
Submitted:
08 August 2024
Posted:
12 August 2024
You are already at the latest version
Abstract
Excess nitrogen in agricultural drainage poses a serious threat to the water quality safety of the Yellow River basin. Utilizing aquatic plants to modify the rhizosphere microbial community structure and facilitate nitrogen transformation is a crucial strategy for mitigating regional water eutrophication. Our results revealed that the denitrification rate (DR), dissimilatory nitrate reduction to ammonium rate (DNRAR) and nitrogen fixation rate (NFR) of rhizosphere sediments exhibited a significant increase in artificially cultivated ditch. Compared with those in natural ditches, the richness of the bacterial community and the relative abundances of Bacteroidota, Verrucomicrobiota, Firmicutes, Anaeromyxobacter and Geobacter, which are involved mainly in the nitrogen cycle, in the rhizosphere sediments of artificially cultivated ditch were significantly greater due to improvements in environmental conditions. In artificially cultivated ditch, the dominant functional genes affecting DRNARs in the rhizosphere sediments of P. australis were nrfC and nrfA, while DRs were driven mainly by norB and napA, which were influenced by the nitrogen and carbon levels; and the dominant functional genes affecting NFRs in the rhizosphere sediments of T. orientalis were nifD, nifK and nifH. Our results provide a scientific basis for the use of aquatic plants for mitigating excess nitrogen levels in agricultural drainage.
Keywords:
denitrification
; DNRA
; functional gene of nitrogen transformation
; rhizosphere sediments
; Phragmites australis
; Typha orientalis
1. Introduction
The oasis irrigation area in Northwest China is an arid inland region that plays a pivotal role in the economic development and food security of the Yellow River Basin [1,2]. The Ningxia Yellow River Irrigation Area is a significant agricultural irrigation region and commodity grain base in Northwest China. Grain crops and agricultural outputs account for more than two-thirds of the overall economic value in Ningxia Province, thereby driving regional economic development [3]. However, extensive agricultural practices, such as the excess use of fertilizers and pesticides, as well as the overdischarge of irrigation water, have resulted in excess nitrogen in the water and sediment of irrigation areas [4]. The discharge of agricultural runoff into the Yellow River has led to water eutrophication in the Yellow River Basin. The third drainage ditch in Ningxia, which is a significant inflow ditch into the Yellow River in Ningxia, exhibits concentrations of ammonium nitrogen (NH4+-N) and total nitrogen (TN) ranging from 7.4 mg/L to 25.4 mg/L and from 17.7 mg/L to 28.2 mg/L, respectively, and has long been classified as a severely polluted class V water body [5]. Therefore, the effective control and reduction of the nitrogen concentration in the water and sediment of farmland drainage ditches in the Yellow River irrigation area of Ningxia is crucial for preventing and controlling eutrophication in irrigation areas and in the Yellow River [6], and it is also a new frontier in watershed ecological environment research and pollution prevention and control technology [7,8].
The drainage ditch system serves as the primary conduit for field drainage and has developed into a dual-function system that not only facilitates water drainage but also supports a unique “plant–sediment–microorganism” wetland ecosystem [9,10]. This system plays a crucial role in the purification of nitrogen, phosphorus, and organic pollutants in field runoff water, thereby enhancing the water quality within the irrigation area. As depicted in Figure 1, in the anaerobic environment of wetlands, the principal processes of the nitrogen cycle, such as denitrification, dissimilatory nitrate reduction to ammonium (DNRA), nitrogen fixation, nitrification, and ammonia oxidation, are predominantly driven by microorganisms [11,12]. Denitrifying microorganisms convert active nitrogen (NO3-, NH4+) into N2 or N2O and then fix it, which is an important way to remove nitrogen from farmland drainage [13]. The nirS, nirK and nosZ genes serve as crucial indicators for assessing the structure and function of denitrifying microbial communities [14,15]. DNRA microorganisms can convert NO3- into NH4+, a more biologically available form of nitrogen. NH4+ can then participate in nitrogen cycling pathways such as ammonia oxidation to produce N2 or N2O [16,17]. Therefore, these microorganisms also have the potential to effectively remove nitrogen from water bodies. The nrfA gene serves as a functional gene marker that accurately reflects the activity of DNRA [18]. Nitrogen fixation is the process through which microorganisms fix nitrogen and sequester it in sediments, thereby supplying water-dwelling organisms with accessible nitrogen [19]. The nifH, nifD and nifK genes serve as pivotal gene markers for nitrogen fixation [20]. Plants play a crucial role in shaping the diversity and composition of microbial communities and in mediating nitrogen transformation in the rhizosphere [21]. Specifically, they influence the structure of nitrogen-transforming microbial communities through root-zone organic carbon [22], thereby impacting the nitrogen cycle. Conversely, the diversity and composition of rhizosphere microbes also have implications for plant health and biochemical processes in rhizosphere soil [23].
Aquatic plants primarily eliminate pollutants from water and sediments through processes such as plant absorption and rhizosphere microbial degradation [24]. The physiological and ecological processes of photosynthesis, respiration, and transpiration in aquatic plants drive nutrient absorption [25]. As aquatic plants release oxygen through photosynthesis, they not only satisfy their own needs but also elevate the DO levels in the water body [26]. Additionally, they create a microenvironment favorable for microbial metabolism in the rhizosphere [27], thereby promoting the microbial conversion and decomposition of nitrogen in the aquatic environment [28]. Therefore, the rhizosphere effect of aquatic plants enhances the nitrogen cycle process of drainage ditches, thereby facilitating water purification and ecological restoration [29]. Studies have demonstrated that emergent plant roots facilitate the transfer of oxygen from the atmosphere to sediments, thereby regulating the DO concentration in the rhizosphere [30] and providing habitats for microorganisms [31]. The concentration of DO in the rhizosphere is directly correlated with the diversity, abundance, and metabolic activity of nitrogen cycle functional bacteria [30]. Furthermore, emergent plants alter the abundance and composition of ammonia-oxidizing microbial communities in sediments through root oxygen secretion [26]. The functional microorganisms that accumulate in the rhizosphere of Phragmites australis facilitate the degradation and transformation of pollutants, thereby enhancing the purification efficiency and ecological stability of wetland ecosystems [32,33].
Plants have been shown to significantly decrease the electrical conductivity (EC) and nutrient content of sediment in ditches, resulting in a notable increase in the abundance of nitrifying bacteria, nitrifying spiral bacteria and denitrifying α-Proteobacteria in sediments [34], which effectively mitigates nitrogen pollution in water [35]. Drainage ditches with floating plants exhibit a greater denitrification rate (DR) than those without vegetation or those with only submerged vegetation [36]. Compared to natural wetlands, artificial wetlands exhibit greater biomass production, greater nutrient content in plant tissue, greater shoot/root ratios, greater transport velocity [37], easier fixation of organic carbon from plants, and more stable microbial metabolic functions [38]. Compared with those in naturally growing P. australis wetlands, the α diversity of the bacterial community in the P. australis rhizosphere of artificial wetlands was greater, the similarity of the bacterial community composition was greater, and the interactions between bacterial groups were more intricate. Although the rhizosphere of P. australis in natural wetlands is significantly enriched in functional bacterial communities related mainly to nutrient cycling and promoting plant growth, the bacterial groups enriched in the rhizosphere of P. australis in artificial wetlands exhibit greater potential for biodegradation [39]. The artificial wetlands in the Dongping Lake Basin exhibit a significant advantage over the natural wetlands in terms of nitrogen metabolism-related gene abundance predicted by PICRUSt, indicating their potential for more complex nitrogen cycling functions and metabolic capabilities [38]. Tax4Fun analysis and prediction revealed a significantly greater abundance of denitrification (napA, norB, nosZ), dissimilatory nitrate reduction (nirB), and nitrogen fixation (nifD, nifK) genes in the artificial wetland of Ximen Island in Yueqing Bay than in the natural wetland, suggesting that the biological activity involved in nitrogen transformation processes may be more active in the artificial wetland [40]. The variations in rhizosphere microbial abundance among different plant species may be attributed to disparities in gas exchange characteristics and oxygen conditions. Compared to Typha orientalis, P. australis exhibits a greater microbial abundance and ammonia-oxidizing bacteria count, as well as a greater removal rate of NH4+-N [41]. The rhizosphere microorganisms P. australis and T. orientalis in artificial wetlands may play crucial roles in regulating nutrient cycling in the sediment by influencing the metabolism of active carbon and nitrogen [42].
Although previous studies have analyzed the structure and function of microbial communities in the rhizosphere sediments of aquatic plants, reports on the nitrogen transformation rate and the composition of key microbial functional communities, including their primary environmental impact factors, are limited. The process of nitrogen removal from agricultural runoff by aquatic plants and their rhizosphere microorganisms is a complex phenomenon that is influenced by a multitude of factors, such as climate, soil composition, hydrological characteristics, and plant species [43].
The Yellow River irrigation area in Ningxia is located in the arid region of the middle temperate zone in Northwest China and is characterized by unique climate, soil and hydrological conditions. This region experiences ample sunshine, significant temperature fluctuations, drought, minimal precipitation, and high evaporation rates. Rivers and lakes in this region exhibit elevated salinity levels, while the ecological attributes of aquatic plants are distinguished by their distinct growth patterns and nutrient absorption mechanisms. No research has reported on whether there are significant differences in rhizosphere nitrogen transformation rates, microbial community structure, or related functional microorganisms between artificially planted aquatic plant communities and naturally occurring aquatic plant communities in agricultural drainage ditches. It is also unknown whether artificially cultivated ditches are more effective at promoting nitrogen transformation and have a stronger nitrogen removal capacity in water bodies.
Therefore, the aim of our study was to investigate the nitrogen transformation rates in rhizosphere sediments of artificial planting and natural farmland drainage ditches and their influencing mechanisms, especially the diversity and structural composition of bacterial communities and key functional genes of the nitrogen cycle, to provide a scientific basis for using aquatic plants to reduce nitrogen pollution in water bodies. Rhizosphere sediments of P. australis and T. orientalis were collected from three drainage ditches in the Yellow River Irrigation Area of Ningxia, including ditches of artificially cultivated P. australis, T. orientalis and Nymphaea tetragona; ditches of naturally growing P. Australis; and ditches of naturally growing T. orientalis. The nitrogen transformation rate, bacterial community diversity, structural composition and key functional genes of the nitrogen cycle in all ditches were determined. We propose the following two hypotheses: (1) Compared with those in rhizosphere sediments in natural ditches, the nitrogen transformation rates in rhizosphere sediments in artificially cultivated ditches are greater, and (2) compared with those in rhizosphere sediments in natural ditches, the bacterial community diversity index of rhizosphere sediments in artificially cultivated ditches is greater, with a more complex composition and greater abundance of nitrogen transformation-related functional bacterial communities and functional genes, particularly for P. Australis rhizosphere soil.
2. Materials and Methods
2.1. Research Site and Sediment Sampling
The study site is located in Changxin township (38°32′64″N, 106°25′74″E), Helan County, Yinchuan city, Ningxia Hui Autonomous Region, China (Figure 2). The research area is a typical Yellow River irrigation area, with an altitude of 1106 m, and has a typical mid-temperate continental arid climate. The mean annual precipitation is 138.8 mm, the mean annual temperature is 9.7 °C, the cumulative temperature is 3280 °C, the mean annual sunshine is 2935.5 hours, and the total annual solar radiation is 140.9 kcal cm-2.
The farmland drainage ditches in the experimental area are trapezoidal earth ditches, and the soil type is irrigation-silted soil. The drainage ditches are dredged every 3-5 years. The three independent drainage ditches chosen for our investigation underwent cleaning in the spring of 2020, and their basic hydrological conditions were consistent: the water surface width was 1.55±0.16 m, the water depth was 0.41±0.27 m, the flow velocity was 0.14±0.073 m s-1, and the ditch length was 220±11.42 m.
For the artificially cultivated drainage ditch, after clearing the weeds in the ditch, according to our previous research results, combined with purification effect, adaptability, and landscape research, P. australis and T. orientalis plants were cross-planted on both sides of the ditch, and N. tetragona was planted in the middle of the ditch, with a row spacing of 30 cm, a spacing of 30 cm, and a density of 16 plants m-2. P. australis, T. orientalis and N. tetragona were purchased from local aquatic plant companies. The other two ditches were P. australis and T. orientalis natural growth drainage ditches. In the P. australis natural growth drainage ditch (natural ditch 1), the dominant species was P. australis, the companion species were Scirpus triqueter and Echinochloa crusgali, and the planting densities were 13 plants m-2, 4 plants m-2, and 1 plant m-2, respectively. In the T. orientalis natural growth drainage ditch (natural ditch 2), the dominant species was T. orientalis, the companion species were P. australis and S. triqueter, and the densities were 10 plants m-2, 3 plants m-2, and 2 plants m-2, respectively. The basic vegetation conditions of the 3 drainage ditches are shown in Table 1.
On July 28, 2022, three small quadrats (i.e., three replicates) with an area of 1 m×4 m were randomly set up in representative areas of each drainage ditch, and the spatial distance between the two small quadrats was at least 50 m. Five well-growing P. australis and T. orientalis plants were selected for collecting rhizosphere sediments in each quadrat. The sediments attached to the roots and within 5 cm of the roots of 5 plants were collected as rhizosphere sediments, and the rhizosphere sediments of the 5 plants in the same quadrat were mixed uniformly in equal proportions as a single sample. Each sediment sample was divided into 3 parts: the first sample was immediately sealed in a freezer tube, placed in liquid nitrogen for transport, and then stored in an ultralow temperature freezer at -80°C for microbial community analysis; the second sample was immediately placed in a portable refrigeration unit, and the nitrogen transformation rates were determined by culturing within 24 hours of collection; and the third sample was placed indoors to air dry naturally for the determination of the physical and chemical properties of the sediment.
Before collecting sediment samples, in situ measurements were conducted for water temperature (T-W), dissolved oxygen concentration (DO-W), pH (pH-W), and oxidation‒reduction potential (ORP-W) near the roots of 5 plant species. Additionally, water samples were obtained for the measurement of total nitrogen (TN-W), ammonium nitrogen (NH4+-W), and nitrate nitrogen (NO3--W) concentrations.
2.2. Nitrogen Transformation Rate in Sediment
The potential denitrification, DNRA, and nitrogen fixation rates were determined using a combination of slurry cultivation experiments and 15N isotope tracing techniques [44,45,46].
- In the slurry preculture experiment, fresh sediment (20 g) was mixed thoroughly with in situ overlying water (1:7 w/v). The mixture was homogenized by rinsing with high-purity helium (purity >99.999%) for 30 minutes and transferred to a 200 mL hermetic vial (Exetainer, Labco, UK). The vial was sealed with a butyl rubber septum and incubated for 24 hours in the dark.
- For the 15N isotope tracing experiment, after the precultivation experiment, the vials were divided into two groups, and 15N-labeled sodium nitrate solution (Na15NO3, 99% 15N, Cambridge Isotope Laboratories, Inc., Tewksbury, USA) was added to achieve a final 15N-labeled solution concentration of 100 μM. One group of vials was injected with 0.2 mL of ZnCl2 solution at a 50% w/v concentration, which was the initial sample. Another group of vials was placed in a constant-temperature incubator for further cultivation for 8 hours. After the cultivation was completed, the groups of vials were injected with ZnCl2 solution as the final samples.
- For the analysis of potential DNRA rates (DNRARs), after the slurry precultivation experiment and 15N isotope tracing experiment were completed, 200 μL of hypobromic acid oxidant was added to the initial and final sample vials. Subsequently, the difference in the concentration of 15NH4+-N in the initial and final sample vials was analyzed using MIMS to calculate the potential DNRARs [48,49].
- For the analysis of potential nitrogen fixation rates (NFRs, after the slurry precultivation experiment was completed, the vials were divided into two groups. One group of vials was injected with 0.2 mL of ZnCl2 solution at a 50% w v-1 concentration, which was the initial sample. Another group of vials was injected with 0.5 mL of 99 atom% 15N-N2 (Campro Scientific, Germany), sealed and cultured at in situ temperature for 24 h. After cultivation was completed, the group of vials was injected with ZnCl2 solution, which was the final sample. Subsequently, all the vials were washed with He for 30 minutes to remove N2, and iodine hypobromide solution was injected to oxidize the 15N label produced by fixing N into N2. Subsequently, the difference in the 15N concentration between the initial and final sample vials was analyzed via MIMS to calculate the potential NFRs [50].
The detailed calculation formulas for the potential DRs, DNRARs and NFRs are listed in the supplementary material.
2.3. Analysis of Sediment and Water Physical and Chemical Properties
The EC of the sediment samples was measured using a conductivity meter (DDS-307, Raymon, Shanghai, China). The total organic carbon (TOC) content in the sediments was determined by the potassium dichromate-concentrated sulfuric acid titration method. The dissolved organic carbon (DOC) content in the sediment was extracted using a 2 mol L-1 KCl solution at a fresh sediment-to-solution ratio of 1:5 and quantified using a TOC analyzer (TOC-L, Shimadzu, Kyoto, Japan). The TN content was determined by the semimicroKjeldahl method. The NH4+-N and NO3--N in the sediments were extracted with 2 mol L-1 KCl at a soil:water ratio of 1:4, and the contents were determined by continuous-flow autoanalyzer measurements (AA3, Seal, UK). The sizes of sediment sand particles and clay particles were determined through a combination of sieving and sediment gravity meter analysis [51].
The T-W, DO-W, pH-W, and ORP-W values were determined using a multiparameter water quality probe (HACH HQ40d, HACH, USA). The TN content in the water samples was determined by alkaline potassium persulfate oxidation and ultraviolet spectrophotometry. The NH4+-W and NO3—W contents in the water samples were measured using a continuous-flow autoanalyzer (AA3, Seal, UK).
2.4. High-Throughput Sequencing and Metagenomic Sequencing
Microbial community genomic DNA was extracted using a soil DNA kit (DP812, Tiangen Biochemical Technology (Beijing) Co., Ltd.) according to the manufacturer’s instructions. The full-length bacterial 16S rRNA gene was amplified using primers 27F_(16S-F) (5′-AGRGTTTGATYNTGGCTCAG-3′) and 1492R_(16S-R) (5′-TASGGHTACCTTGTTASGA CTT-3′) [52] by a PCR instrument (Veriti96well 9902, ABI, USA). After amplification, the PCR products were recovered through 2% agarose gel electrophoresis, purified, detected, and quantified. Subsequently, library construction was performed using the SMRTbell Template Prep Kit. Following the standard protocol of Biomarker Technologies (Beijing, China), sequencing of the samples was conducted on the PacBio Sequel II platform. Usearch software was used to cluster reads at a similarity level of 97.0% and obtain OTUs. Sample alpha diversity indices were evaluated using QIIME2 2020.6 software [53]. Taxonomic annotation was performed using the Silva 138 database (Release 138, https://www.arb-silva.de/documentation/release-138/) [54].
