
Submitted:
05 August 2024
Posted:
06 August 2024
You are already at the latest version
Abstract
The combination of several therapeutic strategies is often seen as a good way to decrease the resistance rates, since bacteria overcome better single-drug treatments than multi-drug ones. This strategy is especially attractive when several targets and subpopulations are affected, as it is the case of Klebsiella pneumoniae persister cells, a subpopulation of bacteria able to transiently survive antibiotic exposures. This work aims to evaluate the potential of a repurposed anticancer drug, mitomycin C, combined with the K. pneumoniae lytic phage vB_KpnM-VAC13 in vitro and the safety in an in vivo murine model against two clinical isolates of this pathogen, one of them exhibiting an imipenem-persister phenotype. At the same time, we verified the absence of toxicity of mitomycin C at the concentration using the human chondrocyte cell line T/C28a2. The viability of these human cells was checked both by cytotoxicity assays and flow cytometry.
Keywords:
persistence
; K. pneumoniae
; repurposing
; mitomycin C
; lytic phage
; cytotoxicity
1. Introduction
Infections caused by Klebsiella pneumoniae represent a rising concern in both hospital settings and the community, most of them being caused by multidrug-resistant (MDR) strains and difficult-to-treat isolates [1]. Many recent outbreaks have evidenced the need for effective treatments against antibiotic-resistant isolates, especially the carbapenemase-producing strains of K. pneumoniae (CRKp) [2,3,4,5].
Frequently underestimated, although clinically relevant, are the antibiotic-persister strains, a subpopulation of bacterial cells that enter into a dormant and latent metabolic state in which they are able to transiently survive antibiotic exposure while exhibiting sensitive minimal inhibitory concentration (MIC) values, that is below the breakpoints established for a specific antibiotic [6,7]. The persister subpopulation exits the metabolically dormant stage once the antibiotic pressure is removed, and is therefore responsible for recalcitrant infections, chronification of these and usually associated with biofilms [8,9]. In the clinical setting, persisters can survive in immunocompromised patients and in those in whom antibiotics did not effectively kill pathogenic bacteria, due to immune-evasion strategies. Examples of these are Pseudomonas aeruginosa and Salmonella enterica surviving in macrophages, together with uropathogenic strains of Escherichia coli [10,11,12,13]. Moreover, they can act as reservoirs for resistant mutants [14]. In vitro, persisters are characterized by a biphasic killing curve in the presence of the antibiotic, since the main susceptible population is rapidly killed, whereas the persister subpopulation has a slower killing time [6].
In this context, innovative strategies such as the repurposing of drugs that have been approved by regulatory agencies with other therapeutic indications rather than antibacterial, together with the use of lytic bacteriophages (or phages), the natural predators of bacteria that specifically infect and kill their hosts, are of special interest [15]. The use of bacteriophages to treat bacterial infections in humans started a century ago and many articles in the scientific literature report their therapeutic successful outcomes [16,17].
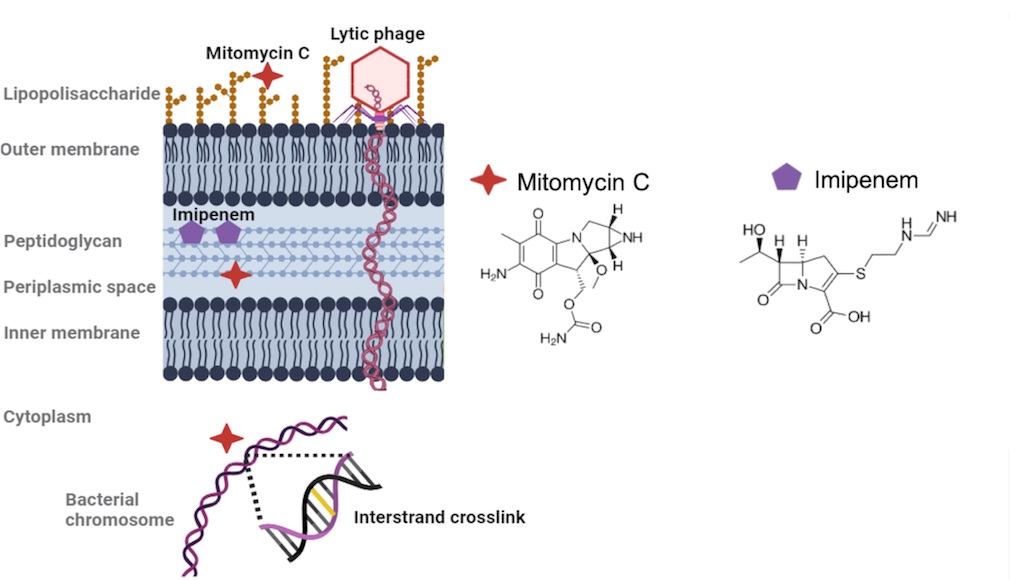
Concerning the repurposing strategy, mitomycin C (MMC), naturally produced by Streptomyces caespitosus, is an FDA-approved anticancer molecule used for the treatment of bladder, gastric, lung and pancreatic cancer, among others [18,19]. In bacteria, MMC passively enters the bacterial cell and gets reduced inside the cytoplasm, which provokes its activation. MMC is an alkylating agent that covalently binds to DNA and induces interstrand crosslinking reactions between guanine adjacent bases in the minor groove of the double helix (5’-GC). These crosslinking reactions are resolved by the UvrABC system in cells in which this mechanism is undamaged and works properly. However, this is not the case for persister cells, where MMC has been proven to kill the subpopulation of these dormant cells in many different pathogens, such as in Acinetobacter baumannii, E. coli and K. pneumoniae [20,21]. As a chemotherapeutic agent, MMC is usually intravenously administered at concentrations ranging from 0.5 to 2 μg/ml, but concerns evaluating the toxicity of this molecule have arisen [22].
Repurposing of this compound as an antibiotic against K. pneumoniae infections, following in vitro and in vivo approach by Galleria meillonella model, was already performed by our research group [23]. In that work, subinhibitory concentrations of mitomycin C and the conventional carbapenem imipenem were combined with the lytic bacteriophage vB_KpnM-VAC13, and tested against i) an imipenem-resistant K. pneumoniae isolate (MICIMP=8 μg/mL), harboring the gene blaOXA-245; and ii) an imipenem-persister isolate of this pathogen (MICIMP=0.5 μg/mL). Based on previous reports that link the lytic phage infection with the entrance into the persister stage [24,25], this work aims to evaluate the safety of anti-persister MMC combined with the phage vB_KpnM-VAC13 in the in vivo murine model to broaden the applicability of this combinatorial approach. At the same time, we verified the absence of toxicity of MMC, at the concentration used against K. pneumoniae, using the human chondrocyte cell line T/C28a2 by cytotoxicity assays and flow cytometry.
2. Results
2.1. Synergy Studies
2.1.1. Optical Density Growth Curves and Viability Assay
Different outcomes in terms of growth were assessed for the three strains tested. In what concerns the imipenem-resistant isolate K2534 (ST437-OXA245), MMC at 3 μg/mL did not affect its growth for the first 8 hours, exerting a slight activity from this time point onwards. The phage vB_KpnM-VAC13 completely inhibited the growth of K2534 for the first 8 hpi, and then a regrowth of a phage-resistant population appeared. In the case of this strain, no synergy was obtained for the combination between MMC and vB_KpnM-VAC13, although a slight delay in the arising of phage-resistant mutants is observed (Figure 1a).
Regarding now the imipenem-persister strain K3325, an inverted effect in terms of susceptibility to MMC and to the vB_KpnM-VAC13 compared to K2534 was obtained: the MMC exhibited a much stronger bactericidal effect than in the OXA245-harbouring isolate (Figure 1b). As already described in a previous work [23], the phage vB_KpnM-VAC13 did not exhibit a very strong lytic activity and the culture regrew at 4 hours post-infection. Importantly, the combination between MMC and vB_KpnM-VAC13 effectively killed the whole bacterial population, exhibiting in this case a strong synergism (Figure 1b).
