
Submitted:
07 August 2024
Posted:
08 August 2024
You are already at the latest version
Abstract
The recent mountain pine beetle outbreaks have caused widespread mortality among lodgepole pine trees in western North America, resulting in a reduced population of surviving trees. While previous studies have focused on the cascading impacts of these outbreaks on the physiology and growth of the surviving trees, there remains a need for a comprehensive study into the interactions among various physiological traits and the growth in post-outbreak stands. Specifically, the relationship between chemical (primarily terpenes) and anatomical (mainly resin ducts) defences, as well as the allocation of non-structural carbohydrates (NSCs) to support these defence modalities, is poorly understood. To address these gaps, we conducted a field survey of surviving lodgepole pine trees in post-mountain pine beetle outbreak stands in western Canada. Our retrospective analysis targeted to determine correlations between the post-outbreak concentrations of monoterpenes, diterpenes, and NSCs in the phloem, and the historical resin duct characteristics and growth traits before and after the outbreak. We detected strong correlations between post-outbreak concentrations of monoterpenes and historical resin duct characteristics, suggesting a possible link between these two defence modalities. Additionally, we found a positive relationship between NSCs and the total concentrations of monoterpenes and diterpenes, suggesting that NSCs likely influence the production of these terpenes in lodgepole pine. Furthermore, historical tree growth patterns showed strong positive correlations with many individual monoterpenes and diterpenes. Interestingly, while surviving trees had enhanced anatomical defences after the outbreak, their growth patterns did not vary before and after the outbreak conditions. The complexity of these relationships emphasizes the dynamics of post-outbreak stand dynamics and resource allocation in lodgepole pine forests, highlighting the need for further research. These findings contribute to a broader understanding of conifer defenses and their coordinated responses to forest insect outbreaks, with implications for forest management and conservation strategies.
Keywords:
carbohydrates
; bark beetle outbreaks
; lodgepole pine
; terpene defenses
; resin ducts
1. Introduction
Periodic outbreaks of bark beetles (Coleoptera: Curculionidae, Scolytinae) have caused extensive tree mortality, significantly altering forest structure and function [1,2,3] and shifting affected areas from carbon sinks to carbon sources [4,5,6]. For instance, recent outbreaks of the mountain pine beetle (MPB, Dendroctonus ponderosae Hopkins) have devastated millions of hectares of mature lodgepole pine (Pinus contorta) forests in western North America [1,2]. Despite this widespread mortality, certain individual trees?pine and non-pine species? have survived these outbreaks [7,8,9]. These surviving trees are crucial for maintaining ecological resilience within bark beetle-affected forests and may serve as a genetic reservoir of traits that confer resistance to future outbreaks, thereby contributing to the overall health and diversity of forest ecosystems [10,11,12,13,14]. Given their importance, numerous studies have reported on the impact of bark beetle outbreaks on these surviving trees, focusing on their post-outbreak defenses (chemical or anatomical) [7,8,9] and post-outbreak growing conditions [10,11,12,13,14]. While several studies have separately examined the impact of bark beetle outbreaks on anatomical and chemical defense mechanisms of the surviving trees [e.g. 7–9,15], few have explored the complex relationship between these defense modalities in the post-outbreak stands. This study aims to retrospectively examine the relationship between the constitutive terpenes in the current year’s phloem of surviving pine trees and the historical resin duct characteristics of the same trees before and after the MPB outbreak in lodgepole pine stands affected by the outbreak.
Interactions between conifers and bark beetles represent well-studied systems, integrating molecular, chemical, and ecological perspectives to elucidate the outcomes of these interactions [16,17,18]. Bark beetles feed, mate, and lay eggs within the subcortical tissues (phloem/cambial zone) of mature trees, making them prominent agents of mortality in coniferous forests [19]. When confronted with bark beetle attacks, conifer trees employ a multifaceted defense strategy, involving both defence compounds and anatomical structures [20,21,22,23,24,25,26]. Resin, a mixture of terpenes, serve as the primary defense against bark beetles for conifers. Monoterpenes and diterpenes, in particular, have received more attention due to their biological effects on bark beetles and their fungal associates [19,27,28,29,30]. Pine phloem, the feeding site for bark beetles, contains large quantities of monoterpenes and diterpenes, which are expressed constitutively and induced [19,23,31]. The production of these terpenes is supported in part by non-structural carbohydrates (NSC), such as soluble sugars and starch [32,33,34,35].
Anatomical defences of conifers involve primarily the synthesis and storage of resin in specialized structures known as resin cells and resin ducts [20,22]. These structures are interconnected axially and radially. Resin ducts are distributed throughout the primary and secondary xylem as well as the cortical tissues. Resin accumulated within these structures is transported to the point of bark beetle attacks. Consequently, the morphology of xylem resin ducts is often associated with the quantity of resin flow in response to wounds in the phloem [37,38,39,40,41]. In some cases, resin ducts are linked to tree survival against bark beetles, with larger duct size and higher duct numbers showing stronger relationships with survival [8,9,42,43,44].
Our earlier investigations demonstrated distinct anatomical defence traits in surviving trees following MPB outbreaks compared to those killed by MPBs [8,9]. Consequently, the surviving trees contained higher concentrations of monoterpenes [7]. However, the interaction between these chemical and anatomical defences remains unknown within the lodgepole pine system. Therefore, our primary objective is to retrospectively investigate whether the constitutive concentrations of post-outbreak monoterpenes and diterpenes in the phloem of surviving pine trees correlate with the historical resin duct characteristics, radial growth, and tree growth traits (age and size) before and after MPB outbreaks in post-outbreak lodgepole pine stands. Additionally, we explored potential relationships between defence traits (chemical or anatomical) and NSCs in the phloem and any trade-offs between monoterpene and diterpene defences following the MPB outbreak. This study aims to shed light on the complex relationship between various defence modalities and tree characteristics amidst the MPB outbreak, providing valuable insights into the legacy impact of bark beetles on host trees.
2. Materials and Methods
2.1. Site Selection
In July 2020, we identified five lodgepole pine-dominated forests in Jasper National Park, Alberta (Canada). Lodgepole pine constituted 45%-59% of the total live and dead tree density (height > 1.3 m) in sites 1-3 and 100% in sites 4 and 5 (Table S1). Douglas-fir (Pseudotsuga menziesii), white spruce (Picea glauca), and aspen (Populus tremuloides) were the other tree species in these sites. All sites had experienced a recent MPB outbreak, as documented in historical records provided by Parks Canada. The outbreaks occurred in sites 1-3 in 2015 and in sites 4 and 5 in 2014. The sites were located at least 500 meters apart. We gathered data on the on-site slope using a clinometer and aspect using a compass. Tree density at a site was calculated using an angle gauge with a basal area factor of 4.6 metric (20 standard). Tree species, health status (live or killed by MPB) and diameter at breast height (DBH, 1.3 m height) were measured for all trees within the plot (Table S1). Trees killed by MPB were identified through characteristic pitch tubes, brood emergence holes on the tree stems, blue stain and oviposition (maternal) galleries under the tree bark [7]. Conversely, the healthy lodgepole pines exhibited no visible symptoms of insect or pathogen attacks.
2.2. Sample Collection
We selected 12 pairs of beetle-killed and healthy lodgepole pine trees in each site, with DBH ranging from 18 to 22 cm (n=60 trees per category). We obtained two phloem samples using a 1.9 cm dia. leather hollow punch from each of the four cardinal directions at DBH on healthy trees (8 samples per tree). The punch was cleaned using 70% ethanol before each sampling. The samples were wrapped in labelled aluminum foil and immediately placed into liquid nitrogen. The samples were stored on dry ice during transportation from the field to the laboratory and put in a -40 °C freezer until chemical analysis.
Furthermore, we obtained one 5 mm and one 12 mm dia. increment cores at 1 m above ground from the north and south aspects of each health tree. The 5 mm dia. core was drilled to the pith of the tree and used to determine its age. The 12 mm dia. cores contained at least 20 years of annual rings dating back from the sampling year and were used to determine the anatomical defence and growth characteristics of trees in the last 15 years. The cores were stored in plastic drinking straws. To determine the year of the outbreak in each site, we took a wedge from the south-facing aspect of each beetle-killed tree at DBH. Wedges contained at least 20 years of growth, counting back from the year of death.
2.3. Chemical Analysis
After removing the outer bark from phloem samples, we ground the samples in liquid N with a mini bead beater at 2,200 rpm for three 20-s intervals. The eight samples from each tree were pooled together before analysis. The samples from each tree were divided into three groups for the following chemical analyses: monoterpenes (100 mg), diterpene (30 mg), and NSCs (30 mg). All these analyses followed the protocols reported by Cale et al. [45].
2.3.1. Monoterpene Analysis
At room temperature, 0.5 mL of hexane with pentadecane (0.004%) (internal standard) was mixed with 100 mg of sample. The mixture was vortexed for 30 s, sonicated for 10 min, and then centrifuged for 15 min at 18,213 g at 4 °C. This process was repeated twice for each sample. The extract was transferred to a 2 mL GC (Gas Chromatograph) vial and stored at -40 °C until analysis. Extracts were analyzed with a GC-MS (GC 7890A, Mass Spectrometry (MS) 5975C, Agilent Tech., Santa Clara, CA, USA) fitted with an Agilent HP-Chiral-20B column (30 m x 0.25 mm ID x 0.25 µm film). The carrier gas was helium at a flow rate of 1.0 mL min-1. The temperature began at 50 °C and increased to 72 °C at a rate of 75 °C min-1, where it was held for 0.5 min, then increased at 30 °C min-1 to 90 °C and held for 2 min. It was then increased to 95 °C at 3 °C min-1 and held for 1 min, then increased to 100 °C at 5 °C min-1 and held for 0.5 min, then to 150 °C at 8 °C min-1, and finally to 175 °C at 15 °C min-1 and held for 1.5 min. The total run time for each sample was 23.6 min. We identified the hexane-extractable compounds, mostly monoterpenes, one phenylpropanoid (4-allyanisole), and one terpene derivative and its acetate ester (borneol and bornyl acetate) (hereafter monoterpenes) by comparing the mass spectra and retention times to those of known standards.
Monoterpenes were quantified using standard curves from four dilutions prepared from analytical standards of (+)-limonene (Chemical Purity: 99%), (-)-limonene (99%), α-terpinene (95%), (+)-α-pinene (98.5%), (-)-α-pinene (98%), myrcene (94%), 3-carene (98.5%), (+)-β-pinene (98%), (-)-β-pinene (99%), β-phellandrene (90%), (+)-camphene (90%), γ-terpinene (97%), terpinolene (90%), 4-allylanisole (98.5%), R-(+)-pulegone (97%), bornyl acetate (97%), (-)-borneol (99%), linalool (98%), ocimene (90%), and α-terpineol (90%). Concentrations were reported as “µg mg-1 of dry weight (DW)” phloem tissue.
