
Submitted:
07 August 2024
Posted:
08 August 2024
You are already at the latest version
Abstract
Glioblastoma (GBL) is one of the more malignant primary brain tumors, currently treated by a multimodality strategy including surgery, radio- and chemotherapy, mainly consisting of Temozolomide(TMZ)-based chemotherapy. Tumor relapse often occurs due to the establishment of TMZ resistance, with a patient median survival <2 years. The identification of natural molecules having strong anti-tumor activity, led to combine these compounds with conventional chemotherapeutic agents, developing protocols of integrated anticancer therapies. Curcumin (CUR), Resveratrol (RES), and its glucoside Polydatin (PLD) are widely employed in the pharmaceutical and nutraceutical fields, and several studies demonstrated that the combination of these natural products was more cytotoxic than the single compound alone in different cancers. Some of us recently demonstrated the synergistic efficacy of the sublingual administration of a new nutraceutical formulation of CUR+PLD in reducing tumor size and improving GBL patient survival. To deepen some mechanistic aspects of these results, we investigated if the pretreatment with a combination of CUR+PLD, can improve TMZ cytotoxicity on GBL cells, by analyzing the effects on cell cycle, viability and morphology, on proteins related to cell proliferation, differentiation, apoptosis or autophagy, and on actin network. Cell viability was assessed by MTT or by the cell counter CytoSmart. CalcuSyn software was used to study the CUR+PLD synergism. Morphology was evaluated by optical and scanning electron microscopy, and protein expression was analyzed by Western Blot. Flow cytometry was used for cell cycle, autophagic flux, and apoptosis analyses. The results obtained provide evidence that CUR and PLD, acting in synergy with each other, strongly improve the efficacy of alkylating antitumor agents such as TMZ, also in drug-resistant GBL cells, through their abilities to affect survival, differentiation, and tumor invasiveness.
Keywords:
curcumin
; polydatin
; temozolomide
; chemoresistance
; glioblastoma
1. Introduction
Glioblastoma (GBL) is a highly proliferative and poorly differentiated primary brain tumor currently treated by surgery and/or by radiotherapy and systemic therapy, mainly consisting of Temozolomide(TMZ)-based chemotherapy. TMZ is an oral alkylating agent that exerts its cytotoxic effect mainly through the methylation of the guanine residues in DNA at the O6 position. This adduct can be removed by the DNA repair protein O6-methylguanine-DNA-methyltransferase (MGMT), heterogeneously expressed in glioblastoma, whose gene transcription is epigenetically silenced by the methylation of its promoter. Thus, hypermethylation of the MGMT gene promoter can determine a greater sensitivity of the tumor cell to pharmacological treatment with TMZ, and the MGMT promoter methylation status is currently used as a predictor for the benefit of TMZ therapy [1,2].
Despite recent advances in the above multimodal protocols, the overall prognosis for GBL patients remains poor, due to the establishment of TMZ resistance and tumor recurrence through infiltrative and invasive residual cancer cells, with a median survival of <2 years [3]. Thus, additional and innovative therapeutic strategies are needed to improve the outcomes of patients with GBL.
In the last decades, the identification of natural compounds which have strong anti-tumor activity by acting as potent modulators of cancer-related pathways [4], has led to the design of therapeutic protocols that combine these compounds with conventional chemotherapeutic agents. In particular, since the available therapeutic protocols for GBL treatment are not very effective, over 50% of patients use complementary and alternative approaches, among which herbal therapies are the most commonly used, but until now with little success.
Curcumin [(1E,6E)-1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione] is the main natural polyphenol extracted from the rhizome of Curcuma longa L., a flowering plant of the ginger family (Zingiberaceae) [5], mainly grown in Asia and Central America. Curcuma longa L., also known by the common name of turmeric, is widely used in traditional medicines such as in Chinese Traditional and Ayurvedic medicine, for different kinds of diseases, due to its antioxidant [6], anti-inflammatory [7], neuroprotective [8], antimicrobial properties [9,10]. Furthermore, its anticancer potential has been widely described, and is still, the topic of numerous studies [11,12,13].
Polydatin (3,5,4-trihydroxystilbene 3-O-beta-D-glucopyranosid, PLD), a glycoside of resveratrol (RES), is a natural compound belonging to the stilbene class, of the polyphenol family. It is extracted from the roots and rhizome of the Polygonum Cuspidatum, originally from Asia but currently also widespread in America and Europe. It is considered the “twin” molecule of RES, with which it shares properties but has some additional advantages. Specifically, PLD is more efficiently absorbed by the oral route and is more resistant to enzymatic oxidation than RES, that penetrates cells passively, while PLD enters cells through an active mechanism based on glucose carriers. These properties make PLD highly more bioavailable than RES [14,15]. PLD is known to have numerous beneficial effects as an antioxidant [16,17], anti-inflammatory [18,19], and immune-modulating agent [20], and as an anti-tumor natural compound in different kinds of cancer cells, also including glioblastoma [21,22,23,24,25].
CUR and PLD (or its derivate RES) are widely used alone or in combination with each other in the pharmaceutical field, such as drugs, and in the nutraceutical field, as food supplements. Several studies demonstrated that combination of low doses of CUR and RES had more potent cytotoxic efficacy than the single compound alone on different types of cancer [26,27], synergistically decreased cancer cells migration [28], shaped immune responses while reduced cancer cell survival [29], and suppressed chemoresistance [30].
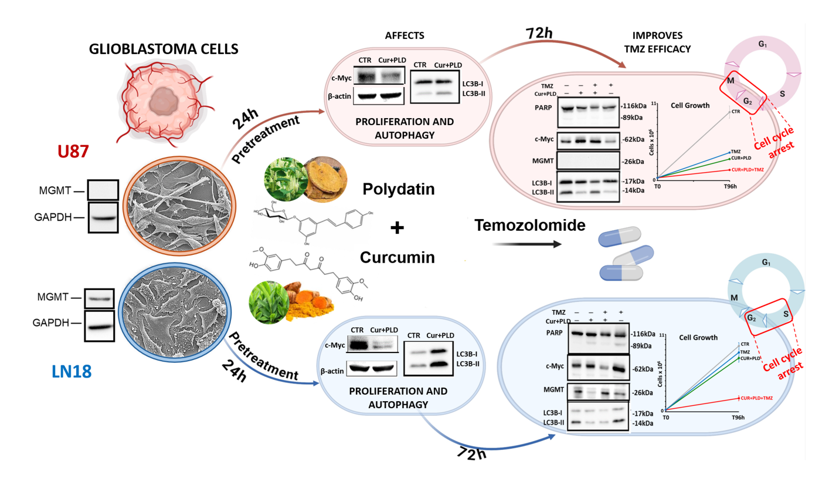
In a recent clinical study performed for a patent application [31] (Applications WO-2020026185-A1, PCT/IB2019/056565), some of us obtained data supporting the synergistic efficacy of the original sublingual administration of a combination of CUR+PLD in the treatment of grade 3rd and 4th GBL, in the context of protocols of integrated anticancer therapies. Specifically, the new nutraceutical formulation of CUR+PLD was able to improve patient survival, and significantly reduce the size of the tumor masses.
These preliminary but stimulating results, prompt us to deepen some mechanistic aspects underlying the action of CUR+PLD on GBL. To this aim, in this study, we assessed if the pretreatment with a combination of CUR and PLD, can improve synergistically TMZ antitumor efficacy in two human glioblastoma cell lines, exhibiting different responsiveness to this chemotherapeutic drug. The effects on cell viability and morphology, on proteins related to cell proliferation, differentiation, apoptosis or autophagy, and on the actin network were analyzed.
2. Results
2.1. Establishment of the Dose- and Time-Dependent Response to TMZ, and of the Synergic Concentrations of CUR and PLD in U87 and LN18 Glioblastoma Cells
Given the heterogeneous expression of the DNA repair protein MGMT in GBM, to verify if the combination of CUR+PLD could be of benefit independently from the methylation levels of the MGMT gene promoter and, consequently, from the sensitivity to TMZ, we used, as cellular models of glioblastoma, the cell lines U87 and LN18, the former negative and the latter positive for the MGMT expression (Figure 1b). Phase contrast microscopy (Figure 1a, top panels) and SEM analyses (Figure 1a, bottom panels) showed that the two cell lines exhibited different morphological features, with the U87 cells being mainly neurite-like shaped while LN18 cells having an epithelial morphology. In preliminary time- and dose-response experiments, TMZ was used at concentrations ranging from 50 to 400µM, and the dose of 200µM for 72 h was selected as the optimal condition for subsequent experiments since this condition reduced cell viability close to 50% vs untreated control in both cell lines (Figure 1c).
To define the concentrations of CUR and PLD which exhibited a synergic effect on cell viability, U87 and LN18 cell lines were treated with CUR and PLD (single or combined), at a concentration range 0.18-10 µg/ml for CUR, and 17.5-300 µg/ml for PLD, and cell viability was tested after 24 h by MTT. The combination index (CI), for all the concentrations and ratios used, was defined by the CalcuSyn software, the analyzer that automatically quantifies synergism or antagonism between drugs. Supplementary Figure S1 reports some representative results obtained by CalcuSyn in both cell lines tested. The concentrations of 10 µg/ml CUR, and 100 µg/ml PLD (ratio 1:10) were selected as the synergic combination to be used in the subsequent experiments since exhibited synergism in both cell lines (CI <0.9) and, more specifically, a strong synergism in U87 cells (CI=0.101) and a moderate synergism in LN18 (CI=0.772).
The effectiveness of the selected combination of CUR+PLD in improving the effect of TMZ on cell viability was preliminarily demonstrated by the MTT test (Figure 2b, c), performed following the experimental protocol schematized in Figure 2a, which showed that the pretreatment with CUR+PLD decreased the number of viable cells by more than 50% compared to the treatment with TMZ alone. The synergistic effect was also confirmed by the higher efficacy of the pretreatments with the combination of CUR+PLD vs those with CUR or PLD alone. Interestingly, for both cell lines, in the samples subjected to 24 h treatment with CUR+PLD and analyzed after additional 72 h of culture in fresh medium (72 h washout), a reduced number of viable cells, comparable to that recorded in samples post-treated with TMZ, was recovered (Figure 2b,c).
2.2. Effects of the Synergic Combination of CUR+PLD on U87 and LN18 Glioblastoma Cells before TMZ Treatment
To assess which modifications were induced by the CUR+PLD combination before the TMZ treatment, both cell lines were analyzed, after 24 h CUR+PLD treatment, for cell morphology, and for the expression levels of proteins related to cell proliferation (c-Myc), astroglial differentiation (GFAP), caspase-dependent apoptosis (cleaved PARP), and autophagy (LC3B). Compared with the untreated control, CUR+PLD-treated U87 cells assumed an astrocyte-like morphology (Figure 3a).
Concomitantly, WB analysis showed a significant decrease in proliferation markers c-Myc, a weak but significant increase in the astroglial differentiation marker GFAP as well as an increment of the autophagosomal marker LC3B-II (Figure 3b), while the cleaved form of PARP was not expressed, suggesting that 24 h CUR+PLD did not induce caspase-dependent apoptosis in U87 cells (Figure 3e). Conversely, LN18 cells subjected to 24 h treatment with CUR+PLD, did not exhibit an astrocyte-like morphology but showed, compared with the untreated control, numerous cells displaying morphological features indicative of apoptosis/autophagy (Figure 3c). WB analysis (Figure 3d, e) showed that GFAP was not modified compared with LN18 control cells (that are almost negative for this astroglial differentiation marker), the autophagosomal marker LC3B-II and the cleaved form of PARP were significantly increased, and, similarly to what recorded in U87 cells, the proliferation marker c-Myc was significantly decreased compared with the untreated control.
