
Submitted:
13 August 2024
Posted:
15 August 2024
You are already at the latest version
Abstract
Mitochondria are dynamic organelles that can adjust and respond to different stimuli within a cell. This plastic ability allows them to effectively coordinate several cellular functions in cells and becomes particularly relevant in highly complex cells such as neurons. An imbalance in mitochondrial dynamics can disrupt mitochondrial function, leading to abnormal cellular function and ultimately to a range of diseases, including neurodegenerative disorders.
Regulation of mRNA transport and local translation inside neurons is crucial for maintaining the proteome of distal mitochondria, which is vital for energy production and synaptic function. A significant portion of the axonal transcriptome is dedicated to mRNAs for mitochondrial proteins, emphasizing the importance of local translation in sustaining mitochondrial function in areas far from the cell body.
In neurons, local translation and the regulation of mRNAs encoding mitochondrial-shaping proteins could be essential for synaptic plasticity and neuronal health. The dynamics of these mRNAs, including their transport and local translation, may influence the morphology and function of mitochondria, thereby affecting the overall energy status and responsiveness of synapses. Comprehending the mitochondria-related mRNAs regulation and local translation, as well as its influence on mitochondrial morphology near the synapses will help to better understand neuronal physiology and neurological diseases where mitochondrial dysfunction and impaired synaptic plasticity play a central role.
Keywords:
Mitochondria
; mitochondrial morphology
; mRNA
; mRNA trafficking
; neuron
; synapse
1. Intro: The Role of Mitochondria in Neurons
The brain’s energy consumption is quite high, accounting for approximately 20% of the body’s total metabolic energy, despite comprising only approximately 2% of its total mass. Neurons are responsible for most of this energy use, primarily to restore ion gradients across the plasma membrane, which are altered by the creation of action potentials and synaptic transmission [1]. Neurons contain numerous mitochondria, which play a crucial role in maintaining neuronal integrity and function under normal conditions, particularly at the synapse [2], namely serving a vital function in energy buffering [3,4].
Mitochondria display distinct morphologies and distributions in axons and dendrites. For example, in pyramidal neurons, the main excitatory neuronal subtype in the cerebral cortex, dendritic mitochondria display long and tubular shapes, forming a complex network filling 70–80% of the dendritic arbor and frequently close to spines [5] (Figure 1). In contrast, axonal mitochondria display a remarkably standard size, are small and punctate, and occupy less than 10% of the axonal volume. In neurons, similar to any given cell type, mitochondrial morphology is mostly controlled by fusion and fission mechanisms, collectively termed mitochondrial dynamics [6] (Figure 2). Therefore, it is hypothesized that the striking differences in mitochondrial morphology between axons and dendrites are the result of a high level of fission in the axon and a high degree of fusion in dendrites. Indeed, fission-dependent regulation of presynaptic mitochondrial size, as well as axon branching, has already been shown in cortical pyramidal neurons [6,7].
Alterations in mitochondrial morphology and function affect neuronal physiology and synaptic transmission, positioning mitochondria as neuromodulators [4] (Figure 3). Specifically at the pre-synaptic terminal, mitochondria are essential for neurotransmitter release by supplying ATP and buffering local calcium (Ca2+) content [8]. Moreover, synaptic vesicle turnover and release are significantly affected when mitochondrial function is inhibited and ATP is depleted [9]. The density and rate of mitochondrial oxidative phosphorylation can affect presynaptic ATP regeneration, and consequently, synaptic vesicle exocytosis [9]. In the post-synaptic compartment, a recent study in C. elegans has shown that mitochondrial ROS signalling regulates excitatory synapse function by altering the recruitment of glutamate receptors, thereby integrating several mitochondrial functions from energy production to Ca2+ buffering and ROS signalling [10].
Electron microscopy has been instrumental in understanding the anatomy and ultrastructure of mitochondria at nanoscale resolution since the first electron micrograph of the mitochondria [11]. Furthermore, it has become widely appreciated that mitochondrial structure is tightly linked to its functional state. This has been shown in studies comparing the ultrastructure of mitochondria in different types of axons, which showed that presynaptic mitochondria in axons from neurons with high activity are larger and contain more densely packed lamellar cristae than those in low-activity neurons [12]. Using electron microscopy, mitochondria in the pre- and post-synaptic compartments of hippocampal neurons were compared, and the results showed that the pre-synaptic mitochondria were typically smaller and darker than the post-synaptic mitochondria [13]. Since 2D snapshots did not allow to fully investigate mitochondria in the complex morphology of axons and dendrites, studies have been conducted to reconstruct in 3D the mitochondrial network in the mouse brain. Authors found that the form of mitochondria varies between different subcellular compartments, and that mitochondria in axons and dendrites generally have distinct shapes, being the pre-synaptic mitochondria generally smaller and shorter and not extending beyond pre-synaptic terminals, while most of the post-synaptic mitochondria are larger and spread through significant portions of the dendrites [14,15]. Besides the compartment-specific differences in mitochondrial morphology, there are also age-related morphological alterations that define the status of the neuronal mitochondrial network [16].
The formation of synaptic mitochondria occurs mostly within the cell body of the neuron, and these mitochondria are transported along axons or dendrites by specific motor proteins that travel along microtubule tracks. Notably, it has been demonstrated that synapse formation promotes the bidirectional transport of mitochondria [17]. Furthermore, it has been argued that synaptic mitochondria may be longer lived than the mitochondria present in the soma of neurons or glial cells, presenting greater cumulative damage from oxidative stress. Accordingly, it was shown that synaptic mitochondria exhibited increased age-associated mtDNA mutations as well as decreased bioenergetic function compared with non-synaptic mitochondria [18,19]. The specificity of synaptic bioenergetics and the synapse-related mitochondrial heterogeneity is not very well understood, but some considerations about the differences in synaptic mitochondria when compared to non-synaptic counterparts have been reviewed in [20]. A more recent study, disregarding the synaptic versus non-synaptic difference though, has shown that some mitochondrial long-lived proteins (LLPs) can persist for months in tissues harbouring long-lived cells, as in the case of brain and neurons [21]. Although some evidence might conflict with the mRNA transport and translation of mitochondrial proteins in neurons (see section below), research has revealed that some long-lasting mitochondrial proteins play a crucial role in regulating the structure of mitochondria in neurons and other long-lived cells. Indeed, Graham et al. reported that discrete proteomic profiles exist dependent upon mitochondrial subcellular localization (synaptic vs. non-synaptic mitochondria) and selective alteration of intrinsic mitochondrial proteins alters synaptic morphology in vivo [22].
Additionally, although not mentioning mitochondria directly, another group found that synapses with varying protein lifespans are unevenly distributed in neurons and different regions of the brain [23]. However, the mechanisms underlying the different properties of synaptic and nonsynaptic mitochondria remain poorly understood. It has been suggested that the more limited ability of synaptic mitochondria to buffer the [Ca2+]i may be due to the punctuate morphology, differently from the tubular mitochondrial morphology observed in other regions of neurons [24]. Mitochondria grouped at the synapses constitute a discrete pool from their non-synaptic counterparts, exhibiting distinct morphological, proteomic, enzymatic, and Ca2+ handling properties [25]. The unique characteristics of the synaptic compartment are likely regulated by the activity within it. Synaptic activity causes a significant increase in Ca2+ influx, further requesting a prompt supply of ATP. Consequently, the synaptic mitochondrial pool may have enhanced its ability to buffer Ca2+ in order to more effectively support synaptic functions.
2. Mitochondrial Morphology and Dynamics
Mitochondria are dynamic organelles that change morphology and intracellular distribution responding to heterogeneous external cues, associated with varied degrees of cellular energetic needs [26]. They undergo cyclic processes of specialized structural alterations in response to external cues, namely fission and fusion, interacting with each other and forming an innate dynamic process that needs constant equilibrium to maintain the physiological function of the cell [27]. The fusion and fission mechanisms depend on the precise regulation of molecular markers. Mitochondria have a double-membrane structure, encompassing an outer mitochondrial membrane (MOM) and an inner mitochondrial membrane (MIM), divided by an intermembrane space (IMS) (Figure 2).
- a)
- Fusion
Mitochondrial fusion is a mechanism described as the close contact tethering and merging of two mitochondria, and the posterior emergence of a unitary larger organelle from the regulated fusion of the membrane of mitochondria. Two major proteins are responsible for the processing of said mechanism, Mitofusin 1 and 2 (Mfn1 and Mfn2) are large GTPases, responsible for the fusing of MOMs from different mitochondria, stabilizing the interaction of the mitochondria in contact by enabling the interaction and rearrangement of MIMs [28]. Then, optic atrophy type 1 protein (OPA1), a mechano-chemical GTPase, physically adapts the inner membrane, granting a functional restructuring of this lipidic structure with the contacted mitochondrion, whilst maintaining the protein content and functionality of this relevant membrane of mitochondrial innate capacity to metabolically support cellular function [29]. Moreover, OPA1 promotes the reshaping of the internal mitochondrial cristae, enabling the maintenance of its function [30]. Fusion is important to synthesize ATP because the fusion of healthy mitochondria leads to an increase in the cristae area, imposing considerably higher pumping of H+, which is processed via ATPases in higher amounts, thereby increasing energy production. In this way, mitochondria support demanding cellular processes such as apoptosis and mitotic progression [31].
- b)
- Fission
Fission is a mitochondrial mechanism that counteracts fusion. It is described as the separation of mitochondrial membranes, mediated by specific molecular players. It is a heterogeneous process within species, but in humans, it is dependent on the endoplasmic reticulum (ER). The ER tags the target mitochondria by interacting with membrane lipids and stimulating the formation of actin bands near the fission site. During this actin rings’ assembly in a putative context, Dynamin-related protein 1 (DRP1) gets activated by phosphorylation and can interact with mitochondrial receptors present at the MOM such as Mitochondrial fission factor (MFF) [32,33]. This interaction is also mediated by regulators such as Mitochondrial fission 1 (Fis1), Mitochondrial dynamics protein 49 and 51 (MiD 49 and MiD51), and MFF itself, receptors of DRP1 [34]. This process of actin band strangling, with the addition of DRP1 GTPase activity on MOM regulated by MFF, leads to the constraining and eventual division of the mitochondrial membrane, forming separate organelles. Fission is thought to be highly important in the synaptic context as it might facilitate mitochondrial transport, boosting the production of energy in confined spaces with decreased accessibility [4]. Besides that, fission is also extremely important to mitochondrial division, enabling the formation of new mitochondria [35].
3. Mitochondrial Morphology in the Synapse
Mitochondria have distinct and equally relevant roles, namely oxidative, metabolic, and Ca2+ buffering functions. Therefore, it’s easily understood that mitochondria operate to regulate and modify synaptic transmission and also related processes of functional and structural plasticity [9,25,36,37,38].
A connection between alterations of mitochondrial function and synaptic activity has been reported long ago for peripheral synapses [39]. Also, some pharmacological studies have shown that inhibiting mitochondrial activity impairs synaptic potentiation and neurotransmission [40,41]. Additionally, acutely blocking mitochondrial function during intense stimulation leads to depressed synaptic transmission [42]. Interestingly, a couple of decades ago Li et al. reported that enhancing mitochondrial respiration can increase the number and plasticity of spines and synapses [43]. Extension or movement of mitochondria into dendritic protrusions correlates with the development and morphological plasticity of spines, whereas molecular manipulations of DRP1 and OPA1 proteins, reducing dendritic mitochondria content, lead to loss of synapses and dendritic spines. Thus, dendritic mitochondria have been found to be crucial for the support of synapses. Reciprocally, synaptic activity modulates the motility and fusion/fission balance of mitochondria and controls mitochondrial distribution in the dendrites. These findings indicate that mitochondria not only respond to synaptic activity in various ways but are also able to regulate synaptic plasticity [44]. Indeed, a recent study from Thomas et al. has shown that post-synaptic mitochondria are critical for the development, plasticity, and maintenance of synaptic inputs in cortical neurons [45]. This study reported that synapses are larger and exhibit greater selectivity to visual stimuli in spines with a mitochondrion in the head or neck, suggesting that mitochondria support the structurally and functionally diverse inputs innervating the basal dendrites. Mitochondrial fusion had already been shown to be necessary for synapse formation in human induced pluripotent stem cells-derived cortical neurons [46]. Mfn2 was demonstrated to be essential for human mitochondrial development in neuronal maturation and differentiation, and Mfn2 knock-down impacted neurogenesis and synapse formation. More recently, Kochan et al. reported that mitochondrial fusion promotes elongated mitochondria in dendrites of new neurons, and that Mfn1 or Mfn2 absence abrogate synaptic plasticity in hippocampal neurons [47]. Moreover, it has been recently shown in vivo that a local regulation of fusion-fission balance in hippocampus is responsible for the distinct mitochondrial morphology in dendrites. Interestingly, Virga et al. have shown that mitochondria within the dendritic arbor display a further degree of subcellular, layer-specific distinct morphology in hippocampus [48]. In the dendrites of these neurons, mitochondria morphology ranges from highly fused and elongated in the apical tuft, to more fragmented in the apical oblique and basal dendritic compartments. Additionally, the authors demonstrated that the regulation of this compartment-specific mitochondrial morphology in dendrites requires the activation of AMPK and its ability to phosphorylate two direct effectors: MFF and the anti-fusion, OPA1-inhibiting protein, mitochondrial fission regulator 1-like protein (Mtfr1l).
