
Submitted:
13 August 2024
Posted:
14 August 2024
You are already at the latest version
Abstract
Gastrointestinal (GI) cancers, such as colorectal and gastric cancers, pose significant global health challenges due to their high rates of incidence and mortality. Even with advancements in treatment and early detection, many patients still face poor outcomes, highlighting the critical need for new biomarkers and therapeutic targets. Telomere length (TL) and telomerase activity (TA) have gained attention in this context. Telomeres, protective nucleotide sequences at chromo-some ends, shorten with each cell division, leading to cellular aging. Telomerase, a ribonucleoprotein enzyme, counteracts this shortening by adding telomeric repeats, a process tightly regulated in normal cells but often dysregulated in cancer. This review critically evaluates the role of TL and TA in the pathogenesis of GI cancers, examining their potential as diagnostic, prognostic, and predictive biomarkers. It explores how alterations in telomere biology contribute to the initiation and progression of GI tumors and assesses the therapeutic implications of targeting telomerase. By integrating findings from diverse studies, this review aims to elucidate the intricate relationship between telomere dynamics and gastrointestinal carcinogenesis, offering in-sights into how TL and TA could be leveraged to enhance the early detection, treatment, and prognosis of GI cancers.
Keywords:
Gastrointestinal (GI) cancers
; biomarkers
; telomerase activity
; Telomere length
1. Introduction
In 2022, an estimated of 20 million new cancer cases were documented worldwide, 26% of which affecting the gastrointestinal (GI) tract. In detail, colorectal cancer (CRC) was the most prevalent malignancy (9.6%), followed by gastric cancer (6.8%) [1,2]. However, despite great efforts from the World Health Organization and national scientific societies to decrease their incidence, malignancies of the GI tract seem to maintain their incidence with CRC being steadily placed in the second place of the most common type of cancer in both sexes. An abundance of evidence suggests that more than half of all the malignancies of the GI tract is attributed to modifiable risk factors such as unhealthy diet habits (high fat, high sugar, low fiber), increased alcohol consumption, smoking, obesity and low physical activity [1,2]. In fact, these risk factors are the key targets of primary prevention strategies that aim to decrease the carcinogenic that modern societies face.
Unfortunately, to a certain extent, these risk factors have become part of the western lifestyle that more and more people follow across the globe and this is the reason why primary prevention has failed to achieve more significant results [1,2,3,4]. Secondary prevention aims to detect premalignant lesions or early state malignancies and treat them in situ. To do so, endoscopy of the upper and lower GI tract has become the modality of choice as it offers the advantage of diagnosis and treatment in the same session. However, evidence suggests that adherence to endoscopy protocols is significantly lower than expected partly due to the invasive nature of the test. For this reason, non-invasive tests have been introduced to assist in an early diagnosis while also providing valuable information as for the prognosis and prediction in case where a cancer has been diagnosed or a specific treatment has been chosen [5,6]. One major step in improving early diagnosis, prognosis and prediction in regard to cancer, is the introduction of biomarkers in everyday clinical practice [7]. A biomarker is a biological molecule found in body fluids or tissues that indicates a normal or abnormal process, condition, or disease [6]. Tumor biomarkers are substances that can be detected in tissues or body fluids such as blood, urine, stool, and saliva [6]. They can generally be synthesized directly by the tumor or by the body in response to the presence of cancer [6,8]. Two promising biomarkers with diagnostic, prognostic or predictive significance in GI tract malignancies are telomere length (TL) and telomerase activity (TA) both of which are important regulators of normal cell function while also playing an important role in carcinogenesis. Overall, the aim of this review paper is to consolidate the findings from published studies on the relationship between TL, TA and their roles as diagnostic, prognostic and predictive biomarkers in gastrointestinal cancer.
1.1. Telomeres
Telomeres, consisting of repetitive nucleotide sequences (5′-TTAGGG-3′), are special DNA structures found at the ends of chromosomes which are made up of repeated hexameric DNA sequences (5’-TTAGGG-3’) and a protein complex called shelterin, formed by six proteins: TRF1, TRF2, TIN2, POT1, TPP1, and RAP1 [9,10]. Telomeres aid in the prevention of degradation and help distinguish chromosome ends from double-strand breaks [11,12]. Working along with shelterin proteins, they play a critical role in maintaining chromosome stability and keeping them from attrition [13]. Telomeric DNA ends with a G-rich single-strand overhang ranging from 50 to 300 nucleotides, folding back to create a "T-loop", encompassing telomeres and shelterins [11T-loops play a crucial role in safeguarding telomeres by preventing DNA repair mechanisms from mistakenly identifying them as double-stranded DNA breaks. With each mitotic cell division, there is a strand of telomere DNA at the chromosome end which lags behind. This cannot be fully replicated by DNA polymerases, resulting in an annual rate of telomere shortening ~20-50 bp,that ultimately leads to cell proliferation arrest and senescence [14]. This phenomenon known as the 'end replication problem' causes a gradual reduction in average telomere length with the rate accelerating after the age of 50 [15,16]. Telomere shortening is influenced by genetic factors like inherited telomere length and telomerase activity, lifestyle such as smoking, diet, and physical activity, and by environmental factors including stress and exposure to pollutants. Chronic inflammation and diseases can also accelerate telomere shortening. [17,18,19,20,21]. Therefore, average TL is suggested as a marker of the 'biological age' of cells and the organism as a whole [16].
With each cell division, telomeres become shorter as the chromosome's 3' end cannot be fully replicated [11]. However, cancer cells can reactivate telomerase, allowing them to divide indefinitely [22]. Telomerase expressed reverse transcriptase (TERT) is silenced to prevent indefinite cell replication, leading to apoptosis or cell senescence. Telomere homeostasis depends on TL and the concentration of active telomerase. Shelterin complex proteins, which typically include TRF1, TRF2, RAP1, POT1, TIN2, and TPP1, can regulate telomere homeostasis by inhibiting telomerase action. TA is restricted to the S phase of the cell cycle, and not all telomeres undergo lengthening after DNA replication—only those prioritized based on their telomere lengths [23].
Genome instability is widely recognized as a key characteristic of carcinogenesis and is considered key in cancer development.
In the accepted model of carcinogenesis, telomeres in somatic cells gradually shorten with every cell division[24]. After roughly 50 to 60 cell divisions, shortened telomeres cause chromosomal instability, leading to replicative senescence through the activation of p53. This process is triggered by DNA damage linked to telomere shortening [24].
A decrease in telomere length with age can contribute to genetic instability and raise the likelihood of cancer. TL measured in peripheral blood of individuals in their seventies are predictive of survival, with the strongest association found between TL and mortality from heart disease and infections. It is said that telomere dysfunction occurs early in the development of cancer in ulcerative colitis, leading to chromosomal instability and the formation of anaphase bridges [25,26]. This process may expedite the molecular progression of carcinogenesis by promoting chromosomal instability in cells [27,28].
1.2. Telomerase
The ribonucleoprotein enzyme telomerase acts to maintain telomere length by employing an RNA template in its reverse-transcriptase function to synthesize telomeric DNA repeats [27]. Telomerase consists of telomerase reverse transcriptase (TERT) and telomeric RNA component (TERC) [12,22,28] (Figure 1).
Although TERT is mostly found in the nucleus, a portion (10–20%) is located in the mitochondria. TERT’s non-telomeric roles include reducing reactive oxygen species (ROS), DNA damage, and apoptosis. Recent research has shown that TERC enters mitochondria, is processed into a shorter form (TERC-53), and then returns to the cytosol. Damage to mitochondria, such as that caused by mitochondrial ROS, can lead to telomere dysfunction. Additionally, ROS from abnormal mitochondrial activity can cause single-strand breaks in telomeres, contributing to their dysfunction. Dysfunctional telomeres suppress PGCα/β in a p53-dependent manner, reducing ATP production, increasing ROS levels, and impairing metabolic functions like gluconeogenesis. This indicates a link between telomeres and mitochondria. Therefore, identifying more mediators in this interaction is essential for understanding the molecular mechanisms underlying carcinogenesis and cancer progression [10,29].
Human telomerase RNA is consistently present across various tissues, making human telomerase reverse transcriptase (hTERT) the critical factor controlling telomere activity. In some cancers, the loss of telomeres can be counterbalanced by the alternative lengthening of telomeres (ALT) pathway, which operates independently of telomerase [12,30]. In the absence of telomerase, the alternative lengthening of telomeres (ALT) pathway relies on homologous recombination for DNA replication to upkeep cellular longevity. While telomerase is present in 85–95% of cancer cells, approximately 5–15% of these cells use the ALT mechanism to sustain their telomeres [24,31,32].
Cellular senescence functions as a crucial safeguard against cancer by inhibiting cells from achieving immortality. In response, cancer cells often elevate their TA to mitigate the natural shortening of telomeres during cell division. This increase is driven by varying levels of hTERT gene expression observed between cancerous and normal cells [28]. It has been observed that telomerase is active in about 90% of human tumors, crucially maintaining telomere lengths and promoting tumor growth [29,33]. Thus, TA will be higher in tumors compared to healthy tissue. An early study by Shay and Bacchetti revealed that increased TA is generally reported in more than 85% of cancer cells [31]. It is intriguing to consider that cells with severely shortened telomeres might activate telomerase under specific conditions to evade cellular senescence, thereby facilitating the development of cancer [34,35].
