
Submitted:
27 September 2024
Posted:
30 September 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
During the last few years, the world witnessed one of the worst calamities affecting humankind- the COVID-19 pandemic. We also witnessed the development of vaccines at an unprecedented pace against the SARS-CoV-2, the causative agent. India, which was one of the countries most impacted by COVID-19, developed its indigenous vaccine in addition to utilizing the ones developed by other countries. Initially, the antibody level and the neutralizing antibody titer against the pathogen were considered as the correlates of immune protection from the disease. The long-term protection from the pathogen is provided by the immune memory cells viz. T and B memory cells and their ability of efficient recall responses. In this regard, global research focused mainly on the mRNA-based vaccines. The studies on immune memory response, particularly- the B cell memory response bestowed by the vaccines given to Indians is relatively obscure. In this work, we examined the level of RBD specific memory B cells in the peripheral circulation of the study participants and the ability of B cells to secrete antigen-specific antibodies among Indians vaccinated with Covaxin (BBV152), Covishield (AZD1222), Corbevax (BECOV2D) and Sputnik Light as well as unvaccinated individuals. Our study revealed that Corbevax and Sputnik Light conferred better antibody-secreting cell (ASC) responses.
Keywords:
Receptor Binding Domain (RBD)
; Memory B-cell (MBC)
; Antibody-secreting cell (ASC)
; Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
; Coronavirus Disease-2019 (COVID-19)
Introduction
Vaccines are used to protect people from different diseases, and to limit their transmission to a larger population. Since the inception of the concept of vaccination in 1796, there have been tremendous advancements in the field of vaccinology. From the early days’ whole-pathogen-inactivated vaccines to the modern mRNA vaccines, the field has shown great progress reaching a crucial turning point during the recent COVID-19 pandemic [1]. Vaccines against SARS-CoV-2 have proved their effectiveness in mitigating severe symptoms and reducing hospitalization, however, the cellular memory responses imparted by them remain largely uncharacterized [2]. Besides antibodies, the generation and maintenance of immune memory is an important element in tackling any future encounters with the pathogen [3]. Generation of immune memory is the change in the immune status of the host so that its footprint lasts for a longer duration [4]. Both T and B cells play critical roles in generating immune memory and help in shaping each other's functionality [5,6]. Cytotoxic CD8+ T cells are important in killing and eliminating the cells infected with an intracellular pathogen such as a virus, while CD4+ T cells generate a cytokine milieu conducive for the maturation of the B and T cells, and concomitantly development of their memory phenotype [7,8]. Long-lasting memory B cells (MBCs) are generated in response to an infection or vaccination and add a further layer of protection [9]. B-cell affinity maturation occurs in the transient but specialized structures known as ‘germinal centres’ [10]. After a proliferative stage where a B-cell clone undergoes somatic hypermutation and antigen-based selection, only those with the ability to effectively bind an antigen survive [11,12]. This process continues for several months after vaccination or an infection. This makes the newer generation B cells produce antibodies with better avidity and superior ability of antigen neutralization. A subset of these B cells- become MBCs that guide the immune response in case of any subsequent encounter with the antigen [5]. The affinity maturation process expands the breadth of recognition with increased affinity of the antibodies and the generated pool of the MBCs are multi-pronged with anticipatory memory potential [12]. Notably, in addition to IgG, IgA and IgM are also capable of neutralizing the viral antigens [13,14].
A comparative assessment of memory B-cell response elicited by different vaccines holds the potential to provide invaluable insights of their relative efficacies in promoting enduring immune memory, thereby facilitating the development of evidence-based vaccination strategies [15]. It is also important to assess the level of memory cells present in the peripheral circulation of the vaccinees over time, as well as their ability to respond to the recurrent invasion by the pathogen [16]. There are concerns regarding potential decline in the vaccine efficacy, as viruses carrying mutations in key neutralizing antibody epitopes propagate in the community, allowing them to partially/completely evade the antibody recognition [17,18]. There are also reports that in certain cases of severe COVID-19 the affinity maturation of the B cells is compromised [19]. In India, three viral vector-based vaccines (Covishield, Sputnik Light and Sputnik V), an inactivated virus-based vaccine (Covaxin)[20], and a protein subunit vaccine (Corbevax) were widely administered [21]. Covaxin, manufactured by Bharat Biotech International Ltd uses the whole SARS-CoV-2 virus in an inactivated form. Corbevax, from Biological E. Limited India, utilizes a yeast (Komagataella phaffii)-produced version of the SARS-CoV-2 spike protein's RBD along with adjuvants (aluminium hydroxide gel and CpG1018) to trigger an effective immune response. It is based on the formulation well-accepted for the recombinant hepatitis B vaccine [22]. Covishield is manufactured by Serum Institute of India Pvt. Ltd., a two-dose COVID-19 vaccine that uses a chimpanzee adenovirus vector called ChAdOx1, designed to deliver SARS-CoV-2 spike protein into the host cell. The Sputnik Light vaccine manufactured by Gamaleya Research Institute of Epidemiology and Microbiology, Russia is a single-dose regimen that also utilizes a recombinant human adenovirus type 26 (rAd26) vector. It is the first dose of the Sputnik V (Gam-COVID-Vac) vaccine used as a standalone for vaccination [23,24]. All these vaccines showed great potential during phase 3 trials with 77.8% efficacy for Covaxin (BBV152) [25], 70.4% for Covishield (AZD1222/ChAdOx1) [26], 91.6% for Sputnik V (Gam-COVID-Vac)[27], and >90% for Corbevax (BECOV2D) [28].
Despite the success shown by these vaccines during the clinical trials, the protection set forth by them had a waning effect. Although it is known that the absence of antibodies need not necessarily correspond to the absence of immune memory, a considerable amount of breakthrough cases were observed in many of the vaccinated cohorts, though with less disease severity [29,30,31,32]. This is partly linked to the immune evasion ability of the virus due to a high mutation rate observed in SARS-CoV-2 genome [33]. However, comprehensive data is not available to convincingly explain this immunological insufficiency that upsurges with time. Moreover, in India, only a few in-depth studies have been conducted on post-vaccination immunology despite India’s COVID-19 vaccination drive being one of the most extensive ones in the world. Therefore, the study of the long-term effects of immunological memory imparted by COVID-19 vaccines given in India is important to understand how immunological memory is established against this virus.
In this study, we chose all the above-mentioned vaccines differing either in their development platform or dose regimen, to study the diverse aspects of the antigen specific B-cell immune response three months after vaccination. MBCs are made to quickly proliferate and differentiate into antibody secreting cells upon re-exposure to the pathogen [34]. Our readouts were based on the Receptor Binding Domain (RBD) of the Spike protein of the SARS-CoV-2 Wuhan strain. A corollary is that immune response against RBD has been shown to be dominant and correlates well with the virus neutralization [16,35,36,37]. We compared the memory B cell response generated by the above-mentioned vaccines in the peripheral circulation of these individuals and the ability of these cells to proliferate and secrete anti-RBD antibodies. We observed a noticeable effect of Corbevax and Sputnik Light vaccines on the ASC response of the vaccinated participants in comparison with the unvaccinated individuals. Some of our findings have also been corroborated by the studies on the T-cell response by other groups [22,38].
Materials and Methods
Ethics statement
This study was conducted at the BRIC-Translational Health Science and Technology Institute, Faridabad, India. Participant enrolment and sample collection were done at ESIC Medical College and Hospital, Faridabad. Written informed assent/consent was obtained from each of the study participants. This study was approved by the Institutional Ethics Committees from both institutes (ESIC Hospital and Medical College, Faridabad File no._134 X/11/13/2021- IEC/43 and BRIC-THSTI Faridabad Ref No: THS 1.8.1/ (130) dated 27th Oct 2021).
Participant details
Samples were collected from a total of 171 participants and their age, sample collection date, and vaccination details were recorded. The participants were of the following categories- Unvaccinated - 35 participants and Vaccinated (Corbevax- 34, Covaxin- 33, Covishield- 40, and Sputnik Light- 29) (Table 1). The sample collection for this study coincided largely with the Omicron-I (December 2021-June 2022) and partially with the Omicron-II (July 2022–October 2023) period.
Inclusion and exclusion criteria
The analysis included participants aged 18-59 years having no documented history of SARS-CoV-2 infection. Eligible individuals were either unvaccinated or had completed a full vaccination course (two doses for BBV152, AZD1222, and BECOV2D; a single dose for Sputnik Light). Participants were excluded if they had obtained a booster vaccine dose prior to specimen collection or had reported a prior infection at the time of enrolment.
Blood collection and sample processing
Eight ml of blood was collected from each participant in the Greiner Bio-One tubes (Cat. No. 22-040-134) coated with sodium heparin. Blood samples were processed immediately to isolate plasma and peripheral blood mononuclear cells (PBMCs) using Lymphoprep density gradient centrifugation. Plasma was stored at -80 °C and PBMCs were resuspended in fetal bovine serum (FBS) supplemented with 10% dimethyl sulfoxide (DMSO) and then stored in liquid nitrogen [34].
Indirect anti-RBD IgG ELISA
96-well Maxisorp plates (Thermo Fisher Scientific- Cat No. 442404) were coated with 100 µl of RBD protein (2 µg/ml in PBS) per well and incubated overnight at 4°C. Plates were washed with PBST (PBS + 0.1% Tween 20) and blocked for two hours at 37°C with the block buffer (PBST + 3% skimmed milk). After incubation, the block buffer was discarded, and 100 µl per well of the diluted Plasma samples were added in duplicates. Plasma samples were diluted in PBST + 3% skimmed milk at a ratio of 1:150. The plate was incubated for 30 minutes at 37°C and then was washed with PBST. Further, 100 µl per well of the secondary antibodies was added. Goat anti-human IgA HRP-conjugated (Southern Biotech 2050-05; 1:5000 dilution), Goat anti-human IgG HRP-conjugated (Jackson Immunoresearch 109-035-088; 1:10000 dilution) and Goat anti-human IgM HRP-conjugated (Jackson Immunoresearch 109-035-129; 1:10000 dilution)] were the secondary antibodies used in this study. The plate was incubated for 30 minutes at room temperature and then washed with PBST. TMB substrate was added in the dark and incubated for 3 minutes. The reaction was stopped with 1N H2SO4 before measuring the optical densities at 450 nm using a microplate reader [39].