While the significance of mitochondrial dynamics is well-documented, the importance of mitochondrial fission is less well-defined compared to that of mitochondrial fusion (Figure 3). Primary cultures of neural cells derived from mice lacking DRP1 exhibited a reduced number of neurites and defective synapse formation, emphasizing the crucial role of DRP1-mediated mitochondrial fission in the highly polarized cells such as neurons [49]. Moreover, the loss of DRP1 also impairs synaptic vesicle release [50]. This further supports the idea that presynaptic mitochondria play a key role in exocytosis. In accordance, it has also been reported that the ATP supply provided by presynaptic mitochondria is essential for fuelling the assembly of the actin cytoskeleton [51]. Additionally, the ATP content impacts the clustering of synaptic vesicles and mitochondria at the synapse [52], as well as the efficient mobilization of synaptic vesicles into the readily releasable pool [53]. Furthermore, mitochondria offer a new level of control over exocytosis by taking up Ca2+ from the cytosol in response to neuronal activity. This activity causes significant increases in intracellular Ca2+ levels in the presynaptic compartment, which act locally to facilitate the exocytosis of synaptic vesicles and influence numerous signalling pathways. Mitochondria regulate these processes by controlling [Ca2+]i levels through their negative membrane potential, which is created by the electron transport chain (ETC), plasma membrane Ca2+-ATPases, and the ER [54]. Nevertheless, mitochondria are not found in all pre-synapses, although their importance in maintaining neuronal homeostasis has been increasingly recognized (reviewed in [55]). In fact, the maintenance of a pool of presynaptic mitochondria was shown to increase the stability of presynaptic strength by maintaining the local ATP homeostasis [56].
The potential for synaptic mitochondria to be exposed to extensive fluctuations in [Ca2+]i may increase the risk of oxidative stress and Ca2+ accumulative damage [57]. Furthermore, when the ETC complex I is inhibited, synaptic mitochondria exhibit a more significant reduction in respiration rates and ATP production compared to non-synaptic mitochondria [58]. These findings indicate the necessity of tailoring the mitochondrial population, specifically for synapses, to maintain regular neuronal function. This also highlights the importance of maintaining a proper mitochondrial proteome in different brain regions and neuron types (excitatory versus inhibitory neurons). Additionally, recent reports have suggested a functional cross-talk between energy sensing and mitochondria anchoring, particularly at the pre-synapse level [59]. Synaptic activity triggers AMP-activated protein kinase (AMPK) activation within the axon, followed by stimulation of the p21-activated kinase (PAK) to recruit mitochondria. The AMPK-PAK signalling pathway results in the phosphorylation of myosin VI, promoting mitochondrial recruitment and anchoring on presynaptic terminals. This maintains presynaptic energy supply and Ca2+ clearance during intense synaptic activity. Postsynaptic mitochondria are also important for local functions, as experimentally disrupting dendritic mitochondria results in the loss of dendritic spines and their synapses [60]. Consequently, it is widely accepted that mitochondrial dysfunction and subsequent synaptic dysfunction contribute to a variety of neurological defects [61].
Dendritic spines in the cerebral cortex and hippocampus of adult brains generally lack the presence of mitochondria [14], and similar observations were reported for cultured hippocampal neurons [43]; although, there are exceptions such as CA3 neurons [62] and olfactory bulb granule cells [63] that frequently contain mitochondria. Researchers have found that depolarization of cultured hippocampal neurons with KCl increases the ratio of fission versus fusion and enhances the distribution of mitochondria in dendritic spines [43]. The activity-dependent recruitment of mitochondria into spines is regulated by Wiskott-Aldrich syndrome protein (WASP)-family verprolin homologous protein 1 (WAVE1), a key regulator of actin polymerization that acts through the activation of the Arp2/3 complex [64].
Several studies have also shown that mitochondrial transport along dendrites is regulated by neuronal activity. For instance, during development of the primary visual cortex, mitochondrial motility is correlated with neuronal activity. Specifically, during the first two postnatal weeks, mitochondria motility within the dendritic compartment decreases with the increase in the frequency of neuronal activity [65]. The transport of mitochondria along dendrites of cultured cerebrocortical and hippocampal neurons is also arrested following stimulation of NMDA receptors, which are coupled to the influx of Ca2+ [66,67,68]. This Ca2+-induced immobilization of mitochondria near the synapse may provide the energy necessary to cope with the increased demand resulting from glutamatergic synapse activity. Accordingly, spines depleted of mitochondria show an impairment in plasticity mechanisms and in translation activity [8,69]. During long-term activity changes, the localization of these synaptic mitochondria can actually change; particularly, the localization of mitochondria at pre-synaptic sites can be regulated by a mechanism dependent on the Ca2+-sensing function of the mitochondrial trafficking protein, Miro1 [70]. By strategically positioning mitochondria near the synapse, Miro1 helps neurons fine-tune the strength of their Ca2+ signals in response to sustained activity changes, supporting a model in which mitochondria are recruited to presynaptic terminals during periods of raised neuronal activity and are involved in rescaling synaptic signals during homeostatic plasticity. The effect of neuronal activity on the distribution of mitochondria has an impact on neurotransmitter release since presynaptic Ca2+ signals are significantly decreased in terminals containing mitochondria, with a consequent downregulation of exocytosis [59,71]. This process, called activity-dependent synaptic repositioning, suggests that high activity attracts mitochondria to the synapse. This dynamic control of mitochondrial distribution is crucial for maintaining a healthy balance in communication between brain cells.
Alterations in mitochondrial dynamics have been investigated upon induction of synaptic plasticity in cultured hippocampal neurons incubated in a Na+-salt solution lacking Mg2+ and supplemented with glycine [72]. This chemical long-term potentiation (cLTP) protocol is dependent on NMDA receptor activation, which enhances spinal volume and upregulates AMPA receptor surface expression. Under these conditions, rapid upregulation of dendritic mitochondrial fission was observed, accompanied by an increase in mitochondrial matrix Ca2+ content. The fission of dendritic mitochondria was triggered by an increase in [Ca2+]i and mediated by Ca2+- and calmodulin-dependent protein kinase II (CaMKII) activation. Furthermore, cLTP-induced mitochondrial fission was mediated by DRP1 and dynamin 2. Notably, inhibition of fission prevented structural alterations in spines, and complementary studies using hippocampal slices showed a role for mitochondrial fission in high-frequency presynaptic stimulation-induced LTP of CA1 synapses [72]. These results are consistent with evidence demonstrating impaired spatial working memory in mice with a DRP1 deletion in postmitotic adult mouse forebrain neurons [73]. A key role of mitochondrial fission in synaptic regulation was also demonstrated by the results obtained in cultured hippocampal neurons transfected with a dominant negative form of DRP1 (DRP1-K38A). Expression of this construct decreased the number of mitochondria in dendrites and in spines, reduced the number of dendritic spines, and blocked activity-induced increase in postsynaptic puncta detected in immunocytochemistry experiments [43]. Moreover, research conducted in mice with a postnatal deletion of DRP1 revealed a decrease in the production of ATP in axons, accompanied by an impairment in synaptic transmission and hippocampus-dependent memory [74]. Notably, the knockdown of DRP1 in D1-medium spiny neurons of the nucleus accumbens prevented cocaine-seeking behaviour after self-administration of the drug [75]. PTEN-induced kinase 1 (PINK1) is a crucial factor in the activation of DRP1 through the phosphorylation of Ser616, which promotes synaptic development and plasticity [76]. In this study, contextual fear memory and spatial memory were found to be impaired in DRP1S616A knock-in mice, highlighting the significance of this phosphorylation site on DRP1 in learning and memory [76].
4. Mitochondrial Morphology: A Role in Disease
Since synapses have high energy demands, and neurons may rely on alternative ways to produce the energy necessary to sustain the presynaptic vesicle cycle, neuronal mitochondria are essential for the maintenance of neurotransmission and synaptic plasticity. Consequently, dysregulation of neuronal mitochondrial function and also mitochondrial morphology have been implicated in neurodegenerative diseases and other neurological disorders [61].
Neurodegenerative diseases, including Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington’s disease (HD), and amyotrophic lateral sclerosis (ALS), are characterised by the loss of selective neuron subtypes in the CNS. Mitochondrial dynamics-related proteins, often part of the so-called mitochondrial quality control (MQC) mechanisms, play a significant role in the progression of these diseases. Considering AD as example, mitochondrial fission and fusion proteins are disturbed in the hippocampus in several AD animal models and patients. Specifically, the fission protein Fis1 is increased, while the fusion proteins Mfn1, Mfn2, and OPA1 are decreased [77]. Additionally, the phosphorylation of DRP1 at Ser616 is higher in the brains of AD patients [77]. Furthermore, it has been shown in vivo that the functional regulation of DRP1 by phosphorylation mediated by PINK1 modulates synaptic development and plasticity, further connecting mitochondrial dynamics and neural circuitry formation and refinement [76]. Another study in vivo found significant mitochondrial fragmentation and irregular distribution of mitochondria in the pyramidal neurons of the brains of mice with AD, before the accumulation of amyloid plaques [78]. By using the mitochondrial division inhibitor-1 (mdivi-1), a pharmacological inhibitor of DRP1, the mitochondrial fragmentation and distribution issues, as well as mitochondrial function, were resolved. Moreover, Mdivi-1 led to a decrease in extracellular amyloid deposits, prevented cognitive decline, and increased cortical levels of synaptophysin [78].
In an animal model of traumatic brain injury (TBI), DRP1-dependent mitochondrial alterations have been associated with intense memory impairments [79]. DRP1 levels in purified hippocampal mitochondria were increased in animals with TBI compared to sham controls. Additionally, cryo-electron micrographs of these mitochondria showed that TBI resulted in an initial increase in the length of hippocampal mitochondria 24 h post-injury, followed by a significant decrease in mitochondrial length 72 h post-injury. Post-TBI administration of Mdivi-1 prevented this decrease in mitochondrial length. Mdivi-1 treatment also reduced the loss of newborn neurons in the hippocampus and improved novel object recognition (NOR) memory and context-specific fear memory [79]. These findings indicate that TBI increases mitochondrial fission, and that inhibition of fission improves hippocampal-dependent learning and memory, suggesting that strategies to reduce mitochondrial fission may have translational value after injury.
Ischemic stroke is another major cause of death and disability of the nervous system. Although reperfusion through revascularization has reduced the mortality rate of ischemic stroke, it also leads to additional damage to the brain tissue, known as ischemia-reperfusion (I/R) injury. Several processes related to mitochondrial quality control (MQC) have been suggested to contribute to I/R damage. During cerebral I/R injury, mitochondrial fission and mitophagy help to remove damaged mitochondria and promote mitochondrial apoptosis of neuron cells [80,81]. Overexpression of Sirtuin 3 (Sirt3) can inhibit mitochondrial fission and attenuate cerebral ischemia-reperfusion injury by triggering pro-survival signals in neurons subjected to I/R injury [82].
Epilepsy is characterized by recurrent seizures that affect neurons without provocation. Research suggests that mitochondrial stress and MQC play a role in the development of this condition. Studies have shown that the levels of OPA1, Mfn2, MFF, and DRP1 are elevated in a mouse model of epilepsy [83]. Additionally, the serine peptidase LONP1, which is also related to MQC, is upregulated in the mitochondria during status epilepticus (SE), and its knockdown has been shown to increase SE-induced mitochondrial apoptosis in neurons [84].
Mitochondrial quality control-related gene mutations or abnormal expression have also been reported to be involved in several brain dysplasias [85]. Heterozygous or de novo mutations in dnm1l gene, coding for DRP1 protein, have been shown to cause microcephaly, optic atrophy, hypoplasia, epileptic encephalopathy and neurodevelopment delay [86,87,88]. Specifically regarding mitochondrial morphology proteins (also reviewed in [61]), the overexpression of Mfn2 mutation in cultured neurons has been reported to disrupt axonal mitochondrial positioning and to promote axon degeneration [89]. Moreover, in a neuron-specific knockout mouse model it has been shown that Mfn2 is required for dendritic outgrowth, axonal projection, and survival [90,91]. In retinal ganglion cells (RGCs), it has been shown that OPA1 can protect neurons from excitotoxicity [92]. OPA1 loss resulted in mitochondrial fragmentation, decreased buffering of cytosolic Ca2+ and sensitization of RGCs to excitotoxic injury. Similarly, the overexpression of pathogenic OPA1 mutants induced mitochondrial fragmentation and reduced motility of mitochondria in neurites of RGCs, and accumulation of cytosolic Ca2+ in proximity of the axonal hillock [93,94]. Additionally, Koch et al. established that MFF loss of function disturbs mitochondrial and peroxisomal dynamics and this leads to early-onset Leigh-like basal ganglia disease [95].
5. RNA-Binding Proteins and Nuclear-Encoded Mitochondrial Transcripts
A significant proportion of the total transcriptome in axons consists of mRNAs that encode for mitochondrial proteins, accounting for approximately one third of the total. Following this, 22% of the transcriptome is composed of signal transduction-related mRNAs, while 5% are for cytoskeleton proteins and 3% are for neurodevelopment-related proteins [96]. The translation of mRNAs encoding proteins for mitochondrial ribosomes, protein import, and respiration has been measured in hippocampal neurites and retinal axon terminals [97,98], providing support for the hypothesis that local translation rejuvenates mitochondria in distal regions away from the cell body, which in turn provide the energy required for the translation of synaptic proteins [8]. To maintain the proteome of distal mitochondria, two key factors are necessary: the availability of mRNA and translation.
- a)
- Mechanisms of mRNA transport
Mitochondria synthesise a small fraction of their proteins from mitochondrial DNA because more than 99% of their proteins are nuclear encoded. These mRNAs (NEM-mRNAs) require a complex system to be transported effectively in polarised cells like neurons. One mechanism of transport is mediated by transport granules, which form membrane-less organelles by liquid-liquid phase separation [99]. There, mRNAs are bound and stabilized by RNA binding proteins (RBPs) and other components important to regulate the content and the (dys-)assembly of the granules for local translation. Interestingly, the blockage of the RNA-binding properties of FUS and TDP-43 mislocalizes them and alters the survival of neurons, suggesting that the correct localization of RBPs may depend on their binding to respective target mRNAs [100,101]. Inhibition of transcription mislocalizes TDP-43 and FUS [102,103], suggesting an additional mechanism of regulation based on the abundance of mRNA. Another mechanism seems to be related to the activity and the binding properties of VCP to RBPs in motor neurons from ALS patients [104]. ALS mutations can also shift FUS to bind mRNAs encoding mitochondrial respiratory chain components [105] or increase the binding of TDP-43 to NEM-mRNAs such as Cox4i1 and Atp5a1, reducing their translation in axons [106]. Transport granules traffic using the molecular motors of microtubules, common to organelles, via direct binding to specific adaptors in anterograde direction [107,108]. However, the adaptors for dynein-mediated retrograde movement are still unknown [109].
