
Submitted:
13 August 2024
Posted:
14 August 2024
You are already at the latest version
Abstract
To evaluate dryland wheat genotypes' performance to different pre-crop and residue management under dryland conditions, a split-split plot experiment based on RCB design with three replications was conducted for two years (2017-2018 and 2018-2019). The site of study's long-term average precipitation, temperature, and relative humidity are 376 mm, 9 ◦C, and 50%, respectively. Wheat-wheat and vetch-wheat cropping systems were considered in the main plots, different wheat and vetch residue levels including 0, 2, and 4 t ha-1 were located in subplots, and five dryland wheat genotypes including Sadra, Hashtroud, Baran, Varan, and Ohadi were allocated in sub-sub plots. Results indicated that leaf chlorophyll content index (CCI) and stomatal conductance (gs) were greater in the vetch-wheat cropping system compared to the wheat monoculture system for all genotypes. Normalized differences vegetative index (NDVI) of genotypes improved by applying the crop residue. Over two years, the application of crop residues resulted in higher variable fluorescence at J and I steps as well as an increase in the photosynthesis performance index (PI). Varan and Baran genotypes stood out as the superior genotype, exhibiting the highest values physiological characteristics and grain yield under the application of 4 t ha-1 of vetch residue. Grain filling rate (GFR) was reduced, while grain filling duration (GFD) was increased with increasing the crop residue levels. The enhanced grain yield of wheat genotypes grown under vetch residue attributed to the factors such as improvement in leave pigments and photosynthetic efficiency which facilitate longer grain filling duration with high grain weight. As a result, it is advisable to adopt a vetch-wheat cropping system with a high proportion of crop residue in dryland regions to achieve increased and sustainable wheat production.
Keywords:
Crop residue
; Dryland Wheat
; Grain Filling
; Chlorophyll fluorescence
1. Introduction
Increasing crop yields in recent decades have been strongly linked to the intensive use of chemical inputs, e.g., fertilizers, and also pesticides [1]. The adoption of intensive farming methods is a direct response to the increase in population size, which has subsequently led to heightened demands on agricultural areas and alterations in the nutritional composition of the soil [2] and soil and plant biodiversity [3]. In contrast to intensive agriculture, the set of management practices implemented in organic farming, e.g., returning crop residues to the soil, using organic and biological fertilizers, controlling weeds bio-mechanically, conservation tillage, applying different crop rotation systems, and intercropping [4], lead to improved recycling of soil nutrients, enhanced soil and crop biodiversity [5], and better yield quality [6].
Crop residues represent a significant renewable resource, rich in essential nutrients. The integration of straw into agricultural practices plays a crucial role in sustainable farming [7]. It is reported that recycling crop residues could maintain soil fertility [8] and support sustainable agriculture in cropping systems [9]. In addition, due to increasing awareness of sustainable agriculture, the area of straw incorporation is becoming larger and larger [10]. In arid regions of Iran, the implementation of crop rotation under no-till conditions emerges as a practical approach to enhance soil organic matter and ensure the stability of crop production by preserving plant residues.
Improving crop yield can be achieved by utilizing mineral and organic fertilizers effectively, managing crop residue, choosing suitable crop varieties for specific environments, and implementing bio-mechanical pest control techniques as an alternative to pesticides [11]. However, it was reported that the use of organic matter (e.g., crop residues) in the soil may reduce grain yield due to the immobilization of nutrients, especially during the initial growth stage of plants. This situation can be improved by the combined use of these organic resources with inorganic fertilizers, which can improve wheat grain yield by positively affecting the biological–chemical properties of the soil [12]. For instance, the utilization of both organic and inorganic nutrient resources in wheat led to an increase in nitrogen supply and a notable enhancement in wheat yield components like 1000-grain weight, tillers, wheat yield, and nitrogen uptake by the grain when compared to wheat that was only provided with inorganic fertilizers [13]. Also, maintenance and build-up of soil C are essential in improving soil functions and offsetting atmospheric CO2 concentration [14], which is a key challenge for humankind [15]. Crop residue management affects CO2 emissions, primarily via altering C and nitrogen (N) cycling [16]. However, multiple factors regulate CO2 emissions from the soil [1]. Soil respiration is influenced by variations in soil temperature and moisture levels, as well as the introduction of organic matter from decomposing residues, which in turn affect faunal and microbial activities [17,18].
Chlorophyll content, which reflects the health of crop leaves, plays a crucial role in the photosynthetic process and, therefore, dry matter production [19]. The slow but steady release of minerals from organic matter resources increases cereal grain yield by improving the photosynthetic capacity and chlorophyll content, especially during flowering and grain filling [1]. Iqbal et al., [19] noted that a combination of organic manure and chemical fertilizer application increases the levels of chlorophyll a and chlorophyll b and also increases the production of photoassimilates. Aside from photosynthesis, crop yields can also be improved by remobilizing dry matter accumulated in vegetative parts before anthesis to the grains during grain filling [20]. However, chlorophyll content and the translocation of photoassimilates within a plant can be affected by a variety of farming practices and climate conditions [21]. Remobilization of the assimilate is the main source of the dryland wheat yield production. Farming management and crop rotation affect crop productivity by influencing factors such as soil properties and plant characteristics such as leaf chlorophyll content and its photosynthetic activity [1]. The production of dry matter through the photosynthetic activity of leaves is the most important source of grain filling in cereals. Hence, the primary aim of this research was to assess pre-crop and residue management using vetch and wheat on physiological and grain yield characteristics in various dryland wheat genotypes.
2. Materials and Methods
2.1. Experimental Site
The field experiment to investigate photosynthesis activity and grain yield of dryland wheat genotypes was initiated in 2017-2019 at Dryland Agricultural Research Institute (latitude 37◦12′N; longitude 46◦20′E; 1730 m a.s.l.), 25 km east of Maragheh, East Azerbaijan Province, Iran. The region's climate is temperate continental with warm summers under the Koeppen’s classification system. The long-term (1989-2016) average precipitation, temperature, and relative humidity of the station are 376 mm, 9 ◦C, and 50%, respectively. Annual precipitation and average temperature per month of the experimental site are mentioned in Table 1. The soil (Fine Mixed, Mesic, Vertic Calcixerepts, USDA system; Calcisols, FAO system) at the study site had a clay loam texture in the 0–15 cm surface layer and a clay texture in the 15–80 cm depth. Before the beginning of the experiment, the physical and chemical properties of the soil were determined and the values of some physical and chemical properties of the soil are given in Table 2.
2.2. Experimental Design and Treatments
A split-split experiment based on RCBD with three replications was conducted. Wheat-wheat and vetch-wheat cropping systems were considered in main plots, three levels of wheat and vetch residues (0, 2, and 4 t ha-1) were located in sub-plots, and five dryland wheat genotypes (Sadra, Hashtroud, Baran, Varan and Ohadi) were allocated in sub-sub-plots. The subplots of the experiment were 44 square meters (10 x 4.4 meters).
An experiment was set up under a conservation agriculture system (No tillage) and the only soil disturbance occurred during the seeding operation. In October of both years before sowing crops, the gathered residues of vetch and wheat were uniformly distributed across the field in accordance with the specified treatments, while all crop residues were removed from the field for the control treatment. The residues were collected in various sizes from the thresher's output. Wheat and vetch (Vicia pannonica), with the rate of 380 and 240 seeds per m2, respectively, were sown by an Aske model 2200 seeder (13 planting rows with a 17.5 cm distance between rows) in 4–5 cm depth of soil. Each plot for both wheat and vetch consisted of 26 rows, spaced 17.5 cm apart. Fertilizers, including 90 kg ha-1 urea, 45 kg ha-1 triple superphosphate used for wheat, and 45 kg ha-1 urea, 30 kg ha-1 triple superphosphate used for vetch along with the sowing time. Weeds were controlled by applying Bromicide MA herbicide (1.5 L ha-1) in the wheat site and Gallant™ Super herbicide (1 L ha-1) in the vetch site.