2.3. Pretreatment with CUR+PLD Significantly Improved the Effect of TMZ Treatment on the Number of Live U87 and LN18 Glioblastoma Cells
To verify the effects of the 24 h pretreatment with CUR+PLD on TMZ cytotoxicity, both cell lines cells were subjected to TMZ treatment and analyzed after additional 72 h, following the experimental protocol schematized in Figure 2a. Phase contrast microscopy on U87 cells showed that in both samples pretreated for 24 h with CUR+PLD (post-treated or not with TMZ), the astrocyte-like morphology observed after 24 h CUR+PLD was still preserved, and dying cells were almost absent (Figure 4a). The analyses by the automated cell counter CytoSmart recorded that U87 viability was not affected (Figure 4d), but the number of live cells, between T0 and 96 h, was significantly reduced in all treated samples compared with the untreated control (Figure 4b, c). Furthermore, in the sample pretreated with CUR+PLD followed by TMZ for 72 h, the number of live cells was significantly lower than in that treated with TMZ alone. Interestingly, U87 cells treated with TMZ alone or combined with CUR+PLD resulted significantly larger than the untreated or CUR+PLD-treated samples (Figure 4e). Conversely, in LN18 cultures, which possess intrinsic resistance to TMZ due to the presence of MGMT gene transcripts, TMZ treatment alone did not result in significant effects on viability or the number of live cells, compared with the untreated control. However, both parameters significantly decreased in the sample treated with CUR+PLD prior to TMZ (Figure 5b, c, d). Differently to U87 cells, in LN18 cells treated with CUR+PLD alone or plus TMZ numerous cells displaying morphological features suggestive of apoptosis/autophagy induction were observed (arrows in Figure 5a), while cell size was not significantly modified (Figure 5e).
2.4. Pretreatment with CUR+PLD Induced a Cell Cycle Arrest between the S and G2/M Phases
Cytofluorimetric analysis of the cell cycle carried out on U87 cells, recorded, in both TMZ-treated samples, an increase in the percentage of cells in the G2/M phase (about 40% of cells vs 11% and 16% in untreated control and CUR+PLD treatment alone, respectively) (Figure 6a and Supplementary Table S1). In LN18 cells, the analysis revealed a slight increase in the percentage of cells in the S/G2 phase, in the sample pre-treated with CUR+PLD plus TMZ compared to the other samples (20% vs 10%, 13%, and 15% in control, CUR+PLD, and TMZ alone, respectively) as well as an increase in the percentage of cells in the sub-G1 peak, typically associated with apoptotic/necrotic cells (20% vs 5%, 11%, and 15% in control, CUR+PLD, and TMZ alone, respectively) (Figure 6b and Supplementary Table S1).
Flow cytometric monitoring of autophagy by CytoID kit (Figure 6c, d), and apoptosis by annexin V/PI staining (Figure 6e, f, and Supplementary Table S2), showed that all treatments did not significantly modify the autophagic flux in both cell lines, and increased very weakly (in U87 cells) or moderately (in LN18 cells) the percentage of cells in early/late apoptosis, consistently with what was obtained by the cell cycle analysis.
2.5. Effects of the Synergic Combination of CUR+PLD on the expression of MGMT and Proteins Related to Cell Proliferation, Apoptosis, or Autophagy
To obtain molecular evidence supporting the data obtained by the viability assays and flow cytometry, the expression levels of proteins related to cell proliferation (c-Myc), caspase-dependent apoptosis (cleaved PARP), and autophagy (LC3B), have been assessed after 72 h TMZ treatment, with or without CUR+PLD pre-treatment, by WB (Figure 7). To verify if the pretreatment with CUR+PLD could affect the transcription of the MGMT gene and, therefore, the methylation level of its promoter, the expression level of MGMT was also analyzed.
In U87 cells (Figure 7a, c), WB showed that, in all samples analyzed, cleavage of PARP was not affected, in line with the cytofluorimetric data, and MGMT gene transcription was not activated, suggesting that the treatments did not modify the methylation levels of the MGMT promoter. Unexpectedly, at the time point analyzed, the levels of c-Myc were significantly decreased vs the control only in the sample treated with TMZ alone, while CUR+PLD (followed or not by TMZ treatment) seemed to boost cells toward proliferation. Furthermore, the autophagosomal marker LC3B-II decreased in samples treated with CUR+PLD or TMZ alone compared with the control, while in CUR+PLD plus TMZ sample LC3B-II resulted significantly higher than in cells treated with TMZ alone. These data, apparently in contrast with those obtained by flow cytometry, could be due to the distinct features of the two methods: while CytoID is more sensitive to dynamic changes in autophagy and, therefore, gives a measure of autophagic flux, WB for LC3B-II provides a direct measure of autophagosome formation and, hence, more specifically quantifies autophagy induction.
In LN18 cells (Figure 7b, d), cleaved PARP was not modified except in CUR+PLD plus TMZ, where it was down-regulated, suggesting that the moderate apoptosis recorded by flow cytometry could be due to a caspase-independent mechanism. Interestingly, in LN18 the MGMT gene transcription was down-regulated in the sample treated with CUR+PLD alone, which seems to affect the methylation levels of the MGMT promoter, thus decreasing the intrinsic resistance of these cells to TMZ. Furthermore, in LN18 cells the pretreatment with CUR+PLD before TMZ downregulated c-Myc expression, which conversely was upregulated in cells treated with TMZ alone. The autophagosomal marker LC3B-II, which was increased in TMZ-treated LN18 cells, was conversely decreased by CUR+PLD pretreatment, suggesting that this synergic combination could inhibit the autophagy-mediated defense mechanism adopted by the cells to survive the TMZ-induced DNA damage [32].
2.6. Pretreatment with CUR+PLD Alters the Actin Network and Ultrastructural Morphology of U87 and LN18 Cells
Actin filaments are one of the main components of the cellular cytoskeleton, which regulates migration and invasiveness of cancer cells, including glioblastoma cells [33,34], where TMZ has been described to moderately alter the actin cytoskeleton regulatory pathways [35]. For this ground, we also preliminary assessed if the pretreatment CUR+PLD could amplify the effect of TMZ on the actin cytoskeleton.
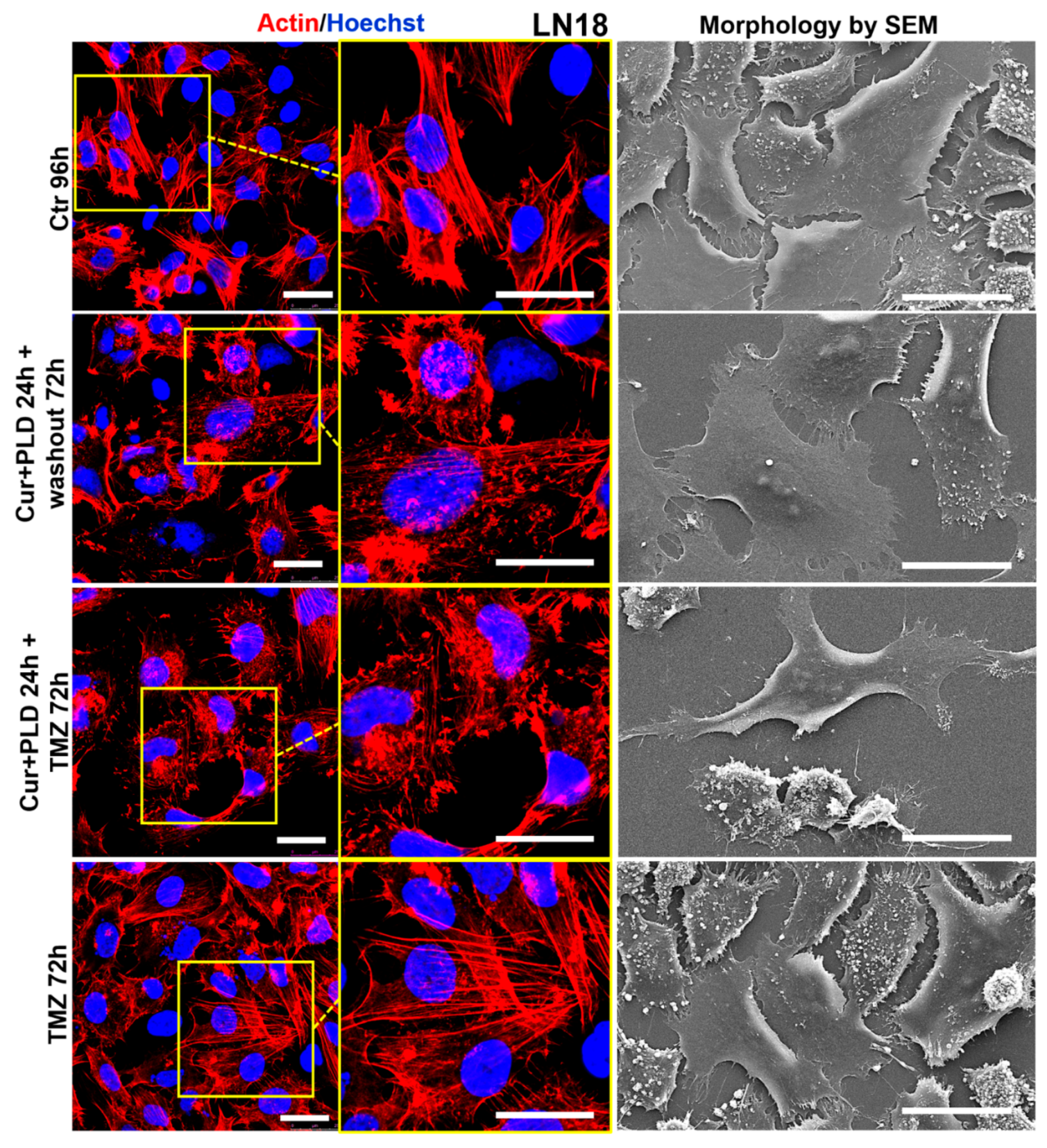
Confocal microscopy (Figure 8 and Figure 9, left panels) showed that control U87 and LN18 cells differed in organization and intracellular distribution of the actin network. Specifically, the actin cytoskeleton was less developed and with the fibrillar form mainly distributed at the cell edges and in the neurite-like structures, in U87 (Figure 8, left panels), while it was highly polymerized forming stress fibers inside almost all the cytoplasm, in LN18 (Figure 9, left panels). These differences in the actin network matched with the different shapes of the two cell lines showed by SEM analysis, the U87 being more elongated and the LN18 having a more polygonal and epithelial-like shape (Figure 8 and 9, right panels). TMZ treatment caused actin depolymerization and cell flattening in the more TMZ-sensitive U87 cells, while the actin network was less affected and cell shape was quite similar to untreated control in the intrinsically more resistant LN18 cells. Pretreatment with CUR+PLD before TMZ increased the TMZ-induced actin clumping in U87 cells, leading to partial detachment from the substrate (Figure 8). Interestingly, in LN18 cells CUR+PLD treatment alone induced significant actin depolymerization and cell flattening. This effect on the actin network was even more pronounced when the synergistic combination was used before TMZ treatment, resulting in cells that appeared less adherent and partially elongated by SEM (Figure 9).
3. Discussion
Herbal preparations have been used since ancient times as the main source of therapeutic principles, and there are many remarkable examples, in the history of medicine, demonstrating how the discovery of natural products profoundly affected advances in biology and inspired drug discovery and therapy.
In the last decades, several naturally occurring dietary agents and a wide variety of products from plant sources have been recognized as promising candidates for developing chemopreventive or therapeutic drugs for cancer [4]. In particular, the anticancer activity of nature-derived polyphenols has been, and still is, the topic of numerous researches worldwide [36]. Among polyphenols, CUR, RES, and more recently its glycoside PLD have been extensively investigated in preclinical in vitro and in vivo studies and have entered some clinical trials [4,37,38]. Potential mechanisms underlying the antitumor efficacy of these nature-derived compounds reside in their antioxidant and anti-inflammatory properties, as well as in their ability to modulate molecular pathways involved in cancer [4,36].