The untranslated regions (UTRs) of mRNAs are determinants of the localisation: for example, longer 3’-UTRs are most likely to localise in neurites whereas short 3’-UTRs reside in the cell body. Interestingly, transcripts with longer 3’-UTRs were found enriched in dendrites after depolarization [110]. The same mRNAs can also be differentially targeted using the 3’-UTR or the 5’-UTR (reviewed in [99]). Cis-regulatory elements (or zipcodes) are motifs found in the 3’-UTR of mRNAs, which are bound by RBPs that target mRNAs to their destination. For example, ZBP1 binds β-actin mRNA using a zipcode of 54 bases at the 3’-UTR and transports β-actin from the perinuclear space to the periphery of the cell repressing the translation [111,112]. As it was difficult to identify conserved sequences for zipcodes in many studies, it has been proposed that additional mechanisms exist to regulate the localization of mRNAs. In a recent study, Loedige et al. [113] demonstrated that mRNA stability is necessary and sufficient to localise mRNAs in axons, following the principle that mRNAs with a short half-life are less likely to travel distally. The stabilisation is regulated by a combination of stabilising (5′ terminal oligopyrimidine tracts and codon optimality) and destabilising parameters (including m6A modifications and AU-rich elements). In general, mRNAs localised in neurites have fewer destabilising elements and relocalise to the soma if they become more stable. Authors propose to use these elements as a prediction of mRNA localization in neurites.
A second mechanism of transport tethers mRNAs to organelles to exploit their bi-directional and saltatory movements, which might be advantageous for responding to rapid signalling stimuli (Figure 4). Docking of organelles is an additional mechanism for regulating their movement (for a review, [114]) and might represent another type of regulation for mRNA motility to investigate. Furthermore, the tethering of mRNAs to organelles also increases the general speed of transport in respect to naked mRNAs [115,116]. Although these measurements could be distinctive and advantageous for travel of mRNAs in general [116], there are exceptions. For example, a minor fraction of Pink1 is not co-transported with mitochondria [115]. This slow-moving Pink1 traffics around one third slower than Pink1-tethered to mitochondria, raising the hypothesis that organelles provide a support for fast-mediated transport of mRNAs whereas independent mRNAs move slower locally. To date, mRNAs have been shown to traffic together with mitochondria, endosomes, and lysosomes. Cox7a is the first transcript described to associate and co-traffic with mitochondria in axons using the coding region [117]. Recently, Pink1 has also been shown to co-traffic with mitochondria using two adaptors: the phosphatidylinositol phosphatase Synaptojanin 2 (SYNJ2) contains an RNA-binding motif and Synaptojanin 2 binding protein (SYNJ2BP) links SYNJ2 to the outer mitochondrial membrane [115]. The association of Pink1 to mitochondria is regulated by the phosphorylation of SYNJ2 at S21 by AMPK [118]. The presence of an RBP in neurites could have different purposes than transport, as stabilising or translating NEM-mRNAs [119] and large datasets of NEM-mRNAs co-associated and transported to mitochondria by different RBPs in neurites and synapses have not been generated. Conversely, a molecular tether and relative mRNA targets have been described for early endosomes. The FERRY complex is composed of Tbck, Ppp1r21, C12orf4, Cryzl1, and Gatd1 proteins, and it tethers NEM-mRNAs to early endosomes through interaction with Rab5 [120]. This complex binds mRNAs encoding components of the mitochondrial matrix, mitochondrial ribosome, cellular respiration, and tricarboxylic acid cycle in HeLa cells. Some of its targets (mdh2 and uchl1) have been validated by FISH to be localised to early endosomes in cell bodies and neurites. Additionally, Splicing Factor Proline and glutamine rich (SFPQ) assembles and traffics Lmnb2 and Bcl2l2 in RNA granules in axons, which colocalize with ribosomes in proximity of mitochondria [121]. SFPQ co-immunoprecipitates with Rab7 and is localised to Rab7a-positive endosomes in axons [116]. An interesting candidate to transport NEM-mRNAs is the BLOC-one-related complex (BORC), which tethers mRNAs to lysosomes in axons [122]. The authors investigated solely the trafficking of Rps7 and Rps27A mRNAs, despite BORC-KO axons being depleted of mRNAs encoding both ribosomal and OXPHOS proteins. The absence of the BORC complex also reduces the translation of these mRNAs, but it is unclear whether this effect is directly mediated by BORC’s transport of NEM-mRNAs or is secondary to the lack of ribosomes.
- b)
- Translation in axons and dendrites
Growth cones, which are the tips of axons devoted to their growth, sense extracellular cues to direct the extension of axons during development. The presence of a local, autonomous translation in neuronal compartments has been shown in experiments where severed neurites attached to the plate stopped responding to chemotropic cues in presence of protein synthesis inhibitors [123]. This so-called local protein synthesis in synapses has been induced in primary neurons using poly-D-lysine coated beads, which trigger the clustering of presynaptic proteins and the formation of pre-synapses. Specifically, these studies showed the local translation of Snap25 and β-catenin [124,125]. Translation in dendrites occurs at 50% near synapses and it is correlated with spontaneous Ca2+ activity following spine stimulation by glutamate uncaging [126]. The majority of studies focused on translation initiation because some of the first evidence regarded the recruitment of initiation factors and activation of LPS in spines in presence of BDNF for long-term potentiation [127,128]. The initiation of translation is regulated by the formation of the 43S pre-initiation complex, which is recruited by the eIF4F complex to form the initiation complex that scans the 5’-UTR until the starting codon (for a review, [129]). The eIF4F complex is composed of the cap-binding protein eIF4E, the RNA helicase eIF4A and the scaffolding protein eIF4G. In absence of exogenous stimuli, G3BP1 interacts with eIF4E and inhibits translation, regulating the genesis of mushroom spines [130]. Upon depolarization, eIF4G2 has been found to bind the 5’-UTR of dendritic transcripts, increasing the ribosome occupancy in uORFs and CDS translation [110] (Figure 4). Of note, Timm23 (the core component of the TIM23 complex that imports proteins across the inner mitochondrial membrane [131]) and Nsun3 (a matrix-localised methyltransferase regulating mitochondrial translation [132]) were more translated in stimulated dendrites. Another study showed that translation may also be resumed on polyribosomes, which are stalled in unstimulated spines, for metabotropic glutamate receptor long-term depression independently of translation initiation [133]. Authors propose that this regulation is more rapid and transcriptionally effective to respond to stimuli with respect to translation initiation.
The organisation of ribosomes in spines is another mechanism to regulate translation (Figure 4). Indeed, ribosome dynamics and activity are subjected to rapid regulation since assembly and translation of ribosomal subunits occurs in seconds after BDNF administration [134]. In isolated dendrites, ribosomal RNAs associate with existing ribosomes to promote ribogenesis and potentially potentiate the synthesis of dendritic proteins [135]. Authors reported also that addition of H2O2 reduced the synthesis of new ribosomal subunits but increased the incorporation of a small subset of ribosomal proteins. This observation suggests that the physiological state of dendrites modulates the composition of ribosomes but also that probably mitochondrial dysfunction (namely oxidative stress) could signal a reshuffling of ribosomal subunits. The hypothesis from these data have not been investigated in this work but it will be interesting to measure if and how the local translatome changes according to different mitochondrial dysfunctions and possibly composition of ribosomes. Ribosomes appeared to be organised in polysomes in electron micrographs of stimulated spines [136,137]. However, an interesting study highlighted that ribosomes are differently organised in neurons, with a prevalence of polysomes in cell bodies and monosomes in neurites [138]. Monosomes – previously considered translationally inactive – probably couple the limited space available in spines to the necessity of translating a plethora of proteins, including preferentially synaptic proteins but also NEM-proteins. As we mentioned in the previous paragraph, NEM-mRNAs are transported together with organelles [116]. Additionally, 70% of the proteins interacting with the FERRY complex are ribosomal subunits, suggesting that FERRY could directly link entire ribosomes to early endosomes [120]. Specifically, ribosomes also co-traffic with these mRNAs forming supercomplexes on organelles devoted as translation platforms to supply new proteins to axonal mitochondria. In developing axons of RGCs, local translation of lmnb2 was measured on Rab7a-positive endosomes paused in contact with mitochondria.
Several lines of evidence suggest that mitochondria are heterogeneous in neurons. In primary neurons, mitochondria decrease their membrane potential and increase reactive oxygen species production with the distance to the soma [139]. Interestingly, the same distal mitochondria imported a mitochondrial reporter slower than perinuclear mitochondria. Moreover, synaptic mitochondria have a different Ca2+ buffering capacity and membrane potential than non-synaptic mitochondria extracted from mouse brains [18,140] and the bioenergetic properties of these mitochondria are prone to decline with age [18,140,141]. A common shared hypothesis is that mitochondria accumulate damage when they traffic to reach distal areas, a process that in humans can last even days [142]. Mitochondria are normally positioned to LPS sites in different neuronal compartments, where they support protein synthesis generating ATP (Figure 4). For instance, the respiration of stalled mitochondria along the axons induces the maturation of filopodia into branches by generating sites of LPS [143] and reduction of the mitochondrial mass by PGC1α inhibits spinogenesis and synaptogenesis [144]. In dendritic spines, the inhibition of mitochondrial function using antimycin or oligomycin decreased newly synthesised proteins [8]. This effect was also observed in single-stimulated spines depleted of functional mitochondria, suggesting that mitochondria are strategically positioned close to spines to sustain LPS during plasticity. In the same study, MFF induced fragmentation of mitochondria in spines and reduced protein synthesis in stimulated spines, pointing to the importance of the morphology of mitochondria for effective translation. LPS also fosters inter-organelle communication. Following axotomy, the glucose regulated protein 75 (Grp75) was translated at the site of injury and increased the tethering of ER and mitochondria, increasing the levels of ATP. Hence, the generation of ATP is controlled locally and promotes the regeneration of severed axons [145]. Interestingly, the ER is also found to be localised near mitochondria at sites of branching [143].
6. Translation and mRNA Regulation of Mitochondria-Shaping Proteins in Synapses
The knowledge about the dynamics of mRNAs encoding for mitochondria-shaping proteins in dendritic spines is limited. The half-life of these transcripts ranges approximately from 3 to 6 hours compatible with other mRNAs found in neurites. These transcripts have few destabilising elements (less than 15 AREs, and Opa1 and Mff have also few m6A modifications), suggesting the presence of mechanisms of transport and local translation [113]. Indeed, Opa1 and Dnm1l (which encodes for DRP1) have been found associated with monosomes in the neuropil of the hippocampus, similarly to the translation pattern of synaptic transcripts [146].
mRNAs encoding for mitochondria-shaping proteins have been hardly detected in axons of motoneurons because they seem to be predominantly enriched in soma and dendrites [147]. Conversely, they are highly enriched in optic nerves [97], raising the question if these mRNAs have different mechanisms of transport and translation among different types of neurons. In support of this hypothesis, the abundance of these mRNA in synaptosomes changes depending on the transcript, with Opa1, Mfn2, Fis1 and Dnm1l as the mostly enriched [148]. Data about the protein content in spines is scarce and conflicting probably due to the limited amount of these proteins. Indeed, several mitochondria-shaping proteins have not been measured in screenings from synaptosomes or were detected only in synaptosomes from specific neuronal populations [149,150]. Interestingly, Jung et al. combined axon-TRAP with metabolic labelling of RNAs to measure the translatome in axons of RGCs [97]. Authors report that mRNAs encoding for mitochondria-shaping proteins in axons are differentially translated in axons independently from having a pro-fusion/fission function. Another observation has been recently reported in dendritic spines [110]. In this study, authors mapped the translatome using a construct expressing TurboID tagged with the postsynaptic protein PSD95, showing that NEM-mRNAs are translationally silent in resting conditions and the translation initiation factor eIF4G2 is required to translate dendritic NEM-mRNAs upon depolarization. However, the translation of Opa1, Mfn1/2 and Dnm1l was not altered. It is possible that other stimuli control the abundance of these proteins, as it has been shown for Pink1, which associates to mitochondria after insulin administration [118].
Up to date, there are no known RBPs regulating the transport or the translation of mRNAs encoding for mitochondria-shaping proteins in dendritic spines. An interesting candidate to regulate fusion and fission proteins in neurons is Clustered mitochondria homolog (CLUH). CLUH binds NEM-mRNAs and components of the translation apparatus, and it prevents peripheral neuropathy [119,151,152]. Among its targets, CLUH binds Opa1, Mff and MiD49 mRNAs in HeLa cells [152,153], regulating the translation of MFF and MiD49 in the cytosol or near mitochondria. Furthermore, CLUH recruits DRP1 to mitochondria to promote fission. The control of CLUH on these targets might be restricted to their translation because the ablation of CLUH does not impact the movement of two of its targets Atp5a1 and Mdh2 in axons. In motoneurons, CLUH is localised in cell bodies and growth cones, and it is essential for axonal growth and translation [153]. However, no data are available about dendrites due to the limited arborisations of these neurons. Another candidate to consider for mitochondrial morphology might be the FERRY complex, which seems to reside in neurites and spines [120]. The FERRY complex does not co-precipitate with mRNAs encoding for mitochondria-shaping proteins but the knocking-out of single FERRY subunits reduces the levels of these mRNAs. Since the FERRY complex associates to ribosomes, it might have an indirect but local effect on the translation of mRNAs related to mitochondria-shaping proteins.
7. Conclusions
The regulation of mRNA transport and local translation within neurons is essential for maintaining the proteome of distal mitochondria, which is crucial for energy production and synaptic function. A significant portion of the axonal transcriptome is dedicated to mRNAs for mitochondrial proteins, highlighting the importance of local translation in sustaining mitochondrial function in regions far from the cell body. This process relies on transport mechanisms involving RBPs and organelles like mitochondria, endosomes, and lysosomes, which ensure the precise localization and translation of these mRNAs. Factors such as untranslated regions (UTRs), cis-regulatory elements, and mRNA stability play key roles in this regulation.