Expression and Purification of the RBD protein of SARS-CoV-2 (Wuhan strain- Hu-1)
The following reagent was contributed by David Veesler for distribution through BEI Resources, NIAID, NIH: Vector pcDNA3.1(-) Containing the SARS-Related Coronavirus 2, Wuhan-Hu-1 Spike Glycoprotein Receptor Binding Domain (RBD), NR-52422as mentioned previously [40,41]. The recombinant his-tagged SARS-CoV-2 ancestral Wu-RBD protein was expressed in transiently transfected Expi293F cells in suspension culture using Expifectamine Transfection kit (Thermo Fisher Scientific, Cat no. A14524) as per manufacturer protocol. Post-transfection and expression, cell culture supernatants were harvested after 5–6 days or until cells showed more than 60% cell death. The supernatant was passed through a Ni–NTA column for protein purification. Bound proteins were eluted with 500 mM imidazole and were concentrated with an Amicon 10 kDa filter (Millipore), protein fractions were aliquoted and stored at −80 °C.
Labelling of RBD protein with the Alexa Fluor-488
Purified RBD was tested using SDS PAGE to verify its purity, and was used for the fluorophore labelling at a concentration of 1 mg/ml. We labelled RBD protein using Alexa Fluor Microscale Protein Labelling kit (Cat No. #A30006; Thermo Fisher Scientific) as per the manufacturer’s instructions. Fluorophore labelled RBDs were aliquoted and stored at -20 °C for further use.
Estimation of Antibody Secreting Cells
Antigen specific antibody secreting cell estimation was performed as described by Crotty et al., 2004 with slight modifications. PBMCs were washed after thawing with complete RPMI medium, counted and cultured at a density of 3 million cells per sample, and added at 0.5 million cells per well in a 24-well plate. An additional 0.5 million cells were cultured in a medium without stimulants as a control. Polyclonal stimulation reagents used were Protein A from Staphylococcus aureus (Sigma Aldrich P7155), Lectin from Phytolacca americana (pokeweed) (L9379), and ODN 2006 (TLR GRADE®) (synthetic) (Enzo Life Sciences- ALX-746-056-M001). The culture plate was kept in a CO2 incubator for 5 days at 37°C.
An ELISpot plate (Sigma Aldrich-MSIPS4510) was activated with 35% Ethanol. After washing with PBST (PBS + 0.05% Tween 20) and PBS respectively, the plate was coated with polyvalent Goat anti-human Ig mix (Thermo Fisher Scientific - H17000) and SARS-CoV-2 spike RBD at a concentration of 10 μg/ml in PBS. The plate was incubated overnight in the dark at 4°C. Following coating, the plate was washed and blocked with 200 µl of blocking buffer per well for at least 2 hours at 37°C or overnight at 4°C. After culturing for 5 days, cells were washed and seeded onto the ELISpot plate ( Figure 3). Subsequent steps included incubation with secondary antibodies [(Goat Anti-human IgA secondary antibody, Biotin (A18785), Goat Anti-human IgG Fc Bio Affinity (Thermo Fisher Scientific - A18821), Goat Anti-human IgM secondary antibody, Biotin (Thermo Fisher Scientific - PA1-86071)], Avidin-D-HRP (Thermo Fisher Scientific 18-4100-51), and substrate solution. The substrate was prepared by mixing 110 µl of AEC-DMF solution (3-Amino-9-Ethylcarbazole in Dimethyl Formamide, 60 mg/mL) with 10 mL of 0.1 M sodium acetate and adding 165 µl of H2O2 [Sigma Aldrich (323381)] after filtering the solution. The plate was incubated for development of spots, rinsed gently with tap water, and dried overnight in the dark.
Spot counting was performed using the Auto-counter feature of the ImmunoSpot 7.0.36.0 device with CTL software, which visualized and recorded the number of spots in each well. Quality control was performed to check for any errors in the counting and corrections were made accordingly. The average of the spot-forming units (SFUs) from the unstimulated control wells was deducted from each of the SFU values.
Estimation of memory B cells in the peripheral circulation
To estimate the percentage of RBD-specific memory B cells in the peripheral circulation of the study participants, a flow-cytometry-based method was utilized [16]. Flow cytometry was performed on a subset of samples only when sufficient PBMCs remained after culturing them for the ELISPOT assay. Due to the limited availability of the cells, not all participants' samples could be analyzed for the estimation of the MBCs in the peripheral circulation. Alexa Fluor-488 labelled RBD was used as a probe for estimation of antigen specific memory B cells. Briefly, 1.5-2.5 x 106 cells were labelled with antibodies against different human B cell molecular markers which included markers for memory phenotype as well (Table 2). Samples were acquired on Canto II Flow cytometer (BD Biosciences) and the data were analysed using FlowJo 10.3 (FlowJo LLC).
Statistics
All the data visualization and statistical analyses were performed using GraphPad Prism 10.2.0. Data was checked for normal distribution before analysis. The outliers were removed using iterative Grubb’s test (Alpha = 0.01). Datasets are visualized as bar charts depicting the median and interquartile range. Depending on the data, appropriate tests from Mann-Whitney U/Wilcoxon rank sum, Kruskal-Wallis’ and Dunn’s multiple comparison tests were used for statistical analyses. A p value of < 0.05 was considered statistically significant.
Results
All participants including unvaccinated individuals showed marked levels of anti-RBD IgG antibodies
We tested the IgG antibody levels of the participants in our study using RBD-specific IgG ELISA [39]. Vaccinated participants from all four vaccines showed a remarkable IgG response towards SARS-CoV-2 RBD, indicating that the vaccinations led to the development of a distinct humoral response against the virus ( Figure 1). However, the unvaccinated participants also showed an elevated RBD-specific IgG response, which suggests their possible exposure to SARS-CoV-2 during the pandemic. For this study, the blood samples from unvaccinated participants were collected during Omicron surge in India, so it is likely that these participants contracted the virus during this period and remained asymptomatic [42]. The presence of IgM antibodies post-recovery phase for a prolonged time among these participants highlights the importance of these multimeric antibodies in tackling the virus. Similar observations have also been made in certain other viral infections. Possibly due to the higher avidity of these antibodies towards the antigens, host continue to make these antibodies in the backdrop of the persistent antigenic challenge [13,43].
Study participants exhibited RBD-specific memory B cells in the peripheral circulation
Our analysis of the MBC-responses to the SARS-CoV-2 RBD in the peripheral blood of the participants yielded mixed results. Flow cytometry analysis on peripheral blood mononuclear cells (PBMCs) from 113 participants (85 vaccinated and 28 unvaccinated) revealed a higher percentage of RBD-specific memory B cells in the unvaccinated group as compared with any of the vaccinated groups. Although we recruited these unvaccinated participants based on their asymptomatic profile during the last three months, they showed a good antibody response against SARS-CoV-2 RBD. This is probably because of a fresh natural (asymptomatic) exposure of the unvaccinated participants to the pathogen at the time of Omicron surge in the country ( Figure 2). All the participants of vaccinated groups showed comparable percentages of RBD specific memory B cells in the peripheral blood. Similar to our observation for the RBD specific antibody levels in the plasma samples, the group of unvaccinated participants had a slightly higher amount of RBD specific memory B cells as compared to all the vaccinated groups in the peripheral circulation. This is probably due to a recent exposure to the virus. However, there was no statistically significant difference seen among participants of vaccinated groups when compared with that of the unvaccinated participants.
Magnitude of the RBD-Specific antibody-secreting B cells among vaccinees from different vaccine groups
To test the magnitude of the RBD-Specific antibody-secreting B cells among vaccinees from different vaccine groups, we did polyclonal stimulation of the PBMCs and tested the ability of the B cells to proliferate and secrete antigen specific antibodies [34]. The schematic diagram of the plate map and a representative image of the B cell ELISPOT plate have been shown in Figure 3 A and B respectively. Our data reveals a distinct distribution of RBD-specific IgM, IgA and IgG antibody secreting B cells among the participants. The percentage of RBD-specific IgM antibody secreting B cells was higher than that of IgA and IgG antibody secreting B cells across all the analyzed groups (Fig 4). However, it was almost similar across all the participant groups. The median percentage of IgA and IgG antibody secreting B cells were slightly higher among participants vaccinated with the Corbevax and Sputnik Light groups as compared with the other groups including the unvaccinated participants. Covaxin also mounted a slightly better IgA response when compared with those of the unvaccinated groups. There were no statistically significant differences in the ASC response between the groups because of a considerable variability as evident from Figure 4. The increase in the median percentage of IgA and IgG antibody-secreting B cells in the Corbevax and Sputnik Light group participants and IgA response also in the Covaxin vaccinated participants is of significance when considering the vaccine induced immune memory response. Despite the fact that these groups had lower RBD-specific IgG antibodies in their plasma samples (Fig.1) and comparatively less RBD-specific memory B cells in circulation compared to the unvaccinated group (Fig.2), a reversal in the pattern was observed in their ASC responses. It should be noted that a single dose Sputnik Light vaccine resulted in a marked level of the level of RBD specific IgG antibody secreting B cells comparable to those produced by other vaccines after two doses.
Temporal Patterns in the RBD-Specific Antibody-Secreting B Cells Post-Vaccination
To test the durability of the ASC response of the participants of different vaccine groups, we categorized the samples of each vaccination group by 4-6 months and beyond 6 months post vaccination. The levels of IgA and IgM-secreting B cells among the Corbevax-immunized participants post 6 months of vaccination, did not differ significantly from the participants in the unvaccinated ( Figure 5A). However, they exhibited a statistically significant increase in RBD specific IgG ASC response as compared to the unvaccinated participants, suggesting an enhanced IgG antibody response among these participants over time ( Figure 5A). However, at early time points a slight enhancement in the median of the ASC response for only IgG was observed among the Corbevax vaccinated participants (Supplementary Figure 1). The ASC response after Covaxin administration remained more or less similar to that of the unvaccinated group among the samples obtained post 6 months of vaccination ( Figure 5B). We observed an increase in the median RBD specific IgA and IgG SFUs at an early time point with a huge inter-individual variation (Supplementary Figure 2). However, the above result cannot be considered conclusive, as the number of samples in the 4-6 months post vaccination group are not sufficient. Participants immunized with Covishield did not show a noticeable trend in the memory B cell immune response for any of the antibodies. The percentage of RBD-specific antibody-secreting B cells remained similar for 4-6 months and afterwards in these participants ( Figure 5C and Supplementary Figure 3). Individuals who received the Sputnik Light vaccine displayed an enhancement in the levels of RBD-specific IgA and IgG antibody-secreting B cells among the participants after 6 months post vaccination ( Figure 5D).Although we did not observe a marked difference in this regard at early time points (Supplementary Figure 4).