In this work, we show that the pretreatment with a synergistic combination of CUR and PLD can increase the anti-tumor efficacy of TMZ in both GBL cell lines tested, as demonstrated by the results from the viability assays. However, the mechanism of action underlying this efficacy seems to be partly different in the two cell lines, which possess different intrinsic sensitivity to TMZ treatment, being one MGMT positive with epithelial morphological characteristics (LN18), and the other MGMT negative, with astroglial features (U87). Specifically, the first interesting result obtained was that the 24 h treatment (at T0 before TMZ addition) with the selected synergistic combination of CUR+PLD is able, per se, to reduce cell proliferation in both GBL cell lines tested, as demonstrated by the decreased levels of c-Myc. However, the effect seems to be mainly a consequence of astroglial differentiation in U87 cells, while it possibly derives from induction of apoptosis/autophagy in the intrinsically more resistant LN18 cells. These different effects exerted by the combination of the two compounds could be in line with the widely described effects of CUR, PLD as well as RES on molecular pathways involved in numerous biological processes, including cell viability and survival, proliferation, differentiation, and apoptosis, such as the Notch and the Wnt/β-catenin pathway [4,39,40]. These pathways are known to be differently dysregulated in tumors depending on cancer cells, and molecules targeting them can revert cancer cell malignity by affecting distinct processes.
Even more interesting results are those that demonstrate that the pretreatment with CUR+PLD significantly improves the effect of TMZ treatment on the number of live cells in both glioblastoma lines, and this effect is more evident in the intrinsically resistant LN18 cells, in which TMZ alone is ineffective. The data from the cytofluorimetric evaluation of autophagic flux, apoptosis, and cell cycle show that after 72 h of treatment with TMZ, in the presence or the absence of CUR+PLD pre-treatment, the autophagic flux is not modified in both cell lines, apoptosis induction is recorded for both experimental conditions only in LN18 cells, but a cell cycle arrest between the S and G2/M phases is recorded in both cell lines. It has been reported that TMZ can cause a shift of glioblastoma cells into G2/M, a prosurvival strategy that the cells adopt to have time for repairing the TMZ-induced DNA damage, and that the co-treatment with RES forces cells through mitosis leading to mitotic catastrophe and senescence, thus increasing the chronic effects of TMZ [41]. Our results from WB, suggest that this is probably what occurs in U87 cells subjected to CUR+PLD plus TMZ, in which, concomitantly to the recorded arrest in G2/M phase, the proliferation marker c-Myc and the autophagosomal marker LC3B-II are unexpectedly higher than in cells treated with TMZ alone. Instead, in CUR+PLD pretreated U87 cells, the increased c-Myc levels observed after 72 h washout period can be due to a gradual reversion of cellular differentiation towards the undifferentiated and proliferative baseline condition once the pharmacological pressure is removed, as supported by a decrease of the astroglial differentiation marker GFAP recorded at the same time point (data not shown). A different mechanism seems to be involved in LN18 cells pretreated with CUR+PLD plus TMZ, since the proliferation marker is lower than that in cells treated with TMZ alone, and the autophagy appears to be inhibited by the pre-treatment. This result suggests that PLD, similar to what was reported for the “twin” compound RES [42], suppresses, in LN18 cells, the cytoprotective autophagy consequent to TMZ-induced DNA damage, thus improving the cytotoxicity of the chemotherapeutic drug.
Finally, the preliminary data on the actin network suggests that the synergic combination could also improve the described effect of TMZ (and for other cytotoxic agents inducing DNA damage) on glioblastoma cell motility and invasiveness [35] and this effect is more marked in the intrinsically more resistant LN18 cells. Indeed, changes in the actin cytoskeleton dynamic are crucial in regulating cell shape and motility and are involved in cancer cell migration and tumor progression. It has been reported that CUR might inhibit cancer cell motility by altering actin polymerization, possibly through direct interaction with this cytoskeleton element [43,44]. Furthermore, PLD can repress glioblastoma cell migration and invasiveness by inhibiting the EGFR-AKT/ERK1/2/STAT3-SOX2/Snail signaling pathway [24]. Our preliminary data on the actin network support these CUR and PLD properties, and stimulate further studies focused on the effect of the synergic combination CUR+PLD on GLB cells migration and invasiveness.
4. Materials and Methods
4.1. Reagents
Temozolomide (TMZ, 3,4-dihydro-3-methyl-4-oxoimidazo [5,1-d]-astetrazine- 8-carboxamide) (Sigma-Aldrich, St. Louis, MO, USA; purity ≥98%, Cas: 85622-93-1), were dissolved in dimethyl sulfoxide (DMSO, Sigma-Aldrich), and stored as 20 mM stock solution. Curcumin extract (CUR) and Polydatin (PLD) were kindly donated by Sherman Tree Nutraceuticals S.r.l. (Rome, Italy). CUR contains more than 85% curcumin by weight and is specifically designed to enhance the water dispersibility of its bioactive compound; PLD has a purity of ≥98%. CUR and PLD were dissolved in DMSO and stored as a 10 mg/ml stock solution.
4.2. Cell Cultures
The human glioblastoma cell lines U87 and LN18 were originally obtained from the American Tissue Culture Collection (ATCC, Manassas, VA, USA). For each experiment, an aliquot of each cell line was revived and then maintained in culture for a maximum of 2 months before usage. Both U87 and LN18 cells were grown as monolayers in Dulbecco’s Modified Eagle Medium (DMEM) plus HAM’s F12 (1:1), supplemented with 10% heat-inactivated Fetal Bovine Serum (FBS), L-glutamine (2 mM), penicillin (100 IU/ml) and streptomycin (100 µg/ml), and maintained at 37°C, in a 5% CO2 humidified atmosphere. For passaging, cells were detached with 0.05% trypsin and 0.002% EDTA solution. All media and supplements for cell cultures were acquired from Gibco (Thermo Fisher Scientific, Inc., Waltham, Massachusetts, USA).
4.3. Cell Treatments and Cell Viability Analyses
In the preliminary dose-response experiments, U87 and LN18 cells were seeded in 96-well plates (5×103 cells/well), and treated after 24 h of adhesion with TMZ (from 50 to 400 µM), or with CUR (from 0.18 to 10 µg/ml) and PLD (from 17.5 to 300 µg/ml) alone or in combination, for 24, 48 and 72 h. At the end of the incubation period, the cells were incubated with 1 mg/ml 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) solution (Sigma-Aldrich) for 3 h at 37 °C. Supernatants were removed from the wells and formazan was solubilized with 150 μL DMSO. Absorbance was read at 570 nm using the Multiskan Labsystem microplate reader (Thermo Fischer Scientific).
To determine the synergistic combination of CUR and PLD, the CalcuSyn software (Biosoft v2.1) was used to calculate the combination index (CI). The concentrations of 10 µg/ml, for CUR, and 100 µg/ml, for PLD, respectively (ratio 1:10) were established as those that exhibited synergism in both cell lines (CI <0.9).
Treatments were performed following the scheme reported in Figure 2a. Briefly, both cell lines were pre-incubated with CUR+PLD for 24 h before the TMZ treatment, and analyzed after 24 h (T0 before TMZ treatment) and after additional 72 h for cell morphology and viability, and for the expression levels of MGMT and proteins related to cell proliferation, differentiation, apoptosis or autophagy. The effects on the actin network were also analyzed by confocal microscopy. The Trypan blue dye exclusion method, followed by the analysis using the automated cell counter CytoSmart (Corning) was used in all the experiments performed to determine cell viability after CUR+PLD plus TMZ combined treatments. CytoSmart was also used to automatically obtain a measure of cell size in treated and untreated samples, in parallel to cell count and viability.
4.4. Evaluation of Cell Morphology
Morphological features of treated and untreated cells were analyzed by phase contrast microscopy, using the Motic AE31 Trinocular inverted microscope (Motic Asia, Hong Kong). Scanning electron microscopy (SEM) was also used for analyzing ultrastructural features in control and treated cultures. In detail, for SEM analyses, U87 and LN18 cells were cultured on 12 mm glass coverslips and treated as schematized in Figure 2a. After 72 h TMZ treatment, the cells were fixed in 2.5% glutaraldehyde in 0.2 M Na-cacodylate buffer (pH 7.4) for 2 h at room temperature. Following three washes with the same buffer, the cells were post-fixed with 1% (w/w) OsO4 for 1 h, dehydrated through an ethanol gradient, and treated to dry by hexamethyldisilazane (HDMS, Sigma-Aldrich). The samples were coated with gold using a sputter coater and analyzed with the SEM FEI Quanta Inspect FeG (FEI Company, Eindhoven, The Netherlands).
4.5. Western Blot (WB) Analysis
Cells were lysed in RIPA lysis buffer containing 50 mM Tris/HCl, pH 8.0, 150 mM NaCl, 1% NP40 nonidet, 0.5% sodium deoxycholate, 0.1% SDS, 1 mM PMSF, 2 mM Na3VO4, 20 mM NaF and 1% protease inhibitor cocktail (Sigma-Aldrich). Lysates were clarified by centrifugation and protein content was determined using Bradford reagent (Bio-Rad, Segrate, Italy). 15-20 µg of each cell extract was separated by 10-15% SDS/PAGE, transferred to a nitrocellulose membrane (Hybond, Amersham GE Healthcare), blocked with 5% non-fat milk (Bio-Rad) in Tris-buffered saline-Tween (TBS-T; 20 mM Tris, 150 mM NaCl, pH 7. 6 and 0.05% Tween-20) and probed with the primary antibodies against MGMT, c-Myc, LC3B, PARP, GFAP, detailed in Supplementary Table S3. Primary antibodies were detected with peroxidase-conjugated secondary antibodies (Bio-Rad, Richmond, CA, United States). The ChemiDoc XRS+ imager (Bio-Rad) was used for membrane exposure and image acquisition. The densities of the bands were quantified by the ImageJ processing program [http://rsbweb.nih.gov/ij/]. Values, normalized to GAPDH, were reported as uncalibrated Optical Density (OD) or fold vs untreated control. Results were from at least three independent experiments and presented as the mean ± SD.
4.6. Confocal Laser Scanning Microscopy (CLSM)
For confocal microscopic analysis of the actin network, cells were grown on the ibiTreat µ-Slide 4 well (Ibidi GmbH, Germany). After treatments performed following the scheme reported in Figure 2a, cells were fixed with 2% paraformaldehyde (Sigma-Aldrich), permeabilized with 0.2% Triton X-100 (Sigma-Aldrich), and stained with Alexa Fluor 546 conjugated phalloidin (Thermo Fisher Scientific; working dilution 1:40) Nuclei were counterstained with Hoechst 33285 (Fluka Biochemika, Buchs, Switzerland, working dilution 1:3000). The analyses were performed using the Stellaris 5 confocal microscope form LEICA Microsystems (Wetzlar, Germany).
4.7. Cytofluorimetric Analyses of Cell Cycle, Apoptosis, and Autophagy
Cell cycle analysis was analyzed in U87 and LN18 cells after 48 and 72 h of TMZ treatment in the absence or the presence of CUR+PLD pretreatment. At these time points, the cells were collected, washed with cold PBS, and fixed in 70% ethanol for 1 h at 4°C. After washing with cold PBS, cells were resuspended in 40 µg/mL propidium iodide (PI) and 100 µg/mL RNAse in PBS, at 37°C for 1 h, and the percentages of cells in the different phases were determined by flow cytometry.
The assessment of apoptosis was performed using a fluorescein isothiocyanate (FITC)-conjugated annexin V (AV) and PI detection kit according to the manufacturer’s protocol (BioViosion Incorporated, Milpitas, CA, USA). Untreated controls and treated cells were collected after 72 h, washed with PBS, and resuspended at 2.5x 105cells/mL. Subsequently, 5 μL annexin V-FITC solution and an equal volume of PI solution were added to 500 μL cell suspension, incubated for 5 min at room temperature in the dark, and analyzed by flow cytometry.