2.3.2. Diterpene Analysis
Diterpenes were extracted from 30 mg of lyophilized ground tissue in 1 mL of methanol. The mixture was vortexed for 30 s, then centrifuged for 10 min at 18,213 rcf at 4 °C. The supernatant was transferred to 2 mL GC vials and kept at -40 °C until analysis. Diterpene acid extractions were analyzed using Ultra-High Performance Liquid Chromatograph (UHPLC, 1290 Infinity, Agilent Tech.), fitted with an InfinityLab Poroshell 120 EC-C18 column (2.1 x 150 mm 1.9 µm, Agilent Tech.) and a Diode Array Detector (UV/Vis, 1290 DAD, Agilent Tech.). Gradient analyses were performed with a binary solvent system: 1.7% v/v acetic acid with ultra-pure water (A) and 100% methanol (HPLC grade) (B) and flowing at 0.3 mL min-1. A 5 µL volume was used. The system started at 75% B for 1 min, then increased to 85% B over 9 min, held for 2 min, decreased to 75% B over 2 min and held at 3 min. The total run time was 17 min. Diterpene resin acids were quantified using wavelengths of 240, 268 and 282 nm as adapted from a multiwavelength detection approach from Kersten et al. [46]. Standard curves calculated from dilutions prepared of pimaric acid (80%), abietic acid (95%), dehydroabietic acid (99%), isopimaric acid (99%), levopimaric acid (95%), neoabietic acid (99%), palustric acid (92%) and sandarocopimaric acid (90%) were used to quantify diterpene acids. Pimaric and isopimaric acids were listed together due to coelution with abietic acid and cannot be quantified individually. Concentrations were reported as “µg mg-1 of DW” phloem tissue.
2.3.3. Non-Structural Carbohydrate (Soluble Sugars and Starch) Analyses
All three soluble sugars (glucose, fructose and sucrose) were extracted from 30 mg of the lyophilized and ground phloem tissue. The sample was mixed with 1.3 mL of ultra-pure water in a 2 mL tube. A marble was placed in the opening of the tube. Tubes were placed above a boiling pot of water and enclosed in steam for 1 hr. A 0.5 mL aliquot was collected and centrifuged. Then, 0.4 mL of supernatant was placed in a new 2 mL tube with 1.0 mL HPLC-grade methanol. The extracts were then incubated at room temperature for one h. Then 0.5 mL of the extract was transferred to 2 mL GC vials and kept at -40 °C until analysis.
Glucose, fructose, and sucrose were analyzed with the UHPLC system that was fitted with an Infinity Lab Poroshell 120 HILIC-Z column (2.1 x 100 mm 2.7 µm, Agilent Tech.) as well as with an Evaporative Light Scattering Detector (ELSD, 1290 ELSD II, Agilent Tech.). A gradient analysis was performed using a binary solvent system of 0.034% v/v ammonium hydroxide to buffer ultra-pure water (A) as well as acetonitrile (HPLC-grade), also buffered with 0.034% v/v ammonium hydroxide (B). The flow rate was set to 0.2 mL min-1. A 4 µL injection volume was used. The solvent system started at 90% B for 2 min, then decreased to 75% B over 3.5 min, where it was held for 1 min. It was then increased to 90% B over 2 min and held there for 0.5 min. The total run time was 9 min. The ELSD settings were set so that the nebulizer temperature was 25 °C, the evaporator tube temperature was 60°C, and the gas flow rate was 1.40 SLM. Dilution prepared from analytical standards of glucose (99%), fructose (99%) and sucrose (99.55%) were used to calculate standard curves to quantify soluble sugars.
For starch extraction, 25 mg of the lyophilized ground phloem tissue was used. Enzymatic digestions were used to convert starch into gluconate-6-phosphate, as described in Cale et al. [45]. As with the soluble sugar extraction, samples were held in steam for 1 hr. Samples were then vortexed for 30 sec. A 0.4 mL aliquot was transferred to a new 2 mL tube with 0.4 mL of alpha-amylase solution of 75 g enzyme (Sigma-Aldrich, St. Louis, MO, USA) in 100 mL ultra-pure water. This enzyme converts starch to large soluble sugars (e.g., maltose). The tubes were vortexed immediately for 30 sec and then incubated in a 50 °C bath for 16 hr. Following the incubation, tubes were removed from the water bath and then inverted twice. The extracts were then centrifuged at 18,213 rcf for 15 min, which removed undigested glucose-containing polymers (e.g., hemicellulose). After which 0.4 mL of the supernatant was transferred to a new 2 mL tube containing 0.4 amyloglucosidase solution consisting of 3 g enzyme (Sigma-Aldrich, St. Louis, MO, USA) in 60 mL sodium acetate buffer (0.1 M, pH 4.5). This enzyme converts maltose and other large sugars to glucose. The tubes were incubated in a 50 °C bath for 16 hr. Tubes were removed from the water bath and inverted twice. The extracts were centrifuged at 18,213 rcf for 15 min. Then, 0.4 mL of the extracts were transferred to 2 mL GC vials and stored at -40 °C until analysis.
We pipetted 0.020 mL of starch extract into a 96-well plate. Next, 0.2 mL of glucose hexokinase (Sigma Aldrich) - isomerase (phosphoglucose isomerase, Sigma Aldrich) was added to each well. This converted starch aliquots into gluconate-6-phosphate, as described in Cale et al. [45]. The well plate was shaken at room temperature for 45 min on an orbital shaker. Then, the amount of gluconate-6-phosphate was measured at an absorbance of 340 nm using the Synergy Microplate Reader H1 (BioTek, Winooski, VT, USA).
The starch concentration was quantified using two calibration curves: the first to estimate the glucose concentration from the amount of gluconate-6-phosphate measured and the second to estimate the starch concentration using the estimated glucose concentration. To do this, the sample concentration of total glucose was standardized by sample dry weight and extraction volume after the first calibration curve. Previously determined concentrations of soluble glucose and fructose (hexoses) were subtracted from the estimated total glucose concentration. A conversion factor was applied to account for potential quantification differences. The second calibration curve used dilutions of pure potato starch standards processed with the same enzymatic digestions as described above. The potential matrix effects were calculated to adjust the final concentrations accordingly.
2.4. Anatomical Defence and Growth Characteristics
Core and wedge samples were glued to wooden mounts and dried for two weeks at room temperature. Samples were then sanded with belt and hand sanders using progressively finer sandpaper. Samples were then scanned at 1,200 dpi to create high-resolution digital images. Annual growth (mm) was measured from pith to last year’s annual ring using ImageJ on all core samples [8]. Calendar years were assigned to each annual ring by visual cross-dating ring characteristics, such as width ratio and the colour contrast between earlywood and latewood per ring. This information was used for wedges to determine the year of death of trees killed by MPB. The earliest death recorded was considered the starting year of the outbreak in a particular site. Among the five sites, two sites experienced the outbreak in 2014, and the remaining three sites had an MPB outbreak in 2015. Therefore, the healthy trees were separated into two outbreak years: 2014 (n=24) and 2015 (n=36).
All samples for resin ducts were examined using ImageJ within a fixed sampling width of 10 mm [8]. To analyze resin ducts, resin duct production (number of resin ducts per 10 mm width per year on a ring [no. yr-1]), resin duct size (mean size of resin ducts per 10 mm width per year on a ring [mm2 yr-1]), total resin duct area (sum of resin duct area per 10 mm width per year on a ring [mm2 yr-1]), resin duct density (total number of resin ducts per year divided by the ring area (10 mm * ring width) for a given year [no. mm2 yr-1], and relative resin duct area (percent area occupied by resin ducts per year within a ring area for a given year [% yr-1]) was calculated for each 12 mm dia. core [13,45]. Ring width (mm yr-1) and basal area increment (BAI) (mm2 yr-1) were used to represent tree radial growth. Tree radii and ring width data were used to calculate BAI, assuming that tree rings are concentric circles. Tree radii were calculated by dividing tree DBH by two.
2.5. Data Analysis
To study the long-term relationship between growth and defence mechanisms in healthy pine trees, we analyzed annual growth and resin duct characteristics across three distinct periods: two pre-outbreak (5-yr and 10-yr before the outbreak) and one post-outbreak (from outbreak onset until 2019 when the last annual ring was formed at the time of sampling) periods.
We conducted Pearson correlation tests to examine correlations between terpenes (21 monoterpenes, six diterpene resin acids, total monoterpenes, and total diterpenes), NSCs (three soluble sugars, starch, and total NSCs) and five anatomical defence traits and two radial growth characteristics. Analysis was performed using R (Version 4.0.5) [47] using Hmisc R package [48]. We selected BAI to represent tree radial growth as it strongly correlates with ring width (Pearson Correlation Coefficient > 0.96 in both the 2014- and 2015- outbreak years). In analyses, we considered overall as well as total and individual concentrations of constitutive monoterpenes and diterpenes.
2.5.1. Gradient Analysis
We employed an indirect gradient analysis to examine the impact of resin duct characteristics, BAI, tree age, and DBH on the overall monoterpenes, diterpenes, and NSCs before and after outbreaks. We utilized non-linear multidimensional scaling (NMDS) with Bray–Curtis dissimilarity to ordinate monoterpenes, diterpenes, and NSCs using the vegan R package [49]. NMDS results were saved as RData files to ensure reproducibility. Subsequently, we projected resin duct characteristics from pre- and post-outbreak periods onto NMDS axes as environmental gradients and visualized these relationships using ordination triplots created with the ggplot2 R package [50]. We followed the same steps when incorporating radial growth characteristics from pre- and post-outbreak periods and tree growth traits (age, DBH) into the NMDS axes for the 2014 and 2015 outbreak years.
2.5.2. Linear Mixed-Effect Models
To identify relationships among defence and growth characteristics, we employed linear mixed-effect models, treating the site as a random effect using the R package lme4 [51]. We visually assessed model assumptions, including normality and homogeneity of residuals, applying transformations and rescaling as needed. Resin duct density and the percent resin duct area were standardized per 1 mm2 and accounted for BAI prior to calculation. Furthermore, we investigated the relationships between total monoterpenes or diterpenes and tree growth traits using linear mixed-effect models. When we observed a statistically significant relationship, we conducted further linear mixed-effect models with the site as a random effect to examine the specific relationship between individual monoterpenes or diterpenes and a particular predictor. In all cases, significance was determined at α = 0.05.
2.5.3. Comparative Analyses
We compared anatomical defences and radial growth responses across pre- and post-outbreak periods within the 2014 and 2015 outbreak years using linear mixed-effect models. Variability across sites was accounted for by nesting trees within sites. Furthermore, we examined the differences in tree age and DBH between the 2014 and 2015 outbreak years using linear mixed-effect models, where the site was considered a random effect to address any unexplained variances among trees.
3. Results
3.1. Do Historical Resin Duct Characteristics Relate to the Current Year’s Terpenes?
3.1.1. Monoterpenes
We observed several significant relationships between some of the resin duct characteristics and overall monoterpenes across both the 2014- and 2015-outbreak years (Figure 1, Table 1). In the 2014 outbreak, the relationship between the total monoterpenes and resin duct production and the total resin duct area was significant during all three periods. Resin duct size was also positively correlated with the total monoterpenes in the post-outbreak and 10-year post-outbreak period. In the 2015 outbreak, the total resin duct area and the resin duct size displayed significant positive relationships with total monoterpenes in all three time periods. Further testing of the relationship between individual monoterpenes and three resin duct predictors (resin duct production, total resin duct area, and individual resin duct size) yielded significant results. Individual monoterpene concentrations across sites are shown in Table S2. All three resin duct characteristics exhibited positive trends with several monoterpenes in the 2014 and 2015 outbreak years (Table 2). In the 2014 outbreak, we observed significant relationships between resin duct characteristics and 16 monoterpenes, while 15 monoterpenes displayed significant relationships with resin duct characteristics in the 2015 outbreak. Among the monoterpenes examined, several had significant relationships with at least one of the resin duct characteristics. Notably, (+)-limonene, (-)-α-pinene, (+)-β-pinene, β-phellandrene, myrcene, (+)-camphene, (-)-borneol and linalool correlated with most of the resin duct characteristics in both outbreak years (Table 2). Additionally, α-terpinene, (+)-α-pinene, 3-carene, γ-terpinene, terpinolene, (R)-(+)-pulegone, bornyl acetate, ocimene, and α-terpineol also exhibited significant relationships with at least one of the resin duct characteristics.
