
Submitted:
12 August 2024
Posted:
14 August 2024
You are already at the latest version
Abstract
Abstract: This study aimed to assess the agronomic performance and yield stability of elite white yam (Dioscorea rotundata) genotypes across diverse Nigerian environments. Twenty-five genotypes were evaluated at three locations in two consecutive growing seasons, 2022 and 2023. The genotypes were assessed for their agronomic performance, tuber yield and related traits using various analytical tools such as AMMI (Additive main effects and multiplicative interaction) and MTSI (Multi-trait stability index). The AMMI analysis revealed significant differences among the genotypes and across the environ-ments for all traits (p
Keywords:
yam
; Genetic variability
; Principal component analysis (PCA)
; Heritability
; Additive main effect and multiplicative interaction (AMMI)
; Multi-trait stability index (MTSI)
1. Introduction
Root and tuber crops (RTB) have been playing more and more important roles in food security and in meeting global dietary requirements [1]. Among the RTB’s, yam (Dioscorea spp.) holds unparalleled significance as both a staple food and a cash crop in the humid and sub-humid tropics of Africa. West Africa accounted for 96% of the world yam production figure of 88.3 Mt. Current estimates put the Nigeria production figure at 61.2 MT representing 72% of West African production [2]. Yam is a multi-species crop consisting of approximately 600 species. Within the diverse array of yam species, White Guinea yam (Dioscorea rotundata Poir) reigns supreme, flourishing extensively in West Africa's tropical regions. Supporting the livelihoods of over 400 million people, its cultivation extends from Cameroon to Sierra Leone, predominantly as subsistence farming heavily reliant on natural environmental inputs. Considering the projected global population of 9.6 billion by 2050, ensuring food security is paramount. Enhancing the productivity of vital food crops like yams while safeguarding the environment becomes imperative. In Nigeria, institutions like the International Institute of Tropical Agriculture (IITA) prioritize yam breeding endeavors aimed at developing novel varieties meeting end-users' preferences, encompassing both farmers and consumers. This focus revolves around critical traits such as consistent tuber quality, high yield, minimal flesh oxidation, reduced hardening, increased dry matter content, and resistance against yam mosaic virus and anthracnose disease [2,3,4].
Breeding is a potent tool for sustainable solutions to enhance yam cultivation and ensure food security. However, progress in trait improvement hinges on comprehensive genetic information and trait behavior across diverse environments. The exploration of yam quantitative traits has consistently illuminated the profound influence of genotype-by-environment interaction. Several investigations [5,6,7] have shed light on this crucial interplay, showcasing its pivotal role in determining yam genotype performance across diverse environmental contexts. This further underscored the necessity of acknowledging this interaction within yam breeding, advocating for multivariate analysis techniques to address its nuances effectively [8]. As breeding strategies take shape, understanding heritable variations and genetic correlations among economically significant traits becomes imperative. This knowledge forms the bedrock for selecting breeding plans, further illuminating the path toward desired progenies within breeding populations.
Genetic variability in quantitative attributes of White Guinea yam is essential as it reveals a vast reservoir of untapped potential for genetic enhancement. Our study aims to estimate variance components, heritability of agro-morphological traits, and scrutinize genotype-environment interaction patterns of some elite white Guinea yam genotypes across multiple environments in Nigeria.
2. Materials and Methods
2.1. Plant materials and Trial Establishment
Twenty-five genotypes from the International Institute of Tropical Agriculture (IITA) yam breeding program were used in this study. Details of the genotypes and testing locations are presented in supplementary Tables 1 and 2. The experiments were carried out at Ikenne located at 6°84′N and 3°69′E; Abuja located at 9°13′N and 7°23′E and Ibadan located at 7°49′N and 3°90′E in Nigeria. The field was ridged, and genotypes were planted in an alpha lattice design of three plants per plot at a spacing of 1mx1m in two replicates at two planting seasons, 2022 and 2023. The trial was staked at 2 meters high and proper weeding was done manually to ensure that the field was kept clean. Fertilizer was applied at the rate of 60 grams per row at 6 weeks after planting.
Agronomic traits were collected at the vegetative stage of the plant and at harvest using “Standard Operating Protocol for Yam Genotype Performance Evaluation Trial” [9]. All agronomic management practices were carried out equally and properly as per the recommendations earlier stated.
The area under the disease progression curve (AUDPC), is a valuable quantitative summary of disease severity for YMV and YAD over time, this was estimated using the trapezoidal method [10] This method represents the time variable and calculates the average disease severity between each pair of adjacent time points:
where n is the number of assessments made, yi is the anthracnose or virus severity score on date i, and t is the time in months between assessments yi and yi + 1.
The rate of pathogen reaction to yam mosaic virus (YMV) and yam anthracnose disease (YAD) (severity scores) were recorded monthly from two to six months after planting following the scale as shown in (Figure 1) below.
%DMC = (dry tuber weight/fresh tuber weight) x 100
TTY = (total tuber weight harvested/effective plot) x 100
2.2. Statistical Analysis
All statistical analyses were performed using various packages in R Studio [11]. The function “find outliers” from the ‘metan’ package was used to ensure there were no outliers and that the data followed a normal distribution. Least square means (LS means) for each trait across environments were calculated using the ‘emmeans’ package version 1.8.7. We fitted the following linear mixed-effect model using the ‘statgen-STA’ package version 1.0.12 [12] to estimate variance components and obtain the best linear unbiased estimations
where y is the response vector of a trait for a given location, is the vector of fixed effects with the design matrix X (relating observations to fixed effects, which include grand mean, replication number), u is the vector of random genetic effects with the design matrix Z (relating trait values to genotype, environment, genotype × environment interaction (GEI)), and e is the residual. A linear model was used to conduct the analysis of variance (ANOVA) using the ‘metan’ package in R, following the alpha lattice design model [13]
where Yijkl represents the response variable, represents the overall mean, represents the effect of the ith genotype on the response variable, represents the effect of jth location on the response variable, (GE)ij represents the interaction between the ith genotype and jth location, (ED)jk represents the interaction effect between jth location, kth replicate within each location and (ED)jkl represents the interaction effect between the jth location, kth replicates and lth block within the replicates and eijkl represents the experimental error.
The principal component analysis (PCA) was conducted using ‘Factoextra package’ [14] to generate Eigen values, percentage of the variation accumulated by the PCA and the load coefficient values between the original characters and respective PCA. Pearson correlation was performed using the function “cor” in R studio. Genetic parameters such as broad-sense heritability and phenotypic coefficients were estimated using the function “get model data” in the metan package in R. Broad-sense heritability across all environments for all traits was calculated as follows:
Phenotypic coefficient of variance
Genotypic coefficient of variance
σ2g is the genotypic variance, σ2e is the residual variance, and σ2 i is the genotype-by-environment interaction variance. Heritability was categorized as low (<30%), moderate (31–60%), and high (>60%) according to Robinson et al. [15].
Genotype by environment interaction (GEI) analysis was performed using the Additive Main Effects and Multiplicative Interaction (AMMI) model [16]
where Yij is the observed mean yield, λk is the singular value for the k-th interaction principal component axis (IPCA); aik is the i-th element of the k-th eigenvector; tjk is the jth element of the kth eigenvector. A residual ρij remains, p IPCA are used, where p≤ min (g−1; e−1). This approach depends on analysis of variance (ANOVA) for estimating genotype and environment main effect, principal component analysis (PCA) for decomposing GEI structure into Interactive Principal Component Axes (IPCAs) and biplot for graphical presentations. AMMI gives a suitable approach in separating genotypic effect from genotype by environment effect with cultivar ranking in mega-environment [17] while GGE is suitable for grouping sites and cultivars without cultivar rank change [18]. Superior cultivars and test environments were selected.
The multi-trait stability index (MTSI) was determined following the formula used by [19] to select the stable genotypes for the studied traits. The genotypes with lowest MTSI were considered as superior genotypes [20].