Nevertheless, it is essential to note that the presence of the longest telomeres does not necessarily correspond to the highest TA. Upon activation, telomerase can maintain telomeres at varying lengths. Crucially, the relationship between TL and TA is indicative of the extent of cancer cell infiltration [36].
2. Telomere Length and Telomerase Activity in Gastrointestinal Cancers
Telomeres serve as crucial chromosome protectors, essential for preserving genomic stability. These DNA-protein complexes consist of repetitive "5’-TTAGGG-3’" sequences and an associated terminal protein complex. Over time, telomeres progressively shorten, leading to chromosomal instability in normal somatic cells. This shortening is linked to various aging-related diseases, including gastrointestinal cancer [11,35]. However, the relationship between TL and cancer risk remains a subject of debate, with studies yielding conflicting results [38].
Pooley et al. observed that significant telomere shortening primarily takes place after a cancer diagnosis, rather than before or during its development [16]. Oxidative stress accelerates telomere shortening by creating single-strand DNA breaks from oxidative or alkylative damage, which are inadequately repaired in the telomeres [39,40,41]. Previous research has shown that people with shorter average telomere lengths might be at a higher risk of dying from various diseases [16].
In 2016, Zhu and colleagues conducted a comprehensive review of studies examining the link between telomere length and cancer risk [42]. The analysis, which included data from 23,379 cancer patients and 68,792 controls across 51 studies, identified a significant correlation between telomere length and gastrointestinal and head and neck cancers. However, no significant connection was found between telomere length and overall cancer risk [42].
Telomerase is commonly activated in cancers to lengthen telomeres (Figure 2). Barthel et al. analyzed telomere lengths in 18,430 samples from 31 types of cancer using data from The Cancer Genome Atlas (TCGA). Their findings revealed that 70% of these cancers had shorter telomeres compared to normal samples, which was associated with increased telomerase activity [43]. In the remaining 30% of cancer types, telomere length was observed to be the same as control or longer due to suspected alternative lengthening of telomeres (ALT) activity [24,43,44]. Conversely, TA is significantly increased in the majority of cancers studied across various malignancies, including colorectal cancer [45,46,47] and pancreatic cancer [45,46]. The distinct contrast in appearance between normal or benign tissues and malignant tissues indicates that telomerase could serve as a universal biomarker for cancer diagnosis and prognosis.
Below, Table 1 provides a collective summary of studies investigating TL and TA in gastrointestinal cancers and their respective bibliographic references.
2.1.1. Esophageal Cancer
The reduction in telomere length begins early in the process of esophageal cancer development, while short telomeres on chromosomes 17p and 12q have been linked to a higher risk of esophageal cancer [51]. Wennerström et al. conducted a study to explore whether individuals with gastroesophageal reflux disease, a known risk factor for esophageal adenocarcinoma, had shorter telomeres in their white blood cells [52]. The researchers found no increased prevalence of either shorter or longer telomeres in individuals with esophageal cancer.
Valdes et al. and O’Sullivan et al. observed a direct correlation between telomere length and advancing age [19,26]. This finding was reinforced by research from Xing and colleagues, who investigated chromosome-specific telomere lengths on chromosomes 17p, 12q, 2p, and 11q in relation to esophageal cancer [51].
In research conducted by Risques et al., telomere length in the blood of Barrett's esophagus patients was found to be a predictor of their likelihood of developing esophageal adenocarcinoma [39]. This association held true even when other factors like gender, age, and smoking were considered. Patients with the shortest leukocyte telomeres had the greatest risk, especially if they were smokers. This highlights the role of short telomeres as a strong indicator of cancer risk, which can be influenced by environmental factors like smoking and oxidative stress [41].
In contrast, Lv et al. reported that longer telomeres are linked to a worse prognosis, establishing them as a standalone negative prognostic factor for esophageal cancer patients [53]. This finding supports the data from Li et al. indicating that inhibition of hTERT expression is able to restrain the migration and invasion abilities of malignant cells in case of esophageal squamous cell carcinoma [54]. In a study by Li et al., it was found that there is increased telomerase activity in esophageal squamous cell carcinoma (ΕSCC) [55]. Ikeguchi et al. and Takubo et al. found that telomerase activity was present in both cancerous tissues and normal esophageal mucosa, indicating that telomerase activity might not serve as a dependable biomarker for esophageal cancer detection [56,57].
Pursuing this further, Mitsui et al. observed in their study that telomerase activity was found in 79.6% of tumors and 59.3% of normal tissues, with significantly higher levels in the tumors [58]. Tumors with extensive blood vessel invasion exhibited increased telomerase activity. Additionally, tumors that responded well to preoperative chemotherapy had notably lower telomerase activity compared to those that did not respond. In an esophageal cancer cell line, treatment with the chemotherapy drug 5-FU led to a reduction in hTERT mRNA expression, resulting in decreased telomerase activity. These findings indicate that telomerase activity is linked to tumor invasion and chemotherapy response in esophageal cancer.
2.1.2. Gastric Cancer
In 2018, gastric cancer ranked as the second most prevalent cancer in the gastrointestinal tract, with more than one million new cases and nearly 800,000 deaths [4].
Based on a long-term study conducted by Shi et al. involving high-population for gastric cancer, the authors reported that individuals with shorter telomeres in cell-free DNA faced a heightened risk of gastric cancer progression [59]. Furthermore, shortened telomeres could be detected over a three-year period prior to the diagnosis of gastric cancer.
In research conducted by Wang et al., involving Chinese participants with an average age of 67, both extremely short and long telomeres were linked to a higher risk of gastric cancer, with increased risks of 63% and 55%, respectively [60]. These findings are consistent with observations made by Hou et al. and Wentzensen et al. [11,50]. The association between telomere length and cancer risk may be due to short telomeres causing chromosomal instability and initiating cancer, while long telomeres could lead to excessive cell division and genetic abnormalities, thereby increasing cancer risk [60,61].
In another study, the authors reported of shorter telomeres in gastric cancer tumor cells at an early stage than those in the surrounding non-cancerous tissue cells [62]. Telomere shortening seems to initiate gastric cancer, while the extension or maintenance of telomere length via telomerase reactivation is crucial for the cancer's invasion and progression. Liu et al. supported these findings by showing that patients with gastric adenocarcinoma had significantly shorter leukocyte telomeres compared to controls. Additionally, progressive telomere shortening was associated with an increased risk of developing the disease [38].
Katayama et al. studied telomerase activity and telomere length in gastric and colorectal cancers [63]. They found that TA was present in 8% of gastric polyps and 22% of colorectal polyps. In contrast, TA was significantly more common in gastric cancer (70%) and colorectal cancer (81%), with p-values below 0.0003 and 0.0001, respectively.
In a similar study, Rathi et al. analyzed human telomerase RNA (hTR) expression in gastric cancer through in situ hybridization on tissue samples [64]. They found that hTR levels were low in normal gastric mucosa, chronic peptic ulcers, and hyperplastic polyps. Tubular adenomas exhibited weak but more widespread hTR expression. In contrast, gastric carcinomas showed moderate to high levels of hTR, with expression intensity increasing as the cancer progressed. These findings suggest that telomerase upregulation occurs early in gastric cancer development and could serve as an important marker for early detection.
Another study by Hu et al. examined the importance of telomerase activity in gastric carcinoma tissues and peritoneal washings [65]. Telomerase activity was present in 89.1% of gastric carcinomas and 47.8% of peritoneal washings, and it was linked to factors such as histological grade, invasion depth, serosal invasion, and peritoneal metastasis. The detection rate in peritoneal washings was significantly higher than those found by cytology (26.1%) and cancer antigen 125 (CA125) (34.8%). The authors proposed that telomerase activity could be an effective diagnostic marker for gastric carcinoma and that identifying it in peritoneal washings via TRAP-ELISA may aid in detecting early peritoneal dissemination in gastric cancer patients. These results were in agreement with findings reported by Svinareva et al. investigating telomerase activity in tissue specimens from patients with gastric adenocarcinomas and gastric lymphoma using a modified TRAP assay [66]. Telomerase activity was present in 16 of 18 (89%) patients with gastric adenocarcinomas and in one patient with gastric lymphoma, but not in the control patient with non-cancerous gastric tissue. The majority of the samples (88%) showed "high" or "very high" levels of telomerase activity. These findings suggest a strong correlation between telomerase activity and malignancy, proposing its potential as a diagnostic marker for gastric cancer [66].
Furthermore, Pascua et al. explored how telomere function affects the prognosis of gastric cancer with different levels of microsatellite instability (MSI) [67]. Their study revealed that patients with high microsatellite instability had significantly better outcomes compared to those with microsatellite stable or low microsatellite instability tumors. They also observed that tumors with the shortest telomeres were associated with a worse prognosis. Similarly, Mushtaq et al. found that shorter telomeres and elevated hTERT expression were related to the progression of gastric cancer [68].
