pollination of Crops; Eds. Springer International Publishing: Cham, 2020. [CrossRef]
- Kapongo, J. P.; Shipp, L.; Kevan, P.; Sutton, J. C. Co-Vectoring of Beauveria Bassiana and Clonostachys Rosea by Bumble Bees (Bombus Impatiens) for Control of Insect Pests and Suppression of Grey Mould in Greenhouse Tomato and Sweet Pepper. Biol. Control 2008, 46 (3), 508–514. [CrossRef]
- Karise, R.; Dreyersdorff, G.; Jahani, M.; Veromann, E.; Runno-Paurson, E.; Kaart, T.; Smagghe, G.; Mänd, M. Reliability of the Entomovector Technology Using Prestop-Mix and Bombus Terrestris L. as a Fungal Disease Biocontrol Method in Open Field. Sci. Rep. 2016, 6 (1), 31650. [CrossRef]
- Crepet, W. L. Advanced (Constant) Insect Pollination Mechanisms: Pattern of Evolution and Implications Vis-a-Vis Angiosperm Diversity. Ann. Mo. Bot. Gard. 1984, 71 (2), 607. [CrossRef]
- Michener, C. D. The Bees of the World, second edition edition.; The Johns Hopkins University Press, 2007.
- Wahengbam, J.; Raut, A.; Pal, S.; Banu, A. N. Role of Bumble Bee in Pollination. Ann. Biol. 2019, 35, 290–295.
- Dara, S. K. Implementation of IPDM in Strawberries and Other Berries. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M. L., Albajes, R., Nicot, P. C., Eds.; Plant Pathology in the 21st Century; Springer International Publishing: Cham, 2020; pp 597–624. [CrossRef]
- Michereff-Filho, M.; Navia, D.; Alexopoulos Quevedo, I.; de Almeida Magalhães, M.; Wagner da Silva Melo, J.; Biaggioni Lopes, R. The Effect of Spider Mite-Pathogenic Strains of Beauveria Bassiana and Humidity on the Survival and Feeding Behavior of Neoseiulus Predatory Mite Species. Biol. Control 2022, 176, 105083. [CrossRef]
- Agence de réglementation de la lutte antiparasitaire du Santé Canada. Projet de décision d’homologation PRD2020-01. 2009, No. H113-9/2009-3F (H113-9/2009-3F-PDF), 89.
- Macharia Kanyi, J.; Gikungu, M.; Karanja, R.; Okoth, S. African Journal of Agricultural Research Managed Bees as Pollinators and Vectors of Bio Control Agent against Grey Mold Disease in Strawberry Plantations. Afr. J. Agric. Res. 2020, Vol. 16(12), 1674–1680. [CrossRef]
- Sabbahi, R. Utilisation du champignon entomopathogène Beauveria bassiana dans une stratégie de gestion phytosanitaire des principaux insectes ravageurs en fraiseraies. phd, Université du Québec, Institut National de la Recherche Scientifique, Québec, 2008. https://espace.inrs.ca/id/eprint/285/ (accessed 2024-02-11).
- Shipp, L.; Kapongo, J. P.; Park, H.-H.; Kevan, P. Effect of Bee-Vectored Beauveria Bassiana on Greenhouse Beneficials under Greenhouse Cage Conditions. Biol. Control 2012, 63 (2), 135–142. [CrossRef]
- Sayed, S.; El Arnaouty, S.; Alotaibi, S.; Salah, M. Pathogenicity and Side Effect of Indigenous Beauveria Bassiana on Coccinella Undecimpunctata and Hippodamia Variegata (Coleoptera: Coccinellidae). Insects 2021, 12, 12. [CrossRef]
Figure 1.
study areas (Leisure farm) located in West Nipissing and the Centre for Applied Research in Biodiversity (CRAB) at Collège Boréal in Sudbury (Images retrieved from Google Earth on January 19, 2024).
Figure 1.
study areas (Leisure farm) located in West Nipissing and the Centre for Applied Research in Biodiversity (CRAB) at Collège Boréal in Sudbury (Images retrieved from Google Earth on January 19, 2024).