Finally, for the reference strain ATCC®10031TM, the lytic phage exhibited a strong lytic effect for the first 10 hours, but a regrowth was visible from this timepoint onwards. A complete arrest of the bacterial growth was obtained in the case of MMC alone and the combination of this compound with the lytic phage (Figure 1c).
In terms of viability, the enumeration of the colony-forming units (CFU) consistently reflected the results obtained with the spectrophotometer: non-statistically significant differences were obtained at 24h for K2534 among the different conditions evaluated (Figure 1d), whereas for K3325 a synergistic effect was achieved in the presence of both MMC and the lytic phage, resulting in a 6-log decrease compared to the control counts. Interestingly, MMC alone only decreased the bacterial counts in 2-log, while the phage produced a 1-log reduction by itself, being the effect of the combination greater than the sum of the individual activities (Figure 1e). Finally, the ATCC®10031TM strain revealed a drastic decrease in the CFU enumerated after exposure to MMC alone and combined with vB_KpnM-VAC13, whereas the phage alone did not produce a statistically significant reduction of the bacterial counts at 24h compared to the control, highly likely due to the arising of phage-resistance.
2.1.2. Frequency of Phage-Resistant Mutants
As previously evoked, the arising of phage-resistant mutants to the combined treatment herein evaluated (MMC and the lytic phage vB_KpnM-VAC13), might compromise the outcome of this approach. Therefore, we assessed the frequency of these mutants not only to the phage alone and combined with MMC, but also to the MMC by itself at the concentration used in the rest of experiments, 3 mg/L.
We observed diverse patterns of resistance to the lytic phage exhibited by the different strains used, ranging from 9.9x10-3 ± 3.6x10-3 for K2534, 7.3Ex10-1 for K3325 (already assessed in our previous work [23]) and 4.1x10-5 ± 1.5x10-5 in the case of the reference strain ATCC®10031TM (Figure 2). In the case of the resistance showed in the presence MMC at 3 mg/L, we obtained an expected high frequency of resistance in the case of K2534 (1.2 ± 0.2), 7.3x10-5 for K3325 and no viable colonies growing for the reference strain, thus a frequency of resistance to MMC of nearly zero. For K2534, the frequency of resistance to the combined MMC and the phage was not statistically significant compared to the phage-resistance alone, whereas it was statistically different comparing the frequency of MMC alone and in combination with the phage (p-value = 0.015). In the case of K3325 the frequency of resistance decreased to nearly zero in the presence of the combination, being statistically significant compared to the phage-resistance to the vB_KpnM-VAC13 alone (p-value = 0.0035). Finally, the reference strain exhibited undifferentiated values of resistance to the MMC alone or combined with the phage (Figure 2).
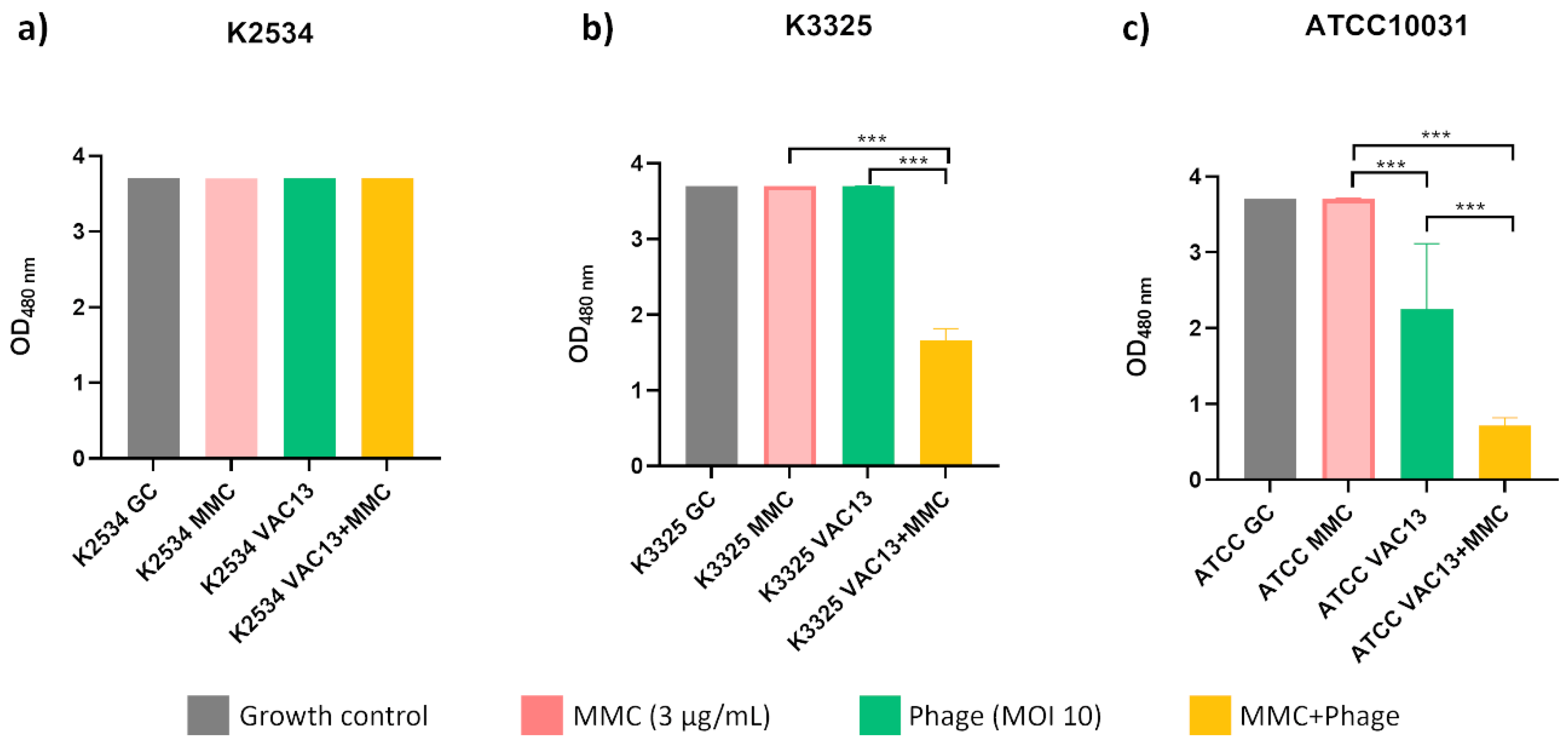
2.1.3. Assessment of the Cellular Respiration In Vitro
The cellular viability in the presence of MMC for every strain tested was comparable to that of the growth control, despite the differences in the growth patterns shown in the growth curves. Besides, this assay indicated that the lytic phage vB_KpnM-VAC13 at MOI of 10 allowed a normal metabolic activity for the two clinical strains (a and b), different from the effect observed in the reference strain (c), in which it significantly reduced the metabolic activity (p < 0.001). For this strain but, interestingly, also for the persister isolate K3325, combinations of phage and MMC were able to significantly reduce the metabolic activity of bacteria compared to the agents separately.
Figure 3.
Respiration assay of K. pneumoniae clinical strains K2534 (A), K3325 (B) and the reference strain ATCC®10031TM (C), using the tetrazolium salt WST-1 (an indicator of NADP+ reduction). *** represents p-values lower than 0.001.
Figure 3.
Respiration assay of K. pneumoniae clinical strains K2534 (A), K3325 (B) and the reference strain ATCC®10031TM (C), using the tetrazolium salt WST-1 (an indicator of NADP+ reduction). *** represents p-values lower than 0.001.