Metagenomic sequencing was performed using the paired-end sequencing method to construct small fragment libraries, which were sequenced on the Illumina Nova Seq 6000 (Illumina, USA) platform. MMseqs2 software (https://github.com/soedinglab/mmseqs2, Version 12-113e3) was used to remove redundancy, with the similarity threshold set at 95% and the coverage threshold set at 90%. By utilizing multiple databases, such as KEGG, for sequence alignment, the predicted gene protein sequences were annotated with functional information. The abundance of related bacteria involved in the nitrogen cycle was analyzed via the KEGG database (https://www.kegg.jp/) [55]. The abundance of functional genes involved in the nitrogen cycle was analyzed via the NCycDB database (https://github.com/qichao1984/NCyc) [56].
2.5. Statistical Analysis
The differences in nitrogen transformation rate and physical and chemical properties between different treatments were determined to be statistically significant at a level of p<0.05 using one-way ANOVA and LSD tests. Pearson correlation analysis was employed to examine the associations among the nitrogen transformation rate, microbial abundance, and environmental factors. Redundancy analysis (RDA) was employed to examine the relationships among the rate of nitrogen transformation, microbial abundance, and environmental factors. All the statistical analyses were carried out using R software (version 3.1.2; R Development Core Team 2016).
3. Results
3.1. Nitrogen Transformation Rates in Rhizosphere Sediments
The DRs of artificially cultivated ditch P. australis (A-Phr) and natural ditch P. australis (N-Phr) were significantly greater than those of artificially cultivated ditch T. orientalis (A-Typ) and natural ditch T. orientalis (N-Typ) (p<0.05), reaching 3.06 and 3.08 μmol N kg-1 h-1, respectively, which were 17.67-22.70% and 18.31-23.36% greater than those of N-Typ and A-Typ, respectively (Figure 3a). The DNRAR of A-Phr was significantly greater than that of A-Typ and N-Typ (p<0.05), reaching 2.14 μmol N kg-1 h-1, which was 31.56%, 29.96%, and 11.07% greater than those of A-Typ, N-Typ, and N-Phr, respectively (Figure 3b). The NFR of A-Typ was significantly greater than that of N-Phr (p<0.05), reaching 0.93 μmol N kg-1 h-1, which was 46.60% and 28.44% greater than those of N-Phr and N-Typ, respectively (Figure 3c).
3.2. Diversity and Composition of the Bacterial Community in Rhizosphere Sediments and Their Driving Factors
A total of 144,287 high-quality sequences were obtained through 16S rRNA sequencing, with each library containing a varying number of sequences ranging from 10,177 to 14,218. The coverage rate for all the samples exceeded 96%. Utilizing a sequence similarity threshold of >97%, high-quality reads were grouped into 5,411 microbial operational taxonomic units (OTUs), resulting in a total of 109,493 sequences (Figure S1). The Chao1 and ACE richness indices of the A-Phr and A-Typ bacterial communities were significantly greater than those of the N-Phr and N-Typ bacterial communities (Figure 4a, 4b). In contrast, the Shannon diversity index of N-Typ was notably greater than that of N-Phr (Figure 4c).
The results of the RDA (Figure 5a) indicate that the first and second principal axes account for 59.14% and 23.85% of the variation, respectively, resulting in a cumulative explanatory rate of 82.99%. The Chao1 and ACE richness indices exhibited significant positive correlations with ORP and NH4+-N values, while the Shannon diversity index showed a significant positive correlation with EC and TOC content. The main environmental factors influencing the richness and diversity of the bacterial communities were ORP-W and NH4+-N and TOC contents (Table S3).
According to the 16S rRNA sequencing results, a total of 37 bacterial phyla were identified, 27 of which exhibited a relative abundance exceeding 1%. Among these, Proteobacteria (18-31%), Bacteroidota (17-28%), Acidobacteriota (3-14%), Verrucomicrobiota (5-10%), and Patescibacteria (2-10%) emerged as the predominant phyla in rhizosphere sediments, displaying a mean relative abundance surpassing 5% and collectively representing 67-70% of bacterial sequences (Figure 4d).
The RDA results (Figure 5b) indicated that the explanatory rates of the first and second axes were 55.82% and 15.38%, respectively, with a cumulative explanatory rate of 71.20%. The predominant bacterial phyla in A-Phr were Bacteroidota, Verrucomicrobiota, Patescibacteria, and Firmicutes; those in A-Typ were Bacteroidota and Firmicutes; those in N-Phr were Proteobacteria and Acidobacteriota; and that in N-Typ was only Acidobacteriota (Figure 4d). The main environmental factors affecting the composition of the bacterial community structure were the sediment DOC, TN, TOC, and sand contents and water DO-W content (Table S3).
The microbial genera involved in the N cycle primarily consisted of Anaeromyxobacter, Pseudomonas, Nitrospira, Geobacter, and Thiobacillus, with relative abundances ranging from 0.26% to 3.66%, 0.22% to 2.22%, 0.22% to 1.69%, 0.14% to 2.01%, and 0.11% to 1.65%, respectively. The relative abundances of Anaeromyxobacter, Geobacter, and Sulfuritalea were significantly greater in the A-Phr group than in the control group (p<0.05), while the relative abundances of Thiobacillus, Hydrogenophaga, and Sulfuritalea were significantly greater in the A-Typ group than in the control group (p<0.05) (Figure 4e). All these microbial genera belong to the Proteobacteria phylum, which is a predominant genus in the denitrification or DNRA pathway.
The RDA results (Figure 5c) demonstrated that the first and second ordination axes accounted for 52.73% and 23.34% of the variation, respectively, resulting in a cumulative explanation rate of 76.07%. Specifically, Anaeromyxobacter and Geobacter exhibited significant positive correlations with the DOC/NO3--N ratio and the NH4+-N content (p<0.05) and a significant negative correlation with the NO3--N content (p<0.05) (Figure 5c). The primary environmental factors influencing the composition of bacterial genera involved in the N cycle are sediment the NH4+-N and DOC contents and the DOC/NO3--N ratio (Table S3).
3.3. Relative Abundance of Nitrogen Transformation Functional Genes in Rhizosphere Sediments and Their Driving Factors
According to the annotation results of the Ncyc database, a total of 68 gene (sub)families were identified, encompassing 8 processes related to the nitrogen cycle. The relative abundances of functional genes involved in organic degradation and synthesis pathways ranged from 57.01% to 61.12%, while those involved in denitrification ranged from 13.89% to 18.61%, those involved in assimilatory nitrate reduction ranged from 8.23% to 12.76%, those involved in dissimilatory nitrate reduction ranged from 8.84% to 10.46%, those involved in nitrogen fixation ranged from 0.58% to 10.05%, those involved in nitrification ranged from 0.10% to 1.62%, those involved in nitrification ranged from 0.19% to 0.60%, and those involved in anammox ranged from 0.003% to 0.07% (Figure 6a).
The results of the RDA (Figure 5d) indicated that the first and second principal axes accounted for 49.71% and 33.76% of the data variation, respectively, resulting in a cumulative explanatory rate of 83.47%. The key environmental factors influencing participation in the N cycle include NH4+-N content, DOC/NO3--N ratio, and NO3--N content (Table S3).
The relative abundances of genes involved in A-Phr denitrification (napA, norB), dissimilatory nitrate reduction (nrfC, nrfA), nitrification (hao), and anammox (hzsA, hzsC) were significantly greater in A-Phr than in A-Typ, N-Phr, and N-Typ (p<0.05). Among these genes, the norB, nrfC, nrfA, hao, hzsA, and hzsC genes were all significantly positively correlated with the NH4+-N content and DOC/NO3--N ratio (p<0.01) but negatively correlated with the NO3--N content (p<0.01) (Figure 6b, 6c; Table S4). The relative abundances of nitrogen fixation genes (nifD, nifK, nifH) in A-Typ were significantly greater than those in A-Phr, N-Phr and N-Typ (p<0.05) and showed a significant positive correlation with the ORP-W and DO-W content (p<0.05). The relative abundances of denitrification (nirK, nosZ, nirS) genes in N-Phr were significantly greater than those in A-Phr, A-Typ and N-Typ (p<0.05) and displayed a significant negative correlation with the TOC and TN contents, ORP-W and DO-W content (p<0.05), among which the nirK and nosZ gene abundances were significantly positively correlated with the DOC content (p<0.05) (Figure 6b, Table S4).
3.4. Influence of Environmental and Microbial Factors on the Nitrogen Conversion Rate of Sediment
The relative abundances of DR-related and functional genes involved in the denitrification pathway exhibited a significant positive correlation (p<0.01), primarily with the relative abundance of norB (p<0.01). Similarly, the relative abundances of DNRAR-related and functional genes involved in the DNRA pathway showed a significant positive correlation (p<0.01), mainly with the relative abundances of nrfC and nrfA (p<0.01). Additionally, the relative abundances of NFR-related and functional genes involved in the nitrogen fixation pathway demonstrated a significant positive correlation (p<0.05), primarily with the relative abundances of nifD, nifK, and nifH (p<0.05) (Table S5).
DR showed significant positive correlations with the DOC/NO3—N ratio, NO3--W content, and DOC content (p<0.05) and significant negative correlations with the NO3--N, TN, and DO-W contents (p<0.05); DNRAR exhibited significant positive correlations with the DOC/NO3--N ratio and NH4+-N content (p<0.05) and a significant negative correlation with the NO3--N content (p<0.01); and NFR demonstrated significant positive correlations with the ORP-W and TOC content (p<0.05) and a significant negative correlation with the DOC content (p<0.05) (Table S5).
The RDA results (Figure 5e) indicated that the first and second principal axes accounted for 67.19% and 29.43% of the data variance, respectively, accounting for a cumulative 96.62% of data variance. The key environmental and microbial factors influencing the nitrogen transformation rate include NO3--N content, DOC/NO3--N ratio, nrfC expression, nrfA expression, DOC content, NH4+-N content, nosZ expression, norB expression, TN content, and nirK expression (Table S3).
4. Discussion
4.1. Differences in Nitrogen Transformation Rates in Rhizosphere Sediments between Artificially Cultivated Ditches and Natural Ditches
The potential DR of sediments reflects the capacity of microorganisms to remove nitrogen [70]; the DNRAR indicates the ability to convert nitrate to NH4+-N, which plays a crucial role in maintaining nitrogen cycle balance [49]; and the NFR signifies the availability of nitrogen in sediments, which is essential for sustaining plant productivity, microbial metabolism, and ecosystem health and stability [71]. Our study revealed that the DRs of A-Phr and N-Phr were significantly greater than those of A-Typ and N-Typ (Figure 3a), and the DNRAR of A-Phr was significantly greater than that of N-Phr (Figure 3b), while the NFR of A-Typ was notably greater than that of N-Typ (Figure 3c). These results are consistent with those of Cheng and White [72] and confirm our first hypothesis, which suggests that artificially planted aquatic plants promote nitrogen transformation in sediments. This can be elucidated from the following perspectives: (1) In artificial planting, to ensure high survival rates, the roots are enveloped in soil rich in nutrients at the nursery to guarantee an ample supply of organic matter and nitrogen [73]; concurrently, the vegetation coverage rate of artificial planting ditches is relatively high, reaching 57% (Table 1), which mitigates water flow velocity and its erosive impact, thereby reducing nutrient loss [74]. The TOC and NH4+-N contents of A-Phr and A-Typ were significantly greater than those of N-Phr and N-Typ (Table S2), providing sufficient substrates for microorganisms to promote nitrogen transformation. The DR and DNRAR were significantly positively correlated with the NH4+-N content, and the NFR was significantly positively correlated with the TOC content (Table S5), further confirming this observation. (2) Management measures for artificially planted ditches, such as weeding and transplanting, have been show to lead to increased sediment disturbance, promote internal nutrient release, and improve the oxygen conditions and water-holding capacity of sediments [75]. This optimization of sediment structure provided more suitable oxidation‒reduction environments for microorganisms. A-Phr and A-Typ exhibited significantly greater ORP-W values than did N-Phr and N-Typ (Table S1), with a significant positive correlation between the NFR and ORP-W (Table S5), consequently leading to an increase in the NFR. (3) Various plant species can alter the physical and chemical properties of sediments and microbial communities, thereby facilitating plant growth through nutrient provision and alleviation of environmental stress [76]. In our investigation, the combination of artificially planted P. australis, T. orientalis and Nymphaea species could form a dense root network in the sediment, increasing the diversity and quantity of functional microorganisms and promoting functional microbial activity [77]. The degradation of root exudates by microbial activity can improve the physicochemical properties of sediments, increase nutrient availability, and impact the structure and function of microbial communities [78], ultimately leading to an increase in nitrogen transformation rates [79]. However, it should be noted that the nitrogen transformation rate of artificially cultivated ditches may not always exceed that of natural ditches, as the biogeochemical process of sediments is inherently complex and dynamic. The nitrogen transformation rate is influenced by various factors, including the physical and chemical properties of sediments, vegetation status, microbial species and activities [80,81,82].
4.2. Diversity and Composition of the Bacterial Community in Rhizosphere Sediments and Their Driving Factors
The diversity and composition of the rhizosphere sediment bacterial community serve as a unique functional conduit at the interface between plants and sediment, facilitating ecological functions such as sediment material cycling and energy flow while also contributing to sediment stability [83,84]. In this investigation, it was observed that the Chao1 and ACE richness indices of the A-Phr and A-Typ bacterial communities were significantly greater than those of the N-Phr and N-Typ bacterial communities (Figure 4a, 4b), with a notably elevated relative abundance of the nitrogen-transforming bacteria Bacteroidota and Firmicutes in the A-Phr and A-Typ when contrasted with their counterparts in the natural ditches (Figure 4d). These findings are largely consistent with previous studies by Hu et al. [39], Fang et al. [38], and Xiao et al. [85], thereby providing support for our second hypothesis.
The high α diversity of the A-Phr and A-Typ bacterial communities may be attributed to the following reasons: (1) Cultivated reeds, cattails, and lotuses release oxygen through their roots, creating an oxidizing environment for microorganisms and resulting in a high ORP [86]. This environment interacts with pH and temperature [87], providing diverse aerobic or anaerobic environments and effective carbon sources for nitrifying/denitrifying bacteria [88,89]. The ORP serves as a fundamental indicator influencing the dynamics of the sediment microbial community [90], and redox processes in sediments are closely associated with the oxygen concentration and vegetation [91,92]. Sediment carbon and nitrogen increase the abundance and diversity of rhizosphere sediment bacteria [93], while nutrient input weakens the competition for resources, which is conducive to maintaining high bacterial alpha diversity [94]. In this study, the primary environmental factors influencing bacterial community richness and diversity were ORP-W and the NH4+-N and TOC contents (Table S3), which supported our hypothesis and was largely consistent with the findings of Wang et al. [95] and Song et al. [40]. (2) There are interspecific differences in nutrient uptake between emergent and floating aquatic plants, which is beneficial for nutrient cycling and water purification in eutrophic wetlands [96,97], reducing water pollutants, improving water clarity and DO concentration, and promoting bacterial growth and reproduction [98,99]. Therefore, the combination of different aquatic plants provides microenvironments with different aerobic/anaerobic and carbon-nitrogen interactions through root oxygen release, root exudates, and nutrient uptake, which are crucial for maintaining sediment microbial diversity and health [77,100]. The ORP-W and NH4+-N and TOC contents in A-Phr and A-Typ were significantly greater than those in N-Phr and N-Typ (Table S1, S2). The Chao1 and ACE richness indices exhibited a significant positive correlation with the ORP-W and NH4+-N content (Figure 5a). These findings collectively support that the Chao1 and ACE richness indices of the bacterial community in the rhizosphere sediments of A-Phr and A-Typ are markedly greater than those in N-Phr and N-Typ carbon, nitrogen and DO in the rhizosphere sediments of emergent plants are the most important factors affecting the structure of the rhizosphere microbial community [38]. When P. australis, T. orientalis and N. tetragona were artificially planted, the introduction of rhizosphere soil significantly increased the TOC and NH4+-N contents of A-Phr and A-Typ (Table S2). Moreover, human management activities have led to dynamic changes in water bodies, causing nutrient release from sediments in a suspended state and enhancing the availability of nutrients [75]. The roots of emergent and floating aquatic plants in the artificially cultivated ditch secreted carbon-rich metabolites and significantly increased the DO-W concentration in the water around the rhizosphere of A-Phr and A-Typ through root oxygen release [26,101] (Table S2). DO is the core component of rhizosphere microbial life activities. Microorganisms utilize organic metabolism to generate energy, which is then converted into carbon and nitrogen nutrients essential for organisms through respiration [102]. Therefore, artificially cultivated ditches can offer more abundant carbon and nitrogen sources for microorganisms. Under appropriate carbon-nitrogen conditions, the rhizosphere sediments of P. australis and T. orientalis promote the enrichment of nitrogen-cycle bacteria such as Proteobacteria, Bacteroidetes, Verrucomicrobiota, and Firmicutes, which are differentially regulated between microhabitats and species [103]. The Proteobacteria phylum harbors a multitude of widely distributed nitrogen-transforming bacteria in various sedimentary environments, including those found in rice fields [104], rivers [105], and lakes [106].
Anaeromyxobacter [57], Geobacter [60], Thiobacillus [61], Hydrogenophaga [62] and Sulfuritalea [66], which belong to the Proteobacteria phylum, are the predominant genera involved in denitrification or the DNRA pathway. Their relative abundances in the rhizosphere sediments of the artificially cultivated ditch exceeded those of the natural ditches (Figure 4e). Among them, Anaeromyxobacter and Geobacter exhibited significant positive correlations with the DOC/NO3--N ratio and the NH4+-N concentration and significant negative correlations with the NO3--N concentration (Figure 5c), consistent with the findings of Chen et al. [107]. NH4+-N and NO3--N serve as crucial substrates or end products in sediment denitrification and DNRA [13,17]. Human intervention in artificially cultivated ditches [75], plant root exudates, and nitrogen uptake and utilization [26,108] create more suitable environments for nitrogen transformation functional microorganisms. The Bacteroidetes phylum includes denitrifying bacteria with nirK or nirS [104], nitrogen fixation bacteria with nifH [106,109], and DNRA bacteria [110]. The Verrucomicrobiota phylum has denitrifying or DNRA functions [110,111]. The Firmicutes phylum plays an important role in sediment denitrification [112] and nitrogen fixation [106,113] processes. Therefore, the dominant bacterial phyla of A-Phr were Bacteroidota, Verrucomicrobiota, and Firmicutes, while the dominant bacterial phyla of A-Typ were Bacteroidota and Firmicutes (Figure 4d). The main influencing factors included the DOC, TN, TOC, Sand, and DO-W contents (Table S3). These results confirmed a significantly greater abundance of the nitrogen cycle bacterial phyla Bacteroidota, Verrucomicrobiota, and Firmicutes in both A-Phr and A-Typ than in N-Phr and N-Typ.