We did not observe any relationship between any of the resin duct characteristics and diterpene profiles or total diterpenes or individual diterpenes in either the pre- or post-outbreak periods within the 2014 and 2015 outbreak years, suggesting no association between phloem diterpenes and xylem anatomical defences (Figure 2).

Table 3.
Results of linear mixed-effect models showing the relationship between total monoterpenes and total diterpenes, and the annual basal area increment (BAI) of lodgepole pine trees in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years and periods.
Table 3.
Results of linear mixed-effect models showing the relationship between total monoterpenes and total diterpenes, and the annual basal area increment (BAI) of lodgepole pine trees in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years and periods.
| Outbreak Years | BAI | Total monoterpenes | Total diterpenes | ||||
| Coefficient | t-value | P-value | Coefficient | t-value | P-value | ||
| 2014 | Post-outbreak | 532.4 | 2.7 | 0.015 | -0.8 | -0.5 | 0.608 |
| 5-yr pre-outbreak | 549.9 | 2.6 | 0.015 | -0.8 | -0.5 | 0.658 | |
| 10-yr pre-outbreak | 665.1 | 3.2 | 0.004 | -1.2 | -0.7 | 0.516 | |
| 2015 | Post-outbreak | 485.9 | 2.6 | 0.013 | 4.9 | 2.7 | 0.012 |
| 5-yr pre-outbreak | 686.8 | 3.7 | 0.002 | 4.9 | 2.4 | 0.024 | |
| 10-yr pre-outbreak | 791.7 | 3.7 | 0.001 | 6.3 | 2.6 | 0.013 | |
Significant P-values (α=0.05) were bolded and italicized.3.1.2. Diterpenes.
3.2. Does NSCs Relate to the Terpenes?
In the 2014 and 2015 outbreaks, there were no significant relationships between any of the NSCs and overall monoterpenes (Figure S1, Table S3) or diterpenes (Figure S2, Table S3). However, when we tested the relationship between the total NSCs and the total monoterpenes or total diterpenes within the 2015 outbreak, the NSCs were significantly and positively related to the total diterpenes (coefficient = 0.701, P = 0.019) and the total monoterpenes (coefficient = 59.25, P = 0.048). We did not observe any significant relationships in the 2014 outbreak year.
3.3. Do the Radial Growth Characteristics Relate to the Terpenes?
Due to the strong correlation between BAI and the annual radial growth rate (ring width), we opted to use BAI as a representation of radial tree growth to account for tree size. Overall, monoterpenes are strongly associated with BAI across all pre- and post-outbreak periods (2014-outbreaK: r2 values ranged from 0.414 to 0.516, P-values from 0.01 to 0.001; 2015-outbreak: r2 values from 0.194 to 0.202, P-values from 0.047 to 0.033; Figure 3). Conversely, the relationship between diterpenes and BAI was marginally significant during the 10-yr pre-outbreak period in the 2015 outbreak (P = 0.05, Figure 4, Table S4).
Our analysis revealed strong positive relationships between individual monoterpenes and BAI (Table 4). BAI had a significantly positive relationship with 65% of individual monoterpenes in all pre-and post-outbreak periods in the 2014 outbreak and 50% of individual monoterpenes in the same periods in the 2015 outbreak. Specifically, (+)-limonene, (-)-α-pinene, myrcene, (+)-β-pinene, β-phellandrene, (+)-camphene, and linalool exhibited consistent positive relationships with BAI across all time periods in both outbreak years. However, this relationship appears to be stronger in the 2014 outbreak. Additionally, within the 5-yr and 10-yr pre-outbreak periods, the relationship between individual monoterpenes and BAI was more pronounced.
The analysis revealed a strong positive relationship between individual diterpenes and BAI across all pre- and post-outbreak periods, specifically within the 2015 outbreak (Table 4). This relationship extended to various diterpenes, including dehydroabietic, levopimaric, palustric, neoabietic, and abietic acids. In contrast, no significant relationship was observed between individual diterpenes and BAI within the 2014 outbreak.
3.4. Do tree DBH and Age Relate to the Terpenes?
Even though the trees sampled had similar DBH in 2014 and 2015 (2014: 22.19±0.59 cm; 2015: 20.76±0.32 cm), there was a notable difference in the mean age of trees between the two outbreak years (2014: 64.27±3.01; 2015: 80.42±3.85, P = 0.005). While DBH did not correlate with monoterpenes in either outbreak year, tree age exhibited a significant relationship with monoterpenes in both the 2014 and 2015 outbreaks (P2014 = 0.001, P2015 = 0.017, Figure 5, Table S5). Tree age negatively correlated with the total (P2014 = 0.043, t-value: -2.2; P2015 = 0.009, t-value: -2.8; Figure 5) and individual (Table 5, Figure 5) monoterpenes. Tree age was linked to 45% of individual monoterpenes in the 2014 outbreak and 40% in the 2015 outbreak. Briefly, (+)-limonene, (-)-α-pinene, γ-terpinene, and linalool strongly correlated with tree age in both outbreak years.
The analysis indicated that tree DBH and age had no significant effects on diterpenes. Additionally, individual diterpenes did not exhibit any relationships with DBH or age (Figure S3) within the 2014 or 2015 outbreak years. Given these findings, further tests examining the relationships between individual diterpenes and tree age or DBH were not pursued, as no significant associations were detected.
3.5. Do NSCs Relate to Anatomical Defence Traits, Radial Growth Rate, and Tree Growth Traits?
The results from the indirect gradient analysis indicated that neither anatomical defences (Figure S4, Table S6) nor radial increment growth rate (BAI, Figure S5, Table S4) within the xylem had any substantial relationships with NSCs in the phloem for both the 2014 and 2015 outbreak years. Similarly, non-significant results were obtained when investigating the relationship between the total NSCs and each resin duct characteristic (Table 1) and BAI (Table 3) for both the 2014 and 2015 outbreaks. Additionally, neither tree age nor DBH appeared to impact the NSC profile, individual NSCs, or total NSCs (Figure S6).
3.6. Is There Any Trade-Off between Monoterpenes and Diterpenes?
3.7. Additional Results
The correlation between anatomical defence and growth characteristics was significant across all time periods. The additional comparisons of anatomical defences and BAI among the post-outbreak, 5-yr pre-outbreak, and 10-yr pre-outbreak periods yielded significant results within both the 2014 (Figure 6) and 2015 (Figure 7) outbreaks. Overall, in both outbreaks, the resin duct characteristics exhibited greater size and relative area during the post-outbreak period compared to the 5-yr and 10-yr pre-outbreak periods. For instance, the annual resin duct size following the 2014 or 2015 outbreaks was larger by approximately 10-20% compared to both pre-outbreak periods. In the 2014 outbreak, the relative resin duct area after the outbreak was about 20% larger than in both the 5-yr and 10-yr pre-outbreak periods, even though BAI did not exhibit a significant response to the outbreak. In the 2015 outbreak, which comprised older pine trees, BAI during the 5-yr pre-outbreak period was 12% larger than in the 10-yr pre-outbreak period. However, no significant growth release was observed following the 2015 outbreak. Moreover, the total resin duct area during the post-outbreak period was 12% larger than during the 5-yr pre-outbreak period but similar to that observed in the 10-yr pre-outbreak period. Notably, resin duct production, density, and size displayed stronger responses following the 2014 and 2015 outbreaks than before. It's important to mention that including the site as a random effect in the linear mixed-effect models represented relatively high variance, which might contribute to the variations observed in terpenes and NSCs.
4. Discussions
We conducted a retrospective analysis of the constitutive monoterpenes and diterpenes, NSCs, and their interactions with anatomical defences and growth characteristics in surviving lodgepole pine trees in post-MPB-outbreak stands. We observed a strong correlation between post-outbreak concentrations of monoterpenes in the phloem and the historical resin duct characteristics in the xylem, along with a positive association between NSCs and total monoterpenes and diterpenes. Despite finding no evidence of growth release (BAI) in surviving pine trees, we observed strong positive relationships between BAI and several individual monoterpenes and diterpenes. Notably, tree age had a negative relationship with monoterpenes. Furthermore, the outbreak enhanced the resin duct characteristics in lodgepole pine. Our analyses, however, revealed no relationships between anatomical defence traits, BAI, tree age, size, and NSCs, nor did we find evidence supporting a trade-off between monoterpenes and diterpenes. These findings offer valuable insights into our broader understanding of conifer defences, particularly concerning how anatomical defences interact in response to outbreaking bark beetle species.
Our study showed strong correlations between monoterpenes and resin duct characteristics. Notably, overall monoterpenes were highly correlated with at least one of the five resin duct characteristics. For instance, the total monoterpenes were linked to the resin duct production, the resin duct size, and the total resin duct area. Furthermore, those monoterpenes that are crucial during MPB host colonization, such as (−)-α-pinene, β-phellandrene, (+)-limonene, and myrcene, had positive relationships with all resin duct characteristics, agreeing with our prior research findings [7,23,27,52,53]. Likewise, Kichas et al. [15] reported strong correlations between resin duct area, resin duct production and monoterpene concentrations in whitebark pine (P. albicaulis). In contrast, we found fewer relationships between diterpenes and resin duct characteristics, suggesting a weaker association between them. Similarly, Mason et al. [24] reported that anatomical defence traits of lodgepole pine differentially influenced the different classes of terpenes. Collectively, these findings highlight that the relationship between chemical and anatomical defences may depend on the classes of defence compounds [15]. This is in line with the conclusion from a meta-analysis, suggesting that trade-offs between monoterpenes and resin ducts are relatively rare across a wide range of plant species despite occasional occurrences in some systems [54]. Nonetheless, these results highlight the complementary role of anatomical and chemical defence structures in post-outbreak lodgepole pine stands, likely in response to the MPB outbreaks [7,8,9,22,24,40,42,43,55].
The second contribution of our study lies in the evidence regarding the positive relationship between total NSCs and total monoterpenes or total diterpenes within the 2015-outbreak. Although such relationships were not consistent among sites, this result suggests that the production of monoterpenes or diterpenes is linked to NSCs, aligning with previous research findings [32,33,34,35,56]. For instance, Mullin et al. [57] reported a positive relationship between starch concentration and total monoterpenes and diterpenes in lodgepole pine trees across varying elevations. Generally, it is well-documented that terpenoid defences require substantial energy demands [58,59,60] and deplete carbohydrate reserves [32,34,60]. However, the absence of a similar relationship between anatomical defences and NSCs suggests that our current understanding of the connections between sinks (defences) and sources (NSCs) in conifers is limited and necessitates further field testing [34,38].