2.1.3. Colorectal Cancer
Numerous cancers associated with shortened telomeres, such as bladder and gastric cancers, are often linked to chronic inflammation or carcinogenic exposures like smoking, in the case of bladder and lung cancers [11]. Chronic inflammation, which is a recognized risk factor for cancers like esophageal, bladder, and gastric cancers, leads to an increased turnover of granulocytes [11]. Since telomeres shorten with each cell division, this accelerated turnover can lead to shorter telomeres in granulocytes and a reduced overall length of leukocyte telomeres [11,69]. Certain cytokines have the ability to activate telomerase, potentially mitigating telomere shortening to some extent. Ulcerative colitis illustrates the complex relationship between inflammation, cancer risk, and telomere dynamics [11,70]. Individuals with this condition face a higher risk of colon cancer, especially those with increased chromosomal instability and shorter telomeres [11,71].
Research indicates a possible link between intestinal microbiota and TL, especially in the context of aging and oxidative stress. This connection highlights the potential of using TL as a diagnostic and prognostic marker, as well as a target for novel anti-cancer therapies, due to its role in colorectal cancer development. Telomeres shorten with each cell division and this process speeds up with age, leading to an increase in senescent cells that release inflammatory cytokines and contribute to age-related diseases [72]. Factors such as poor diet, smoking, and inactivity raise reactive oxygen species (ROS) levels, accelerating telomere shortening [73]. Additionally, aging-related mitochondrial dysfunction, which boosts ROS production, further aggravates telomere damage [72].
Oxidative stress speeds up telomere shortening by damaging telomeric DNA, including single-strand breaks and base alterations, with guanine being particularly vulnerable [72,74]. Antioxidants like peroxiredoxin 1, which protect telomeres, decrease with age, leading to further telomere damage [74]. Intestinal microbiota can influence oxidative stress and inflammation [72; 75]. Certain bacteria associated with inflammatory bowel disease and colorectal cancer, including enterotoxigenic B. fragilis, adherent-invasive E.coli, and Fusobacterium nucleatum among others, increase oxidative stress [76]. Gut bacteria also impact oxidative stress by modulating mitochondrial activity and producing short-chain fatty acids like acetate and butyrate, which are linked to reduced oxidative stress and inflammation [77,78].
Recent studies have indicated that changes in TL independently correlate with the progression and prognosis of colorectal cancer [26,79,80,81,82,83,84,85,86]. However, the abovementioned studies measured telomere length in colonocytes taken from paired samples of cancerous and non-cancerous tissues within the same individuals, compared to peripheral blood leukocyte telomere lengths as described in a study conducted by Zee et al. [87]. Collectively, these findings indicate that telomere behavior in colonocytes varies from that observed in other tissues, including peripheral blood leukocytes.
When considering tumor location, rectal cancers generally have a poorer prognosis compared to colon cancers, and clinical treatments vary accordingly [88,89]. Previous findings have shown that TL differs based on tumor location, with rectal cancers typically exhibiting longer telomeres [89,90]. These results align with the findings of Zöchmeister et al. and Peacock et al., who reported that telomeres were significantly longer in colorectal cancer patients compared to controls [91,92]. Additionally, Kibriya et al. found that among 165 colorectal cancer patients, telomere shortening was more pronounced in lower-grade cancers and in those with microsatellite instability [30]. Other research indicates that microsatellite instability is found in approximately 15% of colorectal cancers, whereas chromosomal instability with stable microsatellites is often linked to TP53 gene mutations [30,93,94].
In Kroupa et al.'s study, telomeres were shorter in tumor tissues compared to adjacent mucosa in 74% of colorectal cancer patients [95]. Shorter telomeres were linked to tumors in the proximal colon, microsatellite instability, mucinous histology, and earlier TNM stages. Similarly, metastatic liver tissues showed reduced telomere length compared to adjacent healthy liver. Patients with a smaller difference in telomere length between tumor and adjacent tissue had better survival outcomes. These findings indicate that telomere length variations, depending on tumor location and characteristics, may influence prognosis and treatment approaches.
In a later study by the same authors expanding the focal point to metastatic colorectal cancer lesions, it was reported that most primary tumors showed shorter telomeres compared to adjacent non-cancerous tissue, indicating a potential role in malignant transformation [96]. Metachronous liver metastases showed shorter telomeres than synchronous ones, with the shortest telomeres found in proximal colon tumors. Neoadjuvant chemoradiotherapy notably shortened telomeres in rectal tumors and nearby tissues. A higher tumor-to-mucosa telomere length ratio was associated with better overall survival. These findings underscore the significance of telomere dynamics in colorectal cancer development.
Ye et al. examined telomere length in colorectal cancer and adenoma cells using quantitative fluorescent in situ hybridization (Q-FISH) [97]. The results indicated that telomere fluorescent intensity units (TFUs) were significantly lower in carcinoma and adenoma cells compared to cancer-associated fibroblasts (CAF), with adenoma cells exhibiting the shortest telomeres. In carcinoma cells, reduced telomere fluorescent intensity units (TFUs) and relative telomere length (RTL) were associated with distant metastases and poorer overall and disease-free survival. These findings suggest that telomere shortening might happen early in colorectal cancer progression and could serve as a prognostic indicator for the disease.
Telomerase has been recognized as a crucial predictor of overall survival in colorectal cancer patients, with those having higher TERT levels showing markedly worse survival rates compared to those with lower TERT levels [93].
Engelhardt et al. examined telomerase levels in various colon tissues, finding that while telomerase is generally low in non-cancerous cells, it was notably higher in colorectal cancer samples. Their study analyzed 130 frozen specimens, including cancerous tissues, adjacent normal tissues, polyps, and colitis. They detected moderate to high telomerase activity in 90% of colorectal tumors and weak activity in the remaining 10%. Normal colon tissues showed no telomerase activity, and polyps and colitis had significantly lower levels than cancerous tissues. Higher telomerase activity was associated with advanced-stage tumors compared to early-stage ones [84]. In a separate study by Garcia-Aranda et al., 91 primary colorectal cancers and their normal counterparts were assessed for telomere length and telomerase activity. This research found that 81.3% of tumors had telomerase activity, and telomeres were shorter in cancer tissues compared to normal ones (p = 0.02). Tumors with shorter telomeres also had higher levels of TRF1 [80].
2.1.4. Liver Cancer
Telomere shortening is a feature observed in intrahepatic cholangiocarcinoma (iCCA), akin to what is seen in hepatocellular carcinoma (HCC). In a healthy liver, cholangiocytes typically have longer telomeres compared to other liver cells, and telomere shortening related to aging is not prominent in the absence of liver disease. However, telomere shortening becomes evident early in the progression of biliary tract carcinoma, beginning with inflammation of the biliary tract and continuing through metaplasia, dysplasia, and carcinoma [97]. Telomerase activity is present in a majority of both HCC and iCCA cases, indicating its importance in telomere maintenance and tumor development. Despite this, the mechanisms behind telomerase reactivation differ between HCC and iCCA. TERT promoter mutations are commonly found in HCC but are rare in iCCA, suggesting that other mechanisms like epigenetic or transcriptional regulation might be more significant in iCCA. These differences highlight the need for further research to identify potential therapeutic targets specific to iCCA [97].
Research has shown that telomere shortening is common in chronic liver disease [98,99]. The impact of telomere length on liver cancer risk varies depending on the cancer's stage. Reduced telomeres can promote liver cancer development [41,100], and the risk of hepatocellular carcinoma (HCC) increases notably during cirrhosis, which is associated with shorter hepatocyte telomeres. Interestingly, while telomeres are generally shorter in HCC patients compared to healthy controls, those with advanced HCC often have longer telomeres [22,101,102].
In normal human liver, there is no significant telomerase activity [103]. In contrast, more than 80% of human HCC exhibit increased level of TA primarily due to the re-expression of hTERT [104,105]. Moreover, increased TA in HCC or surrounding healthy tissue has been identified as an indicator of poor prognosis since it correlates with post-operative recurrence and poor survival [106,107].
2.1.5. Pancreatic Cancer
Telomere length and telomerase activity play significant roles in the development and progression of pancreatic cancer. Unlike colorectal cancer, where shortened telomeres are prevalent, Duell's research suggests that reduced telomere length might increase the risk of pancreatic cancer and its progression [108]. Conversely, Lynch et al. found that longer telomeres were linked to a higher risk of pancreatic cancer [109], a conclusion also supported by Luu et al. and Campa et al. [110,111]. Additionally, Skinner's extensive case-control study demonstrated a clear connection between shorter telomeres and an elevated risk of pancreatic cancer [35]. Telomerase activity helps distinguish pancreatic cancer from other pancreatic diseases [112]. Hiyama et al. noted that nearly 95% of pancreatic adenocarcinoma patients exhibited increased TA [113], whereas benign conditions like pancreatitis had lower TA. Testing for telomerase activity in pancreatic juice has proven useful for differentiating malignant intraductal papillary mucinous neoplasms (IPMNs) from benign ones [114]. Additionally, Nakashima et al. found that about 84% of pancreatic ductal adenocarcinomas (PDACs) expressed hTERT [115], and Hashimoto et al. identified increased TA in 83% of invasive ductal adenocarcinomas (IDCs) and hTERT expression in 88% of cases [116]. These findings suggest that hTERT expression might be a more reliable marker than TA alone. Overall, high hTERT levels and increased TA are linked to poorer outcomes in pancreatic cancer [117].