2.2. Toxicity Studies
2.2.1. In Vitro: Metabolic Activity and Apoptosis Study
Next, we performed a cellular cytotoxic assay using the MTT agent (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide), based on a colorimetric reduction of the tetrazolium salt to purple formazan crystals. This reaction allows the measurement of metabolically active cells, as an indicator of cellular proliferation and metabolism. We visually assessed the conversion of MTT to purple formazan after addition of DMSO (Figure 4a) and calculated the percentage of viability of T/C28a2 cells under the concentrations evaluated (Figure 4b). Low percentages of viability (around 35%) were obtained for 20 and 10 µg/mL MMC, showing non-statistically significant differences between these two concentrations, whereas MMC at 8 µg/mL led to a viability of 43% on average, which represented a statistically significant decrease compared to the viability at MMC=6 µg/mL (around 55%). Interestingly, at 3 µg/mL (the concentration used for the other experiments here reported) a statistically significant increase was observed compared to the viability at a concentration of 6 µg/mL, reaching a 75% (Figure 4b).
These results were confirmed by flow cytometry, which revealed a statistically significant increase in the percentage of live cells (PI and annexin-FITC negative) treated with 3 µg/mL of MMC compared to 10 µg/mL, and a significant decrease in the percentage of apoptotic cells comparing these two concentrations (p<0.001, Figure 4c). Importantly, the percentage of apoptotic cells was not statistically different comparing absence of MMC and MMC at 3 µg/mL. Finally, no difference in the percentage of necrotic cells was observed among the different concentrations of MMC tested.
Representative graphs of each condition (one technical replicate) obtained with the software CytExpert 2.0 are shown in Figure 4d.
2.2.2. In vivo toxicity assay: murine model
Acute toxicity was assessed in a murine model after intraperitoneal injection of a single dose of the following MMC concentrations: 2.4, 1.7 and 0.52 mg/kg, and the lytic bacteriophage vB_KpnM-VAC13 at MOIs of 0.50, 1 and 10. No systemic signs of pain or weight loss were observed.
Cumulative toxicity was assessed with the highest concentrations of MMC and the bacteriophage, thus MMC at 2.4 mg/kg/ip/24h and vB_KpnM-VAC13 at 10 MOI/ip/24h, for 72 hours. After the evaluation at 7 days post treatment, no systemic signs of pain or weight loss were observed.
3. Discussion
The repurposing or repositioning of drugs is the strategy of re-evaluating the therapeutic indications of a pharmaceutic product [26,27]. At a time when new antibiotics are no longer effective, and resistance and persistence to them are increasing, the use of drugs that have already been developed, tested and approved for their use in humans with therapeutic indications other than antibacterial could raise as a promising alternative [28]. Furthermore, it is important to note that antibiotics provoke a disturbance on the healthy microbiota, with a concomitant reduction of bacterial species diversity, altered metabolic activity and the selection of antibiotic-resistant microorganisms [29].
Several studies have already placed MMC as a readily applicable treatment for clinical infections, regarding its bactericidal effects against diverse pathogens [19,20,23,30,31]. In this work we further analyzed the possible synergism between the lytic bacteriophage vB_KpnM-VAC13 in combination with MMC in two different clinical strains of K. pneumoniae and in one reference strain. Furthermore, we verified the absence of toxicity for the repurposed anticancer drug mitomycin C both in vitro and in healthy immunocompetent mice, together with an in vivo toxicity assay with the lytic phage vB_KpnM-VAC13 at its highest concentration.
A synergistic effect between MMC and the lytic phage on the K. pneumoniae imipenem-persister isolate K3325 (Figure 1 and Figure 2) was observed, verified by a decrease of more than 2 log of CFU/mL in the presence of the combination compared with the most active single agent (Figure 2e) [32,33]. The resistant isolate K2534 did not show any affectation of its growth in the presence of MMC, which is consistent with its high MIC to this compound (25 mg/L), already determined in a preliminary study published by our research group [23]. On the other hand, the persister isolate K3325 showed the highest inhibition of its growth in the presence of MMC, consistently with its MIC value of 6.25 mg/L [23]. Besides, the reference strain ATCC®10031TM exhibited a heavy affectation on its growth under all the conditions evaluated: phage alone, mitomycin C alone and the combination of these two (Figure 1c and 1f).
The different growth curves shown by K2534 and K3325 in the presence of this anticancer molecule, alone and combined, could be explained by the distinctive working mechanism of the nucleotide excision repair (NER) UvrABC system, as well as other defense systems that resolve double-strand breaks provoked by MMC [34]. As K2534 is not a persister isolate, it is therefore highly likely that UvrABC system works properly and resolves the DNA crosslinking reactions that MMC induces in its genome, as previously observed [20,21,35], which would explain the absence of bactericidal activity for the conditions in which MMC was added for this isolate.
All in all, we concluded that MMC at 3 μg/mL exhibited a bactericidal activity both in vitro and in vivo (even a synergistic effect when combined with a lytic phage), at least in the case of bacterial persister cells in which NER mechanisms are altered.
Concerning the infection by the phage vB_KpnM-VAC13, different patterns of growth inhibition were observed in vitro (Figure 1). In the case of K2534, OD growth curves revealed a complete inhibition of its growth for the first 8 hpi, and then a regrowth was visible, probably due to the rapid development of phage resistance mutations [36]. This would explain the high frequency of resistant mutants that was assessed for this strain (Figure 2). Interestingly, vB_KpnM-VAC13 did not produce the same effect on the K3325 isolate, being unable to produce a visible lysis at the spectrophotometer (Figure 1b) and confirmed with the unusually high frequency of phage-resistant mutants exhibited by this strain (Figure 2). However, the combination of MMC and the phage successfully reduced the frequency of resistance for both K3325 and the reference strain; differently but expectedly, the K2534 isolate did not show any statistically significant difference in the resistance rates among the three conditions evaluated.
Even if there are toxicity concerns associated with the administration of MMC, the concentrations at which this anticancer exerts bactericidal effects are similar to the therapeutic ones that have been established for cancer treatments: MMC has been safely administered at 2 μg/mL intravenously, and up to 400 μg/mL topically [18,21]. Regarding the toxicity issues that this molecule entails, here we have performed both acute and cumulative toxicity assays, using healthy immunocompetent mice, together with an in vitro assay using cultured human chondrocytes belonging to the cell line T/C28a2.
We then used the human chondrocytes cell line T/C28a2 and exposed it to several concentrations of MMC for 24 h, measured the metabolic activity of the cells and quantified the populations that were alive, dead, or have entered an apoptotic state by flux cytometry (Figure 4). Importantly, at this concentration a high percentage of viability was assessed for human chondrocytes, together with a low level of apoptosis induction, verifying the absence of toxicity of this compound. In what concerns the in vivo model, we did not observe any systemic signs of pain or weight loss for the concentrations evaluated, neither for the acute toxicity experiment nor for the cumulative toxicity at 72h. This confirms the safety of MMC and vB_KpnM-VAC13 at these concentrations in healthy female immunocompetent mice, even if further experiments will be needed to confirm these results.
Taking into consideration that the discovery and development of new antibiotics are currently limited and, in most cases, unsuccessful, the repurposing approach is gaining interest. As it represents a notable reduction of the time, risks and costs associated with the production of new drugs, since the repositioned molecules have already been approved by regulatory agencies and proved to be safe and efficacious, efforts must be made in order to find synergistic relations between repurposed compounds and other antibacterials, such as lytic phages [37,38]. This work represents one example of an exciting synergism between an anticancer compound and a lytic phage, proven to be effective against persister subpopulations of K. pneumoniae and, importantly, exhibiting no toxicity against human cells and healthy, immunocompetent mice.