One of the strongest relationships we observed is that both monoterpenes and diterpenes were positively correlated to tree radial growth, as measured by BAI. In fact, BAI exhibited positive relationships with up to 65% of individual monoterpenes, including (+)-limonene, (−)-α-pinene, myrcene, and β-phellandrene. Similarly, we observed a positive relationship between individual diterpenes, including dehydroabietic, levopimaric, palustric, neoabietic, and abietic acids, and BAI in the 2015 outbreak year. It is worth noting that the varying diterpene-BAI relationships between outbreaks in 2014 and 2015 could be attributed to differences in pine density, as the pine density in the earlier was nearly double that of the latter.
We observed negative correlations between tree age and monoterpenes in the 2014- and 2015-outbreak years. Specifically, the age of trees was linked to approximately 45% of individual monoterpenes in the 2014-outbreak and 40% in the 2015-outbreak. Notably, (+)-limonene, (−)-α-pinene, γ-terpinene, and linalool exhibited the strongest negative associations with tree age. These findings suggest that irrespective of tree size, tree age may be an important factor in determining the allocation of carbon resources to defence mechanisms. It appears that as trees mature, the allocation of resources to defences becomes less of a priority, whereas younger trees may invest more heavily in defence mechanisms. This could be attributed to the fact that younger trees are potentially more vulnerable to herbivory and thus prioritize defence allocation more than older trees, as supported by Swihart and Bryant [62] and Erbilgin and Colgan [63]. In the case of lodgepole pine, it was found that young trees in the range of 30–50 years old exhibited greater resistance to ophiostomoid fungi carried by MPB compared to both younger and older trees [64]. Overall, the relationship between the ontogeny of investment in defence versus growth in long-lived trees remains a complex and unresolved topic within the field of research [2,65].
The MPB outbreak influenced various physiological and growth traits of surviving lodgepole pine trees. Firstly, post-outbreak conditions improved resin duct characteristics compared to pre-outbreak conditions, with resin duct size, relative resin duct area, and total resin duct area all increasing. This suggests that increased investment in anatomical defences following MPB outbreaks, agreeing with the results of earlier studies. For instance, Zhao and Erbilgin [8] reported increased resin duct investment in surviving lodgepole pine trees. Resins serve as the initial line of defence for trees against bark beetle attacks by providing physical and chemical protection [20,30,66,67]. The number and size of xylem resin ducts in pine trees can predict resin production in response to wounding or bark beetle attacks [37,38,40]. Pines that survive MPB attacks tend to have more and larger xylem resin ducts [8,9,42,43,55] and increased pre-attack resin production [68]. Based on the anatomical defence and terpene duct characteristics observed in this and other studies, it would be expected that beetles would show a preference for larger pine trees.
Finally, our study did not provide evidence of growth release (BAI) in the surviving lodgepole pine trees. Furthermore, we found no connections between anatomical defence traits and BAI or evidence suggesting any notable relationships between monoterpenes and diterpenes. These findings contradict those of Kichas et al. [15], who reported significant correlations between BAI and resin duct characteristics in whitebark pine and lodgepole pine. We suspect that the enhanced anatomical defences observed after outbreaks may have required additional resources to support their development and maintenance rather than growth. Alternatively, other conifer species, such as spruce, and understory woody plant species like aspen, may have taken advantage of the available resources following the MPB outbreaks in this study, affecting the growth of surviving lodgepole pine trees. This suggests a complex interplay of factors influencing post-outbreak stand dynamics and resource allocation in these forest ecosystems, highlighting the need for further research to understand these intricate relationships fully.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
GB and NE designed the experiment; GB, JGK, SZ and NE conducted the experiment; GB, SZ, and NE analyzed the data; SZ and NE wrote the manuscript; all authors contributed to editing.
Funding
We acknowledge funding for this research from the Natural Sciences and Engineering Research Council of Canada-Discovery Award to NE.
Data availability Statement
Upon acceptance of the paper, the original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding author.
Acknowledgments
We acknowledge that the research presented herein was conducted according to all applicable laws and rules set forth by the provincial (Alberta) and federal governments and the University of Alberta. All necessary permits were obtained before the research was conducted. The authors declare no conflict of interest.
Conflicts of Interest
The authors disclose that they have no competing interests.
Abbreviations:
BAI: Basal area increment
DBH: Diameter at breast height
DW: Dry weight
ELSD: Evaporative light scattering detector
GC/MS: Gas chromatograph/mass spectrometer
MPB: Mountain pine beetle
NMDS: Non-linear multidimensional scaling
NSCs: Non-structural Carbohydrates
UHPLC: Ultra-High Performance Liquid Chromatograph
References
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale Drivers of Natural Disturbances Prone to Anthropogenic Amplification: The Dynamics of Bark Beetle Eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate Change and Bark Beetles of the Western United States and Canada: Direct and Indirect Effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Kausrud K, Økland B, Skarpaas O, Grégoire JC, Erbilgin N, Stenseth NC (2012) Population dynamics in changing environments: the case of an eruptive forest pest species. Biol Rev. 87: 34-51.
- Seidl R, Schelhaas MJ, Rammer W, Verkerk PJ (2014) Increasing forest disturbances in Europe and their impact on carbon storage. Nat Clim Chang. 4:806–810.
- Ghimire, B.; Williams, C.A.; Collatz, G.J.; Vanderhoof, M.; Rogan, J.; Kulakowski, D.; Masek, J.G. Large carbon release legacy from bark beetle outbreaks across Western United States. Glob. Chang. Biol. 2015, 21, 3087–3101. [Google Scholar] [CrossRef] [PubMed]
- Aldea J, Dahlgren J, Holmström E, Löf M (2023) Current and future drought vulnerability for three dominant boreal tree species. Glob Change Biol. 30: e17079.
- Erbilgin N, Cale JA, Hussain A, Ishangulyyeva G, Klutsch JG, Najar A, Zhao S (2017) Weathering the storm: how lodgepole pine trees survive mountain pine beetle outbreaks. Oecologia. 184:469–478.
- Zhao SY, Erbilgin N (2019) Larger resin ducts are linked to the survival of lodgepole pine trees during mountain pine beetle outbreak. Front Plant Sci. 10:1459 https://doiorg/103389/fpls201901459.
- Zhao SY, Klutsch JG, Cale JA, Erbilgin N (2019) Mountain pine beetle outbreak enhanced resin duct-defenses of lodgepole pine trees. For Ecol Manag. 441: 271-279.
- Amoroso MM, Coates KD, Astrup R (2013) Stand recovery and self-organization following large-scale mountain pine beetle induced canopy mortality in northern forests. For Ecol Manag. 310: 300-311.
- Perovich C, Sibold JS (2016) Forest composition change after a mountain pine beetle outbreak, Rocky Mountain National Park, CO, USA. For Ecol Manag. 1: 366.
- Morris JE, Buonanduci MS, Agne MC, Battaglia MA, Harvey BJ (2022) Does the legacy of historical thinning treatments foster resilience to bark beetle outbreaks in subalpine forests? Ecol Appl. 32 (1): e02474.
- Rodman, K.C.; Andrus, R.A.; Carlson, A.R.; Carter, T.A.; Chapman, T.B.; Coop, J.D.; Fornwalt, P.J.; Gill, N.S.; Harvey, B.J.; Hoffman, A.E.; et al. Rocky Mountain forests are poised to recover following bark beetle outbreaks but with altered composition. J. Ecol. 2022, 110, 2929–2949. [Google Scholar] [CrossRef]
- Lieffers VJ, Benedik J, Stadt K, Macdonald SE (2024) Poor regeneration of pine after mountain pine beetle attack in colder boreal regions of Canada. Can J For Res. 54:168-191.
- Kichas NE, Trowbridge AM, Raffa KF et al (2021) Growth and defense characteristics of whitebark pine (Pinus albicaulis) and lodgepole pine (Pinus contorta var latifolia) in a high-elevation, disturbance-prone mixed-conifer forest in northwestern Montana, USA. For Ecol Manag. 493: 119286.
- Marini L, Økland B, Jönsson AM et al (2017) Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography. 40:1426–1435.
- Thom D, Rammer W, Seidl R (2017) The impact of future forest dynamics on climate: interactive effects of changing vegetation and disturbance regimes. Ecol Monog. 87:665–684.
- Sommerfeld A, Rammer W, Heurich M, Hilmers T, Müller T, Seidl R (2021) Do bark beetle outbreaks amplify or dampen future bark beetle disturbances in Central Europe? J Ecol. 109: 737-749.
- Raffa KF, Aukema BH, Erbilgin N, Klepzig KD, Wallin KF (2005) Interactions among conifer terpenoids and bark beetles across multiple levels of scale: an attempt to understand links between population patterns and physiological processes In T R John (Ed), Recent Advances in Phytochemistry (pp 79–118) Kidlington, Oxford, UK: Elsevier.
- Franceschi VR, Krokene P, Christiansen E, Krekling T (2005) Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 167: 353–76.
- Erbilgin N, Krokene P, Christiansen E, Zeneli G, Gershenzon J (2006) Exogenous application of methyl jasmonate elicits defenses in Norway spruce (Picea abies) and reduces host colonization by the bark beetle Ips typographus. Oecologia. 148: 426–436.
- Vazquez-Gonzalez C, Zas R, Erbilgin N, Ferrenberg S, Rozas V, Sampedro L (2020) Resin ducts as resistance traits in conifers: linking dendrochronology and resin-based defenses. Tree Physiol. 40:1313-1326.
- Erbilgin N (2019) Phytochemicals as mediators for host range expansion of a native invasive forest insect herbivore. New Phytol. 221: 1268–1278.
- Mason CJ, Keefover-Ring K, Villari C, Klutsch JG, Cook S, Bonello P, Erbilgin N, Raffa KF, Townswend PA (2019) Anatomical defenses against bark beetles related to degree of historical exposure between species and are allocated independently of chemical defenses within trees. Plant Cell Environ. 42: 633–646.
- Nagel R, Hammerbacher A, Kunert G, Phillips MA, Gershenzon J, Schmidt A (2022) Bark beetle attack history does not influence the induction of terpene and phenolic defenses in mature Norway spruce (Picea abies) trees by the bark beetle-associated fungus Endoconidiophora polonica. Front Plant Sci. 6:13:892907.
- Mageroy MH, Nagy NE, Steffenrem A, Krokene P, Hietala AM (2023) Conifer defences against pathogens and pests - mechanisms, breeding, and management. Curr For Rep. 9: 429–443.
- Erbilgin N, Ma C, Whitehouse C, Shan B, Najar A, Evenden M (2014) Chemical similarity between historical and novel host plants promotes range and host expansion of the mountain pine beetle in a naïve host ecosystem. New Phytol. 201: 940–950.
- Chiu CC, Keeling CI, Bohlmann J (2017) Toxicity of pine monoterpenes to mountain pine beetle. Sci Reps. 7: 8858 (7). [CrossRef]
- Ullah A, Klutsch JG, Erbilgin N (2021) Production of complementary defense metabolites reflects a co-evolutionary arms race between a host plant and a mutualistic bark beetle-fungal complex. Plant Cell Environ. 44: 3064-3077.
- Chiu CC, Bohlmann J (2022) Mountain pine beetle epidemic: an interplay of terpenoids in host defense and insect pheromones. Ann Rev Plant Biol. 73: 475-494.
- Schiebe C, Hammerbacher A, Witzell J et al (2012) Inducibility of chemical defenses in Norway spruce bark is correlated with unsuccessful mass attacks by the spruce bark beetle. Oecologia. 170:183–198.
