
Submitted:
14 August 2024
Posted:
15 August 2024
You are already at the latest version
Abstract
Soluble inorganic pyrophosphatase (sPPase), a pyrophosphate hydrolase, has been shown to play an important role in several physiological processes such as plant growth and development, metabolic activities, and response to abiotic stresses, but sPPase has been less studied in woody plants. In order to preliminarily verify whether the soluble inorganic pyrophosphatase in Eucommia ulmoides Oliver (E. ulmoides) has a role in resistance to drought stress, we amplified the EuSIP5 (E. ulmoides soluble inorganic pyrophosphatase 5) cDNA sequence from E. ulmoides by RT-PCR. In order to study the drought resistance of the Eucommia ulmoides soluble inorganic pyrophosphatase 5 (EuSIP5) gene, the cDNA sequence of the EuSIP5 gene was amplified from Eucommia ulmoides Oliver by RT-PCR. Bioinformatic analysis indicated that EuSIP5 might be an unstable amphipathic protein, mainly located in the cytoplasm. E. ulmoides, the highest expression of the EuSIP5 gene was found in male leaves and pericarp from April to October, and in leaves of E. ulmoides in July and September. During drought, the expression of EuSIP5 in seedling E. ulmoides leaves was significantly higher than that of the control. An overexpression vector harboring the EuSIP5 was constructed and introduced into the tobacco (Nicotiana tabacum cv. Xanthi). We found that during drought stress, the appearance of wilting was delayed by 4 days in transgenic tobacco plants (OE) compared with wild type plants (WT), and the expression of drought-related genes DET2, CYP85A1, P5CS, ERF1 and F-box was found to be increased in transgenic tobacco leaves, as well as the enzyme activities of the protective enzymes: peroxidase (POD), superoxide dismutase (SOD) and catalase (CAT) were significantly higher than those of the WT, and the malondialdehyde (MDA) content was lower than the WT. These results indicated that the introduction of EuSIP5 gene into tobacco increased the expression of drought-related genes, enhanced the anti-membrane oxidation ability of tobacco, and reduced the oxidative stress damage, thus improving the drought resistance of tobacco.
Keywords:
E. ulmoides
; EuSIP5
; transgenic
; tobacco
; drought resistance
1. Introduction
Drought stress is one of the major abiotic stresses facing plant growth globally, and it has significant negative impacts on plant growth, development and reproduction. Under drought conditions, water uptake in plants is limited, leading to a reduction in intracellular water, damage to cell membrane structure, and reduced cell activity. In addition, drought causes changes in osmoregulatory substances in plants, affecting intracellular ion balance and signalling, which in turn affects plant growth, development and reproduction [1]. Improving the ability of plants to respond to drought stress is essential for agricultural production and the sustainable development of ecosystems. Scientists can improve crop yields and quality under drought conditions by discovering drought-resistant genes in plants and using these genes to breed more drought-tolerant crop varieties [2].
Guizhou Province is located on the eastern side of the Yunnan-Guizhou Plateau in southwestern China, and the region is affected by monsoon climate, which is variable and prone to droughts and floods. Despite the abundant rainfall, droughts caused by engineered water shortages in karst landscapes are frequent, widespread and long-lasting, resulting in reduced food production, frequent secondary disasters and ecological degradation, and crops must be drought-tolerant in order to survive [3].
Eucommia ulmoides Oliver (E. ulmoides) is a rare and endangered plant in China. It has medicinal and edible value [4,5]. Its leaves can be used as tea and its bark as medicinal materials, with anti-hypertension, reduce blood sugar, reduce blood lipids, and anti-cancer and other health effects [6,7,8]. E. ulmoides is mainly distributed in Guizhou, Sichuan, Chongqing, Yunnan, Shaanxi, Hunan, Hubei and so on. It can be successfully introduced into arid and semi-arid areas because of its strong vitality and adaptability. Many functional genes have been cloned and studied in E. ulmoides genome, such as resistance to adversity and insect and disease [9,10,11,12]. By studying the stress-related genes in E. ulmoides, the scientists delved into the expression patterns of E. ulmoides's genes in harsh environments and used these functional genes to improve other crops, to improve the stress resistance of crops [13,14].
In recent decades, researchers have successively focused on the adversity response function of the inorganic pyrophosphatase gene family. PPases are a class of hydrolytic enzymes widely found in plants, animals and microorganisms, and such enzymes play important roles in several physiological processes such as plant growth and development, sugar metabolism, and response to salt and drought stress [15]. PPases is able to hydrolyze inorganic pyrophosphate (PPi) produced during plant growth and metabolism into two inorganic orthophosphates (Pi) if these PPi are not promptly cleared, excessive accumulation may result in cytotoxicity and even cell death [16]. The process by which PPases hydrolyze PPi not only provides energy to plant cells but also helps to maintain the homeostasis of intracellular PPi [17]. PPases may play an important role in plant responses to abiotic stresses. Current research has divided PPases into two broad categories: a group of enzymes that are widely found in plant cells, and they are divided into two main families: membrane-integral pyrophosphatase (H+-PPase) and soluble inorganic pyrophatase (s-PPase) [18]. s-PPase is subdivided into three subtypes in nature, namely, s-PPase I, s-PPase II and s-PPase III. The s-PPase classification in plants belongs to s-PPase I [19]. Chen et al. [20] cloned seven soluble Inorganic pyrophosphatase genes ZmPPases from maize (Zea mays L.) . The expression levels of these genes were up-regulated or down-regulated under salt stress and drought stress, which indicated that ZmPPases might be involved in plant responses to salt stress and drought stress. The expression of soluble inorganic pyrophosphatase genes HbSIP1/2/3 in Brazilian rubber tree [Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg.] under treatments such as ethylene (ET), jasmonic acid (JA), high temperature, and low temperature occurred all up-regulated or down-regulated expression of varying degrees, suggesting a potential role for the inorganic pyrophosphatase genes in the plant response to environmental stresses [21]. George et al. [22] , after silencing the s-PPaseI gene in tobacco, found that PPi accumulation, decreased chlorophyll, Carotenoid and starch content, and decreased ABA content affected stomatal closure in tobacco, thus reducing the drought resistance of tobacco.
The AP2 / ERF (ethylene transcription factor) family is a widespread group of mega-family transcription factors in plants that are involved in regulating plant responses to drought stress through interactions with downstream target genes [23,24,25,26]. MYC transcription factors regulate gene expression associated with drought stress response by participating in the ABA signalling pathway in response to drought stress in plants [27,28]. The mechanism of action of P5CS (pyrroline-5-carboxylic acid synthase) in plant drought tolerance genes mainly involves its role as a key enzyme in the proline biosynthesis pathway, which increases the proline content in plant cells by catalysing the conversion of glutamate to proline [29]. The protein encoded by the CYP85A gene is one of the key enzymes in the biosynthetic pathway of the endogenous plant hormone oleuropein lactones (BRs). Oleuropein lactones are an important class of phytohormones, which play an important role in plant growth and development and in response to environmental stresses [30]. The protein encoded by the DET gene is also a rate-limiting enzyme in the BRs biosynthetic pathway. By affecting the biosynthesis of BRs, it in turn affects the drought tolerance of plants [31]. F-box proteins play an important role in plant drought tolerance; they are key parts of the ubiquitin-26S proteasome system (UPS), which regulates plant adaptation to drought by recognising and degrading specific target proteins [32,33].
In this experiment, we screened and identified EuSIP5, a soluble inorganic pyrophosphatase gene of E. ulmoides, from the E. ulmoides whole gene annotation library constructed in the previous stage of the project team. We then proceeded to perform bioinformatics analysis and predicted the protein structure. Following the transformation of tobacco (Nicotiana tabacum cv. Xanthin) with an overexpression vector constructed using the EuSIP5 gene, preliminary analyses were conducted to ascertain whether the EuSIP5 protein exhibits drought-resistant properties. This involved measuring the relative gene expression in overexpressed tobacco, protective enzyme activities, and phenotypic observations. The experimental findings may provide a partial theoretical basis for soluble inorganic pyrophosphatase in the study of genetic engineering to improve drought resistance in plants.
2. Materials and Methods
2.1. Materials
The material used for genetic transformation and subcellular localization is tobacco, which is kept by the Key Laboratory of Plant Resources Conservation and Germplasm Innovation in Mountainous Region (Ministry of Education) of the Guizhou University. E. ulmoides was grown on the base of the laboratory. We collected young bark, leaves, male flowers and fruits from 18-year-old mature E. ulmoides females and males at the base.