MTSIi=multi-trait stability index for the i-th genotype, =j-th score for the i-th genotype,
=j-th score of ideal genotypes.
3. Results
3.1. Variation in Least Square Means of Quantitative Traits Across Environments for 25 Yam Genotypes
Figure 2 displayed the variation of quantitative traits accessed on 25 yam genotypes across different environments with each plot representing the distribution of LS means for a specific trait in multiple environments. The wider boxes indicate more variation in LS means among genotypes within that environment and the narrower boxes indicate less variation and more consistency among the genotypes. The points outside the whiskers are LS means that are significantly different from others. The median in each environment varies with respect to their traits which implies that the performance of the genotypes for each trait is influenced by the environment. ATW, PLNV, Oxi30, and Oxi180 have outliers, which suggests there are genotypes with high variability among others. Traits such as DMC, TTN, Oxi30, AUDPCYMV, and TTY have their mean higher than the median, which indicates few genotypes with higher performance. Oxi30 has the smallest boxplot which means that the variability is less compared to DMC which has the largest variability.
3.2. Genotypic Coefficients, Phenotypic Coefficients, and Broad-Sense Heritability
This study showed that genotypic coefficient of variation is higher than the phenotypic coefficient of variation except for plant vigour and this means that genetic factors contributed more to the variability observed in the trait compared to environmental factors (Table 3). Genotypic coefficient of variation ranged from 0 for plant vigor to 717.78 for oxidation intensity at 30 minutes. On the other hand, phenotypic coefficient of variation (CVp) values ranged from 23.2 to 108.2 in all the traits. The genotypic variance was also observed to be higher than the phenotypic variance for all the agronomic traits in this study. Low broad sense heritability (H2) (0-30%) was observed for plant vigor and AUDPCYMV while moderate broad sense heritability was observed for all other traits (Table 3).
3.3. Traits importance and contribution
The principal component analysis displayed the contribution of the quantitative traits in the white yam genotypes (Table 4). The first four principal component of the quantitative traits accounted for 73.55% of the total variation. The first principal component (PC1) had total tuber number (TTN), average tuber weight (ATW) and total tuber yield (TTY) which contributed most to the total variation. PC2 had area under disease progression curve for yam mosaic virus (AUDPCYMV), total tuber number (TTN), total tuber yield (TTY), oxidation intensity at 30 and 180 minutes contributed most to the total variation. PC3 had plant vigor and dry matter content which contributed most to the total variation. PC4 had oxidation intensity at 180 minutes, dry matter content and area under disease progression curve yam anthracnose disease which contributed most to the total variation.
3.4. Phenotypic correlation coefficient between the quantitative traits measured.
Pearson correlation coefficient among the agronomic traits for 25 white yam genotypes across six environments displayed (Figure 3) showed the relationship between the agronomic traits evaluated. There was a significant (P<0.001) positive correlation between total tuber number (TTN) and total tuber yield (TTY) (0.99), average tuber weight (ATW) and TTY (0.44), ATW and total tuber number (TTN), AUDPCYMV and AUDPCYAD respectively. There was a negative correlation between AUDPCYAD with oxidation intensity at 30 minutes (-0.25), AUDPCYMV with oxidation intensity at 180 minutes (-0.35) and Oxi30 (-0.49). There was a significant (P<0.05) negative correlation between area under disease progression curve yam mosaic virus (AUDPCYMV) and total tuber yield (TTY) (-0.12), and a positive correlation between plant vigor (PLNV) and AUDPCYMV (0.14). There was a negative correlation (p<0.01) between PLNV and oxidation intensity at 180 minutes.
3.5. Additive main effect and multiplicative interaction on agronomic traits.
The mean squares of AMMI analysis of variance explained for 25 genotypes evaluated agronomic traits in three locations is presented in Table 5. The AMMI analysis revealed significant variation in the main effects of the environment (P<0.001) for all traits except for dry matter content and for average tuber weight (ATW) at (P<0.01). The effect of genotype is significant at (P < 0.05) for all observed traits except AUDPCYMV, PLNV and Oxi30. Genotype X environment interaction (GEI) was decomposed into two principal components, and PC1 mean squares were highly significant (P < 0.001) for all traits except total tuber number (TTN), total tuber yield (TTY) and oxidation intensity at 30 minutes (Oxi30) at p<0.01. PC2 mean squares were significant (P < 0.001) for AUDPCYMV, AUDPCYAD and Oxi180. Mean squares were highly significant (P < 0.01) for TTN, ATW, PLNV, while mean squares were significant (P<0.05) for TTY, Oxi30 and DMC.
3.6. Additive main effects and multiplicative interaction (AMMI) 1 and 2 biplot
Figure 4 shows the AMMI1 biplots where genotypes and environments are depicted as points on a plane with the first PC on the y-axis. The horizontal line within the plot shows the interaction effect and the vertical axis represents main effect. The vertical axis showed interaction between genotypes and the environments while the horizontal showed the main effect of the genotype and environment. The superior genotypes for total tuber yield are the genotypes on the right two quadrants (top and bottom right) of the biplot. The environments were distributed from the low yielding environments on the left quadrants to the high yielding on the right quadrants. IK23, AB23, IK22, and IB23 were identified as high yielding environments (figure 4a). The superior genotypes identified for dry matter were the genotypes on the right two quadrants (top and bottom right) of the biplot. The environments were distributed from the low dry matter content environments on the left quadrants to the high dry matter content on the right quadrants. IK23, AB23, AB22, and IB23 were identified as high dry matter content environments (figure 4b). The genotypes identified as superior for total tuber number were genotypes on the right two quadrants (top and bottom right) of the biplot. The environments were distributed from the low tuber number on the left quadrants to the high tuber number on the right quadrants with IK23, AB23, IK22, and IB23 as high total tuber number environments (figure 4c).
AMMI2 biplot of white Guinea yam genotypes are illustrated in Figure 5. The AMMI 2 revealed the importance of PCA2 scores along the first PCA in explaining the complexity of GEI involving significant multi-environments and identifying the adaptation of genotypes. The PC2 values were 25.9%,12.7% and 27.5% for total tuber yield, dry matter content and total tuber number, respectively. Genotypes and environments near the origin (centre of the plot) have average performance and stability because their performance is less influenced by environmental changes. In contrast, those far from the origin and close to each other indicated strong specific interaction. For total tuber yield, TDr1741021 performed well in AB 23, TDr1719029 and TDr1719020 performed well in IK23, Meccakusa performed well for AB22 and TDr1739046 performed well in IB23 environments. For dry matter content, TDr1738048 performed well in IB23, TDr1739023 and TDr1739058 performed well in IK23, and TDr1741073 performed well in AB22 and A23 environments. For the total tuber number, Meccakusa performs well in AB22, TDr1741021 performs well in AB23, TDr1719029 performed well in IK23 environments.
Figure 6a shows the mean performance and stability of 25 genotypes for total tuber yield across six environments. The arrowed average tester coordination (ATC) lines from the biplot shows the lowest vs. highest and stable vs. unstable genotypes in the experiment [21]. A deviation from the ATC lines indicates stability; therefore, TDr1739023 TDr1719059 and TDr1716069 were among the most stable genotypes. TDr1739046 and TDr1719029 were among the least stable genotypes for total tuber yield because they strongly deviated from the ATC lines. Genotypes close to the arrow are the genotypes with the highest mean performance for total tuber yield. Figure 6b displays the mean performance and stability of 25 genotypes for total tuber number across six environments. A deviation from the ATC lines indicates stability; therefore, TDr1716069, TDr1738003, TDr1719055, TDr1739023 and TDr1719059 were among the most stable genotypes. Dr1739046 and TDr1719029 were among the least stable genotypes for total tuber number because they strongly deviated from the ATC lines. Genotypes close to the arrow are genotypes with the highest mean performance for total tuber number. Figure 6c showes the mean performance and stability of 25 genotypes for dry matter content across six environments where deviation from the ATC lines indicates stability; therefore, TDr1716040 and TDr1716069 were among the most stable genotypes. Genotype TDr1738048 was the least stable for dry matter content because it strongly deviated from the ATC lines. Genotypes close to the arrow are the genotypes with the highest mean performance for dry matter content.