3. Conclusions
Telomeres, which are repetitive hexameric DNA sequences located at the ends of chromosomes, play a crucial role in safeguarding and replicating the genome. The mechanisms that control telomere maintenance are vital in the progression of cancer, positioning them as key focus areas in cancer treatment research. In many cancers, tumors achieve unlimited replicative potential primarily by activating the enzyme telomerase through increased transcription of the TERT gene. Measuring telomerase activity or hTERT expression in blood may have some diagnostic potential, but issues regarding specificity remain problematic. Activated lymphocytes also show telomerase activity, making lymphocyte contamination a potential confounder. While hTR and hTERT have been studied at the RNA transcript level, a major challenge is the absence of reliable hTERT antibodies for immunohistochemistry. Therefore, targeting telomerase is a prominent strategy in the development of cancer therapeutics.
As telomere biology continues to grow as a research field, there are many avenues for future development to explore. The discovery of better risk biomarkers could allow surveillance and prevention efforts to target high-risk patients specifically, potentially reducing costs, patient anxiety, and the risk of complications significantly.
Although telomere shortening is widely acknowledged as an early marker in cancer development, its role in assessing cancer progression through telomere length and telomerase activity remains controversial. While there is evidence suggesting a connection between telomere length and cancer risk, challenges in study design create uncertainties. Variability in the association between cancer and telomere length further complicates the issue. To better understand gastrointestinal cancer biology, combining telomere length and telomerase activity with clinical factors might be beneficial. This approach could lead to the discovery of new biomarkers, aiding in predicting recurrences and informing treatment decisions, thereby improving clinical management of the disease.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fidler, M.M.; Soerjomataram, I.; Bray, F. A global view on cancer incidence and national levels of the human development index. Int. J. Cancer 2016, 139, 2436–2446. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Abnet, C.C.; Neale, R.E.; Vignat, J.; Giovannucci, E.L.; McGlynn, K.A.; Bray, F. Global Burden of 5 Major Types of Gastrointestinal Cancer. Gastroenterology 2020, 159, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Islami, F.; Goding Sauer, A.; Miller, K.D.; Siegel, R.L.; Fedewa, S.A.; Jacobs, E.J.; McCullough, M.L.; Patel, A.V.; Ma, J.; Soerjomataram, I.; et al. Proportion and number of cancer cases and deaths attributable to potentially modifiable risk factors in the United States. CA A Cancer J. Clin. 2017, 68, 31–54. [Google Scholar] [CrossRef] [PubMed]
- Bray F, Laversanne M, Sung H; et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Qu, X. Cancer biomarker detection: recent achievements and challenges. Chem. Soc. Rev. 2015, 44, 2963–2997. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tao, L.; Qiu, J.; Xu, J.; Yang, X.; Zhang, Y.; Tian, X.; Guan, X.; Cen, X.; Zhao, Y. Tumor biomarkers for diagnosis, prognosis and targeted therapy. Signal Transduct. Target. Ther. 2024, 9, 132. [Google Scholar] [CrossRef] [PubMed]
- Sewpersad, S.; Pillay, T.S. Historical perspectives in clinical pathology: Bence Jones protein—early urine chemistry and the impact on modern day diagnostics. J. Clin. Pathol. 2020, 74, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, C.A.; Wang, B.; Demaria, M. Senescence and cancer — role and therapeutic opportunities. Nat. Rev. Clin. Oncol. 2022, 19, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H. Structure and function of telomeres. Nature 1991, 350, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, S.; Xie, W.; Wang, Q.; Luo, Q.; Huang, M.; Gu, M.; Lan, P.; Chen, D. MCCC2 is a novel mediator between mitochondria and telomere and functions as an oncogene in colorectal cancer. Cell. Mol. Biol. Lett. 2023, 28, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wentzensen, I.M.; Mirabello, L.; Pfeiffer, R.M.; Savage, S.A. The Association of Telomere Length and Cancer: A Meta-analysis. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1238–1250. [Google Scholar] [CrossRef] [PubMed]
- Heaphy, C.M.; Meeker, A.K. The potential utility of telomere-related markers for cancer diagnosis. J. Cell. Mol. Med. 2011, 15, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Wright, W.E. Telomeres and telomerase: Three decades of progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef]
- Son, N.H.; Murray, S.; Yanovski, J.; Hodes, R.J.; Weng, N.-P. Lineage-Specific Telomere Shortening and Unaltered Capacity for Telomerase Expression in Human T and B Lymphocytes with Age. J. Immunol. 2000, 165, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.; Wright, W. Hallmarks of telomeres in ageing research. J. Pathol. 2007, 211, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Pooley KA, Sandhu MS, Tyrer J; et al. Telomere length in prospective and retrospective cancer case-control studies. Cancer Res. 2010, 70, 3170–3176. [Google Scholar] [CrossRef] [PubMed]
- Vakonaki, E.; Tsiminikaki, K.; Plaitis, S.; Fragkiadaki, P.; Tsoukalas, D.; Katsikantami, I.; Vaki, G.; Tzatzarakis, M.N.; Spandidos, D.A.; Tsatsakis, A.M. Common mental disorders and association with telomere length (Review). Biomed. Rep. 2018, 8, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.G.; Hall, E.J.; Dhar, S.; Gupta, A.; Rao, P.H.; Pandita, T.K. Telomere stability correlates with longevity of human beings exposed to ionizing radiations. Oncol. Rep. 2003, 10, 1733–1736. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.; Andrew, T.; Gardner, J.; Kimura, M.; Oelsner, E.; Cherkas, L.; Aviv, A.; Spector, T. Obesity, cigarette smoking, and telomere length in women. Lancet 2005, 366, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Yegorov, Y.E.; Poznyak, A.V.; Nikiforov, N.G.; Starodubova, A.V.; Orekhov, A.N. Role of Telomeres Shortening in Atherogenesis: An Overview. Cells 2021, 10, 395. [Google Scholar] [CrossRef] [PubMed]
- Starr, J.M.; McGurn, B.; Harris, S.E.; Whalley, L.J.; Deary, I.J.; Shiels, P.G. Association between telomere length and heart disease in a narrow age cohort of older people. Exp. Gerontol. 2007, 42, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Tsatsakis, A.; Oikonomopoulou, T.; Nikolouzakis, T.K.; Vakonaki, E.; Tzatzarakis, M.; Flamourakis, M.; Renieri, E.; Fragkiadaki, P.; Iliaki, E.; Bachlitzanaki, M.; et al. Role of telomere length in human carcinogenesis (Review). Int. J. Oncol. 2023, 63, 1–24. [Google Scholar] [CrossRef]
- Diotti, R.; Loayza, D. Shelterin complex and associated factors at human telomeres. Nucleus 2011, 2, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Seimiya, H. Revisiting Telomere Shortening in Cancer. Cells 2019, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- O'Sullivan, J.N.; Bronner, M.P.; Brentnall, T.A.; Finley, J.C.; Shen, W.-T.; Emerson, S.; Emond, M.J.; Gollahon, K.A.; Moskovitz, A.H.; Crispin, D.A.; et al. Chromosomal instability in ulcerative colitis is related to telomere shortening. Nat. Genet. 2002, 32, 280–284. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan J, Risques RA, Mandelson MT, Chen L, Brentnall TA, Bronner MP, et al. Telomere Length in the Colon Declines with Age: a Relation to Colorectal Cancer? Cancer Epidemiology, Biomarkers & Prevention. 2006 Mar 1;15(3):573–7.
- Cawthon RM, Smith KR, O’Brien E, Sivatchenko A, Kerber RA. Association between telomere length in blood and mortality in people aged 60 years or older. The Lancet. 2003 Feb;361(9355):393–5.
- Fan HC, Chang FW, Tsai JD, Lin KM, Chen CM, Lin SZ, et al. Telomeres and Cancer. Life. 2021 Dec 16;11(12):1405.
- Gong, P.; Wang, H.; Zhang, J.; Fu, Y.; Zhu, Z.; Wang, J.; Yin, Y.; Wang, H.; Zhou, Z.; Yang, J.; et al. Telomere Maintenance-Associated PML Is a Potential Specific Therapeutic Target of Human Colorectal Cancer. Transl. Oncol. 2019, 12, 1164–1176. [Google Scholar] [CrossRef] [PubMed]
- Kibriya, M.G.; Raza, M.; Kamal, M.; Haq, Z.; Paul, R.; Mareczko, A.; Pierce, B.L.; Ahsan, H.; Jasmine, F. Relative Telomere Length Change in Colorectal Carcinoma and Its Association with Tumor Characteristics, Gene Expression and Microsatellite Instability. Cancers 2022, 14, 2250. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Heaphy, C.M.; Subhawong, A.P.; Hong, S.-M.; Goggins, M.G.; Montgomery, E.A.; Gabrielson, E.; Netto, G.J.; Epstein, J.I.; Lotan, T.L.; Westra, W.H.; et al. Prevalence of the Alternative Lengthening of Telomeres Telomere Maintenance Mechanism in Human Cancer Subtypes. Am. J. Pathol. 2011, 179, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Nikolouzakis, T.K.; Chrysos, E.; Docea, A.O.; Fragkiadaki, P.; Souglakos, J.; Tsiaoussis, J.; Tsatsakis, A. Current and Future Trends of Colorectal Cancer Treatment: Exploring Advances in Immunotherapy. Cancers 2024, 16, 1995. [Google Scholar] [CrossRef] [PubMed]
- Willeit, P.; Willeit, J.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Brandstätter, A.; Kronenberg, F.; Kiechl, S. Telomere Length and Risk of Incident Cancer and Cancer Mortality. JAMA 2010, 304, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Skinner, H.G.; Gangnon, R.E.; Litzelman, K.; Johnson, R.A.; Chari, S.T.; Petersen, G.M.; Boardman, L.A. Telomere Length and Pancreatic Cancer: A Case–Control Study. Cancer Epidemiology Biomarkers Prev. 2012, 21, 2095–2100. [Google Scholar] [CrossRef]
- Tsiaoussis J, Vassilopoulou L, Nikolouzakis T, Rakitskii VN, Vakonaki E, Fragkiadaki P, et al. Biomolecular Profile of Colorectal Cancer - The Role of Telomerase as a Potent Biomarker. Farmacia. 2017 Apr;66(5):643–59.