The monitoring of autophagic flux was performed on live cells using the Cyto-ID Autophagy Detection Kit (Enzo Life Sciences, NY, USA), according to the manufacturer’s instructions. Briefly, untreated controls and treated cells were collected after 72 h, stained with the Cyto-ID staining solution for 30 minutes at 37 °C, and the fluorescence intensities were determined by flow cytometry. For all the analyses, samples were analyzed using a flow cytometry instrument (Gallios Instrument, Beckman Coulter, Brea, CA, USA), and at least 10,000 events were run per sample. Data were analyzed using Kaluza software for analysis v. 2.2 (Beckman Coulter).
4.8. Statistical Analysis
Statistical analysis was performed using the two-tailed Student’s t-test or one-way analysis of variance (ANOVA) followed by the Tukey post-hoc test used to analyze the differences between groups. A p-value < 0.05 was assumed as statistically significant. All results were from at least three independent experiments and presented as means ± SD.
5. Conclusions
The results obtained in this work provide convincing evidence that CUR and PLD, by acting in synergy with each other, combine their abilities to affect several processes crucial for the viability, survival, differentiation, and motility of tumor cells, and might strongly improve the efficacy of alkylating antitumor agents such as TMZ, also in drug-resistant cells, and are stimulating for further studies to deepen some aspects of the described effects. The data also provide experimental evidence on the mechanisms through which the synergic combination CUR+PLD can exert its valuable effects on glioblastoma patients [31], supporting the use of this orodispersible nutraceutical formulation in integrated anticancer therapies. Due to the different mechanisms of action adopted in GBL cells exhibiting different phenotypical features and different TMZ-sensitivity, the use of CUR+PLD as an integrated therapy might be beneficial in a poorly differentiated and multifaceted tumor such as glioblastoma multiforme. Owing the demonstrated ability of CUR+PLD in improving TMZ cytotoxicity in intrinsically resistant MGMT+ cells, the synergic combination of these two natural molecules could be also beneficial in other tumors in which the treatment with alkylating agents often are ineffective because of the presence of high levels of MGMT, such as pancreatic tumor. Actually, from our clinical preliminary data, it seems that the CUR+PLD combination also leads to positive results for pancreatic cancer, and this is therefore already the subject of our scientific and clinical investigations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. PDF file including: Figure S1: Representative data from CalcuSyn software used to define the synergic concentrations of CUR and PLD in U87 and LN18 glioblastoma cells; Table S1: Data from cytofluorometric analysis of cell cycle U87 and LN18 glioblastoma cells; Table S2: Data from cytofluorometric analysis of apoptosis/necrosis by Annexin V and propidium iodide staining in U87 and LN18 glioblastoma cells; Table S3: List of antibodies/reagents used for Western blot analyses.
Author Contributions
Conceptualization, A.S. and MP.F.; methodology, E.K.K., S.R., A.DG., M.C. and ML.D.; formal analysis, A.S., E.K.K., S.R., A.DG., M.C. and ML.D.; investigation, E.K.K. and S.R.; data curation, A.S. and MP.F.; writing—original draft preparation, A.S., E.K.K. and S.R.; writing—review and editing, A.S., M.B., G.R., A.S. and MP.F.; supervision, A.S., E.K.K., A.S. and MP.F.; project administration, A.S. and MP.F.; funding acquisition, G.R.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-financed by ShermanTree Nutraceuticals and by MIMIT (Ministry of Enterprise and Made in Italy), Project Invitalia Brevetti+; GRANT n° BRE0000830. The instrumentation used for cell maintenance and confocal microscopic analysis is part of infrastructure financed to IFT-CNR by the European Union - Next Generation EU, within the PNRR project “Rome Technopole - Innovation Ecosystem”, ECS00000024, CUP B83C22002890005.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data produced in this study are reported in this article. The primary data files are available from the corresponding author upon reasonable request.
Acknowledgments
The Authors would like to thank Dr Massimiliano Romiti, Chief Executive Officer of ShermanTree Nutraceuticals s.r.l., for economically supporting and enthusiastically encouraging our research.
Conflicts of Interest
Ravagnan G. & Bonucci M. are Inventors in the Patent Applications WO-2020026185-A1 and PCT/IB2019/056565. The other Authors declare no conflicts of interest.
References
- Esteller, M.; Garcia-Foncillas, J.; Andion, E.; Goodman, S. N.; Hidalgo, O. F.; Vanaclocha, V.; Baylin, S. B.; Herman, J. G., Inactivation of the DNA-repair gene MGMT and the clinical response of gliomas to alkylating agents. The New England journal of medicine 2000, 343 (19), 1350-4. [CrossRef]
- Hegi, M.E.; Diserens, A.-C.; Godard, S.; Dietrich, P.-Y.; Regli, L.; Ostermann, S.; Otten, P.; Van Melle, G.; de Tribolet, N.; Stupp, R. Clinical Trial Substantiates the Predictive Value of O-6-Methylguanine-DNA Methyltransferase Promoter Methylation in Glioblastoma Patients Treated with Temozolomide. Clinical cancer research: an official journal of the American Association for Cancer Research 2004, 10, 1871–1874. [CrossRef]
- Tan, A.C.; Ashley, D.M.; López, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of glioblastoma: State of the art and future directions. CA Cancer J. Clin. 2020, 70, 299–312. [CrossRef]
- Sferrazza G, C. M., Brusotti G, Pierimarchi P, Temporini C, Serafino A, Calleri E, Nature-derived compounds modulating Wnt/β-catenin pathway: a preventive and therapeutic opportunity in neoplastic diseases. Acta Pharmaceutica Sinica B 2020, 10 (10), 1814-1834. [CrossRef]
- Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Ramirez-Tortosa, M., Curcumin and Health. Molecules 2016, 21 (3), 264. [CrossRef]
- Abrahams, S.; Haylett, W.L.; Johnson, G.; Carr, J.A.; Bardien, S. Antioxidant effects of curcumin in models of neurodegeneration, aging, oxidative and nitrosative stress: A review. Neuroscience 2019, 406, 1–21. [CrossRef]
- Kahkhaie, K.R.; Mirhosseini, A.; Aliabadi, A.; Mohammadi, A.; Mousavi, M.J.; Haftcheshmeh, S.M.; Sathyapalan, T.; Sahebkar, A. Curcumin: a modulator of inflammatory signaling pathways in the immune system. Inflammopharmacology 2019, 27, 885–900. [CrossRef]
- Ferreira, N.; Saraiva, M.J.; Almeida, M.R. Uncovering the Neuroprotective Mechanisms of Curcumin on Transthyretin Amyloidosis. Int. J. Mol. Sci. 2019, 20, 1287. [CrossRef]
- Prasad, S.; Tyagi, A.K. Curcumin and its analogues: a potential natural compound against HIV infection and AIDS. Food Funct. 2015, 6, 3412–3419. [CrossRef]
- Zhang, D.; Yang, Y.; Yao, B.; Hu, T.; Ma, Z.; Shi, W.; Ye, Y. Curcumin inhibits Aspergillus flavus infection and aflatoxin production possibly by inducing ROS burst. Food Res. Int. 2023, 167, 112646. [CrossRef]
- Willenbacher, E.; Khan, S.Z.; Mujica, S.C.A.; Trapani, D.; Hussain, S.; Wolf, D.; Willenbacher, W.; Spizzo, G.; Seeber, A. Curcumin: New Insights into an Ancient Ingredient against Cancer. Int. J. Mol. Sci. 2019, 20, 1808. [CrossRef]
- Luthra, P.M.; Lal, N. Prospective of curcumin, a pleiotropic signalling molecule from Curcuma longa in the treatment of Glioblastoma. Eur. J. Med. Chem. 2016, 109, 23–35. [CrossRef]
- Dhandapani, K. M.; Mahesh, V. B.; Brann, D. W., Curcumin suppresses growth and chemoresistance of human glioblastoma cells via AP-1 and NFkappaB transcription factors. Journal of neurochemistry 2007, 102 (2), 522-38. [CrossRef]
- Du, Q.-H.; Peng, C.; Zhang, H. Polydatin: A review of pharmacology and pharmacokinetics. Pharm. Biol. 2013, 51, 1347–1354. [CrossRef]
- Ye, P.; Wu, H.; Jiang, Y.; Xiao, X.; Song, D.; Xu, N.; Ma, X.; Zeng, J.; Guo, Y. Old dog, new tricks: Polydatin as a multitarget agent for current diseases. Phytotherapy Res. 2021, 36, 214–230. [CrossRef]
- Fabris, S.; Momo, F.; Ravagnan, G.; Stevanato, R. Antioxidant properties of resveratrol and piceid on lipid peroxidation in micelles and monolamellar liposomes. Biophys. Chem. 2008, 135, 76–83. [CrossRef]
- Li, Z.; Chen, X.; Liu, G.; Li, J.; Zhang, J.; Cao, Y.; Miao, J. Antioxidant Activity and Mechanism of Resveratrol and Polydatin Isolated from Mulberry (Morus alba L.). Molecules 2021, 26, 7574. [CrossRef]
- Lanzilli, G.; Cottarelli, A.; Nicotera, G.; Guida, S.; Ravagnan, G.; Fuggetta, M.P. Anti-inflammatory Effect of Resveratrol and Polydatin by In Vitro IL-17 Modulation. Inflammation 2011, 35, 240–248. [CrossRef]
- Ravagnan, G.; De Filippis, A.; Carteni, M.; De Maria, S.; Cozza, V.; Petrazzuolo, M.; Tufano, M. A.; Donnarumma, G., Polydatin, a natural precursor of resveratrol, induces beta-defensin production and reduces inflammatory response. Inflammation 2013, 36 (1), 26-34. [CrossRef]
- Fuggetta, M.; Mattivi, F. The immunomodulating activities of resveratrol glucosides in humans.. Recent Patents Food, Nutr. Agric. 2011, 3, 81–90. [CrossRef]
- De Maria, S.; Scognamiglio, I.; Lombardi, A.; Amodio, N.; Caraglia, M.; Cartenì, M.; Ravagnan, G.; Stiuso, P. Polydatin, a natural precursor of resveratrol, induces cell cycle arrest and differentiation of human colorectal Caco-2 cell. J. Transl. Med. 2013, 11, 264–264. [CrossRef]
- Jin, Y.; Zhan, X.; Zhang, B.; Chen, Y.; Liu, C.; Yu, L., Polydatin Exerts an Antitumor Effect Through Regulating the miR-382/PD-L1 Axis in Colorectal Cancer. Cancer Biother Radiopharm 2020, 35 (2), 83-91. [CrossRef]
- De Gregorio, A.; Krasnowska, E.K.; Zonfrillo, M.; Ravagnan, G.; Bordignon, V.; Bonmassar, E.; Fuggetta, M.P. Influence of Polydatin on the Tumor Microenvironment In Vitro: Studies with a Colon Cancer Cell Model. Int. J. Mol. Sci. 2022, 23, 8442. [CrossRef]
- Chen, Y.; Niu, J.; Li, L.; Li, Z.; Jiang, J.; Zhu, M.; Dong, T.; Zhang, J.; Shi, C.; Xu, P.; Lu, Y.; Jiang, Y.; Liu, P.; Chen, W., Polydatin executes anticancer effects against glioblastoma multiforme by inhibiting the EGFR-AKT/ERK1/2/STAT3-SOX2/Snail signaling pathway. Life sciences 2020, 258, 118158. [CrossRef]
- Klinger, N.V.; Mittal, S. Therapeutic Potential of Curcumin for the Treatment of Brain Tumors. Oxidative Med. Cell. Longev. 2016, 2016, 9324085. [CrossRef]
- DU, Q.; Hu, B.; An, H.-M.; Shen, K.-P.; Xu, L.; Deng, S.; Wei, M.-M. Synergistic anticancer effects of curcumin and resveratrol in Hepa1-6 hepatocellular carcinoma cells. Oncol. Rep. 2013, 29, 1851–1858. [CrossRef]
- Majumdar, A.P.N.; Banerjee, S.; Nautiyal, J.; Patel, B.B.; Patel, V.; Du, J.; Yu, Y.; Elliott, A.A.; Levi, E.; Sarkar, F.H. Curcumin Synergizes With Resveratrol to Inhibit Colon Cancer. Nutr. Cancer 2009, 61, 544–553. [CrossRef]
- Wang, P.; Wu, Q.; Peng, Q.; Kang, Z.; Xiao, S.; Zheng, P.; Li, J.; Chen, Y. Comparative analysis of the molecular mechanism of inhibiting proliferation and migration in cervical cancer HeLa cell by curcumin and resveratrol. Nat. Prod. Res. 2023, 37, 4032–4037. [CrossRef]
- Focaccetti, C.; Palumbo, C.; Benvenuto, M.; Carrano, R.; Melaiu, O.; Nardozi, D.; Angiolini, V.; Lucarini, V.; Kërpi, B.; Masuelli, L.; et al. The Combination of Bioavailable Concentrations of Curcumin and Resveratrol Shapes Immune Responses While Retaining the Ability to Reduce Cancer Cell Survival. Int. J. Mol. Sci. 2023, 25, 232. [CrossRef]
- Muhanmode, Y.; Wen, M.K.; Maitinuri, A.; Shen, G. Curcumin and resveratrol inhibit chemoresistance in cisplatin-resistant epithelial ovarian cancer cells via targeting P13K pathway. Hum. Exp. Toxicol. 2022, 41. [CrossRef]
- Ravagnan, G.; Bonucci, M. Composition Comprising Resveratrolosides and Curcumins. WO2020026185-A1, February 2020, 2020.