The Escherichia coli (E. coli) strain DH5α, Agrobacterium tumefaciens (A. tumefaciens) GV3101 and overexpression of the empty vector (pSH737) were preserved and provided by our laboratory. The subcellular localisation empty carrier (pEGOEP35S-H-GFP) was purchased from Wuhan Aidijing Biotechnology (Wuhan, China).
2.2. Method
2.2.1. The Process of Gene Cloning and Subsequent Bioinformatics Analysis
The EuSIP5 protein-coding gene was identified from the whole genome annotation database of E. ulmoides, constructed by this project team. Primers specific to the EuSIP5 gene were designed for amplification and sequencing by NCBI (Table S1). The coding sequence was amplified by PCR and sequenced, resulting in the EuSIP5 cDNA sequence, which was determined to be 552 bp in length.
The use of online websites and software to bioinformatically analyse the cloned sequences and predict the physical and chemical properties, functions and phylogeny of the proteins: The physico-chemical properties of proteins were analysed using the ‘protparam’ programme on the online website ExPASy (https://web.expasy.org/protparam/); The hydrophilicity and hydrophobicity of proteins were analysed using ProtScale (https://web.expasy.org/protscale/); The structural analysis of conservative domains employs the ‘CD-Search’ tool, which is hosted by the National Center for Biotechnology Information Bureau (NCBI) (https://www.ncbi.nlm.nih.gov/cdd/); The analysis of protein phosphorylation sites was conducted using the ‘NetPhos’ tool (http://www.cbs.dtu.dk/services/NetPhos/); The protein signal peptide sequence was predicted using the web-based tool ‘SignalP’ (http://www.cbs.dtu.dk/services/SignalP/); The protein transmembrane regions are predicted using the ‘TMHMM’ server software version 2.0 (http://www.cbs.dtu.dk/services/TMHMM/); The protein 3D structures are predicted using the SWISS-MODEL software (https://swissmodel.expasy.org); The amino acid sequences of EuSIP5 were compared with those of other plant s-PPases using the software program DNAMAN. A phylogenetic tree was constructed using Mega 7 to illustrate the evolutionary relationships of soluble inorganic pyrophosphatase family genes. The soluble inorganic pyrophosphatase sequences of the other species are shown in Table S2.
2.2.2. Expression Analysis of the EuSIP5 Gene in E. ulmoides
The materials were collected from the field perennial E. ulmoides. In the same year, different tissues (young stem bark, young leaves, flowers, fruits) of male and female plants were collected at different month (from April to October), total RNA was extracted and reverse transcribed into cDNA. RNA was extracted using the OMEGA Plant Total RNA Extraction Kit and reverse transcription was performed using the Sevier Reverse Transcription Kit (Wuhan, China).
2.2.3. Vector Construction
Based on the cloned EuSIP5 cDNA sequence, specific primers eGFP-F and eGFP-R were designed (Table S1), the target fragment was amplified and recovered, and the target fragment and plasmid empty vector were assembled with pEGOEP35S-H-eGFP as the original vector using the Golden Gate cloning method. pEGOEP35S-H-EuSIP5-eGFP fusion vector was transferred into E. coli DH5α by heat-stimulation method. 35S-F and eGFP-cx primers (Table S1) were designed to verify and sequence the colonies, and plasmids were extracted and sequenced after picking out the positive spots and culturing in an expanded culture to determine the success of constructing the target fragments into the expression vectors.
Vector selection of GUS reporter genes as marker genes. The target gene fragments were integrated into the pSH737 vector using EcoRⅠand XbaⅠas digestion sites. The EuSIP5 plant overexpression vector pSH737-35S-EuSIP5-Nos was constructed by a one-step cloning method using the original vector containing the laboratory constructed Kana resistance screening pSH737. E. coli DH5α containing plasmid pSH737 was inoculated into LB solid medium containing 100 mg·L-1 Kana, and the plasmid DNA of the Kana-screened positive E. coli single colony was extracted, and the collected recombinant plasmid DNA was stored at -20℃ for backup use. The previously obtained EuSIP5 gene target fragment and pSH737 plasmid were double digested, and the obtained target fragment was gelatinised, and the digested target fragment and digested pSH737 plasmid were ligated. After correct sequencing, the target fragment was transferred to Agrobacterium GV3101 and stored at -80°C.
2.2.4. Subcellular Localization
Transient expression of the EuSIP5 fusion protein was achieved by injection into tobacco leaves. The constructed pEGOEP35S-H-EuSIP5-eGFP vector plasmid was electrotransformed into A. tumefaciens GV3101, and the A. tumefaciens colonies were scraped from the solid Petri dishes with inoculation loop after overnight incubation at 37°C.Tobacco plants were selected at 3-4 weeks of age, and the back of the leaf was gently picked up with a 1 mL defoaming syringe needle and injected from the lower epidermal surface of the leaf using a syringe with the needle removed and marking the waterlogged area. The water-labelled areas were marked and the injected tobacco plants were incubated at 25±2 °C, 16/8 h in low light for 2 days for observation, and the labelled tobacco leaves were removed and made into slides, which were observed and photographed under a laser confocal microscope.
2.2.5. Genetic Transformation and Identifcation of Tobacco
The leaf disc method was used to genetically transform wild-type (WT) aseptic tobacco seedlings cultured for 3 ~ 4 weeks. The leaves of the WT tobacco were cut into leaf discs of approximately 1.0 cm2 and A. tumefaciens strain GV3101 containing the pSH737-35S-EuSIP5-NOS vector was cultured overnight in YEP liquid medium at 28°C with shaking (180 rpm). When the bacteria were collected by centrifugation and resuspension, the final optical density (OD) at 600 nm was 0.5. Leaf discs were inoculated with resuspended bacterial solution, soaked for 5 min and then drained of excess bacterial solution with sterile filter paper, co-cultured under dark light for 2 d, transferred to screening medium containing Kana antibiotic and the medium was changed every 2 weeks; when the differentiated shoots grew to 1 ~ 2 cm, they were then transferred to rooting medium to induce rooting and to cultivate strong seedlings for subsequent treatments (The incubation conditions were 25±2 °C, 16/8 h diurnal photoperiod). Transgenic plants are eventually recovered by high temperature and pressure inactivation and incineration.
Since the overexpression vector pSH737 uses a GUS reporter gene as a marker gene, we performed a preliminary validation using GUS chemical tissue staining. We soaked the leaves of generation tobacco seedlings grown on antibiotic-containing Kana medium with GUS stain for 15 min in 25 Kpa vacuum before transferring them to a 37°C constant-temperature incubator for overnight incubation. The leaves were then eluted with ethanol and the results of GUS staining were observed. DNA from the positive GUS-stained transgenic tobacco lines was extracted for further verification using the TIANGEN Plant Genomic DNA Kit (Beijing, China). After extraction, 1 μL of DNA template was taken for PCR using specific primers EuSIP5-F and EuSIP5-R to verify whether the EuSIP5 gene was integrated in OE tobacco.
2.2.6. Quantitative Fluorescence Real-Time PCR (qRT-PCR)
qRT-PCR specific primers were designed using Primer Premier 5 software. Total plant RNA was extracted according to the OMEGA kit instructions, and the RNA reverse transcription to cDNA step was performed according to the Sevier Reverse Transcription Kit. The fluorescent dyes used were Genstar fluorescent dyes, and the most suitable thermal cycling system for qRT-PCR was obtained by mapping as follows: 95°C for 5 min pre-denaturation and then enter the cycle. 95°C (30s) and 58°C (30s), 40 cycles, and fluorescent signals were collected at each annealing and extension (58°C). After all qRT-PCR cycles were completed, a melting curve with a temperature gradient of 65-95°C (0.1°C/s) was obtained and used to determine the uniqueness of the product
2.2.7. Drought Stress Treatment of Transgenic Tobacco
To verify the response of EuSIP5 transgenic tobacco to drought stress, wild-type plants (WT) and transgenic plants (OE) of the same strain with the same growth conditions were collected and subjected to natural drought treatment without watering for 10 d. Sampling times were set as 0 d, 2 d, 4 d, 6 d and 8 d. Samples were watered every 3 days during the normal growth period of the material in the climate chamber, and the third day after the last watering was treated as 0 d of drought treatment, and then samples were taken every 2 days and observed and photographed. In the artificial climate chamber, the material was watered every 3 days during the normal growth period, and the third day from the last watering was treated as 0 d of drought treatment, and then the samples were sampled every 2 days and observed and photographed, and the samples were stored in the refrigerator at -80°C after quick freezing in liquid nitrogen. RNA was extracted and reverse transcribed into cDNA for qRT-PCR analysis. The tobacco NtEF1α gene was used as an internal reference gene. Three replicates were set up for each sample, and the relative expression of different genes in tobacco was calculated using the 2-ΔΔCt method.