- Goodsman DW, Lusebrink I, Landhäusser SM, Erbilgin N, Lieffers VJ (2013) Variation in carbon availability, defense chemistry and susceptibility to fungal invasion along the stems of mature trees New Phytol 197: 586–594.
- Roth M, Hussain A, Cale JA, Erbilgin N (2018) Successful colonization of lodgepole pine trees by mountain pine beetle increased monoterpene production and exhausted carbohydrate reserves. J Chem Ecol. 44: 209–214.
- Erbilgin, N.; Zanganeh, L.; Klutsch, J.G.; Chen, S.; Zhao, S.; Ishangulyyeva, G.; Burr, S.J.; Gaylord, M.; Hofstetter, R.; Keefover-Ring, K.; et al. Combined drought and bark beetle attacks deplete non-structural carbohydrates and promote death of mature pine trees. Plant, Cell Environ. 2021, 44, 3866–3881. [Google Scholar] [CrossRef] [PubMed]
- Huang JB, Kautz M, Trowbridge AM et al (2020) Tree defense and bark beetles in a drying world: carbon partitioning, functioning and modeling. New Phytol. 225: 26-36.
- Zas R, Moreira X, Ramos M et al (2015) Intraspecific variation of anatomical and chemical defensive traits in Maritime pine (Pinus pinaster) as factors in susceptibility to the pinewood nematode (Bursaphelenchus xylophilus). Trees-Struct Func. 29:663–673.
- Blanche CA, Lorio P L, Sommers RA, Hodges JD, Nebeker T E (1992) Seasonal cambial growth and development of loblolly pine: xylem formation, inner bark chemistry, resin ducts, and resin flow. For Ecol Manag. 49: 151–65.
- Lombardero MJ, Ayres MP, Lorio Jr, PL, Ruel JJ (2000) Environmental effects on constitutive and inducible resin defences of Pinus taeda. Ecol Let. 3: 329–339.
- Rodríguez-García A, López R, Martín JA, Pinillos F, Gil L (2014) Resin yield in Pinus pinaster is related to tree dendrometry, stand density and tapping-induced systemic changes in xylem anatomy. For Ecol Manag. 313: 47–54.
- Hood S, Sala A (2015) Ponderosa pine resin defenses and growth: metrics matter. Tree Physiol. 35: 1223–1235.
- Westbrook JW, Walker AR, Neves LG et al (2015) Discovering candidate genes that regulate resin canal number in Pinus taeda stems by integrating genetic analysis across environments, ages, and populations. New Phytol. 205: 627–641.
- Kane JM, Kolb TE (2010) Importance of resin ducts in reducing ponderosa pine mortality from bark beetle attack. Oecologia. 164: 601–609.
- Ferrenberg S, Kane JM, Mitton JB (2014) Resin duct characteristics associated with tree resistance to bark beetles across lodgepole and limber pines. Oecologia. 174: 1283–92.
- Hood S, Sala A, Heyerdahl EK, Boutin M (2015) Low-severity fire increases tree defense against bark beetle attacks. Ecology. 96: 1846–1855.
- Cale AC, Klutsch GJ, Dykstra BC, Peters B, Erbilgin N (2019) Pathophysiological responses of pine defensive metabolites largely lack differences between pine species but vary with eliciting ophiostomatoid fungal species. Tree Physiol. 39: 1121–1135.
- Kersten PJ, Kopper BJ, Raffa KF, Illman BL (2006) Rapid analysis of abietanes in conifers J Chem Ecol. 22: 1367–1388.
- R Core Team, R, 2018 R: A language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria https://wwwR-projectorg/.
- Harrell, F.E., Jr. HMISC: Harrell Miscellaneous. R Package Version 4.1-1. 2018. Available online: http://CRAN.R-project.org/package=Hmisc (accessed on 21 March 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H.; Vegan: Community Ecology Package. R Package Version 2.4-6. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 27 April 2018).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Soft 67(1): 1-48.
- Klutsch JG, Erbilgin N (2018) Dwarf mistletoe infection in jack pine alters growth-defense relationships. Tree Physiol. 38:1538–1547.
- Erbilgin N (2019) Phytochemicals as mediators for host range expansion of a native invasive forest insect herbivore. New Phytol. 221: 1268–1278.
- Moles AT, Peco B, Wallis IR et al (2013) Correlations between physical and chemical defences in plants: tradeoffs, syndromes, or just many different ways to skin a herbivorous cat? New Phytol. 198: 252–263.
- Gaylord ML, Kolb TE, McDowell NG (2015) Mechanisms of piñon pine mortality after severe drought: a retrospective study of mature trees. Tree Physiol. 35: 806–816.
- Dietze MC, Sala A, Carbone MS, Czimczik CI, Mantooth JA, Richardson AD, Vargas R (2014) Nonstructural carbon in woody plans. Ann Rev Plant Biol. 65: 667–687.
- Mullin M, Klutsch JG, Cale JA, Hussain A, Zhao S, Whitehouse C, Erbilgin N (2021) Primary and secondary metabolite profiles of lodgepole pine trees change with elevation, but not with latitude. J Chem Ecol. 47: 280–293.
- Herms DA, Mattson WJ (1992) The dilemma of plants: to grow or defend. The Quar Rev Biol. 67: 283–335.
- Gershenzon J (1994) Metabolic costs of terpenoid accumulation in higher plants. J Chem Ecol. 20: 1281–1328.
- Wiley E, Rogers BJ, Hodgkinson R, Landhäusser SM (2016) Nonstructural carbohydrate dynamics of lodgepole pine dying from mountain pine beetle attack. New Phytol. 209: 550–562.
- Kolb TE, Keefover-Ring K, Burr SJ, Hofstetter R, Gaylord M, Raffa K F (2019) Drought-mediated changes in tree physiological processes weaken tree defenses to bark beetle attack. J Chem Ecol. 45:888–900.
- Swihart RK, Bryant JP (2001) Importance of biogeography and ontogeny of woody plants in winter herbivory by mammals. J Mammal. 82: 1–21.
- Erbilgin N, Colgan LJ (2012) Differential effects of plant ontogeny and damage type on phloem and foliage monoterpenes in jack pine (Pinus banksiana). Tree Physiol. 32: 946–957.
- Shrimpton DM (1973) Age- and size-related response of lodgepole pine to inoculation with Europhium clavigerum. Can J Bot. 51:1155–1160.
- Hartmann H, Moura CF, Anderegg WR et al (2018) Research frontiers for improving our understanding of drought-induced tree and forest mortality. New Phytol. 218:15–28.
- Seybold SJ, Huber DPW, Lee JC, Graves AD, Bohlmann J (2006) Pine monoterpenes and pine bark beetles: a marriage of convenience for defense and chemical communication. Phytochem Rev. 5: 143–178.
- Reid ML, Purcell JRC (2011) Condition-dependent tolerance of monoterpenes in an insect herbivore. Arthropod-Plant Inter. 5: 331–337.
- Shrimpton DM, Reid RW (1973) Change in resistance of lodgepole pine to mountain pine beetle between 1965 and 1972. Can J For Res. 3:430–43.
Figure 1.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between monoterpenes and the annual resin duct characteristics of lodgepole pine trees in post-outbreak (2014A, 2015A) and 5-yr and 10-yr pre-outbreak (2014B, 2015B) periods in Jasper National Park (Alberta, Canada). RDP: Resin duct production; RDA: Total resin duct area; RDS: Individual resin duct size; RDD: Resin duct density; RDA%: Relative resin duct area; 5yB: 5-yr pre-outbreak; 10yB: 10-yr pre-outbreak.
Figure 1.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between monoterpenes and the annual resin duct characteristics of lodgepole pine trees in post-outbreak (2014A, 2015A) and 5-yr and 10-yr pre-outbreak (2014B, 2015B) periods in Jasper National Park (Alberta, Canada). RDP: Resin duct production; RDA: Total resin duct area; RDS: Individual resin duct size; RDD: Resin duct density; RDA%: Relative resin duct area; 5yB: 5-yr pre-outbreak; 10yB: 10-yr pre-outbreak.

Figure 2.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between diterpenes and the annual resin duct characteristics of lodgepole pine trees in post-outbreak (2014A, 2015A) and 5-yr and 10-yr pre-outbreak (2014B, 2015B) periods in Jasper National Park (Alberta, Canada). RDP: Resin duct production; RDA: Total resin duct area; RDS: Individual resin duct size; RDD: Resin duct density; RDA%: Relative resin duct area; 5yB: 5-yr pre-outbreak; 10yB: 10-yr pre-outbreak.
Figure 2.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between diterpenes and the annual resin duct characteristics of lodgepole pine trees in post-outbreak (2014A, 2015A) and 5-yr and 10-yr pre-outbreak (2014B, 2015B) periods in Jasper National Park (Alberta, Canada). RDP: Resin duct production; RDA: Total resin duct area; RDS: Individual resin duct size; RDD: Resin duct density; RDA%: Relative resin duct area; 5yB: 5-yr pre-outbreak; 10yB: 10-yr pre-outbreak.
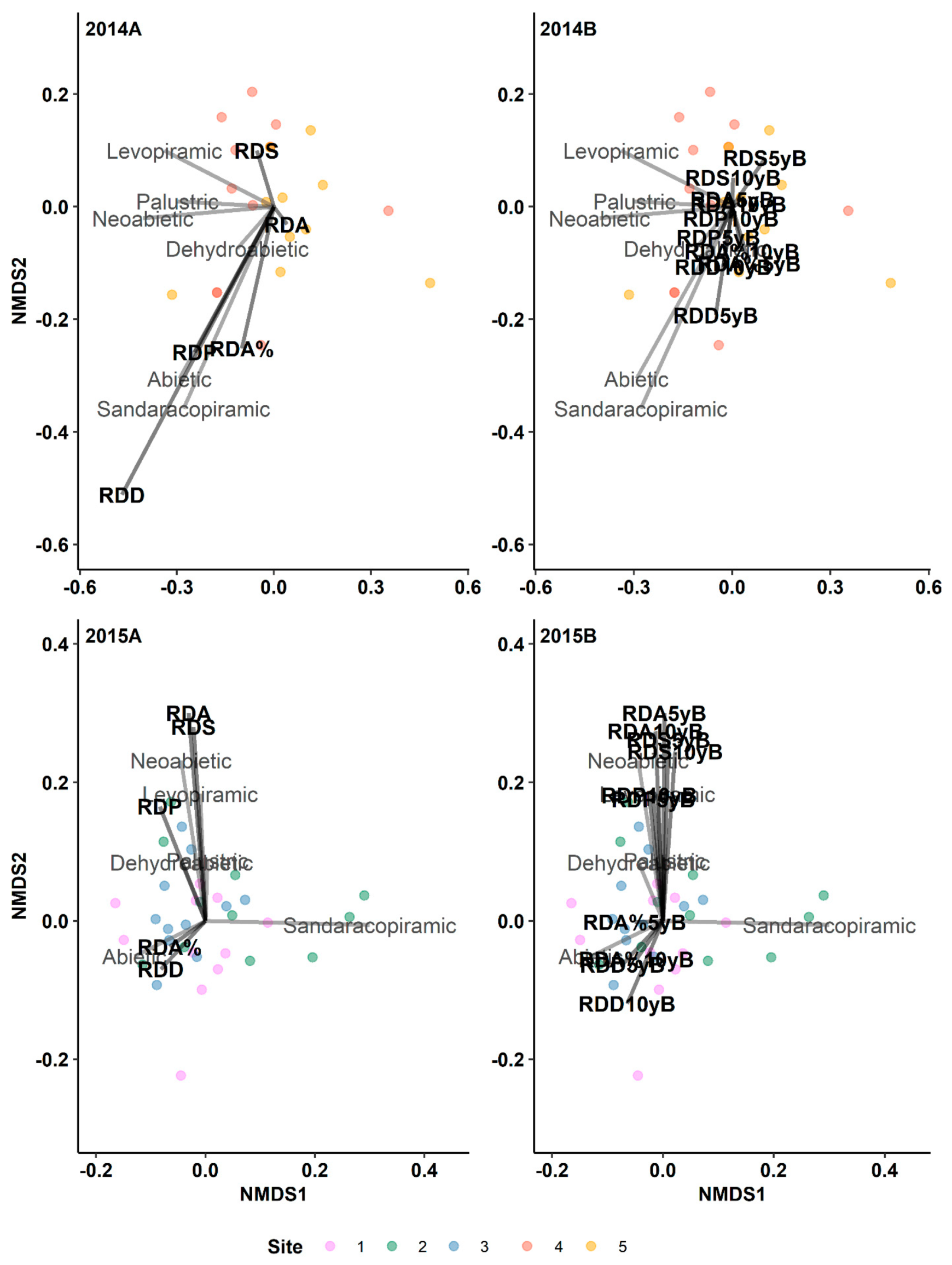
Figure 3.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between monoterpenes and the annual basal area increment (BAI, mm2 yr-1) of lodgepole pine trees in post-outbreak (2014A, 2015B) and 5-yr and 10-yr pre-outbreak (2014B, 2015B) periods in Jasper National Park (Alberta, Canada). 5yB: 5-yr pre-outbreak; 10yB: 10-yr pre-outbreak.