3.7. Multi-trait selection for agronomic traits
Selection of the traits under study was performed through a multi-trait stability index (MTSI). The selection differential was negative for all the traits. The selection differential for WAASB (Weighted Average of Absolute Scores) index was 1.09%, 9.08%, 19.2%, 19.3%, 22.6%, 24.3%, 26.2%,26.4% and 39% for DMC, AUDPCYAD, TTY, AUDPCYMV, TTN, ATW, PLNV, Oxi180 and Oxi30 respectively (Table 6). The four selected genotypes with the lowest MTSI values (Supplementary Table 3) were TDr1739049, TDr1739058 TDr1741044 and TDr1739023 (figure 7a). Figure 7b provides a visual representation of the strengths and weaknesses of the selected genotypes based on the contribution of each factor in the MTSI index. The lower the share justified by a factor (close to the outer edge), the closer the attributes within that factor are to the stable ideal state. Genotypes TDr1741044, TDr1739049, and TDr1739058, which had the lowest value in the first factor, were close to the ideal genotype for the total tuber number and total tuber yield. Genotypes TDr1741044 and TDr1739049 had the lowest share of the second factor, indicating their proximity to the ideal genotype in terms of AUDPCYMV and AUDPCYAD. Genotypes TDr1739023, TDr1739049 and TDr1739058 had the lowest share of the third factor, indicating their proximity to the ideal genotype in terms of Oxi30 and Oxi180. Genotypes TDr1739058 and TDr1739023 had the lowest share of the fourth factor, indicating their proximity to the ideal genotype in terms of PLNV and ATW.
* (Xo) = original mean, (Xs) = mean of the selected accessions, SD = selection differential, %=percentage.
4. Discussion
This study assessed the genetic potential of twenty-five genotypes of white Guinea yam (Dioscorea rotundata) for its agronomic attributes at three different locations. The analysis of agronomic traits revealed a significant genotype-by-environment interaction. The width of the boxes in the boxplots reflects the variability in the least square (LS) means among genotypes within each environment. The outliers denotes LS means that are significantly different from the majority, highlighting genotypes with extreme performance values. The varying median values across environments for each trait indicated that genotypic performance is influenced by environmental conditions. These findings underscore the importance of considering environmental influence when assessing genotypic performance for quantitative traits. Traits with high variability may require environment-specific strategies for selection and breeding, whereas traits with low variability suggest more stable performance across different conditions. Similar findings were also reported in yam and rice [22,23]
Genetic variance serves as a cornerstone in the process of selecting superior genotypes and enhancing desirable traits within a population. In our investigation, we discerned notable genotype effects across various traits, save for plant vigor. This observation underscores a promising avenue for harnessing genetic diversity to drive improvements through hybridization and selective breeding strategies. Our findings are in line with the findings of Cynthia et al. [24] in their examination of cassava, where they also identified similar trends in certain traits. This parallel further corroborates the potential efficacy of targeted breeding efforts to amplify genetic gains and foster trait enhancement across diverse agricultural environments.
The low broad-sense heritability (0-30%) for plant vigor and yam mosaic virus (AUDPCYMV) observed in this study suggests that a relatively small proportion of the phenotypic variation in these traits can be attributed to genetic differences among the genotypes. This may require more intensive selection pressure or the incorporation of additional genetic or environmental factors to achieve improvements. On the other hand, the moderate broad-sense heritability observed for all other traits indicated that genetic factors played a more substantial role in determining the variation in these traits. This implies that traits such as total tuber number, tuber yield and other agronomic characteristics are more strongly influenced by genetic differences among the genotypes. However, these findings appear to deviate from the reported results of high broad-sense heritability for tuber yield and yam mosaic disease [25], as well as from the moderate to high broad-sense heritability in various measured traits across wild and cultivated yam [26,27], further adding to the complexity of interpreting genetic influences on yam traits.
The principal component analysis outcomes offer valuable insights into the underlying factors driving variation in quantitative traits among the genotypes. These revelations align with similar studies that uncovered distinct associations between white yam genotypes and key traits such as fresh tuber yields, tuber dry matter content, and susceptibility to yam mosaic virus [27]. Such genotype-trait associations unveiled by this study provide a wealth of information that holds great promise for breeders, particularly in the meticulous selection of parental candidates for targeted breeding initiatives aimed at enhancing specific traits [28]. Moreover, several authors have highlighted the significance of phenotypic traits in unraveling the diversity and differentiation in yam [29,30,31]. These collective insights not only enrich our understanding of yam genetics and agronomy but also pave the way for more informed and effective breeding strategies tailored to meet the evolving demands of yam cultivation and utilization.
The correlations among the studied traits showed the strength of their linear associations. This implies that certain traits can serve as reliable proxy selection parameters for evaluating different genotypes. This correlation analysis offers valuable insights into the interplay among diverse traits and has the potential to significantly enhance the precision of selecting and breeding yam genotypes tailored to exhibit specific attributes.
The AMMI1 and AMMI2 biplots provided a comprehensive view of the genotype-environment interactions for tuber yield, dry matter content, and tuber number. The Additive Main Effects and Multiplicative Interaction (AMMI) model is a crucial tool for evaluating the significance of genotype-by-environment (G × E) interactions in agronomic traits across multi-environment trials. This model allows for a detailed understanding of the relationships between genotypes and the environments in which they are tested. AMMI model 1 biplot is widely used for identifying high potential yield and stability [32]. In the AMMI1 biplot, when mean environments and genotypes are placed on lines parallel to the ordinate, it indicates that they have similar performance [33,34]. Additionally, genotypes positioned on the right side of the center of the biplot exhibit higher yields compared to those on the left side. Our findings align with this, as genotypes located in the right-hand quadrants for tuber yield, dry matter content, and tuber number reflect superior performance.
The IPCA1 and IPCA2 scores are essential for understanding the function of genotype-by-environment interaction and the adaptability of genotypes in the test environments. The first two PCA interactions for tuber yield, dry matter content, and tuber number accounted for the total variation of the G + G × E interaction for the traits considered. Among the genotypes, TDr1719029 and TDr1719020 stand out for total tuber yield, TDr1739023, TDr1739058, and TDr1719055 for dry matter content, and TDr1719029 and TDr1719020 for tuber number, indicating their similar response patterns. In the test years, Ibadan was relatively close to the biplot origin, indicating it has a less interactive location and could be a suitable location for selecting genotypes with average adaptation.
The MTSI index selected four ideal genotypes TDr1739049, TDr1739058, TDr1741044 and TDr1739023 as the best performing genotypes among the twenty-five genotypes studied. Genotypes with lower values are assumed to have better performance and stability based on the traits assessed. The strength and weakness view revealed the four selected genotypes performance towards their traits. FA1 showed the contributing factor of TTN and TTY to the selection of the genotypes, while AUDPCYMV and AUDPCYAD were found to be more contributing to the selected genotypes FA2. FA1 and FA4 are the group contributing less to the genotype selection, while FA2 and FA3 are the strength of the genotypes. Multi-trait stability index had been successfully used in previous studies to evaluate yield and other agronomic traits in various genotypes and has been applied to evaluate several crops to identified superior genotypes in terms of yield, yield stability, and other agronomic traits [35,36,37].
5. Conclusions
This study delved into the genetic potential of twenty-five genotypes of white Guinea yam (Dioscorea rotundata) across six environments, shedding light on their agronomic attributes. The findings revealed significant genotype and environment interactions for these traits, with genetic factors playing a predominant role in trait variability. Notably, each genotype exhibited unique agronomic characteristics, offering promising avenues for targeted selection and breeding programs to enhance specific traits of interest. The application of the Multi-Trait Stability Index (MTSI) corroborated previous studies, demonstrating its efficacy in evaluating yield and other agronomic traits across different genotypes. These findings contributed more to the broader understanding of yam genetics and agronomy, laying the groundwork for informed breeding strategies tailored to meet the evolving demands of yam cultivation and utilization.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: list of genotypes; Table S2: list of environments; Table S3: MTSI selection list.
