
Submitted:
14 August 2024
Posted:
15 August 2024
You are already at the latest version
Abstract
Butirosins are naturally occurring aminoglycoside (AG) antibiotics featuring a 4,5-disubstituted 2-deoxystreptamine (2-DOS) with a (2S)-4-amino-2-hydroxybutyrate (AHBA) side chain. This side chain has been shown to confer resistance against AG modifying enzymes, leading to ongoing studies on the butirosin biosynthetic pathway and the corresponding enzymes. Butirosin is produced by Niallia (formerly Bacillus) circulans and Bacillus vitellinus, with most research focused on the former strain. To date, no whole genome analysis has been performed on B. vitellinus. In this study, we sequenced the complete genome of B. vitellinus NBRC 13296 and performed a comparative analysis of different butirosin biosyntheric gene clusters (BGCs), including those from N. circulans. The complete genome of B. vitellinus NBRC 13296 comprises a 6,331,192 base circlular chromosome with a GC content of 52.68%. The annotation revealed the presence of 5,605 CDSs, 70 tRNA genes, 30 rRNA genes, and 3 ncRNA genes in NBRC 13296. The highest dDDH and ANI values between NBRC 13296 and the most closely related type strain, Paenibacillus chitinolyticus KCCM 41400, were 97.8% and 98.66%, respectively. Based on these genome-based comparative analyses, we propose reclassifying B. vitellinus NBRC 13296 as P. chitinolyticus. Genome mining revealed 18 gene clusters encoding the biosynthesis of diverse secondary metabolites in the genome of B. vitellinus NBRC 13296, indicating the enormous biosynthetic potential of this strain. The predicted structural diversity of secondary metabolites includes aminoglycoside, PKS, NRPS, PKS-NRPS hybrids, metallophores, phosphonates, terpenes, β-lactones, and RiPP peptides. We then comparatively characterized butorosin BGCs previously studied in several N. circulans strains. Additionally, comparative genome analysis revealed complete butirosin BGCs identified from P. chitinolyticus KCCM 41400, P. chitinolyticus NRRL B-23119, P. chitinolyticus NRRL B-23120, P. chitinolyticus B-14908, P. chitinolyticus YSY-3.1, P. chitinolyticus JMW06, Paenibacillus sp. GbtcB18, Paenibacillus sp. HGH0039, Paenibacillus sp. MZ04-78.2. Finally, we identified the core region consisting of BtrS, BtrN, BtrM, BtrL, BtrA, BtrB, BtrC, BtrD, BtrD, BtrE, BtrF, BtrG, BtrH, BtrI, BtrI, BtrJ, BtrK, BtrO, BtrP, and BtrV, followed by an upstream region organizing BtrQ, BtrW, BtrX, BtrY, and BtrZ in the same transcriptional direction and sequential genetic arrangement, and a downstream region organizing various proteins based on BtrT, BtrR2, BtrU, and BtrR1. Our study provided insights into the reclassification of B. vitellinus NBRC 13296 to P. chitinolyticus and suggested the need for continued studies on butirosin biosynthesis from an enzymatic perspective.
Keywords:
Bacillus vitellinus NBRC 13296
; butirosin BGC
; Paenibacillus chitinolyticus
; reclassification
1. Introduction
Aminoglycosides are an important class of antibiotics used to treat serious infections caused by several Gram-negative and Gram-positive bacteria, despite their nephrotoxic and ototoxic side effects. The mechanism of action of aminoglycosides involves binding to the 30S ribosomal subunit, which interferes with protein synthesis by causing errors in translation, specifically in codon reading, thereby disrupting protein synthesis [1,2]. Aminoglycosides consist of highly functionalized aminosugars and deoxysugars linked by glycosidic bonds to an aminocyclitol aglycone, with most aglycones containing a 2-deoxystreptamine (2-DOS) moiety, to which other amino sugars are attached at 4 and 5 or 4 and 6 positions of the 2-DOS carbons. Aminoglycoside producers primarily belong to the actinomycetes group of bacteria, but butirosin is one of the few aminoglycosides produced by a non-actinomycete bacterium [3,4].
The (S)-4-amino-2-hydroxybutyrate (AHBA) group attached to the C-1 amine of the 2-DOS moiety in butirosin has long been of interest because this AHBA moiety can prevent the action of several aminoglycoside-modifying enzymes (AMEs), such as aminoglycoside N-acetyltransferases (AACs), which can theoretically or practically cause aminoglycoside resistance. For example, amikacin is a semisynthetic aminoglycoside with an AHBA group attached to the C-1 of the 2-DOS aglycone of kanamycin, and is considered one of the few antibiotics that can overcome various AME mechanisms [5-7]. Butirosin was identified as a pair of epimers, butirosins A and B, from Niallia (formerly Bacillus) circulans in 1971 and was subsequently discovered in Bacillus vitellinus [8,9].