4. Materials and Methods
4.1. Bacterial Strains, Phage and Growth Culture Conditions
K. pneumoniae clinical strains K2534 (imipenem-resistant) and K3325 (imipenem-persister) were isolated and stored at the National Center of Microbiology (CNM, Madrid, Spain). K. pneumoniae K2534 (ST437-OXA245) was isolated from a rectal sample, whereas K3325 was isolated from a blood sample. K. pneumoniae subsp. pneumoniae reference strain ATCC®10031TM was also employed for several experiments. All the strains were cultured in Luria-Bertani (LB) growth medium (1% tryptone, 0.5% NaCl and 0.5% yeast extract) at 37˚C and, most of the time, in shaking conditions (180 rpm).
The lytic bacteriophage vB_KpnM-VAC13 was isolated from sewage water and phenotypically and genomically characterized by our group in recent works [23,39]. MMC was purchased at SigmaAldrich® and diluted in deionized water or filter-sterilized saline buffer.
Characteristics of the K. pneumoniae strains and the lytic phage used throughout this work are reported in Table 1:
4.2. Optical Density Growth Curves
Overnight cultures of K. pneumoniae clinical strains K2534 and K3325 and the reference strain K. pneumoniae ATCC®10031TM were cultured in LB medium in 96-well flat-bottom plates (200 μl final volume) in the presence of the lytic bacteriophage vB_KpnM-VAC13 at multiplicities of infection (MOI) of 10, in presence of 3 μg/mL of MMC, or with both together. In every case, MMC was added 1-hour post-infection (hpi) to allow the entrance into the persistent state. A row of LB exclusively inoculated with bacteria was used as growth control, while other two rows containing either non-inoculated LB, or MMC at 3 μg/mL, were included as blanks. The plates were incubated with continuous shaking and read using an EPOCH Microplate Reader (BioTek®) at an optical density of 600 nm (OD600 nm), measured every 15 minutes for 24 hpi.
4.3. Time-Kill Curves Assay
The same setup as described in the aforementioned section was used for the viability assay. At 24h, the cultures were serially diluted in saline buffer and plated in duplicate on LB-agar plates (1% tryptone, 0.5% NaCl and yeast extract, 2% agar). Briefly, curves started with an initial inoculum of 107 CFU/mL and samples were taken at 0 and 24 h. Bactericidal activity was defined as a decrease of ≥3 log10 CFU/mL from the initial inoculum, whereas synergy was defined as a decrease ≥2 log10 CFU/mL for the combination compared with the most active single agent [32].
4.4. Frequency of Resistant Mutants
Overnight cultures of three different clones belonging to K. pneumoniae isolates K2534, K3325 and ATCC®10031TM were diluted 1:100 in LB broth medium until an optical density equivalent to 107 CFU/mL was reached. Dilutions of the cultures were performed in saline buffer, then 100 µL were plated on LB-agar plates containing 3 mg/L of MMC to assess the resistance arising to this molecule alone. To calculate the frequency of phage-resistant mutants to the phage vB_KpnM-VAC13 alone or combined with MMC, 100 µL of a solution containing 109 plaque-forming units (PFU)/mL were added to 100 µL of the diluted cultures, then plated by the top-agar method (or double-layer method) onto TA-agar plates (1% tryptone, 0.5% NaCl, 1.5% agar), or onto MMC-containing TA plates (supplemented with MMC at 3 mg/L). The top-agar method has been previously described in [41,42] and the protocol of determination of the phage-resistance frequency is based on previous works as well [43].
4.5. Respiration Assay Using the Tetrazolium Salt WST-1
K. pneumoniae clinical strains K2534 and K3325 and the reference strain K. pneumoniae ATCC®10031TM were cultured in LB medium in flat-bottom 96-well plates (200 μl final volume) in the presence of either the phage at MOI = 10, MMC (3 μg/mL), or both (MMC added 1 hpi). The reagent WST-1 (Roche®), a tetrazolium salt that produces a color change in the medium when reduced by the presence of NAD(P)H, was diluted 1:100 in every well, including the growth control and the blanks. NADPH is considered an indicator of bacterial metabolic activity. The plates were incubated without shaking and read within an EPOCH Microplate Reader (BioTek®). The OD at a wavelength of 480 nm was measured every 15 minutes during 24 hpi for K2534 and ATCC®10031TM and at 12 hpi for K3325.
4.6. Cytotoxicity Assay Using Human Chondrocytes T/C28a2 Cell Line
4.6.1. MTT-Cytotoxic Assay
Cell viability was determined by an MTT-based cytotoxicity assay on human cells, specifically the T/C28a2 chondrocyte cell line. MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide) produces purple formazan by the mitochondrial enzymes of viable cells. Cultured T/C28a2 chondrocytes at a concentration of 6,500 cells/well were seeded in a 96-well cell-culture plate (Corning®) and allowed to form a monolayer for 24 h. After visually assessing that the cells had reached a confluence around 80% (medium cell density), they were exposed to 200 μl of DMEM High Glucose-Pyruvate medium (Gibco®) supplemented with 10% fetal bovine serum (FBS, Gibco®) and 1% penicillin/streptomycin, containing various concentrations of MMC (firstly, 0.3 to 300 μg/mL were assessed in 10-fold, then the range was restraint to 3, 6, 8, 10 and 20 mg/L of MMC) for 24 h and 48 h. A non-supplemented medium (DMEM + 0% FBS) was included as a control to verify that the observed effect was indeed due to the presence of the drug.
Then, 200 μL of MTT diluted in DMEM was added to the wells. After 3 h incubation at 37˚C in a 5% CO2 incubator, the dye solution was carefully removed, and the crystals of purple formazan were solubilized with 100 μL DMSO (Sigma Aldrich). The plate was then incubated for 15 min at room temperature on a shaker, and absorbance at 570 nm and 630 nm (reference) was measured by NanoQuant microplate reader. The experiment was done using six technical replicates from three biological replicates.
4.6.2. Apoptosis Study Using Flow Cytometry
Cultured T/C28a2 chondrocytes at a concentration of 15,000 cells/well were seeded in a 48-well culture plate (Corning®) and allowed to form a monolayer for 24 h. After checking that the cells had reached a medium cell confluence, they were exposed to 300 μl of DMEM medium supplemented with 10% FBS and 1% penicillin/streptomycin, containing a range of MMC concentrations (0 -only supplemented DMEM-, 3, 6 and 10 mg/L), and were incubated for 24 h at 5% CO2 and 37˚C. Six wells were included as technical replicates and the experiment was done using biological replicates.
After 24 h of incubation with the MMC, the medium was aspirated and kept on previously labelled Eppendorf tubes to take into consideration any naturally occurring dead cells. The wells were rinsed with filtered saline buffer and 100 µL of 2x Trypsin-EDTA 0.5% (Gibco®) were added to each well. The plate was incubated at 37˚C for 5 min to produce the detachment of the chondrocytes. Meantime, the Eppendorf tubes were centrifuged at 1500 rpm for 5 min and the supernatants were discarded. The trypsinized cells were added to the corresponding pellets obtained after the last centrifugation, and the tubes were again centrifuged for 5 min at 1500 rpm. After removing the supernatants once again, cells were washed with 300 µL of saline buffer and this step was repeated once more. Finally, pelleted cells were resuspended with the Binding Buffer (1x) of the “Dead Cell Apoptosis Kit with Annexin V FITC & Propidium Iodide for Flow Cytometry” (InvitrogenTM) containing 1 µL of propidium iodide (PI) at 1.5 mM in deionized water, and 5 µL of annexin V-FITC (25mM HEPES, 140 mM NaCl, 1mM EDTA pH 7.4, 0.1% BSA) per reaction. Half of the tubes were resuspended in saline buffer without any PI or annexin V-FITC, and used as controls to define the gates at the flow cytometer CytoFLEX S (Beckman Coulter).