Figure 3.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between monoterpenes and the annual basal area increment (BAI, mm2 yr-1) of lodgepole pine trees in post-outbreak (2014A, 2015B) and 5-yr and 10-yr pre-outbreak (2014B, 2015B) periods in Jasper National Park (Alberta, Canada). 5yB: 5-yr pre-outbreak; 10yB: 10-yr pre-outbreak.
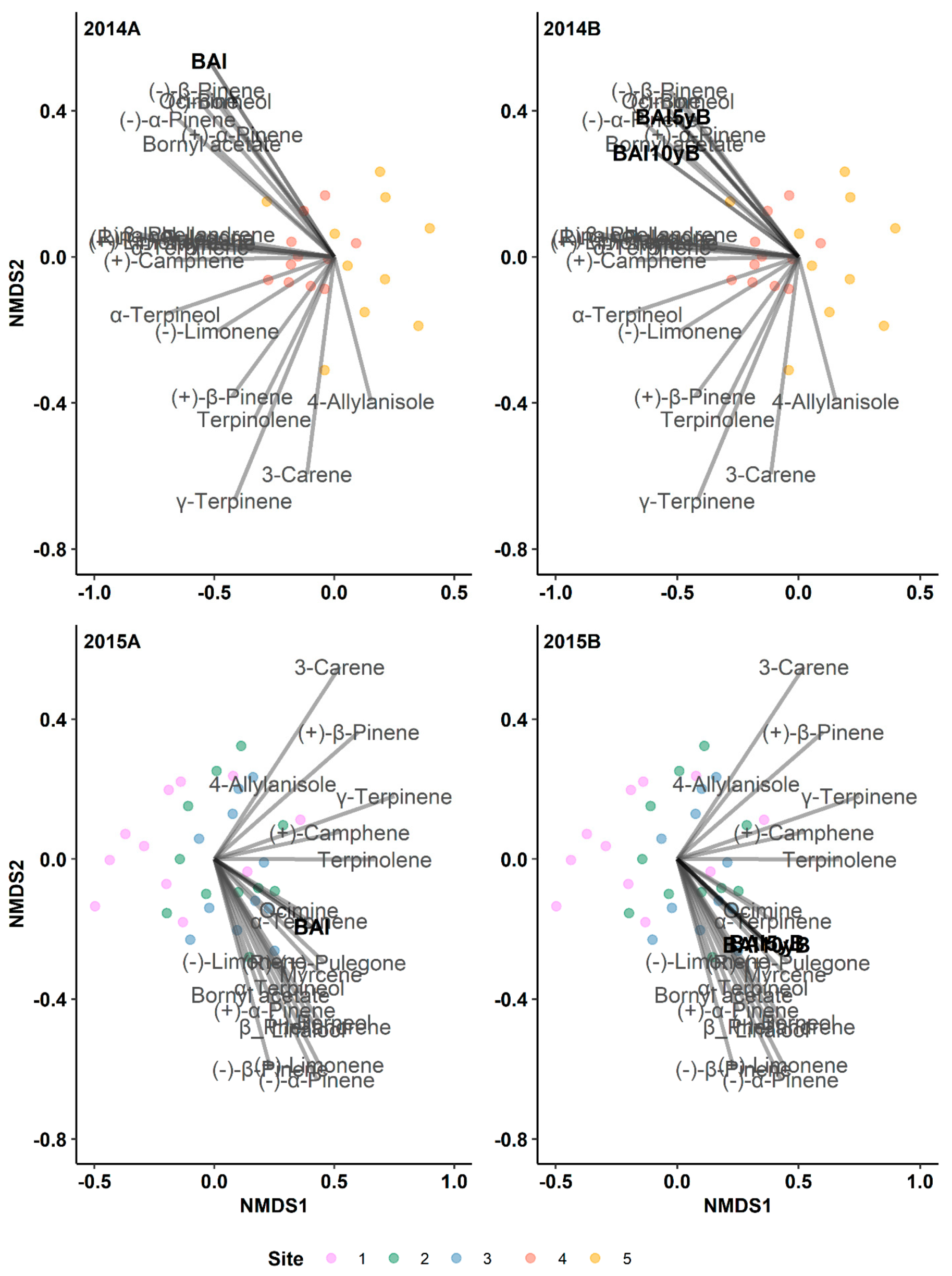
Figure 4.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between diterpenes and the annual basal area increment (BAI, mm2 yr-1) of lodgepole pine trees in post-outbreak (2014A, 2015B) and 5-yr and 10-yr pre-outbreak (2014B, 2015B) periods in Jasper National Park (Alberta, Canada). 5yB: 5-yr pre-outbreak; 10yB: 10-yr pre-outbreak.
Figure 4.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between diterpenes and the annual basal area increment (BAI, mm2 yr-1) of lodgepole pine trees in post-outbreak (2014A, 2015B) and 5-yr and 10-yr pre-outbreak (2014B, 2015B) periods in Jasper National Park (Alberta, Canada). 5yB: 5-yr pre-outbreak; 10yB: 10-yr pre-outbreak.
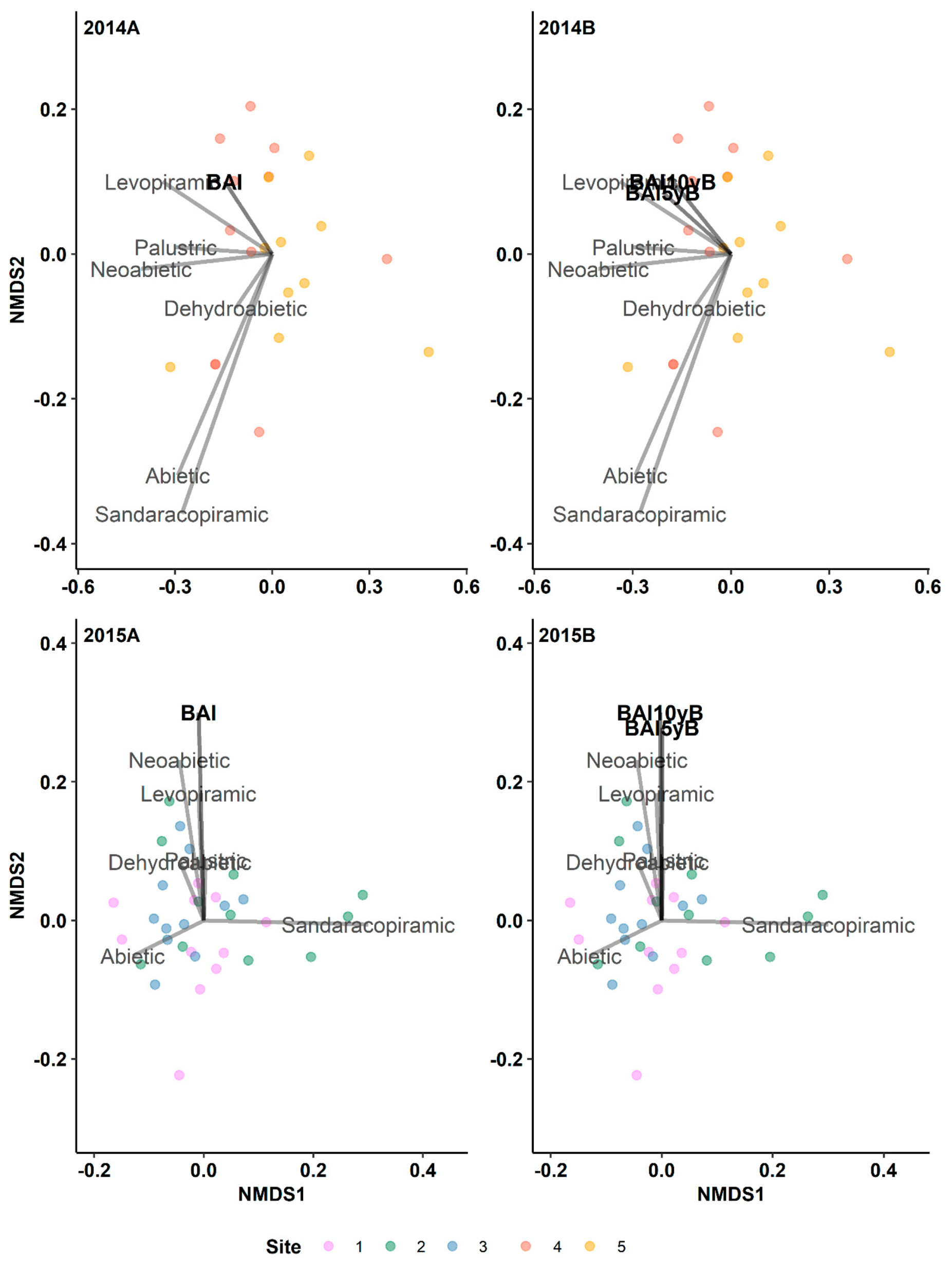
Figure 5.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between monoterpenes and diameter at breast height (DBH) and age of lodgepole pine trees in Jasper National Park (Alberta, Canada) for different outbreak years.
Figure 5.
Results of indirect gradient analysis by NMDS with Bray–Curtis dissimilarity showing the relationship between monoterpenes and diameter at breast height (DBH) and age of lodgepole pine trees in Jasper National Park (Alberta, Canada) for different outbreak years.