- Calado RT, Young NS. Telomere diseases. N Engl J Med. 2009 Dec 10;361(24):2353–65.
- Liu, Y.; Lei, T.; Zhang, N.; Zheng, Y.; Kou, P.; Shang, S.; Yang, M. Leukocyte telomere length and risk of gastric cardia adenocarcinoma. Sci. Rep. 2018, 8, 14584. [Google Scholar] [CrossRef] [PubMed]
- Risques, R.A.; Vaughan, T.L.; Li, X.; Odze, R.D.; Blount, P.L.; Ayub, K.; Gallaher, J.L.; Reid, B.J.; Rabinovitch, P.S. Leukocyte Telomere Length Predicts Cancer Risk in Barrett's Esophagus. Cancer Epidemiology Biomarkers Prev. 2007, 16, 2649–2655. [Google Scholar] [CrossRef]
- von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem Sci. 2002 Jul;27(7):339–44.
- Zeng, H.; Wu, H.-C.; Wang, Q.; Yang, H.-I.; Chen, C.-J.; Santella, R.M.; Shen, J. Telomere Length and Risk of Hepatocellular Carcinoma: A Nested Case–Control Study in Taiwan Cancer Screening Program Cohort. Anticancer. Res. 2017, 37, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Han, W.; Xue, W.; Zou, Y.; Xie, C.; Du, J.; Jin, G. The association between telomere length and cancer risk in population studies. Sci. Rep. 2016, 6, 22243. [Google Scholar] [CrossRef] [PubMed]
- Barthel, F.P.; Wei, W.; Tang, M.; Martinez-Ledesma, E.; Hu, X.; Amin, S.B.; Akdemir, K.C.; Seth, S.; Song, X.; Wang, Q.; et al. Systematic analysis of telomere length and somatic alterations in 31 cancer types. Nat. Genet. 2017, 49, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Pickett, H.A. Telomeric replication stress: the beginning and the end for alternative lengthening of telomeres cancers. Open Biol. 2022, 12, 220011. [Google Scholar] [CrossRef]
- Okayasu, I.; Mitomi, H.; Yamashita, K.; Mikami, T.; Fujiwara, M.; Kato, M.; Oshimura, M. Telomerase activity significantly correlates with cell differentiation, proliferation and lymph node metastasis in colorectal carcinomas. J. Cancer Res. Clin. Oncol. 1998, 124, 444–449. [Google Scholar] [CrossRef]
- Yoshida, R.; Kiyozuka, Y.; Ichiyoshi, H.; Senzaki, H.; Takada, H.; Hioki, K.; Tsubura, A. Change in telomerase activity during human colorectal carcinogenesis. . 1999, 19, 2167–72. [Google Scholar]
- Bertorelle, R.; Briarava, M.; Rampazzo, E.; Biasini, L.; Agostini, M.; Maretto, I.; Lonardi, S.; Friso, M.L.; Mescoli, C.; Zagonel, V.; et al. Telomerase is an independent prognostic marker of overall survival in patients with colorectal cancer. Br. J. Cancer 2013, 108, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Hata, T.; Ishida, M.; Motoi, F.; Yamaguchi, T.; Naitoh, T.; Katayose, Y.; Egawa, S.; Unno, M. Telomerase activity in pancreatic juice differentiates pancreatic cancer from chronic pancreatitis: A meta-analysis. Pancreatology 2016, 16, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bao, G.; Huo, T.; Wang, Z.; He, X.; Dong, G. Constitutive telomere length and gastric cancer risk: Case-control analysis in Chinese Han population. Cancer Sci. 2009, 100, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Savage, S.A.; Blaser, M.J.; Perez-Perez, G.; Hoxha, M.; Dioni, L.; Pegoraro, V.; Dong, L.M.; Zatonski, W.; Lissowska, J.; et al. Telomere Length in Peripheral Leukocyte DNA and Gastric Cancer Risk. Cancer Epidemiology Biomarkers Prev. 2009, 18, 3103–3109. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Ajani, J.A.; Chen, M.; Izzo, J.; Lin, J.; Chen, Z.; Gu, J.; Wu, X. Constitutive Short Telomere Length of Chromosome 17p and 12q but not 11q and 2p Is Associated with an Increased Risk for Esophageal Cancer. Cancer Prev. Res. 2009, 2, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Wennerström, E.C.M.; Risques, R.A.; Prunkard, D.; Giffen, C.; Corley, D.A.; Murray, L.J.; Whiteman, D.C.; Wu, A.H.; Bernstein, L.; Ye, W.; et al. Leukocyte telomere length in relation to the risk of Barrett's esophagus and esophageal adenocarcinoma. Cancer Med. 2016, 5, 2657–2665. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Zhang, Y.; Li, X.; Ren, X.; Wang, M.; Tian, S.; Hou, P.; Shi, B.; Yang, Q. Long telomere length predicts poor clinical outcome in esophageal cancer patients. Pathol. - Res. Pr. 2017, 213, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dong, G.; Song, J.; Tan, G.; Wu, X. Telomerase inhibition decreases esophageal squamous carcinoma cell migration and invasion. Oncol. Lett. 2020, 20, 2870–2880. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, M.-Y.; Liang, Y.-R.; Wu, X.-Y. Correlation between expression of human telomerase subunits and telomerase activity in esophageal squamous cell carcinoma. World J. Gastroenterol. 2003, 9, 2395–2399. [Google Scholar] [CrossRef] [PubMed]
- Ikeguchi, M.; Unate, H.; Maeta, M.; Kaibara, N. Detection of telomerase activity in esophageal squamous cell carcinoma and normal esophageal epithelium. Langenbeck's Arch. Surg. 1999, 384, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Takubo, K.; Nakamura, K.-I.; Izumiyama, N.; Mafune, K.-I.; Tanaka, Y.; Miyashita, M.; Sasajima, K.; Kato, M.; Oshimura, M. Telomerase activity in esophageal carcinoma. J. Surg. Oncol. 1997, 66, 88–92. [Google Scholar] [CrossRef]
- Mitsui, A.; Kuwabara, Y.; Iwase, H.; Mitani, M.; Shinoda, N.; Sato, A.; Toyama, T.; Sugiura, M.; Suzuki, T.; Kato, J.; et al. Telomerase activity in esophageal squamous cell carcinoma: Down-regulation by chemotherapeutic agent. J. Surg. Oncol. 2001, 79, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, Y.; Zhang, L.; Ma, J.-L.; Zhou, T.; Li, Z.-X.; Liu, W.-D.; Li, W.-Q.; Deng, D.-J.; You, W.-C.; et al. Telomere Length of Circulating Cell-Free DNA and Gastric Cancer in a Chinese Population at High-Risk. Front. Oncol. 2019, 9, 1434. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Koh, W.-P.; Jin, A.; Wang, R.; Yuan, J.-M. Telomere length and risk of developing gastric adenocarcinoma: The Singapore Chinese Health Study. Gastric Cancer 2017, 21, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Cesare, A.J.; Reddel, R.R. Alternative lengthening of telomeres: models, mechanisms and implications. Nat. Rev. Genet. 2010, 11, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Zhang, Q.; Mei, L.; Liu, X.; Yang, W.; Yu, J. Telomere shortening occurs early during gastrocarcinogenesis. Med Oncol. 2011, 29, 893–898. [Google Scholar] [CrossRef] [PubMed]
- Katayama, S.; Shiota, G.; Oshimura, M.; Kawasaki, H. Clinical usefulness of telomerase activity and telomere length in the preoperative diagnosis of gastric and colorectal cancer. J. Cancer Res. Clin. Oncol. 1999, 125, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Rathi, A.; Hur, K.; Gazdar, A.F.; Bae, J.-S.; Jang, J.-J.; Kim, D.-Y. Telomerase RNA expression during progression of gastric cancer. Hum. Pathol. 1999, 30, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wu, H.; Zhang, S.; Yuan, H.; Cao, L. Clinical Significance of Telomerase Activity in Gastric Carcinoma and Peritoneal Dissemination. J. Int. Med Res. 2009, 37, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Svinareva L, V. , Glukhov AI, Zimnik O V., Bykov II, Khorobrykh T V., Shvets VI. The study of telomerase activity in gastric cancer. Biochem Mosc Suppl B Biomed Chem. 2011 Jun 24;5(2):188–92.