4.7. Acute and Cumulative of MMC and Lytic Bacteriophage vB_KpnM-VAC13 in Healthy Female C57BL6/J Mice
Immunocompetent C57BL/6J mice weighing 20 g were used. The mice had murine pathogen-free sanitary status and were assessed for genetic authenticity. This study was conducted in accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals (National Research Council. Guide for the Care and Use of Laboratory Animals. The National Academies Press 2011). Experiments were approved by the Committee on the Ethics of Animal Experiments of the Regional Ministry of Agriculture, Livestock, Fisheries and Development, Spain (04/07/2022/099).
For acute toxicity, groups of 6 healthy female mice, 7-week-old, were intraperitoneally (ip) treated with a single dose of MMC at concentrations of 2.4, 1.7 and 0.52 mg/L and vB_KpnM-VAC13 at MOI of 0.5, 1 and 10. After the doses and during seven days, the following indicative signs of pain were assessed: reduced water (dehydration) or food intake; isolation; self-mutilation; tremors/spasms; dyspnea; physical activity (increased/reduced); chromodacryorrhoea; muscle stiffness; piloerection; teeth grinding; or weight loss. For the cumulative toxicity, groups of 6 healthy C57BL/6J female mice received for 72 hours, the highest dose that implied no signs of toxicity in the acute toxicity studies previously evaluated, which was 2.4 mg/kg/ip/q24h and MOI10/ip/q24h, for MMC and/or vB_KpnM-VAC13, respectively. The same systematic signs of pain, listed above, were monitored.
Author Contributions
O.P., developed the experiments and wrote the original manuscript; A.B-P., C.L-C., L.A., collaborated in the developed the experiments; I.B., O.P., L.F-G., analyzed the references as well collaborated in writing the manuscript; C.O-C., C.I-Q., observed the results and help to design the illustrations; R.C., A.O., re-written the manuscript; and finally. M.T., supervised all results, validated the work and obtained funding for the research.
Funding
This study was funded by grant PI22/00323 awarded to M. Tomás, within the State Plan for R+D+I 2013-2016 (National Plan for Scientific Research, Technological Development and Innovation 2008-2011) and co-financed by the ISCIII-Deputy General Directorate for Evaluation and Promotion of Research - European Regional Development Fund “A Way of Making Europe” and Instituto de Salud Carlos III FEDER, Spanish Network for the Research in Infectious Diseases (REIPI, RD16/0016/0006 and CIBER CB21/13/00012, CB21/13/00084 and CB21/13/00095), Instituto de Salud Carlos III FEDER and by a grant from the Instituto de Salud Carlos III (MePRAM Project, PMP22/00092), Instituto de Salud Carlos III, Ministerio de Ciencia e Innovación, funded by NextGeneration European Union funds that support the actions of the Resilience and Recovery Facility. L. Fernandez-Garcia was supported by a Fulbright Scholar Fellowship and a Xunta de Galicia Postdoctoral Grant.
Institutional Review Board Statement
The animal study protocol used in this work was conducted in accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals (National Research Council. Guide for the Care and Use of Laboratory Animals. The National Academies Press 2011). Experiments were approved by the Committee on the Ethics of Animal Experiments of the Regional Ministry of Agriculture, Livestock, Fisheries and Development, Spain (04/07/2022/099).
Data Availability Statement
Genomic data of the clinical strains and the lytic phage used in this work can be found in GenBank (NCBI) under these accession numbers: WRXG00000000.1, SAMEA3649525 and MZ322895.1, respectively.
Acknowledgments
We thank PhD Mercedes Fernández Moreno and PhD Carlos Vaamonde García for kindly providing us with the human chondrocytes cell line T/C28a2. We also thank PhD Juan Antonio Fafián Labora for his useful help with the flux cytometer.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Clegg, S.; Murphy, C.N. Epidemiology and Virulence of Klebsiella pneumoniae. Microbiol Spectr 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Dong, N.; Zheng, Z.; Lin, D.; Huang, M.; Wang, L.; Chan, E.W.; Shu, L.; Yu, J.; Zhang, R.; et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: a molecular epidemiological study. Lancet Infect Dis 2018, 18, 37–46. [Google Scholar] [CrossRef]
- Zeng, L.; Yang, C.; Zhang, J.; Hu, K.; Zou, J.; Li, J.; Wang, J.; Huang, W.; Yin, L.; Zhang, X. An Outbreak of Carbapenem-Resistant Klebsiella pneumoniae in an intensive care unit of a major teaching hospital in Chongqing, China. Front Cell Infect Microbiol 2021, 11, 656070. [Google Scholar] [CrossRef] [PubMed]
- Duman, Y.; Ersoy, Y.; Gursoy, N.C.; Altunisik Toplu, S.; Otlu, B. A silent outbreak due to Klebsiella pneumoniae that co-produced NDM-1 and OXA-48 carbapenemases, and infection control measures. Iran J Basic Med Sci 2020, 23, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Di Pilato, V.; Principe, L.; Andriani, L.; Aiezza, N.; Coppi, M.; Ricci, S.; Giani, T.; Luzzaro, F.; Rossolini, G.M. Deciphering variable resistance to novel carbapenem-based β-lactamase inhibitor combinations in a multi-clonal outbreak caused by Klebsiella pneumoniae carbapenemase (KPC)-producing Klebsiella pneumoniae resistant to ceftazidime/avibactam. Clin Microbiol Infect 2023, 29, 537.e531–537.e538. [Google Scholar] [CrossRef]
- Huemer, M.; Mairpady Shambat, S.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic resistance and persistence-Implications for human health and treatment perspectives. EMBO Rep 2020, 21, e51034. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Merrin, J.; Chait, R.; Kowalik, L.; Leibler, S. Bacterial persistence as a phenotypic switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef]
- Jayaraman, R. Bacterial persistence: some new insights into an old phenomenon. J Biosci 2008, 33, 795–805. [Google Scholar] [CrossRef]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial persister cell formation and dormancy. Appl Environ Microbiol 2013, 79, 7116–7121. [Google Scholar] [CrossRef]
- Helaine, S.; Cheverton, A.M.; Watson, K.G.; Faure, L.M.; Matthews, S.A.; Holden, D.W. Internalization of Salmonella by macrophages induces formation of nonreplicating persisters. Science 2014, 343, 204–208. [Google Scholar] [CrossRef]
- Koeva, M.; Gutu, A.D.; Hebert, W.; Wager, J.D.; Yonker, L.M.; O'Toole, G.A.; Ausubel, F.M.; Moskowitz, S.M.; Joseph-McCarthy, D. An Antipersister Strategy for Treatment of Chronic Pseudomonas aeruginosa Infections. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Goneau, L.W.; Yeoh, N.S.; MacDonald, K.W.; Cadieux, P.A.; Burton, J.P.; Razvi, H.; Reid, G. Selective target inactivation rather than global metabolic dormancy causes antibiotic tolerance in uropathogens. Antimicrob Agents Chemother 2014, 58, 2089–2097. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Xie, X.; Tian, W.; Bahar, A.A.; Lin, N.; Song, F.; An, J.; Ren, D. (Z)-4-bromo-5-(bromomethylene)-3-methylfuran-2(5H)-one sensitizes Escherichia coli persister cells to antibiotics. Appl Microbiol Biotechnol 2013, 97, 9145–9154. [Google Scholar] [CrossRef] [PubMed]