2.3. Filed Detected Physiological Traits
Normalized differential vegetation index (NDVI) from each plot, stomatal conductance (gs), and chlorophyll content index (CCI) from three plants in each plot were recorded in both years at the anthesis stage (65 from Zadoks scale) by portable chlorophyll meter (SPAD 502 plus), GreenSiker (Trimber), and Porometer (AF4), respectively.
Photosynthesis activity and PSII performance were monitored with a Handy-PEA portable fluorimeter (Hansatech Instruments Ltd., Norfolk, UK) at the anthesis stage (65 from Zadoks scale). Three plants from each plot were randomly selected and the flag leaves of plants were dark-adapted for 30 min using a leaf clip provided by the producer of handy-PEA. Measurements were performed on the middle of plant flag leaves following the standard protocol with illumination with continuous red light (peak in 650 nm wavelength; the spectral line half width of 22 nm) provided by an array of three light-emitting diodes. The light pulse intensity used was 3500 μmol (photon) m-2 s -1 and the duration of the light pulse was 1 s. The measured data were used to calculate the photosynthetic parameters using Biolyzer v. 3.06 HP software (software provided with handy-PEA). Some of the parameters we discussed in this article due to their significance are Fo = minimum fluorescence, FM = maximum fluorescence, FV/FM = the maximum quantum efficiency of PSII, VJ = the relative variable fluorescence in step J after 2 ms, VI = the relative variable fluorescence in step I after 30 ms, PIABS = performance index.
2.4. Grain Filling Rate and Duration
Spikes of all genotypes were harvested after flowering with 7 days’ distance in five stages. Then, grains were removed from the spike and, subsequently, grains were dried, and the mean grain weight of each sample was determined. A two-piece regression model was applied to determine maximum grain and grain filling duration (GFD).
Where, W is grain weight, a is the intercept, b is the slope, t is days after anthesis, and tm is the end of the grain-filling period (time of mass maturity). Subsequently, GFR was calculated as:
GFR= MGW/GFD
Where, GFR is grain-filling rate, MGW is maximum grain weight and GFD is grain-filling duration [22].
2.5. Grain Yield and Weight
Five samples of 3 square meters (35% of the plot) were taken from each plot when grain in spike had around 14–15% moisture. After being harvested, each plot’s worth of grain was packed and threshed. Then, grains of the whole plot were weighted and the yield for 15 m2 was determined. According to that, the grain yield (GY) was calculated in Kg per hectare. For measuring 1000 seed weight, an automatic seed counter was used with eight repetitions.
2.6. Data Analysis
All the data were analyzed based on experimental design, using SAS v. 9.1 software. The mean value of each trait was compared according to the Duncan multiple range test at p ≤ 0.05.
3. Results
3.1. Physiological Traits: CCI, gs and NDVI
In both years of the experiment, the leaf chlorophyll content index (CCI) and stomatal conductance (gs) were found to be greater in the vetch-wheat cropping system compared to the wheat-wheat for all genotypes. The highest and lowest values of the CCI and gs in both years of the experiment and pre-crop treatments were recorded for the Varan and Baran genotypes, respectively. The CCI of the Hashtroud and Ohadi genotypes was statistically similar to the Varan genotype (Table 3).
The responses to crop residue levels among different genotypes did not exhibit any significant differences. The CCI of all genotypes increased with crop residue application. The enhancement in CCI of the Sadra (16%) and Hashtroud (17%) was greater than that of other genotypes. CCI in almost all genotypes under both 2 and 4 t ha-1 crop residue treatments was statistically similar. However, the CCI of the Varan genotype increased with increasing crop residue levels. The highest CCI was obtained from the Varan genotype under 4 t ha-1 crop residue (Figure 1).
Normalized differences vegetative index (NDVI) of wheat genotypes were improved in both rates of 2 and 4 t ha-1 of wheat and vetch residues compared to the control condition. The NDVI of the genotypes significantly decreased at 4 t ha-1 of wheat residue. However, NDVI of all genotypes at 4 t ha-1 of the vetch residues increased. The highest NDVI was recorded for the Varan and Baran genotypes with vetch residue treatments. The Baran and Ohadi genotypes exhibited a greater degree of change in their NDVI values compared to other genotypes when subjected to both wheat and vetch pre-crop residue. In contrast, the Hashtroud genotype did not display a heightened sensitivity to crop residue (Table 4). The gs activity of the genotypes in the vetch pre-crop treatment generally exhibited higher levels compared to the wheat monoculture system. The gs of all genotypes increased by increasing the residue from both wheat and vetch crops. However, the gs of all genotypes were suspended/reduced at the 4 t ha-1 of wheat residue. The highest gs were recorded for Varan and Baran genotypes with 4 t ha-1 of vetch pre-crop treatment (Table 4).
3.2. Chlorophyll Fluorescence Parameters
In both years of the study, there was a significant decrease observed in the light absorption per active reaction center (ABS/RC) at 2 t ha-1 of wheat residue and 2 and 4 t ha-1 of vetch residue levels. In the initial year, the ABS/RC of samples from both the control and the 4 t ha-1 of the wheat residue exhibited no significant difference. However, in the subsequent year, the ABS/RC value surpassed that of the control treatment when 4 t ha-1 of wheat residue was applied (Table 5). Over two years, the utilization of wheat and vetch residue resulted in higher variable fluorescence at J and I steps as well as an increase in the photosynthesis performance index (PI), in comparison to the control treatment. However, these increments under 4 t ha-1 of wheat residue were lower than 2 t ha-1. The greatest values for all chlorophyll fluorescence parameters were recorded with 4 t ha-1 of vetch residue during the first year (Table 5).
The application of crop residue from wheat and vetch resulted in an increase in the chlorophyll fluorescence parameters across all genotypes. Nevertheless, when wheat residue was applied at a rate of 4 t ha-1, it caused a reduction in the value of these parameters in all genotypes. Generally, chlorophyll fluorescence parameters of the samples under vetch residue were higher than that of wheat residue, especially with 4 t ha-1 of vetch residue. In terms of chlorophyll fluorescence parameters, the Varan genotype stood out as the superior one, exhibiting the highest values for Fv/Fo, Fv/Fm, VJ, VI, and PI under the application of 4 t ha-1 of vetch residue. However, there were no significant differences between Baran and Varan genotypes for Fv/Fo and VJ parameters (Table 6).
In the second year of the study, the values of the Fv/Fo, Fv/Fm, and VI were significantly reduced (Figure 2A, B, and C). Varan had the highest Fv/Fm value in both years. Differences among Sadra, Hashtroud, and Ohadi in the first year and between Sadra and Hashtroud in the second year in terms of Fv/Fm were not significant (Figure 2B). Wheat genotypes had more VI in vetch pre-crop than wheat monoculture treatment during two years. The statistical similarity of the responses among almost all genotypes was observed in both years under the wheat pre-cropping system as well as in the second year under the vetch pre-cropping system for VI. However, the highest VI value was obtained from Varan under vetch pre-crop to wheat genotypes in the first year (Figure 2 C).
3.3. Grain Yield and Components
With the increases in crop residue levels, the 1000 grain weight (GW) of all genotypes increased. This increment for the vetch-wheat was more than the wheat-wheat cropping system. The highest grain weight was recorded for Varan and Baran genotypes under 4 t ha-1 vetch residue. The grain-filling rate (GFR) of the genotypes was reduced with increasing the crop residue levels. This reduction in genotypes was different. The highest rate of grain filling was obtained from Varan and Baran under the vetch-wheat system when the crop residue was removed (control treatment). In contrast, the grain filling duration (GFD) of all genotypes increased with increasing vetch residue levels. GRD of genotype under 4 t ha-1 of wheat residue decreased or remained unchanged in comparison to the control. The duration of grain filling was the longest for the Ohadi, Varan, and Baran genotypes when subjected to a vetch residue of 4 t ha-1. The genotypes exhibited higher grain yield as the wheat residue progressively increased up to 2 t ha-1, while the vetch residue showed a similar trend up to 4 t ha-1. The grain yield of wheat either decreased or remained unchanged depending on different genotypes under 4 t ha-1 of wheat residue. Varan and Baran achieved the highest grain yield when subjected to 4 t ha-1 of vetch residue (Table 7).