- Pascua, I.; Fernández-Marcelo, T.; Sánchez-Pernaute, A.; de Juan, C.; Head, J.; Torres-García, A.-J.; Iniesta, P. Prognostic value of telomere function in gastric cancers with and without microsatellite instability. Eur. J. Gastroenterol. Hepatol. 2015, 27, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, I.; Bhat, G.R.; Rah, B.; Besina, S.; Zahoor, S.; Wani, M.A.; Shah, M.A.; Bashir, S.; Farooq, M.; Rather, R.A.; et al. Telomere Attrition With Concomitant hTERT Overexpression Involved in the Progression of Gastric Cancer May Have Prognostic and Clinical Implications in High-Risk Population Group From North India. Front. Oncol. 2022, 12, 919351. [Google Scholar] [CrossRef] [PubMed]
- Rufer, N.; Brümmendorf, T.H.; Kolvraa, S.; Bischoff, C.; Christensen, K.; Wadsworth, L.; Schulzer, M.; Lansdorp, P.M. Telomere Fluorescence Measurements in Granulocytes and T Lymphocyte Subsets Point to a High Turnover of Hematopoietic Stem Cells and Memory T Cells in Early Childhood. J. Exp. Med. 1999, 190, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, U.; Griese, E.-U.; Schwab, M.; Fritz, P.; Thon, K.-P.; Klotz, U. Telomere length in different tissues of elderly patients. Mech. Ageing Dev. 2000, 119, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Fern, L.; Pallis, M.; Carter, G.I.; Seedhouse, C.; Russell, N.; Byrne, J. Clonal haemopoiesis may occur after conventional chemotherapy and is associated with accelerated telomere shortening and defects in the NQO1 pathway; possible mechanisms leading to an increased risk of t-AML/MDS. Br. J. Haematol. 2004, 126, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Barnes, R.P.; Fouquerel, E.; Opresko, P.L. The impact of oxidative DNA damage and stress on telomere homeostasis. Mech. Ageing Dev. 2019, 177, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.B.; Plotnikov, E.Y.; Silachev, D.N.; Zorova, L.D.; Pevzner, I.B.; Zorov, S.D.; Babenko, V.A.; Jankauskas, S.S.; Popkov, V.A.; Savina, P.S. Microbiota and mitobiota. Putting an equal sign between mitochondria and bacteria. Biochem. (Moscow) 2014, 79, 1017–1031. [Google Scholar] [CrossRef] [PubMed]
- Aeby, E.; Ahmed, W.; Redon, S.; Simanis, V.; Lingner, J. Peroxiredoxin 1 Protects Telomeres from Oxidative Damage and Preserves Telomeric DNA for Extension by Telomerase. Cell Rep. 2016, 17, 3107–3114. [Google Scholar] [CrossRef] [PubMed]
- Al Bander, Z.; Nitert, M.D.; Mousa, A.; Naderpoor, N. The Gut Microbiota and Inflammation: An Overview. Int. J. Environ. Res. Public Heal. 2020, 17, 7618. [Google Scholar] [CrossRef] [PubMed]
- Zeng MY, Inohara N, Nuñez G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017 Jan;10(1):18–26.
- Rivière, A.; Gagnon, M.; Weckx, S.; Roy, D.; De Vuyst, L. Mutual Cross-Feeding Interactions between Bifidobacterium longum subsp. longum NCC2705 and Eubacterium rectale ATCC 33656 Explain the Bifidogenic and Butyrogenic Effects of Arabinoxylan Oligosaccharides. Appl. Environ. Microbiol. 2015, 81, 7767–7781. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Guo, H.-L.; Deng, X.; Zhu, T.-T.; Xiong, J.-F.; Xu, Y.-H.; Xu, Y. Short-Chain Fatty Acids Inhibit Oxidative Stress and Inflammation in Mesangial Cells Induced by High Glucose and Lipopolysaccharide. Exp. Clin. Endocrinol. Diabetes 2017, 125, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Raynaud, C.M.; Jang, S.J.; Nuciforo, P.; Lantuejoul, S.; Brambilla, E.; Mounier, N.; Olaussen, K.A.; André, F.; Morat, L.; Sabatier, L.; et al. Telomere shortening is correlated with the DNA damage response and telomeric protein down-regulation in colorectal preneoplastic lesions. Ann. Oncol. 2008, 19, 1875–1881. [Google Scholar] [CrossRef]
- Garcia-Aranda C, de Juan C, Diaz-Lopez A, Sanchez-Pernaute A, Torres A, Diaz-Rubio E, et al. Correlations of telomere length, telomerase activity, and telomeric-repeat binding factor 1 expression in colorectal carcinoma. Cancer. 2006 Feb 30;106(3):541–51.
- Gertler, R.; Rosenberg, R.; Stricker, D.; Friederichs, J.; Hoos, A.; Werner, M.; Ulm, K.; Holzmann, B.; Nekarda, H.; Siewert, J.-R. Telomere Length and Human Telomerase Reverse Transcriptase Expression As Markers for Progression and Prognosis of Colorectal Carcinoma. J. Clin. Oncol. 2004, 22, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Tatsumoto, N.; Hiyama, E.; Murakami, Y.; Imamura, Y.; Shay, J.W.; Matsuura, Y.; Yokoyama, T. High telomerase activity is an independent prognostic indicator of poor outcome in colorectal cancer. . 2000, 6, 2696–701. [Google Scholar] [PubMed]
- Engelhardt, M.; Drullinsky, P.; Guillem, J.; A Moore, M. Telomerase and telomere length in the development and progression of premalignant lesions to colorectal cancer. . 1997, 3, 1931–41. [Google Scholar] [PubMed]
- Engelhardt, M.; Albanell, J.; Drullinsky, P.; Han, W.; Guillem, J.; I Scher, H.; Reuter, V.; A Moore, M. Relative contribution of normal and neoplastic cells determines telomerase activity and telomere length in primary cancers of the prostate, colon, and sarcoma. . 1997, 3, 1849–57. [Google Scholar] [PubMed]
- Hastie, N.D.; Dempster, M.; Dunlop, M.G.; Thompson, A.M.; Green, D.K.; Allshire, R.C. Telomere reduction in human colorectal carcinoma and with ageing. Nature 1990, 346, 866–868. [Google Scholar] [CrossRef] [PubMed]
- Zee RYL, Castonguay AJ, Barton NS, Buring JE. Mean telomere length and risk of incident colorectal carcinoma: a prospective, nested case-control approach. Cancer Epidemiol Biomarkers Prev. 2009 Aug;18(8):2280–2.
- Li F-Y, Lai M-D. Colorectal cancer, one entity or three. J Zhejiang Univ Sci B. 2009 Mar 8;10(3):219–29.
- Fernández-Marcelo T, Sánchez-Pernaute A, Pascua I, De Juan C, Head J, Torres-García AJ, et al. Clinical Relevance of Telomere Status and Telomerase Activity in Colorectal Cancer. PLoS One. 2016 Feb 25;11(2): e0149626.
- Rampazzo, E.; Bertorelle, R.; Serra, L.; Terrin, L.; Candiotto, C.; Pucciarelli, S.; Del Bianco, P.; Nitti, D.; De Rossi, A. Relationship between telomere shortening, genetic instability, and site of tumour origin in colorectal cancers. Br. J. Cancer 2010, 102, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Zöchmeister, C.; Brezina, S.; Hofer, P.; Baierl, A.; Bergmann, M.M.; Bachleitner-Hofmann, T.; Karner-Hanusch, J.; Stift, A.; Gerger, A.; Leeb, G.; et al. Leukocyte telomere length throughout the continuum of colorectal carcinogenesis. Oncotarget 2018, 9, 13582–13592. [Google Scholar] [CrossRef] [PubMed]
- Peacock, S.D.; Massey, T.E.; Vanner, S.J.; King, W.D. Telomere length in the colon is related to colorectal adenoma prevalence. PLOS ONE 2018, 13, e0205697. [Google Scholar] [CrossRef] [PubMed]
- Bertorelle, R.; Rampazzo, E.; Pucciarelli, S.; Nitti, D.; De Rossi, A. Telomeres, telomerase and colorectal cancer. World J. Gastroenterol. 2014, 20, 1940–1950. [Google Scholar] [CrossRef] [PubMed]
- Nikolouzakis, T.K.; Vassilopoulou, L.; Fragkiadaki, P.; Sapsakos, T.M.; Papadakis, G.Z.; Spandidos, D.A.; Tsatsakis, A.M.; Tsiaoussis, J. Improving diagnosis, prognosis and prediction by using biomarkers in CRC patients (Review). Oncol. Rep. 2018, 39, 2455–2472. [Google Scholar] [CrossRef] [PubMed]
- Kroupa, M.; Rachakonda, S.K.; Liska, V.; Srinivas, N.; Urbanova, M.; Jiraskova, K.; Schneiderova, M.; Vycital, O.; Vymetalkova, V.; Vodickova, L.; et al. Relationship of telomere length in colorectal cancer patients with cancer phenotype and patient prognosis. Br. J. Cancer 2019, 121, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Kroupa, M.; Kubecek, O.; Tomasova, K.; Hanak, P.; Krupova, M.; Cervena, K.; Siskova, A.; Rosendorf, J.; Hosek, P.; Vodickova, L.; et al. The dynamics of telomere length in primary and metastatic colorectal cancer lesions. Sci. Rep. 2023, 13, 1–10. [Google Scholar] [CrossRef]
- Ye, X.; Li, J.; Song, C.; Chen, W. Telomere in colorectal cancer associated with distant metastases and predicted a poor prognosis. Transl. Cancer Res. 2021, 10, 2906–2917. [Google Scholar] [CrossRef] [PubMed]
- der Stroth, L.I.; Tharehalli, U.; Günes, C.; Lechel, A. Telomeres and Telomerase in the Development of Liver Cancer. Cancers 2020, 12, 2048. [Google Scholar] [CrossRef] [PubMed]
- Kitada, T.; Seki, S.; Kawakita, N.; Kuroki, T.; Monna, T. Telomere Shortening in Chronic Liver Diseases. Biochem. Biophys. Res. Commun. 1995, 211, 33–39. [Google Scholar] [CrossRef]
- Urabe Y, Nouso K, Higashi T, Nakatsukasa H, Hino N, Ashida K, et al. Telomere length in human liver diseases. Liver. 1996 Oct 10;16(5):293–7.