2.2.8. Expression Analysis of Genes Related to Drought Resistance in Transgenic Tobacco
The expression of genes related to drought tolerance in transgenic tobacco was analysed by qRT-PCR, and tobacco NtEF1α was selected as an internal reference gene. Genes associated with drought tolerance in tobacco: NtERF1 (>D38123.1), NtP5CS (>GBEA01096483.1), NtCYP85A1 (>DQ649022.1), NtDET2 (>GBEA01057209.1), NtF-Box (>GBEA01054484.1), and qRT-PCR specific primers were designed using Primer Premier 5 software (Table S1).
2.2.9. Determination of Protective Enzyme Activity
Three WT plants and three OE plants of the same strain under the same growth conditions were each taken, and the leaves of the same part were drought-treated for 0d, 2d, 4d, 6d and 8d. The activities of catalase (CAT), superoxide dismutase (SOD), peroxidase (POD) and the content of malondialdehyde (MDA) were measured according to the instructions, and the activities and contents of WT plants were taken as the control group. The CAT enzyme activity assay kit and malondialdehyde (MDA) content assay kit were purchased from Servicebio (Wuhan, China), the SOD enzyme activity assay kit was purchased from Nanjing Jianjian (Nanjing, China), and the POD enzyme activity assay kit was purchased from Solarbio (Beijing, China).
2.2.10. Statistical Analysis
All data are expressed as the sample mean ± standard error of the three biological replicates and their respective technical replicates. Statistical analyses were performed using Two-way ANOVA, and multiple comparisons were tested using Dunnett's test (for data on expression of drought-related genes in tobacco) and Tukey's test (for relative expression of the EuSIP5 gene in E. ulmoides). The Shapiro-Wilk test for normal distribution and Levene's test of variance were used for the residuals of each variance. Significance was indicated by both asterisks (*) and letters; in the asterisk method, * indicates a significant difference and ns indicates a nonsignificant difference; in the letter method, different letters indicate a significant difference and the same letter indicates a nonsignificant difference. which was conducted using SPSS 26.0 software (SPSS Inc., Chicago, IL, USA). All figures were plotted in the GraphPad Prism 9.0.
3. Results
3.1. Cloning and Analysis of the EuSIP5 Gene
Specific primers were designed according to the cDNA sequence of the EuSIP5 gene, and the cDNA of E. ulmoides cDNA was used as a template for PCR amplification. The image result of 1% agarose gel electrophoresis showed a bright band between 500bp and 750bp. The sequencing result was 552bp, which matched the sequence comparison result in the database (Figure S1).
According to the sequencing results, the cDNA sequence from which the gene was cloned was 552 bp, and bioinformatic analysis showed that the sequence contained an open reading frame (ORF) of 381 bp, encoding 126 amino acid residues, and bioinformatic analysis showed that the theoretical relative molecular mass (MW) of the EuSIP5 protein was 20.651 kDa, and the isoelectric point (pI) was 6.44; The total number of acidic amino acid residues (Asp+Glu) was 26, the total number of basic amino acid residues (Arg+Lys) was 24, the instability index was 41.51, the aliphatic index was 85.82, the grand average of hydropathicity (GRAVY) was -0.334, indicating that this protein is an unstable amphiphilic protein; The CELLO prediction system server was used to predict the subcellular localisation of the protein in the cytoplasmic and nuclear membranes; the conserved structural domains of the protein were analysed using the NCBI online tool CD-search; Conserved domain analysis of EuSIP5 shows that it is a soluble inorganic pyrophosphatase of the pyrophosphatase superfamily (Figure 1a). Comparison of EuSIP5 with homologous proteins of other species. The results showed that EuSIP5 shared 88.10% similarity with the homologous proteins of other species (Figure 1b). Phylogenetic analysis of the amino acid sequence of EuSIP5 with sequences from other species. The results showed that the soluble inorganic pyrophosphatase EuSIP5 in E. ulmoides was closest in evolutionary relationship to the soluble inorganic pyrophosphatases in Camellia lanceoleosa and Cornus florida (Figure 1c).
3.2. Spatio-Temporal Expression Characteristics of the EuSIP5 Gene
In order to understand the expression characteristics of the EuSIP5 gene in male and female strains of E. ulmoides, the relative expression of the gene in different tissues collected at different times was analysed using qRT-PCR. It can be seen that the EuSIP5 gene was expressed to varying degrees in all tissues of both male and female Eucommia plants and was tissue specific. Male leaves showed the highest expression, followed by male stem bark > female leaves > female stem bark > female fruit > female root > male root > female flower > male flower, suggesting that the gene may function mainly in nutrient organs (Figure 2a). The overall level of EuSIP5 gene expression was higher than that of female stems and male stems in both female and male leaves from April to October, and the highest expression of the gene in female leaves was reached in July, which was 9.63 times higher than that of female stems in the same month. In male leaves, EuSIP5 gene expression was highest in September, which was 12.51 times higher than that of male stems in the same month (Figure 2c-d). The expression of the EuSIP5 gene in different parts of the plant showed obvious spatial and temporal specificity, especially in the period and part of the plant where metabolism was strong, and it was suggested that this gene was closely related to nutritional growth and metabolic activities. In natural drought treatment of Eucommia seedling for 15 d, the expression of EuSIP5 gene was significantly increased (P<0.05) compared with the initial level, and the expression was 1.41 times of the initial level. It indicated that EuSIP5 gene would respond to drought stress (Figure 2b).
3.3. Subcellular Localization
Six variants of soluble inorganic pyrophosphatase (AtPPA1-AtPPA6) have been identified in Arabidopsis thaliana [34,35], and subcellular localisation experiments using green fluorescent protein (GFP) as a marker have shown that AtPPA1-AtPPA5 are predominantly distributed in the cytoplasm, whereas AtPPA6 is located in the chloroplasts [36]. Twelve soluble inorganic pyrophosphatases (PbrPPA1-PbrPPA12) have been identified in Pear, of which GFP-tagged subcellular localisation experiments on the PbrPPA1, PbrPPA2, PbrPPA5, PbrPPA9 and PbrPPA10 proteins showed that the PbrPPA1, PbrPPA2 and PbrPPA5 fusion proteins were localised in the cytoplasmic lysate and bound to the nuclear membrane, whereas the PbrPPA9-GFP and PbrPPA10-GFP fusion proteins were detected in the chloroplasts [37].
We used EcoR I and Hind III as cleavage sites for double digestion verification, and the vector was named pEGOEP35S-H-EuSIP5-eGFP after successful vector verification (Figure S2a). We found that EuSIP5 protein fluorescence was expressed on the cytoplasm of cells by laser confocal microscopy, and that the soluble inorganic pyrophosphatase EuSIP5 may act on the cytoplasm (Figure 3).
3.4. Genetically Modified Tobacco and Identification of Transgenic Positive Plants
The overexpression vector of the EuSIP5 gene was constructed and named pSH737-35S-EuSIP5-Nos after correct sequencing (Figure S2b). Agrobacterium containing pSH737-35S-EuSIP5-Nos vector was introduced into tobacco, the Agrobacterium-infected tobacco leaves differentiated into small green shoots after 4 weeks, and the green shoots were cut off and transferred to rooting medium, and then transplanted to soil after the small shoots grew and the root system was sufficiently developed (Figure S3a-d). Obtain transgenic tobacco plants.
GUS histochemistry showed that the leaves of WT tobacco plants were not stained blue, whereas the leaves of OE plants were stained blue (Figure S3e), providing preliminary confirmation that pSH737-35S-EuSIP5 has been transferred into tobacco. PCR characterisation showed that the transgenic tobacco plants amplified a gene-specific band of 552 bp, whereas the WT and OE tobacco plants did not, indicating that the EuSIP5 gene had been transferred into the tobacco genome (Figure S3f). This indicates that the EuSIP5 gene has been integrated into the tobacco genome. Among all the transgenic lines, three transgenic lines, OE3, OE5 and OE10, with the best growth and highest expression were selected for subsequent experiments.