4. Discussion
4.1. Physiological Parameters: CCI, NDVI, and gs
The increase in the content of chlorophyll under the vetch-wheat system (Table 3) may be due to the gradual release of nutrients such as nitrogen and manganese from the vetch (legume) residue. The chlorophyll content is an index used to evaluate the health of leaves and the photosynthetic ability of crops, which is highly influenced by environmental conditions such as nutrition [23]. The conditions for the development of microorganisms are more favourable in the legume cropping system, which results in higher soil organic matter deposition and higher exudation of organic compounds by the roots [24]. These conditions provide N, resulting in increases in the chlorophyll content. Increasing the gs under the vetch-wheat system (Table 3) is related to the effects of the legume pre-crop on nutrient availability. Nitrogen availability can influence stomatal conductance in plants. Increased nitrogen availability can lead to higher stomatal conductance, as nitrogen is a crucial nutrient for plant growth and development [25].
The relationship between crop residue quantity and CCI is determined by how residue impacts soil water storage and nutrient availability for plant growth. Chlorophyll content in plants can vary depending on factors such as light intensity, temperature, and nutrient availability. Higher CCI under different levels of the crop residue (Figure 1) indicated that N in the root zone soil is often higher in cereal–legume cropping systems than in cereal monoculture [26]. Lotfi and Pessarakli [27] demonstrated that maintaining the residues, especially in the vetch–wheat cropping system, increasing the soil moisture storage, and also enhancing soil fertility through the rotation of legumes and cereals contributes to the improvement of plant growth by enhancing plant pigment production.
The NDVI index is employed for assessing the health status of plants, and the elevated NDVI values observed in the wheat genotypes within the vetch-wheat cropping system (Table 4) signify the presence of favorable conditions conducive to plant growth. Lotfi and Pessarakli [27] found that plants from the vetch–wheat rotation had higher NDVI than chickpea–wheat and safflower–wheat treatments. NDVI has previously been associated with plant water status by Zhang et al., [28], even though it is primarily affected by the relative water content and cell wall elasticity of leaf tissues. Spectral vegetation indexes, such as NDVI, could successfully estimate the photosynthetic efficiency of the crop [27] and it is sensitive to the leaf area index and green biomass [28]. By estimating the mid-season N requirement, the NDVI sensor could act as a framework for rational N management in cereals [28]. A robust relationship between NDVI and grain yield showed higher NUEs than the conventional approach of N recommendation [29].
4.2. Chlorophyll Fluorescence Parameters
The impairment of PSII reaction centers during stressful conditions has been impacted by electron transport and energy conversion. Certain OJIP-related parameters, like ABS/RC, have been computed to pinpoint the areas of damage on the acceptor side of PSII [30]. Reducing the ABS/RC at 2 t ha-1 of wheat residue and 2 and 4 t ha-1 of vetch residue levels (Table 5) showed plants have not been under stress conditions. Under stressed conditions, ABS/RC dramatically increased, implying the dissipation of most of the absorbed energy by the PSII reaction center [31]. According to Strasser et al. [30], increasing ABS/RC under stress might indicate a change in the number of light-harvesting complexes (HLC) per PSII reaction center or inactivation of some reaction centers, i.e., a reduction in active QA-reducing reaction centers. Additional factors leading to the increase in ABS/RC levels include the deterioration of chlorophyll due to early leaf senescence caused by severe stress and the relocation of antennae from inactive PSII RCs to active ones [32]. In our research, the reduction of the ABS/RC was supported by the NDVI and CCI results (Figure 1, Table 4). Fv/Fo is the most sensitive component in the photosynthetic electron transport chain [32]. The increase in Fv/Fo (Table 5) as a consequence of crop residue application, results from photosynthetic electron transport construction. The maximal quantum efficiency of PSII (Fv/Fm) and the photosynthetic performance index (PI) are widely utilized metrics for evaluating the condition of PSII in response to abiotic stressors [33]. However, there is conclusive evidence that advises against reliance on Fv/Fm and PI as the most important parameters [32]. Increasing Fv/Fm value under crop residue (Table 5) indicates high efficiency of the electron transport and quantum yield of the PSII as Kalaji et al. [33] reported that the reduction in Fv/Fm under stress conditions could prevent the transfer of electrons from PSII reaction center to electron transporters. Improving the Fv/Fm can be interpreted as a reversible effect of light inhibition, which has a protective and regulatory function for the photosynthetic system of leaves [32]. The reduction in Fv/Fm under high levels of wheat residue might indicate the fact that PSII RCs were impaired or not functioning effectively in photosynthesis.
4.3. Grain Yield and Component
Environmental stresses during grain filling can lead to a reduction in mean grain weight [34]. Genetic factors can also contribute to grain weight, influencing the rate and duration of grain filling and their interaction [35]. Our results indicated that Baran and Varan had the highest grain filling duration and weight (Table 6). Environmental fluctuations such as temperature, water availability, light intensity, and nutrition caused by different crop rotations and residue levels have a significant impact on grain weight and yield. This is likely related to high environmental variations during the relatively long grain-filling period [36]. In some cases, the grain weight was more related to the grain-filling duration (GFD), while in others it was more related to the maximum grain-filling rate [37]. Our results confirm that crop residue levels had more significant effects on grain-filling duration than grain-filling rate (Table 7). GFD and GFR have major effects on final grain weight and can have significant effects on final yield [38]. Differential responses may easily emerge when the genotypes are exposed to a sufficiently wide range of growing conditions. In our study, we demonstrated that the determination of the grain weight and grain-filling parameters was influenced by both genotypes and environment. Crop residue by affecting the soil properties like moisture content and temperature can affect plants growing conditions. Lotfi and Pessarakli [27] reported that soil moisture storage is influenced by crop rotation and tillage methods in dryland conditions. In addition, several studies have demonstrated that genotype determines the grain-filling rate, whereas environmental factors, such as temperature, affect the duration of the grain-filling period [38]. Higher grain yield (Table 7) of wheat genotypes under vetch crop residue is related to improvement in NDVI (Table 4), photosynthetic efficiency (Table 6), grain-filling duration, and grain weight (Table 7). Chlorophyll is a key component in the process of photosynthesis. The findings of our study indicate that reduced ABS/RC levels contribute to the maintenance of chlorophyll in leaves by delaying senescence. Additionally, increased values of Fv/Fo and Fv/Fm are associated with higher photosynthetic efficiency in plants. So, higher values of CCI and /or NDVI especially during grain filling suppliers’ great requirements of photosynthates during grain development [39]. A longer duration of grain filling facilitates more dry matter accumulation, which ultimately results in high yield [40].
Possible factors for the higher crop performance under forage legumes are related to soil moisture availability and nutrient uptake. Soil texture and/or initial nutrient status or residue quality can affect plant responses to the different types of residue [8]. The literature on the effect of residue management on nitrogen (N) or phosphorus (P) uptake by plants is equally dispersed with no (for N: [41]); positive (for N: [42]) or negative (for N: [43]) effects reported by different authors. N is a key factor in the reaction of grains following legumes compared with grains following nonlegumes [44]. Geng et al. [45] conducted a global meta-analysis on legume-wheat rotation systems effect and indicate that legume-wheat rotations significantly increased wheat yields compared to gramineae–wheat rotations (P < 0.05). This is consistent with the results of Cernay et al. [46], whose meta-analysis found that previous legume crops significantly increased subsequent grain yields by 29 % on average.
The potential decline or cessation in the growth and productivity of wheat genotypes when exposed to 4 t ha-1 of wheat residue (Table 7) could potentially be attributed to the allelopathic impacts of the material. Allelopathy in wheat has been deeply studied and demonstrated by a consolidated scientific literature [47]. There are many allelochemicals involved in wheat allelopathy, belonging to three main chemical classes: phenolic acids (e.g., p-hydroxybenzoic, ferulic, syringic, vanillic, p-coumaric, etc.), benzoxazinoids (DIMBOA, MBOA, HMBOA, DIMBOA glycoside, BOA) and short-chain fatty acids (e.g., propionic, acetic, butyric, etc.) [47]. Scavo et al., [48] reported that ear extracts of durum wheat had the most active phytotoxicity and they suggest that wheat residue could be left on the soil surface or soil incorporated after harvest for possible weed control. Reduction of the physiological properties like CCI (Figure 1) content and chlorophyll fluorescence values (Table 6) of the genotypes under 4 t ha-1 wheat residue might be the allelochemical's adverse effect on the uptake of nutrients. For example, ferulic acid and cinnamic acid decrease the availability of Fe, P, and other nutrients [47]. This reduction is attributed to altered cell membrane permeability due to the presence of these allelochemicals [48]. Several physiological effects induced in response to allelochemicals include a reduction in overall plant growth [48]. Khaliq et al. [44] found that wheat straw-amended and infested-rhizosphere soil had an allelopathic effect on T. portulacastrum and significantly reduced its germination, seedling growth, and biochemical attributes.