In dendrites, local translation and the regulation of mRNAs encoding mitochondrial-shaping proteins could be crucial for synaptic plasticity and neuronal health. The dynamics of these mRNAs, including their transport and local translation, may influence the morphology and function of mitochondria, thereby affecting the overall energy status and responsiveness of synapses. However, the knowledge about the specific mechanisms regulating these transcripts in dendrites remains limited. The presence of mitochondria-shaping protein mRNAs in synaptosomes suggest a local regulation. For instance, extracellular cues, which trigger local translation initiation and ribosome reorganization, could induce the rapid synthesis of these proteins at synapses. However, data on the dynamics and local translation of mRNAs for mitochondrial-shaping proteins in dendrites are scarce and further research is necessary to fully elucidate the stimuli and pathways regulating the translation of these proteins.Understanding these regulatory processes is vital for comprehending how neurons maintain their function and adaptability, especially under conditions of stress or injury. Such insights could reveal new therapeutic targets for neurological diseases where mitochondrial dysfunction and impaired synaptic plasticity play a central role.
Author Contributions
All authors contributed to the preparation of the manuscript.
Funding
The work was supported by grants from the Portuguese Science and Technology Foundation (FCT), the European Regional Development Fund (ERDF), through the Centro 2020 Regional Operational Program (under project CENTRO-01–0145- FEDER-000008:BrainHealth 2020), the Operational Programme for Competitiveness and Internationalization—COMPETE 2020 grant (PTDC/MED-NEU/3736/2020 [to F.V.D.] and UIDB/04539/2020).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rossi, M.J.; Pekkurnaz, G. Powerhouse of the mind: mitochondrial plasticity at the synapse. Curr. Opin. Neurobiol. 2019, 57, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.J.; Jolivet, R.; Attwell, D. Synaptic Energy Use and Supply. Neuron 2012, 75, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Garcia, G.C.; Bartol, T.M.; Phan, S.; Bushong, E.A.; Perkins, G.; Sejnowski, T.J.; Ellisman, M.H.; Skupin, A. Mitochondrial morphology provides a mechanism for energy buffering at synapses. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Duarte, F.V.; Ciampi, D.; Duarte, C.B. Mitochondria as central hubs in synaptic modulation. Cell. Mol. Life Sci. 2023, 80, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.-X.; Tan, X.; Fang, H.; Lau, P.-M.; Wang, X.; Cheng, H.; Bi, G.-Q. Dendritic mitoflash as a putative signal for stabilizing long-term synaptic plasticity. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Lee, A.; Hirabayashi, Y.; Kwon, S.-K.; Lewis, T.L.; Polleux, F. Emerging roles of mitochondria in synaptic transmission and neurodegeneration. Curr. Opin. Physiol. 2018, 3, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T.L.; Kwon, S.-K.; Lee, A.; Shaw, R.; Polleux, F. MFF-dependent mitochondrial fission regulates presynaptic release and axon branching by limiting axonal mitochondria size. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Rangaraju, V.; Lauterbach, M.; Schuman, E.M. Spatially Stable Mitochondrial Compartments Fuel Local Translation during Plasticity. Cell 2019, 176, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Ivannikov, M.V.; Sugimori, M.; Llinás, R.R. Synaptic Vesicle Exocytosis in Hippocampal Synaptosomes Correlates Directly with Total Mitochondrial Volume. J. Mol. Neurosci. 2012, 49, 223–230. [Google Scholar] [CrossRef]
- R. L. Doser, K. M. R. L. Doser, K. M. Knight, E. W. Deihl, and F. J. Hoerndli, “Activity-dependent mitochondrial ROS signaling regulates recruitment of glutamate receptors to synapses,” Elife, vol. 13, Mar. 2024.
- Palade, G.E. The fine structure of mitochondria. Anat. Rec. 1952, 114, 427–451. [Google Scholar] [CrossRef] [PubMed]
- Cserép, C.; Pósfai, B.; Schwarcz, A.D.; Dénes. Mitochondrial Ultrastructure Is Coupled to Synaptic Performance at Axonal Release Sites. eneuro 2018, 5. [Google Scholar] [CrossRef]
- Freeman, D.W.; Petralia, R.S.; Wang, Y.-X.; Mattson, M.P.; Yao, P.J. Mitochondria in hippocampal presynaptic and postsynaptic compartments differ in size as well as intensity. Matters 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Popov, V.; Medvedev, N.I.; Davies, H.A.; Stewart, M.G. Mitochondria form a filamentous reticular network in hippocampal dendrites but are present as discrete bodies in axons: A three-dimensional ultrastructural study. J. Comp. Neurol. 2005, 492, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Delgado, T.; Petralia, R.S.; Freeman, D.W.; Sedlacek, M.; Wang, Y.-X.; Brenowitz, S.D.; Sheu, S.-H.; Gu, J.W.; Kapogiannis, D.; Mattson, M.P.; et al. Comparing 3D ultrastructure of presynaptic and postsynaptic mitochondria. Biol. Open 2019, 8, bio044834. [Google Scholar] [CrossRef]
- J. Faitg et al., “3D neuronal mitochondrial morphology in axons, dendrites, and somata of the aging mouse hippocampus,” Cell Rep., vol. 36, no. 6, p. 109509, Aug. 2021.
- Badal, K.K.; Akhmedov, K.; Lamoureux, P.; Liu, X.-A.; Reich, A.; Fallahi-Sichani, M.; Swarnkar, S.; Miller, K.E.; Puthanveettil, S.V. Synapse Formation Activates a Transcriptional Program for Persistent Enhancement in the Bi-directional Transport of Mitochondria. Cell Rep. 2019, 26, 507–517. [Google Scholar] [CrossRef]
- Brown, M.R.; Sullivan, P.G.; Geddes, J.W. Synaptic Mitochondria Are More Susceptible to Ca2+Overload than Nonsynaptic Mitochondria. J. Biol. Chem. 2006, 281, 11658–11668. [Google Scholar] [CrossRef]
- Stauch, K.L.; Purnell, P.R.; Fox, H.S. Quantitative Proteomics of Synaptic and Nonsynaptic Mitochondria: Insights for Synaptic Mitochondrial Vulnerability. J. Proteome Res. 2014, 13, 2620–2636. [Google Scholar] [CrossRef] [PubMed]
- Fedorovich, S.V.; Waseem, T.V.; Puchkova, L.V. Biogenetic and morphofunctional heterogeneity of mitochondria: the case of synaptic mitochondria. Prog. Neurobiol. 2017, 28, 363–373. [Google Scholar] [CrossRef]
- Bomba-Warczak, E.; Edassery, S.L.; Hark, T.J.; Savas, J.N. Long-lived mitochondrial cristae proteins in mouse heart and brain. J. Cell Biol. 2021, 220. [Google Scholar] [CrossRef]
- Graham, L.C.; Eaton, S.L.; Brunton, P.J.; Atrih, A.; Smith, C.; Lamont, D.J.; Gillingwater, T.H.; Pennetta, G.; Skehel, P.; Wishart, T.M. Proteomic profiling of neuronal mitochondria reveals modulators of synaptic architecture. Mol. Neurodegener. 2017, 12, 1–16. [Google Scholar] [CrossRef]
- Bulovaite, E.; Qiu, Z.; Kratschke, M.; Zgraj, A.; Fricker, D.G.; Tuck, E.J.; Gokhale, R.; Koniaris, B.; Jami, S.A.; Merino-Serrais, P.; et al. A brain atlas of synapse protein lifetime across the mouse lifespan. Neuron 2022, 110, 4057–4073. [Google Scholar] [CrossRef] [PubMed]
- Seager, R.; Lee, L.; Henley, J.M.; Wilkinson, K.A. Mechanisms and roles of mitochondrial localisation and dynamics in neuronal function. Neuronal Signal. 2020, 4, NS20200008. [Google Scholar] [CrossRef]
- Pekkurnaz, G.; Wang, X. Mitochondrial heterogeneity and homeostasis through the lens of a neuron. Nat. Metab. 2022, 4, 802–812. [Google Scholar] [CrossRef]
- J. Bereiter-Hahn and M. Vöth, “Dynamics of mitochondria in living cells: Shape changes, dislocations, fusion, and fission of mitochondria,” Microsc. Res. Tech., vol. 27, no. 3, pp. 198–219, Feb. 1994.
- Protasoni, M.; Zeviani, M. Mitochondrial Structure and Bioenergetics in Normal and Disease Conditions. Int. J. Mol. Sci. 2021, 22, 586. [Google Scholar] [CrossRef]
- G. Chandhok, M. Lazarou, and B. Neumann, “Structure, function, and regulation of mitofusin-2 in health and disease,” Biol. Rev., vol. 93, no. 2, pp. 933–949, May 2018.
- Pernas, L.; Scorrano, L. Mito-Morphosis: Mitochondrial Fusion, Fission, and Cristae Remodeling as Key Mediators of Cellular Function. Annu. Rev. Physiol. 2016, 78, 505–531. [Google Scholar] [CrossRef] [PubMed]
- von der, Malsburg; et al. , “Structural mechanism of mitochondrial membrane remodelling by human OPA1,” Nature, vol. 620, no. 7976, pp. 1101–1108, Aug. 2023.
- Quintana-Cabrera, R.; Scorrano, L. Determinants and outcomes of mitochondrial dynamics. Mol. Cell 2023, 83, 857–876. [Google Scholar] [CrossRef]
- Knott, A.B.; Perkins, G.; Schwarzenbacher, R.; Bossy-Wetzel, E. Mitochondrial fragmentation in neurodegeneration. Nat. Rev. Neurosci. 2008, 9, 505–518. [Google Scholar] [CrossRef]
- Kraus, F.; Roy, K.; Pucadyil, T.J.; Ryan, M.T. Function and regulation of the divisome for mitochondrial fission. Nature 2021, 590, 57–66. [Google Scholar] [CrossRef]
- Atkins, K.; Dasgupta, A.; Chen, K.-H.; Mewburn, J.; Archer, S.L. The role of Drp1 adaptor proteins MiD49 and MiD51 in mitochondrial fission: implications for human disease. Clin. Sci. 2016, 130, 1861–1874. [Google Scholar] [CrossRef]
- Egner, J.M.; Nolden, K.A.; Harwig, M.C.; Bonate, R.P.; De Anda, J.; Tessmer, M.H.; Noey, E.L.; Ihenacho, U.K.; Liu, Z.; Peterson, F.C.; et al. Structural studies of human fission protein FIS1 reveal a dynamic region important for GTPase DRP1 recruitment and mitochondrial fission. J. Biol. Chem. 2022, 298, 102620. [Google Scholar] [CrossRef]
- Su, B.; Ji, Y.-S.; Sun, X.-L.; Liu, X.-H.; Chen, Z.-Y. Brain-derived Neurotrophic Factor (BDNF)-induced Mitochondrial Motility Arrest and Presynaptic Docking Contribute to BDNF-enhanced Synaptic Transmission. J. Biol. Chem. 2014, 289, 1213–1226. [Google Scholar] [CrossRef]
- Cheng, A.; Hou, Y.; Mattson, M.P. Mitochondria and neuroplasticity. ASN Neuro 2010, 2, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Faria-Pereira and V., A. Morais, “Synapses: The Brain’s Energy-Demanding Sites,” Int. J. Mol. Sci., vol. 23, no. 7, p. 3627, Mar. 2022.
- Nguyen, P.V.; Atwood, H.L.; Marin, L. Altered impulse activity modifies synaptic physiology and mitochondria in crayfish phasic motor neurons. J. Neurophysiol. 1994, 72, 2944–2955. [Google Scholar] [CrossRef] [PubMed]
- Alnaes, E.; Rahamimoff, R. On the role of mitochondria in transmitter release from motor nerve terminals. J. Physiol. 1975, 248, 285–306. [Google Scholar] [CrossRef]
- Tang, Y.-G.; Zucker, R.S. Mitochondrial Involvement in Post-Tetanic Potentiation of Synaptic Transmission. Neuron 1997, 18, 483–491. [Google Scholar] [CrossRef]
- Billups, B.; Forsythe, I.D. Presynaptic Mitochondrial Calcium Sequestration Influences Transmission at Mammalian Central Synapses. J. Neurosci. 2002, 22, 5840–5847. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Okamoto, K.-I.; Hayashi, Y.; Sheng, M. The Importance of Dendritic Mitochondria in the Morphogenesis and Plasticity of Spines and Synapses. Cell 2004, 119, 873–887. [Google Scholar] [CrossRef]
- Mattson, M.P. Mitochondrial Regulation of Neuronal Plasticity. Neurochem. Res. 2006, 32, 707–715. [Google Scholar] [CrossRef] [PubMed]
- C. I. Thomas, M. A. C. I. Thomas, M. A. Ryan, N. Kamasawa, and B. Scholl, “Postsynaptic mitochondria are positioned to support functional diversity of dendritic spines,” Elife, vol. 12, pp. 1–17, Dec. 2023.