4.3. Relative Abundance of Nitrogen Transformation Functional Genes in Rhizosphere Sediments and Their Driving Factors
Aquatic plants improve the rhizosphere environment, promote the growth of nitrogen transformation-related microorganisms, accelerate the denitrification of eutrophic wetlands, and optimize the effective utilization of biological nitrogen [114,115]. In this study, the primary pathways for nitrogen transformation in rhizosphere sediments included denitrification, assimilative nitrate reduction, dissimilatory nitrate reduction, and nitrogen fixation. The relative abundances of nitrification and anammox were less than 1% (Figure 6a). Due to the low levels of DO ranging from 2.58 mg/L to 7.50 mg/L in the water surrounding the rhizosphere, the presence of facultative anaerobic conditions in the sediment environment, relatively high ECs ranging from 270 μs/cm to 518 μs/cm, and relatively low concentrations of NH4+-N ranging from 1.35 mg/kg to 2.90 mg/kg (Table S1, S2), nitrification and anammox were inhibited [116,117]. The abundances of denitrification genes (napA and norB), dissimilatory nitrate reduction genes (nrfC and nrfA), nitrification genes (hao) and anammox genes (hzsA and hzsC) in A-Phr were significantly greater than those in N-Phr, and the abundance of nitrogen fixation genes (nifD, nifK, and nifH) in A-Typ was also significantly greater than that in N-Typ (Figure 6b, 6c), which is consistent with the findings of previous studies by Song et al. [40] and Fang et al. [38], confirming part of our second hypothesis.
Under facultative anaerobic conditions, denitrifying microorganisms can reduce NO3--N to N2O or N2 [118]. The periplasmic nitrate reductase napA, which converts NO3- to NO2-; the nitrite reductases nirS and nirK, which convert NO2- to NO; the nitric oxide reductase subunit B norB; and the nitrous oxide reductase nosZ, which converts NO to N2 [119,120], are widely used as marker genes for analyzing denitrifying microbial communities [14,121]. In this study, the relative abundances of the nirS, nirK, and nosZ genes of N-Phr involved in the denitrification pathway were greater than those of A-Phr, A-Typ, and N-Typ, while the relative abundances of the napA and norB genes of A-Phr were greater than those of A-Typ, N-Phr, and N-Typ (Figure 6b).
This can be attributed to the following factors: (1) In the natural ditch with P. australis, there was minimal human interference, and the sediments remained in a prolonged anaerobic state. The nirS gene plays a crucial role in the anaerobic denitrification pathway [122]. Our study revealed a significant negative correlation between the denitrification pathway, nirS gene, and DO-W content (Figure 5d, Table S4), confirming the high abundance of the denitrification functional gene nirS under anaerobic conditions. (2) Human management has led to changes in the DO, carbon, and nitrogen contents in the sediments of artificially cultivated ditches. A-Phr exhibited a high abundance of the aerobic denitrification gene napA [123], which is primarily supported by DOC as an electron donor and energy source [124]. The denitrification pathway showed a significant positive correlation with the DOC content (Figure 5d), and the napA gene also exhibited a significant positive correlation with the DOC/NO3--N ratio (Table S4), providing further confirmation. (3) Artificial cultivation has led to an increase in the diversity of ecological niches in sediments, resulting in nearly equal distributions of nirK/nirS in the A-Phr and A-Typ sediments, which is consistent with the findings of Saarenheimo et al. [125]. (4) Denitrifying bacteria, as heterotrophic microorganisms, tend to accumulate in facultative anaerobic and anaerobic zones due to their distinct oxygen and nutrient requirements [126]. These bacteria can utilize nitrogen oxides as terminal electron acceptors in cellular bioenergy processes [127] while using carbon sources as electron donors and energy sources [123]. Anaerobic environments with high carbon sediment are considered hotspots for denitrification [128]. The DO-W contents of A-Phr and N-Phr were relatively low, whereas the DOC content was relatively high (Table S1, S2). Compared to the root system of T. orientalis, the root system of P. australis secretes more oxygen and sugary exudates, creating diverse anaerobic/aerobic environments and providing effective carbon sources for denitrifying bacteria [26,89]. Consequently, the denitrification functional genes of A-Phr and N-Phr exhibited higher expression levels than did those of T. orientalis (Figure 6a).
Under anaerobic conditions, DNRA initially reduces NO3− to NO2− and subsequently directly converts NO2− to biologically more available NH4+, thereby enhancing nitrogen utilization [129,130]. The expression of the nitrite reductase (cytochrome c-552) nrfA is positively associated with DNRA activity [131], and nrfA serves as a marker gene [18]. In this study, A-Phr exhibited a greater relative abundance of the nrfA gene, which is involved in the DNRA pathway (Figure 6b). There are several potential explanations for this observation: (1) As previously mentioned, A-Phr exhibited a higher DOC/NO3--N ratio than N-Phr, which was advantageous for the distribution of nitrate in the DNRA process [132], leading to the dominance of the DNRA process. Additionally, the elevated DOC/NO3--N ratio had a positive impact on the interactions among rare microorganisms involved in the DNRA process [133,134]. In this study, we observed a significant positive correlation between the DNRA pathway and the DOC/NO3—N ratio, as well as a negative correlation with the NO3--N content (Figure 5d), further confirming this finding. (2) In this study, we observed a significant positive correlation between the DNRA pathway, the nrfA gene, and DOC content (Figure 5d, Table S4), suggesting that the higher DOC content of A-Phr serves as an important carbon source for DNRA. This increase in DOC content has been found to enhance the presence of the nrfA gene and promote DNRA occurrence [135,136]. (3) The exudates in the rhizosphere of P. australis provide ample carbon sources for the DNRA process [137], leading to increased complexity and stability in microbial interactions, thereby enhancing the DNRA process in rhizosphere sediments [138].
Nitrogen-fixing microorganisms convert N2 into NH4+ to provide plants with nutrients for absorption and utilization [139] and represent the primary source of biologically available nitrogen. The nifH gene serves as a crucial indicator for identifying nitrogen-fixing biological species and analyzing their nitrogen-fixing activity [140]. In this study, A-Typ exhibited a relatively high abundance of genes involved in the nitrogen fixation pathway, including nifD, nifK, and nifH (Figure 6b). The potential factors may include the following: (1) The positive correlation between the nitrogen-fixing gene and ORP-W (Figure 5d, Table S4) may be attributed to the exudation of oxygen from roots, resulting in an increase in the ORP of A-Typ, thereby facilitating the acceleration of nitrogen fixation [26]. (2) The positive correlation between nitrogen-fixing genes and DO-W (Figure 5d, Table S4) suggests a potentially abundant population of aerobic nitrogen-fixing bacteria in rhizosphere sediments [141,142], indicating the enhancement of an aerobic nitrogen-fixing microbial community by DO [143]. (3) The genes associated with nitrogen fixation showed a significant negative correlation with DOC (Figure 5d, Table S4). This suggests that DOC, acting as a substrate, may enhance the proliferation of denitrifying bacteria and DNRA bacteria [124,136] while inhibiting nitrogen-fixing bacteria.
4.4. Influence of Environmental and Microbial Factors on the Nitrogen Conversion Rate of Sediment
Previous research has predominantly focused on the influence of environmental factors such as carbon and nitrogen on nitrogen transformation rates, highlighting the significance of abiotic factors [144,145]. Additionally, investigations have demonstrated that microorganisms play a central role in driving nitrogen transformation, suggesting that both biotic and abiotic factors regulate nitrogen transformation rates [146,147]. This study, which monitors both environmental and microbial factors in real time, can more effectively reveal the regulatory mechanisms of sediment nitrogen transformation [80].
The nitrogen removal capacity of agricultural ditch systems in the Yellow River irrigation area is primarily evaluated through the measurement of the sediment denitrification rate [148]. The denitrification rate is influenced mainly by environmental factors such as the NO3—N, DOC/NO3—N, TN, NO3--W, and DOC contents, while the key microbial factor is the norB gene (Table S5). (1) The microenvironment of the rhizosphere varies among different plant species, leading to differences in the potential denitrification rates and abundance of denitrification functional genes [13]. The cultivation of plants in drainage ditches has been shown to increase the DR [149]. Due to the superior competitive advantage of a single plant community in terms of nitrogen uptake, it diminishes the available nitrate for denitrification. As a result, a multiplant community may not necessarily lead to an increase in the DR [150]. Therefore, the difference in denitrification rates between A-Phr and N-Phr was not statistically significant (Figure 3a). However, the root system of P. australis can secrete more oxygen and sugar exudates than that of T. orientalis [26,89], resulting in higher denitrification rates and abundances of denitrification functional genes in A-Phr and N-Phr than in T. orientalis (Figure 3a, Figure 6a). (2) DOC is the most important carbon source for denitrifying microorganisms [152]. The Weihe River in Xi’an, which is heavily influenced by agriculture, shows a significant positive correlation between the sediment denitrification rate and DOC content, as well as a significant positive correlation with the abundance of the nirS and nroB functional genes for denitrification [86]. Organic carbon provides electrons and energy for nitrogen cycle microorganisms [153,154]. Due to agricultural fertilization, the NO3--N load in rivers has increased [155], resulting in alterations in sediment nitrogen cycling and the transformation of wetlands from active nitrogen “sources” to “sinks”. The elevated NO3--N concentration in the overlying water and reduced TOC in the sediments led to a decrease in the nitrogen fixation rate and an increase in the denitrification rate [156]. (3) Agricultural rivers have a relatively high denitrification rate due to their high nitrogen load [157]. The potential denitrification rate of sediments in the Huaihe River is positively correlated with the abundance of the nirS, nirK, and nosZ genes, while the NH4+-N and TN contents contribute more to the variation in the denitrification rate [147]. However, the rate of denitrification may be affected by the suppression of microorganisms, and the DR in river sediments influenced by agriculture is related to changes in microbial community composition and the abundance of the nosZ gene, which is mainly inhibited by high NO3--N concentrations [158]. Therefore, variations in denitrification rates are driven by both denitrifying microorganisms and sediment physical and chemical properties, potentially due to different responses of denitrifying bacteria to environmental variables (NO3--N, TN, DOC) [159]. Furthermore, since the samples in this study were collected at only one time point, they only reflect the denitrification activity at a specific period, which has certain limitations. Further research is needed to study the temporal sequence changes in the structure of the denitrifying bacterial community and the DR.
The DNRAR is primarily influenced by environmental factors such as the NO3--N content, the DOC/NO3—N ratio and the NH4+-N content, as well as microbial factors such as nrfA and nrfC genes (Table S5). A-Phr contains a high concentration of the electron donor DOC and a low concentration of the electron acceptor NO3--N (Table S1, S2), creating a more favorable environment for DNRA [160,161]. Moreover, the DOC/NO3--N ratio, organic carbon, and nitrate are important factors affecting the bacterial community and functional genes of DNRA [81,162]. DNRA bacteria have a high affinity for NO3--N and are adapted to environments with high carbon and limited NO3--N [163]. Therefore, environmental and microbial factors jointly promote the DNRAR; for example, the DNRAR of sediments in the riparian zone is positively correlated with DOC/NO3--N and significantly positively correlated with the abundance of the functional gene nrfA [86].
The primary environmental factors influencing the rate of nitrogen fixation included the ORP-W, TOC content, and DOC content, while the key microbial factors were the nifD, nifK, and nifH genes (Table S5). Because A-Typ has a high ORP-W and TOC content (Table S1, S2), it provides oxidation‒reduction conditions for nitrogen-fixing microorganisms and supplements energy for heterotrophic organisms [164,165]. Therefore, the nitrogen fixation rate of plant roots is related to the ORP, TOC content, and abundance of nitrogen-fixing microorganisms [26,166]. The findings of Spinette et al. [167] suggest that the enrichment of organic matter in estuarine sediments promotes biological nitrogen fixation. Additionally, Luo et al. [168] demonstrated a positive correlation between the rate of sediment nitrogen fixation and the relative abundance of the nitrogen-fixing genes nifD, nifK, and nifH.
In this study, the rate of nitrogen transformation was found to be significantly positively correlated with the abundance of functional genes (Table S5), which is consistent with findings in the Yangtze River estuary [169], the Weihe River bank [86], and high-altitude rivers [170], suggesting that the abundance of functional genes is a good predictor of the nitrogen transformation rate [171]. This finding also indicates that the nitrogen transformation rate in sediments is jointly regulated by different functional microorganisms and environmental factors and that different functional microorganisms are driven by environmental factors [146]. Therefore, the key nitrogen transformation rate of artificially cultivated ditches in the Yellow River irrigation area is related to microorganisms, vegetation conditions and sediment physical and chemical properties. The main environmental and microbial factors affecting the nitrogen transformation rate were the NO3--N content, DOC/NO3--N ratio, nrfC expression, nrfA expression, DOC content, NH4+-N content, nosZ expression, norB expression, TN content, and nirK expression (Table S3).
The reasons for this may be as follows: (1) In an artificially cultivated ditch, P. australis and T. orientalis transfer oxygen from the air through their stems and roots, which are rich in aerated tissues, to water-saturated sediments, controlling the DO concentration in the rhizosphere and ultimately creating habitats for microorganisms that degrade water pollutants together with sediments [31]. N. tetragona possesses well-developed rhizomes, which also offer an expanded surface area for the attachment of microorganisms. The planting of different eco-types of plants in artificially cultivated ditches maximizes the utilization of various environmental factors in the vertical space, particularly the electron donor DOC and electron acceptor NO3--N necessary for denitrification and DNRA [82,172]. This creates a more suitable habitat for the functional DNRA genes (nrfC, nrfA) and denitrification genes (nosZ, norB, nirK). (2) A-Phr had a greater DNRAR and DR than N-Phr, and the functional genes involved in denitrification (napA, norB) and dissimilatory nitrate reduction (nrfC, nrfA) had an advantage (Figure 6b). In addition, the expression of functional genes involved in nitrification (hao) and anammox (hzsA, hzsC) also increased (Figure 6c), mainly due to the increase in the NO3--N content, DOC/NO3—N ratio and NH4+-N content in the rhizosphere (Table S4), which can gradually progress through a more complete nitrogen cycle (Figure 7). Through artificial vegetation restoration, the carbon and nitrogen effectiveness of riverbeds has significantly improved, and the denitrification of sediment has increased [75]. Denitrification in vegetated ditches can reduce the nitrogen load by 28-56% [173,174]. Moreover, DNRA can convert unstable nitrates into ammonium, which plants can absorb, thereby reducing nitrogen losses in agricultural ecosystems [175] by competing with denitrification. When the sediment carbon-nitrogen ratio is low, denitrification processes have a competitive advantage; when the ratio is high, DNRA predominates. Therefore, a higher abundance of DOC and a more limited NO3--N content favor the growth of DNRA bacteria [86,129]. Other studies have shown that oxygen is an important factor affecting the competition between denitrification and DNRA [176]. While both denitrification and DNRA prefer anoxic environments, DNRA bacteria are more likely to survive in deposit environments with relatively high oxygen levels [177]. The nitrification and anammox pathways, which are important links in the nitrogen cycle, are closely coupled with other processes and play a crucial role in maintaining the sediment nitrogen cycle and ecosystem health [148,178]. (3) A-Typ had a greater nitrogen fixation rate, and the expression of nifD, nifK, and nifH functional genes involved in the nitrogen fixation pathway increased (Figure 6b). This observation was mainly related to the ORP-W, DOC content, and DO-W content (Table S4), which provide more available nitrogen for biological metabolism. Nitrogen fixation and denitrification are two important processes for maintaining the balance of nitrogen in sediments. To some extent, nitrogen fixation compensates for the nitrogen loss caused by denitrification [179]. Plant roots can release oxygen and unstable carbon into sediments, stimulating denitrification and enhancing nitrogen removal through the biological conversion of nitrate to N2 [180]. Furthermore, aquatic vegetation has the capacity to mitigate soil erosion, stabilize sediments, and facilitate the sequestration of carbon and nitrogen in sedimentary environments [181]. The combination of emerged and floating aquatic plants in artificially cultivated ditches creates a unique environment for rhizosphere microorganisms, which is conducive to nitrogen fixation and denitrification processes, thus significantly improving water quality [179,182]. Therefore, the establishment of drainage ditches featuring aquatic plant-dominated habitats holds significant importance for mitigating regional water eutrophication.
5. Conclusions
The predominant pathways for nitrogen transformation in the sediments of drainage ditches within the Yellow River irrigation area included denitrification, DNRA, and nitrogen fixation. A-Phr exhibited a high DR and DNRAR, while A-Typ demonstrated a high rate of nitrogen fixation. The richness indices of the A-Phr and A-Typ bacterial communities, as well as the relative abundances of bacterial phyla (Bacteroidota, Verrucomicrobiota, Firmicutes) and bacterial genera (Anaeromyxobacter, Geobacter, Thiobacillus, Hydrogenophaga, Sulfuritalea) involved in nitrogen cycling, were all significantly greater than those of the natural ditches. The functional genes nrfC, nrfA, nosZ, norB, and nirK, as well as the environmental factors NO3--N content, DOC/NO3--N ratio, DOC content, NH4+-N content, and TN content, interact with each other to collectively influence the rates of nitrogen transformation in rhizosphere sediments.
The artificial cultivation of aquatic plants primarily improves the sediment microbial environment through human management, vegetation coverage, and root oxygen exhalation and secretion, leading to an increase in the abundance of functional genes related to denitrification (napA, norB), dissimilatory nitrate reduction (nrfC, nrfA), and nitrogen fixation (nifD, nifK, nifH), thereby facilitating nitrogen transformation and enhancing the capacity to remove nitrogen loads from water bodies while also stimulating biological metabolism and the effective cycling of nitrogen. Our research findings demonstrate the disparities in nitrogen transformation rates between artificially cultivated and naturally occurring aquatic plants in drainage ditches within the Yellow River irrigation area, as well as the influential effects of microbial and environmental factors. These findings provide a scientific foundation for utilizing aquatic plants in the restoration of agricultural runoff. In future research, greater emphasis should be placed on monitoring the dynamic changes in nitrogen transformation processes and functional microbial communities, as well as further exploration of the microbial mechanisms involved in the nitrogen cycle within artificially cultivated ditch water and sediments.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org. Detailed calculation formulas for the potential DRs, DNRARs and NFRs. Table S1: Physical and chemical properties of water around roots. Table S2: Physical and chemical properties of all sediment samples (n = 3). Table S3: RDA single environmental factor difference significance (envfit test). Table S4: Pearson’s correlation coefficients between the relative abundances of major functional genes involved in nitrogen cycling pathways and environmental factors. Table S5: Pearson’s correlation coefficient between critical nitrogen conversion rates, environmental factors and the relative abundance of genes involved in nitrogen cycling pathways. Figure S1: Number of high-quality sequences (Effective-CCS (a)), the number of features (OUT Num (b)) and the number of reads corresponding to the feature (Seq num (c)) obtained through 16S rRNA sequencing.