Author Contributions
Design and study conceptualization, O.A.A, A.A. and P.A.; study methodology and manuscript drafting, O.A.A.; implementation, O.A.A and A.A.; formal data curation and analysis, O.A.A.; manuscript reviewing and editing, A.A., P.A, A.S.G, K.D.T.; supervision, A.A, A.S.G and K.D.T; coordination and fund acquisition, A.A and P.A
Funding
This research was supported by the Bill & Melinda Gates Foundation AfricaYam project grant number: (INV-003446).
Data Availability Statement
The data that supports the findings of this study are available in the supplementary material of this article.
Acknowledgments
We thank the International Institute of Tropical Agriculture (IITA) for their support of the first author's PhD research in crop production at the Federal University of Technology Minna, Nigeria. Special gratitude goes out to the entire Yam Breeding Team Unit of the IITA Abuja and Ibadan. We are grateful to our reviewers for their insightful comments and suggestions that improved the manuscript.
Conflicts of Interest
The authors declare no conflict of interest in this work.
References
- Q. Liu, J. Q. Liu, J. Liu, P. Zhang, S. He, Root and Tuber Crops, Editor(s): Neal K. 2014; -61. [Google Scholar] [CrossRef]
- Nayer, N.M. The Contribution of Tropical Tuber Crops Towards Food Security, J. Rt crp. 2015. [Google Scholar]
- FAOSTAT Food and Agriculture Organization. Agricultural Production Statistics. 2022. Available at https://www.fao.org/faostat/en/#data/QCL.
- Darkwa, K.; Are, P.; Olasanmi, B.; Iseki, K.; Matsumoto, R.; Powell, A.; Bauchet, G.; De Koeyer, D. ; Muranaka, S; Adebola, P. Comparative assessment of genetic diversity matrices and clustering methods in white uinea yam (Dioscorea rotundata) based on morphological and molecular markers. Sci rep, 2020, 10, 13191. [Google Scholar]
- Agre, P.; Norman, P.E.; Asiedu, R.; Asfaw, A. Identification of Quantitative Trait Nucleotides and Candidate Genes for Tuber Yield and Mosaic Virus Tolerance in an Elite Population of White Guinea Yam (Dioscorea rotundata) Using Genome-Wide Association Scan. BMC Plant Biology. 2021, 21, 552. [Google Scholar] [CrossRef] [PubMed]
- Asfaw, A.; Aderonmu, D.S.; Darkwa, K.; De Koeyer, D.; Agre, P.; Abe, A.; Olasanmi, B.; Adebola, P.; Asiedu, R. Genetic parameters, prediction, and selection in a white Guinea yam early-generation breeding population using pedigree information. Crop Sci., 2021, 61, 1038–1051. [Google Scholar] [CrossRef] [PubMed]
- Adjei, E.A; Esuma, W.; Alicai, T.; Bhattacharjee, R.; Dramadri, I.O.; Agaba, R.; Emmanuel, B.C. ; Phenotypic Diversity within Ugandan Yam (Dioscorea species) Germplasm Collection. Int. J of Agron. 2022. [CrossRef]
- Norman, P.E.; Tongoona, P.B.; Danquah, A.; Danquah, E.Y.; Agre, P.A.; Agbona, A.; Asiedu, R.; Asfaw, A. Genetic parameter estimation and selection in advanced breeding population of white Guinea yam. J. Crop Improv. 2021, 35, 790–815. [Google Scholar] [CrossRef]
- Akinyosoye, S.T.; Adetumbi, J.A.; Amusa, O.D.; Agbeleye, A.; Anjorin, F.; Olowolafe, M.O.; Omodele, T. ; Bivariate analysis of the genetic variability among some accessions of African Yam Bean (Sphenostylis stenocarpa (Hochst ex A. Rich) Harms) 2017, Vol. 109, No. 3. [CrossRef]
- Norman, P.; Tongoona, P.; Danquah, A.; Danquah, E.; Agre, P.; Agbona, A.; Asiedu, R.; Asfaw, A. Genetic Analysis of Agronomic and Quality Traits from Multi-Location white Yam Trials using Mixed Model with Genomic Relationship Matrix. G. J. Bot. Sci. 2022. [CrossRef]
- Asfaw, A. Standard operating protocol for yam variety performance evaluation trial. Ibadan, Nigeria: IITA. 2016. [Google Scholar]
- Campbell, C.I.; Madden, L.V. Introduction to plant disease epidemiology. 1990. [Google Scholar]
- R Core Team. An Introduction to R: A Programming Environment for Data Analysis and Graphics Version 3.6.1; R Foundation for Statistical Computing: Vienna, Austria. 2022.
- van Rossum, BJ.; van Eeuwijk, F. ; Boer M, statgenSTA https://CRAN.Rproject.org/package=statgenSTA (2022).
- Patterson, H.D.; Williams, E.R. A new class of resolvable incomplete block design. Biometrica, 1976, 63, 83–92. [Google Scholar] [CrossRef]
- Resende, M.D.V. Genética Biométrica e Estatística no Melhoramento de Plantas Perenes; Embrapa: Brasília, DF, Brazil. 2002. [Google Scholar]
- Robinson, H. F.; Comstock, R. E.; Harvey, P. H. “Estimates of heritability and the degree of dominance in corn. ” Agr. J., 1955, 41, 353–359. [Google Scholar] [CrossRef]
- Gauch, H.G. Statistical analysis of regional yield trials: AMMI analysis of factorial designs. Elsevier, Amsterdam. 1992. [Google Scholar]
- Hagos, H.G.; Abay, F. Ammi and gge biplot analysis of bread wheat genotypes in the northern part of Ethiopia. Journal of plant breeding. Genetics. 2013, 1, 12–18. [Google Scholar]
- Yan, W.; Hunt, LA. Interpretation of genotype × environment interaction for winter wheat yield in Ontario. Crp. Sci. J. 2001, 41: 19–25.
- Olivoto, T. : A.D.C. L'ucio.; J.A.G. da silva.; B.G. Sari.; M.I. Diel. Mean performance and stability in multi-environment trials II: Selection based on multiple traits. Agron. J. 2019, 111:2961-2969. [Google Scholar] [CrossRef]
- Sellami, M.H.; Pulvento, C.; Lavini, A. Selection of suitable genotypes of lentil (Lens culinaris medik.) under rainfed conditions in south Italy using multi-trait stability index (MTSI) Agron. J., 2021, 11 10. 3390. [Google Scholar]
- Frashadfar, E.; Safari, H.; Jamshidi, B. GGE biplot analysis of adaptation in wheat substitution lines. International Journal of Agriculture and crop science 2012, 4, 877–881. [Google Scholar]
- Chukwu, V. A.; Osuafor, O. O.; Ukwuaba, I.; Enyigwe, J.; Chinenye, C. P. "Farmers’ Adaptation Strategies to the Effect of Climate Variation on Yam Production in Ebonyi State, Nigeria." J. Agric. Crp, 2023; vol. 9, pp. 240–249.