- Bordin, D.L.; Lima, M.; Lenz, G.; Saffi, J.; Meira, L.B.; Mésange, P.; Soares, D.G.; Larsen, A.K.; Escargueil, A.E.; Henriques, J.A. DNA alkylation damage and autophagy induction. Mutat. Res. Mol. Mech. Mutagen. 2013, 753, 91–99. [CrossRef]
- Pollard, T.D.; Cooper, J.A. Actin, a Central Player in Cell Shape and Movement. Science 2009, 326, 1208–1212. [CrossRef]
- Kitambi, S.S.; Masoumi, S.; Harishankar, A.; Gracias, A.; Bachinger, F.; Fufa, T.; Chandrasekar, G.; Gaunitz, F.; Walfridsson, J. Understanding cytoskeleton regulators in glioblastoma multiforme for therapy design. Drug Des. Dev. Ther. 2016, ume 10, 2881–2897. [CrossRef]
- Avci, N. G.; Ebrahimzadeh-Pustchi, S.; Akay, Y. M.; Esquenazi, Y.; Tandon, N.; Zhu, J. J.; Akay, M., NF-kappaB inhibitor with Temozolomide results in significant apoptosis in glioblastoma via the NF-kappaB(p65) and actin cytoskeleton regulatory pathways. Scientific reports 2020, 10 (1), 13352. [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [CrossRef]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: multitargeting for multiple chronic diseases. Br. J. Pharmacol. 2017, 174, 1325–1348. [CrossRef]
- Kotecha, R.; Takami, A.; Espinoza, J.L. Dietary phytochemicals and cancer chemoprevention: a review of the clinical evidence. Oncotarget 2016, 7, 52517–52529. [CrossRef]
- Cai, J.; Qiao, Y.; Chen, L.; Lu, Y.; Zheng, D. Regulation of the Notch signaling pathway by natural products for cancer therapy. J. Nutr. Biochem. 2024, 123, 109483. [CrossRef]
- Chen, X. J.; Shen, Y. S.; He, M. C.; Yang, F.; Yang, P.; Pang, F. X.; He, W.; Cao, Y. M.; Wei, Q. S., Polydatin promotes the osteogenic differentiation of human bone mesenchymal stem cells by activating the BMP2-Wnt/beta-catenin signaling pathway. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2019, 112, 108746. [CrossRef]
- Filippi-Chiela, E.C.; Thomé, M.P.; e Silva, M.M.B.; Pelegrini, A.L.; Ledur, P.F.; Garicochea, B.; Zamin, L.L.; Lenz, G. Resveratrol abrogates the Temozolomide-induced G2 arrest leading to mitotic catastrophe and reinforces the Temozolomide-induced senescence in glioma cells. BMC Cancer 2013, 13, 147–147. [CrossRef]
- Lin, C.-J.; Lee, C.-C.; Shih, Y.-L.; Lin, T.-Y.; Wang, S.-H.; Lin, Y.-F.; Shih, C.-M. Resveratrol enhances the therapeutic effect of temozolomide against malignant glioma in vitro and in vivo by inhibiting autophagy. Free. Radic. Biol. Med. 2011, 52, 377–391. [CrossRef]
- Holy, J. Curcumin inhibits cell motility and alters microfilament organization and function in prostate cancer cells. Cell Motil. Cytoskelet. 2004, 58, 253–268. [CrossRef]
- Dhar, G.; Chakravarty, D.; Hazra, J.; Dhar, J.; Poddar, A.; Pal, M.; Chakrabarti, P.; Surolia, A.; Bhattacharyya, B. Actin–Curcumin Interaction: Insights into the Mechanism of Actin Polymerization Inhibition. Biochemistry 2015, 54, 1132–1143. [CrossRef]
Figure 1.
Features of cell lines used (a, b) and dose- and time-dependent response to TMZ in U87 and LN18 cells (c). (a) Phase contrast microscopy (top panels) and scanning electron microscopy (bottom panels) illustrating the morphological features of U87 and LN18 glioblastoma cell lines. The scale bars represent 100 μm. (b) Baseline expression by WB of the DNA repair protein O6-methylguanine-DNA-methyltransferase (MGMT), in U87 and LN18 cells. GAPDH was used as the loading control. (c) Time- and dose-response experiments performed by MTT assay, for defining the optimal condition for TMZ treatment on U87 and LN18 cells. Results are the mean ± SD from three independent experiments (n = 3). Significance vs untreated control (two-tailed Student’s t-test): *p < 0.05; **p < 0.01; ***p < 0.001.
Figure 1.
Features of cell lines used (a, b) and dose- and time-dependent response to TMZ in U87 and LN18 cells (c). (a) Phase contrast microscopy (top panels) and scanning electron microscopy (bottom panels) illustrating the morphological features of U87 and LN18 glioblastoma cell lines. The scale bars represent 100 μm. (b) Baseline expression by WB of the DNA repair protein O6-methylguanine-DNA-methyltransferase (MGMT), in U87 and LN18 cells. GAPDH was used as the loading control. (c) Time- and dose-response experiments performed by MTT assay, for defining the optimal condition for TMZ treatment on U87 and LN18 cells. Results are the mean ± SD from three independent experiments (n = 3). Significance vs untreated control (two-tailed Student’s t-test): *p < 0.05; **p < 0.01; ***p < 0.001.
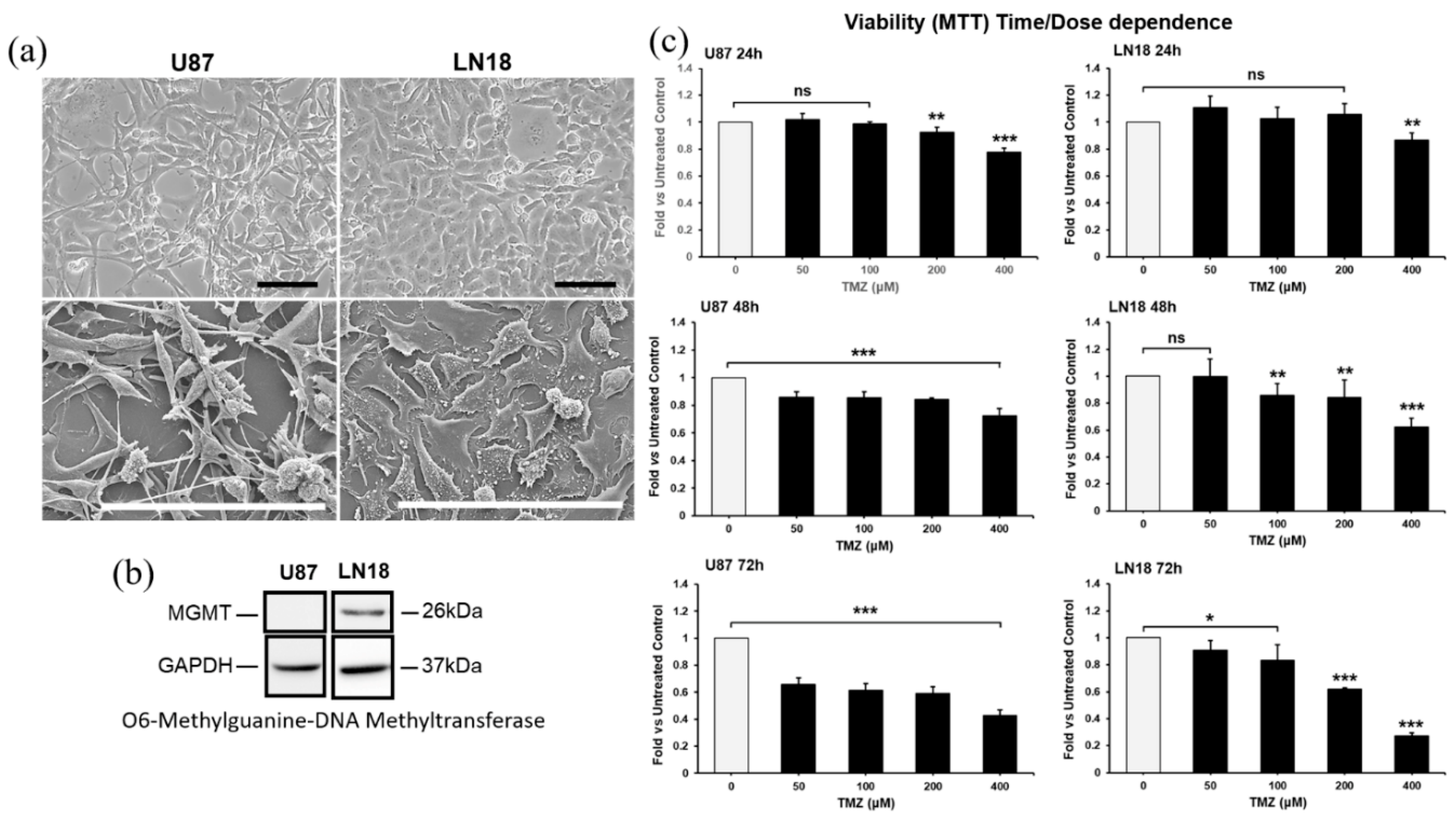
Figure 2.
Experimental scheme (a) and MTT cell viability assay (b, c) used for the preliminary assessment of the efficacy of the selected synergic combination of CUR+PLD. (a) Scheme of the protocol used for evaluating the effects of treatments on morphology, viability, and protein expression in both cell lines, as detailed in the Method section. (b, c) Viability assay performed by MTT on U87 and LN18 subjected to TMZ treatments in the presence or absence of CUR and PLD single or combined pretreatments. Values are the mean ± SD from three independent experiments (n = 3). Significance vs untreated control (two-tailed Student’s t-test): ***p < 0.001.
Figure 2.
Experimental scheme (a) and MTT cell viability assay (b, c) used for the preliminary assessment of the efficacy of the selected synergic combination of CUR+PLD. (a) Scheme of the protocol used for evaluating the effects of treatments on morphology, viability, and protein expression in both cell lines, as detailed in the Method section. (b, c) Viability assay performed by MTT on U87 and LN18 subjected to TMZ treatments in the presence or absence of CUR and PLD single or combined pretreatments. Values are the mean ± SD from three independent experiments (n = 3). Significance vs untreated control (two-tailed Student’s t-test): ***p < 0.001.