- Fang, D.; Yan, S.; Yu, Q.; Chen, D.; Yan, S.S. Mfn2 is Required for Mitochondrial Development and Synapse Formation in Human Induced Pluripotent Stem Cells/hiPSC Derived Cortical Neurons. Sci. Rep. 2016, 6, 31462. [Google Scholar] [CrossRef]
- Kochan, S.M.; Malo, M.C.; Jevtic, M.; Jahn-Kelleter, H.M.; Wani, G.A.; Ndoci, K.; Pérez-Revuelta, L.; Gaedke, F.; Schäffner, I.; Lie, D.C.; et al. Enhanced mitochondrial fusion during a critical period of synaptic plasticity in adult-born neurons. Neuron 2024, 112, 1997–2014. [Google Scholar] [CrossRef]
- Virga, D.M.; Hamilton, S.; Osei, B.; Morgan, A.; Kneis, P.; Zamponi, E.; Park, N.J.; Hewitt, V.L.; Zhang, D.; Gonzalez, K.C.; et al. Activity-dependent compartmentalization of dendritic mitochondria morphology through local regulation of fusion-fission balance in neurons in vivo. Nat. Commun. 2024, 15, 1–21. [Google Scholar] [CrossRef]
- Ishihara, N.; Nomura, M.; Jofuku, A.; Kato, H.; Suzuki, S.O.; Masuda, K.; Otera, H.; Nakanishi, Y.; Nonaka, I.; Goto, Y.-I.; et al. Mitochondrial fission factor Drp1 is essential for embryonic development and synapse formation in mice. Nat. Cell Biol. 2009, 11, 958–966. [Google Scholar] [CrossRef]
- Singh, M.; Denny, H.; Smith, C.; Granados, J.; Renden, R. Presynaptic loss of dynamin-related protein 1 impairs synaptic vesicle release and recycling at the mouse calyx of Held. J. Physiol. 2018, 596, 6263–6287. [Google Scholar] [CrossRef]
- Cardanho-Ramos, C.; Faria-Pereira, A.; Morais, V.A. Orchestrating mitochondria in neurons: Cytoskeleton as the conductor. Cytoskeleton 2019, 77, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Peng, H.B. The Function of Mitochondria in Presynaptic Development at the Neuromuscular Junction. Mol. Biol. Cell 2008, 19, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Cai, Q.; Lu, W.; Sheng, Z.-H.; Mochida, S. KIF5B Motor Adaptor Syntabulin Maintains Synaptic Transmission in Sympathetic Neurons. J. Neurosci. 2009, 29, 13019–13029. [Google Scholar] [CrossRef]
- Rizzuto, R. Intracellular Ca2+ pools in neuronal signalling. Curr. Opin. Neurobiol. 2001, 11, 306–311. [Google Scholar] [CrossRef]
- Devine, M.J.; Kittler, J.T. Mitochondria at the neuronal presynapse in health and disease. Nat. Rev. Neurosci. 2018, 19, 63–80. [Google Scholar] [CrossRef]
- Sun, T.; Qiao, H.; Pan, P.-Y.; Chen, Y.; Sheng, Z.-H. Motile Axonal Mitochondria Contribute to the Variability of Presynaptic Strength. Cell Rep. 2013, 4, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Lores-Arnaiz, S.; Bustamante, J. Age-related alterations in mitochondrial physiological parameters and nitric oxide production in synaptic and non-synaptic brain cortex mitochondria. Neuroscience 2011, 188, 117–124. [Google Scholar] [CrossRef]
- Davey, G.P.; Peuchen, S.; Clark, J.B. Energy Thresholds in Brain Mitochondria. J. Biol. Chem. 1998, 273, 12753–12757. [Google Scholar] [CrossRef]
- Li, S.; Xiong, G.-J.; Huang, N.; Sheng, Z.-H. The cross-talk of energy sensing and mitochondrial anchoring sustains synaptic efficacy by maintaining presynaptic metabolism. Nat. Metab. 2020, 2, 1077–1095. [Google Scholar] [CrossRef] [PubMed]
- E. Schuman and D. Chan, “Fueling synapses.,” Cell, vol. 119, no. 6, pp. 738–40, Dec. 2004.
- Chen, W.; Zhao, H.; Li, Y. Mitochondrial dynamics in health and disease: mechanisms and potential targets. Signal Transduct. Target. Ther. 2023, 8, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Chicurel, M.E.; Harris, K.M. Three-dimensional analysis of the structure and composition of CA3 branched dendritic spines and their synaptic relationships with mossy fiber boutons in the rat hippocampus. J. Comp. Neurol. 1992, 325, 169–182. [Google Scholar] [CrossRef]
- Cameron, H.A.; Kaliszewski, C.K.; Greer, C.A. Organization of mitochondria in olfactory bulb granule cell dendritic spines. Synapse 1991, 8, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.Y.; Engmann, O.; Teylan, M.A.; Nairn, A.C.; Greengard, P.; Kim, Y. WAVE1 controls neuronal activity-induced mitochondrial distribution in dendritic spines. Proc. Natl. Acad. Sci. 2008, 105, 3112–3116. [Google Scholar] [CrossRef]
- AP Silva, C.; Yalnizyan-Carson, A.; Busch, M.V.F.; van Zwieten, M.; Verhage, M.; Lohmann, C.; Neurogenomics, C.F. ; Netherlands Activity-dependent regulation of mitochondrial motility in developing cortical dendrites. eLife 2021, 10. [Google Scholar] [CrossRef]
- MacAskill, A.F.; Rinholm, J.E.; Twelvetrees, A.E.; Arancibia-Carcamo, I.L.; Muir, J.; Fransson, A.; Aspenstrom, P.; Attwell, D.; Kittler, J.T. Miro1 Is a Calcium Sensor for Glutamate Receptor-Dependent Localization of Mitochondria at Synapses. Neuron 2009, 61, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Rintoul, G.L.; Filiano, A.J.; Brocard, J.B.; Kress, G.J.; Reynolds, I.J. Glutamate Decreases Mitochondrial Size and Movement in Primary Forebrain Neurons. J. Neurosci. 2003, 23, 7881–7888. [Google Scholar] [CrossRef]
- MacAskill, A.F.; Kittler, J.T. Control of mitochondrial transport and localization in neurons. Trends Cell Biol. 2009, 20, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Bapat, O.; Purimetla, T.; Kruessel, S.; Shah, M.; Fan, R.; Thum, C.; Rupprecht, F.; Langer, J.D.; Rangaraju, V. VAP spatially stabilizes dendritic mitochondria to locally support synaptic plasticity. Nat. Commun. 2024, 15, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, V.; Devine, M.J.; Higgs, N.F.; Kittler, J.T. Miro1-dependent mitochondrial positioning drives the rescaling of presynaptic Ca 2+ signals during homeostatic plasticity. Embo Rep. 2016, 18, 231–240. [Google Scholar] [CrossRef] [PubMed]
- R. S. Zucker and W. G. Regehr, “Short-Term Synaptic Plasticity,” Annu. Rev. Physiol., vol. 64, no. 1, pp. 355–405, Mar. 2002.
- Divakaruni, S.S.; Van Dyke, A.M.; Chandra, R.; LeGates, T.A.; Contreras, M.; Dharmasri, P.A.; Higgs, H.N.; Lobo, M.K.; Thompson, S.M.; Blanpied, T.A. Long-Term Potentiation Requires a Rapid Burst of Dendritic Mitochondrial Fission during Induction. Neuron 2018, 100, 860–875. [Google Scholar] [CrossRef] [PubMed]
- Oettinghaus, B.; Schulz, J.M.; Restelli, L.M.; Licci, M.; Savoia, C.; Schmidt, A.; Schmitt, K.; Grimm, A.; Morè, L.; Hench, J.; et al. Synaptic dysfunction, memory deficits and hippocampal atrophy due to ablation of mitochondrial fission in adult forebrain neurons. Cell Death Differ. 2016, 23, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Shields, L.Y.; Kim, H.; Zhu, L.; Haddad, D.; Berthet, A.; Pathak, D.; Lam, M.; Ponnusamy, R.; Diaz-Ramirez, L.G.; Gill, T.M.; et al. Dynamin-related protein 1 is required for normal mitochondrial bioenergetic and synaptic function in CA1 hippocampal neurons. Cell Death Dis. 2015, 6, e1725–e1725. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Engeln, M.; Schiefer, C.; Patton, M.H.; Martin, J.A.; Werner, C.T.; Riggs, L.M.; Francis, T.C.; McGlincy, M.; Evans, B.; et al. Drp1 Mitochondrial Fission in D1 Neurons Mediates Behavioral and Cellular Plasticity during Early Cocaine Abstinence. Neuron 2017, 96, 1327–1341. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Tian, R.; Han, H.; Slone, J.; Wang, C.; Ke, X.; Zhang, T.; Li, X.; He, Y.; Liao, P.; et al. PINK1-mediated Drp1S616 phosphorylation modulates synaptic development and plasticity via promoting mitochondrial fission. Signal Transduct. Target. Ther. 2022, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Su, B.; Lee, H.-G.; Li, X.; Perry, G.; Smith, M.A.; Zhu, X. Impaired Balance of Mitochondrial Fission and Fusion in Alzheimer's Disease. J. Neurosci. 2009, 29, 9090–9103. [Google Scholar] [CrossRef]
- Wang, W.; Yin, J.; Ma, X.; Zhao, F.; Siedlak, S.L.; Wang, Z.; Torres, S.; Fujioka, H.; Xu, Y.; Perry, G.; et al. Inhibition of mitochondrial fragmentation protects against Alzheimer’s disease in rodent model. Hum. Mol. Genet. 2017, 26, 4118–4131. [Google Scholar] [CrossRef]
- Fischer, T.D.; Hylin, M.J.; Zhao, J.; Moore, A.N.; Waxham, M.N.; Dash, P.K. Altered Mitochondrial Dynamics and TBI Pathophysiology. Front. Syst. Neurosci. 2016, 10, 29–29. [Google Scholar] [CrossRef]
- Zhao, H.; Pan, W.; Chen, L.; Luo, Y.; Xu, R. Nur77 promotes cerebral ischemia–reperfusion injury via activating INF2-mediated mitochondrial fragmentation. Histochem. J. 2018, 49, 599–613. [Google Scholar] [CrossRef]
- Kumar, R.; Bukowski, M.J.; Wider, J.M.; Reynolds, C.A.; Calo, L.; Lepore, B.; Tousignant, R.; Jones, M.; Przyklenk, K.; Sanderson, T.H. Mitochondrial dynamics following global cerebral ischemia. Mol. Cell. Neurosci. 2016, 76, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Luo, Y.; Chen, L.; Zhang, Z.; Shen, C.; Li, Y.; Xu, R. Sirt3 inhibits cerebral ischemia-reperfusion injury through normalizing Wnt/β-catenin pathway and blocking mitochondrial fission. Cell Stress Chaperon- 2018, 23, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.; Zeigler, M.; Mizuno, S.; Morrison, R.S.; Totah, R.A.; Barker-Haliski, M. Reductions in Hydrogen Sulfide and Changes in Mitochondrial Quality Control Proteins Are Evident in the Early Phases of the Corneally Kindled Mouse Model of Epilepsy. Int. J. Mol. Sci. 2022, 23, 1434. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Park, H.; Kim, T.-H.; Kang, T.-C. LONP1 Regulates Mitochondrial Accumulations of HMGB1 and Caspase-3 in CA1 and PV Neurons Following Status Epilepticus. Int. J. Mol. Sci. 2021, 22, 2275. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Sun, P.; Zhang, H.; Yang, H. Mitochondrial quality control in the brain: The physiological and pathological roles. Front. Neurosci. 2022, 16, 1075141. [Google Scholar] [CrossRef] [PubMed]
- Verrigni, D.; Di Nottia, M.; Ardissone, A.; Baruffini, E.; Nasca, A.; Legati, A.; Bellacchio, E.; Fagiolari, G.; Martinelli, D.; Fusco, L.; et al. Clinical-genetic features and peculiar muscle histopathology in infantileDNM1L-related mitochondrial epileptic encephalopathy. Hum. Mutat. 2019, 40, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Vanstone, J.R.; Smith, A.M.; McBride, S.; Naas, T.; Holcik, M.; Antoun, G.; Harper, M.-E.; Michaud, J.; Sell, E.; Chakraborty, P.; et al. DNM1L-related mitochondrial fission defect presenting as refractory epilepsy. Eur. J. Hum. Genet. 2015, 24, 1084–1088. [Google Scholar] [CrossRef]
- Waterham, H.R.; Koster, J.; van Roermund, C.W.; Mooyer, P.A.; Wanders, R.J.; Leonard, J.V. A Lethal Defect of Mitochondrial and Peroxisomal Fission. New Engl. J. Med. 2007, 356, 1736–1741. [Google Scholar] [CrossRef]
- Misko, A.L.; Sasaki, Y.; Tuck, E.; Milbrandt, J.; Baloh, R.H. Mitofusin2 Mutations Disrupt Axonal Mitochondrial Positioning and Promote Axon Degeneration. J. Neurosci. 2012, 32, 4145–4155. [Google Scholar] [CrossRef]
- Pham, A.H.; Meng, S.; Chu, Q.N.; Chan, D.C. Loss of Mfn2 results in progressive, retrograde degeneration of dopaminergic neurons in the nigrostriatal circuit. Hum. Mol. Genet. 2012, 21, 4817–4826. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Sterky, F.H.; Mourier, A.; Terzioglu, M.; Cullheim, S.; Olson, L.; Larsson, N.-G. Mitofusin 2 is necessary for striatal axonal projections of midbrain dopamine neurons. Hum. Mol. Genet. 2012, 21, 4827–4835. [Google Scholar] [CrossRef] [PubMed]
- E Kushnareva, Y.; A Gerencser, A.; Bossy, B.; Ju, W.-K.; White, A.D.; Waggoner, J.; Ellisman, M.H.; Perkins, G.; Bossy-Wetzel, E. Loss of OPA1 disturbs cellular calcium homeostasis and sensitizes for excitotoxicity. Cell Death Differ. 2012, 20, 353–365. [Google Scholar] [CrossRef]
- Zaninello, M.; Palikaras, K.; Sotiriou, A.; Tavernarakis, N.; Scorrano, L. Sustained intracellular calcium rise mediates neuronal mitophagy in models of autosomal dominant optic atrophy. Cell Death Differ. 2021, 29, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Zaninello, M.; Palikaras, K.; Naon, D.; Iwata, K.; Herkenne, S.; Quintana-Cabrera, R.; Semenzato, M.; Grespi, F.; Ross-Cisneros, F.N.; Carelli, V.; et al. Inhibition of autophagy curtails visual loss in a model of autosomal dominant optic atrophy. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- J. Koch et al., “Disturbed mitochondrial and peroxisomal dynamics due to loss of MFF causes Leigh-like encephalopathy, optic atrophy and peripheral neuropathy,” J. Med. Genet., vol. 53, no. 4, pp. 270–278, Apr. 2016.