- Cheng, Y.; Yu, C.; Huang, M.; Du, F.; Song, C.; Ma, Z.; Zhai, X.; Yang, Y.; Liu, J.; Bei, J.-X.; et al. Genetic association of telomere length with hepatocellular carcinoma risk: A Mendelian randomization analysis. Cancer Epidemiology 2017, 50, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.-J.; Wang, X.-Y.; Duan, M.; Liu, L.-Z.; Shi, J.-Y.; Dong, L.-Q.; Yang, L.-X.; Wang, Z.-C.; Ding, Z.-B.; Ke, A.-W.; et al. Telomere length variation in tumor cells and cancer-associated fibroblasts: potential biomarker for hepatocellular carcinoma. J. Pathol. 2017, 243, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.; Sun, Y.; Saddozai, U.A.K.; Hayyat, S.; Munir, M.U.; Akbar, M.U.; Khawar, M.B.; Ren, Z.; Ji, X.; Khan, M.I.U. Circulating tumor DNA and its role in detection, prognosis and therapeutics of hepatocellular carcinoma. Chin. J. Cancer Res. 2024, 36, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Choi, J.Y.; Byun, B.H.; Cho, C.H.; Kim, H.S.; Kim, B.S. Telomerase is strongly activated in hepatocellular carcinoma but not in chronic hepatitis and cirrhosis. Exp. Mol. Med. 1998, 30, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, J.-I.; Tahara, H.; Tahara, E.; Saito, M.; Ito, K.; Nakamura, H.; Nakanishi, T.; Nakanishi, E.; Ide, T.; Ishikawa, F. Telomerase activation by hTRT in human normal fibroblasts and hepatocellular carcinomas. Nat. Genet. 1998, 18, 65–68. [Google Scholar] [CrossRef]
- Satyanarayana, A.; Manns, M.P.; Rudolph, K.L. Telomeres and telomerase: A dual role in hepatocarcinogenesis. Hepatology 2004, 40, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Kubota, K.; Takayama, T.; Makuuchi, M. Telomerase activity as a predictive marker for recurrence of hepatocellular carcinoma after hepatectomy. Am. J. Surg. 2001, 181, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi T, Sugawara Y, Shi YZ, Makuuchi M. Telomerase Expression and p53 Status in Hepatocellular Carcinoma. American Journal of Gastroenterology. 2002 Dec;97(12):3166–71.
- Duell, E.J. Telomere length and pancreatic cancer risk: breaking down the evidence. Gut 2016, 66, 1–1. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.M.; Major, J.M.; Cawthon, R.; Weinstein, S.J.; Virtamo, J.; Lan, Q.; Rothman, N.; Albanes, D.; Stolzenberg-Solomon, R.Z. A prospective analysis of telomere length and pancreatic cancer in the alpha-tocopherol beta-carotene cancer (ATBC) prevention study. Int. J. Cancer 2013, 133, 2672–2680. [Google Scholar] [CrossRef] [PubMed]
- Luu, H.N.; Huang, J.Y.; Wang, R.; Adams-Haduch, J.; Jin, A.; Koh, W.-P.; Yuan, J.-M. Association between leukocyte telomere length and the risk of pancreatic cancer: Findings from a prospective study. PLOS ONE 2019, 14, e0221697. [Google Scholar] [CrossRef] [PubMed]
- Campa D, Mergarten B, De Vivo I, Boutron-Ruault MC, Racine A, Severi G, et al. Leukocyte Telomere Length in Relation to Pancreatic Cancer Risk: A Prospective Study. Cancer Epidemiology, Biomarkers & Prevention. 2014 Nov 1;23(11):2447–54.
- Mizumoto, K.; Suehara, N.; Muta, T.; Kitajima, S.; Hamasaki, N.; Tominaga, Y.; Shimura, H.; Tanaka, M. Semi-quantitative analysis of telomerase in pancreatic ductal adenocarcinoma. J. Gastroenterol. 1996, 31, 894–897. [Google Scholar] [CrossRef]
- Hiyama, E.; Kodama, T.; Shinbara, K.; Iwao, T.; Itoh, M.; Hiyama, K.; Shay, J.W.; Matsuura, Y.; Yokoyama, T. Telomerase activity is detected in pancreatic cancer but not in benign tumors. . 1997, 57, 326–31. [Google Scholar] [PubMed]
- Inoue, H.; Tsuchida, A.; Kawasaki, Y.; Fujimoto, Y.; Yamasaki, S.; Kajiyama, G. Preoperative diagnosis of intraductal papillary-mucinous tumors of the pancreas with attention to telomerase activity. Cancer 2001, 91, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Murakami, Y.; Uemura, K.; Hayashidani, Y.; Sudo, T.; Hashimoto, Y.; Ohge, H.; Oda, M.C.; Sueda, T.; Hiyama, E. Usefulness of Human Telomerase Reverse Transcriptase in Pancreatic Juice as a Biomarker of Pancreatic Malignancy. Pancreas 2009, 38, 527–533. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Murakami, Y.; Uemura, K.; Hayashidani, Y.; Sudo, T.; Ohge, H.; Fukuda, E.; Sueda, T.; Hiyama, E. Detection of human telomerase reverse transcriptase (hTERT) expression in tissue and pancreatic juice from pancreatic cancer. Surgery 2008, 143, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Srinivasan, R.; Vasishta, R.K.; Wig, J.D. Positive Regulation of Human Telomerase Reverse Transcriptase Gene Expression and Telomerase Activity by DNA Methylation in Pancreatic Cancer. Ann. Surg. Oncol. 2009, 16, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Telomere Lengthening by Telomerase. “Created with Biorender.com”.
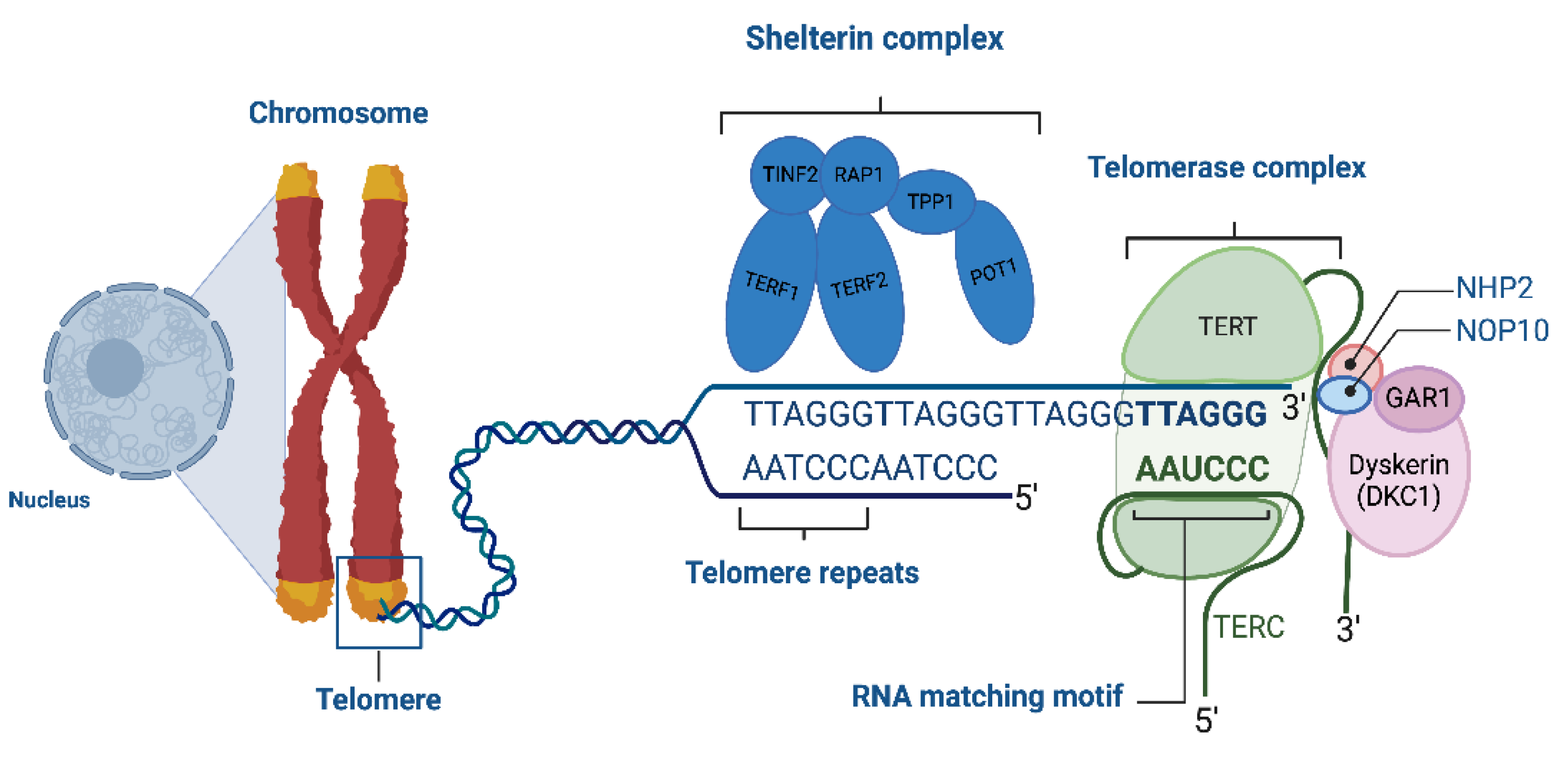
Figure 2.