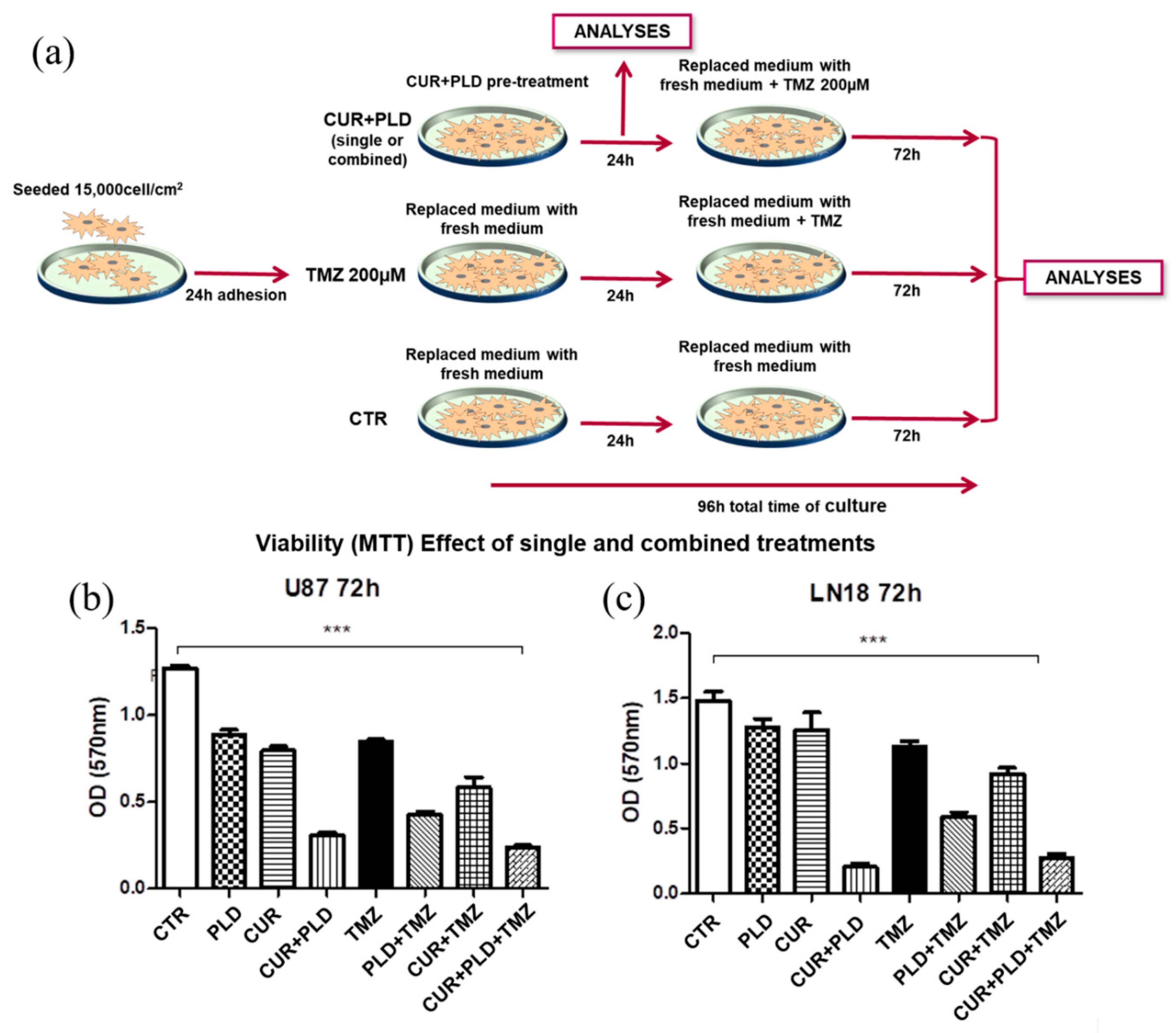
Figure 3.
Effects of the synergic combination of CUR+PLD before TMZ treatment on U87 and LN18 cells. (a, c) Phase contrast microscopy showing the morphological changes induced by 24 h CUR+PLD treatment on U87 (a) and LN18 (c) cells. The scale bars represent 100 μm. (b, d, e) WB and analysis of modifications induced by 24 h CUR+PLD treatment on expression levels of the proliferation marker c-Myc, the astroglial differentiation marker GFAP, the autophagosomal marker LC3B and the maker of caspase-dependent apoptosis PARP. Histograms in (b, d) report the densitometric analysis for GFAP, c-Myc and LC3B expressions; values were normalized to GAPDH. Results are the mean ± SD from three independent experiments (n = 3). Significance vs control (two-tailed Student’s t test): *, p ˂ 0.05; **, p ˂ 0.01; ns: not significant.
Figure 3.
Effects of the synergic combination of CUR+PLD before TMZ treatment on U87 and LN18 cells. (a, c) Phase contrast microscopy showing the morphological changes induced by 24 h CUR+PLD treatment on U87 (a) and LN18 (c) cells. The scale bars represent 100 μm. (b, d, e) WB and analysis of modifications induced by 24 h CUR+PLD treatment on expression levels of the proliferation marker c-Myc, the astroglial differentiation marker GFAP, the autophagosomal marker LC3B and the maker of caspase-dependent apoptosis PARP. Histograms in (b, d) report the densitometric analysis for GFAP, c-Myc and LC3B expressions; values were normalized to GAPDH. Results are the mean ± SD from three independent experiments (n = 3). Significance vs control (two-tailed Student’s t test): *, p ˂ 0.05; **, p ˂ 0.01; ns: not significant.
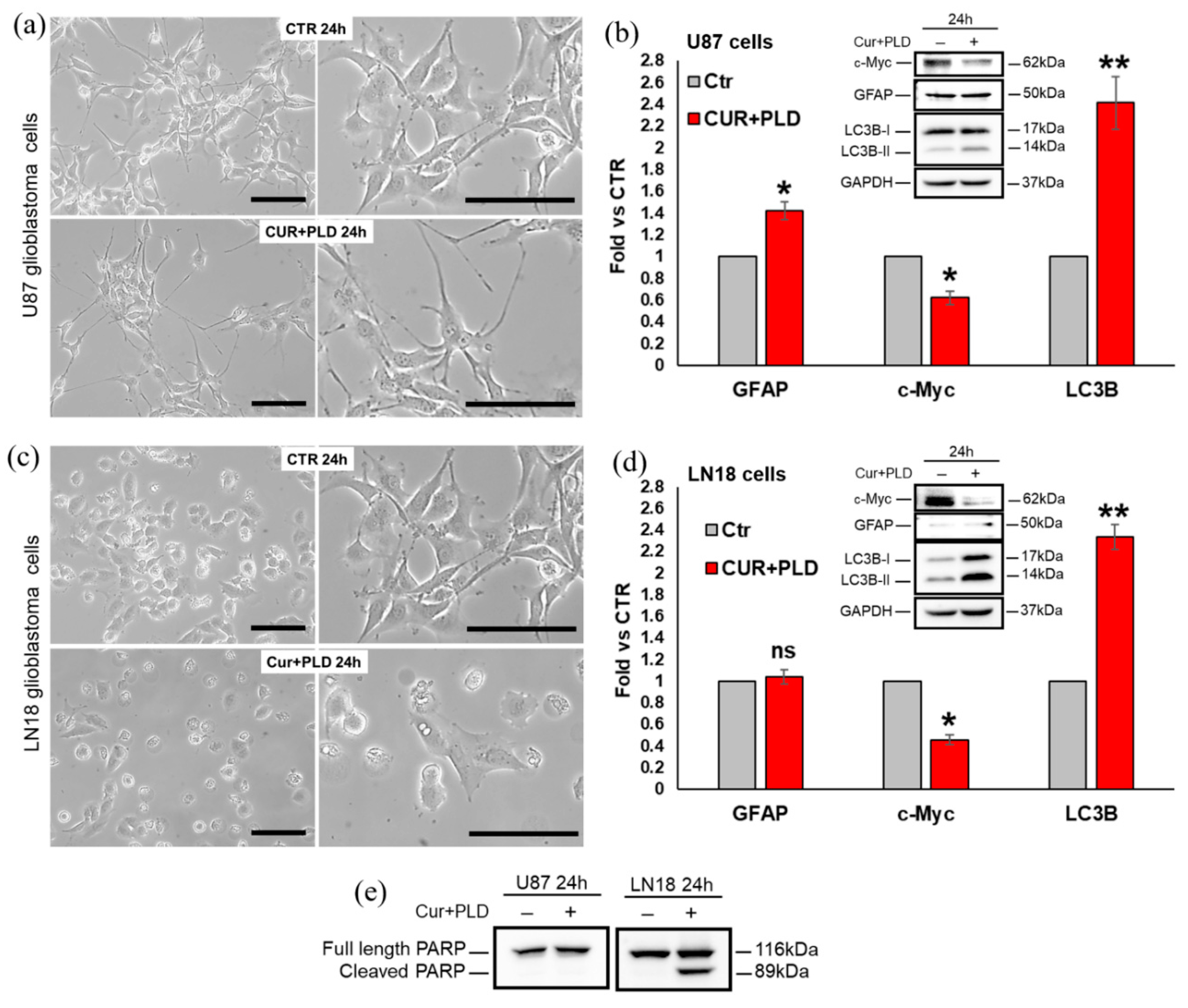
Figure 4.
Effects of the pretreatment with the synergic combination of CUR+PLD on TMZ-induced effects on U87 cells. (a) Phase contrast microscopy showing the morphological modification induced in untreated controls and in samples pre-treated for 24 h with CUR+PLD post-treated or not with TMZ. Arrows point to rare cells displaying morphological features suggestive of apoptosis/autophagy. The scale bars represent 100 μm. (b-e) Analyses by the automated cell counter CytoSmart of the number of live cells (b), cell growth between T0 and T 96h (c), percent of the viable cell (d) and cell size. Results are the mean ± SD from three independent experiments (n = 3). Significance (One-way ANOVA + Tukey multiple comparison test): *vs untreated control (CTR), #vs TMZ alone or pretreated with CUR+PLD: **p ˂ 0.01; ***p ˂ 0.001; ns: not significant.
Figure 4.
Effects of the pretreatment with the synergic combination of CUR+PLD on TMZ-induced effects on U87 cells. (a) Phase contrast microscopy showing the morphological modification induced in untreated controls and in samples pre-treated for 24 h with CUR+PLD post-treated or not with TMZ. Arrows point to rare cells displaying morphological features suggestive of apoptosis/autophagy. The scale bars represent 100 μm. (b-e) Analyses by the automated cell counter CytoSmart of the number of live cells (b), cell growth between T0 and T 96h (c), percent of the viable cell (d) and cell size. Results are the mean ± SD from three independent experiments (n = 3). Significance (One-way ANOVA + Tukey multiple comparison test): *vs untreated control (CTR), #vs TMZ alone or pretreated with CUR+PLD: **p ˂ 0.01; ***p ˂ 0.001; ns: not significant.