- Andreassi, C.; Zimmermann, C.; Mitter, R.; Fusco, S.; De Vita, S.; Saiardi, A.; Riccio, A. An NGF-responsive element targets myo-inositol monophosphatase-1 mRNA to sympathetic neuron axons. Nat. Neurosci. 2010, 13, 291–301. [Google Scholar] [CrossRef]
- Jung, J.; Ohk, J.; Kim, H.; Holt, C.E.; Park, H.J.; Jung, H. mRNA transport, translation, and decay in adult mammalian central nervous system axons. Neuron 2022, 111, 650–668. [Google Scholar] [CrossRef]
- C. Glock et al., “The translatome of neuronal cell bodies, dendrites, and axons,” Proc. Natl. Acad. Sci., vol. 118, no. 43, pp. 1–11, Oct. 2021.
- Fernandopulle, M.S.; Lippincott-Schwartz, J.; Ward, M.E. RNA transport and local translation in neurodevelopmental and neurodegenerative disease. Nat. Neurosci. 2021, 24, 622–632. [Google Scholar] [CrossRef]
- Voigt, A.; Herholz, D.; Fiesel, F.C.; Kaur, K.; Müller, D.; Karsten, P.; Weber, S.S.; Kahle, P.J.; Marquardt, T.; Schulz, J.B. TDP-43-Mediated Neuron Loss In Vivo Requires RNA-Binding Activity. PLOS ONE 2010, 5, e12247. [Google Scholar] [CrossRef]
- Daigle, J.G.; Lanson, J.N.A.; Smith, R.B.; Casci, I.; Maltare, A.; Monaghan, J.; Nichols, C.D.; Kryndushkin, D.; Shewmaker, F.; Pandey, U.B. RNA-binding ability of FUS regulates neurodegeneration, cytoplasmic mislocalization and incorporation into stress granules associated with FUS carrying ALS-linked mutations. Hum. Mol. Genet. 2012, 22, 1193–1205. [Google Scholar] [CrossRef]
- H. Ederle et al., “Nuclear egress of TDP-43 and FUS occurs independently of Exportin-1/CRM1,” Sci. Rep., vol. 8, no. 1, p. 7084, May 2018.
- Tsai, Y.-L.; Mu, Y.C.; Manley, J.L. Nuclear RNA transcript levels modulate nucleocytoplasmic distribution of ALS/FTD-associated protein FUS. Sci. Rep. 2022, 12, 1–14. [Google Scholar] [CrossRef]
- Ziff, O.J.; Harley, J.; Wang, Y.; Neeves, J.; Tyzack, G.; Ibrahim, F.; Skehel, M.; Chakrabarti, A.M.; Kelly, G.; Patani, R. Nucleocytoplasmic mRNA redistribution accompanies RNA binding protein mislocalization in ALS motor neurons and is restored by VCP ATPase inhibition. Neuron 2023, 111, 3011–3027. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-L.; Coady, T.H.; Lu, L.; Zheng, D.; Alland, I.; Tian, B.; Shneider, N.A.; Manley, J.L. ALS/FTD-associated protein FUS induces mitochondrial dysfunction by preferentially sequestering respiratory chain complex mRNAs. Genes Dev. 2020, 34, 785–805. [Google Scholar] [CrossRef] [PubMed]
- Altman, T.; Ionescu, A.; Ibraheem, A.; Priesmann, D.; Gradus-Pery, T.; Farberov, L.; Alexandra, G.; Shelestovich, N.; Dafinca, R.; Shomron, N.; et al. Axonal TDP-43 condensates drive neuromuscular junction disruption through inhibition of local synthesis of nuclear encoded mitochondrial proteins. Nat. Commun. 2021, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, Y.; Pazyra-Murphy, M.F.; Silagi, E.S.; Tasdemir-Yilmaz, O.E.; Li, Y.; Rose, L.; Yeoh, Z.C.; Vangos, N.E.; Geffken, E.A.; Seo, H.-S.; et al. Binding and transport of SFPQ-RNA granules by KIF5A/KLC1 motors promotes axon survival. J. Cell Biol. 2020, 220. [Google Scholar] [CrossRef] [PubMed]
- Baumann, S.; Komissarov, A.; Gili, M.; Ruprecht, V.; Wieser, S.; Maurer, S.P. A reconstituted mammalian APC-kinesin complex selectively transports defined packages of axonal mRNAs. Sci. Adv. 2020, 6, eaaz1588. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.C.; Grawenhoff, J.; Baumann, S.J.; Lorenzon, N.; Maurer, S.P. Mammalian Neuronal mRNA Transport Complexes: The Few Knowns and the Many Unknowns. Front. Integr. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Hacisuleyman, E.; Hale, C.R.; Noble, N.; Luo, J.-D.; Fak, J.J.; Saito, M.; Chen, J.; Weissman, J.S.; Darnell, R.B. Neuronal activity rapidly reprograms dendritic translation via eIF4G2:uORF binding. Nat. Neurosci. 2024, 27, 822–835. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.F.; Oleynikov, Y.; Kislauskis, E.H.; Taneja, K.L.; Singer, R.H. Characterization of a β-Actin mRNA Zipcode-Binding Protein. Mol. Cell. Biol. 1997, 17, 2158–2165. [Google Scholar] [CrossRef]
- Hüttelmaier, S.; Zenklusen, D.; Lederer, M.; Dictenberg, J.; Lorenz, M.; Meng, X.; Bassell, G.J.; Condeelis, J.; Singer, R.H. Spatial regulation of β-actin translation by Src-dependent phosphorylation of ZBPNature 2005, 438, 512–515. [CrossRef]
- Loedige, I.; Baranovskii, A.; Mendonsa, S.; Dantsuji, S.; Popitsch, N.; Breimann, L.; Zerna, N.; Cherepanov, V.; Milek, M.; Ameres, S.; et al. mRNA stability and m6A are major determinants of subcellular mRNA localization in neurons. Mol. Cell 2023, 83, 2709–2725. [Google Scholar] [CrossRef]
- Zaninello, M.; Bean, C. Highly Specialized Mechanisms for Mitochondrial Transport in Neurons: From Intracellular Mobility to Intercellular Transfer of Mitochondria. Biomolecules 2023, 13, 938. [Google Scholar] [CrossRef]
- Harbauer, A.B.; Hees, J.T.; Wanderoy, S.; Segura, I.; Gibbs, W.; Cheng, Y.; Ordonez, M.; Cai, Z.; Cartoni, R.; Ashrafi, G.; et al. Neuronal mitochondria transport Pink1 mRNA via synaptojanin 2 to support local mitophagy. Neuron 2022, 110, 1516–1531. [Google Scholar] [CrossRef] [PubMed]
- J.-M. M. Cioni et al., “Late Endosomes Act as mRNA Translation Platforms and Sustain Mitochondria in Axons,” Cell, vol. 176, no. 1–2, pp. 56-72.e15, Jan. 2019. [CrossRef]
- Cohen, B.; Altman, T.; Golani-Armon, A.; Savulescu, A.F.; Ibraheem, A.; Mhlanga, M.M.; Perlson, E.; Arava, Y.S. Co-transport of the nuclear-encoded Cox7c mRNA with mitochondria along axons occurs through a coding-region-dependent mechanism. J. Cell Sci. 2022, 135. [Google Scholar] [CrossRef] [PubMed]
- Hees, J.T.; Wanderoy, S.; Lindner, J.; Helms, M.; Mahadevan, H.M.; Harbauer, A.B. Insulin signalling regulates Pink1 mRNA localization via modulation of AMPK activity to support PINK1 function in neurons. Nat. Metab. 2024, 6, 514–530. [Google Scholar] [CrossRef] [PubMed]
- Zaninello, M.; Schlegel, T.; Nolte, H.; Pirzada, M.; Savino, E.; Barth, E.; Klein, I.; Wüstenberg, H.; Uddin, T.; Wolff, L.; et al. CLUH maintains functional mitochondria and translation in motoneuronal axons and prevents peripheral neuropathy. Sci. Adv. 2024, 10, eadn2050. [Google Scholar] [CrossRef] [PubMed]
- Schuhmacher, J.S.; Dieck, S.T.; Christoforidis, S.; Landerer, C.; Gallesio, J.D.; Hersemann, L.; Seifert, S.; Schäfer, R.; Giner, A.; Toth-Petroczy, A.; et al. The Rab5 effector FERRY links early endosomes with mRNA localization. Mol. Cell 2023, 83, 1839–1855. [Google Scholar] [CrossRef] [PubMed]
- E Cosker, K.; Fenstermacher, S.J.; Pazyra-Murphy, M.F.; Elliott, H.L.; A Segal, R. The RNA-binding protein SFPQ orchestrates an RNA regulon to promote axon viability. Nat. Neurosci. 2016, 19, 690–696. [Google Scholar] [CrossRef]
- R. De Pace et al., “Messenger RNA transport on lysosomal vesicles maintains axonal mitochondrial homeostasis and prevents axonal degeneration,” Nat. Neurosci., vol. 27, no. 6, pp. 1087–1102, Jun. 2024.
- Campbell, D.S.; E Holt, C. Chemotropic Responses of Retinal Growth Cones Mediated by Rapid Local Protein Synthesis and Degradation. Neuron 2001, 32, 1013–1026. [Google Scholar] [CrossRef]
- Taylor, A.M.; Wu, J.; Tai, H.-C.; Schuman, E.M. Axonal Translation of β-Catenin Regulates Synaptic Vesicle Dynamics. J. Neurosci. 2013, 33, 5584–5589. [Google Scholar] [CrossRef] [PubMed]
- Batista, A.F.; Martínez, J.C.; Hengst, U. Intra-axonal Synthesis of SNAP25 Is Required for the Formation of Presynaptic Terminals. Cell Rep. 2017, 20, 3085–3098. [Google Scholar] [CrossRef]
- Sun, C.; Nold, A.; Fusco, C.M.; Rangaraju, V.; Tchumatchenko, T.; Heilemann, M.; Schuman, E.M. The prevalence and specificity of local protein synthesis during neuronal synaptic plasticity. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Smart, F.M.; Edelman, G.M.; Vanderklish, P.W. BDNF induces translocation of initiation factor 4E to mRNA granules: Evidence for a role of synaptic microfilaments and integrins. Proc. Natl. Acad. Sci. 2003, 100, 14403–14408. [Google Scholar] [CrossRef]
- Kang, H.; Schuman, E.M. A Requirement for Local Protein Synthesis in Neurotrophin-Induced Hippocampal Synaptic Plasticity. Science 1996, 273, 1402–1406. [Google Scholar] [CrossRef] [PubMed]
- Querido, J.B.; Díaz-López, I.; Ramakrishnan, V. The molecular basis of translation initiation and its regulation in eukaryotes. Nat. Rev. Mol. Cell Biol. 2023, 25, 168–186. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Li, X.; Flores, A.D.; Lai, K.-O. The translation initiating factor eIF4E and arginine methylation underlie G3BP1 function in dendritic spine development of neurons. J. Biol. Chem. 2023, 299, 105029. [Google Scholar] [CrossRef]
- S. Callegari, L. D. Cruz-Zaragoza, and P. Rehling, “From TOM to the TIM23 complex – handing over of a precursor,” Biol. Chem., vol. 401, no. 6–7, pp. 709–721, May 2020.
- L. Van Haute et al., “Deficient methylation and formylation of mt-tRNAMet wobble cytosine in a patient carrying mutations in NSUN3,” Nat. Commun., vol. 7, no. 1, p. 12039, Jun. 2016.
- Graber, T.E.; Hébert-Seropian, S.; Khoutorsky, A.; David, A.; Yewdell, J.W.; Lacaille, J.-C.; Sossin, W.S. Reactivation of stalled polyribosomes in synaptic plasticity. Proc. Natl. Acad. Sci. 2013, 110, 16205–16210. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Moradi, M.; Reinhard, S.; Ji, C.; Jablonka, S.; Hennlein, L.; Lüningschrör, P.; Doose, S.; Sauer, M.; Sendtner, M. Dynamic remodeling of ribosomes and endoplasmic reticulum in axon terminals of motoneurons. J. Cell Sci. 2021, 134. [Google Scholar] [CrossRef] [PubMed]
- Fusco, C.M.; Desch, K.; Dörrbaum, A.R.; Wang, M.; Staab, A.; Chan, I.C.W.; Vail, E.; Villeri, V.; Langer, J.D.; Schuman, E.M. Neuronal ribosomes exhibit dynamic and context-dependent exchange of ribosomal proteins. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- E Ostroff, L.; Fiala, J.C.; Allwardt, B.; Harris, K.M. Polyribosomes Redistribute from Dendritic Shafts into Spines with Enlarged Synapses during LTP in Developing Rat Hippocampal Slices. Neuron 2002, 35, 535–545. [Google Scholar] [CrossRef]
- Chirillo, M.A.; Waters, M.S.; Lindsey, L.F.; Bourne, J.N.; Harris, K.M. Local resources of polyribosomes and SER promote synapse enlargement and spine clustering after long-term potentiation in adult rat hippocampus. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Biever, A.; Glock, C.; Tushev, G.; Ciirdaeva, E.; Dalmay, T.; Langer, J.D.; Schuman, E.M. Monosomes actively translate synaptic mRNAs in neuronal processes. Science 2020, 367, 526. [Google Scholar] [CrossRef]
- Baranov, S.V.; Baranova, O.V.; Yablonska, S.; Suofu, Y.; Vazquez, A.L.; Kozai, T.D.Y.; Cui, X.T.; Ferrando, L.M.; Larkin, T.M.; Tyurina, Y.Y.; et al. Mitochondria modulate programmed neuritic retraction. Proc. Natl. Acad. Sci. 2018, 116, 650–659. [Google Scholar] [CrossRef]
- Lores-Arnaiz, S.; Lombardi, P.; Karadayian, A.G.; Orgambide, F.; Cicerchia, D.; Bustamante, J. Brain cortex mitochondrial bioenergetics in synaptosomes and non-synaptic mitochondria during aging. Neurochem. Res. 2016, 41, 353–363. [Google Scholar] [CrossRef]
- C. Espino de la Fuente-Muñoz, M. C. Espino de la Fuente-Muñoz, M. Rosas-Lemus, P. Moreno-Castilla, F. Bermúdez-Rattoni, S. Uribe-Carvajal, and C. Arias, “Age-Dependent Decline in Synaptic Mitochondrial Function Is Exacerbated in Vulnerable Brain Regions of Female 3xTg-AD Mice,” Int. J. Mol. Sci., vol. 21, no. 22, p. 8727, Nov. 2020.