- Oumarou, S.; Massaoudou, H.; Mahamadou, S.; Baboucarr, M.; Eric, D.; Kwadwo, O. Environmental Impact on Early Generations Selection and Its Implications on Breeding Strategies in Rice. International J. Adv Res Bot. 3. [CrossRef]
- Cynthia, I.A. , Siraj, I.K., Bussie, M., Saviour, J.E., Isaac, A., Samuel, K.O. Ismail, Y.R. Exploring genetic variability, heritability, and trait correlations in gari and eba quality from diverse cassava varieties in Nigeria, J. F. Sci. Agri, 2023, 104(8):4860-4871. [CrossRef]
- Bandana, P.; Arup, K.M.; Sangram, K.M.; Sangram, K.L.; Debabrata, P. Genetic variability and inter species relationship between wild and cultivated yams (Dioscorea spp. 2019. [Google Scholar]
- Norman, P.E.; Tongoona, P.B.; Danquah, A.; Danquah, E.Y.; Agre, P.A.; Agbona, A.; Asiedu, R.; Asfaw, A. Genetic parameter estimation and selection in advanced breeding population of white Guinea yam. J. Crop Improv. 2021, 35, 790–815. [Google Scholar] [CrossRef]
- Yan, W.; Frégeau-Reid, J. Breeding Line Selection Based on Multiple Traits. Crop Sci. [CrossRef]
- Norman, PE.; Tongoona, P.; Shanahan, P.E. Diversity of the morphological traits of yam (Dioscorea spp.) genotypes from Sierra Leone. J. Ap. Biosci, 3058. [Google Scholar]
- Anokye, M.; Tetteh, JP.; Otoo, E. Morphological characterization of some water yam (Dioscorea alata L.) germplasm in Ghana. J.Agri. Sci.
- Agre, P. , Asibe, F., Darkwa, K., Darkwa K., Edemodu A., Bauchet G., Asiedu R., Adebola P. and Asfaw A. Phenotypic and molecular assessment of genetic structure and diversity in a panel of winged yam (Dioscorea alata) clones and cultivars. Scientific Reports – Nature Research 9, 18221 (2019). [CrossRef]
- Kılıç, H. Additive main effects and multiplicative interactions (AMMI) analysis of grain yield in barley genotypes across environments. J. Agric. Sci. 2014, 20, 337–344. [Google Scholar] [CrossRef]
- Vincent, I. E.; Grace, O.O.; Timothy, O.O.; Idowu, A.O. AMMI and GGE biplot analyses of Bambara groundnut [Vigna subterranea (L.) Verdc.] for agronomic performances under three environmental conditions. 2023. [Google Scholar]
- Sharifi, P.; Abbasian, A.; Mohaddesi, A. Evaluation the mean performance and stability of rice genotypes by combining features of AMMI and BLUP techniques and selection based on multiple traits. Plant Genet. [CrossRef]
- Rajabi, A.; Ahmadi, M.; Bazrafshan, M.; Hassani, M.; Saremirad, A. Evaluation of resistance and determination of stability of different sugar beet (Beta vulgaris L.) genotypes in rhizomania-infected conditions. F. Sci. J. Nutr.2022, 11:1403–1414. [CrossRef]
- Mahdi, H.; Seyed, B.M.; Ali, S.; Dariush, T. Genotype by environment and genotype by yield*trait interactions in sugar beet: analyzing yield stability and determining key traits association. 2024. [Google Scholar]
Figure 1.
Visual scale for yam mosaic virus disease scoring.

Figure 2.
Boxplots of quantitative traits accessed on 25 yam genotypes *. *PLNV: plant vigour; AUDPCYAD: area under disease progression curve yam anthracnose disease; AUDPCYMV: area under disease progression curve yam mosaic virus, TTW: total tuber weight per plant; ATW: average tuber weight; TTY: total tuber weight; OXi30 and OXi180: intensity of tuber oxidation at 30 and 180 minutes and DMC: dry matter content.
Figure 2.
Boxplots of quantitative traits accessed on 25 yam genotypes *. *PLNV: plant vigour; AUDPCYAD: area under disease progression curve yam anthracnose disease; AUDPCYMV: area under disease progression curve yam mosaic virus, TTW: total tuber weight per plant; ATW: average tuber weight; TTY: total tuber weight; OXi30 and OXi180: intensity of tuber oxidation at 30 and 180 minutes and DMC: dry matter content.
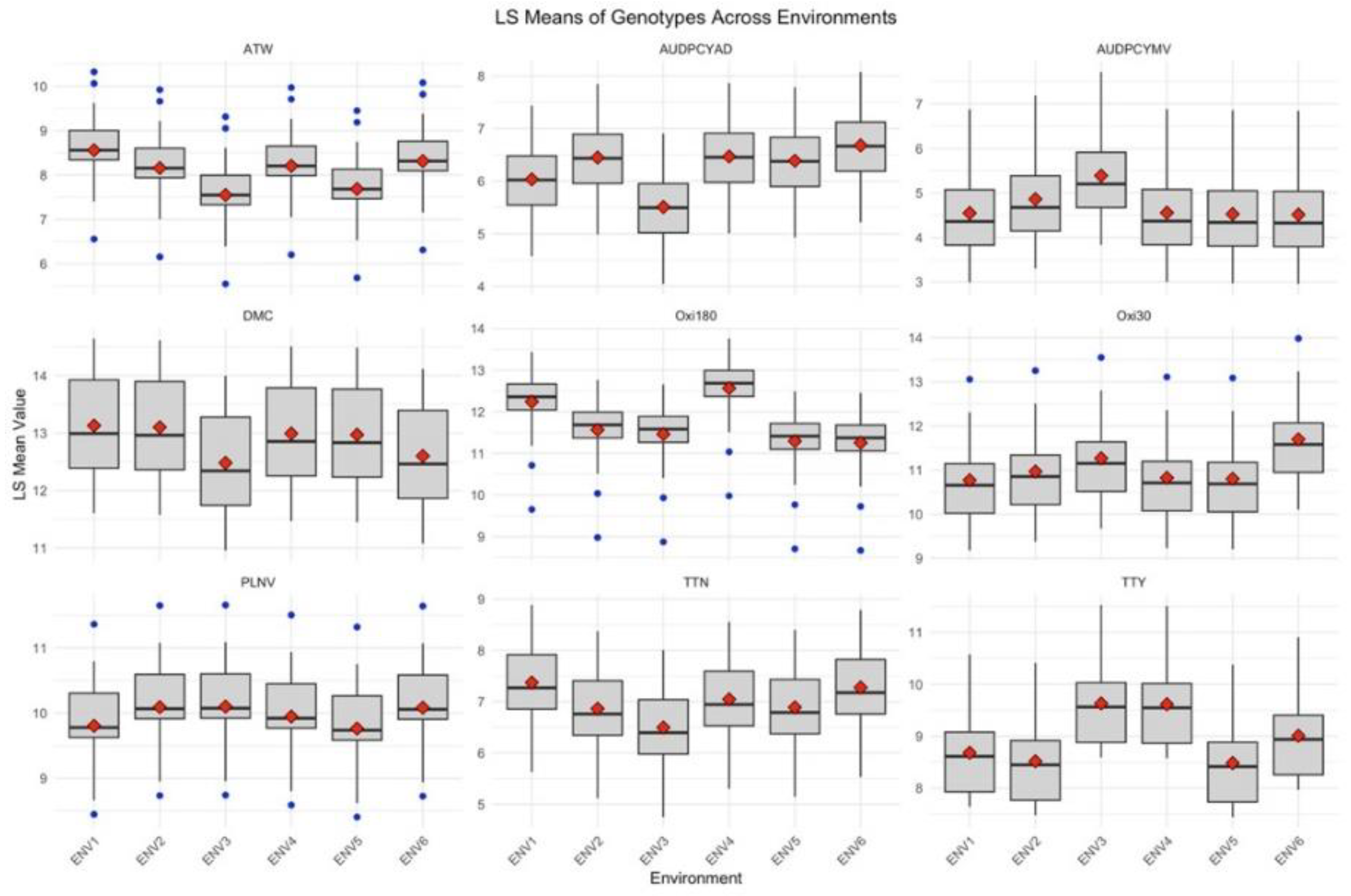
Figure 3.
Pearson correlation coefficient among the agronomic traits for 25 white yam genotypes*. *PLNV: plant vigor; AUDPCYAD: area under disease progression curve yam anthracnose disease; AUDPCYMV: area under disease progression curve yam mosaic virus, TTW: total tuber weight per plant; ATW: average tuber weight; TTY: total tuber weight; OXi30 and OXi180: intensity of tuber oxidation at 30 and 180 minutes and DMC: dry matter content.