3.5. Effect of Trans-EuSIP5 Gene on Drought Tolerance in Tobacco
The expression of the EuSIP5 gene and tobacco drought-related genes in WT and OE plants from 0-8 d of natural drought treatment was analysed by qRT-PCR. The experimental results showed that the expression of DET2 gene in OE plants showed a decreasing and then increasing trend, and the expression was significantly lower than that of WT plants at 2 d of drought treatment, and then the expression increased continuously, reaching 6.63-fold of that of WT plants at 8 d after treatment (Figure 4a); The expression level of the CYP85A1 gene in OE plants was unaffected during 0-2 d of treatment, and the expression showed an increasing and then decreasing trend after 2 d. The highest value was reached at 4 d of treatment, which was 5.73 times higher than that of WT plants (Figure 4b); The expression of P5CS, ERF1 and F-box genes in OE plants was higher than that of the WT plants after treatment, with F-box having the highest expression at d 6 of treatment, which was 1.75 times higher than that of the WT (Figure 4c); The expression of P5CS and ERF1 reached the highest levels at 4 d of treatment, which were 2.82 and 5.90 times higher than the WT, respectively (Figure 4d-e).
Observation of the phenotypes showed that wilting and stem collapse began on day 4 of natural drought in the WT tobacco, whereas wilting of the leaf tips began on day 8 of treatment in the OE tobacco, but the overall symptoms were less severe (Figure 5).
3.6. Effect of Trans-EuSIP5 on the Activity of Tobacco Protective Enzymes
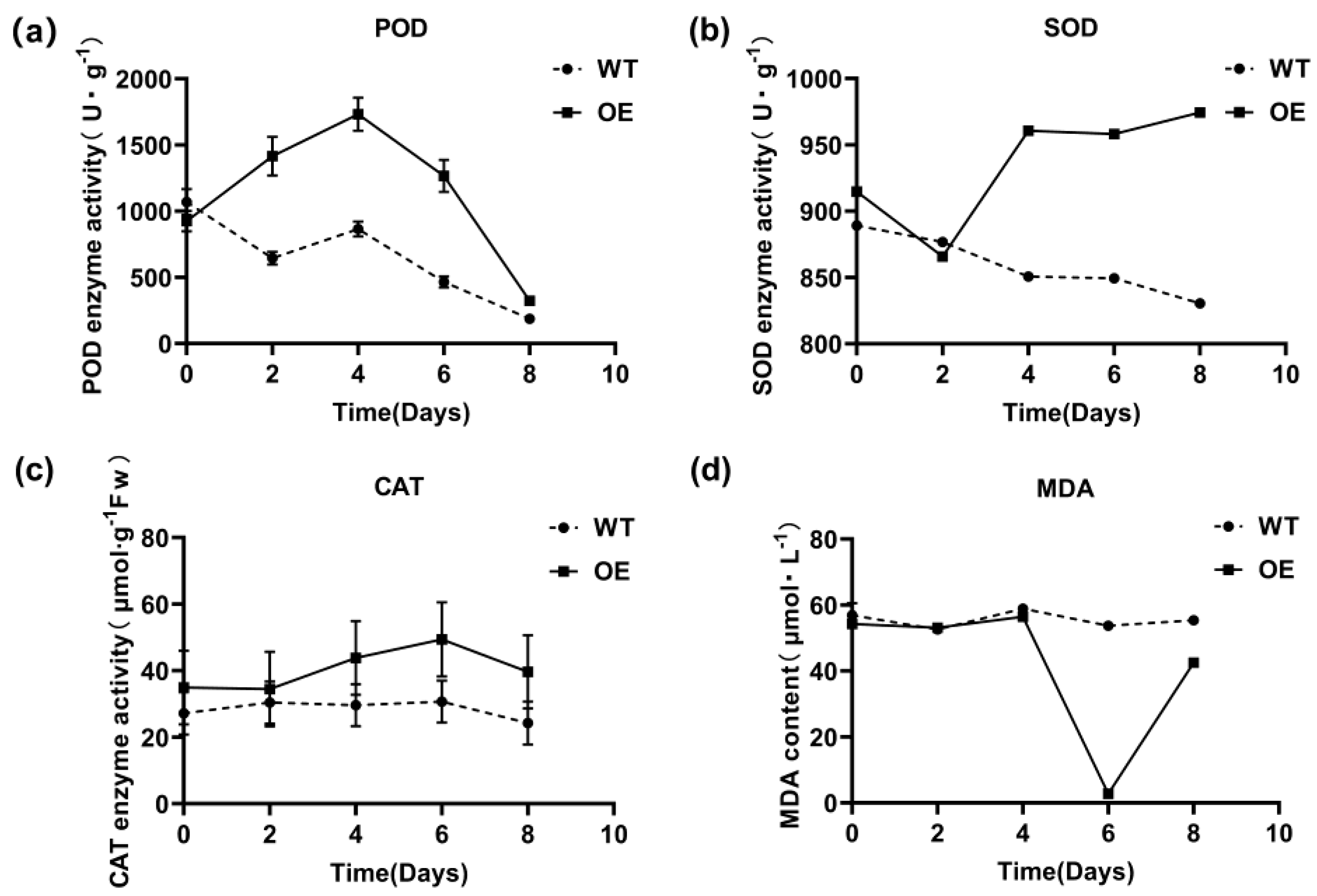
The protective enzyme activities of WT and OE tobacco plants were quantified following a 0-8 d period of natural drought treatment. The experimental results demonstrated that the activities of the POD, SOD and CAT enzymes were elevated in the OE tobacco plants relative to the WT. The POD enzyme exhibited the highest activity, reaching 1732.65 U·g-1 at the 4 d mark, which was 2.00 times higher than that of the WT (Figure 6a); The highest activity of the SOD enzyme was 960.663 U·g-1 after 8 days of treatment. This was 1.13 times higher than that of the WT (Figure 6b); The highest level of CAT activity was observed at day 6 of the treatment period, with an activity value of 49.4174 μmol·g-1Fw, which was 1.60 times higher than that of the WT control (Figure 6c); The MDA content in OE tobacco plants was found to be lower than that of the WT following treatment. However, after 6 d of drought treatment, the MDA content was drastically reduced, with a content of 2.77 μmol·L-1 observed, which was 19.36-fold lower than that in the WT (Figure 6d).
4. Discussion
Globally, drought is the primary driver of reduced agricultural output, with annual losses attributed to drought accounting for 53% of the total, a figure that exceeds the combined impact of other natural disasters [38]. The objective of this study was to examine the potential of the EuSIP5 gene, a soluble inorganic pyrophosphatase from E. ulmoides, in enhancing plant drought tolerance. Through the cloning, bioinformatics analysis, overexpression vector construction and expression of the EuSIP5 gene in tobacco, it was found that the overexpression of the EuSIP5 gene could improve the tolerance of tobacco to drought stress.
The secondary structure of the protein encoded by the ZmPPases gene, a soluble inorganic pyrophosphatase isolated from maize, is predominantly composed of α helices and β folds. This structural feature enables s-PPase to carry out its catalytic reaction in an efficient manner. The molecular weight of the protein in question ranges from 22.8 to 25.6 kDa, while the isoelectric point ranges from 4.84 to 6.24 [20]. In contrast, the results of the present study demonstrated that the secondary structure of EuSIP5 protein comprises three distinct secondary structures, namely α helix, extended linkage, and irregular coiled, with a molecular mass of 20,108.47 kDa and an isoelectric point of 9.79. This diversity may be attributed to the evolution of s-PPase genes among different species. Furthermore, structural domain analysis revealed that the EuSIP5 protein contains a soluble inorganic pyrophosphatase structural domain. Subsequent sequence comparison and phylogenetic analysis demonstrated that the protein exhibited high correlation with the amino acid sequence of homologous proteins and possessed the structural motif of inorganic pyrophosphatase. Subcellular localisation demonstrated that EuSIP5 was localised to the cytoplasm, a result consistent with that of Gutiérrez-Luna, Navarro de la Sancha, Valencia-Turcotte, Vázquez-Santana and Rodríguez-Sotres [36].The preceding findings led to the hypothesis that the EuSIP5 gene is a soluble inorganic pyrophosphatase gene in E. ulmoides.
In a study of the EuERD16 gene in perennial adult E. ulmoides, Li Bo [39] observed that expression was higher in March and May viviparous leaves and July fruits. The EuEFD16 gene was found to be consistently upregulated in tissues that divide and grow faster at the early stage of growth and development. In this study, we analysed the spatio-temporal expression characteristics of the EuSIP5 gene in E. ulmoides. The results demonstrated that EuSIP5 plays a role in the growth and development of E. ulmoides. In addition, the expression of this gene was found to be tissue-specific, with the highest expression observed in the leaves of male plants. Furthermore, the alterations in the expression of the EuSIP5 gene across different months indicated that it might be influenced by environmental factors. The rapid growth and development of E. ulmoides in July-September was accompanied by an increase in plant respiration and photosynthesis, accompanied by a significant rise in the expression of the EuSIP5 gene in the nutrient organs. In August, the higher temperatures resulted in a reduction in the respiration and transpiration of plants, accompanied by a decline in the expression of the EuSIP5 gene.