Author Contributions
Conceptualization, Y.H.; Data curation, Y.H.; Formal analysis, Y.H.; Investigation, Y.H. and Z.H; Methodology, Y.H.; Project administration, R.L.; Supervision, X.F.; Writing – original draft, Y.H.; Writing – review & editing, R.L., W.X., P.L. and X.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ningxia Key R&D Program (grant no. 2022BEG02007), the Ningxia Agricultural High-quality Development and Ecological Protection Technology Innovation Demonstration Project (grant no. NGSB-2021-11), and Ningxia Youth Talent Project (grant no. 2018).
Data Availability Statement
The data are not publicly available due to privacy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, T.; Zhang, Q.; Wang, G.; Singh, V.P.; Zhao, J.; Sun, S.; Wang, D.; Liu, T.; Duan, L. Ecological Degradation in the Inner Mongolia Reach of the Yellow River Basin, China: Spatiotemporal Patterns and Driving Factors. Ecological Indicators 2023, 154, 110498. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Qi, J.; Wang, Q. Evaluation of the Stability and Suitable Scale of an Oasis Irrigation District in Northwest China. Water 2020, 12, 2837. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y. Associations among ecosystem services from local perspectives. Science of The Total Environment 2019, 690, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lei, Q.; Liang, X.; Lindsey, S.; Luo, J.; Pei, W.; Du, X.; Wu, S.; An, M.; Qiu, W.; et al. Optimization of the N Footprint Model and Analysis of Nitrogen Pollution in Irrigation Areas: A Case Study of Ningxia Hui Autonomous Region, China. Journal of Environmental Management 2023, 340, 118002. [Google Scholar] [CrossRef]
- Tian, L. Analysis and Investigation of the Third Drainage Ditch Pollution Situation in Ningxia Based on Multivariate Statistics. E3S Web Conf. 2020, 145, 02082. [Google Scholar] [CrossRef]
- Pei, W.; Yan, T.; Lei, Q.; Zhang, T.; Fan, B.; Du, X.; Luo, J.; Lindsey, S.; Liu, H. Spatio-Temporal Variation of Net Anthropogenic Nitrogen Inputs (NANI) from 1991 to 2019 and Its Impacts Analysis from Parameters in Northwest China. Journal of Environmental Management 2022, 321, 115996. [Google Scholar] [CrossRef] [PubMed]
- Carstensen, M.V.; Hashemi, F.; Hoffmann, C.C.; Zak, D.; Audet, J.; Kronvang, B. Efficiency of Mitigation Measures Targeting Nutrient Losses from Agricultural Drainage Systems: A Review. Ambio 2020, 49, 1820–1837. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Zhang, J.; Liu, L.; Li, Y.; Li, X. A Source-Sink Landscape Approach to Mitigation of Agricultural Non-Point Source Pollution: Validation and Application. Environmental Pollution 2022, 314, 120287. [Google Scholar] [CrossRef]
- Li, S.; Wu, M.; Jia, Z.; Luo, W.; Fei, L.; Li, J. Influence of Different Controlled Drainage Strategies on the Water and Salt Environment of Ditch Wetland: A Model-Based Study. Soil and Tillage Research 2021, 208, 104894. [Google Scholar] [CrossRef]
- Clifford, C.C.; Heffernan, J.B. B. North Carolina Coastal Plain Ditch Types Support Distinct Hydrophytic Communities. Wetlands 2023, 43, 1–27. [Google Scholar] [CrossRef]
- Hou, L.; Gao, D.; Zheng, Y.; Li, X.; Yin, G.; Dong, H.; Liang, X.; Liu, M. Nitrogen Cycling in Aquatic Environments of China: Progress and Future Challenges. Progress in Physical Geography: Earth and Environment 2022, 46, 846–868. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The Microbial Nitrogen-Cycling Network. Nat Rev Microbiol 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Li, Y.; Yang, W.; Huang, J.; Hou, K.; Zhang, L.; Song, H.; Yang, L.; Tian, C.; Rong, X.; et al. A Comparison of the Mechanisms and Performances of Acorus Calamus, Pontederia Cordata and Alisma Plantagoaquatica in Removing Nitrogen from Farmland Wastewater. Bioresource Technology 2021, 332, 125105. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, W.; Zhao, S.; Wang, X.; Hefting, M.M.; Schwark, L.; Zhu, G. Anammox and Denitrification Separately Dominate Microbial N-Loss in Water Saturated and Unsaturated Soils Horizons of Riparian Zones. Water Research 2019, 162, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Sanford, R.A.; Chee-Sanford, J.; Ooi, S.K.; Löffler, F.E.; Konstantinidis, K.T.; Yang, W.H. Beyond Denitrification: The Role of Microbial Diversity in Controlling Nitrous Oxide Reduction and Soil Nitrous Oxide Emissions. Global Change Biology 2021, 27, 2669–2683. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.-W.; Chen, D.; He, J.-Z. Microbial Regulation of Terrestrial Nitrous Oxide Formation: Understanding the Biological Pathways for Prediction of Emission Rates. FEMS Microbiology Reviews 2015, 39, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Castro-Barros, C.M.; Jia, M.; Van Loosdrecht, M.C.M.; Volcke, E.I.P.; Winkler, M.K.H. Evaluating the Potential for Dissimilatory Nitrate Reduction by Anammox Bacteria for Municipal Wastewater Treatment. Bioresource Technology 2017, 233, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J.; Sanford, R.A.; Connor, L.; Yang, W.H.; Chee-Sanford, J. Optimization of PCR Primers to Detect Phylogenetically Diverse NrfA Genes Associated with Nitrite Ammonification. Journal of Microbiological Methods 2019, 160, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Hester, E.R.; Harpenslager, S.F.; Lüke, C.; Lücker, S.; Welte, C.U. Linking Nitrogen Load to the Structure and Function of Wetland Soil and Rhizosphere Microbial Communities. 2018, 3, e00214–17. [Google Scholar] [CrossRef]
- Uesaka, K.; Banba, M.; Chiba, S.; Fujita, Y. Restoration of the Functional Nif Gene Cluster by Complex Recombination Events during Heterocyst Development in the Nitrogen-Fixing Cyanobacterium Calothrix Sp. NIES-4101. Plant And Cell Physiology 2024, 65, 1050–1064. [Google Scholar] [CrossRef]
- He, R.; Zeng, J.; Zhao, D.; Wang, S.; Wu, Q.L. Decreased Spatial Variation and Deterministic Processes of Bacterial Community Assembly in the Rhizosphere of Phragmites Australis across the Middle–Lower Yangtze Plain. Molecular Ecology 2022, 31, 1180–1195. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Jia, M.; Xun, M.; Wang, X.; Chen, K.; Yang, H. Nitrogen Transformation and Microbial Community Structure Varied in Apple Rhizosphere and Rhizoplane Soils under Biochar Amendment. J Soils Sediments 2021, 21, 853–868. [Google Scholar] [CrossRef]
- Sun, L.; Tsujii, Y.; Xu, T.; Han, M.; Li, R.; Han, Y.; Gan, D.; Zhu, B. Species of Fast Bulk-soil Nutrient Cycling Have Lower Rhizosphere Effects: A Nutrient Spectrum of Rhizosphere Effects. Ecology 2023, 104, e3981. [Google Scholar] [CrossRef]
- Supreeth, M. Enhanced remediation of pollutants by microorganisms-plant combination. International Journal of Environmental Science and Technology 2021, (5), 1–12. [Google Scholar] [CrossRef]
- Cong, S.; Yuelu, J. Physiological and Ecological Responses and Changes of Phaeodactylum Tricornutum under Long-Term Stress of Naphthalene. IOP Conf. Ser.: Earth Environ. Sci. 2019, 310, 052021. [Google Scholar] [CrossRef]
- Fang, J.; Zhao, R.; Cao, Q.; Quan, Q.; Sun, R.; Liu, J. Effects of Emergent Aquatic Plants on Nitrogen Transformation Processes and Related Microorganisms in a Constructed Wetland in Northern China. Plant Soil 2019, 443, 473–492. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, M.; Zhang, Q.; Bao, Y.; Yang, N.; Huo, Y.; He, P. Epiphytic Bacterial Community Composition on the Surface of the Submerged Macrophyte Myriophyllum Spicatum in a Low-Salinity Sea Area of Hangzhou Bay. Oceanological and Hydrobiological Studies 2019, 48, 43–55. [Google Scholar] [CrossRef]
- Yang, J.; Pei, H.; Lü, J.; Liu, Q.; Nan, F.; Liu, X.; Xie, S.; Feng, J. Effects of Phytoplankton Community and Interaction between Environmental Variables on Nitrogen Uptake and Transformations in an Urban River. J. Ocean. Limnol. 2022, 40, 1012–1026. [Google Scholar] [CrossRef]
- Wei, W.; Tong, J.; Hu, B.X. Study on Ecological Dynamic Model for Phytoremediation of Farmland Drainage Water. Journal of Hydrology 2019, 578, 124026. [Google Scholar] [CrossRef]
- Bai, S.; Chen, J.; Guo, M.; Ren, N.; Zhao, X. Vertical-Scale Spatial Influence of Radial Oxygen Loss on Rhizosphere Microbial Community in Constructed Wetland. Environment International 2023, 171, 107690. [Google Scholar] [CrossRef]
- Duan, Z.; Xie, S.; Chen, Z.; Wang, J.; Li, Z. Effects of Aquatic Plants in Constructed Wetlands to Removal of Water Pollutants. In Proceedings of the World Environmental and Water Resources Congress 2018; American Society of Civil Engineers: Minneapolis, Minnesota,, 31 May 2018; pp. 239–245. [Google Scholar] [CrossRef]
- Riva, V.; Mapelli, F.; Syranidou, E.; Crotti, E.; Choukrallah, R.; Kalogerakis, N.; Borin, S. Root Bacteria Recruited by Phragmites Australis in Constructed Wetlands Have the Potential to Enhance Azo-Dye Phytodepuration. Microorganisms 2019, 7, 384. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Mao, W.; Pang, L.; Li, R.; Li, S. Influence of Phragmites Communis and Zizania Aquatica on Rhizosphere Soil Enzyme Activity and Bacterial Community Structure in a Surface Flow Constructed Wetland Treating Secondary Domestic Effluent in China. Environ Sci Pollut Res 2020, 27, 26141–26152. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Zhang, X.; Cai, M.; Zhou, L.; Chen, G.; Zou, G. Roles of Vegetation in Nutrient Removal and Structuring Microbial Communities in Different Types of Agricultural Drainage Ditches for Treating Farmland Runoff. Ecological Engineering 2020, 155, 105941. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, F.; Xiao, R.; Li, Y.; He, Y.; Wu, J. Effects of Vegetation on Ammonium Removal and Nitrous Oxide Emissions from Pilot-Scale Drainage Ditches. Aquatic Botany 2016, 130, 37–44. [Google Scholar] [CrossRef]
- Veraart, A.J.; Dimitrov, M.R.; Schrier-Uijl, A.P.; Smidt, H.; De Klein, J.J.M. Abundance, Activity and Community Structure of Denitrifiers in Drainage Ditches in Relation to Sediment Characteristics, Vegetation and Land-Use. Ecosystems 2017, 20, 928–943. [Google Scholar] [CrossRef]
- Meuleman, A.F.M.; Beekman, J.Ph.; Verhoeven, J.T.A. Nutrient Retention and Nutrient-Use Efficiency in Phragmites Australis Stands after Wasterwater Application. Wetlands 2002, 22, 712–721. [Google Scholar] [CrossRef]
- Fang, J.; Yang, R.; Cao, Q.; Dong, J.; Li, C.; Quan, Q.; Huang, M.; Liu, J. Differences of the Microbial Community Structures and Predicted Metabolic Potentials in the Lake, River, and Wetland Sediments in Dongping Lake Basin. Environ Sci Pollut Res 2020, 27, 19661–19677. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; He, R.; Zeng, J.; Zhao, D.; Wang, S.; He, F.; Yu, Z.; Wu, Q.L. Lower Compositional Variation and Higher Network Complexity of Rhizosphere Bacterial Community in Constructed Wetland Compared to Natural Wetland. Microb Ecol 2023, 85, 965–979. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wang, Q.; Zhu, Y.; Christakos, G.; Wu, J. Changes to the Structure and Function of Microbial Communities in Spartina Alterniflora and Kandelia Obovata Sediments as a Factor of Stand Age. Applied Soil Ecology 2022, 177, 104544. [Google Scholar] [CrossRef]
- Ding, J.; Jia, Y.; Zhao, C.; Bo, W.; Xu, X.; Lv, R.; Zhou, G.; Kong, Q.; Du, Y.; Xu, F.; et al. Microbial Abundance and Community i46n Constructed Wetlands Planted with Phragmites Australis and Typha Orientalis in Winter. International Journal of Phytoremediation 2021, 23, 1476–1485. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, F.; Li, X.; Li, C.; Zhao, Y.; Gao, Y.; Liu, J. Effects of Plants and Soil Microorganisms on Organic Carbon and the Relationship between Carbon and Nitrogen in Constructed Wetlands. Environ Sci Pollut Res 2023, 30, 62249–62261. [Google Scholar] [CrossRef] [PubMed]
- Walton, C.R.; Zak, D.; Audet, J.; Petersen, R.J.; Lange, J.; Oehmke, C.; Wichtmann, W.; Kreyling, J.; Grygoruk, M.; Jabłońska, E.; et al. Wetland Buffer Zones for Nitrogen and Phosphorus Retention: Impacts of Soil Type, Hydrology and Vegetation. Science of The Total Environment 2020, 727, 138709. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.P. Denitrification in Sediment Determined from Nitrogen Isotope Pairing. FEMS Microbiology Letters 1992, 86, 357–362. [Google Scholar] [CrossRef]
- Hou, L.; Liu, M.; Carini, S.A.; Gardner, W.S. Transformation and Fate of Nitrate near the Sediment–Water Interface of Copano Bay. Continental Shelf Research 2012, 35, 86–94. [Google Scholar] [CrossRef]
- Montoya, J.P.; Voss, M.; Kahler, P.; Capone, D.G. A Simple, High-Precision, High-Sensitivity Tracer Assay for N(Inf2) Fixation. Appl Environ Microbiol 1996, 62, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Rysgaard, S.; Glud, R.N.; Risgaard-Petersen, N.; Dalsgaard, T. Denitrification and Anammox Activity in Arctic Marine Sediments. Limnology & Oceanography 2004, 49, 1493–1502. [Google Scholar] [CrossRef]
- Yang, Z.; Lu, L.; Cheng, Z.; Xian, J.; Yang, Y.; Liu, L.; Xu, X. Dissimilatory Nitrate Reduction in Urban Lake Ecosystems: A Comparison Study between Closed and Open Lakes in Chengdu, China. Water Research 2022, 214, 118218. [Google Scholar] [CrossRef]
- Deng, F.; Hou, L.; Liu, M.; Zheng, Y.; Yin, G.; Li, X.; Lin, X.; Chen, F.; Gao, J.; Jiang, X. Dissimilatory Nitrate Reduction Processes and Associated Contribution to Nitrogen Removal in Sediments of the Yangtze Estuary. JGR Biogeosciences 2015, 120, 1521–1531. [Google Scholar] [CrossRef]
- Hou, L.; Wang, R.; Yin, G.; Liu, M.; Zheng, Y. Nitrogen Fixation in the Intertidal Sediments of the Yangtze Estuary: Occurrence and Environmental Implications. JGR Biogeosciences 2018, 123, 936–944. [Google Scholar] [CrossRef]
- Li, D.; Gao, G.; Lü, Y.; Fu, B. Multi-Scale Variability of Soil Carbon and Nitrogen in the Middle Reaches of the Heihe River Basin, Northwestern China. CATENA 2016, 137, 328–339. [Google Scholar] [CrossRef]
- Guo, L.; Wang, X.; Lin, Y.; Yang, X.; Ni, K.; Yang, F. Microorganisms That Are Critical for the Fermentation Quality of Paper Mulberry Silage. Food and Energy Security 2021, 10, e304. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat Biotechnol 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Research 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. The KEGG Resource for Deciphering the Genome. Nucleic Acids Research 2004, 32, 277D–280. [Google Scholar] [CrossRef]
- Tu, Q.; Lin, L.; Cheng, L.; Deng, Y.; He, Z. NCycDB: A Curated Integrative Database for Fast and Accurate Metagenomic Profiling of Nitrogen Cycling Genes. Bioinformatics 2019, 35, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- You, A.; Hua, L.; Hu, J.; Tian, J.; Ding, T.; Cheng, N.; Hu, L. Patters of Reactive Nitrogen Removal at the Waters in the Semi-Constructed Wetland. Journal of Environmental Management 2023, 344, 118733. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhang, W.; Gao, J.; Hu, X.; Zhang, C.; He, Q.; Yang, F.; Wang, H.; Wang, X.; Zhan, X. A pilot-scale study on the treatment of landfill leachate by a composite biological system under low dissolved oxygen conditions: Performance and microbial community. Bioresource Technology 2020, 296, 122344. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Poursat, B.A.J.; De Ridder, D.; Smidt, H.; Van Der Wal, A.; Sutton, N.B. Unravelling the Contribution of Nitrifying and Methanotrophic Bacteria to Micropollutant Co-Metabolism in Rapid Sand Filters. Journal of Hazardous Materials 2022, 424, 127760. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bu, C.; Yang, H.; Qiao, Z.; Ding, S.; Ni, S.-Q. Survey of Dissimilatory Nitrate Reduction to Ammonium Microbial Community at National Wetland of Shanghai, China. Chemosphere 2020, 250, 126195. [Google Scholar] [CrossRef]
- Chen, C.; Yang, X.; Luo, H.; Zeng, D.; Sima, M.; Huang, S. Linking Microbial Community and Biological Functions to Redox Potential during Black-Odor River Sediment Remediation. Environ Sci Pollut Res 2020, 27, 40392–40404. [Google Scholar] [CrossRef]