- Misgeld, T.; Schwarz, T.L. Mitostasis in Neurons: Maintaining Mitochondria in an Extended Cellular Architecture. Neuron 2017, 96, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Spillane, M.; Ketschek, A.; Merianda, T.T.; Twiss, J.L.; Gallo, G. Mitochondria Coordinate Sites of Axon Branching through Localized Intra-axonal Protein Synthesis. Cell Rep. 2013, 5, 1564–1575. [Google Scholar] [CrossRef]
- Cheng, A.; Wan, R.; Yang, J.-L.; Kamimura, N.; Son, T.G.; Ouyang, X.; Luo, Y.; Okun, E.; Mattson, M.P. Involvement of PGC-1α in the formation and maintenance of neuronal dendritic spines. Nat. Commun. 2012, 3, 1250. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Wang, W.; Hwang, J.; Namgung, U.; Min, K.-T. Increased ER–mitochondria tethering promotes axon regeneration. Proc. Natl. Acad. Sci. 2019, 116, 16074–16079. [Google Scholar] [CrossRef]
- Cajigas, I.J.; Tushev, G.; Will, T.J.; Dieck, S.T.; Fuerst, N.; Schuman, E.M. The Local Transcriptome in the Synaptic Neuropil Revealed by Deep Sequencing and High-Resolution Imaging. Neuron 2012, 74, 453–466. [Google Scholar] [CrossRef]
- Nijssen, J.; Aguila, J.; Hoogstraaten, R.; Kee, N.; Hedlund, E. Axon-Seq Decodes the Motor Axon Transcriptome and Its Modulation in Response to ALS. Stem Cell Rep. 2018, 11, 1565–1578. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Cao, W.; Wang, Y.; Zhu, Q.; Luo, J.; Wang, B.; Zheng, H.; Weitz, D.A.; Zong, C. Droplet-based transcriptome profiling of individual synapses. Nat. Biotechnol. 2023, 41, 1332–1344. [Google Scholar] [CrossRef]
- Gonzalez-Lozano, M.A.; Koopmans, F.; Sullivan, P.F.; Protze, J.; Krause, G.; Verhage, M.; Li, K.W.; Liu, F.; Smit, A.B. Stitching the synapse: Cross-linking mass spectrometry into resolving synaptic protein interactions. Sci. Adv. 2020, 6, eaax5783. [Google Scholar] [CrossRef] [PubMed]
- M. van Oostrum et al., “The proteomic landscape of synaptic diversity across brain regions and cell types,” Cell, vol. 186, no. 24, pp. 5411-5427.e23, Nov. 2023.
- Sen, A.; Cox, R.T. Clueless is a conserved ribonucleoprotein that binds the ribosome at the mitochondrial outer membrane. Biol. Open 2016, 5, 195–203. [Google Scholar] [CrossRef]
- Gao, J.; Schatton, D.; Martinelli, P.; Hansen, H.; Pla-Martin, D.; Barth, E.; Becker, C.; Altmueller, J.; Frommolt, P.; Sardiello, M.; et al. CLUH regulates mitochondrial biogenesis by binding mRNAs of nuclear-encoded mitochondrial proteins. J. Cell Biol. 2014, 207, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Sibilla, C.; Liu, R.; Yun, J.; Hay, B.A.; Blackstone, C.; Chan, D.C.; Harvey, R.J.; Guo, M. Clueless/CLUH regulates mitochondrial fission by promoting recruitment of Drp1 to mitochondria. Nat. Commun. 2022, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mitochondrial fusion and fission. Mitochondria display distinct morphologies and distributions in axons and dendrites. Generally, dendritic mitochondria display long and tubular shapes, forming a complex network within the dendritic arbor, while axonal mitochondria display a remarkably small sand punctate morphology, occupying a small fraction of the axonal volume. Created using BioRender.
Figure 1.
Mitochondrial fusion and fission. Mitochondria display distinct morphologies and distributions in axons and dendrites. Generally, dendritic mitochondria display long and tubular shapes, forming a complex network within the dendritic arbor, while axonal mitochondria display a remarkably small sand punctate morphology, occupying a small fraction of the axonal volume. Created using BioRender.
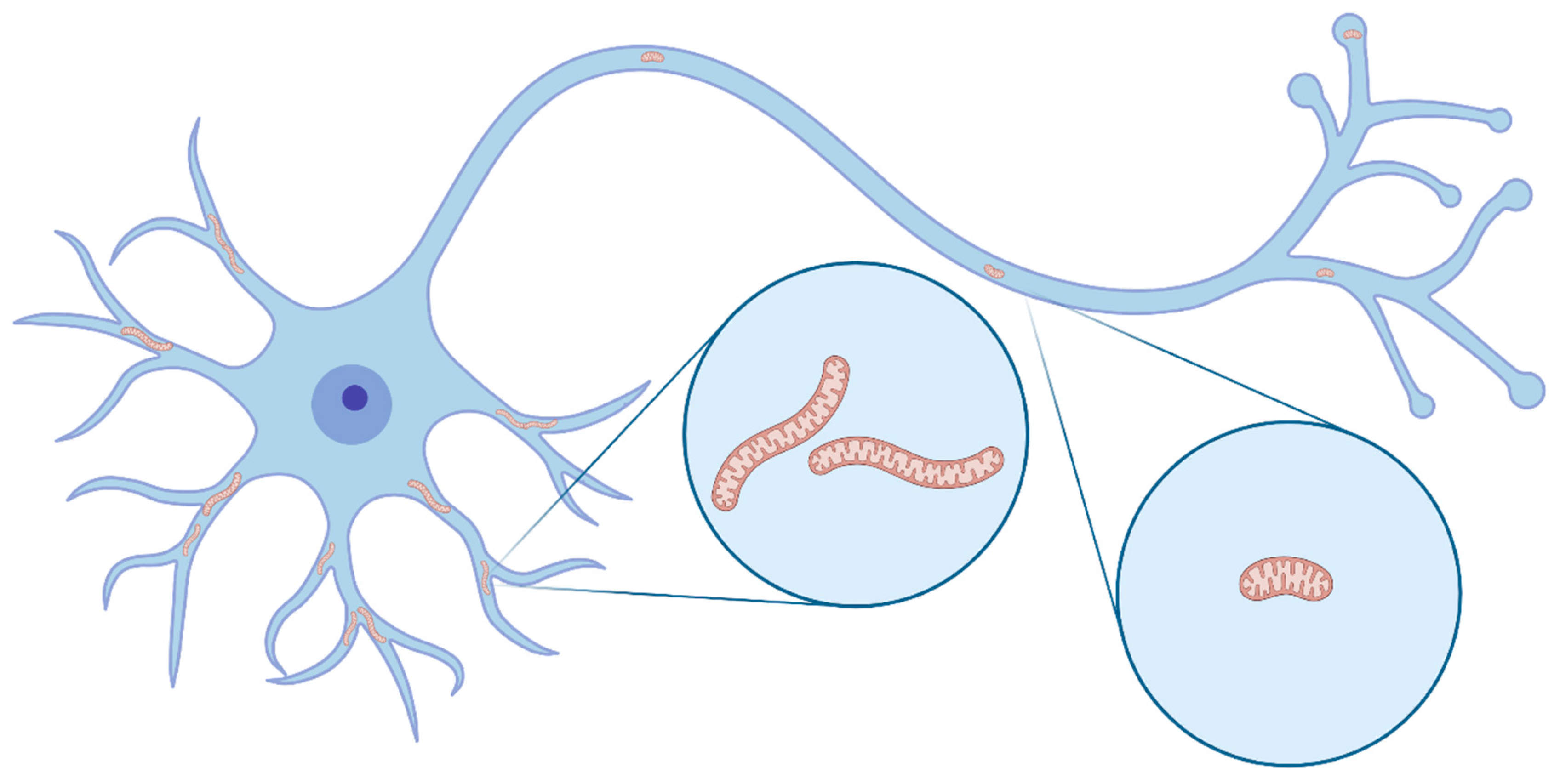
Figure 2.
Mitochondrial fusion and fission. Mitochondria are dynamic organelles that change morphology. They undergo cyclic processes of specialized structural alterations, namely fission and fusion that depend on particular proteins pro-fusion or pro-fission. Fusion is dependent on the tight control and activity of Mitofusins and OPA1, leading to the fusing of MIM and MOM, respectively. Regarding fission, a synchronous regulation of actin nucleation aided by the ER and the recruitment of DRP1 for the mitochondrial outer membrane, via the various described adaptors, is needed to rupture mitochondria. This cyclic process is crucial for the well-being of mitochondria, hence also for the physiologic functioning of neurons. Created using BioRender.
Figure 2.
Mitochondrial fusion and fission. Mitochondria are dynamic organelles that change morphology. They undergo cyclic processes of specialized structural alterations, namely fission and fusion that depend on particular proteins pro-fusion or pro-fission. Fusion is dependent on the tight control and activity of Mitofusins and OPA1, leading to the fusing of MIM and MOM, respectively. Regarding fission, a synchronous regulation of actin nucleation aided by the ER and the recruitment of DRP1 for the mitochondrial outer membrane, via the various described adaptors, is needed to rupture mitochondria. This cyclic process is crucial for the well-being of mitochondria, hence also for the physiologic functioning of neurons. Created using BioRender.
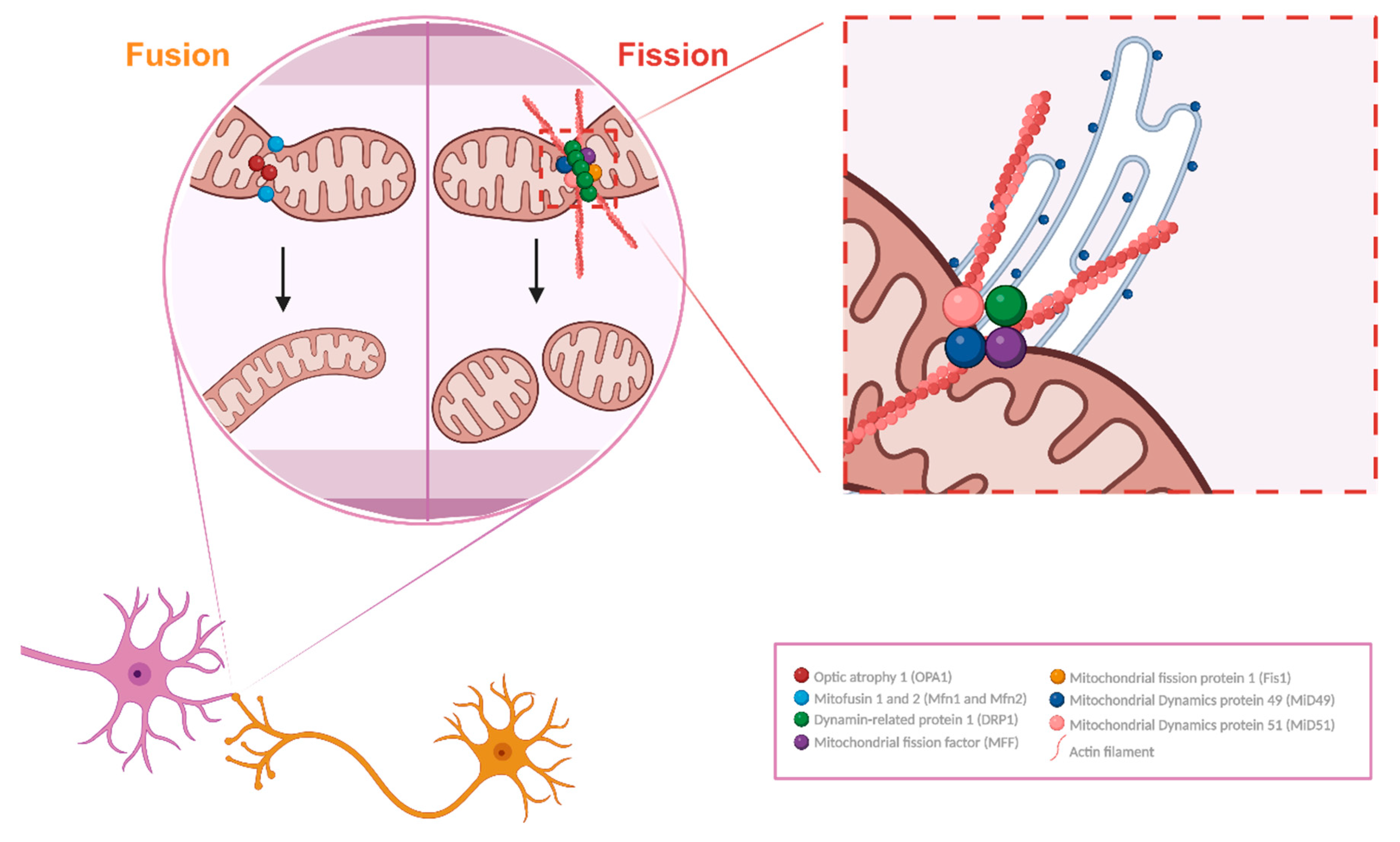
Figure 3.
Mitochondrial morphology-related events and neuronal regulation. Mitochondrial fusion has been widely implicated in several processes, such as neuronal differentiation, dendritic arborization or spine formation. Particularly, Mfn2-dependent mitochondrial fusion has been implicated in neuronal modulation. Mitochondrial fission, and particularly DRP1-dependent fission, has also been reported to regulate neuronal function and synaptic activity. Created using BioRender.
Figure 3.
Mitochondrial morphology-related events and neuronal regulation. Mitochondrial fusion has been widely implicated in several processes, such as neuronal differentiation, dendritic arborization or spine formation. Particularly, Mfn2-dependent mitochondrial fusion has been implicated in neuronal modulation. Mitochondrial fission, and particularly DRP1-dependent fission, has also been reported to regulate neuronal function and synaptic activity. Created using BioRender.