Genetic transformation of tobacco was employed to overexpress the EuSIP5 gene in tobacco. A comparison of the phenotypes of plants observed under drought stress revealed that the OE plants exhibited significantly enhanced growth and development compared to the WT plants. Furthermore, the transgenic tobacco exhibited markedly improved drought tolerance, as evidenced by a reduction in the rate of wilting. Further analysis of the changes in the expression of tobacco drought-related genes, including DET2, CYP85A1, P5CS, ERF1 and F-box, indicated that the EuSIP5 gene may enhance the plant's response to drought by affecting the expression of drought-induced response genes within tobacco, thereby increasing antioxidant enzyme activity and decreasing MDA content.
When the normal growth and metabolism of plants are inhibited by drought, the content of membrane lipid peroxidation product MDA in plant tissues increases dramatically [40]. This results in the inhibition of plant photosynthesis, which in turn activates the protective enzyme system, which consists of protective enzymes such as SOD. The POD and CAT enzymes are capable of playing a role in protecting the cell membranes. Among these, SOD is considered to be the core of the protective system, and is positively correlated with plant antioxidant stress capacity [41,42]. SOD can mitigate membrane lipid peroxidation damage by dismutation reactions using superoxide anion (O2-·) as a substrate [43], POD scavenges harmful free radicals and degrades toxic substances within cells [44]. CAT effectively breaks down excess H₂O₂ in the tissues [45]. The coordinated functioning of various antioxidant enzymes maintains low intracellular levels of reactive oxygen species (ROS), thereby allowing plant cells to be protected from or mitigated against damage from oxidative stress [46]. The determination of protective enzyme activities demonstrated that the activities of the POD, SOD and CAT enzymes were higher in OE plants than in the WT, while the MDA content was lower than in the WT. The results indicate that the overexpression of EuSIP5 enhances the resistance to lipid oxidation and reduces oxidative stress damage in tobacco, thereby increasing the drought tolerance of the plant.
In conclusion, this study provides new insights into the potential of genetic engineering technology to enhance drought tolerance in plants and identifies promising genetic resources for the development of drought-tolerant crop varieties in the future. This could contribute to the sustainability of agricultural production and ecosystems. However, further studies are needed to elucidate the specific role of the EuSIP5 gene in the mechanism of drought tolerance in plants.
5. Conclusions
In this study, we cloned and found that EuSIP5 may be an unstable amphipathic protein, which exists in the cytoplasm. The spatio-temporal expression and tissue expression specificity of the EuSIP5 gene in different tissues and developmental stages of E. ulmoides were revealed, suggesting that the gene may be closely related to the nutritional growth and metabolic activities of the plant. Furthermore, the transfer of the EuSIP5 gene into tobacco was found to enhance the drought resistance of tobacco plants.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: List of the primers used for the experiments; Table S2: The list shows soluble inorganic pyrophosphatase sequences retrieved from NCBI from different plant species; Figure S1: Target gene band test; Figure S2: Constructing carrier maps; Figure S3: Tobacco genetic transformation process and GUS staining and PCR identification.
Author Contributions
D-G.Z. and C.L. conceived and designed the experiments; L-Y.L. and C.X. performed the experiments and ana-lyzed the data; L-Y.L. wrote the Manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (No. 31870285), Guizhou Academy of Agricultural Sciences Talent Special Project (No. 2023-02), and by Talent Base for Germplasm Resources Utilization and Innovation of Characteristic Plant in Guizhou (RCJD2018-14).
Data Availability Statement
The data that support the fundings of this study are available from the corresponding author on reasonable request.
Acknowledgments
We thank all the colleagues that helped with the development of different parts of this manuscript and Key Laboratory of Mountain Plant Resources Protection and Germplasm Innovation of the Ministry of Education and Guizhou Plant Conservation Technology Center, Guizhou Key Laboratory for providing equipment and technical support for this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science (New York, N.Y.) 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Critical Reviews in Biotechnology 2021, 41, 669–691. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.X.; Shi, J.W.; Li, R.; An, Y.C.; Yang, B.L. Effects of extreme drought on plant species in Karst area of Guizhou Province, Southwest China. Chinese Journal of Applied Ecology 2011, 22, 1127–1134. [Google Scholar] [CrossRef]
- Ren, N.; Gong, W.W.; Zhao, Y.C.; Zhao, D.G.; Xu, Y.W. Innovation in sweet rice wine with high antioxidant activity: Eucommia ulmoides leaf sweet rice wine. 2023, 9. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Huang, X.; Zhao, Y.; Zhao, D. Cloning and Function Identification of a Phytoene Desaturase Gene from Eucommia ulmoides. 2023, 92, 1377–1389. [Google Scholar] [CrossRef]
- Sasaki, Y.; Chiba, A.; Murakami, M.; Sekihashi, K.; Tanaka, M.; Takahoko, M.; Moribayashi, S.; Kudou, C.; Hara, Y.; Nakazawa, Y.; et al. Antimutagenicity of Tochu tea (an aqueous extract of Eucommia ulmoides leaves): 2. Suppressing effect of Tochu tea on the urine mutagenicity after ingestion of raw fish and cooked beef. Mutation Research/Genetic Toxicology 1996, 371, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Choi, M.S.; Kim, M.J.; Jung, U.J.; Kim, H.J.; Park, K.K.; Noh, H.J.; Park, H.M.; Park, Y.B.; Lee, J.S.; et al. Hypoglycemic and hypolipidemic action of Du-zhong (Eucommia ulmoides Oliver) leaves water extract in C57BL/KsJ-db/db mice. Journal of Ethnopharmacology 2006, 107, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Hsieh, C.L. Reactive Oxygen Species Scavenging Activity of Du-zhong (Eucommia ulmoides Oliv.) and Its Active Compounds. Journal of Agricultural and Food Chemistry 2000, 48, 3431–3436. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Li, B.; Zhao, Y.C.; Zhao, D.G. Cloning and characterization of the DIR1 promoter from Eucommia ulmoides Oliv and its response to hormonal and abiotic stress. Plant Cell, Tissue and Organ Culture (PCTOC) 2021, 146, 313–322. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Liu, Y.Q.; Dong, X.; Liu, J.J.; Zhao, D.G. Identification of a novel laccase gene EuLAC1 and its potential resistance against Botrytis cinerea. Transgenic Research 2022, 31, 215–225. [Google Scholar] [CrossRef]
- Dong, X.; Zhao, Y.C.; Ran, X.; Guo, L.X.; Zhao, D.G. Overexpression of a New Chitinase Gene EuCHIT2 Enhances Resistance to Erysiphe cichoracearum DC. in Tobacco Plants. International Journal of Molecular Sciences 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhao, D.G. Cloning, Characterization, and Functional Analysis of EuTIL1, a Gene-Encoding Temperature-Induced Lipocalin in Eucommia ulmoides Oliv. Horticulturae 2023, 9. [Google Scholar] [CrossRef]
- Lu, Y.X.; Dong, X.; Huang, X.Z.; Zhao, D.G.; Zhao, Y.C.; Peng, L. Combined analysis of the transcriptome and proteome of Eucommia ulmoides Oliv. (Duzhong) in response to Fusarium oxysporum. 2022, 10. [Google Scholar] [CrossRef]
- Wang, L.; Wang, R.; Li, Y.; Zhao, Y.; Zhao, D. Overexpression of β-1,4-Glucanase Gene EuEG1 Improves Micrografting of Eucommia ulmoides. 2023, 92, 3063–3075. [Google Scholar] [CrossRef]
- Gloria, S.B.; Agustín, H.; Guillermo, L.L.; José Román, P.C.; Plácido, N.; Aurelio, S. Inorganic Pyrophosphatase Defects Lead to Cell Cycle Arrest and Autophagic Cell Death through NAD+ Depletion in Fermenting Yeast*. Journal of Biological Chemistry 2013, 288, 13082–13092. [Google Scholar] [CrossRef]
- Serrano-Bueno, G.; Hernández, A.; López-Lluch, G.; Pérez-Castiñeira, J.R.; Navas, P.; Serrano, A. Inorganic pyrophosphatase defects lead to cell cycle arrest and autophagic cell death through NAD+ depletion in fermenting yeast. Journal of Biological Chemistry 2013, 288, 13082–13092. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Zeng, R.Z.; Xiao, X.Z. Advances on the Research of Pyrophosphatase in Plants. Life Science Research 2004, 83–87. [Google Scholar] [CrossRef]
- Xian, J.H.; Zhang, M.P.; Sun, C.Y.; Wang, Y.F.; Wang, K.Y.; Chen, J.; Zhao, M.Z.; Wang, Y. Research Progress of Soluble Pyrophosphatase. Jilin Agriculture University, 2019.