This observation suggests the development of an enduring and broader antibody response over time following Sputnik Light vaccination. A limitation in drawing definitive conclusions about impact of various vaccines on temporal dynamics of ASC responses is the relatively small number of the participants vaccinated with Covaxin, Covishield and Sputnik Light within the 4–6-month time window (Supplementary Figure 1-4).
No gender-specific disparities observed in serological or memory immune responses
A considerable number of males and females participated in this study, as evident from the Table 1B. We performed a sex-disaggregated analysis of the immune responses to evaluate any potential disparities between males and females. As shown in Figure 6, our analysis revealed no statistically significant differences in serological immune responses or B cell memory responses between male and female participants. This finding indicates that gender does not appear to influence the B cell immune response to vaccination. Despite the lower number of vaccinated females compared to males, our data suggest that the immune responses are comparable across the two genders.
Discussion and Conclusion
Understanding the immune memory response is crucial in getting insights into the longevity of the protection against a pathogen. The memory response of T and B cells and their functionality are shaped not only by the antigens of a pathogen and their presentations but also by the interactions between these two cell types [44]. The B cell memory phenotype is generated as a result of multiple iterations of antigen-based selection of antibody-secreting B-cell populations [45]. The instability of the viral genome and the resulting mutations in the antigens, particularly the Spike protein in case of the SARS-CoV-2, diminishes the impact of B-cell memory response during a reinfection or a breakthrough infection [46]. The efficacy of various vaccination platforms against SARS-CoV-2 and natural infection in conferring immunity has been a subject of extensive research and debate [21]. It has been shown that MBC responses were more pronounced when individuals experience a natural infection followed by a single vaccine dose as opposed to vaccination alone [47]. These studies show the impact of various vaccines on generating immune memory, or enhancing the one already produced due to a natural infection.
We observed that the unvaccinated individuals also exhibited antibody response as well as memory B cells with proliferative capability in their peripheral circulation. This is probably due to the asymptomatic infections they might have had during the peak of the Omicron wave in the country [42]. Studies regarding memory B cells in the peripheral circulation and the estimation of ASCs have been done among the BNT162b2 vaccinated and naturally infected individuals. The generation and durability of the MBCs has been linked to the severity of the disease, where moderate to severe infections have resulted in generating ability to mount a better B cell recall response. Several studies show that the overall antibody titers decline rapidly a few months after infection and vaccination eventually after attaining a peak in the initial days [48,49,50]. Relatively few studies have been performed to gain insight into the protection provided by memory B cells. Most of the studies in this field have focused on the mRNA-based vaccines [51,52,53,54,55,56]. A study carried out with a cohort of BNT162b2 vaccinees noted an increase in the percentage of pathogen specific memory B cells eight months after the second dose, and even further increase after a booster dose [57,58]. Nayak et al. observed a positive correlation between RBD-specific memory B cells in peripheral circulation and RBD-specific IgG titers. They also mentioned that the individuals with a low amount of neutralizing antibodies also had a low amount of memory B cells in their peripheral circulation [16]. Various aspects of immunological memory produced by BNT162b2 vaccination against SARS-CoV-2 have been widely studied [55,57,59]. In India where the BNT162b2 vaccine was rarely administered, such studies are lacking.
In this study, blood samples particularly of the unvaccinated groups were collected during the Omicron surge in India and our results suggest that these unvaccinated participants were exposed to the virus. We didn’t have a study group of uninfected and unvaccinated participants to serve as a perfect comparator for our study. Despite this limitation, these results give us a glimpse into the potential complexity of the B cell immune response. Among the vaccinated groups, the fraction of ASCs shows variation from vaccine to vaccine. BECOV2D and Gam-COVID-Vac showed promising results with slight increase in the number of ASCs six months after vaccination- like what was previously observed in BNT162b2. On the other hand, though BBV152 and AZD1222 induced a considerable number of ASCs after vaccination, they did not show any significant change in the B cell recall response over time. These vaccines induced a B-cell recall response like a mild infection, but its temporal dynamics depended on the specific vaccine. These findings highlight the complex interplay between vaccination, prior infection and the pathogen-specific memory B cell generation and their maintenance. We would like to mention here that these responses are probably the mixed result of natural infection and vaccination.
Although the ASC estimation is tedious to perform and takes almost a week to the results, the study needs to be carried out on a larger sample size for developing better understanding of the B cell memory and its recall response. Larger sample size would have also helped us in understanding temporal dynamics of the ASC response mounted by different vaccines with a clearer conclusion. A limitation in our study is the exposure status of the unvaccinated participants. It would have been better to have PBMCs and blood plasma collected from the people before the pandemic to have an actual comparison between the vaccinated and particularly unvaccinated groups. We observed a substantial interindividual variation among these responses with heterogeneity at the level of RBD-specific memory B cells in the peripheral circulation as well as in the secretion of antigen-specific antibodies Our study is one among a few done to understand the memory B cell response, particularly the ability of immune recall responses after vaccination against COVID-19 [60,61]. This aspect of evidence-based understanding was lacking for those vaccines administered to Indians. The study of the memory B cell response is essential in understanding the long-term protection offered by vaccinations. The memory response of the B cells mounted by the vaccines to an infection, its durability, and effectiveness, play a key role in preventing the recurrence of the disease. Our study highlights the immune memory response, particularly the B cell memory response generated after vaccination against COVID-19. The levels of memory B cells generated by vaccination, or the natural infection serve as a better immune correlate of protection than the antibody levels in the blood circulation [62].
A high percentage of individuals infected during the Omicron wave in India remained asymptomatic [42]. Possibly due to asymptomatic exposure to the virus, even the unvaccinated individuals exhibited high levels of RBD-specific IgG antibodies as observed in this study. They also showed the presence of RBD-specific B cells in their peripheral circulation. However, the vaccinated people, especially those vaccinated with Corbevax and Sputnik Light showed better response in terms of secreting RBD-specific antibodies after polyclonal stimulation. This suggests an effective and durable B cell memory response generated by these vaccines. This knowledge could also help in making policy decisions by the Government bodies, preferential use of a particular vaccine development platform or whether we need a booster vaccination for such infectious diseases. This study is also unique in guiding tailored and more equitable vaccination strategies based on the understanding of the durability of the memory response of the B cells, its ability to proliferate and secrete antigen specific antibodies.
Data availability
Authors will provide data keeping the participants’ details confidential and as per the journal policy.
Acknowledgments
Authors wish to acknowledge BRIC-THSTI, Faridabad for intramural funding support to this work. We would like to acknowledge Dr. Sankalp and Dr. Rohit Dhaka from ESIC Medical College and Hospital, Faridabad for their help in participant enrollment. We also thank Amit Kumar Yadav, Aftab Hussain, Richa Kumari, Deepak Rathore, Akshay Binayake, Aymaan Jaheer, Manas Ranjan Tripathy and Jitender Chandilla, Immunobiology and Immunotherapy laboratory, BRIC-THSTI, Faridabad for the suggestions and help. We are thankful to the participants for donating their blood samples which made this study feasible. We are also thankful to the BEI resources for providing the RBD-Wuhan plasmid NR-52309. The figure for the graphical abstract is created by using BioRender.com
Author’s contribution
Atharv Athavale- performed the experiments, made tables and figures, wrote, reviewed and edited the different versions of the manuscript; Anmol Gaur and Nafees Ahmed- collected samples and performed the experiments; Adarsh Subramaniam, Jyotsana Dandotiya and Sneha Raj- performed the experiments; Santosh K. Upadhyay- edited and reviewed the different versions of the manuscript; Sweety Samal- provided RBD reagent, reviewed the manuscript; Anil Kumar Pandey- provided samples for the study, reviewed the different versions of the manuscript; Ramesh Chandra Rai- conceptualized the study, wrote the manuscript, reviewed and edited the different versions of the manuscript, tables and figures; Amit Awasthi- conceptualized the study, and reviewed the different versions of the manuscript; All authors read and approved the final version of the manuscript.
Conflicts of Interest
Authors declare no known competing financial interests that could have influenced this work.