- Windels, E.M.; Michiels, J.E.; Fauvart, M.; Wenseleers, T.; Van den Bergh, B.; Michiels, J. Bacterial persistence promotes the evolution of antibiotic resistance by increasing survival and mutation rates. Isme j 2019, 13, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Pacios, O.; Blasco, L.; Bleriot, I.; Fernandez-Garcia, L.; Gonzalez Bardanca, M.; Ambroa, A.; Lopez, M.; Bou, G.; Tomas, M. Strategies to Combat Multidrug-Resistant and Persistent Infectious Diseases. Antibiotics (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Uyttebroek, S.; Chen, B.; Onsea, J.; Ruythooren, F.; Debaveye, Y.; Devolder, D.; Spriet, I.; Depypere, M.; Wagemans, J.; Lavigne, R.; et al. Safety and efficacy of phage therapy in difficult-to-treat infections: a systematic review. Lancet Infect Dis 2022, 22, e208–e220. [Google Scholar] [CrossRef]
- Pirnay, J.P.; Djebara, S.; Steurs, G.; Griselain, J.; Cochez, C.; De Soir, S.; Glonti, T.; Spiessens, A.; Vanden Berghe, E.; Green, S.; et al. Retrospective, observational analysis of the first one hundred consecutive cases of personalized bacteriophage therapy of difficult-to-treat infections facilitated by a Belgian consortium. 2023. [CrossRef]
- Bradner, W.T. Mitomycin C: a clinical update. Cancer Treat Rev 2001, 27, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Verweij, J.; Pinedo, H.M. Mitomycin C: mechanism of action, usefulness and limitations. Anticancer Drugs 1990, 1, 5–13. [Google Scholar]
- Cruz-Muñiz, M.Y.; López-Jacome, L.E.; Hernández-Durán, M.; Franco-Cendejas, R.; Licona-Limón, P.; Ramos-Balderas, J.L.; Martinéz-Vázquez, M.; Belmont-Díaz, J.A.; Wood, T.K.; García-Contreras, R. Repurposing the anticancer drug mitomycin C for the treatment of persistent Acinetobacter baumannii infections. Int J Antimicrob Agents 2017, 49, 88–92. [Google Scholar] [CrossRef]
- Kwan, B.W.; Chowdhury, N.; Wood, T.K. Combatting bacterial infections by killing persister cells with mitomycin C. Environ Microbiol 2015, 17, 4406–4414. [Google Scholar] [CrossRef]
- Huang, D.; Zhang, M.; Zhang, H.; Cui, Z.; Luo, D.; Li, T.; Li, X.; He, Y.; Zhang, S.L. Design and synthesis of TPP+-Mitomycin C conjugate with reduced toxicity. Bioorg Med Chem Lett 2022, 77, 129036. [Google Scholar] [CrossRef] [PubMed]
- Pacios, O.; Fernández-García, L.; Bleriot, I.; Blasco, L.; González-Bardanca, M.; López, M.; Fernández-Cuenca, F.; Oteo, J.; Pascual, Á.; Martínez-Martínez, L.; et al. Enhanced Antibacterial Activity of Repurposed Mitomycin C and Imipenem in Combination with the Lytic Phage vB_KpnM-VAC13 against Clinical Isolates of Klebsiella pneumoniae. Antimicrob Agents Chemother 2021, 65, e0090021. [Google Scholar] [CrossRef] [PubMed]
- Bleriot, I.; Blasco, L.; Pacios, O.; Fernández-García, L.; Ambroa, A.; López, M.; Ortiz-Cartagena, C.; Cuenca, F.F.; Oteo-Iglesias, J.; Pascual, Á.; et al. The role of PemIK (PemK/PemI) type II TA system from Klebsiella pneumoniae clinical strains in lytic phage infection. Sci Rep 2022, 12, 4488. [Google Scholar] [CrossRef] [PubMed]
- L, F.-G.; M, T.; TK, W. Ribosome inactivation by Escherichia coli GTPase RsgA inhibits T4 phage. 2023, 14. [CrossRef]
- Farha, M.A.; Brown, E.D. Drug repurposing for antimicrobial discovery. Nat Microbiol 2019, 4, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tong, Z.; Shi, J.; Li, R.; Upton, M.; Wang, Z. Drug repurposing for next-generation combination therapies against multidrug-resistant bacteria. Theranostics 2021, 11, 4910–4928. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: progress, challenges and recommendations. Nat Rev Drug Discov 2019, 18, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.; Guarner, F.; Bustos Fernandez, L.; Maruy, A.; Sdepanian, V.L.; Cohen, H. Antibiotics as Major Disruptors of Gut Microbiota. Front Cell Infect Microbiol 2020, 10, 572912. [Google Scholar] [CrossRef]
- Soo, V.W.; Kwan, B.W.; Quezada, H.; Castillo-Juárez, I.; Pérez-Eretza, B.; García-Contreras, S.J.; Martínez-Vázquez, M.; Wood, T.K.; García-Contreras, R. Repurposing of Anticancer Drugs for the Treatment of Bacterial Infections. Curr Top Med Chem 2017, 17, 1157–1176. [Google Scholar] [CrossRef]
- Domalaon, R.; Ammeter, D.; Brizuela, M.; Gorityala, B.K.; Zhanel, G.G.; Schweizer, F. Repurposed Antimicrobial Combination Therapy: Tobramycin-Ciprofloxacin Hybrid Augments Activity of the Anticancer Drug Mitomycin C Against Multidrug-Resistant Gram-Negative Bacteria. Front Microbiol 2019, 10, 1556. [Google Scholar] [CrossRef]
- Herrera-Espejo, S.; Del Barrio-Tofiño, E.; Cebrero-Cangueiro, T.; López-Causapé, C.; Álvarez-Marín, R.; Cisneros, J.M.; Pachón, J.; Oliver, A.; Pachón-Ibáñez, M.E. Carbapenem Combinations for Infections Caused by Carbapenemase-Producing Pseudomonas aeruginosa: Experimental In Vitro and In Vivo Analysis. Antibiotics (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Ayerbe-Algaba, R.; Gil-Marqués, M.L.; Jiménez-Mejías, M.E.; Sánchez-Encinales, V.; Parra-Millán, R.; Pachón-Ibáñez, M.E.; Pachón, J.; Smani, Y. Synergistic Activity of Niclosamide in Combination With Colistin Against Colistin-Susceptible and Colistin-Resistant Acinetobacter baumannii and Klebsiella pneumoniae. Front Cell Infect Microbiol 2018, 8, 348. [Google Scholar] [CrossRef]
- Weng, M.W.; Zheng, Y.; Jasti, V.P.; Champeil, E.; Tomasz, M.; Wang, Y.; Basu, A.K.; Tang, M.S. Repair of mitomycin C mono- and interstrand cross-linked DNA adducts by UvrABC: a new model. Nucleic Acids Res 2010, 38, 6976–6984. [Google Scholar] [CrossRef]
- Chowdhury, N.; Wood, T.L.; Martínez-Vázquez, M.; García-Contreras, R.; Wood, T.K. DNA-crosslinker cisplatin eradicates bacterial persister cells. Biotechnol Bioeng 2016, 113, 1984–1992. [Google Scholar] [CrossRef]
- Moulton-Brown, C.E.; Friman, V.P. Rapid evolution of generalized resistance mechanisms can constrain the efficacy of phage-antibiotic treatments. Evol Appl 2018, 11, 1630–1641. [Google Scholar] [CrossRef]
- Rangel-Vega, A.; Bernstein, L.R.; Mandujano-Tinoco, E.A.; García-Contreras, S.J.; García-Contreras, R. Drug repurposing as an alternative for the treatment of recalcitrant bacterial infections. Front Microbiol 2015, 6, 282. [Google Scholar] [CrossRef]
- Peyclit, L.; Baron, S.A.; Rolain, J.M. Drug Repurposing to Fight Colistin and Carbapenem-Resistant Bacteria. Front Cell Infect Microbiol 2019, 9, 193. [Google Scholar] [CrossRef]
- Pacios, O.; Fernández-García, L.; Bleriot, I.; Blasco, L.; Ambroa, A.; López, M.; Ortiz-Cartagena, C.; Cuenca, F.F.; Oteo-Iglesias, J.; Pascual, Á.; et al. Phenotypic and Genomic Comparison of Klebsiella pneumoniae Lytic Phages: vB_KpnM-VAC66 and vB_KpnM-VAC13. Viruses 2021, 14. [Google Scholar] [CrossRef]
- Available online: https://genomes.atcc.org/genomes/f629ae6e6bcd483e (accessed on 15 January 2024).