Telomerase Activity in Healthy Somatic Cells vs. Cancer Cells. Telomerase, an enzyme essential for maintaining telomere length, comprises the TERT subunit, which acts as a reverse transcriptase, and the TERC subunit, which provides a template for telomere extension. In healthy somatic cells, TERT is typically suppressed to prevent unchecked cell division, leading to cellular aging and programmed cell death. In contrast, cancer cells often reactivate telomerase, enabling continuous proliferation and the accumulation of genetic mutations. “Created with Biorender.com”.
Figure 2.
Telomerase Activity in Healthy Somatic Cells vs. Cancer Cells. Telomerase, an enzyme essential for maintaining telomere length, comprises the TERT subunit, which acts as a reverse transcriptase, and the TERC subunit, which provides a template for telomere extension. In healthy somatic cells, TERT is typically suppressed to prevent unchecked cell division, leading to cellular aging and programmed cell death. In contrast, cancer cells often reactivate telomerase, enabling continuous proliferation and the accumulation of genetic mutations. “Created with Biorender.com”.
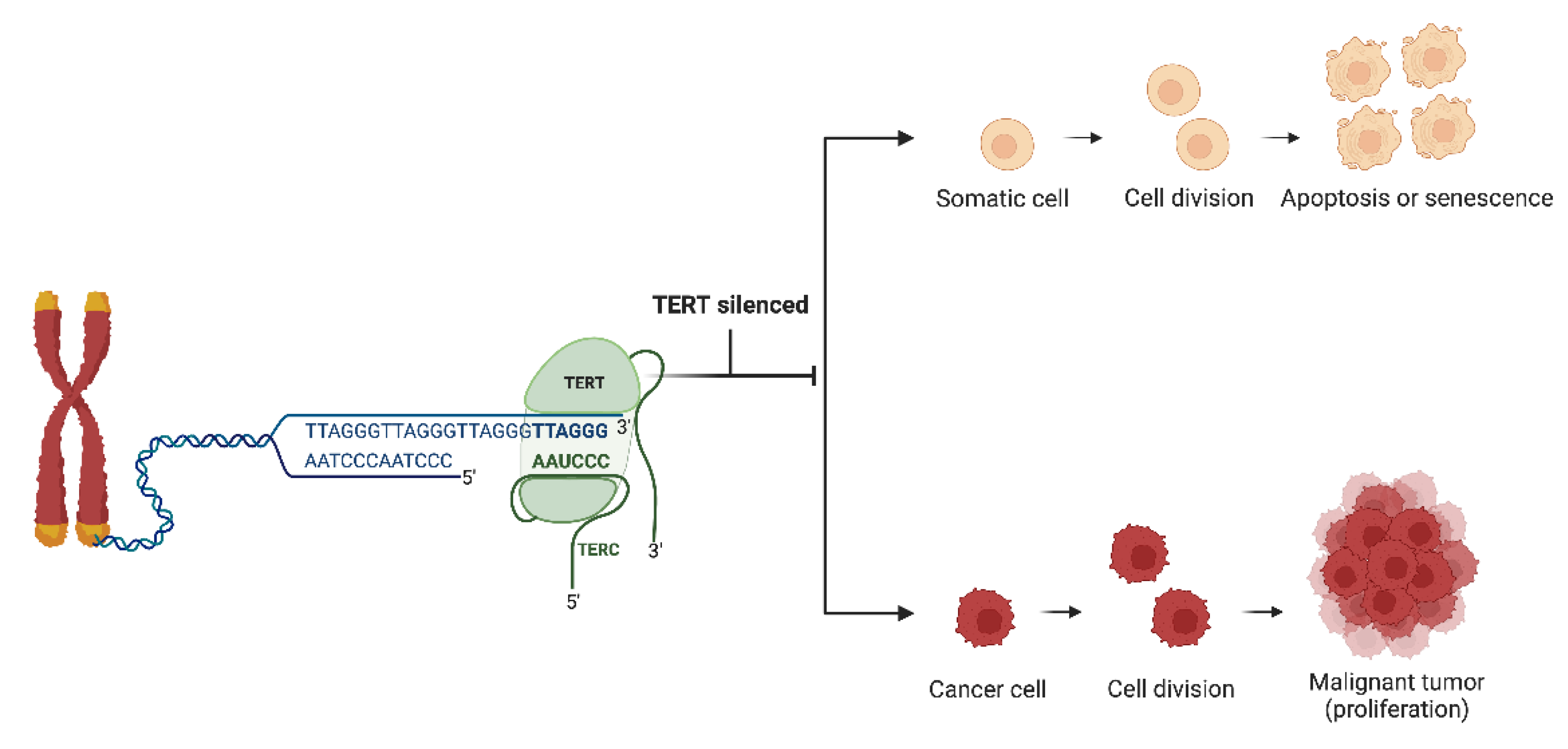

Table 1.
Studies investigating the association between TL, TA and gastrointestinal cancer.
| Tumor site | References | |
|---|---|---|
| Telomere length | Telomerase activity | |
| Stomach | Liu et al. 2018 [38] Liu et al. 2009 [49] Hou et al. 2009 [50] Wang et al. 2018 [60] Shi et al. 2019 [59] Wentzensen et al. 2011 [11] Cesare et al. 2010 [61] Mushtaq et al. 2022 [68] |
Katayama et al. 1999 [63] Rathi et al. [64] Hu et al. [65] Svinareva et al. [66] |
| Esophagus | Wentzensen et al. 2011 [11] Risques et al. 2007 [39] Xing et al. 2009 [51] Wennerström et al. 2016 [52] O’Sullivan et al. 2006 [26] Zeng et al. 2017 [41] Lv et al. 2017 [53] Li et al. 2020 [54] |
Ikeguchi et al. [56] Takubo et al. [57] Mitsui et al. [58] |
| Colorectum | Pooley et al. 2010 [16] Hastie et al. 1990 [85] Engelhardt et al. 1997 [83,84] Tatsumoto et al. 2000 [82] Gertler et al. 2004 [81] Garcia-Aranda et al. 2006 [80] Raynaud et al. 2008 [79] O’Sullivan et al. 2006 [26] Rampazzo et al. 2010 [89] Wennerström et al. 2016 [52] Zöchmeister et al. 2018 [90] Zee et al. 2009 [86] Li and Lai 2009 [87] Fernández-Marcelo et al. 2016 [88] Peacock et al. 2018 [91] Nikolouzakis et al. 2018 [93] Kroupa et al. [94] Kroupa et al. [95] Ye et al. [96] |
Engelhardt et al. 1997 [83,84] Tatsumoto et al. 2000 [82] Bertorelle 2013 [92] Katayama et al. 1999 [63] Garcia-Aranda et al. 2006 [80] Nikolouzakis et al. 2024 [33] |
| Liver | Urabe et al. 1996 [99] Ma et al. 2017 [101] Zeng et al. 2017 [41] Cheng et al. 2017 [100] Rashid et al. 2024 [102] Tsatsakis et al. 2023 [22] |
Park et al. 1998 [103] Nakayama et al. 1998 [104] Satyanarayana et al. 2004 [105] Kobayashi et al. 2001 [106] Kobayashi et al. 2002 [107] |
| Pancreas | Skinner et al. 2012 [35] Campa et al. 2019 [111] Duell 2017 [108] Luu et al. 2019 [110] Campa et al. 2014 [111] Lynch et al. 2013 [109] Zeng et al. 2017 [41] Ma et al. 2017 [101] |
Lynch et al. 2013 [109] Luu et al. 2019 [110] Campa et al. 2014 [111] Mizumoto et al. 1996 [112] Hiyama et al. 1997 [113] Inoue et al. 2001 [114] Nakashima et al. 2009 [115] Hashimoto et al. 2008 [116] Kumari et al. 2009 [117] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