- Deng, M.; Li, L.; Dai, Z.; Senbati, Y.; Song, K.; He, X. Aerobic Denitrification Affects Gaseous Nitrogen Loss in Biofloc-Based Recirculating Aquaculture System. Aquaculture 2020, 529, 735686. [Google Scholar] [CrossRef]
- Xu, T.; Tao, Y.; Song, L.; Wang, H.; Ren, B. A Unique Microbiome in a Highly Polluted and Alkalic Lake in a Seasonally Frozen Area. Environmental Research 2022, 204, 112056. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, L.; Yang, L.; Liu, C.; Li, J.; Li, N. Combined Impact of Organic Matter, Phosphorus, Nitrate, and Ammonia Nitrogen on the Process of Blackwater. Environ Sci Pollut Res 2021, 28, 32831–32843. [Google Scholar] [CrossRef]
- Sarkodie, E.K.; Jiang, L.; Li, K.; Guo, Z.; Yang, J.; Shi, J.; Peng, Y.; Wu, X.; Huang, S.; Deng, Y.; et al. The Influence of Cysteine in Transformation of Cd Fractionation and Microbial Community Structure and Functional Profile in Contaminated Paddy Soil. Science of The Total Environment 2024, 906, 167535. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Wang, Q.; She, Z.; Gao, M.; Zhao, Y.; Guo, L.; Jin, C. Nitrogen Removal Pathway and Dynamics of Microbial Community with the Increase of Salinity in Simultaneous Nitrification and Denitrification Process. Science of The Total Environment 2019, 697, 134047. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.R.L.; De Alcantara Neto, F.; Dutra, A.F.; Mendes, L.W.; Antunes, J.E.L.; Melo, V.M.M.; Oliveira, F.A.S.; Rocha, S.M.B.; Pereira, A.P.D.A.; Prado, R.D.M.; et al. Silicon Application Influences the Prokaryotic Communities in the Rhizosphere of Sugarcane Genotypes. Applied Soil Ecology 2023, 187, 104818. [Google Scholar] [CrossRef]
- Yun, L.; Yu, Z.; Li, Y.; Luo, P.; Jiang, X.; Tian, Y.; Ding, X. Ammonia Nitrogen and Nitrite Removal by a Heterotrophic Sphingomonas Sp. Strain LPN080 and Its Potential Application in Aquaculture. Aquaculture 2019, 500, 477–484. [Google Scholar] [CrossRef]
- Ma, H.; Gao, X.; Chen, Y.; Zhu, J.; Liu, T. Fe(II) Enhances Simultaneous Phosphorus Removal and Denitrification in Heterotrophic Denitrification by Chemical Precipitation and Stimulating Denitrifiers Activity. Environmental Pollution 2021, 287, 117668. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, F.; Luo, P.; Xiao, R.; Chen, J.; Chen, L.; Wu, J. Does rice straw application reduce N2O emissions from surface ffow CWs for swine wastewater treatment? Chemosphere 2019, 226, 273–281. [Google Scholar] [CrossRef]
- Yu, H.; Liu, X.; Yang, C.; Peng, Y.; Yu, X.; Gu, H.; Zheng, X.; Wang, C.; Xiao, F.; Shu, L.; et al. Co-Symbiosis of Arbuscular Mycorrhizal Fungi (AMF) and Diazotrophs Promote Biological Nitrogen Fixation in Mangrove Ecosystems. Soil Biology and Biochemistry 2021, 161, 108382. [Google Scholar] [CrossRef]
- Cheng, J.Z.; White, J.R. Dredge-Material Created Coastal Marshes Are More Effective at Improving Water Quality than Natural Marshes in Early-Stage Development. Ecological Engineering 2022, 185, 106814. [Google Scholar] [CrossRef]
- Ma, L.; Xiong, Z.; Yao, L.; Liu, G.; Zhang, Q.; Liu, W. Soil Properties Alter Plant and Microbial Communities to Modulate Denitrification Rates in Subtropical Riparian Wetlands. Land Degrad Dev 2020, 31, 1792–1802. [Google Scholar] [CrossRef]
- Wang, W.; Pan, X.; Shu, X.; Tan, X.; Zhao, B.; Zhang, Q. Direct Evidence Indicates That Revegetation Improves Organic Carbon Limitation in Sediment Denitrification in a Eutrophic Headwater River. Ecological Engineering 2024, 198, 107132. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, Z.; Li, X.; Long, Z.; Pei, Y. Comprehensive Analysis of the Migration and Transformation of Nutrients between Sediment and Overlying Water in Complex Habitat Systems. Science of The Total Environment 2022, 852, 158433. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, C.J.; Brock, M.T.; Van Diepen, L.T.A.; Maignien, L.; Ewers, B.E.; Weinig, C. The Plant Circadian Clock Influences Rhizosphere Community Structure and Function. The ISME Journal 2018, 12, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.; Li, J.; Liu, H.; Li, B.; Yu, X.; Cao, X.; Liu, D.; Wen, L.; Zhuo, Y.; Wang, L. Characteristics of Bacterial Biodiversity and Community Structure in Non-Rhizosphere Soils along Zonal Distribution of Plants within Littoral Wetlands in Inner Mongolia, China. Global Ecology and Conservation 2020, 24, e01310. [Google Scholar] [CrossRef]
- Zhang, J.; Fan, S.; Qin, J.; Dai, J.; Zhao, F.; Gao, L.; Lian, X.; Shang, W.; Xu, X.; Hu, X. Changes in the Microbiome in the Soil of an American Ginseng Continuous Plantation. Front. Plant Sci 2020, 11, 572199. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yang, Q.; Feng, J.; Yang, Z.; Yu, C.; Zhang, J.; Ling, J.; Dong, J. Introduction of Exotic Species Sonneratia Apetala Alters Diazotrophic Community and Stimulates Nitrogen Fixation in Mangrove Sediments. Ecological Indicators 2022, 142, 109179. [Google Scholar] [CrossRef]
- Zhang, M.; Zha, J.; Dong, Y.; Zhang, Q.; Pang, S.; Tian, S.; Sun, Q. Regulation of Potential Denitrification Rates in Sediments by Microbial-Driven Elemental Coupled Metabolisms. Journal of Environmental Management 2023, 348, 119320. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Lu, Z.; Tobias, C.R.; Wang, Y.; Xiao, K.; Yu, Q.; Lin, J.; Huang, G.; Chen, N. Salt Marsh Expansion into Estuarine Mangrove Mudflats Reduces Nitrogen Removal Capacity. CATENA 2023, 232, 107459. [Google Scholar] [CrossRef]
- Wang, S.; Pi, Y.; Song, Y.; Jiang, Y.; Zhou, L.; Liu, W.; Zhu, G. Hotspot of Dissimilatory Nitrate Reduction to Ammonium (DNRA) Process in Freshwater Sediments of Riparian Zones. Water Research 2020, 173, 115539. [Google Scholar] [CrossRef] [PubMed]
- Ling, N.; Wang, T.; Kuzyakov, Y. Rhizosphere Bacteriome Structure and Functions. Nat Commun 2022, 13, 836. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Zeng, J.; Zhao, D.; Huang, R.; Yu, Z.; Wu, Q.L. Contrasting Patterns in Diversity and Community Assembly of Phragmites Australis Root-Associated Bacterial Communities from Different Seasons. Appl Environ Microbiol 2020, 86, e00379–20. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Zhang, S.; Yan, P.; Huo, J.; Aurangzeib, M. Microbial Community and Their Potential Functions after Natural Vegetation Restoration in Gullies of Farmland in Mollisols of Northeast China. Land 2022, 11, 2231. [Google Scholar] [CrossRef]
- Ye, F.; Duan, L.; Sun, Y.; Yang, F.; Liu, R.; Gao, F.; Wang, Y.; Xu, Y. Nitrogen Removal in Freshwater Sediments of Riparian Zone: N-Loss Pathways and Environmental Controls. Front. Microbiol. 2023, 14, 1239055. [Google Scholar] [CrossRef]
- Rankovi’c, V.; Radulovi’c, J.; Radojevi’c, I.; Ostoji’c, A.; Čomić, L. Neural network modeling of dissolved oxygen in the Gruža reservoir, Serbia. Ecological Modelling 2010, 221, 1239–1244. [Google Scholar] [CrossRef]
- Lu, B.; Xu, Z.; Li, J.; Chai, X. Removal of Water Nutrients by Different Aquatic Plant Species: An Alternative Way to Remediate Polluted Rural Rivers. Ecological Engineering 2018, 110, 18–26. [Google Scholar] [CrossRef]
- Wu, H.; Wang, X.; He, X.; Zhang, S.; Liang, R.; Shen, J. Effects of Root Exudates on Denitrifier Gene Abundance, Community Structure and Activity in a Micro-Polluted Constructed Wetland. Science of The Total Environment 2017, 598, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Husson, O. Redox Potential (Eh) and PH as Drivers of Soil/Plant/Microorganism Systems: A Transdisciplinary Overview Pointing to Integrative Opportunities for Agronomy. Plant Soil 2013, 362, 389–417. [Google Scholar] [CrossRef]
- Xia, P.; Zhang, J.; Liu, J.; Yu, L. Shifts of Sediment Bacterial Community and Respiration along a Successional Gradient in a Typical Karst Plateau Lake Wetland (China). J. Ocean. Limnol. 2021, 39, 880–891. [Google Scholar] [CrossRef]
- Guo, J.; Wang, X.; Cao, X.; Qi, W.; Peng, J.; Liu, H.; Qu, J. The Influence of Wet-to-Dry Season Shifts on the Microbial Community Stability and Nitrogen Cycle in the Poyang Lake Sediment. Science of The Total Environment 2023, 903, 166036. [Google Scholar] [CrossRef] [PubMed]
- Enebe, M.; Babalola, O. The Influence of Soil Fertilization on the Distribution and Diversity of Phosphorus Cycling Genes and Microbes Community of Maize Rhizosphere Using Shotgun Metagenomics. Genes 2021, 12, 1022. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Deng, Y.; Zhang, P.; Xue, K.; Liang, Y.; Van Nostrand, J.D.; Yang, Y.; He, Z.; Wu, L.; Stahl, D.A.; et al. Stochasticity, Succession, and Environmental Perturbations in a Fluidic Ecosystem. Proc. Natl. Acad. Sci. U.S.A. 2014, 111. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-H.; Yuan, S.-W.; Che, F.-F.; Wan, X.; Wang, Y.-F.; Yang, D.-H.; Yang, H.-J.; Zhu, D.; Chen, P. Strong Bacterial Stochasticity and Fast Fungal Turnover in Taihu Lake Sediments, China. Environmental Research 2023, 237, 116954. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Ping, Y.; Cui, L.; Zhang, X.; Li, W.; Hu, Y.; Cornelissen, J.H.C. Nutrient Resorption from Leaves of Wetland Plants in a Constructed Wetland Depends on Green Leaf Nutrient Content and Life Form. Wetlands 2020, 40, 983–991. [Google Scholar] [CrossRef]
- Hu, J.; Yu, H.; Li, Y.; Wang, J.; Lv, T.; Liu, C.; Yu, D. Variation in resource allocation strategies and environmental driving factors for different life-forms of aquatic plants in cold temperate zones. Journal of Ecology 2021, 109, 3046. [Google Scholar] [CrossRef]
- Bao, Y.; Huo, Y.; Duan, Y.; He, P.; Wu, M.; Yang, N.; Sun, B. Growth and Nutrient Uptake of Myriophyllum Spicatum under Different Nutrient Conditions and Its Potential Ecosystem Services in an Enclosed Sea Area in the East China Sea. Marine Pollution Bulletin 2020, 151, 110801. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Li, J.; Ma, X.; Lv, T.; Wang, L.; Li, J.; Liu, C. Community Structure and Function of Epiphytic Bacteria Attached to Three Submerged Macrophytes. Science of The Total Environment 2022, 835, 155546. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, J.; Liu, X.; Zhong, H.; Wang, B.; Kong, Z.; Wu, L. Tracking the Changes of Wetland Soil Bacterial Community and Metabolic Potentials under Drought and Flooding Conditions in Experimental Microcosms. J Soils Sediments 2021, 21, 2404–2417. [Google Scholar] [CrossRef]
- Wang, X.; Jain, A.; Chen, B.; Wang, Y.; Jin, Q.; Yugandhar, P.; Xu, Y.; Sun, S.; Hu, F. Differential Efficacy of Water Lily Cultivars in Phytoremediation of Eutrophic Water Contaminated with Phosphorus and Nitrogen. Plant Physiology and Biochemistry 2022, 171, 139–146. [Google Scholar] [CrossRef]
- Xu, C.; Chen, L.; Chen, S.; Chu, G.; Wang, D.; Zhang, X. Effects of Rhizosphere Oxygen Concentration on Root Physiological Characteristics and Anatomical Structure at the Tillering Stage of Rice. Annals of Applied Biology 2020, 177, 61–73. [Google Scholar] [CrossRef]
- Pietrangelo, L.; Bucci, A.; Maiuro, L.; Bulgarelli, D.; Naclerio, G. Unraveling the Composition of the Root-Associated Bacterial Microbiota of Phragmites Australis and Typha Latifolia. Front. Microbiol. 2018, 9, 1650. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Isobe, K.; Nishizawa, T.; Zhu, L.; Shiratori, Y.; Ohte, N.; Koba, K.; Otsuka, S.; Senoo, K. Higher Diversity and Abundance of Denitrifying Microorganisms in Environments than Considered Previously. The ISME Journal 2015, 9, 1954–1965. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Li, G.; Li, X.; Li, C.; Li, J.; Zhao, C.; Cui, J.; Du, C.; Tian, Z.; Shi, Y.; et al. Effect of Microbial Network Complexity and Stability on Nitrogen and Sulfur Pollutant Removal during Sediment Remediation in Rivers Affected by Combined Sewer Overflows. Chemosphere 2023, 331, 138832. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, F.; Chen, Y.; Zou, W.; Zhu, Z. Diazotrophic Community in the Sediments of Poyang Lake in Response to Water Level Fluctuations. Front. Microbiol. 2024, 15, 1324313. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhao, Z.; Li, J.; Wang, H.; Guo, G.; Wu, W. Effects of Muddy Water Irrigation with Different Sediment Particle Sizes and Sediment Concentrations on Soil Microbial Communities in the Yellow River Basin of China. Agricultural Water Management 2022, 270, 107750. [Google Scholar] [CrossRef]
- Hu, J.; Yu, H.; Li, Y.; Wang, J.; Lv, T.; Liu, C.; Yu, D. Variation in resource allocation strategies and environmental driving factors for different life-forms of aquatic plants in cold temperate zones. Journal of Ecology 2021, 109, 3046. [Google Scholar] [CrossRef]
- Wang, L.; Xing, P.; Li, H.; Zhou, L.; Wu, Q.L. Distinct Intra-Lake Heterogeneity of Diazotrophs in a Deep Oligotrophic Mountain Lake. Microb Ecol 2020, 79, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, T.; McIlroy, S.J.; Tyson, G.W.; Guo, J. Phylogenetic and Metabolic Diversity of Microbial Communities Performing Anaerobic Ammonium and Methane Oxidations under Different Nitrogen Loadings. ISME Communications 2023, 3, 39. [Google Scholar] [CrossRef]
- Pan, Y.; Sun, R.-Z.; Wang, Y.; Chen, G.-L.; Fu, Y.-Y.; Yu, H.-Q. Carbon Source Shaped Microbial Ecology, Metabolism and Performance in Denitrification Systems. Water Research 2023, 243, 120330. [Google Scholar] [CrossRef]
- Shao, Y.; He, Q.; Fu, Y.; Zhang, G.; Liu, Y. Environmental impact and variation analysis of different CaO2 and Ca (NO3)2 dosing modes on microbial community in black-odorous sediment. Process Safety and Environmental Protection 2022, 167, 641–650. [Google Scholar] [CrossRef]
- Tian, L.Q.; Jiang, H.L.; Bai, L.L.; Wang, C.L.; Xu, S.Q. Q. Biological Nitrogen Fixation in Sediments of a Cyanobacterial Bloom-Occurring Bay in One Eutrophic Shallow Lake: Occurrence and Related Environmental Factors. Journal of Geophysical Research: Biogeosciences 2021, 126, 1–15. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, D.; Guo, Y.; Zhao, J.; Bao, Z. Rhizosphere-Associated Anammox Bacterial Diversity and Abundance of Nitrogen Cycle-Related Functional Genes of Emergent Macrophytes in Eutrophic Wetlands. Curr Microbiol 2024, 81, 107. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, Y.; Peng, L.; Liu, G.; Wan, X.; Hua, Y.; Zhu, D.; Hamilton, D.P. Diversity of Anammox Bacteria and Abundance of Functional Genes for Nitrogen Cycling in the Rhizosphere of Submerged Macrophytes in a Freshwater Lake in Summer. J Soils Sediments 2019, 19, 3648–3656. [Google Scholar] [CrossRef]
- Fang, J.; Lü, T.; Liu, J.; He, S.; Yang, X.; Dou, H.; Zhang, H. Responses of Nitrogen Cycling and Related Microorganisms to Brackish Wetlands Formed by Evapotranspiration. Pedosphere 2024, 34, 252–266. [Google Scholar] [CrossRef]
- Zhang, S.; Qin, W.; Xia, X.; Xia, L.; Li, S.; Zhang, L.; Bai, Y.; Wang, G. Ammonia Oxidizers in River Sediments of the Qinghai-Tibet Plateau and Their Adaptations to High-Elevation Conditions. Water Research 2020, 173, 115589. [Google Scholar] [CrossRef] [PubMed]
- Wallenstein, M.D.; Myrold, D.D.; Firestone, M.; Voytek, M. Environmental Controls on Denitrifying Communities And Denitrification Rates: Insights From Molecular Methods. Ecological Applications 2006, 16, 2143–2152. [Google Scholar] [CrossRef]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiology and Molecular Biology Reviews 1997, 61, 533–616. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Wang, F.; Ren, L.; Ma, S.; Li, S.; Liu, L.; Wang, J. Ratio of Nitrate to Ammonium Mainly Drives Soil Bacterial Dynamics Involved in Nitrate Reduction Processes. Applied Soil Ecology 2022, 169, 104164. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, C.; Nelson, W.C.; Shi, L.; Xu, F.; Liu, Y.; Yan, A.; Zhong, L.; Thompson, C.; Fredrickson, J.K.; et al. Effect of Water Chemistry and Hydrodynamics on Nitrogen Transformation Activity and Microbial Community Functional Potential in Hyporheic Zone Sediment Columns. Environ. Sci. Technol. 2017, 51, 4877–4886. [Google Scholar] [CrossRef]