Figure 3.
Pearson correlation coefficient among the agronomic traits for 25 white yam genotypes*. *PLNV: plant vigor; AUDPCYAD: area under disease progression curve yam anthracnose disease; AUDPCYMV: area under disease progression curve yam mosaic virus, TTW: total tuber weight per plant; ATW: average tuber weight; TTY: total tuber weight; OXi30 and OXi180: intensity of tuber oxidation at 30 and 180 minutes and DMC: dry matter content.
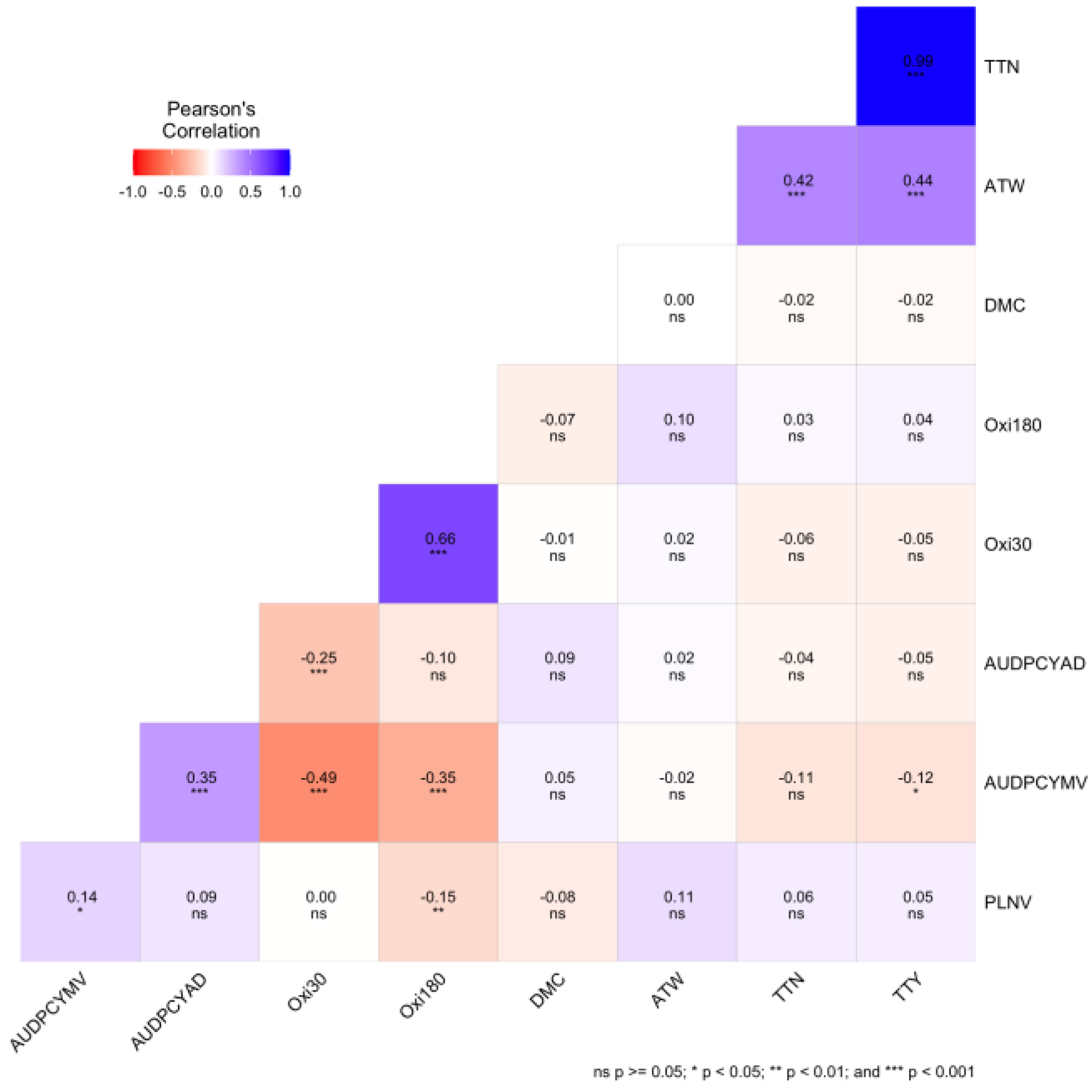
Figure 4.
Ammi1 biplot view for best genotypes across six environments (a) Total tuber yield (b)Dry matter content, (c)total tuber number.
Figure 4.
Ammi1 biplot view for best genotypes across six environments (a) Total tuber yield (b)Dry matter content, (c)total tuber number.
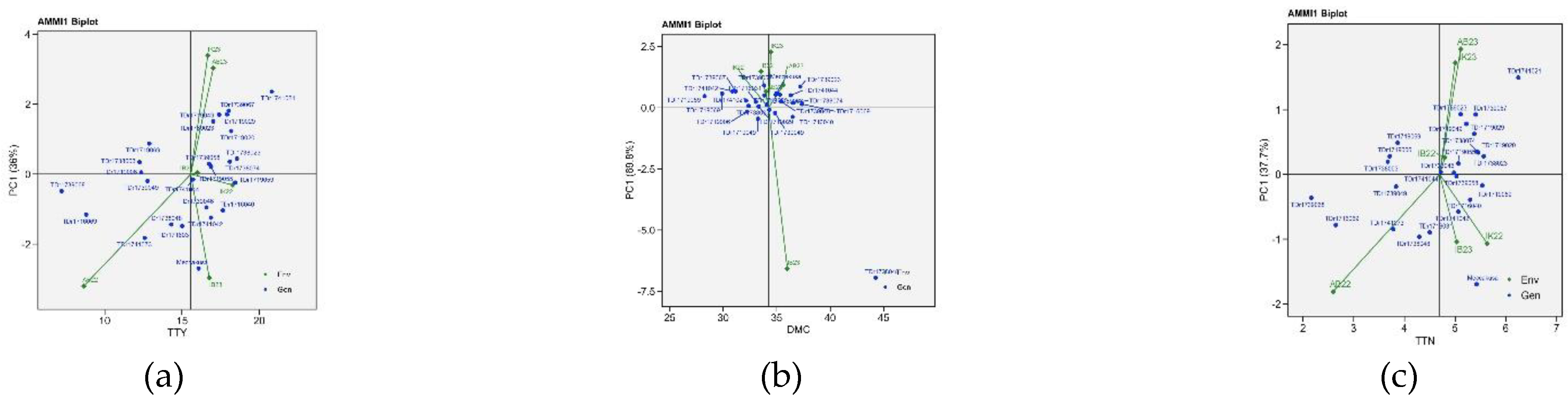
Figure 5.
Ammi2 biplot polygon view for best genotypes across six environments (a) total tuber yield, (b) dry matter content, (c) total tuber number.
Figure 5.
Ammi2 biplot polygon view for best genotypes across six environments (a) total tuber yield, (b) dry matter content, (c) total tuber number.