To date, the whole genome sequence (WGS) of N. circulans in the NCBI (National Center for Biotechnology Information) databases has been completed for the reference strain N. circulans NBRC 13626 (SAMD00046991), with additional WGS reported for N. circulans DC10 (SAMN18824981), isolated from textile dye-contaminated soil sediments in India, and N. circulans GN 3 (SAMN14847575), isolated from purple rhizosphere soil growing the cabbage Brassica campestris in China. However, there are no reports of complete genome information for B. vitellinus, one of the main producer strains of butirosin. Here, we report the complete genome sequence of B. vitellinus NBRC 13296 to better understand the biosynthesis of butirosin and other bioactive secondary metabolites. The genome was sequenced using long-read PacBio technologies to obtain high-quality genomic sequences [10,11].
Meanwhile, bioinformatics software such as antiSMASH, a platform for automated genome mining of secondary metabolite producers, can be used to rapidly identify, analyze, and annotate BGCs from genome-wide information and identify similar characterized gene clusters in the MIBiG repository [12,13]. Finally, comparative genomic analysis using next-generation sequencing (NGS) techniques and bioinformatics software confirmed the presence of a butirosin BGC.
Interestingly, during the BLASTP search using antiSMASH, butirosin BGCs were identified as biosynthetic enzymes in the type strain Paenibacillus chitinolyticus KCCM 41400 (SAMN08222605), as well as in P. chitinolyticus NRRL B-23119 (SAMN27675096), P. chitinolyticus NRRL B-23120 (SAMN27675097), P. chitinolyticus YSY-3.1 (SAMD00444452), P. chitinolyticus JMW06 (SAMN19998407), P. chitinolyticus B-14908 (SAMN33770086), Paenibacillus sp. GbtcB18 (SAMN18679176), Paenibacillus sp. MZ04-78.2 (SAMN28689874), Paenibacillus sp. HGH0039 (SAMN02596731).), N. circulans ATCC 21557 (AJ781030), N. circulans ATCC 21558 (LC571042.1), and N. circulans SANK 72073 (AB097196). The full genome-derived 16S rRNA gene sequences, average nucleotide identity (ANI), and digital DNA-DNA hybridization (dDDH) suggest that all the strains mentioned above should be classified as members of the same species. We propose the reclassification of B. vitellinus NBRC 13296 as P. chitinolyticus NBRC 13296 and an emended description of P. chitinolyticus. Finally, in this article, we aim to provide scientists with a comparative analysis of butirosin BGCs to assist in the study of the butirosin biosynthetic pathway and corresponding enzymes.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
Bacillus vitellinus NBRC 13296 was obtained from the NITE Biological Research Center (NBRC, Tokyo, Japan). The cells were grown in a tryptic soy broth (TSB) medium containing 1.7% tryptone (pancreatic digest of casein), 0.3% soytone (peptic digest of soybean), 0.25% glucose, 0.5% sodium chloride, and 0.25% dipotassium phosphate (BD Biosciences, Franklin Lakes, NJ, USA) at 30°C for 24 h.
2.2. Genome De Novo Sequencing, Assembly, and Annotation
The genomic DNA of B. vitellinus was extracted and purified using the Nanobind DNA extraction kit, which produces HiFi reads on the PacBio systems (Pacific Biosciences, Menlo Park, CA, USA). The gDNA obtained from the extraction was first sheared using the g-TUBE (Covaris) following the manufacturer’s protocol. SMRTbell libraries were created using the PacBio SMRTbell Express template preparation kit (v1.0). SMRTbell templates were annealed using the PacBio DNA/Polymerase Binding Kit P6. The PacBio DNA Sequencing Kit 4.0 and 8 SMRT cells were used for sequencing. SMRT cells (Pacific Biosciences, Menlo Park, CA, USA) using C4 chemistry and 240 min movies were captured for each SMRT cell using the PacBio Sequel II (Pacific Biosciences) sequencing platform. These sequences were assembled de novo using the CANU ver. 2.2 workflow. The assembly was circularized by Circulator (v. 1.5.5) and rearranged to start at the dnaA gene. [14]. Regarding gene prediction and annotation, protein-coding, tRNA, rRNA genes, and repeat regions were predicted using the NCBI PGAP (Prokaryotic Genome Annotation Process). Gene annotation was performed using RefSeq and the HMM library (TIGRFAM, Pfam, PRK HMMs) database [15, 16].
2.3. Phylogenetic Analysis
Phylogenetic trees were constructed based on the complete 16S rRNA gene sequence derived from the whole genome information of B. vitellinus NBRC 13296. The evolutionary tree based on 16S rRNA sequences was constructed with the MEGA 11 package [17], applying neighbor-joining and maximum-likelihood algorithms after multiple alignments of the sequence data by the ClustalW program. This process primarily used BlastN homology searches and referenced BacDive, the worldwide largest database for standardized bacterial information, for type strains. The confidence of the tree topologies was assessed by 1000 bootstrap replicates.