- Kang, P.; Chen, S.; Huang, T.; Zhang, H.; Shang, P.; Zhao, Z.; Tan, X. Denitrification Characteristics and Functional Genes of Denitrifying Bacteria Under Aerobic or Anaerobic Conditions. Huan Jing Ke Xue 2018, 8, 3789–3796. [Google Scholar] [CrossRef]
- Wang, F.; Cui, Q.; Liu, W.; Jiang, W.; Ai, S.; Liu, W.; Bian, D. Synergistic Denitrification Mechanism of Domesticated Aerobic Denitrifying Bacteria in Low-Temperature Municipal Wastewater Treatment. npj Clean Water 2024, 7, 6. [Google Scholar] [CrossRef]
- Maruyama, R.; Yasumoto, K.; Mizusawa, N.; Iijima, M.; Yasumoto-Hirose, M.; Iguchi, A.; Hermawan, O.R.; Hosono, T.; Takada, R.; Song, K.-H.; et al. Metagenomic Analysis of the Microbial Communities and Associated Network of Nitrogen Metabolism Genes in the Ryukyu Limestone Aquifer. Sci Rep 2024, 14, 4356. [Google Scholar] [CrossRef] [PubMed]
- Saarenheimo, J.; Rissanen, A.J.; Arvola, L.; Nykänen, H.; Lehmann, M.F.; Tiirola, M. Genetic and Environmental Controls on Nitrous Oxide Accumulation in Lakes. PLoS ONE 2015, 10, e0121201. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, Y.; Lv, J.; He, X.; Wang, J.; Teng, D.; Jiang, L.; Wang, H.; Lv, G. Rhizosphere Effect Alters the Soil Microbiome Composition and C, N Transformation in an Arid Ecosystem. Applied Soil Ecology 2022, 170, 104296. [Google Scholar] [CrossRef]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiology and Molecular Biology Reviews 1997, 61, 533–616. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.Y.; Van Meter, K.J.; Byrnes, D.K.; Basu, N.B. Maximizing US Nitrate Removal through Wetland Protection and Restoration. Nature 2020, 588, 625–630. [Google Scholar] [CrossRef]
- Wang, S.; Liu, C.; Wang, X.; Yuan, D.; Zhu, G. Dissimilatory Nitrate Reduction to Ammonium (DNRA) in Traditional Municipal Wastewater Treatment Plants in China: Widespread but Low Contribution. Water Research 2020, 179, 115877. [Google Scholar] [CrossRef]
- Wang, C.; He, T.; Zhang, M.; Zheng, C.; Yang, L.; Yang, L. Review of the Mechanisms Involved in Dissimilatory Nitrate Reduction to Ammonium and the Efficacies of These Mechanisms in the Environment. Environmental Pollution 2024, 345, 123480. [Google Scholar] [CrossRef]
- Huang, X.; Luoluo; Xie, D.; Li, Z. Dissimilatory Nitrate Reduction to Ammonium in Four Pseudomonas Spp. under Aerobic Conditions. Heliyon 2023, 9, e14983. [Google Scholar] [CrossRef]
- Wei, Z.; Jin, K.; Li, C.; Wu, M.; Shan, J.; Yan, X. Environmental Factors Controlling Dissimilatory Nitrate Reduction to Ammonium in Paddy Soil. J Soil Sci Plant Nutr 2022, 22, 4241–4248. [Google Scholar] [CrossRef]
- Pang, Y.; Ji, G. Biotic Factors Drive Distinct DNRA Potential Rates and Contributions in Typical Chinese Shallow Lake Sediments. Environmental Pollution 2019, 254, 112903. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Md.M.; Roberts, K.L.; Grace, M.R.; Kessler, A.J.; Cook, P.L.M. Role of Organic Carbon, Nitrate and Ferrous Iron on the Partitioning between Denitrification and DNRA in Constructed Stormwater Urban Wetlands. Science of The Total Environment 2019, 666, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Kasak, K.; Espenberg, M.; Anthony, T.L.; Tringe, S.G.; Valach, A.C.; Hemes, K.S.; Silver, W.L.; Mander, Ü.; Kill, K.; McNicol, G.; et al. Restoring Wetlands on Intensive Agricultural Lands Modifies Nitrogen Cycling Microbial Communities and Reduces N2O Production Potential. Journal of Environmental Management 2021, 299, 113562. [Google Scholar] [CrossRef] [PubMed]
- Broman, E.; Zilius, M.; Samuiloviene, A.; Vybernaite-Lubiene, I.; Politi, T.; Klawonn, I.; Voss, M.; Nascimento, F.J.A.; Bonaglia, S. Active DNRA and Denitrification in Oxic Hypereutrophic Waters. Water Research 2021, 194, 116954. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Bai, Y.; Qu, J. The Phragmites Root-Inhabiting Microbiome: A Critical Review on Its Composition and Environmental Application. Engineering 2022, 9, 42–50. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, S.; Lu, J.; Duan, R.; Chen, H.; Ma, Y.; Si, T.; Luo, M. Reed Restoration Decreased Nutrients in Wetlands with Dredged Sediments: Microbial Community Assembly and Function in Rhizosphere. Journal of Environmental Management 2023, 344, 118700. [Google Scholar] [CrossRef]
- Singh, R.P.; Reddy, C.R.K. Seaweed–microbial interactions: key functions of seaweed-associated bacteria. FEMS Microbiology Ecology 2014, 88, 213–230. [Google Scholar] [CrossRef] [PubMed]
- Zehr, J.P.; Braun, S.; Chen, Y.; Mellon, M. Nitrogen Fixation in the Marine Environment: Relating Genetic Potential to Nitrogenase Activity. Journal of Experimental Marine Biology and Ecology 1996, 203, 61–73. [Google Scholar] [CrossRef]
- Barraquio, W.L.; Watanabe, I. Occurrence of Aerobic Nitrogen Fixing Bacteria in Wetland and Dryland Plants. Soil Science and Plant Nutrition 1981, 27, 121–125. [Google Scholar] [CrossRef]
- Ketelboeter, L.M.; Mitra, S.; Gyaneshwar, P. A Thiamine Transporter Is Required for Biofilm Formation by Rhizobium Sp. IRBG74. FEMS Microbiology Letters 2023, 370, fnad046. [Google Scholar] [CrossRef]
- Luo, Z.; Qiu, Z.; Wei, Q.; Du Laing, G.; Zhao, Y.; Yan, C. Dynamics of Ammonia-Oxidizing Archaea and Bacteria in Relation to Nitrification along Simulated Dissolved Oxygen Gradient in Sediment–Water Interface of the Jiulong River Estuarine Wetland, China. Environ Earth Sci 2014, 72, 2225–2237. [Google Scholar] [CrossRef]
- Deng, D.; Pan, Y.; Liu, G.; Liu, W.; Ma, L. Seeking the Hotspots of Nitrogen Removal: A Comparison of Sediment Denitrification Rate and Denitrifier Abundance among Wetland Types with Different Hydrological Conditions. Science of The Total Environment 2020, 737, 140253. [Google Scholar] [CrossRef]
- Li, Y.; Jin, H.; Chen, J.; Wang, D.; Yang, Z.; Wang, B.; Zhuang, Y.; Wang, R. Nitrogen Removal through Sediment Denitrification in the Yangtze Estuary and Its Adjacent East China Sea: A Nitrate Limited Process during Summertime. Science of The Total Environment 2021, 795, 148616. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhou, Y.; Yang, F.; Guan, Q.; Li, Q.; Liang, H.; Zhao, J. Impact of Environmental Factors on the Diversity of Nitrogen-Removal Bacteria in Wetlands in the Sanmenxia Reservoir of the Yellow River. J Soils Sediments 2023, 23, 512–525. [Google Scholar] [CrossRef]
- Zhang, M.; Daraz, U.; Sun, Q.; Chen, P.; Wei, X. Denitrifier Abundance and Community Composition Linked to Denitrification Potential in River Sediments. Environ Sci Pollut Res 2021, 28, 51928–51939. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; She, D.; Pan, Y.; Abulaiti, A.; Huang, Y.; Liu, R.; Wang, F.; Xia, Y.; Shan, J. Ditch Level-Dependent N Removal Capacity of Denitrification and Anammox in the Drainage System of the Ningxia Yellow River Irrigation District. Science of The Total Environment 2024, 916, 170314. [Google Scholar] [CrossRef]
- Nifong, R.L.; Taylor, J.M. Vegetation and Residence Time Interact to Influence Metabolism and Net Nutrient Uptake in Experimental Agricultural Drainage Systems. Water 2021, 13, 1416. [Google Scholar] [CrossRef]
- Rachel, L.N.; Jason, M.T. Vegetation and Residence Time Interact to Influence Metabolism and Net Nutrient Uptake in Experimental Agricultural Drainage Systems. Water 2021, 13, 1416. [Google Scholar] [CrossRef]
- Kong, Y.; Zhang, H.; Tian, L.; Yuan, J.; Chen, Y.; Li, Y.; Chen, J.; Chang, S.X.; Fang, Y.; Tavakkoli, E.; et al. Relationships between Denitrification Rates and Functional Gene Abundance in a Wetland: The Roles of Single- and Multiple-Species Plant Communities. Science of The Total Environment 2023, 863, 160913. [Google Scholar] [CrossRef]
- Surey, R.; Kaiser, K.; Schimpf, C.M.; Mueller, C.W.; Böttcher, J.; Mikutta, R. Contribution of Particulate and Mineral-Associated Organic Matter to Potential Denitrification of Agricultural Soils. Front. Environ. Sci. 2021, 9, 640534. [Google Scholar] [CrossRef]
- Davidsson, T.E.; Ståhl, M. The Influence of Organic Carbon on Nitrogen Transformations in Five Wetland Soils. Soil Science Soc of Amer J 2000, 64, 1129–1136. [Google Scholar] [CrossRef]
- Sigleo, A.C. Denitrification Rates Across a Temperate North Pacific Estuary, Yaquina Bay, Oregon. Estuaries and Coasts 2019, 42, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Man, Z.; Xie, C.; Qin, Y.; Che, S. Effects of Revetments on Nitrification and Denitrification Potentials in the Urban River–Riparian Interface. Land 2024, 13, 333. [Google Scholar] [CrossRef]
- Li, S.; Twilley, R.R.; Hou, A. Heterotrophic Nitrogen Fixation in Response to Nitrate Loading and Sediment Organic Matter in an Emerging Coastal Deltaic Floodplain within the Mississippi River Delta Plain. Limnology & Oceanography 2021, 66, 1961–1978. [Google Scholar] [CrossRef]
- Lin, X.; Li, X.; Gao, D.; Liu, M.; Cheng, L. Ammonium Production and Removal in the Sediments of Shanghai River Networks: Spatiotemporal Variations, Controlling Factors, and Environmental Implications. JGR Biogeosciences 2017, 122, 2461–2478. [Google Scholar] [CrossRef]
- Semedo, M.; Song, B. From Genes to Nitrogen Removal: Determining the Impacts of Poultry Industry Wastewater on Tidal Creek Denitrification. Environ. Sci. Technol. 2020, 54, 146–157. [Google Scholar] [CrossRef]
- Tsiknia, M.; Paranychianakis, N.V.; Varouchakis, E.A.; Nikolaidis, N.P. Environmental Drivers of the Distribution of Nitrogen Functional Genes at a Watershed Scale. FEMS Microbiology Ecology 2015, 91. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Peng, Y.; Gao, H. Enhanced Long-Term Advanced Denitrogenation from Nitrate Wastewater by Anammox Consortia: Dissimilatory Nitrate Reduction to Ammonium (DNRA) Coupling with Anammox in an Upflow Biofilter Reactor Equipped with EDTA-2Na/Fe(II) Ratio and PH Control. Bioresource Technology 2020, 305, 123083. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yin, G.; Hou, L.; Liu, M.; Zheng, Y.; Han, P.; Dong, H.; Liang, X.; Gao, D.; Liu, C. Nitrogen Removal Processes Coupled with Nitrification in Coastal Sediments off the North East China Sea. J Soils Sediments 2021, 21, 3289–3299. [Google Scholar] [CrossRef]
- Hu, Y.; Hong, Y.; Ye, J.; Wu, J.; Wang, Y.; Ye, F.; Chang, X.; Long, A. Shift of DNRA Bacterial Community Composition in Sediment Cores of the Pearl River Estuary and the Impact of Environmental Factors. Ecotoxicology 2021, 30, 1689–1703. [Google Scholar] [CrossRef]
- Zhao, B.; Li, X.; Wang, Y.; Tan, X.; Qi, W.; Li, H.; Wei, J.; You, Y.; Shi, W.; Zhang, Q. Dissimilatory Nitrate Reduction and Functional Genes in Two Subtropical Rivers, China. Environ Sci Pollut Res 2021, 28, 68155–68173. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, N.; Shao, X.; Gao, D.; Xia, J.; Cui, Q.; Zhang, D. Effects of Coastal Marsh Conversion to Shrimp Aquaculture Ponds on Sediment Nitrogen Fixation. Front. Mar. Sci. 2022, 9, 1034145. [Google Scholar] [CrossRef]
- Wang, R.; Li, X.; Hou, L.; Liu, M.; Zheng, Y.; Yin, G.; Yang, Y. Nitrogen Fixation in Surface Sediments of the East China Sea: Occurrence and Environmental Implications. Marine Pollution Bulletin 2018, 137, 542–548. [Google Scholar] [CrossRef]
- Huang, X.; Feng, J.; Yang, Q.; Chen, L.; Zhang, J.; Yang, B.; Tang, X.; Yu, C.; Ling, J.; Dong, J. High Site Elevation Enhanced Nitrogen Fixation and the Stability of Diazotrophic Community in Planted Sonneratia Apetala Mangrove Sediments. Applied Soil Ecology 2023, 191, 105059. [Google Scholar] [CrossRef]
- Spinette, R.; Brown, S.; Ehrlich, A.; Puggioni, G.; Deacutis, C.; Jenkins, B. Diazotroph Activity in Surface Narragansett Bay Sediments in Summer Is Stimulated by Hypoxia and Organic Matter Delivery. Mar. Ecol. Prog. Ser. 2019, 614, 35–50. [Google Scholar] [CrossRef]
- Luo, Z.; Zhong, Q.; Han, X.; Hu, R.; Liu, X.; Xu, W.; Wu, Y.; Huang, W.; Zhou, Z.; Zhuang, W.; et al. Depth-Dependent Variability of Biological Nitrogen Fixation and Diazotrophic Communities in Mangrove Sediments. Microbiome 2021, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Hou, L.; Zhang, Z.; Ge, J.; Li, M.; Yin, G.; Han, P.; Dong, H.; Liang, X.; Gao, J.; et al. Overlooked Contribution of Water Column to Nitrogen Removal in Estuarine Turbidity Maximum Zone (TMZ). Science of The Total Environment 2021, 788, 147736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qin, W.; Bai, Y.; Zhang, Z.; Wang, J.; Gao, H.; Gu, J. Linkages between anammox and denitrifying bacterial communities and nitrogen loss rates in highelevation rivers. Limnology and Oceanography 2020, 9999, 1–14. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Q.; Yang, A.; Hou, L.; Zheng, Y.; Zhai, W.; Gong, J. Incorporation of Microbial Functional Traits in Biogeochemistry Models Provides Better Estimations of Benthic Denitrification and Anammox Rates in Coastal Oceans. JGR Biogeosciences 2018, 123, 3331–3352. [Google Scholar] [CrossRef]
- Wang, Z.; Ahmad, H.A.; Teng, Z.-J.; Sun, M.; Ismail, S.; Qiao, Z.; Ni, S.-Q. Widespread but Overlooked DNRA Process in a Full-Scale Simultaneous Partial Nitrification, Anammox, and Denitrification Plant. ACS EST Water 2022, 2, 1360–1369. [Google Scholar] [CrossRef]
- Taylor, J.M.; Moore, M.T.; Scott, J.T. Contrasting Nutrient Mitigation and Denitrification Potential of Agricultural Drainage Environments with Different Emergent Aquatic Macrophytes. J. Environ. Qual. 2015, 44, 1304–1314. [Google Scholar] [CrossRef] [PubMed]
- Speir, S.L.; Taylor, J.M.; Thad, S.J. Seasonal differences in relationships between nitrate concentration and denitrification rates in ditch sediments vegetated with rice cutgrass. Journal of Environmental Quality 2017, 46, 1500–1509. [Google Scholar] [CrossRef] [PubMed]
- Pandey, C.B.; Kumar, U.; Kaviraj, M.; Minick, K.J.; Mishra, A.K.; Singh, J.S. DNRA: A Short-Circuit in Biological N-Cycling to Conserve Nitrogen in Terrestrial Ecosystems. Science of The Total Environment 2020, 738, 139710. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Dai, H.; Yuan, S. The Competition between Heterotrophic Denitrification and DNRA Pathways in Hyporheic Zone and Its Impact on the Fate of Nitrate. Journal of Hydrology 2023, 626, 130175. [Google Scholar] [CrossRef]
- Quick, A.M.; Reeder, W.J.; Farrell, T.B.; Tonina, D.; Feris, K.P.; Benner, S.G. Nitrous Oxide from Streams and Rivers: A Review of Primary Biogeochemical Pathways and Environmental Variables. Earth-Science Reviews 2019, 191, 224–262. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, F.; Huang, Z.; Xiao, R.; Zhu, H.; Wu, J. Are vegetated drainage ditches effective for nitrogen removal under cold temperatures? Bioresource Technology 2020, 301, 122744. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Yan, Z.; Wang, C.; Xu, S.; Jiang, H. Habitat Heterogeneity Induces Regional Differences in Sediment Nitrogen Fixation in Eutrophic Freshwater Lake. Science of The Total Environment 2021, 772, 145594. [Google Scholar] [CrossRef] [PubMed]
- Aoki, L.; McGlathery, K. Restoration Enhances Denitrification and DNRA in Subsurface Sediments of Zostera Marina Seagrass Meadows. Mar. Ecol. Prog. Ser. 2018, 602, 87–102. [Google Scholar] [CrossRef]
- Hansen, J.; Reidenbach, M. Wave and Tidally Driven Flows in Eelgrass Beds and Their Effect on Sediment Suspension. Mar. Ecol. Prog. Ser. 2012, 448, 271–287. [Google Scholar] [CrossRef]
- Aoki, L.R.; McGlathery, K.J.; Oreska, M.P.J. Seagrass Restoration Reestablishes the Coastal Nitrogen Filter through Enhanced Burial. Limnology & Oceanography 2020, 65, 1–12. [Google Scholar] [CrossRef]
Figure 1.
Conceptual model for the N cycle in aquatic ecosystems. Arrows represent the key N cycling processes.
Figure 1.
Conceptual model for the N cycle in aquatic ecosystems. Arrows represent the key N cycling processes.