References
- Pollard, A.J.; Bijker, E.M. A Guide to Vaccinology: From Basic Principles to New Developments. Nat Rev Immunol 2021, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, K. The Success of SARS-CoV-2 Vaccines and Challenges Ahead. Cell Host & Microbe 2021, 29, 1111–1123. [Google Scholar] [CrossRef]
- Pušnik, J.; König, J.; Mai, K.; Richter, E.; Zorn, J.; Proksch, H.; Schulte, B.; Alter, G.; Streeck, H. Persistent Maintenance of Intermediate Memory B Cells Following SARS-CoV-2 Infection and Vaccination Recall Response. J Virol 2022, 96, e0076022. [Google Scholar] [CrossRef] [PubMed]
- Quast, I.; Tarlinton, D. B Cell Memory: Understanding COVID-19. Immunity 2021, 54, 205–210. [Google Scholar] [CrossRef]
- Kurosaki, T.; Kometani, K.; Ise, W. Memory B Cells. Nat Rev Immunol 2015, 15, 149–159. [Google Scholar] [CrossRef]
- Inoue, T.; Moran, I.; Shinnakasu, R.; Phan, T.G.; Kurosaki, T. Generation of Memory B Cells and Their Reactivation. Immunological Reviews 2018, 283, 138–149. [Google Scholar] [CrossRef]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T Cells in Cancer and Cancer Immunotherapy. Br J Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef]
- Laidlaw, B.J.; Craft, J.E.; Kaech, S.M. The Multifaceted Role of CD4(+) T Cells in CD8(+) T Cell Memory. Nat Rev Immunol 2016, 16, 102–111. [Google Scholar] [CrossRef]
- Palm, A.-K.E.; Henry, C. Remembrance of Things Past: Long-Term B Cell Memory After Infection and Vaccination. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Tarlinton, D.; Good-Jacobson, K. Diversity among Memory B Cells: Origin, Consequences, and Utility. Science 2013, 341, 1205–1211. [Google Scholar] [CrossRef]
- Mesin, L.; Ersching, J.; Victora, G.D. Germinal Center B Cell Dynamics. Immunity 2016, 45, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, M.; Obara, M.; Chiyyeadu, A.; Costa, B.; Salam, A.; Ziegler, A.; Waltl, I.; Pavlou, A.; Bonifacius, A.; Hoffmann, M.; et al. Memory B Cells Anticipate SARS-CoV-2 Variants through Somatic Hypermutation. J Infect 2024, 88, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Chua, C.-L.; Sam, I.-C.; Chiam, C.-W.; Chan, Y.-F. The Neutralizing Role of IgM during Early Chikungunya Virus Infection. PLoS ONE 2017, 12, e0171989. [Google Scholar] [CrossRef] [PubMed]
- Lizeng, Q.; Nilsson, C.; Sourial, S.; Andersson, S.; Larsen, O.; Aaby, P.; Ehnlund, M.; Björling, E. Potent Neutralizing Serum Immunoglobulin A (IgA) in Human Immunodeficiency Virus Type 2-Exposed IgG-Seronegative Individuals. J Virol 2004, 78, 7016–7022. [Google Scholar] [CrossRef]
- Hartley, G.E.; Edwards, E.S.J.; O’Hehir, R.E.; Van Zelm, M.C. New Insights into Human Immune Memory from SARS-CoV -2 Infection and Vaccination. Allergy 2022, 77, 3553–3566. [Google Scholar] [CrossRef]
- Nayak, K.; Gottimukkala, K.; Kumar, S.; Reddy, E.S.; Edara, V.V.; Kauffman, R.; Floyd, K.; Mantus, G.; Savargaonkar, D.; Goel, P.K.; et al. Characterization of Neutralizing versus Binding Antibodies and Memory B Cells in COVID-19 Recovered Individuals from India. Virology 2021, 558, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, A. Targeting Spike Protein: Modified Antibody for Broad-Spectrum Binding to Coronaviruses: An In Silico Study. VIJ 2023, 7, 1–15. [Google Scholar] [CrossRef]
- Ejemel, M.; Li, Q.; Hou, S.; Schiller, Z.A.; Tree, J.A.; Wallace, A.; Amcheslavsky, A.; Kurt Yilmaz, N.; Buttigieg, K.R.; Elmore, M.J.; et al. A Cross-Reactive Human IgA Monoclonal Antibody Blocks SARS-CoV-2 Spike-ACE2 Interaction. Nat Commun 2020, 11, 4198. [Google Scholar] [CrossRef]
- Hajilooi, M.; Keramat, F.; Moazenian, A.; Rastegari-Pouyani, M.; Solgi, G. The Quantity and Quality of Anti-SARS-CoV-2 Antibodies Show Contrariwise Association with COVID-19 Severity: Lessons Learned from IgG Avidity. Med Microbiol Immunol 2023, 212, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Vadrevu, K.M.; Ganneru, B.; Reddy, S.; Jogdand, H.; Raju, D.; Sapkal, G.; Yadav, P.; Reddy, P.; Verma, S.; Singh, C.; et al. Persistence of Immunity and Impact of Third Dose of Inactivated COVID-19 Vaccine against Emerging Variants. Sci Rep 2022, 12, 12038. [Google Scholar] [CrossRef]
- Ahmed, N.; Athavale, A.; Tripathi, A.H.; Subramaniam, A.; Upadhyay, S.K.; Pandey, A.K.; Rai, R.C.; Awasthi, A. To Be Remembered: B Cell Memory Response against SARS-CoV-2 and Its Variants in Vaccinated and Unvaccinated Individuals. Scandinavian Journal of Immunology 2024, 99, e13345. [Google Scholar] [CrossRef] [PubMed]
- Thuluva, S.; Paradkar, V.; Gunneri, S.; Yerroju, V.; Mogulla, R.; Suneetha, P.V.; Turaga, K.; Kyasani, M.; Manoharan, S.K.; Adabala, S.; et al. Immunogenicity and Safety of Biological E’s CORBEVAXTM Vaccine Compared to COVISHIELDTM (ChAdOx1 nCoV-19) Vaccine Studied in a Phase-3, Single Blind, Multicentre, Randomized Clinical Trial. Hum Vaccin Immunother 2023, 19, 2203632. [Google Scholar] [CrossRef] [PubMed]
- Tukhvatulin, A.I.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S.; Botikov, A.G.; et al. An Open, Non-Randomised, Phase 1/2 Trial on the Safety, Tolerability, and Immunogenicity of Single-Dose Vaccine “Sputnik Light” for Prevention of Coronavirus Infection in Healthy Adults. Lancet Reg Health Eur 2021, 11, 100241. [Google Scholar] [CrossRef] [PubMed]
- Russian Direct Investment Fund Sputnik Light: A Single-Component Stand-Alone Vaccine against COVID-19 and a Perfect Booster; 2021.
- Ella, R.; Vadrevu, K.M.; Jogdand, H.; Prasad, S.; Reddy, S.; Sarangi, V.; Ganneru, B.; Sapkal, G.; Yadav, P.; Abraham, P.; et al. Safety and Immunogenicity of an Inactivated SARS-CoV-2 Vaccine, BBV152: A Double-Blind, Randomised, Phase 1 Trial. The Lancet Infectious Diseases 2021, 21, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and Efficacy of the ChAdOx1 nCoV-19 Vaccine (AZD1222) against SARS-CoV-2: An Interim Analysis of Four Randomised Controlled Trials in Brazil, South Africa, and the UK. The Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatulin, A.I.; Shcheblyakov, D.V.; Dzharullaeva, A.S.; Grousova, D.M.; Erokhova, A.S.; Kovyrshina, A.V.; Botikov, A.G.; et al. Safety and Immunogenicity of an rAd26 and rAd5 Vector-Based Heterologous Prime-Boost COVID-19 Vaccine in Two Formulations: Two Open, Non-Randomised Phase 1/2 Studies from Russia. The Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Biological, E. Limited CORBEVAX Is India’s 1st Indigenously Developed Protein Sub-Unit COVID-19 Vaccine 2021.
- Krishna, B.; Gupta, A.; Meena, K.; Gaba, A.; Krishna, S.; Jyoti, R.; Aeron, N.; Prashanth, S.; Samriti, null; Ganapathy, U. Prevalence, Severity, and Risk Factor of Breakthrough Infection after Vaccination with Either the Covaxin or the Covishield among Healthcare Workers: A Nationwide Cross-Sectional Study. J Anaesthesiol Clin Pharmacol 2022, 38, S66–S78. [Google Scholar] [CrossRef]
- Malhotra, A. Curing the Pandemic of Misinformation on COVID-19 mRNA Vaccines through Real Evidence-Based Medicine - Part 2. Journal of Insulin Resistance 2022, 5, 72. [Google Scholar] [CrossRef]
- Vallejo, A.; Vizcarra, P.; Martín-Hondarza, A.; Gómez-Maldonado, S.; Haemmerle, J.; Velasco, H.; Casado, J.L. Impact of SARS-CoV-2-Specific Memory B Cells on the Immune Response after mRNA-Based Comirnaty Vaccine in Seronegative Health Care Workers. Front Microbiol 2022, 13, 1002748. [Google Scholar] [CrossRef]
- Cox, R.J.; Brokstad, K.A. Not Just Antibodies: B Cells and T Cells Mediate Immunity to COVID-19. Nat Rev Immunol 2020, 20, 581–582. [Google Scholar] [CrossRef]