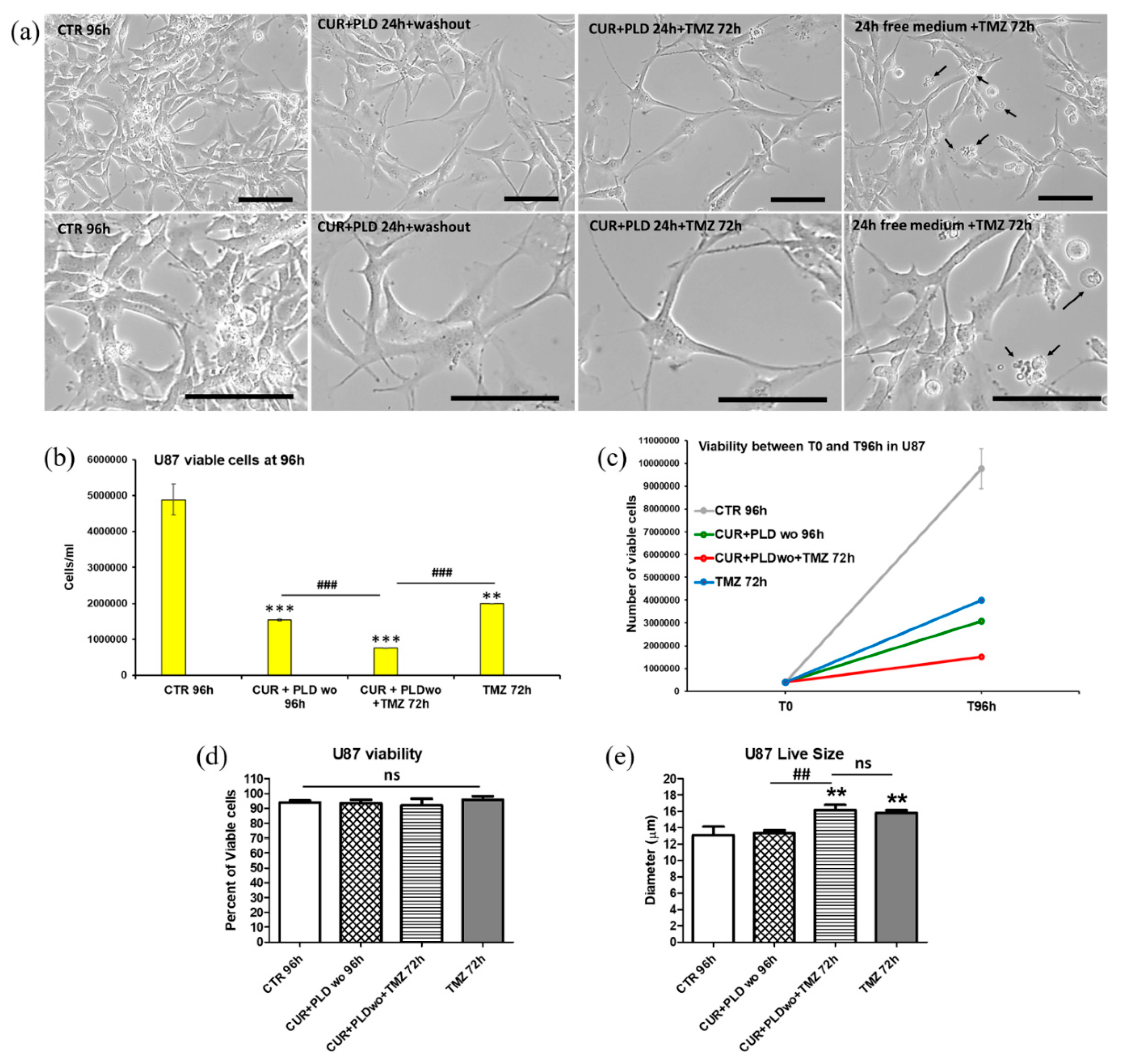
Figure 5.
Effects of the pretreatment with the synergic combination of CUR+PLD on TMZ-induced effects on LN18 cells. (a) Phase contrast microscopy showing the morphological modification induced in untreated controls and in samples pre-treated for 24 h with CUR+PLD post-treated or not with TMZ. Arrows point to numerous cells displaying morphological features suggestive of apoptosis/autophagy. The scale bars represent 100 μm. (b-e) Analyses by the automated cell counter CytoSmart of the number of live cells (b), cell growth between T0 and T 96h (c), percent of the viable cell (d) and cell size. Results are the mean ± SD from three independent experiments (n = 3). Significance (One-way ANOVA + Tukey multiple comparison test): *vs untreated control (CTR), #vs TMZ alone or pretreated with CUR+PLD: *p ˂ 0.05; **p ˂ 0.01; ***p ˂ 0.001; ns: not significant.
Figure 5.
Effects of the pretreatment with the synergic combination of CUR+PLD on TMZ-induced effects on LN18 cells. (a) Phase contrast microscopy showing the morphological modification induced in untreated controls and in samples pre-treated for 24 h with CUR+PLD post-treated or not with TMZ. Arrows point to numerous cells displaying morphological features suggestive of apoptosis/autophagy. The scale bars represent 100 μm. (b-e) Analyses by the automated cell counter CytoSmart of the number of live cells (b), cell growth between T0 and T 96h (c), percent of the viable cell (d) and cell size. Results are the mean ± SD from three independent experiments (n = 3). Significance (One-way ANOVA + Tukey multiple comparison test): *vs untreated control (CTR), #vs TMZ alone or pretreated with CUR+PLD: *p ˂ 0.05; **p ˂ 0.01; ***p ˂ 0.001; ns: not significant.
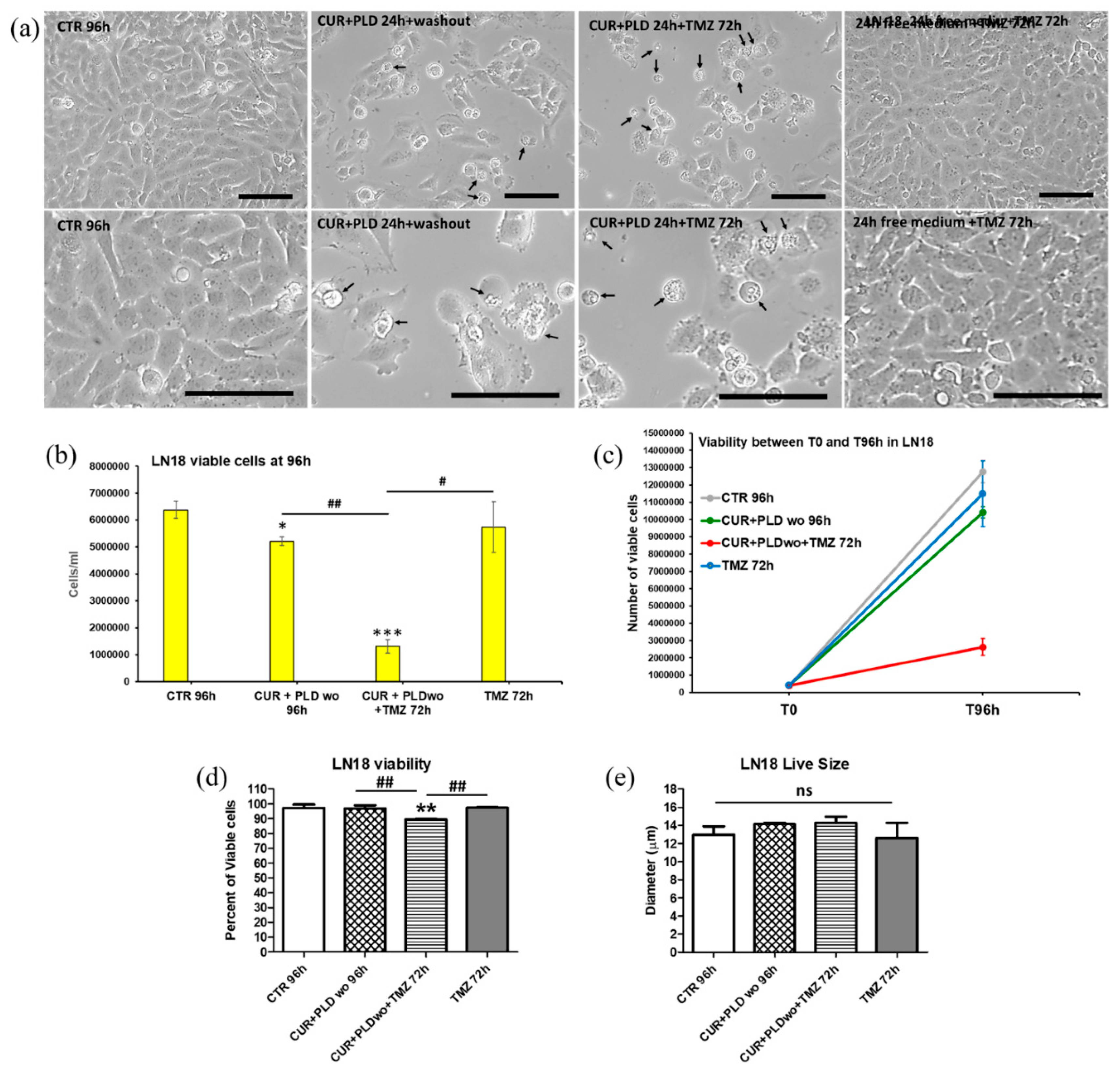
Figure 6.
Effects of the TMZ treatment with or without pretreatment with CUR+PLD on cell cycle, autophagic flux, and apoptosis. (a, b) The bar graphs illustrate the cytofluorimetric analysis of cell cycle in U87 (a) and LN18 (b) cells between 48 and 72h of culture. (c, d) Cytofluorimetric analysis of autophagy performed by the Cyto-ID autophagy detection kit after 72 h TMZ challenging in U87 (c) and LN18 (d) cells; ns: not significant. (e, f) Cytofluorimetric evaluation of apoptosis by Annexin V (AV) staining in U87 (e) and LN18 (f) cells; PI–/AV+, PI+/AV+, PI+/AV– counts identified the early apoptotic, late apoptotic, and necrotic cells, respectively.
Figure 6.
Effects of the TMZ treatment with or without pretreatment with CUR+PLD on cell cycle, autophagic flux, and apoptosis. (a, b) The bar graphs illustrate the cytofluorimetric analysis of cell cycle in U87 (a) and LN18 (b) cells between 48 and 72h of culture. (c, d) Cytofluorimetric analysis of autophagy performed by the Cyto-ID autophagy detection kit after 72 h TMZ challenging in U87 (c) and LN18 (d) cells; ns: not significant. (e, f) Cytofluorimetric evaluation of apoptosis by Annexin V (AV) staining in U87 (e) and LN18 (f) cells; PI–/AV+, PI+/AV+, PI+/AV– counts identified the early apoptotic, late apoptotic, and necrotic cells, respectively.
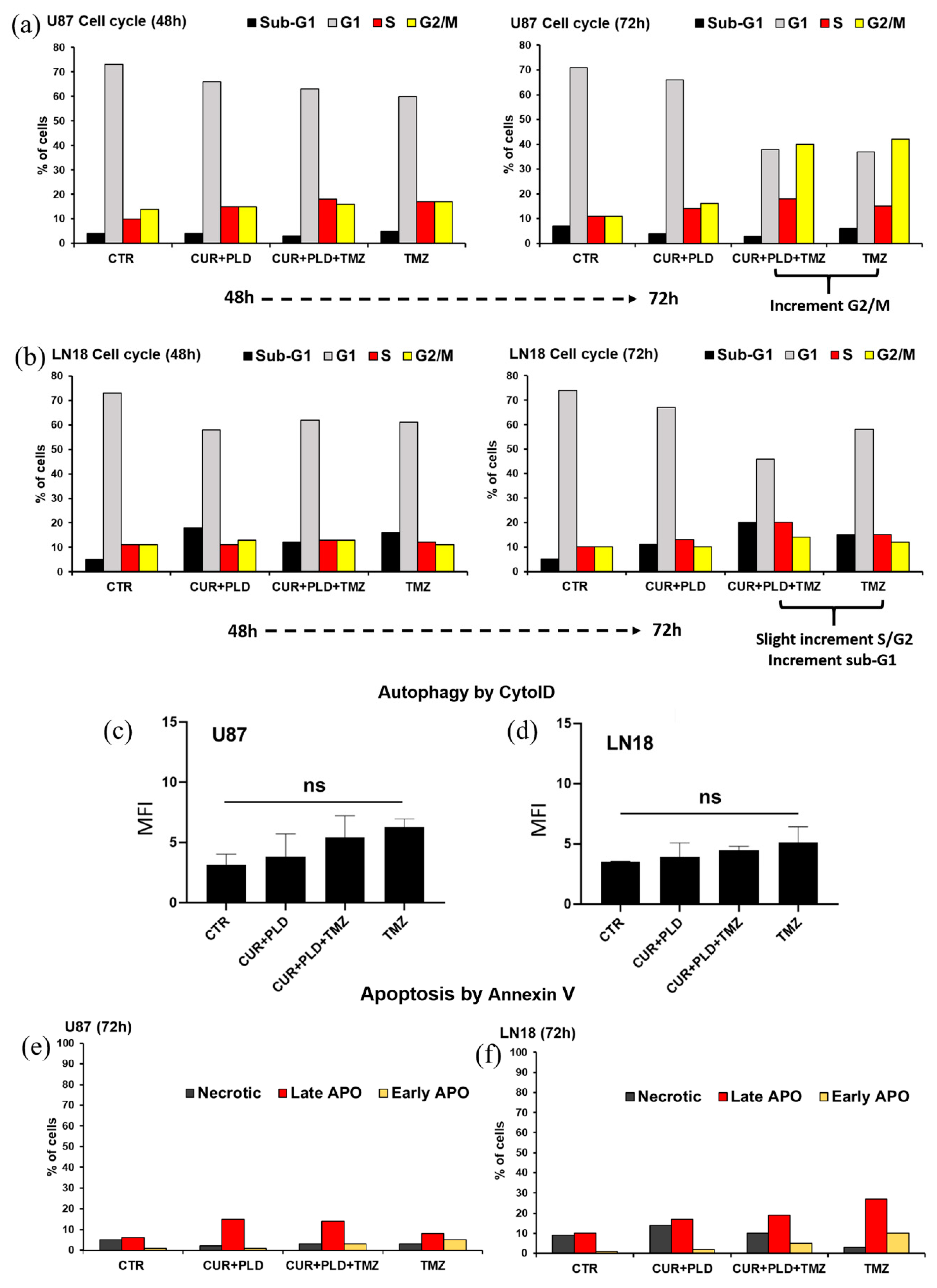
Figure 7.