- Huang, H.; Patskovsky, Y.; Toro, R.; Farelli, J.D.; Pandya, C.; Almo, S.C.; Allen, K.N.; Dunaway-Mariano, D. Divergence of structure and function in the haloacid dehalogenase enzyme superfamily: Bacteroides thetaiotaomicron BT2127 is an inorganic pyrophosphatase. Biochemistry 2011, 50, 8937–8949. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, Y.R.; Wang, X.L.; Fang, T.; Zhen, S.H.; Lu, J.W.; Zhang, J.; Fu, J.J. Cloning and Expression Analysis of Soluble Inorganic Pyrophosphatase Family Genes in Maize (Zea mays L.). Journal of Plant Genetic Resources 2021, 22, 455–465. [Google Scholar] [CrossRef]
- Zhu, J.H.; Xu, J.; Yu, X.H.; Chang, W.J.; Zhang, Z.L. Prokaryotic Expression for Three Soluble Inorganic Pyrophosphatase Genes from Hevea brasiliensis. Chinese Journal of Tropical Crops 2013, 34, 41–45. [Google Scholar]
- George, G.M.; van der Merwe, M.J.; Nunes-Nesi, A.; Bauer, R.; Fernie, A.R.; Kossmann, J.; Lloyd, J.R. Virus-induced gene silencing of plastidial soluble inorganic pyrophosphatase impairs essential leaf anabolic pathways and reduces drought stress tolerance in Nicotiana benthamiana. Plant physiology 2010, 154, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Song, Q.; Wei, H.; Wang, Y.; Lin, M.; Sun, K.; Zhang, Y.; Yang, J.; Li, C.; Luo, K.J.N.P. The AP2/ERF transcription factor PtoERF15 confers drought tolerance via JA-mediated signaling in Populus. 2023, 240, 1848–1867. [Google Scholar] [CrossRef]
- Wang, H.; Ni, D.; Shen, J.; Deng, S.; Xuan, H.; Wang, C.; Xu, J.; Zhou, L.; Guo, N.; Zhao, J.J.F.i.P.S. Genome-wide identification of the AP2/ERF gene family and functional analysis of GmAP2/ERF144 for drought tolerance in soybean. 2022, 13, 848766. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yu, M.; Zhang, S.; Song, T.; Zhang, M.; Zhou, H.; Wang, Y.; Xiang, J.; Zhang, X. Transcriptomic Identification of Wheat AP2/ERF Transcription Factors and Functional Characterization of TaERF-6-3A in Response to Drought and Salinity Stresses. International Journal of Molecular Sciences 2022, 23. [Google Scholar] [CrossRef]
- Ma, Z.; Hu, L.; Jiang, W. Understanding AP2/ERF Transcription Factor Responses and Tolerance to Various Abiotic Stresses in Plants: A Comprehensive Review. International Journal of Molecular Sciences 2024, 25. [Google Scholar] [CrossRef]
- Abe, H.; Yamaguchi-Shinozaki, K.; Urao, T.; Iwasaki, T.; Hosokawa, D.; Shinozaki, K. Role of arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. The Plant Cell 1997, 9, 1859–1868. [Google Scholar]
- Van Moerkercke, A.; Duncan, O.; Zander, M.; Šimura, J.; Broda, M.; Vanden Bossche, R.; Lewsey, M.G.; Lama, S.; Singh, K.B.; Ljung, K.J.P.o.t.N.A.o.S. A MYC2/MYC3/MYC4-dependent transcription factor network regulates water spray-responsive gene expression and jasmonate levels. 2019, 116, 23345–23356. [Google Scholar] [CrossRef]
- Yang, D.; Ni, R.; Yang, S.; Pu, Y.; Qian, M.; Yang, Y.; Yang, Y. Functional Characterization of the Stipa purpurea P5CS Gene under Drought Stress Conditions. International Journal of Molecular Sciences 2021, 22. [Google Scholar] [CrossRef]
- Duan, F.M.; Ding, J.; Lee, D.S.; Lu, X.L.; Feng, Y.Q.; Song, W.W. Overexpression of SoCYP85A1, a spinach cytochrome p450 gene in transgenic tobacco enhances root development and drought stress tolerance. Frontiers in plant science 2017, 8, 1909. [Google Scholar] [CrossRef]
- Zheng, B.F.; Ma, Y.Z.; Lu, H.D.; An, W.K.; Zhang, F.C. Effects of Spraying Brassinolide on Expression of Steroid 5-Alpha Reductase Gene(Gh DET2) in Cotton under Drought Stress. Genomics and Applied Biology 2018, 37, 859–866. [Google Scholar] [CrossRef]
- An, J.; Li, Q.X.; Yang, J.J.; Zhang, G.Q.; Zhao, Z.X.; Wu, Y.Z.; Wang, Y.; Wang, W. Wheat F-box protein TaFBA1 positively regulates plant drought tolerance but negatively regulates stomatal closure. Frontiers in Plant Science 2019, 10, 1242. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.H.; Zhao, Y.; Zhao, Y.; Feng, C.; Zhang, Y.H.; Wang, F.W.; Li, X.W.; Gao, H.T.; Liu, W.C.; Jing, Y. Soybean F-box-like protein GmFBL144 interacts with small heat shock protein and negatively regulates plant drought stress tolerance. Frontiers in plant science 2022, 13, 823529. [Google Scholar] [CrossRef] [PubMed]
- Navarro-De la Sancha, E.; Coello-Coutiño, M.P.; Valencia-Turcotte, L.G.; Hernández-Domínguez, E.E.; Trejo-Yepes, G.; Rodríguez-Sotres, R. Characterization of two soluble inorganic pyrophosphatases from Arabidopsis thaliana. Plant Science 2007, 172, 796–807. [Google Scholar] [CrossRef]
- Schulze, S.; Mant, A.; Kossmann, J.; Lloyd, J.R. Identification of an Arabidopsis inorganic pyrophosphatase capable of being imported into chloroplasts. FEBS letters 2004, 565, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Luna, F.M.; Navarro de la Sancha, E.; Valencia-Turcotte, L.G.; Vázquez-Santana, S.; Rodríguez-Sotres, R. Evidence for a non-overlapping subcellular localization of the family I isoforms of soluble inorganic pyrophosphatase in Arabidopsis thaliana. Plant Science 2016, 253, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Qiao, X.; Zhu, X.X.; Khan, W.; Wu, J.; Zhang, S.L. Expression and evolutionary analysis of soluble inorganic pyrophosphatase gene family in pear and four other Rosaceae species. Plant Systematics and Evolution 2020, 306, 46. [Google Scholar] [CrossRef]
- Wang, J.; Kang, L.Y.; Liu, Z.B.; Lv, J.H.; Liu, Y.H.; Zou, X.X. Research Progress on the Impact of Drought on Plant. Hunan Agricultural Sciences, 2017; 123-126+130. [Google Scholar] [CrossRef]
- Li, B.; Zeng, Q.; Zhao, D.; Zhao, D.G. Cloning and Function Analysis of EuERD16 Gene in Eucommia ulmoides. Genomics and Applied Biology 2023, 42, 373–383. [Google Scholar] [CrossRef]
- Chen, S.Y. Membrane lipid peroxidation and plant adversity stresses. Bulletin of Botany 1989, 212–215. [Google Scholar]
- Li, M.Q. Physiological and Molecular Regulatory Mechanisms of Soybean in Response to Drought. Shenyang Agricultural University, 2023.
- Wang, Q.M. Effects of Drought Stress on Protective Enzymes Activities and Membrane Lipid Peroxidation in Leaves of Soybean Seedlings. Journal of Agro-Environment Science 2006, 918–921. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Zheng, A.Z.; Liu, C.P.; Shen, Z.G. Effect of Cadmium on MDA Content, POD and SOD Activities of Brassica Pekinensis and Brassica Chinensis. Hubei Agricultural Sciences 2005, 67–69. [Google Scholar]
- Song, X.G.; She, X.P. The Generation and the Role of Hydrogen Peroxide in Plant. Journal of Lianyungang Teachers College 2010, 27, 99–103. [Google Scholar] [CrossRef]
- Türkan, İ.; Bor, M.; Özdemir, F.; Koca, H. Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Science 2005, 168, 223–231. [Google Scholar] [CrossRef]
Figure 1.