- Kudriavtsev, A.V.; Vakhrusheva, A.V.; Novoseletsky, V.N.; Bozdaganyan, M.E.; Shaitan, K.V.; Kirpichnikov, M.P.; Sokolova, O.S. Immune Escape Associated with RBD Omicron Mutations and SARS-CoV-2 Evolution Dynamics. Viruses 2022, 14, 1603. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S.; Aubert, R.D.; Glidewell, J.; Ahmed, R. Tracking Human Antigen-Specific Memory B Cells: A Sensitive and Generalized ELISPOT System. Journal of Immunological Methods 2004, 286, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Barnes, C.O.; Jette, C.A.; Abernathy, M.E.; Dam, K.-M.A.; Esswein, S.R.; Gristick, H.B.; Malyutin, A.G.; Sharaf, N.G.; Huey-Tubman, K.E.; Lee, Y.E.; et al. SARS-CoV-2 Neutralizing Antibody Structures Inform Therapeutic Strategies. Nature 2020, 588, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 Spike Receptor-Binding Domain Bound to the ACE2 Receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Wang, Q.; Du, Q.; Guo, B.; Mu, D.; Lu, X.; Ma, Q.; Guo, Y.; Fang, L.; Zhang, B.; Zhang, G.; et al. A Method To Prevent SARS-CoV-2 IgM False Positives in Gold Immunochromatography and Enzyme-Linked Immunosorbent Assays. J Clin Microbiol 2020, 58, e00375–20. [Google Scholar] [CrossRef]
- Thuluva, S.; Paradkar, V.; Gunneri, S.R.; Yerroju, V.; Mogulla, R.; Turaga, K.; Kyasani, M.; Manoharan, S.K.; Medigeshi, G.; Singh, J.; et al. Evaluation of Safety and Immunogenicity of Receptor-Binding Domain-Based COVID-19 Vaccine (Corbevax) to Select the Optimum Formulation in Open-Label, Multicentre, and Randomised Phase-1/2 and Phase-2 Clinical Trials. eBioMedicine 2022, 83, 104217. [Google Scholar] [CrossRef]
- Mehdi, F.; Chattopadhyay, S.; Thiruvengadam, R.; Yadav, S.; Kumar, M.; Sinha, S.K.; Goswami, S.; Kshetrapal, P.; Wadhwa, N.; Chandramouli Natchu, U.; et al. Development of a Fast SARS-CoV-2 IgG ELISA, Based on Receptor-Binding Domain, and Its Comparative Evaluation Using Temporally Segregated Samples From RT-PCR Positive Individuals. Front Microbiol 2020, 11, 618097. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Khatri, R.; Parray, H.A.; Siddiqui, G.; Chiranjivi, A.K.; Raj, S.; Kaul, R.; Maithil, V.; Samal, S.; Ahmed, S. Biophysical and Biochemical Characterization of the Receptor Binding Domain of SARS-CoV-2 Variants. Protein J 2022, 41, 457–467. [Google Scholar] [CrossRef]
- Madhavan, R.; Paul, J.S.; Babji, S.; Thamizh, I.; Kumar, D.; Khakha, S.A.; Rennie, A.; Kumar, K.; Dhanapal, P.; Saravanan, P.; et al. SARS-CoV-2 Infections before, during, and after the Omicron Wave: A 2-Year Indian Community Cohort Study. The Lancet Regional Health - Southeast Asia 2024, 28, 100470. [Google Scholar] [CrossRef]
- Skountzou, I.; Satyabhama, L.; Stavropoulou, A.; Ashraf, Z.; Esser, E.S.; Vassilieva, E.; Koutsonanos, D.; Compans, R.; Jacob, J. Influenza Virus-Specific Neutralizing IgM Antibodies Persist for a Lifetime. Clin Vaccine Immunol 2014, 21, 1481–1489. [Google Scholar] [CrossRef]
- Takemori, T.; Kaji, T.; Takahashi, Y.; Shimoda, M.; Rajewsky, K. Generation of Memory B Cells inside and Outside Germinal Centers. European Journal of Immunology 2014, 44, 1258–1264. [Google Scholar] [CrossRef] [PubMed]
- Shlomchik, M.J.; Weisel, F. Germinal Center Selection and the Development of Memory B and Plasma Cells. Immunological Reviews 2012, 247, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.L.; Essigmann, H.T. Original Antigenic Sin: The Downside of Immunological Memory and Implications for COVID-19. mSphere 2021, 6, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Sasikala, M.; Shashidhar, J.; Deepika, G.; Ravikanth, V.; Krishna, V.V.; Sadhana, Y.; Pragathi, K.; Reddy, D.N. Immunological Memory and Neutralizing Activity to a Single Dose of COVID-19 Vaccine in Previously Infected Individuals. International Journal of Infectious Diseases 2021, 108, 183–186. [Google Scholar] [CrossRef]
- Yamayoshi, S.; Yasuhara, A.; Ito, M.; Akasaka, O.; Nakamura, M.; Nakachi, I.; Koga, M.; Mitamura, K.; Yagi, K.; Maeda, K.; et al. Antibody Titers against SARS-CoV-2 Decline, but Do Not Disappear for Several Months. EClinicalMedicine 2021, 32, 100734. [Google Scholar] [CrossRef]
- Jo, D.-H.; Minn, D.; Lim, J.; Lee, K.-D.; Kang, Y.-M.; Choe, K.-W.; Kim, K.-N. Rapidly Declining SARS-CoV-2 Antibody Titers within 4 Months after BNT162b2 Vaccination. Vaccines 2021, 9, 1145. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, H.R.; Parai, D.; Chandra Dash, G.; Kshatri, J.S.; Mishra, N.; Choudhary, P.K.; Pattnaik, D.; Panigrahi, K.; Behera, S.; Ranjan Sahoo, N.; et al. Persistence of Antibodies Against Spike Glycoprotein of SARS-CoV-2 in Healthcare Workers Post Double Dose of BBV-152 and AZD1222 Vaccines. Front Med (Lausanne) 2021, 8, 778129. [Google Scholar] [CrossRef]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. mRNA Vaccines Induce Durable Immune Memory to SARS-CoV-2 and Variants of Concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef]
- Kaku, C.I.; Bergeron, A.J.; Ahlm, C.; Normark, J.; Sakharkar, M.; Forsell, M.N.E.; Walker, L.M. Recall of Preexisting Cross-Reactive B Cell Memory after Omicron BA.1 Breakthrough Infection. Sci Immunol 2022, 7, eabq3511. [Google Scholar] [CrossRef]
- Wang, Z.; Muecksch, F.; Raspe, R.; Johannsen, F.; Turroja, M.; Canis, M.; ElTanbouly, M.A.; Santos, G.S.S.; Johnson, B.; Baharani, V.A.; et al. Memory B Cell Development Elicited by mRNA Booster Vaccinations in the Elderly. J Exp Med 2023, 220, e20230668. [Google Scholar] [CrossRef] [PubMed]
- Morales-Núñez, J.J.; García-Chagollán, M.; Muñoz-Valle, J.F.; Díaz-Pérez, S.A.; Torres-Hernández, P.C.; Rodríguez-Reyes, S.C.; Santoscoy-Ascencio, G.; Sierra García de Quevedo, J.J.; Hernández-Bello, J. Differences in B-Cell Immunophenotypes and Neutralizing Antibodies Against SARS-CoV-2 After Administration of BNT162b2 (Pfizer-BioNTech) Vaccine in Individuals with and without Prior COVID-19 - A Prospective Cohort Study. J Inflamm Res 2022, 15, 4449–4466. [Google Scholar] [CrossRef] [PubMed]
- Terreri, S.; Piano Mortari, E.; Vinci, M.R.; Russo, C.; Alteri, C.; Albano, C.; Colavita, F.; Gramigna, G.; Agrati, C.; Linardos, G.; et al. Persistent B Cell Memory after SARS-CoV-2 Vaccination Is Functional during Breakthrough Infections. Cell Host Microbe 2022, 30, 400–408. [Google Scholar] [CrossRef]
- Geropeppa, M.; Papadatou, I.; Sarantis, P.; Tzanoudaki, M.; Ntanasis-Stathopoulos, I.; Bagratuni, T.; Terpos, E.; Spoulou, V. Receptor-Binding-Domain-Specific B Cell Responses Induced by mRNA Immunization against SARS-CoV-2. Vaccines 2023, 11, 1148. [Google Scholar] [CrossRef] [PubMed]
- Busà, R.; Miele, M.; Sorrentino, M.C.; Amico, G.; Timoneri, F.; Miceli, V.; Di Bella, M.; Russelli, G.; Gallo, A.; Zito, G.; et al. Long-Term Effectiveness of BNT162b2 Pfizer-BioNTech mRNA-Based Vaccine on B Cell Compartment: Efficient Recall of SARS-CoV-2-Specific Memory B Cells. Int J Mol Sci 2022, 23, 15046. [Google Scholar] [CrossRef]
- Mise-Omata, S.; Ikeda, M.; Takeshita, M.; Uwamino, Y.; Wakui, M.; Arai, T.; Yoshifuji, A.; Murano, K.; Siomi, H.; Nakagawara, K.; et al. Memory B Cells and Memory T Cells Induced by SARS-CoV-2 Booster Vaccination or Infection Show Different Dynamics and Responsiveness to the Omicron Variant. The Journal of Immunology 2022, 209, 2104–2113. [Google Scholar] [CrossRef]
- Brewer, R.C.; Ramadoss, N.S.; Lahey, L.J.; Jahanbani, S.; Robinson, W.H.; Lanz, T.V. BNT162b2 Vaccine Induces Divergent B Cell Responses to SARS-CoV-2 S1 and S2. Nat Immunol 2022, 23, 33–39. [Google Scholar] [CrossRef]
- Bednarski, E.; Del Rio Estrada, P.M.; DaSilva, J.; Boukadida, C.; Zhang, F.; Luna-Villalobos, Y.A.; Rodríguez-Rangel, X.; Pitén-Isidro, E.; Luna-García, E.; Díaz Rivera, D.; et al. Antibody and Memory B-Cell Immunity in a Heterogeneously SARS-CoV-2-Infected and -Vaccinated Population. mBio 13, e00840-22. [CrossRef]
- Zhang, Z.; Mateus, J.; Coelho, C.H.; Dan, J.M.; Moderbacher, C.R.; Gálvez, R.I.; Cortes, F.H.; Grifoni, A.; Tarke, A.; Chang, J.; et al. Humoral and Cellular Immune Memory to Four COVID-19 Vaccines. Cell 2022, 185, 2434–2451.e17. [Google Scholar] [CrossRef]
- Fryer, H.A.; Hartley, G.E.; Edwards, E.S.J.; O’Hehir, R.E.; van Zelm, M.C. Humoral Immunity and B-Cell Memory in Response to SARS-CoV-2 Infection and Vaccination. Biochem Soc Trans 2022, 50, 1643–1658. [Google Scholar] [CrossRef]
Figure 1.
IgG antibody levels against SARS-CoV-2 receptor binding domain (RBD) of Spike protein among unvaccinated and vaccinated participants.
Figure 1.
IgG antibody levels against SARS-CoV-2 receptor binding domain (RBD) of Spike protein among unvaccinated and vaccinated participants.