5. Conclusions
In both years of the experiment, the CCI and gs were greater in the vetch-wheat cropping system compared to the wheat-wheat for all genotypes. NDVI of wheat genotypes improved in both rates of 2 and 4 t ha-1 of wheat and vetch residues compared to that of the control condition. The gs activity of the genotypes grown in vetch pre-crop generally exhibited higher levels compared to the wheat pre-crop. The gs activity of all phenotypes did not show an increase when the level of wheat residue reached 4 t ha-1. Over two years, the utilization of wheat and vetch residue resulted in higher variable fluorescence at J and I steps as well as an increase in PI, but ABS/RC declined in comparison to the control treatment. Varan genotype stood out as the superior one, exhibiting the highest values for Fv/Fo, Fv/Fm, VJ, VI, and PI under the application of 4 t ha-1 of vetch residue. The highest GW grains were recorded for Varan and Baran genotypes under 4 t ha-1 of vetch residue. The GFR of the genotypes reduced with increasing the crop residue levels. In contrast, the GFD of all genotypes increased with increasing vetch residue levels. Higher GY of genotypes under vetch residue is related to improving in NDVI, photosynthetic efficiency, grain-filling duration, and grain weight. It can be concluded that a vetch-wheat cropping system with crop residue management in dryland areas is one of the key sustainable practices for wheat production, but this concept could be expanded by assessing the rotation of additional crops in conjunction with various tillage systems.
Author Contributions
R.L.—investigation, writing—original draft preparation; S.G.- investigation, formal analysis; A.A.-software; R.R.- data curation; A.M.- methodology; M.P.—writing—review and editing. All authors have read and agreed with the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All data presented in the Results and Discussion are available upon request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Geng, Y.; Cao, G.; Wang, L.; Wang, S. Effects of equal chemical fertilizer substitutions with organic manure on yield, dry matter, and nitrogen uptake of spring maize and soil nitrogen distribution. PLoS ONE 2019, 14, e0219512. [Google Scholar] [CrossRef]
- Berthrong, S.T.; Buckley, D.H.; Drinkwater, L.E. Agricultural management and labile carbon additions affect soil microbial community structure and interact with carbon and nitrogen cycling. Micro. Ecol. 2013, 66, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends in Ecol. & Evol. 2003, 18, 182–188. [Google Scholar]
- Shennan, C.; Krupnik, T.J.; Baird, G.; Cohen, H.; Forbush, K.; Lovell, R.J.; Olimpi, E.M. Organic and conventional agriculture: A useful framing? Ann. Rev. Environ. and Reso. 2017, 42, 317–346. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ahnström, J.; Weibull, A.C. The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J. App. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Hammad, H.M.; Khaliq, A.; Abbas, F.; Farhad, W.; Fahad, S.; Aslam, M.; Shah, G.M.; Nasim, W.; Mubeen, M.; Bakhat, H.F. Comparative effects of organic and inorganic fertilizers on soil organic carbon and wheat productivity under the arid region. Comm. in Soil Sci. & Plant Anal. 2020, 51, 406–422. [Google Scholar]
- Xu, X.; Pang, D.; Chen, J.; Luo, Y.; Zheng, M.; Yin, Y.; Yi, L.; Wang, Z. Straw return accompanies by low nitrogen moderately promoted deep root. Field Crops Res. 2018, 221: 71–80.
- Chen, J.; Gong, Y.; Wang, S.; Guan, B.; Balkovic, J.; Kraxner, F. To burn or retain crop residues on croplands? An integrated analysis of crop residue management in China. Sci. Total Environ. 2019, 662, 141–150. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Xu, M.; Feng, G.; Zhang, W.; Lu, C. Crop yield and soil organic matter after long–term straw return to soil in China. Nutr. Cycl. Agroecosys. 2015, 102, 371–381. [Google Scholar] [CrossRef]
- Xiaobing, L.; Zhi, W.; Linjie, M.; Nan, C.; Yali, M.; Zhiguo, Z. Crop residue incorporation combined with potassium fertilizer increased cotton canopy apparent photosynthesis and seed cotton yield in a barley-cotton rotation system, Archi. Agron. & Soil Sci. 2021. 67, 300-312.
- Petrescu-Mag, R.M.; Banatean-Dunea, I.; Vesa, S.C.; Copacinschi, S.; Petrescu, D.C. What do Romanian farmers think about the effects of pesticides? Perceptions and willingness to pay for bio-pesticides. Sustain. 2019, 11, 3628. [Google Scholar]
- Agegnehu, G.; Tsigie, A.; Tesfaye, A. Evaluation of crop residue retention, compost, and inorganic fertilizer application on barley productivity and soil chemical properties in the central Ethiopian highlands. Ethiop. J. Agric. Sci. 2012, 22, 45–61. [Google Scholar]
- Sarwar, G.; Schmeisky, H.; Hussain, N.; Muhammad, S.; Tahir, M.A.; Saleem, U. Variations in nutrient concentrations of wheat and paddy as affected by different levels of compost and chemical fertilizer in normal soil. Pak. J. Bot. 2009, 41, 2403–2410. [Google Scholar]
- Delgado-Baquerizo, M.; Eldridge, D.; Maestre, F.; Karunaratne, S.; Trivedi, P.; Reich, P.; Singh, B. 2017. Climate legacies drive global soil carbon stocks in terrestrial ecosystems. Sci. Adv. 2017, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Comino, J. ; Lopez-Vicente, ´ M. ; Kumar, V.; Rodríguez-Seijo, A.; Valko, ´ O.; Rojas, C.; Pourghasemi, H.R.; Salvati, L.; Bakr, N.; Vaudour, E. 2020. Soil science challenges in a new era: a transdisciplinary overview of relevant topics. Air, Soil & Water Res. 2020, 13, 1178622120977491. [Google Scholar]
- Nawaz, A.; Lal, R.; Shrestha, R.K.; Farooq, M. Mulching affects soil properties and greenhouse gas emissions under long-term no-till and plough-till systems in Alfisol of Central Ohio. Land Degrad. & Develop. 2017, 28 (2), 673–681.
- Dong, W.; Liu, E.; Wang, J.; Yan, C.; Li, J.; Zhang, Y. Impact of tillage management on the short-and long-term soil carbon dioxide emissions in the dryland of Loess Plateau in China. Geoder. 2017, 307, 38–44. [Google Scholar] [CrossRef]
- Toosi, E.R.; Kravchenko, A.N.; Guber, A.K.; Rivers, M.L. 2017. Pore characteristics regulate the priming and fate of carbon from plant residue. Soil Biol. Biochem. 113, 219–230.
- Iqbal, A.; He, L.; Ali, I.; Ullah, S.; Khan, A.; Khan, A.; Akhtar, K.; Wei, S.; Zhao, Q.; Zhang, J. Manure combined with chemical fertilizer increases rice productivity by improving soil health, post-anthesis biomass yield, and nitrogen metabolism. PLoS ONE 2020, 15, e0238934. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zi, Y.; Li, C.; Peng, Y.; Zhu, X.; Guo, W. Dry matter accumulation, partitioning, and remobilization in high-yielding wheat under rice–wheat rotation in China. Agron. J. 2016, 108, 604–614. [Google Scholar] [CrossRef]
- Smith, M.R.; Rao, I.M.; Merchant, A. Source-sink relationships in crop plants and their influence on yield development and nutritional quality. Front. Plant Sci. 2018, 9, 1889. [Google Scholar] [CrossRef]
- Lotfi R, Ghassemi-Golezani K, Najafi N. Grain filling and yield of mung bean affected by salicylic acid and silicon under salt stress. J. Plant Nutr. 2018, 41 (14): 1778-1785.