Prediction of conserved structural domains of EuSIP5, multiple sequence alignment of EuSIP5 with other species and construction of the EuSIP5 phylogenetic tree. (a) Conserved domain analysis of EuSIP5 shows that it is a soluble inorganic pyrophosphatase of the pyrophosphatase superfamily; (b) EuSIP5 has 88.10% similarity to homologous proteins of other species; (c) The results showed that the soluble inorganic pyrophosphatase EuSIP5 in E. ulmoides was closest in evolutionary relationship to the soluble inorganic pyrophosphatases in Camellia lanceoleosa and Cornus florida.
Figure 1.
Prediction of conserved structural domains of EuSIP5, multiple sequence alignment of EuSIP5 with other species and construction of the EuSIP5 phylogenetic tree. (a) Conserved domain analysis of EuSIP5 shows that it is a soluble inorganic pyrophosphatase of the pyrophosphatase superfamily; (b) EuSIP5 has 88.10% similarity to homologous proteins of other species; (c) The results showed that the soluble inorganic pyrophosphatase EuSIP5 in E. ulmoides was closest in evolutionary relationship to the soluble inorganic pyrophosphatases in Camellia lanceoleosa and Cornus florida.
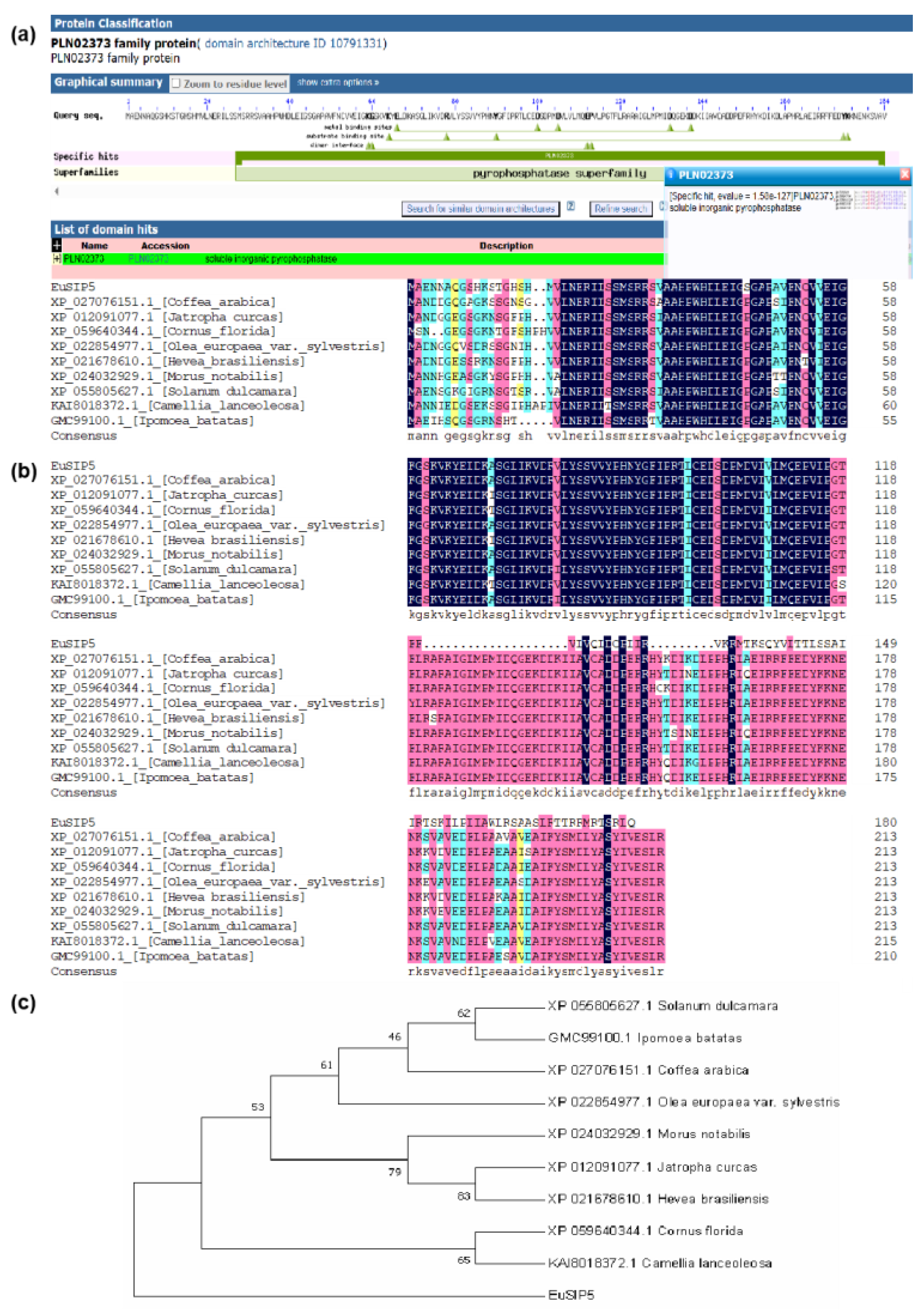
Figure 2.
Relative expression of the EuSIP5 genes in different tissues of E. ulmoides. (a) Expression patterns of EuSIP5 gene in different tissues of E. ulmoides; (b) Expression of EuSIP5 gene under drought treatment; (c) Comparison of the relative expression of the EuSIP5 gene in different tissues of female E. ulmoides in different months; (d) Comparison of the relative expression of the EuSIP5 gene in different tissues of male E. ulmoides in different months. The Error bars indicate standard error (SE); *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
Figure 2.
Relative expression of the EuSIP5 genes in different tissues of E. ulmoides. (a) Expression patterns of EuSIP5 gene in different tissues of E. ulmoides; (b) Expression of EuSIP5 gene under drought treatment; (c) Comparison of the relative expression of the EuSIP5 gene in different tissues of female E. ulmoides in different months; (d) Comparison of the relative expression of the EuSIP5 gene in different tissues of male E. ulmoides in different months. The Error bars indicate standard error (SE); *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
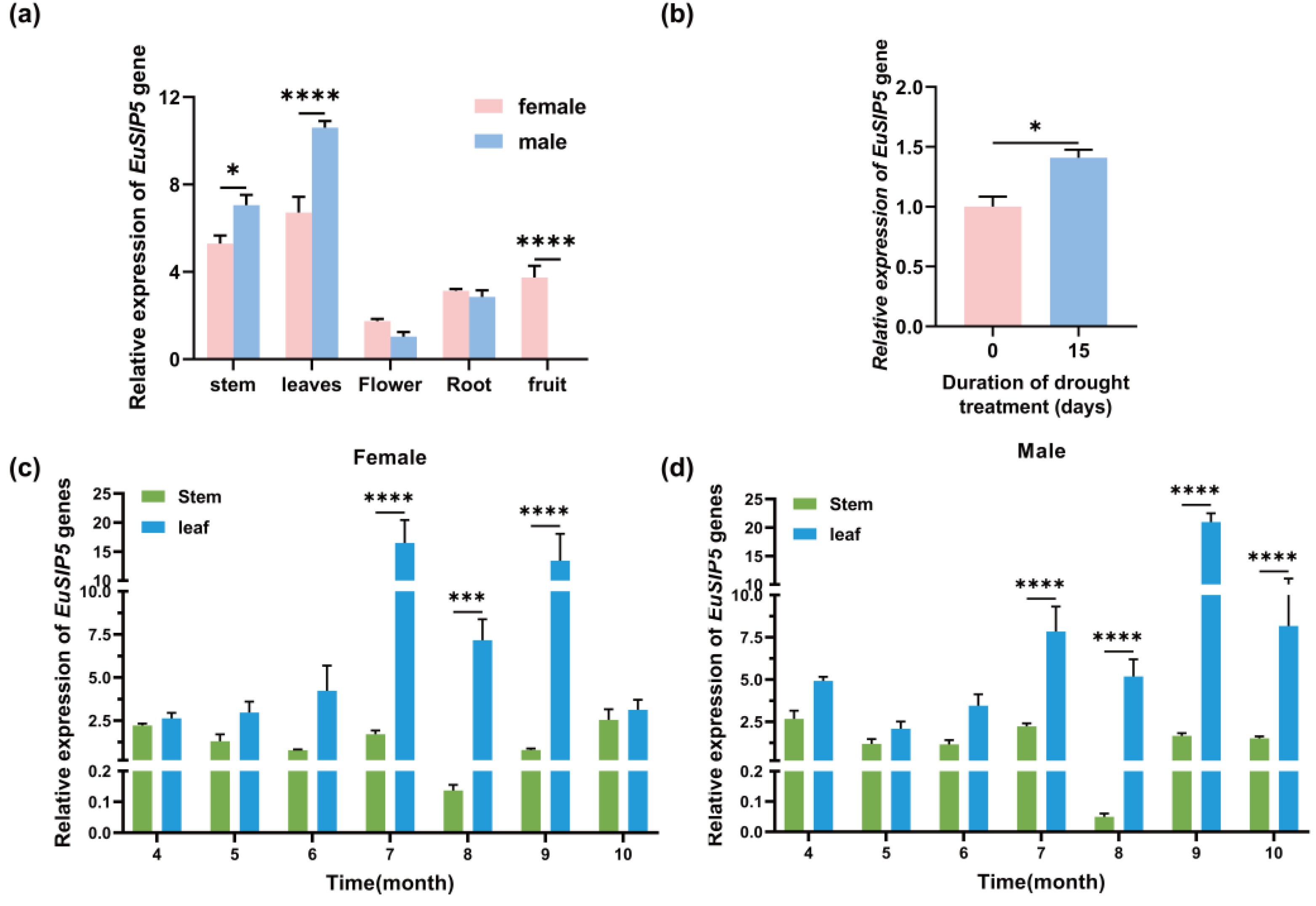
Figure 3.