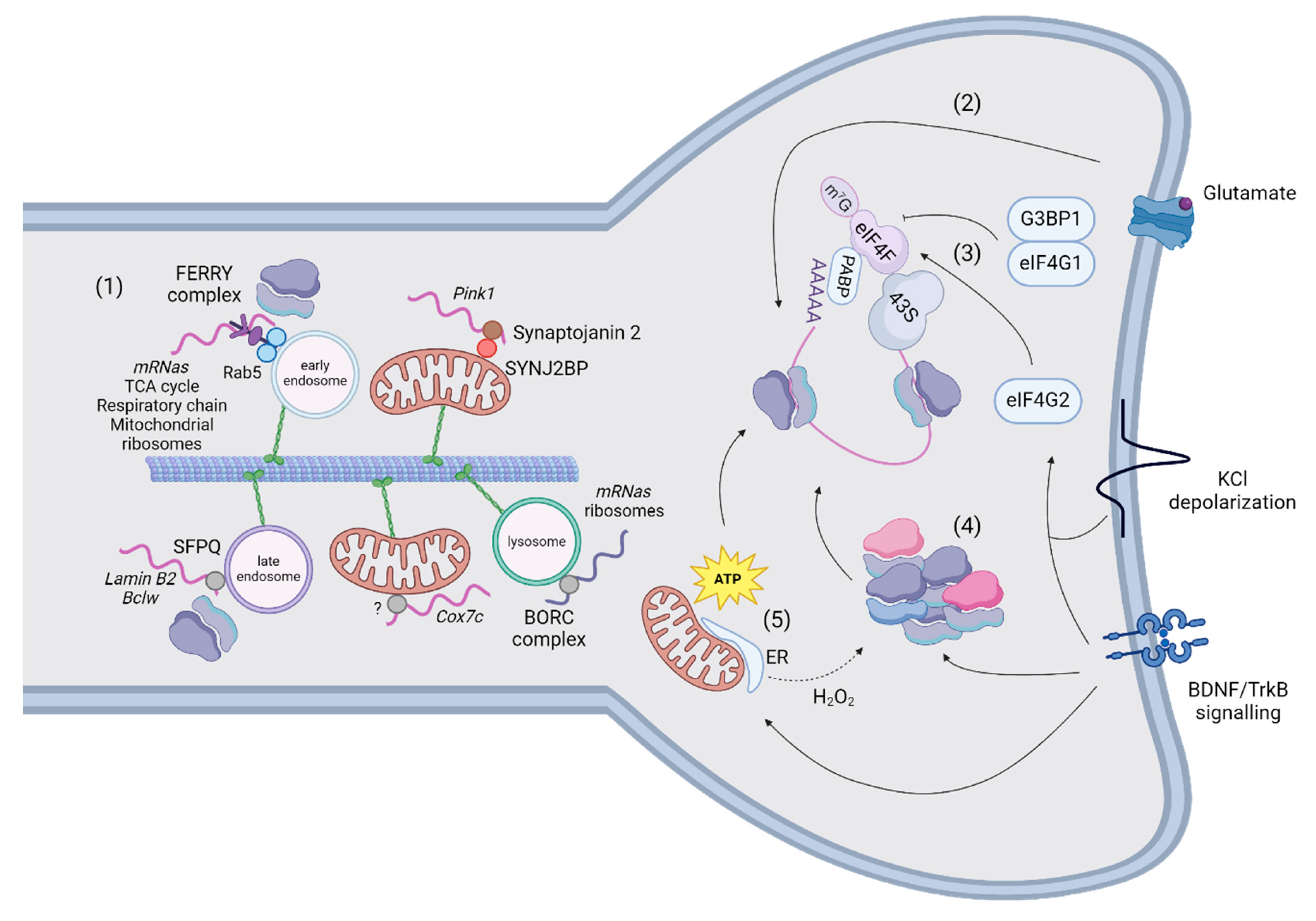
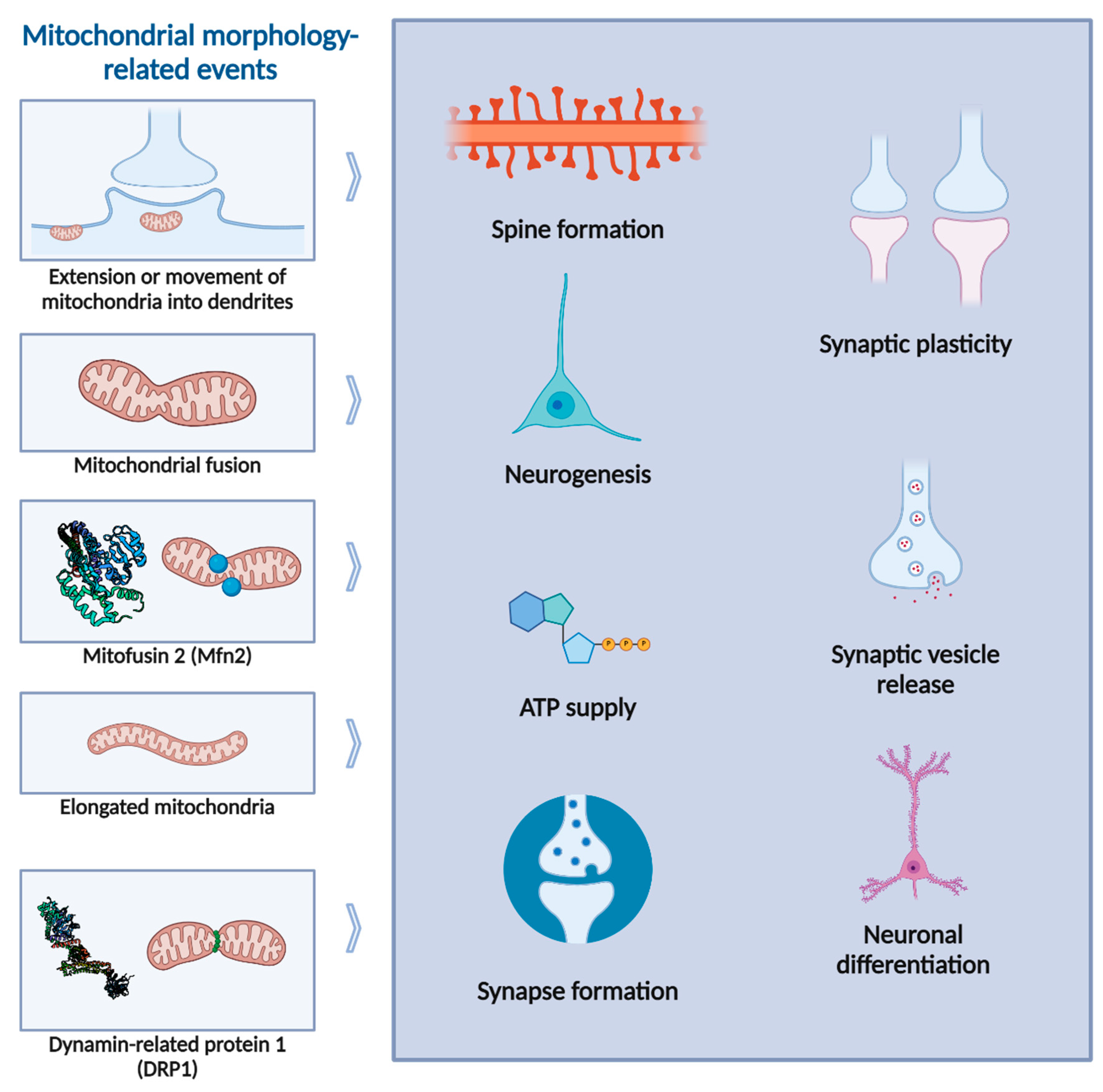
Figure 4.
Transport and translation in neurites and spines. The mechanisms of transport of NEM-mRNAs in dendrites is still scarce (1). In neurites, NEM-mRNAs are transported using adaptors together with mitochondria (Synaptojanin 2), early (FERRY complex) and late endosomes (SFPQ). The NEM-mRNAs analysed in studies so far encode for proteins involved in mitochondrial metabolism, translation and clearance. Endosomes and lysosomes can also transport ribosomes or mRNAs encoding for ribosomal subunits using the FERRY or the BORC complexes. Finally, endosomes in contact with mitochondria act as platforms to translate NEM-mRNAs. Once reached their destination, mRNAs are translated according to the stimulus. In glutamate-dependent long-term depression, translation is resumed in polyribosomes (2). It has been shown that translating ribosomes in neurites are also organised as monosomes. G3BP1 sequesters eIF4G1 and blocks the initiation of translation in absence of exogenous stimuli. However, BDNF or KCl-depolarization mobilises eIF4G2 to transcripts to increase the occupancy of ribosomes (3) and induces the synthesis and assembly of ribosomal subunits (4). Mitochondria are located in proximity of spines, where they produce ATP to sustain local synthesis (5). The tethering to the ER might potentiate translation by increasing the production of ATP. Of note, oxidative stress reshuffles the composition of ribosomal subunits (4, 5). Created using BioRender.
Figure 4.
Transport and translation in neurites and spines. The mechanisms of transport of NEM-mRNAs in dendrites is still scarce (1). In neurites, NEM-mRNAs are transported using adaptors together with mitochondria (Synaptojanin 2), early (FERRY complex) and late endosomes (SFPQ). The NEM-mRNAs analysed in studies so far encode for proteins involved in mitochondrial metabolism, translation and clearance. Endosomes and lysosomes can also transport ribosomes or mRNAs encoding for ribosomal subunits using the FERRY or the BORC complexes. Finally, endosomes in contact with mitochondria act as platforms to translate NEM-mRNAs. Once reached their destination, mRNAs are translated according to the stimulus. In glutamate-dependent long-term depression, translation is resumed in polyribosomes (2). It has been shown that translating ribosomes in neurites are also organised as monosomes. G3BP1 sequesters eIF4G1 and blocks the initiation of translation in absence of exogenous stimuli. However, BDNF or KCl-depolarization mobilises eIF4G2 to transcripts to increase the occupancy of ribosomes (3) and induces the synthesis and assembly of ribosomal subunits (4). Mitochondria are located in proximity of spines, where they produce ATP to sustain local synthesis (5). The tethering to the ER might potentiate translation by increasing the production of ATP. Of note, oxidative stress reshuffles the composition of ribosomal subunits (4, 5). Created using BioRender.
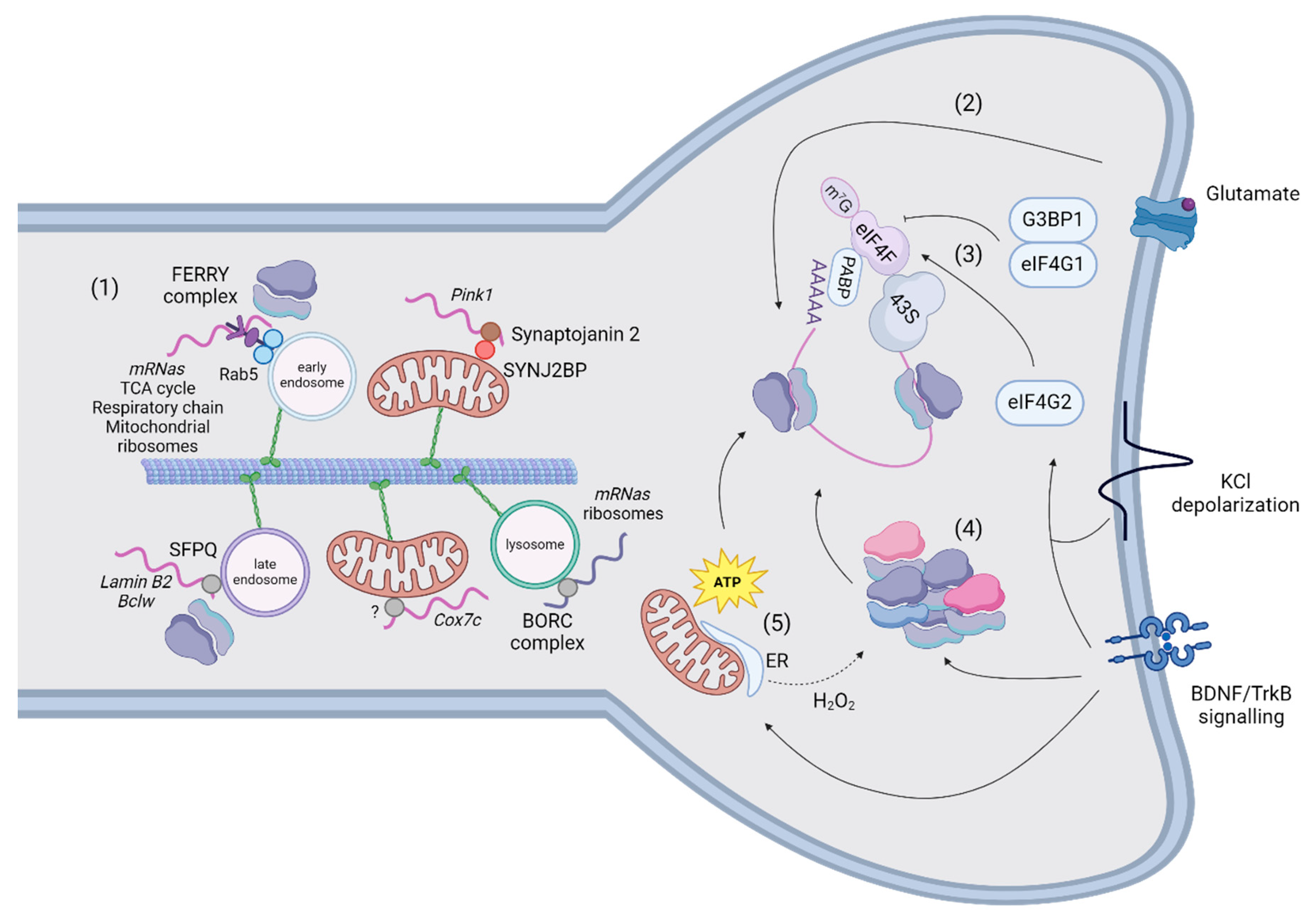
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