- Gorooei, A.; Gaiser, T.; Aynehband, A.; Rahnama, A.; Kamali, B. The Effect of Farming Management and Crop Rotation Systems on Chlorophyll Content, Dry Matter Translocation, and Grain Quantity and Quality of Wheat (Triticum aestivum L.) Grown in a Semi-Arid Region of Iran. Agron. 2023, 13, 1007. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco J., M. Regulation and function of root exudates. Plant Cell and Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Zhu, Y.; Yao, X.; Tian, Y.; Liu, X.; Cao, W. Analysis of common canopy vegetation indices for indicating leaf nitrogen accumulations in wheat and rice. Inter. J. Appl. Earth Observ. 2008, 10, 1–10. [Google Scholar] [CrossRef]
- Dalal, R.C.; Strong, W.M.; Doughton, J.A.; Weston, E.J.; Copper, J.E.; Wildermuth, G.B.; Lehane, K.J.; King, A.J.; Holmes, J.E. Sustaining Productivity of a Vertisol at Warra, Queensland, with Fertilizer, No-tillage, or Legumes. Aust. J. Agric. Res. 1998, 38, 489–501. [Google Scholar] [CrossRef]
- Lotfi, R.; Pessarakli, M. Effects of Crop Rotation and Tillage on Winter Wheat Growth and Yield under Cold Dryland Conditions. Crops 2023, 3, 88–100. [Google Scholar] [CrossRef]
- Yin, X.; McClure, M.A. Relationship of Corn Yield, Biomass, and Leaf Nitrogen with Normalized Difference Vegetation Index and Plant Height. Agron. J. 2013, 105, 1005–1016. [Google Scholar] [CrossRef]
- Chu, X.; Guo, Y.; He, J.; Yao, X.; Zhu, Y.; Cao, W.; Cheng, T.; Tian, Y. Comparison of Different Hyperspectral Vegetation Indices for Estimating Canopy Leaf Nitrogen Accumulation in Rice. Agron. J. 2014, 106, 1911–1920. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient, pp. 321–362. In G.C. Papageorgiou and Govindjee (eds.). Chlorophyll a Fluorescence: A Signature of Photosynthesis. Springer. 2004, Dordrecht, the Netherlands.
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, R.; Abbasi, A.; Kalaji, H.M.; Eskandari, I.; Sedghieh, V.; Khorsandi, H.; Sadeghian, N.; Yadav, S.; Rastogi, A. The role of potassium on drought resistance of winter wheat cultivars under cold dryland conditions: Probed by chlorophyll a fluorescence. Plant Phys. & Biochem., 2022, 182(3), 45-54.
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Center, M.D.; Allakhverdiev, S.I.; Goltsev. V. Identification of nutrient deficiency in maize and tomato plants by in vivo chlorophyll a fluorescence measurements. Plant Phys. & Biochem., 2014, 81, 16–25. [Google Scholar]
- Serrago, R.A.; Alzueta, I.; Savin, R.; Slafer, G.A. Understanding grain yield responses to source-sink ratios during grain filling in wheat and barley under contrasting environments. Field Crop Res. 2013, 150, 42–51. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Pruneddu, G. Has long-term selection for yield in durum wheat also induced changes in leaf and canopy traits? Field Crops Res. 2008, 106, 68–76. [Google Scholar] [CrossRef]
- Wu, X.L.; Tang, Y.L.; Li, C.S.; Wu, C.; Huang, G.; Rong, M.A. (2014). Characteristics of grain filling in wheat growing in Sichuan Basin. Acta Agron. Scien. 2014, 40(2), 337−345.
- Cossani, C.M.; Slafe, G.A.; Savin, R. Do barley and wheat (bread and durum) differ in grain weight stability through seasons and water–nitrogen treatments in a Mediterranean location? Field Crops Res. 2011, 121, 240–247. [Google Scholar] [CrossRef]
- Motzo, R.; Giunta, F.; Pruneddu, G. The response of rate and duration of grain filling to long-term selection for yield in Italian durum wheat. Crop & Past. Sci. 2010, 61, 162–169. [Google Scholar]
- Hazratkulova, S.; Sharma, R.; Alikulov, S.; Islomov, S.; Yuldashev, T.; Ziyaev, Z.; Khalikulov, Z.; Ziyadullaev, Z.; Turok, J. Analysis of genotypic variation for normalized difference vegetation index and its relationship with grain yield in winter wheat under terminal heat stress. Plant Breed. 2012, 131, 716–721. [Google Scholar] [CrossRef]
- Kumar, M.; Mishra, V.K.; Chand, R.; Sharma, S.; Kumar, U.; Jaiswal, J.P.; Choudhary, M.; Mahato, A.; Ashutosh. ; Singh, P.; Joshi, A. NDVI and grain fill duration are important to be considered in breeding for terminal heat stress tolerance in wheat. J. Agron. & Crop Sci. 2023, 209, 489–501. [Google Scholar]
- Brennan, J.; Hackett, R.; McCabe, T.; Grant, J.; Fortune, R.A.; Forristal, P.D. The effect of tillage system and residue management on grain yield and nitrogen use efficiency in winter wheat in a cool Atlantic climate. Euro. J. Agron. 2014, 54:61–69.
- Malhi, S.S.; Nyborg, M.; Solberg, E.D.; Dyck, M.F.; Puurveen, D. Improving crop yield and N uptake with long-term straw retention in two contrasting soil types. Field Crops Res. 2011, 124, 378–391. [Google Scholar] [CrossRef]
- Damon, P.M.; Bowden, B.; Rose, T.; Rengel, Z. Crop residue contributions to phosphorus pools in agricultural soils: a review. Soil Biol. & Biochem. 2014, 74, 127–137. [Google Scholar]
- Evans, J.; Fettell, N.A.; Coventry, D.R.; O’Connor, G.E.; Walsgott, D.N.; Mahoney, J.; Armstrong, E.L. Wheat response after temperate crop legumes in south-eastern Australia. Aust. J. Agric. Res. 1991, 42, 31–43. [Google Scholar] [CrossRef]
- Geng, S.; Tan, J.; Li, L.; Miao, Y.; Wang, Y. Legumes can increase the yield of subsequent wheat with or without grain harvesting compared to Gramineae crops: A meta-analysis. Euro. J. Agron. 142, 126643.
- Cernay, C.; Makowski, D.; Pelzer, E. Preceding cultivation of grain legumes increases cereal yields under low nitrogen input conditions. Environ. Chem. Let. 2017, 16, 631–636. [Google Scholar] [CrossRef]
- Aslam, F.; Khaliq, A.; Matloo, A.; Tanveer, A.; Hussain, S.; Zahir, Z.H. 2017. Allelopathy in agro-ecosystems: a critical review of wheat allelopathy-concepts and implications. Chemoecol. 2017, 27, 1–24. [Google Scholar] [CrossRef]
- Mohammadi, R.; Amri, A. Genotype x environment interaction for durum wheat grain yield and selection for drought tolerance in irrigated and drought environments in Iran. J. Crop Sci. & Biotech. 2011, 14(4), 265–274.
Figure 1.
Changes in chlorophyll content index (CCI) of the wheat genotypes under different crop residue levels. Means followed by the same letter are not significantly different at P<0.05 according to the Duncan multiple range test. * and ** are significant at 5% and 1% probability levels, respectively.
Figure 1.
Changes in chlorophyll content index (CCI) of the wheat genotypes under different crop residue levels. Means followed by the same letter are not significantly different at P<0.05 according to the Duncan multiple range test. * and ** are significant at 5% and 1% probability levels, respectively.