Subcellular localisation of EuSIP5 in tobacco cells. (A) Localisation map of pEGOEP-35S-EuSIP5-eGFP in tobacco cells; (B) Localisation map of pEGOEP-35S-eGFP in tobacco cells; (a) Green fluorescence from labelled proteins; (b) chloroplast self-illumination; (c) bright field; (d) superposition field. Scale bar: 50 μm.
Figure 3.
Subcellular localisation of EuSIP5 in tobacco cells. (A) Localisation map of pEGOEP-35S-EuSIP5-eGFP in tobacco cells; (B) Localisation map of pEGOEP-35S-eGFP in tobacco cells; (a) Green fluorescence from labelled proteins; (b) chloroplast self-illumination; (c) bright field; (d) superposition field. Scale bar: 50 μm.
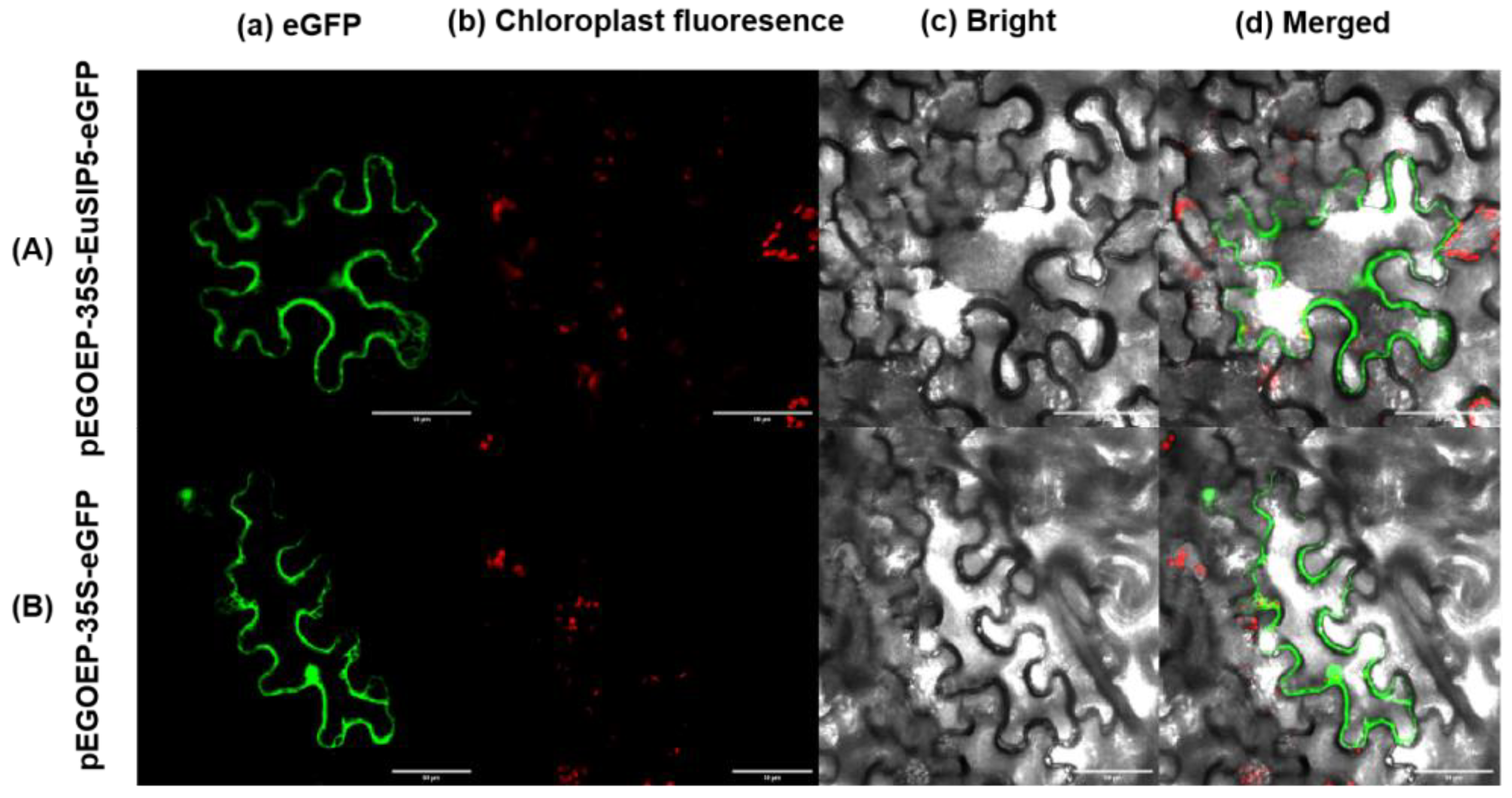
Figure 4.
Expression level analysis of genes related to tobacco drought resistance. (a) Relative expression of NtDET2 gene in tobacco; (b) Relative expression of NtCYP85A1 gene in tobacco; (c) Relative expression of NtF-box gene in tobacco. (d) Relative expression of NtP5CS gene in tobacco; (e) Relative expression of NtERF1 gene in tobacco. WT: wild-type plants; OE: transgenic plant; Error bars indicate standard error (SE); *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
Figure 4.
Expression level analysis of genes related to tobacco drought resistance. (a) Relative expression of NtDET2 gene in tobacco; (b) Relative expression of NtCYP85A1 gene in tobacco; (c) Relative expression of NtF-box gene in tobacco. (d) Relative expression of NtP5CS gene in tobacco; (e) Relative expression of NtERF1 gene in tobacco. WT: wild-type plants; OE: transgenic plant; Error bars indicate standard error (SE); *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
Figure 5.
Phenotype comparison of tobacco drought treatment (0-8 d). (a) Drought treatment for 0 days, WT and OE tobacco were in good condition; (b) Wilt in wild-type tobacco at 4 d of drought treatment; (c) Drought treatment for 8 d, Transgenic tobacco began to wilt. WT: wild-type plants; OE: transgenic plant; scale bar: 2 cm.
Figure 5.
Phenotype comparison of tobacco drought treatment (0-8 d). (a) Drought treatment for 0 days, WT and OE tobacco were in good condition; (b) Wilt in wild-type tobacco at 4 d of drought treatment; (c) Drought treatment for 8 d, Transgenic tobacco began to wilt. WT: wild-type plants; OE: transgenic plant; scale bar: 2 cm.

Figure 6.
Analysis of tobacco protective enzyme viability. (a) POD enzyme activity; (b) SOD enzyme activity; (c) CAT enzyme activity; (d) MDA content. Error bars indicate standard error (SE).
Figure 6.
Analysis of tobacco protective enzyme viability. (a) POD enzyme activity; (b) SOD enzyme activity; (c) CAT enzyme activity; (d) MDA content. Error bars indicate standard error (SE).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