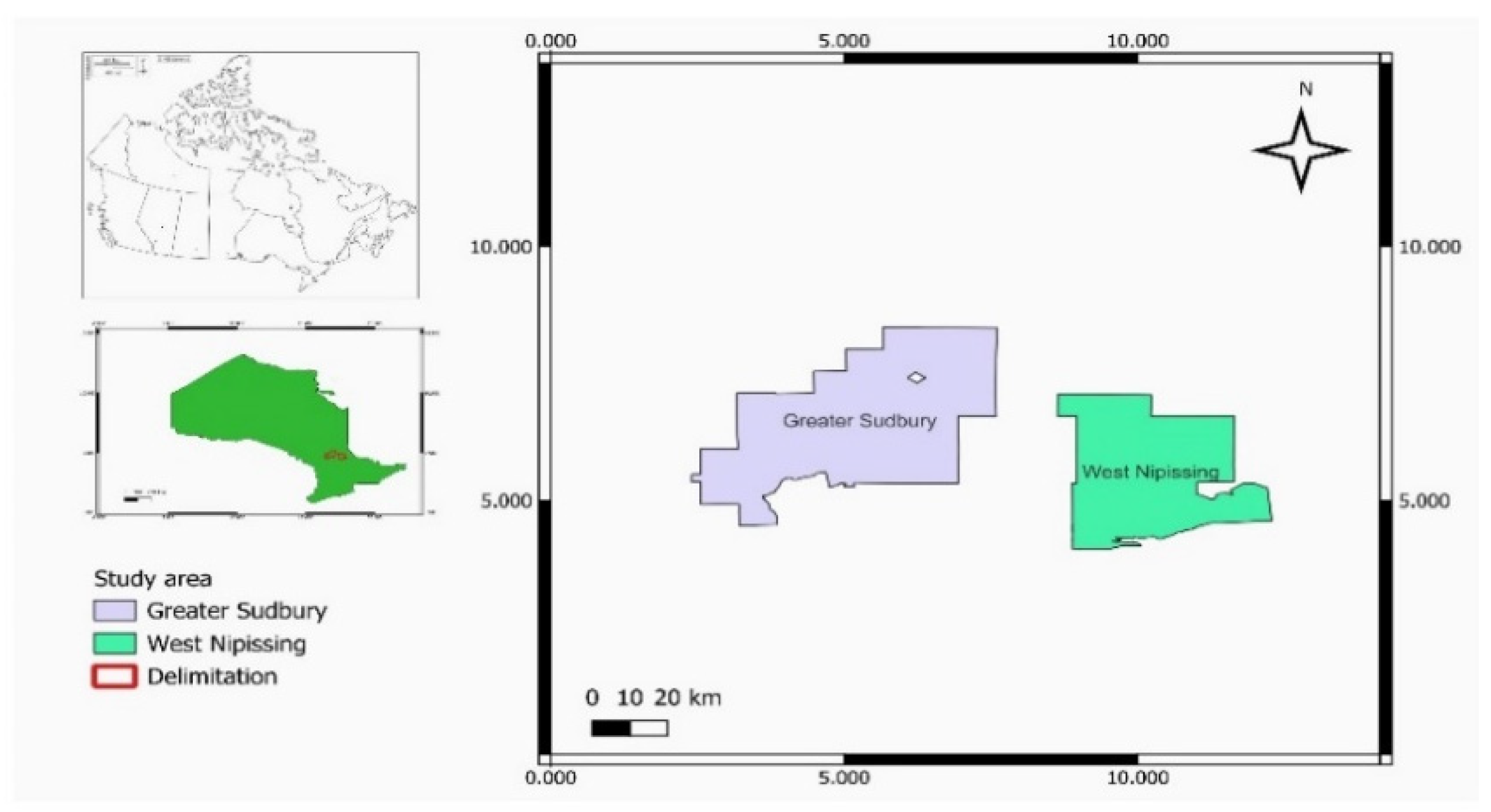
Figure 2.
Bumblebees provided by Biobest Canada Ltd. in 2023 and disposal in the fields.

Figure 3.
average number of dead mites caused by different concentrations of conidia and neem oil. T1: control without application; T2: Control (Bumblebees + 25g corn flour); T3: Bumblebees + 3.3 x 1013 conidia/g + 25g corn flour; Size 4: Bumblebees + 2.2 x 1013 conidia/g + 50g corn flour; T5: Bumblebees + 1.1 x 1013 conidia/g + 75g corn flour); T6 (14.19 Neem + 1 liter of water) and T7 (3.3 x 1013 conidia/g + 1 liter of water).