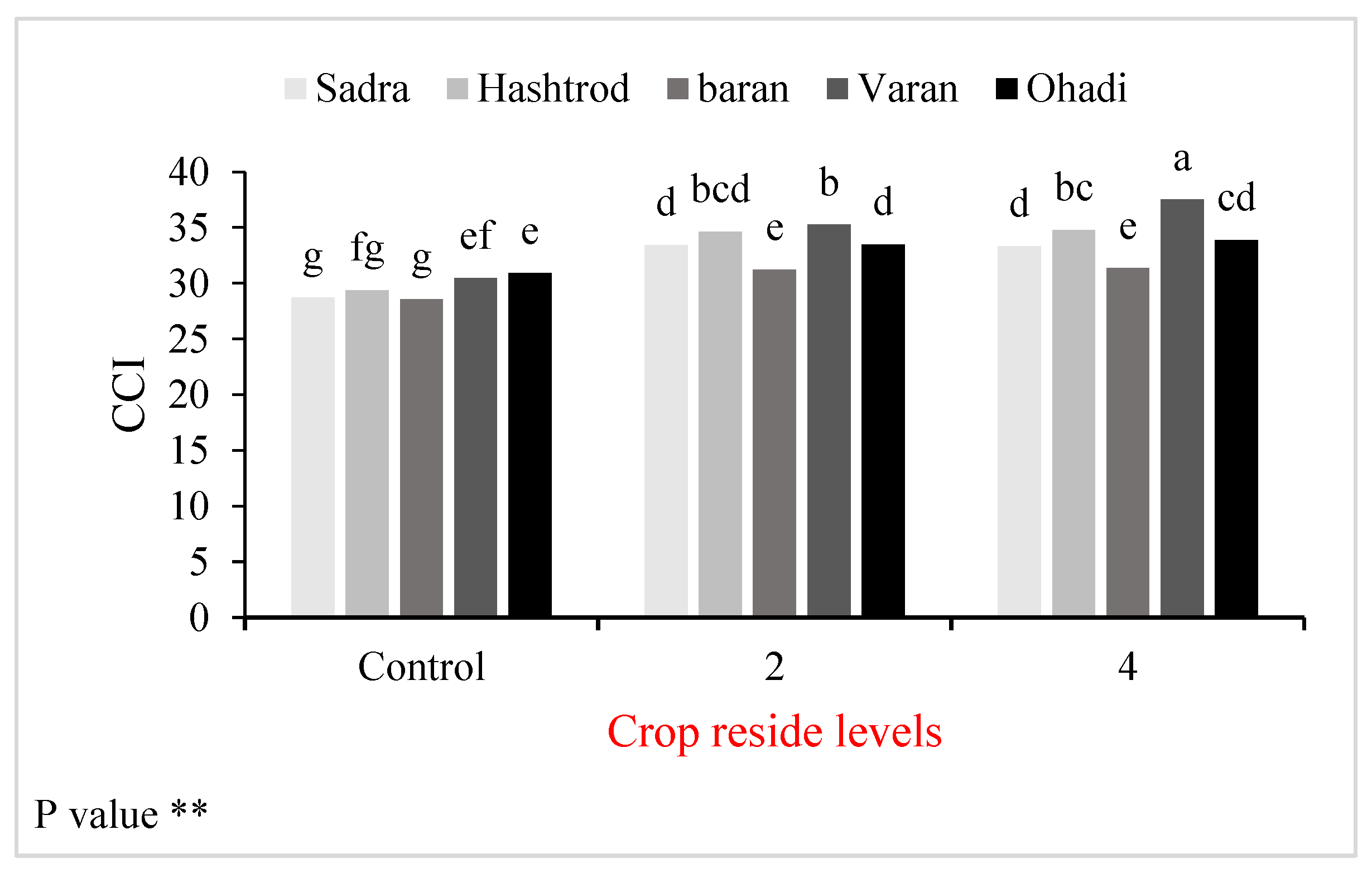
Figure 2.
Changes in Fv/Fo (A), Fv/Fm (B), and VI (C) of wheat genotypes under wheat-wheat (w-w) and forage (vetch)-wheat (f-w) rotation during 2017-2019. Means followed by the same letter are not significantly different at P<0.05 according to the Duncan multiple range test. Fv,Fo, FV/Fm, and VI are water-splitting complex in PSII, the maximum quantum efficiency of the PSII, and relative variable fluorescence at I step, respectively.
Figure 2.
Changes in Fv/Fo (A), Fv/Fm (B), and VI (C) of wheat genotypes under wheat-wheat (w-w) and forage (vetch)-wheat (f-w) rotation during 2017-2019. Means followed by the same letter are not significantly different at P<0.05 according to the Duncan multiple range test. Fv,Fo, FV/Fm, and VI are water-splitting complex in PSII, the maximum quantum efficiency of the PSII, and relative variable fluorescence at I step, respectively.
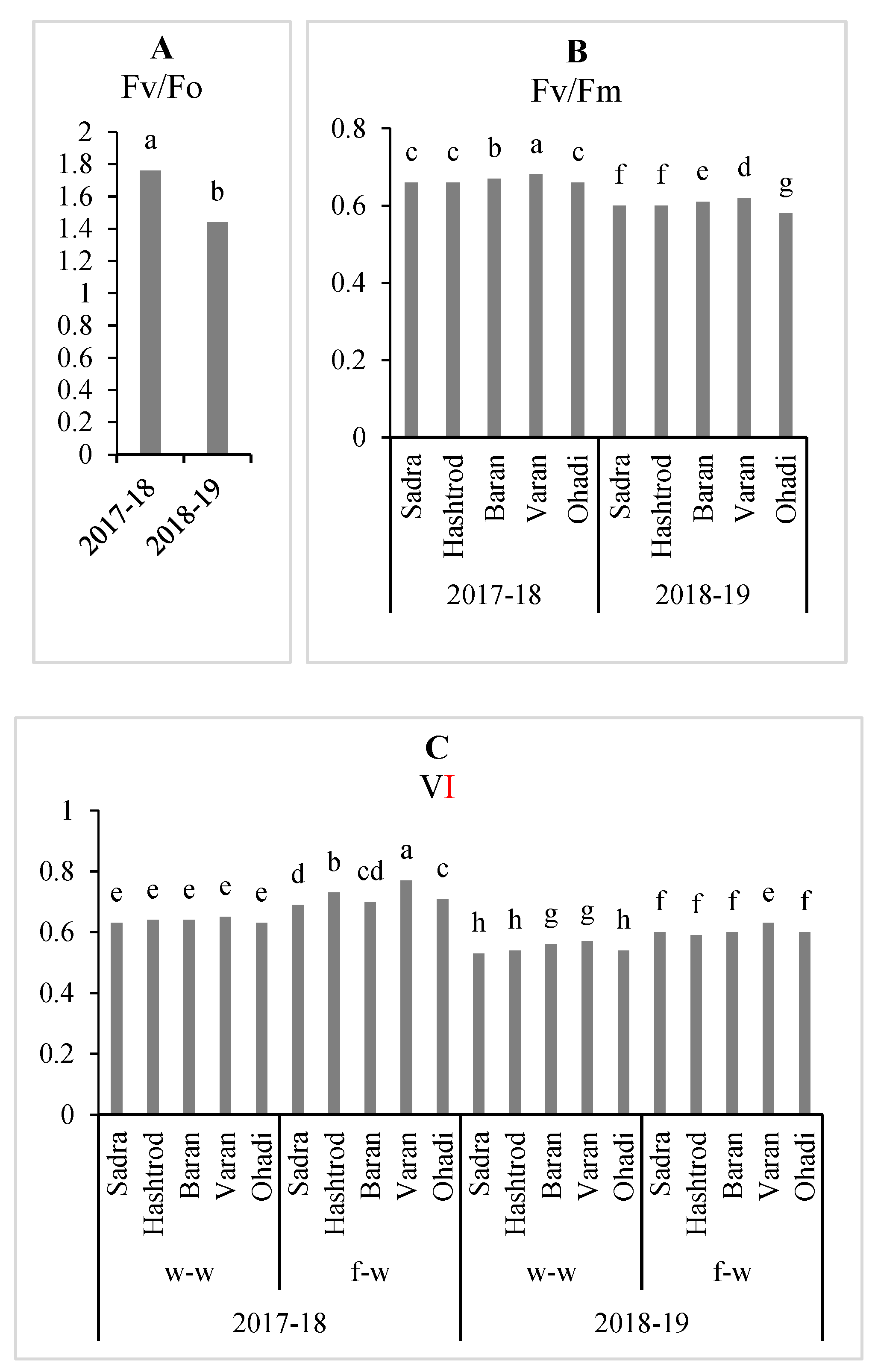

Table 1.