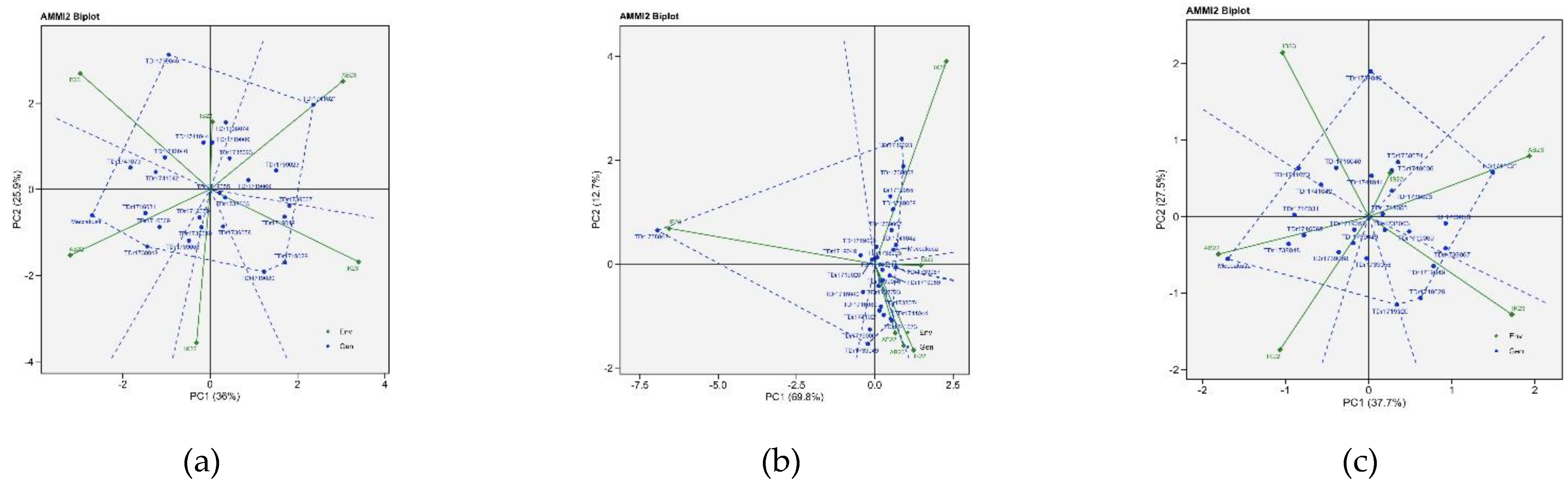
Figure 6.
Mean and stability biplot for total tuber yield (a), total tuber number (b) and dry matter content (c).
Figure 6.
Mean and stability biplot for total tuber yield (a), total tuber number (b) and dry matter content (c).
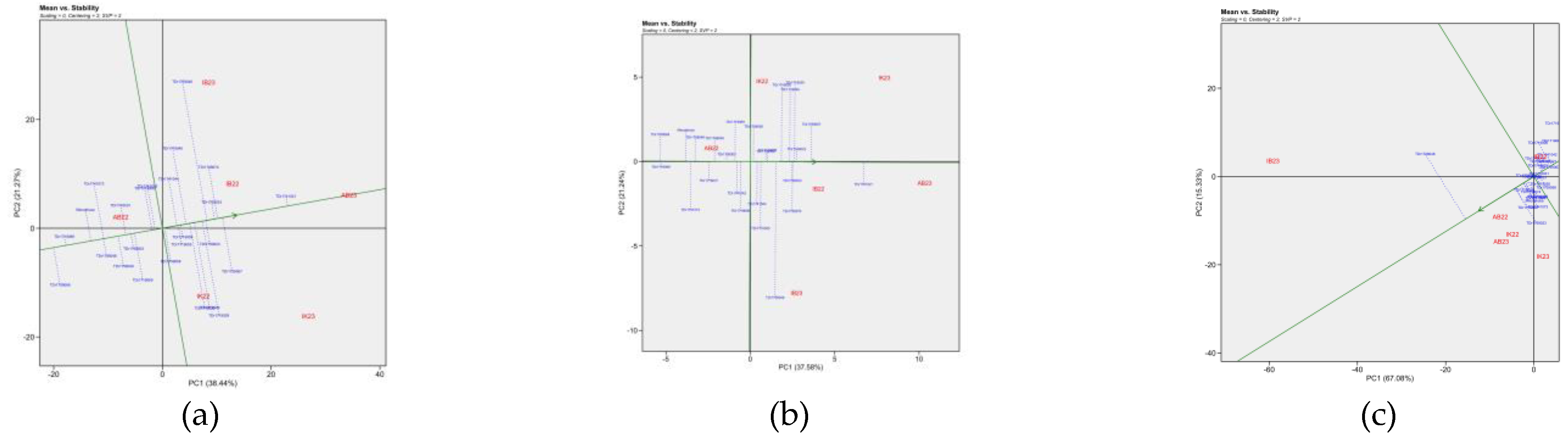
Figure 7.
Twenty-five genotypes of white yam selected by multi-trait stability index.
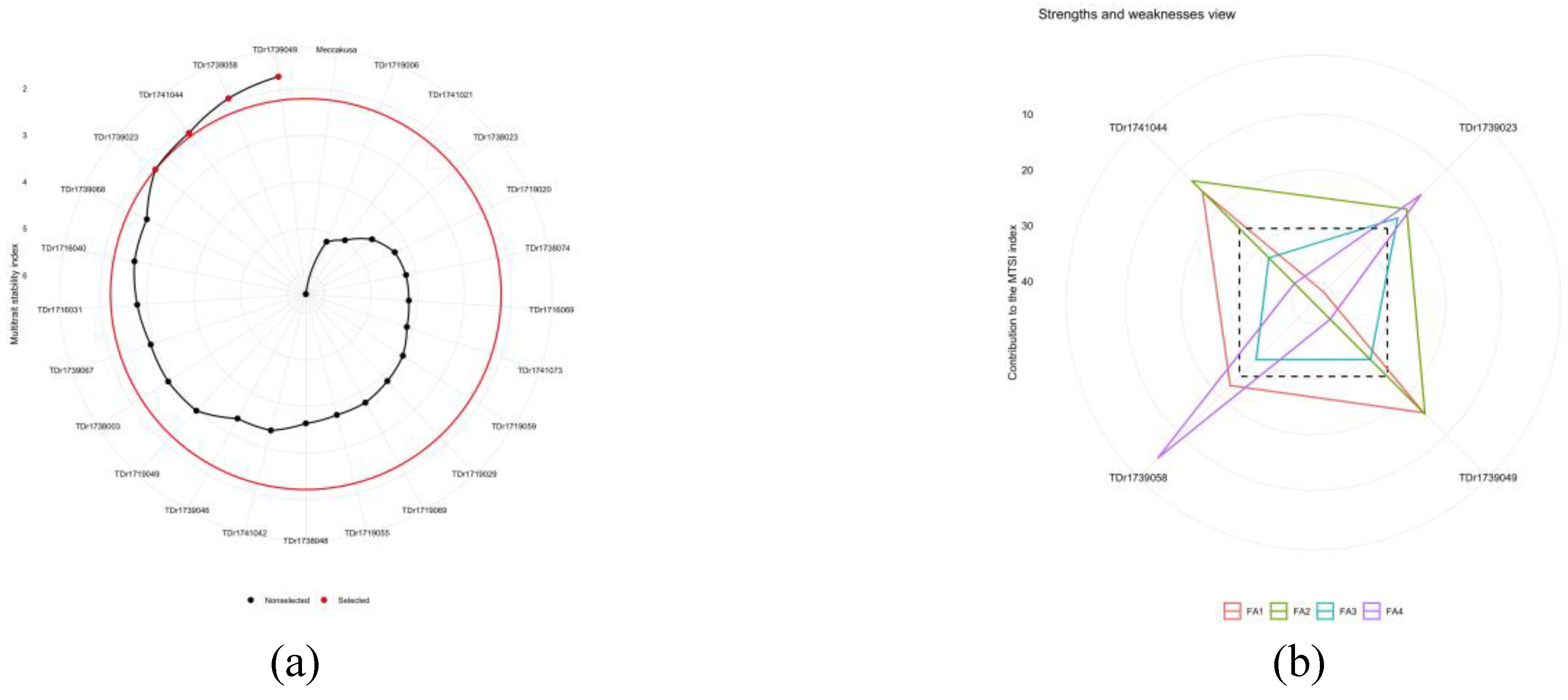

Table 1.