Figure 3.
average number of dead mites caused by different concentrations of conidia and neem oil. T1: control without application; T2: Control (Bumblebees + 25g corn flour); T3: Bumblebees + 3.3 x 1013 conidia/g + 25g corn flour; Size 4: Bumblebees + 2.2 x 1013 conidia/g + 50g corn flour; T5: Bumblebees + 1.1 x 1013 conidia/g + 75g corn flour); T6 (14.19 Neem + 1 liter of water) and T7 (3.3 x 1013 conidia/g + 1 liter of water).
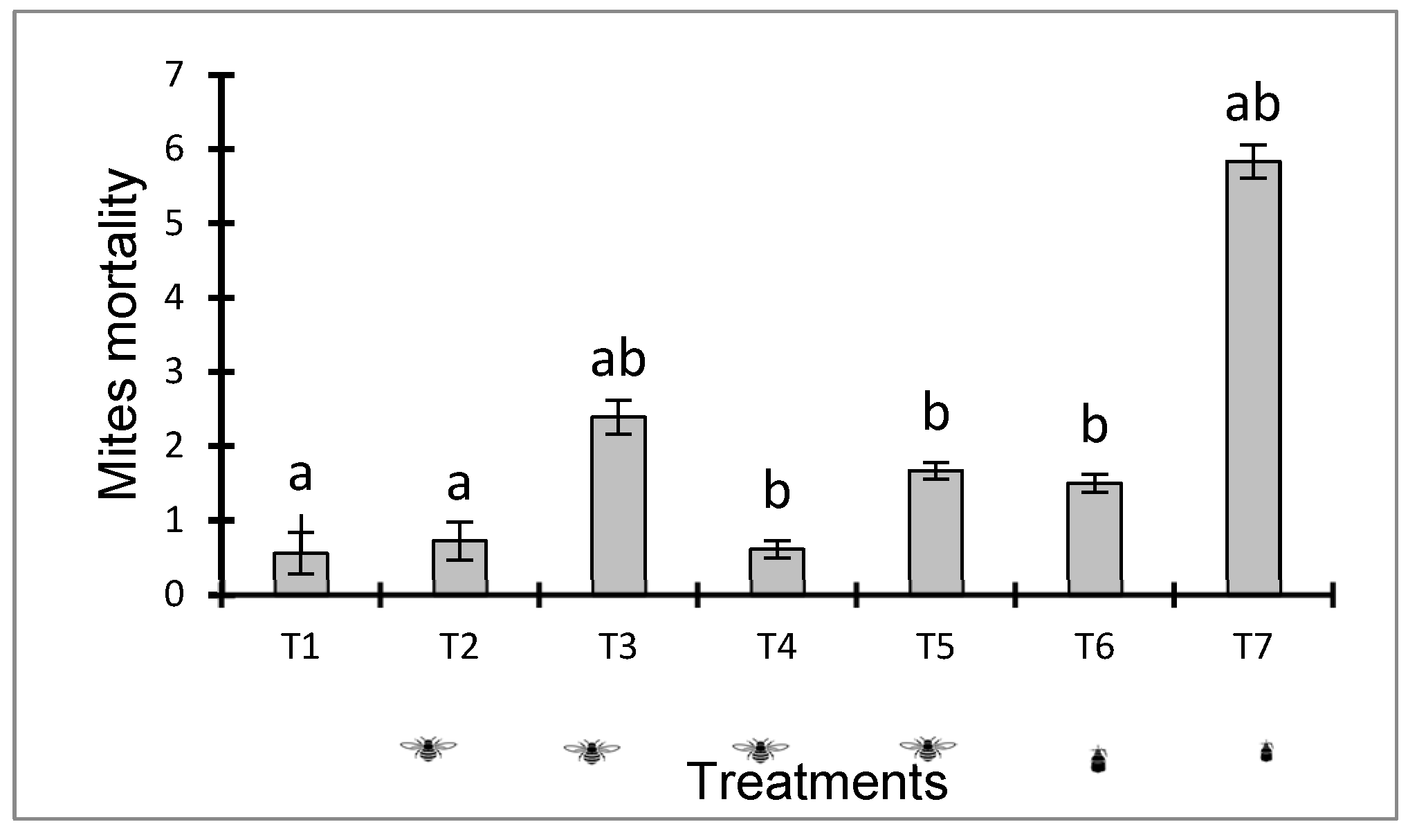
Figure 4.
effect of treatments on beneficial insects.