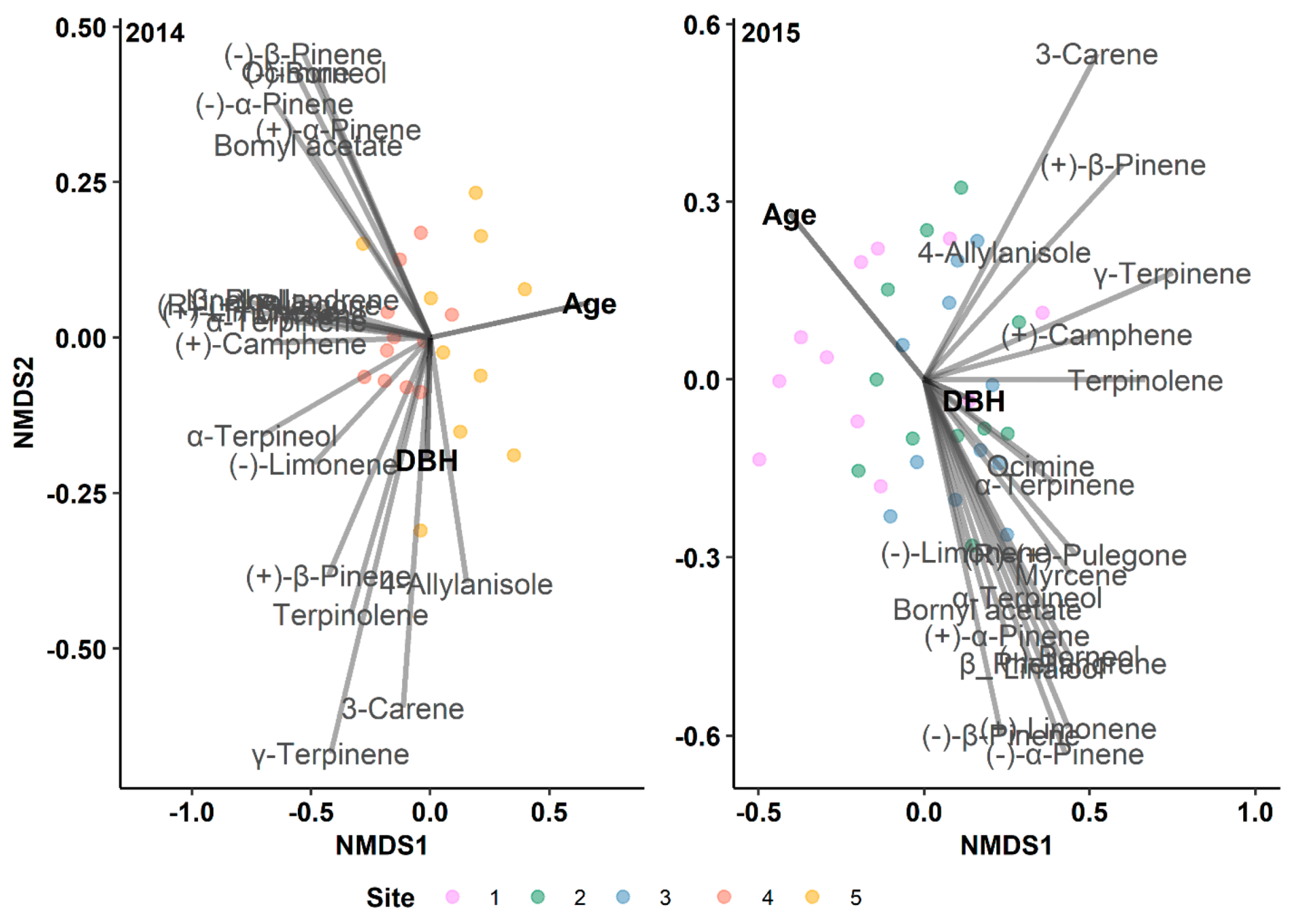
Figure 6.
Mean (±SE) resin duct production (a), total resin duct area (b), resin duct size (c), resin duct density (d), relative resin duct area (e), and basal area increment (f) of lodgepole pine trees in 5-yr and 10-yr prior- and post-outbreak periods in the 2014 outbreak. Significant differences among periods were indicated by different letters.
Figure 6.
Mean (±SE) resin duct production (a), total resin duct area (b), resin duct size (c), resin duct density (d), relative resin duct area (e), and basal area increment (f) of lodgepole pine trees in 5-yr and 10-yr prior- and post-outbreak periods in the 2014 outbreak. Significant differences among periods were indicated by different letters.
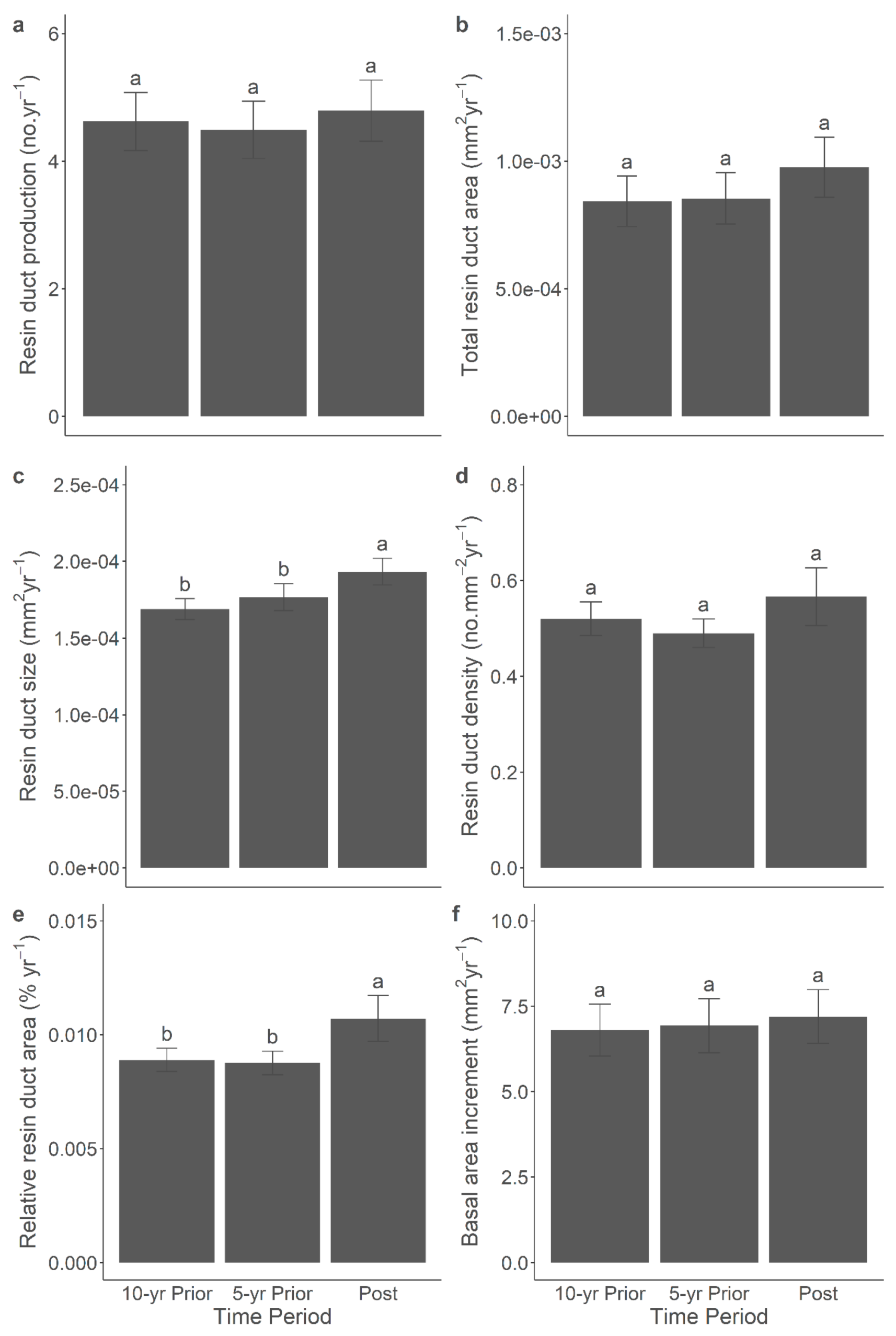
Figure 7.
Mean (±SE) resin duct production (a), total resin duct area (b), resin duct size (c), resin duct density (d), relative resin duct area (e), and basal area increment (f) of lodgepole pine trees in 5-yr and 10-yr prior- and post-outbreak periods in the 2015 outbreak. Significant differences among periods were indicated by different letters.
Figure 7.
Mean (±SE) resin duct production (a), total resin duct area (b), resin duct size (c), resin duct density (d), relative resin duct area (e), and basal area increment (f) of lodgepole pine trees in 5-yr and 10-yr prior- and post-outbreak periods in the 2015 outbreak. Significant differences among periods were indicated by different letters.
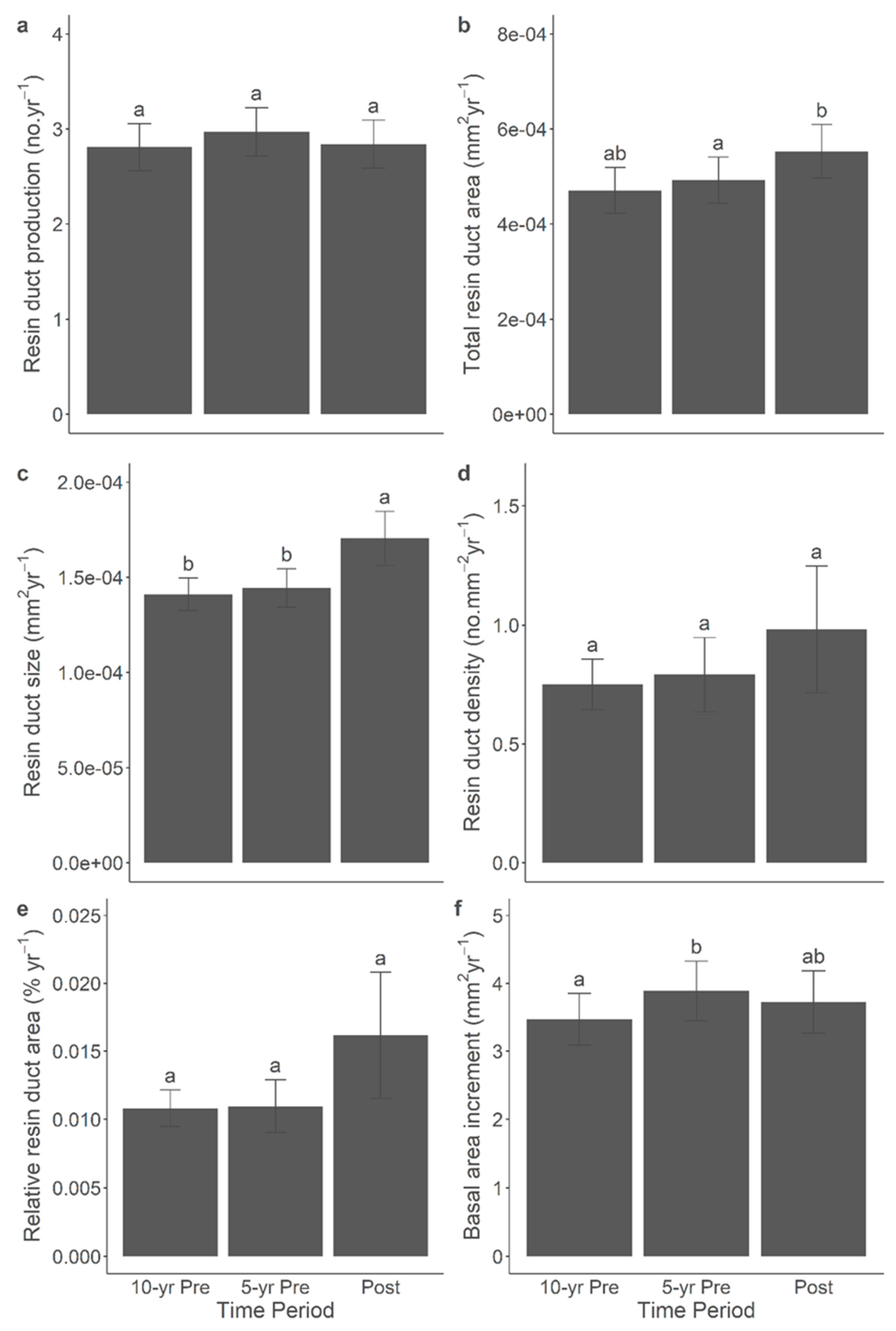
Table 1.