Agronomic traits, description and time recorded for this study.
| S/N | Traits | Full names | Description | Time recorded |
| 1 | AUDPCYAD | Area under disease progression刘curve yam anthracnose disease | The rating of symptoms caused by anthracnose over a period of 2-5MAP | Over the period of 2-5MAP |
| 2 | AUDPCYMV | Area under disease progression 刘curve yam mosaic virus | The rating of symptoms caused by virus over a period of 2-5MAP | Over the period of 2-5MAP |
| 3 | PLNV | Plant Vigor | How vigorous the plants appear at 3MAP | 3 MAP |
| 4 | INTOX30 | Intensity of tuber oxidation 30 minutes | Visual tuber oxidation was accessed at harvest | At harvest |
| 5 | INTOX180 | Intensity of tuber oxidation 180minutes | Visual tuber oxidation was accessed at harvest | At harvest |
| 6 | TTY | Total yield per plot | Yield was estimated per plot using the formula total tuber weight divided by the effective plot multiplied by ten | At harvest |
| 7 | DM | Dry matter content | Percentage of dry matter content of tuber | At harvest |
| 8 | ATW | Average tuber weight | Average weight of tuber per plot was accessed at harvest | At harvest |
All traits information was accessed using the Standard Operating Protocol for Yam Genotype Performance Evaluation Trial” [9].
Table 3.
Genetic parameters estimate in the selected white yam genotypes *.
| Genetic parameters | AUDPCYMV | AUDPCYAD | TTN | ATW | TTY | PLNV | Oxi30 | Oxi180 | DMC |
| GV | 54055.75 | 117840.00 | 365.58 | 36.68 | 3840.65 | 0.00 | 260.80 | 326.09 | 3672.45 |
| PV | 2350.25 | 2946.00 | 8.70 | 0.94 | 93.67 | 0.26 | 5.93 | 8.36 | 89.57 |
| H2 | 23.00 | 40.00 | 42.00 | 39.00 | 41.00 | 0.00 | 44.00 | 39.00 | 41.00 |
| CVg | 137.85 | 147.41 | 407.68 | 417.93 | 398.26 | 0.00 | 717.78 | 498.90 | 176.99 |
| CVp | 28.75 | 23.32 | 62.87 | 66.86 | 62.18 | 20.06 | 108.20 | 79.80 | 27.64 |
| Mean | 168.65 | 232.86 | 4.69 | 1.45 | 15.56 | 2.54 | 2.25 | 3.62 | 34.24 |
*GV=genotypic variance, PV=phenotypic variance, H2 broad sense heritability, CVg=Genotypic coefficient of variance, CVP=phenotypic coefficient of variance, PLNV: plant vigour; AUDPCYAD: area under disease progression curve yam anthracnose disease; AUDPCYMV: area under disease progression curve yam mosaic virus, TTW: total tuber weight per plant; ATW: average tuber weight; TTY: total tuber weight; OXi30 and OXi180: intensity of tuber oxidation at 30 and 180 minutes and DMC: dry matter content.
Table 4.
Principal component analysis and variables contribution on each factor for quantitative traits *.
Table 4.
Principal component analysis and variables contribution on each factor for quantitative traits *.
| Variables | PC1 | PC2 | PC3 | PC4 |
| AUDPCYMV | -0.29 | 0.42 | -0.08 | -0.20 |
| AUDPCYAD | -0.17 | 0.28 | -0.01 | -0.63 |
| TTN | 0.55 | 0.30 | 0.09 | 0.07 |
| ATW | 0.38 | 0.19 | -0.10 | -0.28 |
| TTY | 0.56 | 0.30 | 0.09 | 0.07 |
| PLNV | 0.01 | 0.18 | -0.71 | -0.22 |
| Oxi30 | 0.23 | -0.54 | -0.14 | -0.28 |
| Oxi180 | 0.25 | -0.46 | -0.06 | -0.39 |
| DMC(%) | -0.06 | 0.05 | 0.66 | -0.44 |
| eigenvalue | 2.35 | 2.14 | 1.08 | 1.05 |
| variance(%) | 26.08 | 23.76 | 12.02 | 11.69 |
| cumulative(%) | 26.08 | 49.84 | 61.87 | 73.55 |
*PLNV: plant vigor; AUDPCYAD: area under disease progression curve yam anthracnose disease; AUDPCYMV: area under disease progression curve yam mosaic virus, TTW: total tuber weight per plant; ATW: average tuber weight; TTY: total tuber weight; OXi30 and OXi180: intensity of tuber oxidation at 30 and 180 minutes and DMC: dry matter content, PC=principal component.
Table 5.
Mean squares of Additive main effect and multiplicative interaction (AMMI) analysis of variance for 25 genotypes evaluated for agronomic traits in six environments1.
Table 5.
Mean squares of Additive main effect and multiplicative interaction (AMMI) analysis of variance for 25 genotypes evaluated for agronomic traits in six environments1.
| Sources | df | AUDPCYMV | AUDPCYAD | TTN | ATW | TTY | PLNV | Oxi30 | Oxi180 | DMC |
| Environment(E) | 5 | 38808*** | 27679.4*** | 28.3268*** | 1.26671** | 300.556*** | 1.34567*** | 49.651*** | 41.826*** | 54.909 |
| Genotype(G) | 24 | 1057 | 1698.6* | 5.5966* | 0.65483* | 61.42* | 0.12718 | 3.151 | 5.313* | 55.97* |
| Interaction (G*E) | 120 | 798 | 1016.3 | 3.3071 | 0.37295 | 36.548 | 0.12981 | 2.054 | 3.263 | 34.021 |
| PC1 | 28 | 1705*** | 1864.4*** | 5.2033** | 0.70452*** | 53.8** | 0.2399*** | 3.045** | 5.526*** | 101.611*** |
| PC2 | 26 | 948*** | 1414.4*** | 4.339** | 0.46769** | 45.594* | 0.15599** | 2.557* | 4.634*** | 19.983* |
| Residuals | 66 | 354 | 499.7 | 2.0961 | 0.19496 | 25.665 | 0.0728 | 1.436 | 1.764 | 10.877 |
1PLNV: plant vigor; AUDPCYAD: area under disease progression curve yam anthracnose disease; AUDPCYMV: area under disease progression curve yam mosaic virus, TTW: total tuber weight per plant; ATW: average tuber weight; TTY: total tuber weight; OXi30 and OXi180: intensity of tuber oxidation at 30 and 180 minutes and DMC: dry matter content, PC=principal component.
Table 6.
Selection differential of the WAASB index for D50F, TSW, SY in sesame accessions*.
| Variable | Factor | FA1 | FA2 | FA3 | FA4 | Xo | Xs | SD | SD% | Communality |
| Oxi30 | FA 3 | -0.35 | 0.21 | -0.69 | -0.14 | 0.22 | 0.14 | -0.09 | -39.00 | 0.67 |
| Oxi180 | FA 3 | 0.21 | -0.20 | -0.76 | 0.14 | 0.30 | 0.22 | -0.08 | -26.40 | 0.68 |
| PLNV | FA 4 | 0.10 | 0.06 | -0.41 | -0.76 | 0.12 | 0.09 | -0.03 | -26.20 | 0.76 |
| ATW | FA 4 | -0.17 | -0.20 | 0.28 | -0.75 | 0.22 | 0.16 | -0.05 | -24.30 | 0.72 |
| TTN | FA 1 | -0.96 | -0.12 | -0.04 | -0.02 | 0.34 | 0.26 | -0.08 | -22.60 | 0.94 |
| AUDPCYMV | FA 2 | -0.17 | -0.89 | -0.19 | -0.03 | 1.29 | 1.04 | -0.25 | -19.30 | 0.85 |
| TTY | FA 1 | -0.99 | -0.02 | 0.03 | 0.01 | 0.66 | 0.53 | -0.13 | -19.20 | 0.97 |
| AUDPCYAD | FA 2 | -0.04 | -0.92 | 0.07 | -0.05 | 1.48 | 1.35 | -0.14 | -9.08 | 0.86 |
| DMC | FA 2 | -0.10 | 0.31 | -0.13 | 0.25 | 0.21 | 0.21 | 0.00 | -1.09 | 0.18 |
| Average | 0.7368696 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