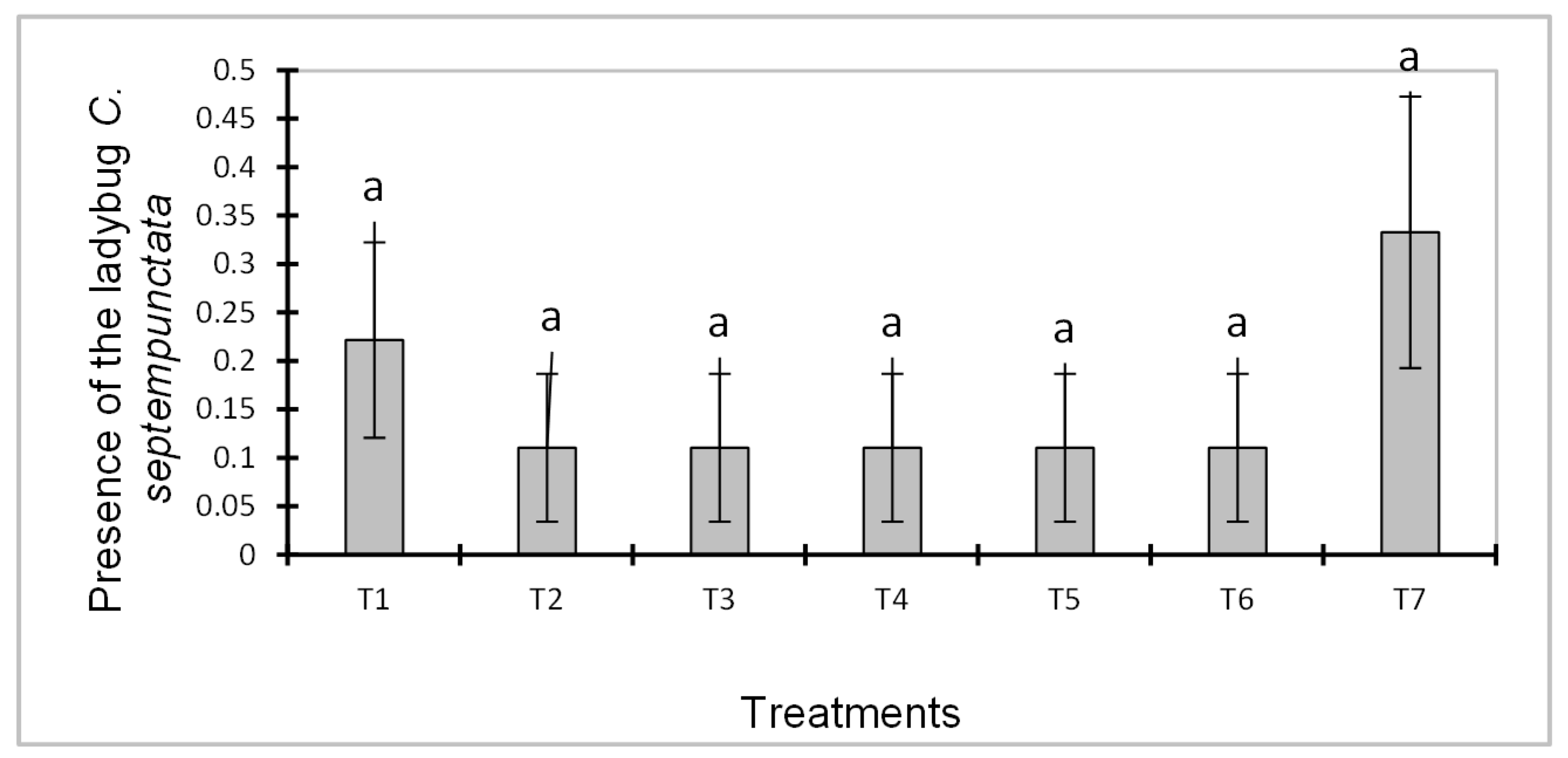

Table 1.
Average rate of damage observed on the plant (curled, stunted and dried leaves).
| Time after application of treatments | Leaves and flowers | |
| **% P Damage. pallidus | *Number of dead mites of P. pallidus | |
| 7JHA | 56,571 ± 0.041ab | 1.119 ± 0.38ab |
| 12JHA | 22,571 ± 0.054a | 2.524 ± 0.81a |
| 17JHA | 56,571 ± 0.041ab | 2,048 ± 0.57ab |
Values followed by the same letter are not significantly different at the 5% level according to the analysis of variance followed by SNK.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