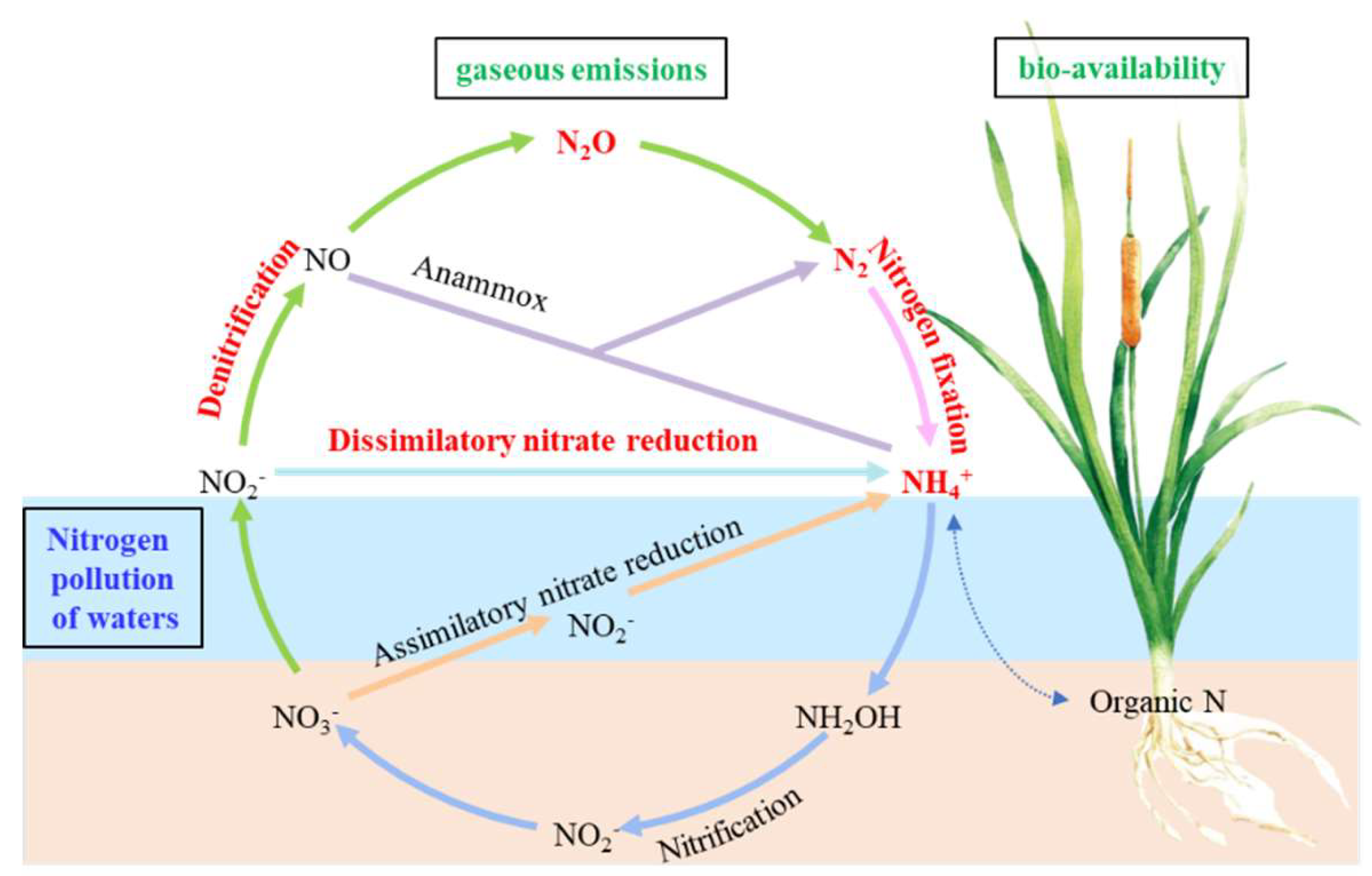
Figure 2.
The geographical location and distribution of the sample plots in various restored stands. A-Phr, artificially cultivated ditch with P. australis rhizosphere sediment; A-Typ, artificially cultivated ditch with T. orientalis rhizosphere sediment; N-Phr, natural ditch with P. australis rhizosphere sediment; N-Typ, natural ditch with T. orientalis rhizosphere sediment.
Figure 2.
The geographical location and distribution of the sample plots in various restored stands. A-Phr, artificially cultivated ditch with P. australis rhizosphere sediment; A-Typ, artificially cultivated ditch with T. orientalis rhizosphere sediment; N-Phr, natural ditch with P. australis rhizosphere sediment; N-Typ, natural ditch with T. orientalis rhizosphere sediment.
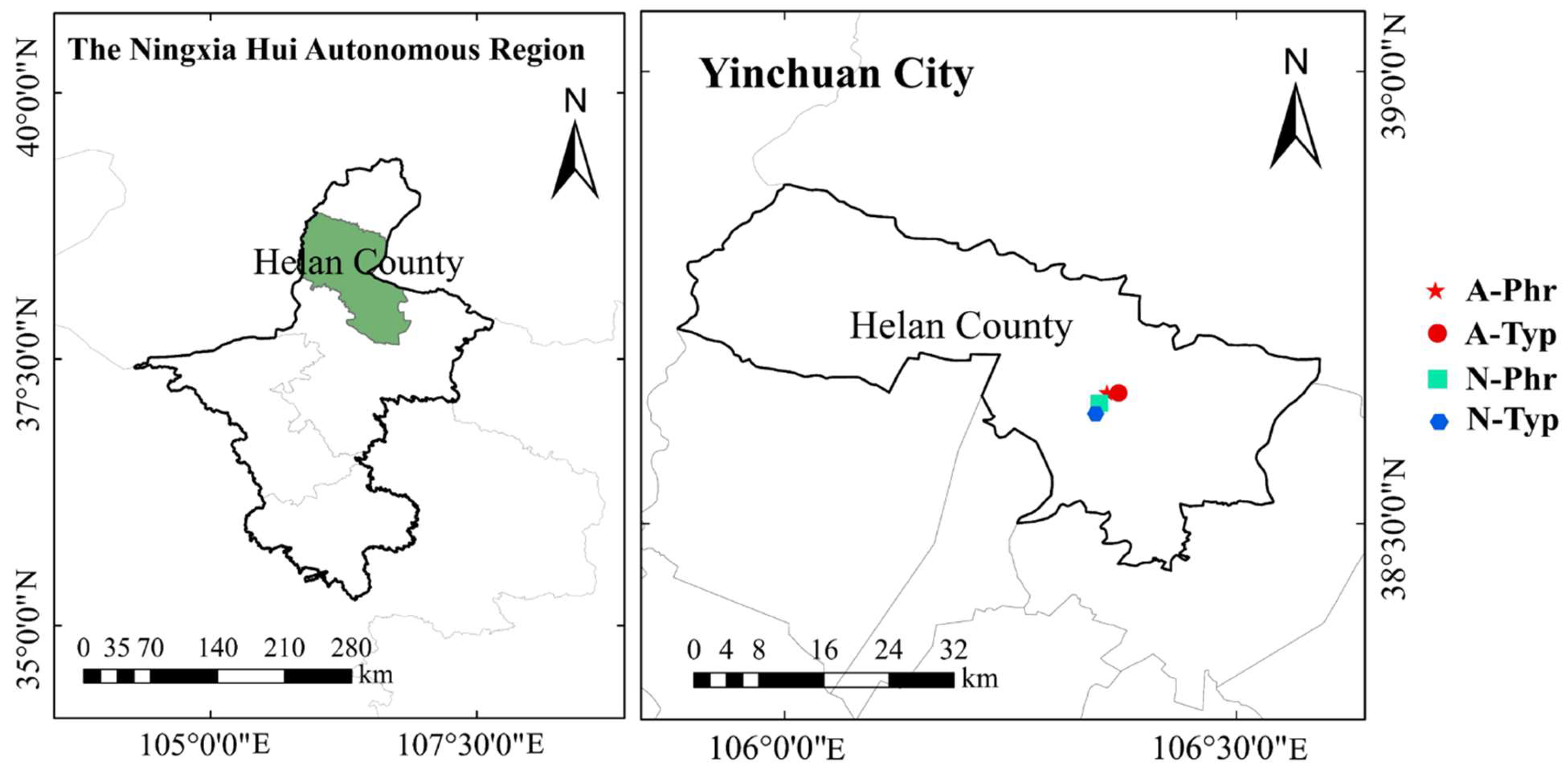
Figure 3.
Nitrogen denitrification rate (a), dissimilatory nitrate reduction to ammonium rate (b), and nitrogen fixation rate (c) in all sediment samples. DR is the nitrogen denitrification rate, DNRAR is the dissimilatory nitrate reduction to ammonium rate, and NFR is the nitrogen fixation rate.
Figure 3.
Nitrogen denitrification rate (a), dissimilatory nitrate reduction to ammonium rate (b), and nitrogen fixation rate (c) in all sediment samples. DR is the nitrogen denitrification rate, DNRAR is the dissimilatory nitrate reduction to ammonium rate, and NFR is the nitrogen fixation rate.
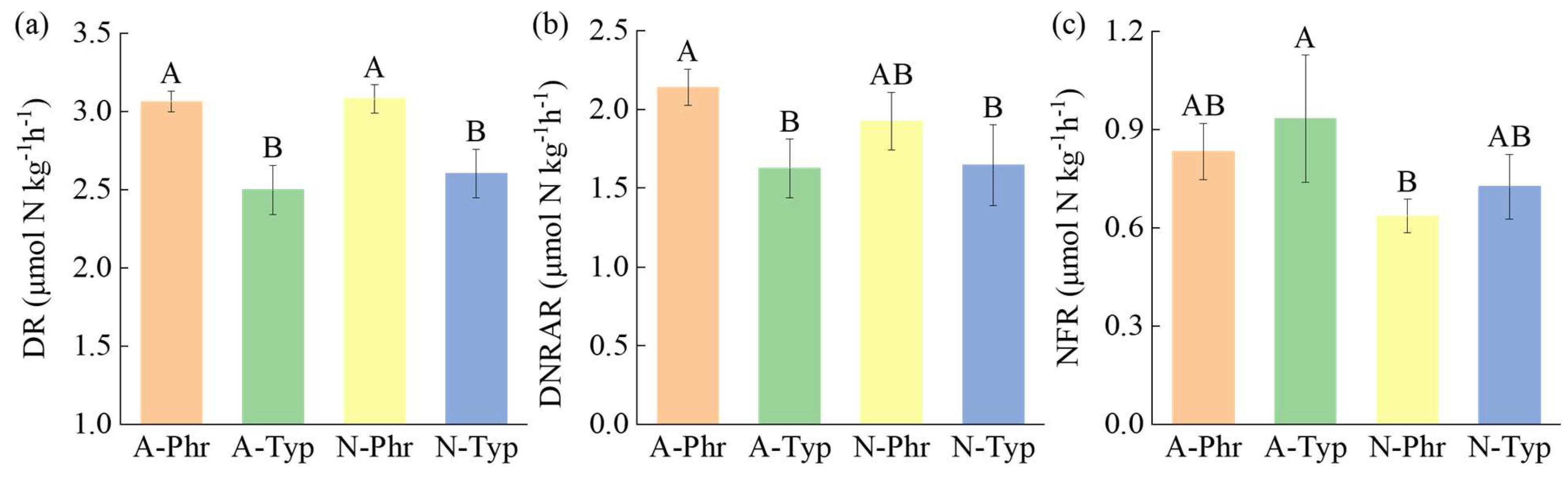
Figure 4.
Alpha diversity indices, including Chao1 (a), ACE (b), and Shannon (c) indices; relative abundance at the phylum level of bacteria (d); and functional and relative abundance at the genus level of bacteria involved in the nitrogen cycle (e) in different rhizosphere sediment samples.
Figure 4.
Alpha diversity indices, including Chao1 (a), ACE (b), and Shannon (c) indices; relative abundance at the phylum level of bacteria (d); and functional and relative abundance at the genus level of bacteria involved in the nitrogen cycle (e) in different rhizosphere sediment samples.
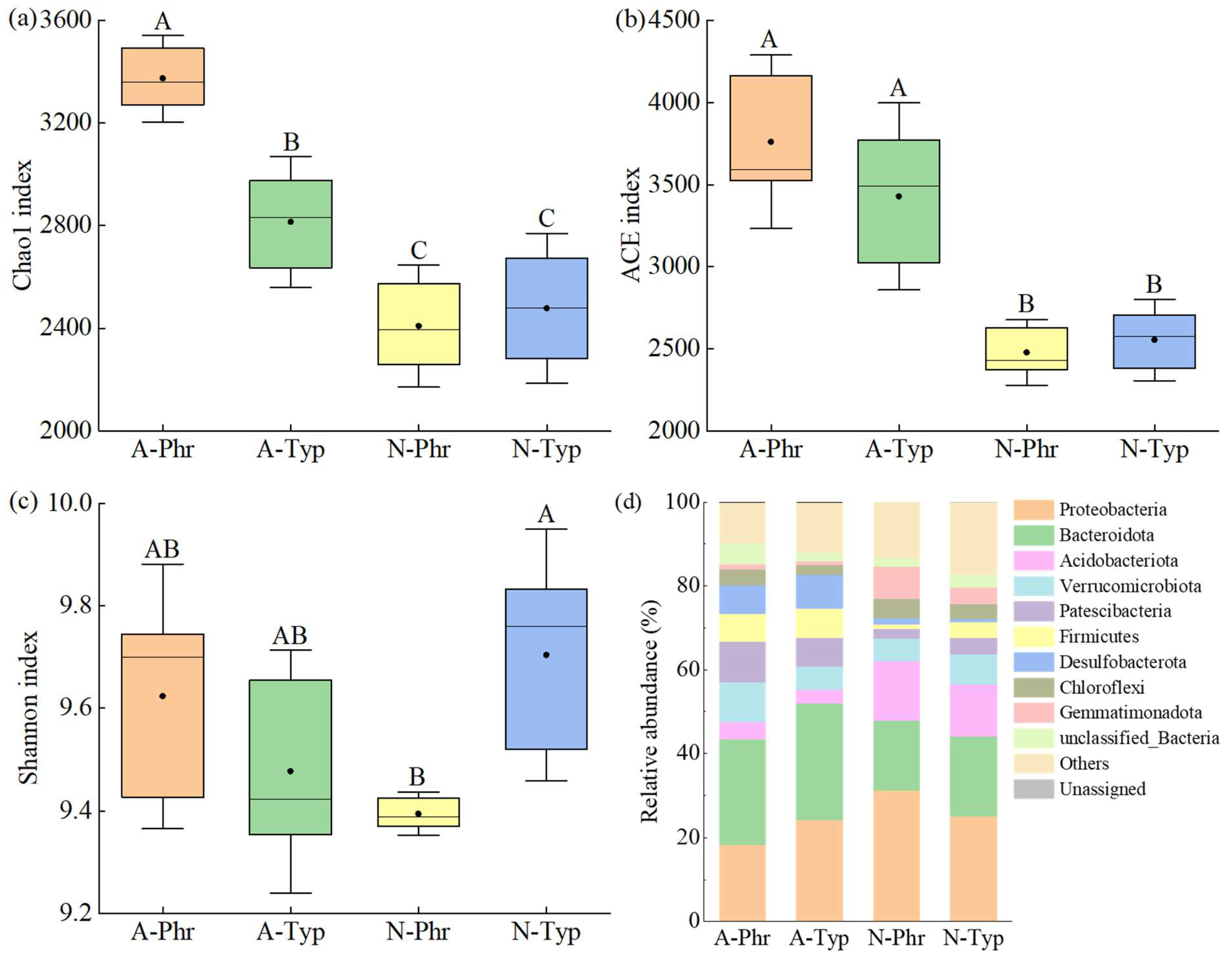
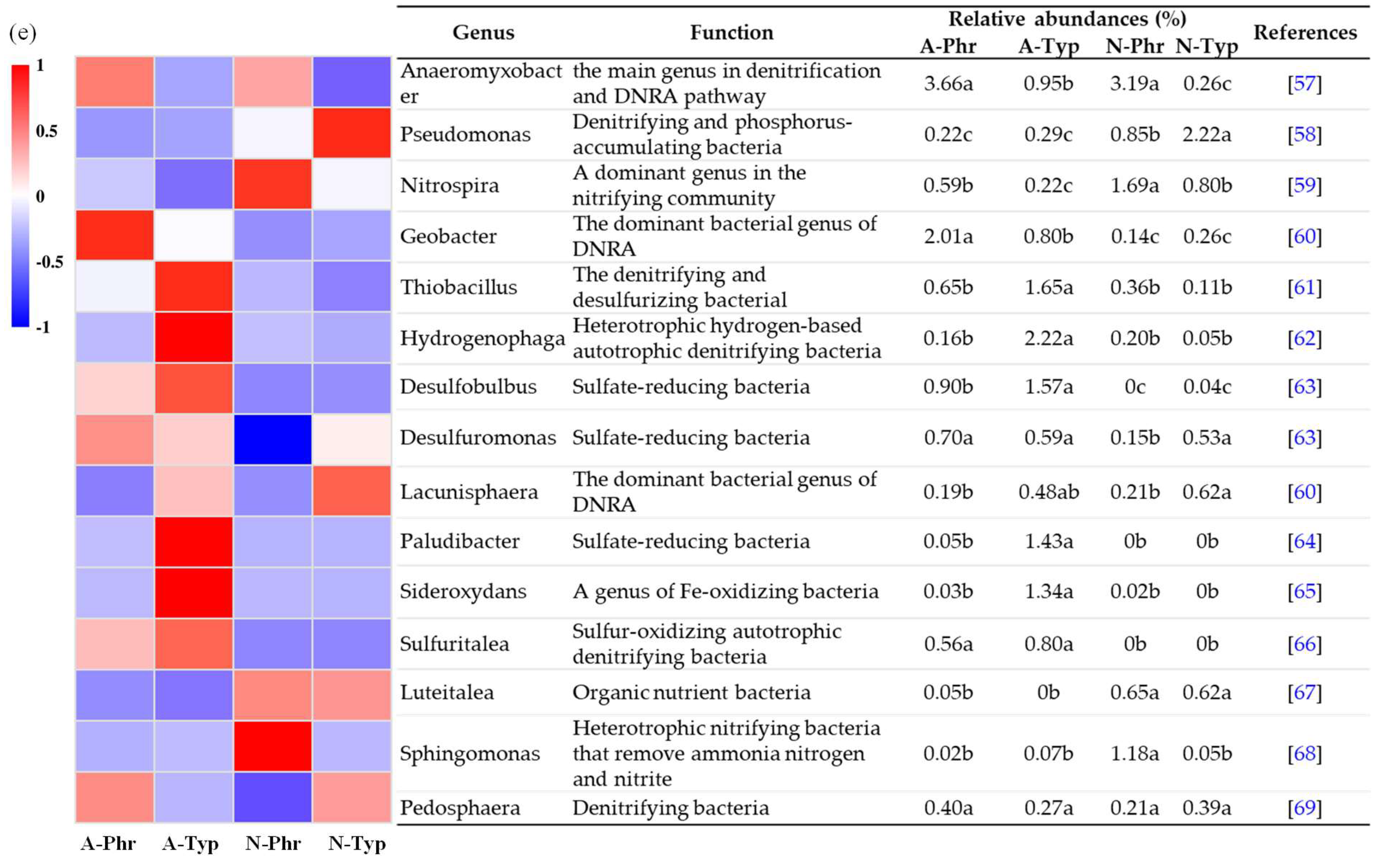
Figure 5.
Redundancy analysis (RDA) was used to identify the relationships among nitrogen conversion rates and microbial and environmental factors. (a) The alpha diversity index of bacteria and environmental factors; (b) the phylum level of bacteria and environmental factors; (c) the genus level of bacteria and environmental factors; (d) the nitrogen cycling pathways and environmental factors; (e) the nitrogen conversion rates and their environmental and microbial factors. DO is the dissolved oxygen in water; ORP is the oxidation‒reduction potential in water; EC is the electrical conductivity of the sediments; TOC is the total organic carbon in the sediments; DOC is the dissolved organic carbon in the sediments; TN is the total nitrogen in the sediments; NH4+-N is the ammonium nitrogen in the sediments; NO3--N is the nitrate nitrogen in the sediments; DOC/NO3--N is the ratio of dissolved organic carbon to nitrate nitrogen in the sediments; Sand is the sand content (the sediment particle size fraction is 2~0.02 mm) in the sediments.
Figure 5.