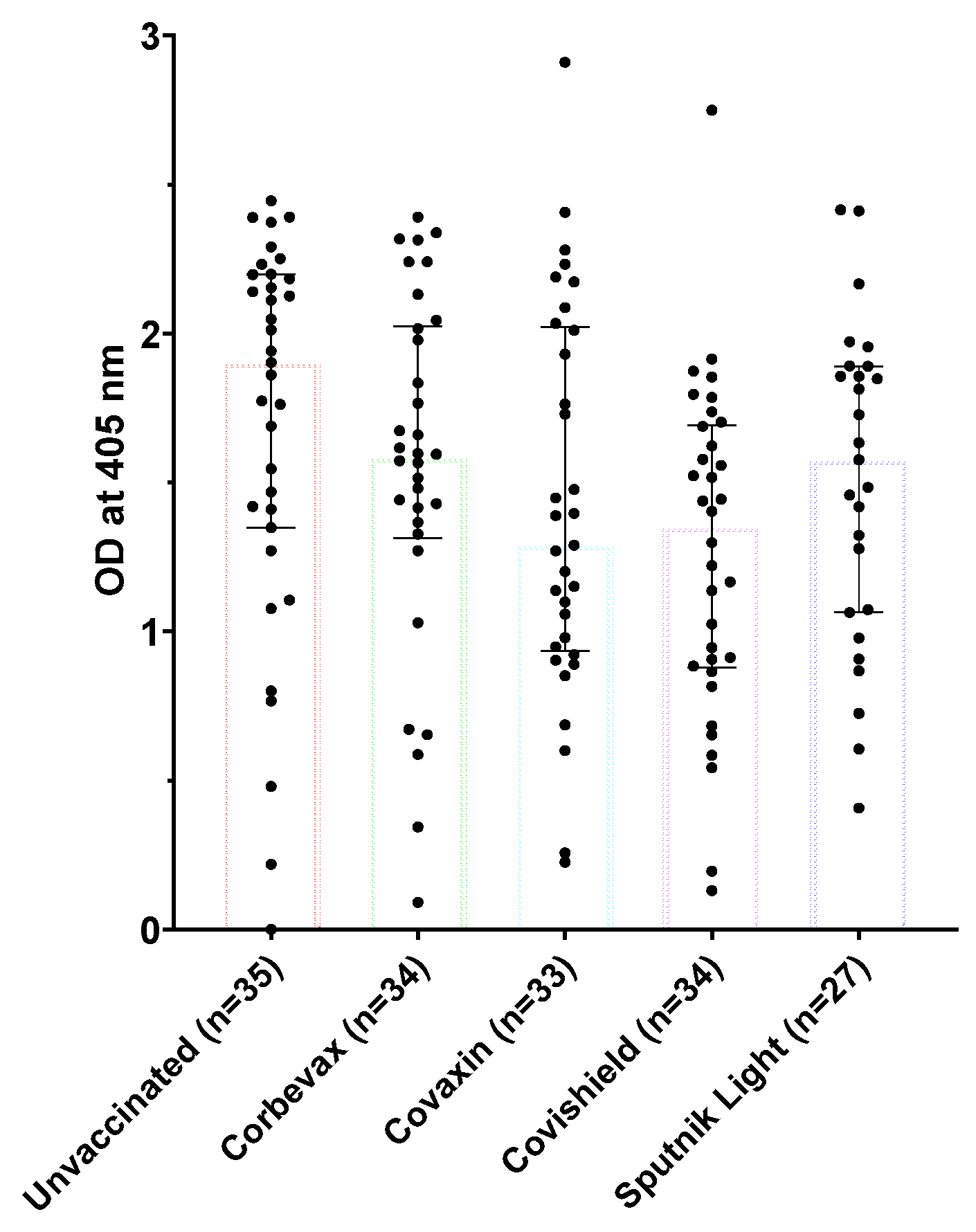
Figure 2.
(A) Representative flow cytometry gating strategy for SARS-CoV-2 RBD-specific memory B cells. (B) Graphical representation comparing the percentage of RBD specific memory B cells in vaccinated and unvaccinated participants [Outliers removed using iterative Grubb’s test, alpha = 0.01].
Figure 2.
(A) Representative flow cytometry gating strategy for SARS-CoV-2 RBD-specific memory B cells. (B) Graphical representation comparing the percentage of RBD specific memory B cells in vaccinated and unvaccinated participants [Outliers removed using iterative Grubb’s test, alpha = 0.01].
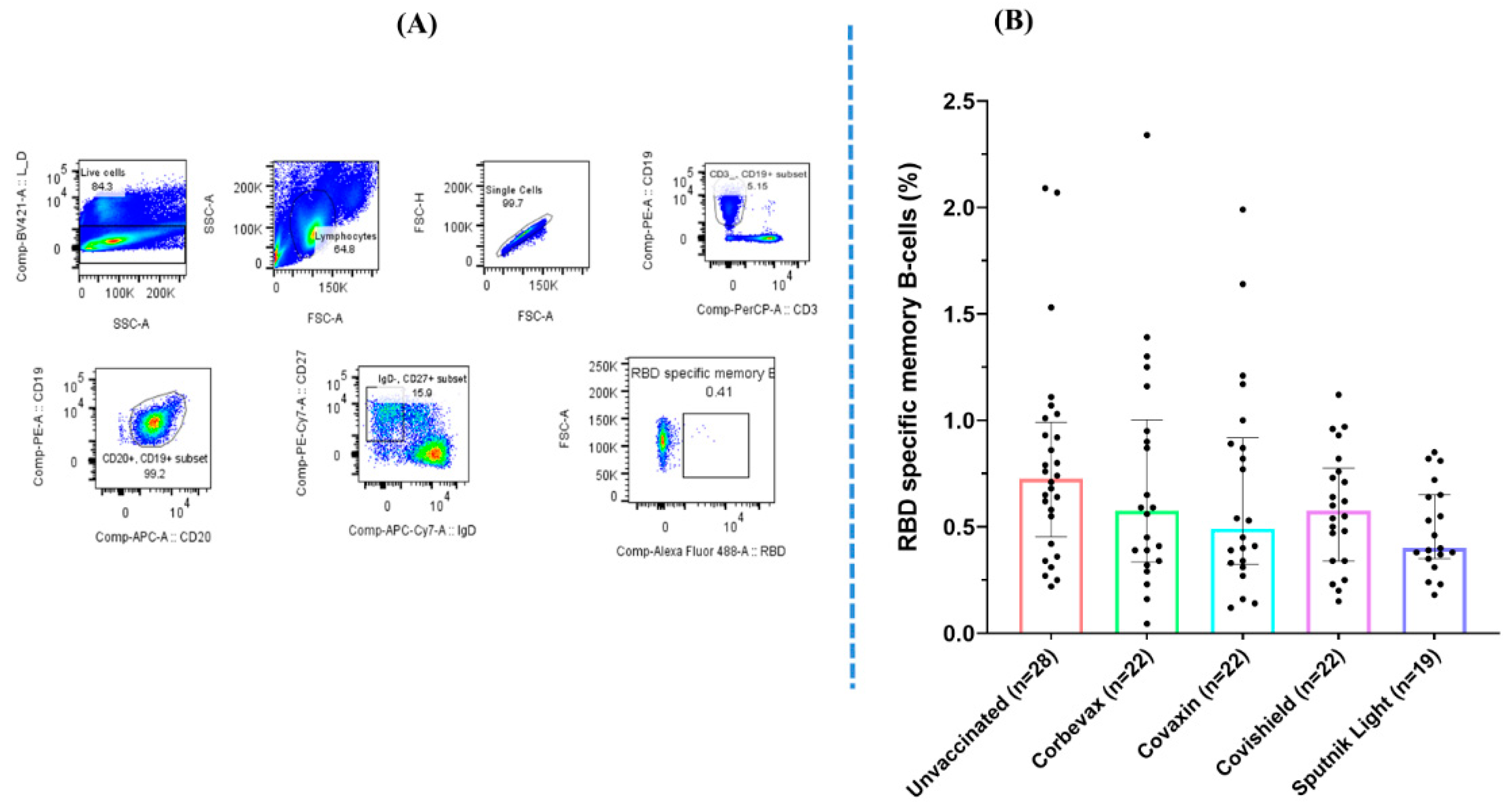
Figure 3.
(A) Schematic ELISPOT assay plate map (created using BioRender). (B) Representative plate image showing the total and RBD-specific antibody secreting cells (ASCs) among the cells collected from an unvaccinated and a vaccinated individual.
Figure 3.
(A) Schematic ELISPOT assay plate map (created using BioRender). (B) Representative plate image showing the total and RBD-specific antibody secreting cells (ASCs) among the cells collected from an unvaccinated and a vaccinated individual.
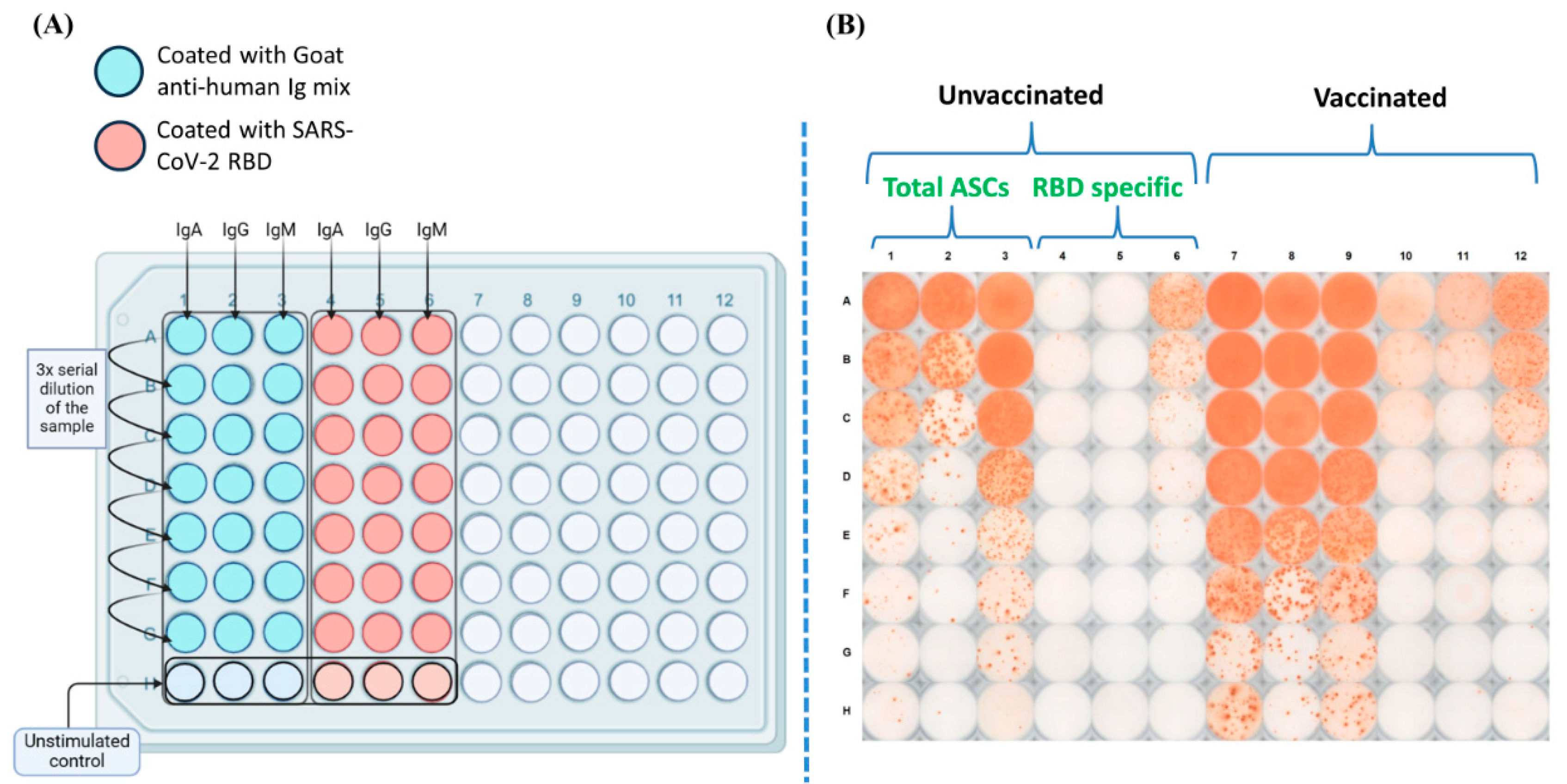
Figure 4.