Effects on MGMT and proteins related to cell proliferation, apoptosis, or autophagy. Western Blot (a, b) and densitometric analysis (c, d) of the expression levels of MGMT, the proliferation marker c-Myc, the astroglial differentiation marker GFAP, the autophagosomal marker LC3B, and the maker of caspase-dependent apoptosis PARP, evaluated after 72 h TMZ treatment with or without pretreatment with CUR+PLD, in U87 (a, c) and LN18 (b, d) cells; values were normalized to GAPDH. Results are the mean ± SD from three independent experiments (n = 3). Significance (One-way ANOVA + Tukey multiple comparison test): *vs untreated control (CTR), #vs TMZ alone or pretreated with CUR+PLD: *p ˂ 0.05; **p ˂ 0.01; ns: not significant.
Figure 7.
Effects on MGMT and proteins related to cell proliferation, apoptosis, or autophagy. Western Blot (a, b) and densitometric analysis (c, d) of the expression levels of MGMT, the proliferation marker c-Myc, the astroglial differentiation marker GFAP, the autophagosomal marker LC3B, and the maker of caspase-dependent apoptosis PARP, evaluated after 72 h TMZ treatment with or without pretreatment with CUR+PLD, in U87 (a, c) and LN18 (b, d) cells; values were normalized to GAPDH. Results are the mean ± SD from three independent experiments (n = 3). Significance (One-way ANOVA + Tukey multiple comparison test): *vs untreated control (CTR), #vs TMZ alone or pretreated with CUR+PLD: *p ˂ 0.05; **p ˂ 0.01; ns: not significant.
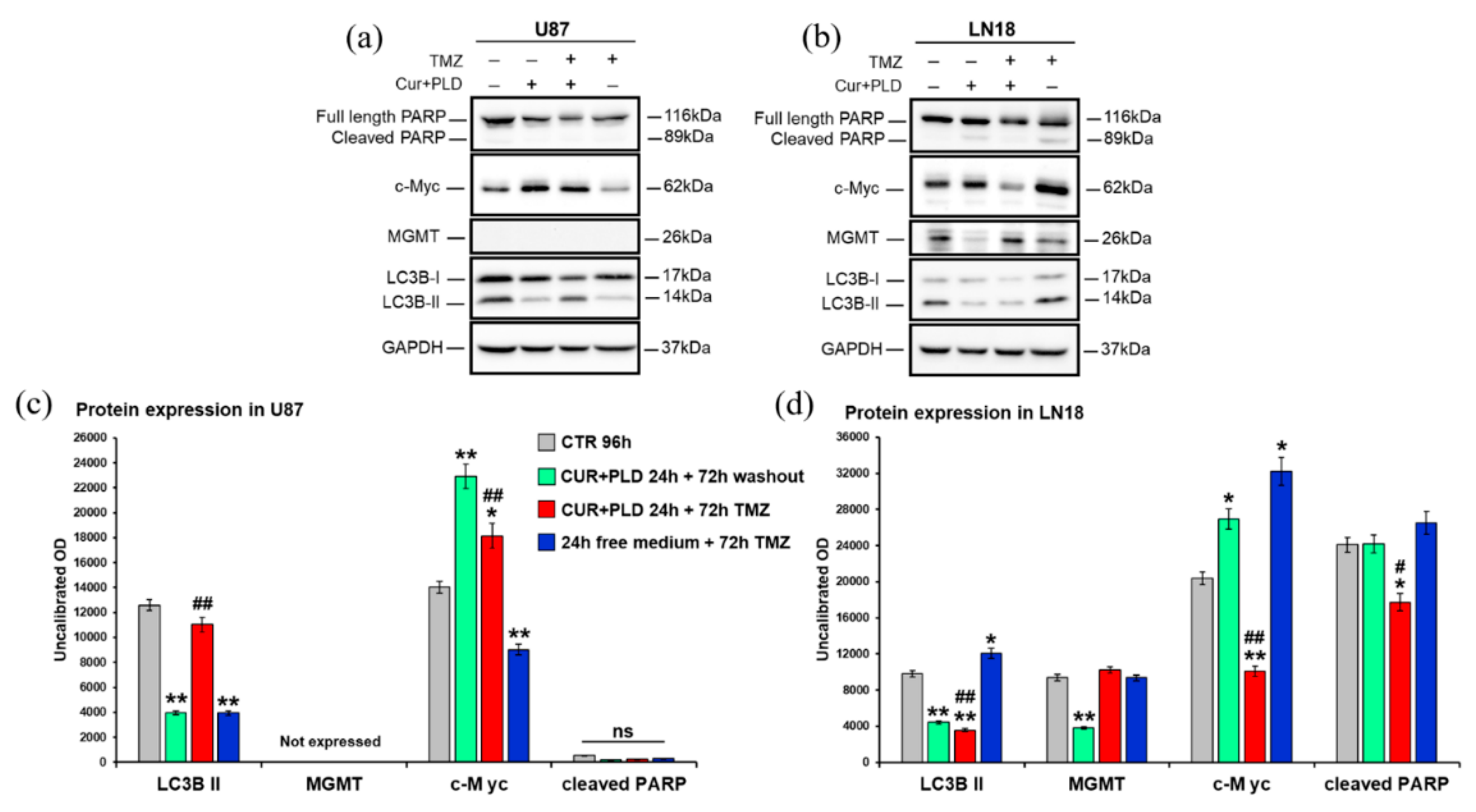
Figure 8.
Effect of pre-treatment with CUR+PLD on the actin network and ultrastructural morphology of U87 cells. Confocal microscopic analysis of the actin network (left panels) and morphology by SEM (right panels), evaluated after 72 h TMZ treatment with or without pretreatment with CUR+PLD. The scale bars represent 25 μm.
Figure 8.
Effect of pre-treatment with CUR+PLD on the actin network and ultrastructural morphology of U87 cells. Confocal microscopic analysis of the actin network (left panels) and morphology by SEM (right panels), evaluated after 72 h TMZ treatment with or without pretreatment with CUR+PLD. The scale bars represent 25 μm.
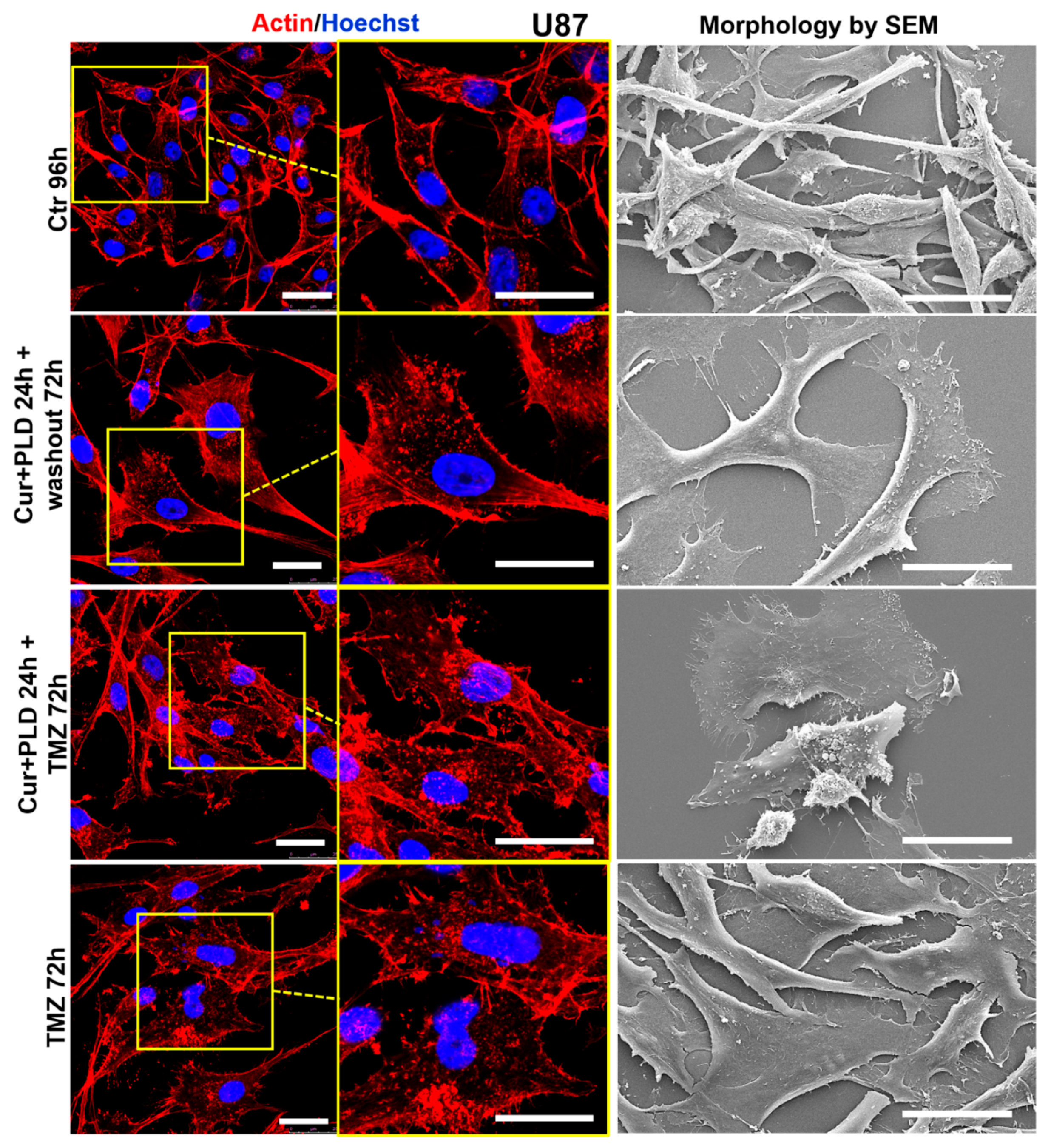
Figure 9.
Effect of pre-treatment with CUR+PLD on the actin network and ultrastructural morphology of LN18 cells. Confocal microscopic analysis of the actin network (left panels) and morphology by SEM (right panels), evaluated after 72 h TMZ treatment with or without pretreatment with CUR+PLD. The scale bars represent 25 μm.
Figure 9.
Effect of pre-treatment with CUR+PLD on the actin network and ultrastructural morphology of LN18 cells. Confocal microscopic analysis of the actin network (left panels) and morphology by SEM (right panels), evaluated after 72 h TMZ treatment with or without pretreatment with CUR+PLD. The scale bars represent 25 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