Monthly rainfall distribution and mean temperature all over the year during 2017-2019.
| Month | 2017-18 | 2018-19 | ||
| Precipitation (mm) |
Average temperature (˚C) | Precipitation (mm) |
Average temperature (˚C) |
|
| October | 0.2 | 11.4 | 9.7 | 13.7 |
| November | 36 | 8.5 | 47 | 5.5 |
| December | 48 | -0.8 | 91 | 2.3 |
| January | 29 | 1.4 | 41 | -2.5 |
| February | 85 | -0.9 | 86 | -1.2 |
| March | 80 | 4.3 | 56 | 0.2 |
| April | 55 | 8.6 | 116 | 5.2 |
| May | 67 | 10.4 | 43 | 9.6 |
| June | 23 | 16.7 | 4.2 | 18.1 |
| July | 0 | 24.6 | 0 | 22.7 |
| August | 0 | 24.8 | 0.5 | 23.6 |
| September | 1.8 | 20.0 | 0 | 19.2 |
Table 2.
Physical and chemical properties of the soils (from the field of the experiment site).
| Soil Depth (cm) | Soil Texture (%) | pH | K | P | TN | OC | CaCO3 | SP | ||
| Sand | Silt | Clay | mg kg-1 | % | ||||||
| 0-30 | 39 | 42 | 19 | 7.6 | 453 | 7.33 | 0.13 | 0.66 | 7.1 | 51 |
K: potassium; P: phosphorus; TN: total nitrogen; OC: organic carbon; CaCO3: calcium carbonate; SP: saturation percentage.
Table 3.
Leaf chlorophyll content index (CCI) and stomatal conductance (gs) of the wheat genotypes under two wheat and vetch pre-cropping systems during 2017-2019.
Table 3.
Leaf chlorophyll content index (CCI) and stomatal conductance (gs) of the wheat genotypes under two wheat and vetch pre-cropping systems during 2017-2019.
| CCI | 2017-18 | 2018-19 | ||
| P value ** | wheat-wheat | vetch-wheat | wheat-wheat | vetch-wheat |
| Sadra | 31.67 EFG | 32.74 DEF | 31.24 FG | 31.59 EFG |
| Hashtrod | 33.83 CD | 34.64 BC | 30.74 GH | 32.48 DEF |
| Baran | 29.61 HI | 32.38 DEF | 29.26 I | 30.27 GHI |
| Varan | 37.16 A | 35.99 AB | 31.44 EFG | 33.01 DE |
| Ohadi | 34.99 BC | 33.03 DE | 31.16 FG | 31.84 EFG |
| gs (mmol m-2 S-1) | ||||
| P value ** | ||||
| Sadra | 12.55 H | 13.92 E | 12.60 H | 14.51 D |
| Hashtrod | 13.34 FG | 14.34 D | 13.34 FG | 14.61 D |
| Baran | 13.77 E | 15.44 AB | 13.77 E | 15.22 BC |
| Varan | 13.72 EF | 15.66 A | 13.57 EFG | 15.79 A |
| Ohadi | 13.24 G | 15.15 BC | 13.84 E | 15.00 C |
Means followed by the same letter in each column are not significantly different at P<0.05 according to the Duncan multiple range test. * and ** are significant at 5% and 1% probability levels, respectively.
Table 4.
Normalized differences in vegetative index (NDVI) and stomatal conductance (gs) of the wheat genotypes under different levels of the wheat and vetch residue levels.
Table 4.
Normalized differences in vegetative index (NDVI) and stomatal conductance (gs) of the wheat genotypes under different levels of the wheat and vetch residue levels.
| NDVI | wheat-wheat | vetch-wheat | ||||||
| P value * | control | 2 T ha-1 | 4 T ha-1 | control | 2 T ha-1 | 4 T ha-1 | ||
| Sadra | 0.47 FG | 0.51 BC | 0.49 DE | 0.45 HI | 0.47 FG | 0.49 DE | ||
| Hashtrod | 0.45 HI | 0.48 EF | 0.45 HI | 0.45 HI | 0.48 EF | 0.50 CD | ||
| Baran | 0.46 GH | 0.51 BC | 0.48 EF | 0.46 GH | 0.52 AB | 0.52 AB | ||
| Varan | 0.48 EF | 0.52 AB | 0.48 EF | 0.49 DE | 0.53 A | 0.53 A | ||
| Ohadi | 0.43 J | 0.48 EF | 0.44 IJ | 0.44 IJ | 0.49 DE | 0.51 BC | ||
| gs | ||||||||
| P value ** | ||||||||
| Sadra | 11.23 N | 13.45 H-K | 13.06 KL | 12.16 M | 14.97 E | 15.53 D | ||
| Hashtrod | 12.85 L | 13.75 G-J | 13.43 IJK | 13.14 KL | 14.27 F | 16.03 C | ||
| Baran | 13.21 KL | 14.23 FG | 13.87 F-I | 13.33 JKL | 15.41 DE | 17.26 A | ||
| Varan | 12.98 KL | 14.02 FG | 13.95 FGH | 12.88 L | 16.78 B | 17.52 A | ||
| Ohadi | 13.22 KL | 13.95 FGH | 13.45 H-K | 13.37 I-L | 15.12 DE | 16.74 B | ||
Means followed by the same letter in each column are not significantly different at P<0.05 according to the Duncan multiple range test, and * and ** are significant at 5% and 1% probability levels, respectively.
Table 5.
Dryland wheat chlorophyll fluorescence parameters’ changes under different levels of wheat and vetch residues during 2017-2019.
Table 5.
Dryland wheat chlorophyll fluorescence parameters’ changes under different levels of wheat and vetch residues during 2017-2019.
| Treatments | ABS/RC | VJ | VI | PI | ||
| P value | ** | ** | ** | ** | ||
| 2017-18 | wheat-wheat | control | 2.206 GH | 0.45 G | 0.53 F | 0.34 F |
| 2 T ha-1 | 3.314 B | 0.66 D | 0.77 B | 0.61 B | ||
| 4 T ha-1 | 3.558 A | 0.50 F | 0.61 E | 0.48 E | ||
| vetch-wheat | control | 2.250 FG | 0.42 H | 0.51 F | 0.35 F | |
| 2 T ha-1 | 2.924 D | 0.73 C | 0.74 BC | 0.62 B | ||
| 4 T ha-1 | 2.236 G | 0.82 A | 0.92 A | 0.67 A | ||
| 2018-19 | wheat-wheat | control | 2.1673 HI | 0.42 H | 0.46 G | 0.32 F |
| 2 T ha-1 | 3.219 C | 0.62 E | 0.66 D | 0.51 DE | ||
| 4 T ha-1 | 3.333 B | 0.50 F | 0.53 F | 0.51 DE | ||
| vetch-wheat | control | 2.280 F | 0.43 H | 0.43 G | 0.32 F | |
| 2 T ha-1 | 2.7493 E | 0.66 D | 0.66 D | 0.54 CD | ||
| 4 T ha-1 | 2.129 I | 0.75 B | 0.72 C | 0.56 C | ||
Means followed by the same letter in each column are not significantly different at P<0.05 according to the Duncan multiple range test, and * and ** are significant at 5% and 1% probability levels, respectively. ABS/RC, VJ, VI, and PI are the light absorption per active reaction center, relative variable fluorescence at J step, relative variable fluorescence at I step, and photosynthesis performance index, respectively.
Table 6.
Chlorophyll fluorescence parameters’ changes of dryland wheat genotypes under different pre-cropping systems of wheat and vetch and their residue levels.
Table 6.