Fraction of the RBD specific ASC (IgA, IgG and IgM) among vaccinated versus unvaccinated individuals. [Outliers removed using iterative Grubb’s test, alpha = 0.01]
Figure 4.
Fraction of the RBD specific ASC (IgA, IgG and IgM) among vaccinated versus unvaccinated individuals. [Outliers removed using iterative Grubb’s test, alpha = 0.01]
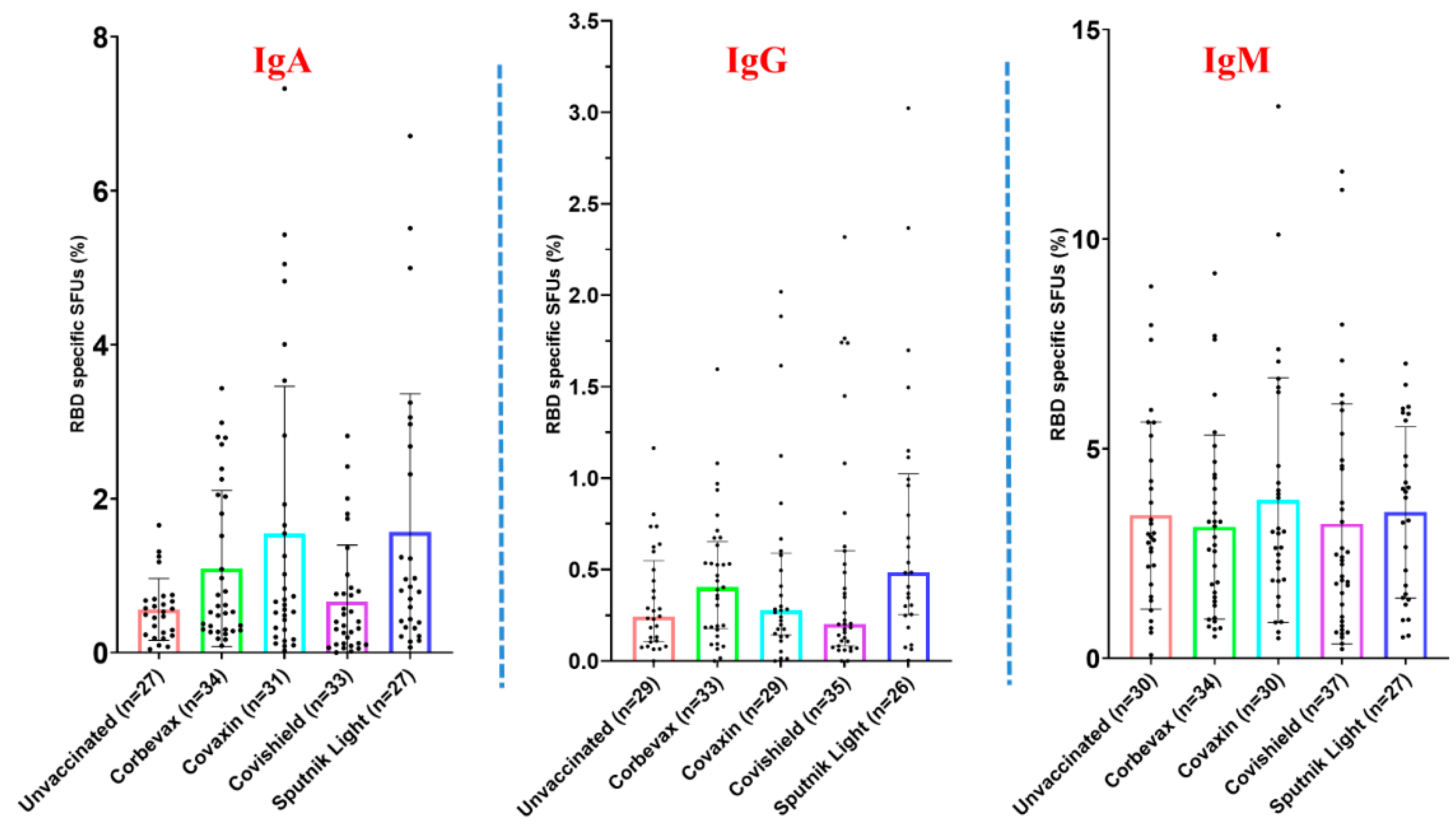
Figure 5.
Fraction of the RBD specific ASC (IgA, IgG and IgM) to depict temporal changes (for participants vaccinated more than 6 months prior to sample collection): A. among Corbevax vaccinated versus unvaccinated individuals. B. among Covaxin vaccinated versus unvaccinated individuals. C. among Covishield vaccinated versus unvaccinated individuals. D. among Sputnik Light vaccinated versus unvaccinated individuals. [Mann Whitney U test; *p < 0.05, **p < 0.01; outliers were removed using iterative Grubb’s test, alpha = 0.01].
Figure 5.
Fraction of the RBD specific ASC (IgA, IgG and IgM) to depict temporal changes (for participants vaccinated more than 6 months prior to sample collection): A. among Corbevax vaccinated versus unvaccinated individuals. B. among Covaxin vaccinated versus unvaccinated individuals. C. among Covishield vaccinated versus unvaccinated individuals. D. among Sputnik Light vaccinated versus unvaccinated individuals. [Mann Whitney U test; *p < 0.05, **p < 0.01; outliers were removed using iterative Grubb’s test, alpha = 0.01].
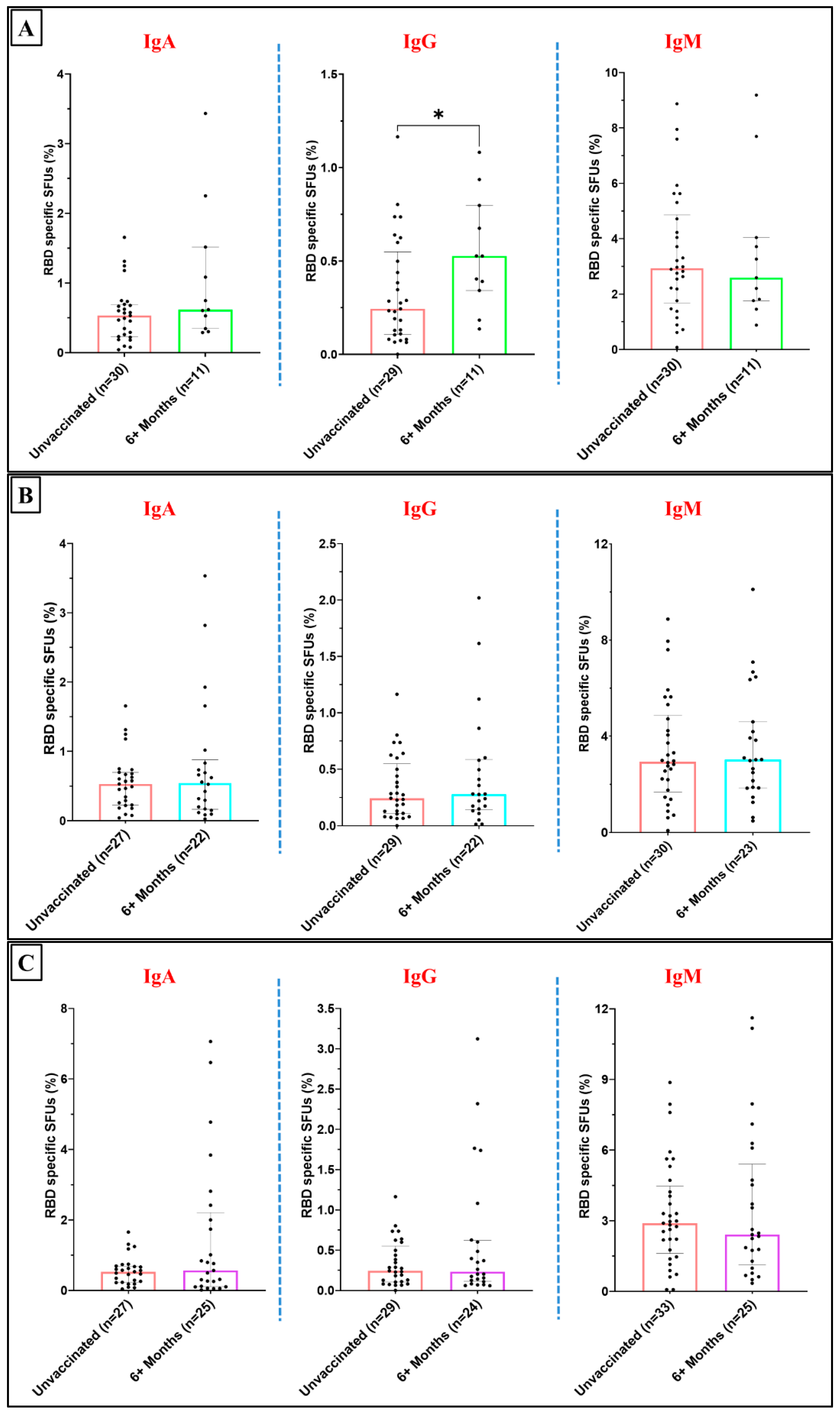
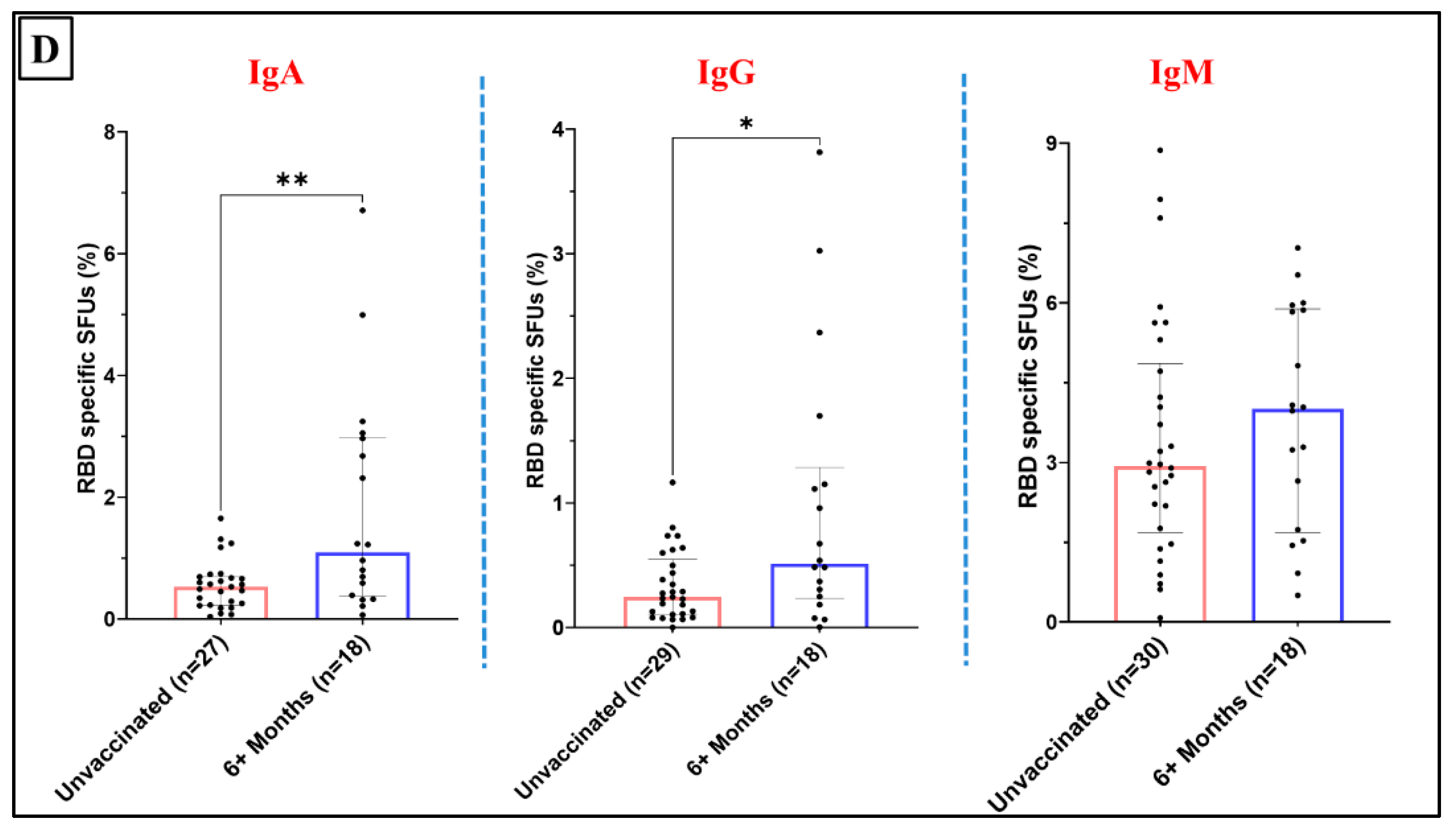
Figure 6.
Gender-disaggregated analysis of immune response and memory B cell response. 6A- IgG antibody levels against SARS-CoV-2 spike RBD compared between male and female participants. 6B. Percentage of RBD specific memory B cells in the peripheral circulation of male and female participants segregated based on the vaccines given. 6C. Percentage of RBD-specific IgA (i), IgG (ii) and IgM (iii) spot forming units in unvaccinated and vaccinated males and females.