Redundancy analysis (RDA) was used to identify the relationships among nitrogen conversion rates and microbial and environmental factors. (a) The alpha diversity index of bacteria and environmental factors; (b) the phylum level of bacteria and environmental factors; (c) the genus level of bacteria and environmental factors; (d) the nitrogen cycling pathways and environmental factors; (e) the nitrogen conversion rates and their environmental and microbial factors. DO is the dissolved oxygen in water; ORP is the oxidation‒reduction potential in water; EC is the electrical conductivity of the sediments; TOC is the total organic carbon in the sediments; DOC is the dissolved organic carbon in the sediments; TN is the total nitrogen in the sediments; NH4+-N is the ammonium nitrogen in the sediments; NO3--N is the nitrate nitrogen in the sediments; DOC/NO3--N is the ratio of dissolved organic carbon to nitrate nitrogen in the sediments; Sand is the sand content (the sediment particle size fraction is 2~0.02 mm) in the sediments.
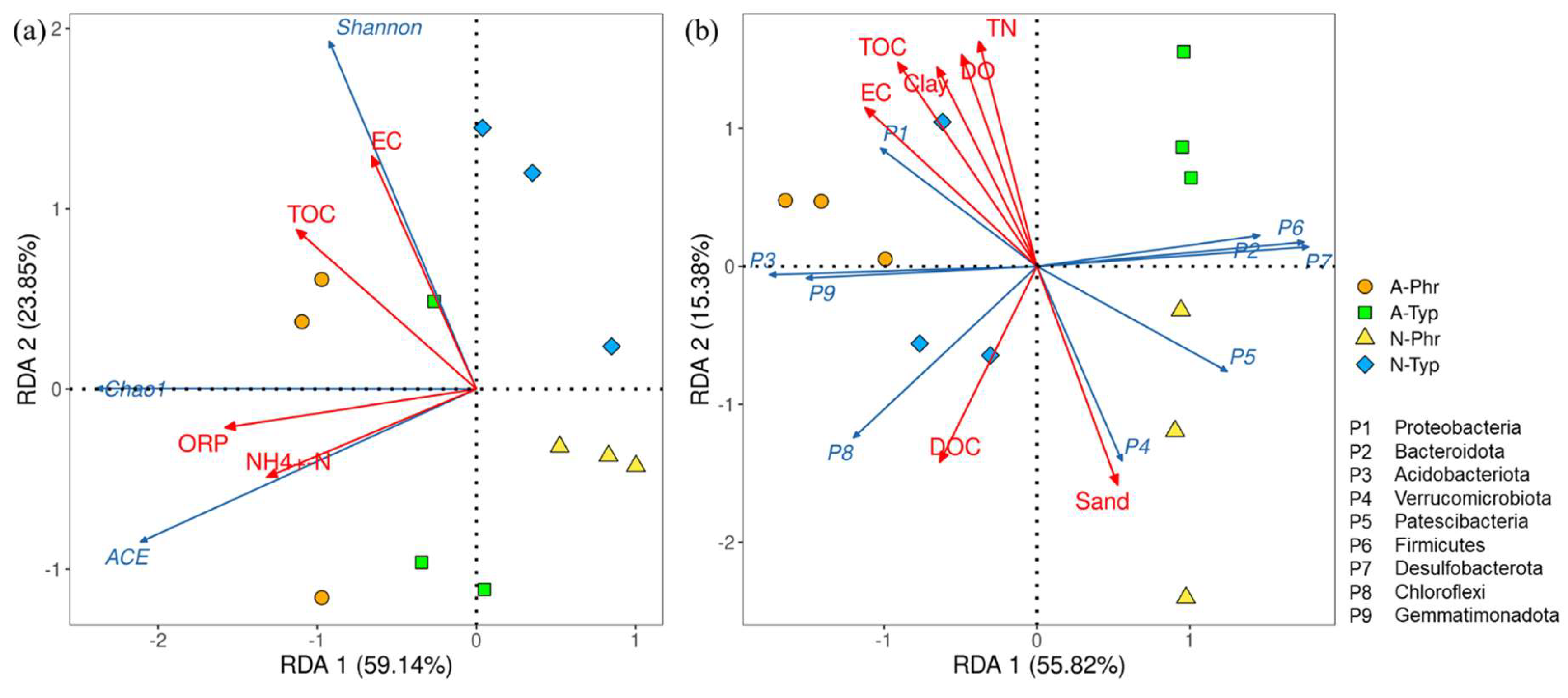
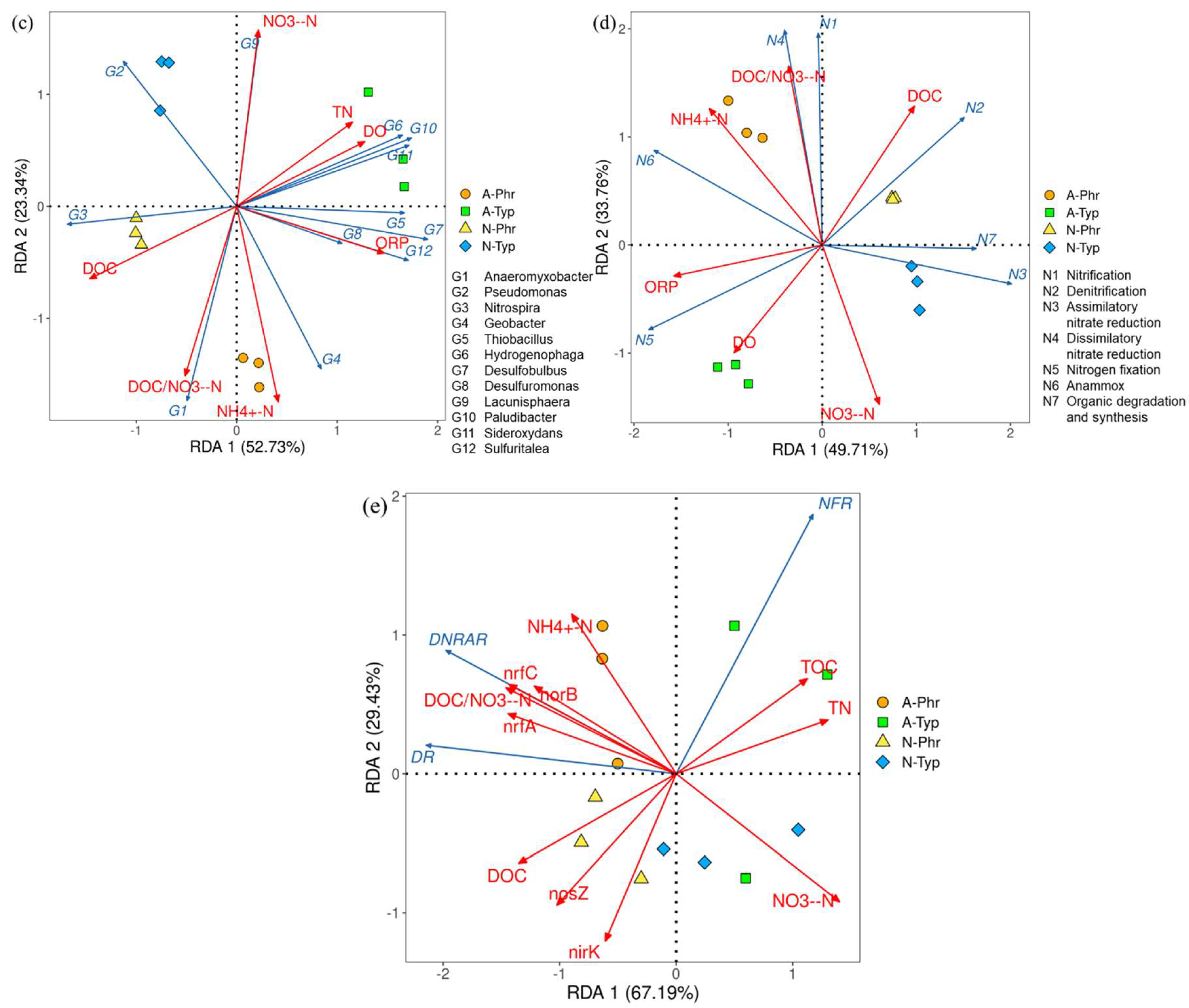
Figure 6.
The relative abundance of nitrogen cycling pathway genes (a); functional genes involved in denitrification, dissimilatory nitrate reduction and nitrogen fixation pathways (b); and functional genes involved in nitrification and anammox pathways (c) in different rhizosphere sediment samples.
Figure 6.
The relative abundance of nitrogen cycling pathway genes (a); functional genes involved in denitrification, dissimilatory nitrate reduction and nitrogen fixation pathways (b); and functional genes involved in nitrification and anammox pathways (c) in different rhizosphere sediment samples.

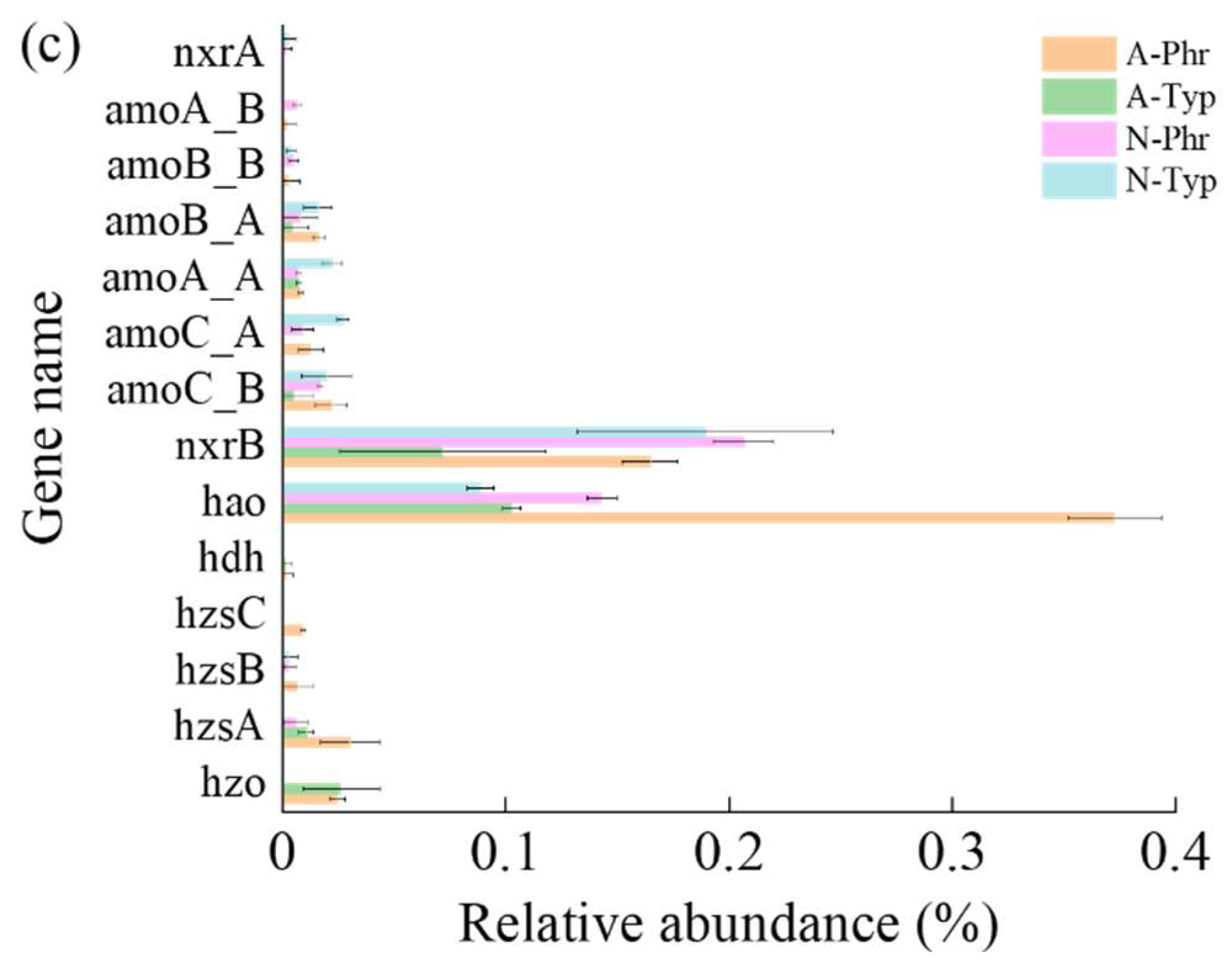
Figure 7.
Schematic diagram of nitrogen cycling at the gene level in different rhizosphere sediment samples from drainage ditches. DR is the nitrogen denitrification rate; DNRAR is the dissimilatory nitrate reduction to ammonium rate; NFR is the nitrogen fixation rate; DO is the dissolved oxygen in water; ORP is the oxidation‒reduction potential in water; TOC is the total organic carbon in the sediments; DOC is the dissolved organic carbon in the sediments; TN is the total nitrogen in the sediments; NH4+-N is the ammonium nitrogen in the sediments; NO3--N is the nitrate nitrogen in the sediments; and DOC/NO3--N is the ratio of dissolved organic carbon to nitrate nitrogen in the sediments. The numerical values within parentheses denote the different rates, with units of μmol N kg-1 h-1. The numerical values within parentheses after the genes of various functions denote their respective relative abundance. * indicates significant differences among the different rhizosphere sediment samples (p<0.05).
Figure 7.
Schematic diagram of nitrogen cycling at the gene level in different rhizosphere sediment samples from drainage ditches. DR is the nitrogen denitrification rate; DNRAR is the dissimilatory nitrate reduction to ammonium rate; NFR is the nitrogen fixation rate; DO is the dissolved oxygen in water; ORP is the oxidation‒reduction potential in water; TOC is the total organic carbon in the sediments; DOC is the dissolved organic carbon in the sediments; TN is the total nitrogen in the sediments; NH4+-N is the ammonium nitrogen in the sediments; NO3--N is the nitrate nitrogen in the sediments; and DOC/NO3--N is the ratio of dissolved organic carbon to nitrate nitrogen in the sediments. The numerical values within parentheses denote the different rates, with units of μmol N kg-1 h-1. The numerical values within parentheses after the genes of various functions denote their respective relative abundance. * indicates significant differences among the different rhizosphere sediment samples (p<0.05).
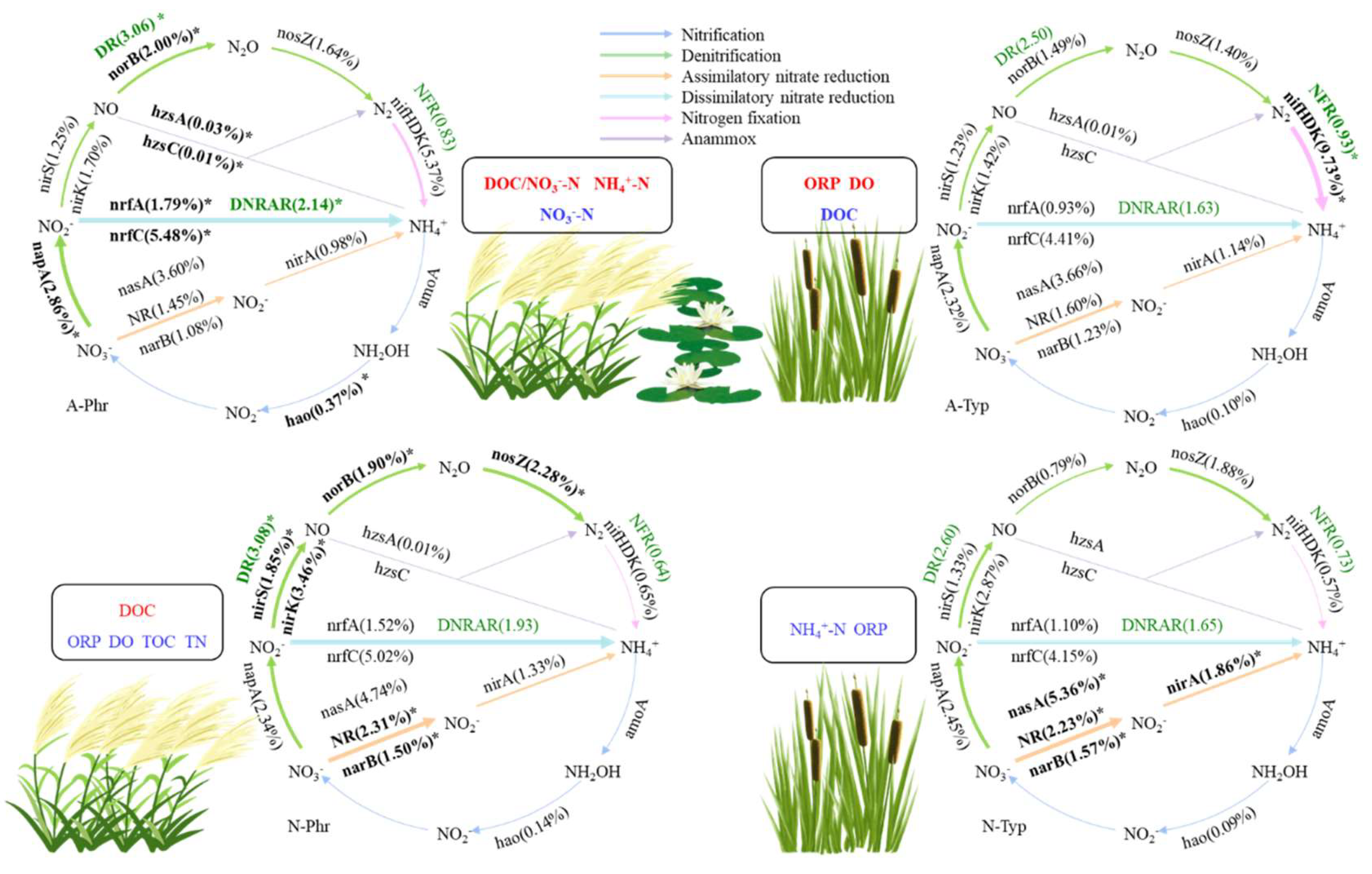

Table 1.
Composition of aquatic plant species in agricultural drainage ditches.
| Type of ditch | Principal species | Associated species | Relative coverage | Sample location | Abbreviation |
|---|---|---|---|---|---|
| Artificial cultivated ditch | Phragmites australis, Typha orientalis, Nymphaea tetragona | - | 57% | Phragmites australis- Rhizosphere | A-Phr |
| Typha orientalis - Rhizosphere | A-Typ | ||||
| Natural ditch1 | Phragmites australis | Scirpus triqueter, Echinochloa crusgali | 37.5% | Phragmites australis- Rhizosphere | N-Phr |
| Natural ditch2 | Typha orientalis | Phragmites australis, Scirpus triqueter | 30% | Typha orientalis - Rhizosphere | N-Typ |
Note: A-Phr, artificially cultivated ditch with P. australis rhizosphere sediment; A-Typ, artificially cultivated ditch with T. orientalis rhizosphere sediment; N-Phr, natural ditch with P. australis rhizosphere sediment; N-Typ, natural ditch with T. orientalis rhizosphere sediment.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