- Abedon, S.T.; Yin, J. Bacteriophage plaques: theory and analysis. Methods Mol Biol 2009, 501, 161–174. [Google Scholar] [CrossRef]
- Raya, R.R.; H'bert, E.M. Isolation of Phage via Induction of Lysogens. Methods Mol Biol 2009, 501, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.; Pereira, C.; Almeida, A. Sequential Combined Effect of Phages and Antibiotics on the Inactivation of Escherichia coli. Microorganisms 2018, 6. [Google Scholar] [CrossRef]
Figure 1.
a, b and c: Optical density growth curves performed with a microtiter plate reader and incubator at a wavelength of 600 nm every 15 minutes, using K. pneumoniae clinical strains K2534 (A), K3325 (B) and the reference strain ATCC®10031TM (C), all infected with vB_KpnM-VAC13; d, e and f: Viability of these three strains at the tested conditions, assessed after CFU enumeration. Only statistically significant comparisons have been depicted (** represents a p-value < 0.01).
Figure 1.
a, b and c: Optical density growth curves performed with a microtiter plate reader and incubator at a wavelength of 600 nm every 15 minutes, using K. pneumoniae clinical strains K2534 (A), K3325 (B) and the reference strain ATCC®10031TM (C), all infected with vB_KpnM-VAC13; d, e and f: Viability of these three strains at the tested conditions, assessed after CFU enumeration. Only statistically significant comparisons have been depicted (** represents a p-value < 0.01).
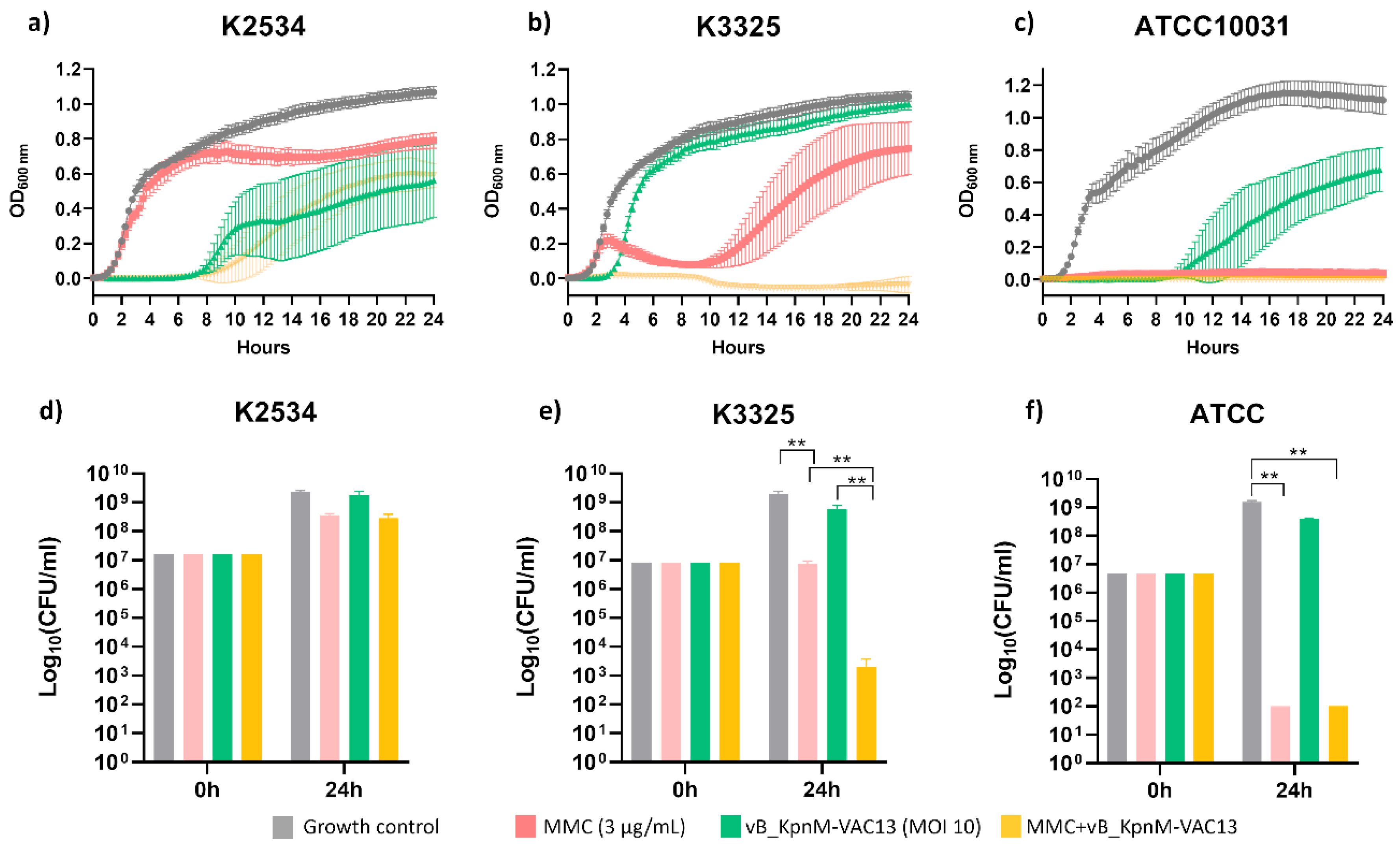
Figure 2.
Frequency of resistant mutants in the presence of the phage alone, mitomycin C (MMC) at 3 mg/L and both combined.
Figure 2.
Frequency of resistant mutants in the presence of the phage alone, mitomycin C (MMC) at 3 mg/L and both combined.
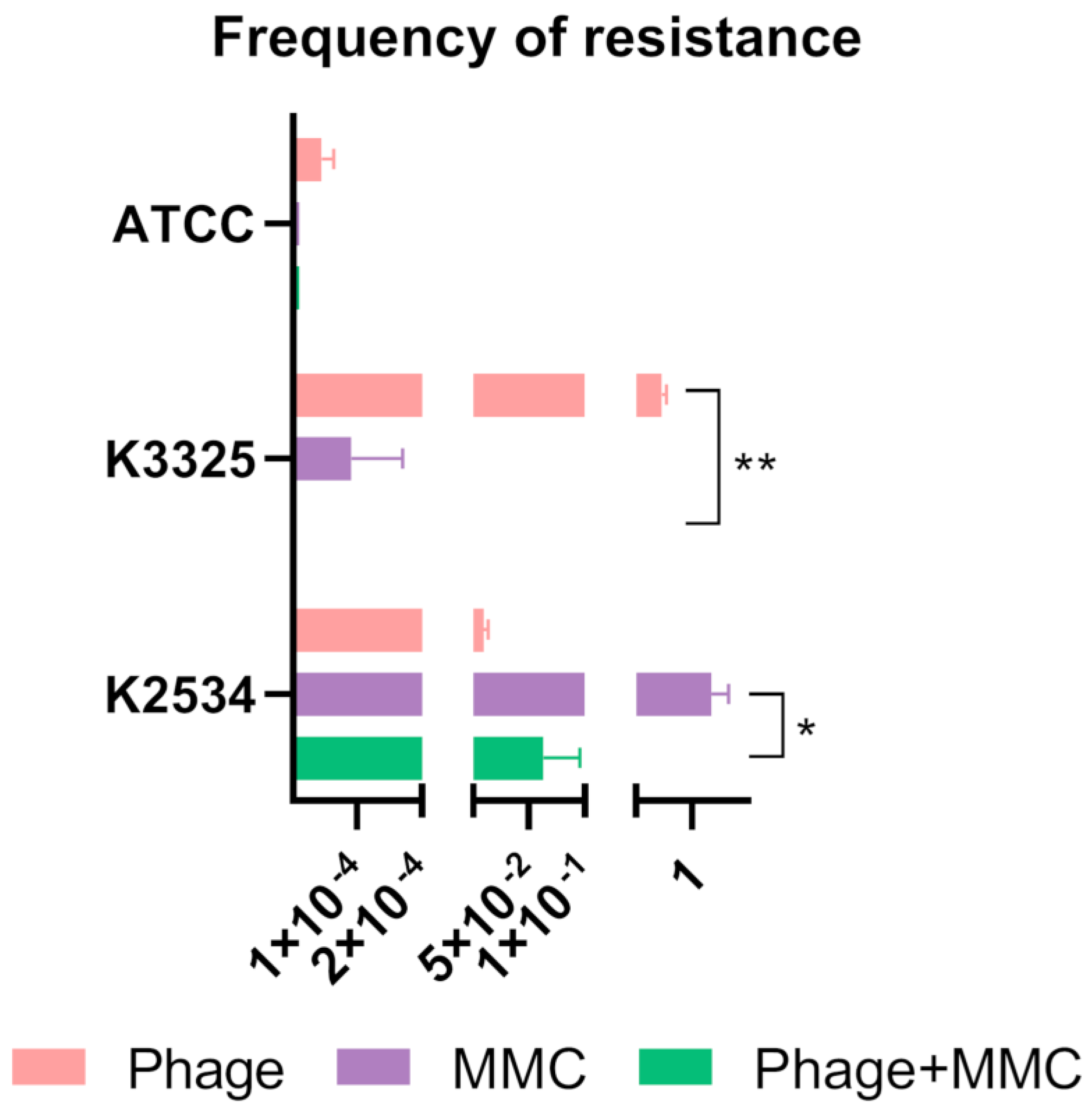
Figure 4.