Chlorophyll fluorescence parameters’ changes of dryland wheat genotypes under different pre-cropping systems of wheat and vetch and their residue levels.
| Treatments | Fv/Fo | Fv/Fm | VJ | VI | PI | ||
| P value | ** | ** | ** | ** | ** | ||
| wheat-wheat | control | Sadra | 1.38 IJ | 0.59 KLM | 0.42 IJ | 0.49 KL | 0.34 L |
| Hashtrod | 1.46 HI | 0.60 JKL | 0.44 HI | 0.49 KL | 0.34 L | ||
| Baran | 1.28 J-M | 0.61 IJK | 0.44 HI | 0.50 K | 0.33 L | ||
| Varan | 1.29 JKL | 0.60 JKL | 0.43 HIJ | 0.5 K | 0.33 L | ||
| Ohadi | 1.19 LM | 0.58 LM | 0.43 HIJ | 0.48 KLM | 0.33 L | ||
| 2 T ha-1 | Sadra | 1.67 FG | 0.63 GHI | 0.63 F | 0.70 HI | 0.56 EF | |
| Hashtrod | 1.65 G | 0.64 FGH | 0.63 F | 0.71 H | 0.55 FG | ||
| Baran | 1.46 HI | 0.65 EFG | 0.65 E | 0.72 GH | 0.56 EF | ||
| Varan | 1.52 H | 0.67 CDE | 0.66 E | 0.75 EF | 0.60 BCD | ||
| Ohadi | 1.45 HI | 0.64 FGH | 0.63 F | 0.70 HI | 0.54 FG | ||
| 4 T ha-1 | Sadra | 1.18 M | 0.63 GHI | 0.51 G | 0.56 J | 0.53 GH | |
| Hashtrod | 1.18 M | 0.63 GHI | 0.51 G | 0.58 J | 0.49 IJ | ||
| Baran | 1.22 LM | 0.62 HIJ | 0.50 G | 0.57 J | 0.48 JK | ||
| Varan | 1.20 LM | 0.61 IJK | 0.51 G | 0.58 J | 0.51 HI | ||
| Ohadi | 1.26 LKM | 0.60 JKL | 0.49 G | 0.56 J | 0.46 K | ||
| vetch-wheat | control | Sadra | 1.53 H | 0.59 KLM | 0.45 H | 0.47 LM | 0.34 L |
| Hashtrod | 1.51 H | 0.58 LM | 0.41 J | 0.47 LM | 0.34 L | ||
| Baran | 1.37 IJ | 0.59 KLM | 0.42 IJ | 0.47 LM | 0.34 L | ||
| Varan | 1.26 KLM | 0.6 JKL | 0.42 IJ | 0.48 KLM | 0.33 L | ||
| Ohadi | 1.33 JK | 0.57 M | 0.42 IJ | 0.46 M | 0.33 L | ||
| 2 T ha-1 | Sadra | 1.83 E | 0.67 CDE | 0.70 CD | 0.68 I | 0.59 CD | |
| Hashtrod | 1.94 D | 0.67 CDE | 0.71 CD | 0.70 HI | 0.58 DE | ||
| Baran | 1.76 EF | 0.68 BCD | 0.69 D | 0.71 H | 0.58 DE | ||
| Varan | 1.97 D | 0.70 AB | 0.72 C | 0.74 FG | 0.59 CD | ||
| Ohadi | 1.66 G | 0.66 DEF | 0.66 E | 0.68 I | 0.58 DE | ||
| 4 T ha-1 | Sadra | 2.30 B | 0.68 BCD | 0.79 A | 0.79 CD | 0.61 BC | |
| Hashtrod | 2.25 B | 0.68 BCD | 0.77 B | 0.81 BC | 0.61 BC | ||
| Baran | 2.33 AB | 0.69 BC | 0.80 A | 0.77 DE | 0.62 B | ||
| Varan | 2.40 A | 0.72 A | 0.81 A | 0.89 A | 0.66 A | ||
| Ohadi | 2.13 C | 0.67 CDE | 0.77 B | 0.82 B | 0.61 BC | ||
Means followed by the same letter in each column are not significantly different at P<0.05 according to the Duncan multiple range test, and * and ** are significant at 5% and 1% probability levels, respectively. Fv/Fo, Fv/Fm, VJ, VI, and PI are water-splitting complexes in PSII, the maximum quantum efficiency of the PSII, relative variable fluorescence at the J step, relative variable fluorescence at the I step, and photosynthesis performance index, respectively.
Table 7.
Changes in grain weight, filling rate, filling duration, and yield of wheat genotypes under different pre-cropping systems and residue levels.
Table 7.
Changes in grain weight, filling rate, filling duration, and yield of wheat genotypes under different pre-cropping systems and residue levels.
| Treatments | 1000 GW (g) |
GFR (mg grain-1 day-1) |
GFD (day) |
GY (t ha-1) |
||
| P value | ** | ** | ** | * | ||
| wheat-wheat | control | Sadra | 35.26 S | 0.77 DE | 37.52 FG | 2.2941 K |
| Hashtrod | 35.78 R | 0.77 DE | 37.49 FG | 2.5258 J | ||
| Baran | 36.86 NO | 0.79 BC | 37.28 FGH | 2.5910 IJ | ||
| Varan | 37.07 L-N | 0.80 B | 37.27 FGH | 2.7381 GH | ||
| Ohadi | 36.51 PQ | 0.77 DE | 37.58 F | 1.8849 M | ||
| 2 T ha-1 | Sadra | 36.33 Q | 0.70 HI | 36.59 I | 2.5832 IJ | |
| Hashtrod | 36.43 Q | 0.71 GH | 36.92 F-I | 2.6409 HIJ | ||
| Baran | 37.74 HI | 0.77 DE | 36.64 HI | 2.9173 EF | ||
| Varan | 38.04 FG | 0.78 CD | 37.22 F-I | 3.0400 DE | ||
| Ohadi | 36.97 MNO | 0.75 F | 38.15 E | 2.1031 L | ||
| 4 T ha-1 | Sadra | 37.30 JKL | 0.61 L | 36.62 HI | 2.5365 J | |
| Hashtrod | 37.57 IJ | 0.64 K | 37.1 F-I | 2.6107 HIJ | ||
| Baran | 38.62 CD | 0.72 G | 37.11 F-I | 2.8448 FG | ||
| Varan | 38.86 C | 0.71 GH | 37.16 F-I | 3.0761 CD | ||
| Ohadi | 37.26 KLM | 0.71 GH | 36.89 GHI | 2.0758 L | ||
| vetch-wheat | control | Sadra | 36.73 OP | 0.75 F | 37.22 F-I | 2.6216 HIJ |
| Hashtrod | 37.247 LM | 0.76 EF | 37.21 F-I | 2.6599 HIJ | ||
| Baran | 37.61 HIJ | 0.82 A | 35.63 J | 2.7254 GHI | ||
| Varan | 37.95 GH | 0.83 A | 35.45 J | 2.6518 HIJ | ||
| Ohadi | 37.75 HI | 0.79 BC | 36.7 HI | 1.9110 M | ||
| 2 T ha-1 | Sadra | 37.73 HI | 0.69 I | 38.64 DE | 3.0793 CD | |
| Hashtrod | 37.66 HIJ | 0.69 I | 38.63 DE | 3.1949 C | ||
| Baran | 38.81 GHI | 0.75 F | 39.11 CD | 3.3286 B | ||
| Varan | 38.67 CD | 0.76 EF | 39.35 BC | 3.4491 B | ||
| Ohadi | 37.507 IJK | 0.72 G | 39.83 AB | 2.7091 G-I | ||
| 4 T ha-1 | Sadra | 38.243 EF | 0.65 K | 38.76 CD | 3.3456 B | |
| Hashtrod | 38.443 DE | 0.67 J | 38.89 CD | 3.3667 B | ||
| Baran | 40.107 A | 0.69 I | 39.85 AB | 3.7032 A | ||
| Varan | 40.387 A | 0.71 GH | 39.82 AB | 3.8099 A | ||
| Ohadi | 39.353 B | 0.69 I | 40.09 A | 2.8307 FG | ||
Means in each column followed by the same letter are not significantly different at P<0.05 according to the Duncan multiple range tests and * and ** are significant at 5% and 1% probability levels, respectively. 1000 GW, GFR, GFD, and GY are 1000 grain weight, grain filling rate, grain filling duration, and grain yield, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