Results of linear mixed-effect models showing the relationship between total monoterpenes and each of the five annual resin duct characteristics of lodgepole pine trees in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years and periods.
Table 1.
Results of linear mixed-effect models showing the relationship between total monoterpenes and each of the five annual resin duct characteristics of lodgepole pine trees in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years and periods.
| Outbreak Years | Period | Variables* | Total monoterpenes | ||
| Coefficient | t-value | P-value | |||
| 2014 | Post-outbreak | Resin duct production | 1.1e3 | 4.3 | <0.001 |
| Total resin duct area | 4.8e6 | 4.8 | <0.001 | ||
| Individual resin duct size | 4.2e7 | 2.4 | 0.029 | ||
| Resin duct density | 1.4e3 | 0.5 | 0.626 | ||
| Relative resin duct area (%) | 2.2e5 | 1.4 | 0.187 | ||
| 5-yr pre-outbreak | Resin duct production | 1.1e3 | 3.1 | 0.005 | |
| Total resin duct area | 4.8e6 | 3.2 | 0.005 | ||
| Individual resin duct size | 3.0e7 | 1.7 | 0.107 | ||
| Resin duct density | 2.7e3 | 0.5 | 0.637 | ||
| Relative resin duct area (%) | 4.2e5 | 1.4 | 0.189 | ||
| 10-yr pre-outbreak | Resin duct production | 1.4e3 | 4.2 | <0.001 | |
| Total resin duct area | 6.2e6 | 4.1 | <0.001 | ||
| Individual resin duct size | 6.0e7 | 2.4 | 0.025 | ||
| Resin duct density | 1.5e3 | 0.3 | 0.761 | ||
| Relative resin duct area (%) | 4.2e5 | 1.3 | 0.219 | ||
| 2015 | Post-outbreak | Resin duct production | 78.7 | 0.2 | 0.831 |
| Total resin duct area | 3.4e6 | 2.1 | 0.045 | ||
| Individual resin duct size | 2.3e7 | 4.2 | <0.001 | ||
| Resin duct density | -100.8 | -0.3 | 0.767 | ||
| Relative resin duct area (%) | -1.3e3 | -0.1 | 0.946 | ||
| 5-yr pre-outbreak | Resin duct production | 584.0 | 1.6 | 0.127 | |
| Total resin duct area | 5.8e6 | 3.3 | 0.002 | ||
| Individual resin duct size | 3.0e7 | 3.7 | 0.002 | ||
| Resin duct density | -127.4 | -0.2 | 0.830 | ||
| Relative resin duct area (%) | -8.0e3 | -0.2 | 0.868 | ||
| 10-yr pre-outbreak | Resin duct production | 741.6 | 1.8 | 0.081 | |
| Total resin duct area | 6.2e6 | 3.3 | 0.002 | ||
| Individual resin duct size | 4.0e7 | 4.1 | <0.001 | ||
| Resin duct density | -280.3 | -0.3 | 0.762 | ||
| Relative resin duct area (%) | -6.8e3 | -0.1 | 0.926 | ||
Significant P-values (α=0.05) were bolded and italicized.
Table 2.
Results of linear mixed-effect models showing the direction of relationships between individual monoterpenes and the selected anatomical defense characteristics of lodgepole pine in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years and periods.
Table 2.
Results of linear mixed-effect models showing the direction of relationships between individual monoterpenes and the selected anatomical defense characteristics of lodgepole pine in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years and periods.
| Monoterpenes | 2014-Outbreak | 2015-Outbreak | ||||||||||||
| Post-outbreak | 5-yr pre-outbreak | 10-yr pre-outbreak | Post-outbreak | 5-yr pre-outbreak | 10-yr pre-outbreak | |||||||||
| RDP | RDA | RDS | RDP | RDA | RDP | RDA | RDS | RDA | RDS | RDA | RDS | RDA | RDS | |
| 4-Allylanisole | ||||||||||||||
| (-)-Borneol | ++ | ++ | ++ | ++ | + | + | + | + | ++ | + | + | + | ++ | |
| Bornyl acetate | + | + | + | + | + | + | ||||||||
| (+)-Camphene | ++ | ++ | ++ | ++ | ++ | ++ | + | +++ | + | +++ | +++ | |||
| 3-Carene | + | + | + | |||||||||||
| (+)-Limonene | +++ | +++ | + | ++ | ++ | +++ | +++ | + | +++ | ++ | ++ | ++ | +++ | |
| (-)-Limonene | ||||||||||||||
| Linalool | + | ++ | ++ | +++ | +++ | ++ | ++ | + | ++ | + | ++ | ++ | ++ | |
| Myrcene | ++ | +++ | ++ | ++ | +++ | +++ | + | +++ | ++ | +++ | ++ | ++ | ||
| Ocimene | ++ | ++ | +++ | +++ | +++ | +++ | ||||||||
| β-Phellandrene | ++ | ++ | + | + | ++ | +++ | +++ | + | +++ | + | ++ | + | +++ | |
| (+)-α-Pinene | + | ++ | + | + | + | |||||||||
| (-)-α-Pinene | +++ | +++ | + | +++ | +++ | +++ | +++ | + | + | +++ | +++ | ++ | +++ | +++ |
| (+)-β-Pinene | + | ++ | + | + | + | + | + | ++ | + | + | + | + | + | ++ |
| (-)-β-Pinene | ++ | +++ | ++ | + | ++ | ++ | ++ | + | ++ | +++ | ||||
| β-Phellandrene | ++ | ++ | + | + | ++ | +++ | +++ | + | +++ | + | ++ | + | +++ | |
| (R)-(+)-Pulegone | + | + | ++ | ++ | + | ++ | ++ | |||||||
| α-Terpinene | + | + | + | |||||||||||
| γ-Terpinene | + | + | + | ++ | + | + | ++ | ++ | + | ++ | ||||
| α-Terpineol | +++ | ++ | + | +++ | ||||||||||
| Terpinolene | ++ | +++ | + | + | + | |||||||||
RDP: Resin duct production; RDA: Total resin duct area; RDS: Individual resin duct size. + denotes positive and – denotes negative. *+: P-value ≤ 0.05; ++**: P-value ≤ 0.01; +++***: P-value ≤ 0.001.
Table 4.
Results of linear mixed-effect models showing the direction of relationships between individual monoterpenes or diterpenes and the annual basal area increment (BAI) of lodgepole pine in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years and periods. Since none of the results for diterpenes were statistically different in the 2014-outbreak year, only the results from the 2015 outbreak year were presented.
Table 4.
Results of linear mixed-effect models showing the direction of relationships between individual monoterpenes or diterpenes and the annual basal area increment (BAI) of lodgepole pine in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years and periods. Since none of the results for diterpenes were statistically different in the 2014-outbreak year, only the results from the 2015 outbreak year were presented.
| Terpene classes | 2014-Outbreak | 2015-Outbreak | ||||
| Post-outbreak BAI |
5-yr pre-outbreak BAI |
10-yr pre-outbreak BAI |
Post-outbreak BAI |
5-yr pre-outbreak BAI |
10-yr pre-outbreak BAI |
|
| Monoterpenes | ||||||
| 4-Allylanisole | ||||||
| (-)-Borneol | +++ | + | ++ | |||
| Bornyl acetate | ++ | + | + | |||
| (+)-Camphene | ++ | ++ | ++ | + | + | + |
| 3-Carene | ||||||
| (+)-Limonene | + | + | ++ | ++ | +++ | +++ |
| (-)-Limonene | ||||||
| Linalool | +++ | +++ | +++ | +++ | ++ | +++ |
| Myrcene | ++ | ++ | +++ | ++ | +++ | +++ |
| Ocimene | + | + | ++ | |||
| β-Phellandrene | + | ++ | ++ | ++ | ++ | +++ |
| (+)-α-Pinene | ++ | + | ||||
| (-)-α-Pinene | +++ | ++ | +++ | ++ | +++ | +++ |
| (+)-β-Pinene | + | + | + | + | + | + |
| (-)-β-Pinene | ++ | + | ++ | ++ | ++ | |
| (R)-(+)-Pulegone | ++ | +++ | +++ | |||
| α-Terpinene | ||||||
| γ-Terpinene | + | ++ | ++ | |||
| α-Terpineol | + | +++ | +++ | +++ | ||
| Terpinolene | + | + | + | |||
| Diterpenes | ||||||
| Abietic | + | + | + | |||
| Dehydroabietic | ++ | ++ | +++ | |||
| Levopiramic | +++ | +++ | +++ | |||
| Neoabietic | +++ | +++ | +++ | |||
| Palustric | ++ | ++ | +++ | |||
| Sandaracopiramic | ||||||
+ denotes positive and – denotes negative relationship. +*: P-value ≤ 0.05; ++**: P-value ≤ 0.01; +++***: P-value ≤ 0.001.Furthermore, we detected a positive relationship between the total monoterpenes and BAI during pre- and post-outbreak periods in both the 2014 and 2015 outbreaks (Table 3). BAI also exhibited a significant and positive relationship with the total diterpenes across all pre- and post-outbreak periods in the 2015 outbreak (Table 3); however, in the 2014 outbreak, we did not detect any significant relationship between BAI and the total diterpenes.
Table 5.
Results of linear mixed-effect models showing the direction of relationships between individual monoterpenes and age of lodgepole pine trees in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years.
Table 5.
Results of linear mixed-effect models showing the direction of relationships between individual monoterpenes and age of lodgepole pine trees in Jasper National Park (Alberta, Canada) for different mountain pine beetle outbreak years.
| Tree age | ||
| Monoterpenes | 2014-Outbreak | 2015-Outbreak |
| 4-Allylanisole | ||
| (-)-Borneol | ||
| Bornyl acetate | ||
| (+)-Camphene | - | |
| 3-Carene | ||
| (+)-Limonene | - | -- |
| (-)-Limonene | ||
| Linalool | --- | -- |
| Myrcene | - | |
| Ocimene | ||
| β-Phellandrene | - | |
| (+)-α-Pinene | ||
| (-)-α-Pinene | - | -- |
| (+)-β-Pinene | - | |
| (-)-β-Pinene | - | |
| (R)-(+)-Pulegone | - | |
| α-Terpinene | - | |
| γ-Terpinene | -- | -- |
| α-Terpineol | -- | |
| Terpinolene | - | |
+ denotes positive and – denotes negative relationship. -: P-value ≤ 0.05; --: P-value ≤ 0.01; ---: P-value ≤ 0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