Figure 6.
Gender-disaggregated analysis of immune response and memory B cell response. 6A- IgG antibody levels against SARS-CoV-2 spike RBD compared between male and female participants. 6B. Percentage of RBD specific memory B cells in the peripheral circulation of male and female participants segregated based on the vaccines given. 6C. Percentage of RBD-specific IgA (i), IgG (ii) and IgM (iii) spot forming units in unvaccinated and vaccinated males and females.
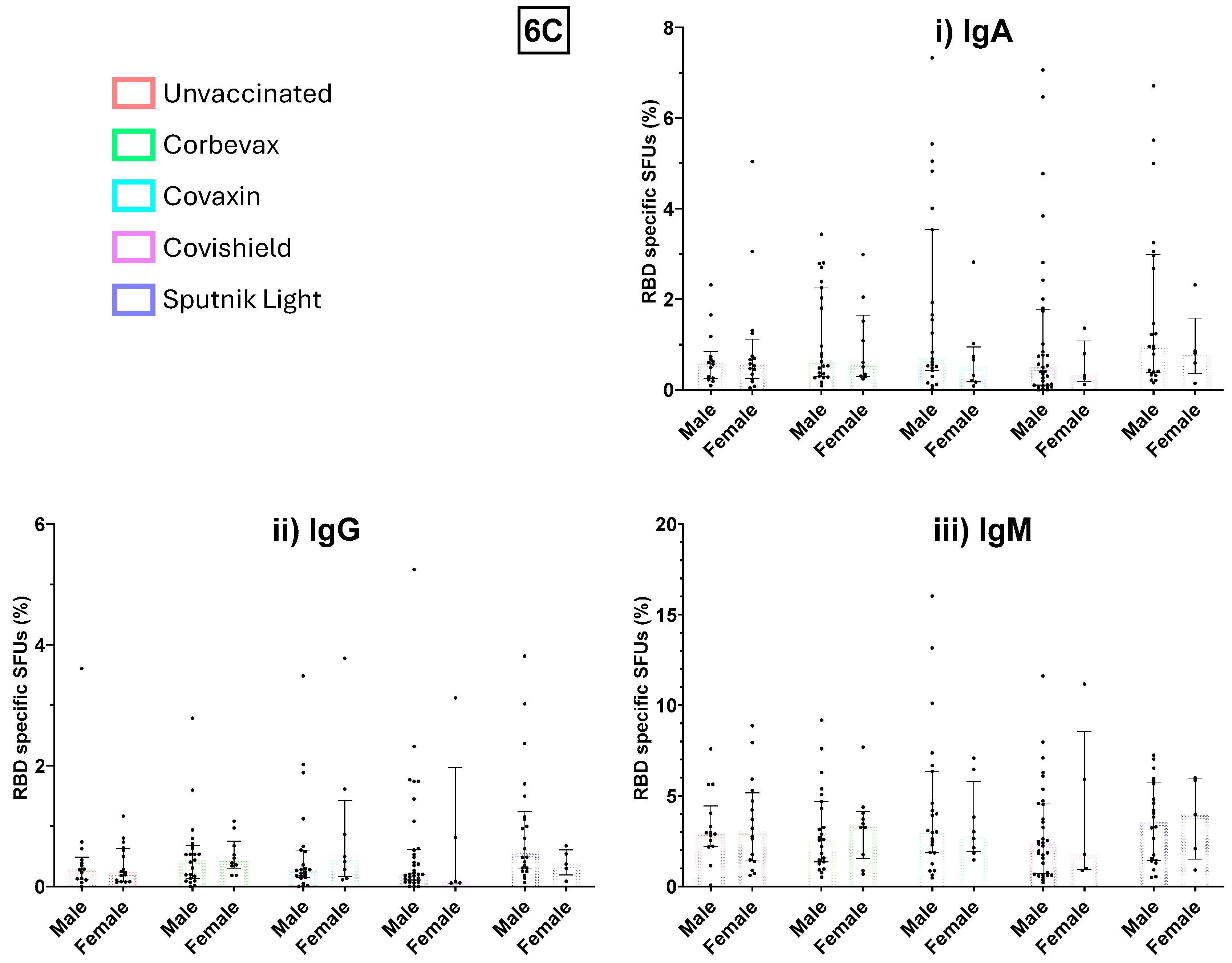

Table 1.
Participant details (A) and their demography (B).
| (A) | |||||||
| Sr. No. | Status of vaccination | Number | |||||
| 1 | Unvaccinated | 35 | |||||
| 2 | Vaccinated (Corbevax) | 34 | |||||
| 3 | Vaccinated (Covaxin) | 33 | |||||
| 4 | Vaccinated (Covishield) | 40 | |||||
| 5 | Vaccinated (Sputnik Light) | 29 | |||||
| (B) | |||||||
| Unvaccinated | Corbevax | Covishield | Covaxin | Sputnik Light | |||
| Sex | Male | 17 | 24 | 33 | 24 | 25 | |
| Female | 18 | 10 | 7 | 9 | 4 | ||
| Age | Range | 37 (18-55) | 26 (20-46) | 34 (19-53) | 33 (21-54) | 39 (20-59) | |
| Median | 28 | 29 | 26 | 26 | 30 | ||
| Months since vaccination | Range | - | 5 (3-8) | 12 (3-15) | 12 (3-15) | 5 (6-11) | |
| Median | - | 6 | 8 | 11 | 8 | ||
Table 2.
Molecular markers used for the memory B cell estimation in the peripheral circulation of the study participants.
Table 2.
Molecular markers used for the memory B cell estimation in the peripheral circulation of the study participants.
| SN. | Molecular marker | Fluorophore | Make and Catalogue number |
|---|---|---|---|
| 1 | Fixable viability dye | Violet Dye | Invitrogen #L34964 |
| 2 | CD3 | PerCP | BioLegend #344814 |
| 3 | CD19 | PE | BioLegend #302208 |
| 4 | CD20 | APC | BioLegend #302310 |
| 5 | CD27 | PE-Cy7 | BioLegend #356412 |
| 6 | IgD | APC-Cy7 | BioLegend #348218 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