In vitro evaluation of the toxicity of MMC using the human chondrocyte cell line T/C28a2. A) 96-well microplate containing T/C28a2 cells after MTT-incubation and DMSO addition. B) MTT cytotoxic assay represented as the percentage of viability after measurement of the OD570nm at 24h of incubation with the corresponding concentrations of MMC. *** represents p-values lower than 0.001. C) Viability assay using the Live-Dead Apoptosis Kit, with propidium iodide and annexin V-FITC and using the flow cytometer CytoFLEX (Beckman Coulter). *represents p-values between 0.05 and 0.01; ** represents p-values between 0.01 and 0.001; *** represents p-values lower than 0.001. D) Representative graphs after flow cytometry analysis using the CytExpert 2.0 software (PI: propidium iodide; FITC: fluorescein isothiocyanate).
Figure 4.
In vitro evaluation of the toxicity of MMC using the human chondrocyte cell line T/C28a2. A) 96-well microplate containing T/C28a2 cells after MTT-incubation and DMSO addition. B) MTT cytotoxic assay represented as the percentage of viability after measurement of the OD570nm at 24h of incubation with the corresponding concentrations of MMC. *** represents p-values lower than 0.001. C) Viability assay using the Live-Dead Apoptosis Kit, with propidium iodide and annexin V-FITC and using the flow cytometer CytoFLEX (Beckman Coulter). *represents p-values between 0.05 and 0.01; ** represents p-values between 0.01 and 0.001; *** represents p-values lower than 0.001. D) Representative graphs after flow cytometry analysis using the CytExpert 2.0 software (PI: propidium iodide; FITC: fluorescein isothiocyanate).
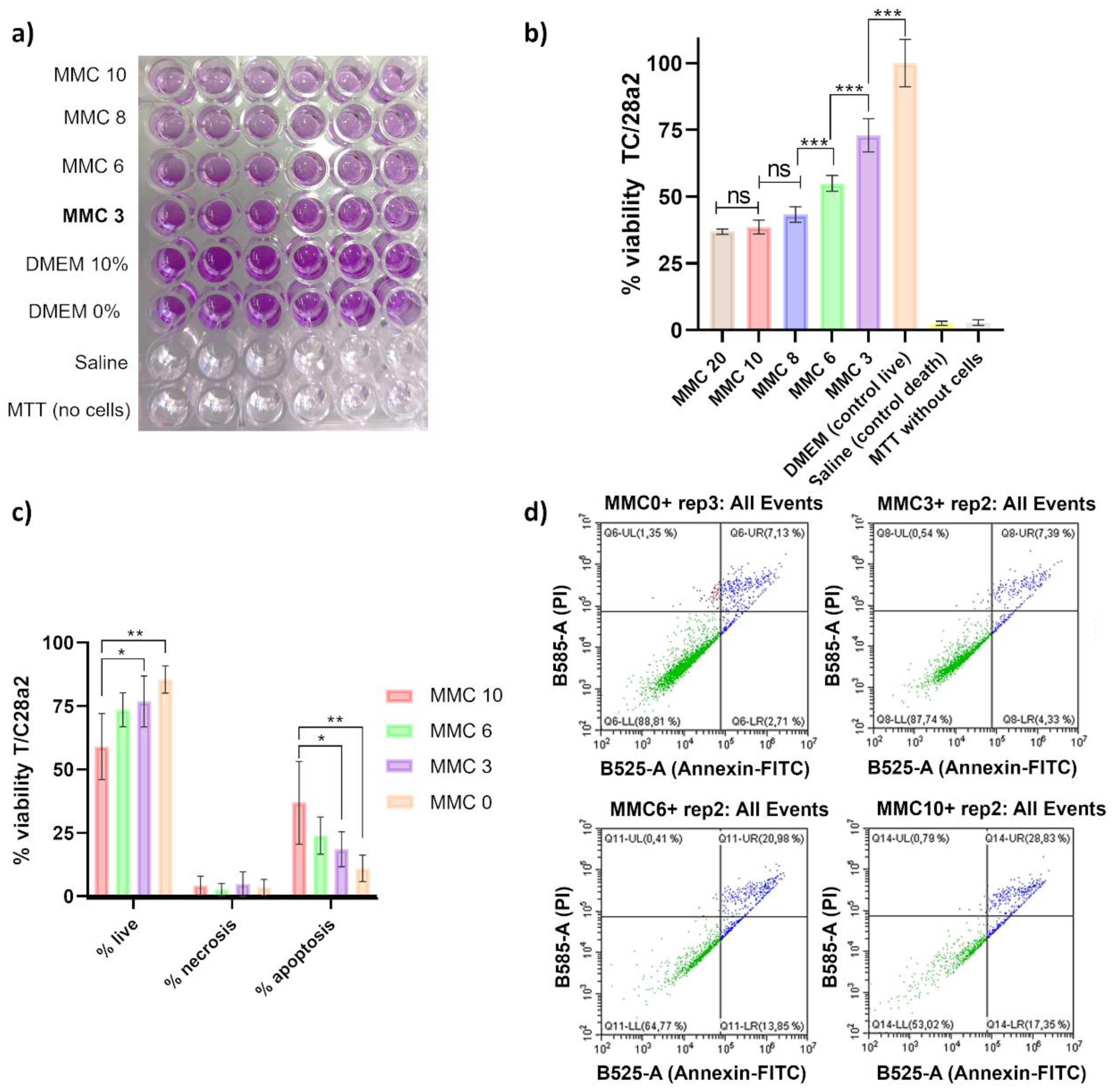

Table 1.
List of strains and bacteriophages used in this study.
| K. pneumoniae clinical strain | Sequence-type | Carbapenemase | Biological origin | GenBank acc. Number | Reference |
|---|---|---|---|---|---|
| K2534 | ST437 | OXA-245 | Rectal | WRXG00000000.1 | [23] |
| K3325 | ST42 | - | Blood | SAMEA3649525 | [23] |
| ATCC | ST131 | - | Commercial | [40] | |
| K. pneumoniae lytic bacteriophage | Morphotype | Genus | Genome size (bp) | GenBank accession number | Characterization (reference) |
| vB_KpnM-VAC13 | Myoviridae | Slopekvirus | 174826 | MZ322895.1 | [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