2.4. Comparative Genomic Studies and Whole Genome Relatedness
Digital DNA-DNA hybridization (dDDH) values of the B. vitellinus NBRC 13296 genome and its neighbors were calculated using the Genome-to-Genome Distance Calculator (GGDC 4.0) within the Type (Strain) Genome Server (TYGS) of the Leibniz Institute DSMZ [18]. The average nucleotide identity (ANI) values between B. vitellinus NBRC 13296 and its nearest neighbors were computed using the ANI calculator of Ezbiocloud, an online tool used to compare two prokaryotic genome sequences [19].
2.5. Secondary Metabolite and Butirosin BGCs Analysis
To discover BGCs involved in the production of secondary metabolites, including butirosin, various computational programs such as Known ClusterBlast, ClusterBlast, SubClusterBlast, ActiveSiteFinder, and Cluster PFam analysis were employed. The PRISM 4 and BAGEL 4 tools were implemented with default settings. Anti-SMASH 7.0 facilitated the identification, annotation, and analysis of secondary metabolite BGCs across the genome [12]. RiPPs and bacteriocins were mined by BAGEL 4 [20], while PRISM 4 version 4.4.5 was used for the comprehensive analysis of secondary metabolite structures and biological activities [21]. These sophisticated computational tools provide accurate predictions of microbial secondary metabolite encoding potential and putative structures.
3. Results and Discussion
3.1. Sequencing, Assembly, Phylogenetic Analysis, and Genomic Characteristics
In whole-genome sequencing based on the PacBio Sequel IIe platform, the complete genome sequence of B. vitellinus NBRC 13296 was composed of 1 contig with a total length of 6,331,192 bps and an average G + C content of 50.98%. A total of 5,731 genes were identified in its genome, including 5,605 annotated protein-coding genes, 92 tRNA, 30 rRNA, 3 ncRNA, and 1 tmRNA genes. Ten copies of 16SrRNAs were identified in the genome of B. vitellinus NBRC 13296, and variations at seven positions were also identified by alignment between the 16S rRNA sequences, totaling 1,544 bp, using ClustalW. No plasmid was detected. In the 16S rRNA gene sequence analysis, B. vitellinus NBRC 13296 showed sequentially 99.66%, 95.71%, 95.60%, and 94.96% similarities to the type strains Paenibacillus chitinolyticus HSCC 596 (NR_040854.1), Paenibacillus gansuensis B518 (NR_043219.1), Paenibacillus lutrae N10 (NR_173496.1), and Paenibacillus favisporus GMP01 (NR_029071.1), respectively (Table S1). The phylogenetic tree constructed from the BLASTN, BacDive, and EzBioCloud 16S database [22] using maximum-likelihood and neighbor-joining methods in the MEGA 11 application with 1000 bootstrap values showed B. vitellinus NBRC 13296 forming a clade with P. chitinolyticus as shown in Figure 1 and Figure S1.
Comparative genomic analysis revealed that the Digital DNA-DNA hybridization (dDDH) values between B. vitellinus NBRC 13296 and the closely related strain P. chitinolyticus KCCM 41400 were far above the 70% threshold value for species evaluation [23]. The average nucleotide identity (ANI) values were also within the threshold range (94–96%) for species identification [24]. The dDDH and ANI values based on genome sequences between B. vitellinus NBRC 13296 and P. chitinolyticus KCCM 41400 were 97.8% and 98.66%, respectively, providing strong evidence that these strains may belong to the same taxonomic species.
3.2. Prediction of Secondary Metabolite Biosynthetic Gene Clusters (BGCs)
Using the standard cluster rule-based approach in antiSMASH analysis, a variety of natural product classes were identified in B. vitellinus NBRC 13296. Approximately 12.4% of the B. vitellinus NBRC 13296 genome is dedicated to secondary metabolism. The secondary metabolite gene cluster are concentrated in the center of the chromosome, spanning regions 5 to 16 (Figure S2). In total, 18 putative biosynthetic gene clusters were identified (Table 1). Interestingly, region 15 contained two-copies of an opine-like-metallophore gene cluster, consisting of opine metallophore biosynthesis dehydrogenase, nicotianamine synthase family protein, and nickel/cobalt ABC transporter substrate-binding protein/permease (Figure S3). Thus, a total of 19 BGCs were identified through antiSMASH analysis. Next, we used BAGEL to analyze the genome sequence of NBRC 13296 and found a total of 2 BGCs for different NRPS/LAP and sactipeptides. The former cluster identified by BAGEL overlaps with region 1 of antiSMASH, while the latter sactipeptides cluster is novel (Table S2 and S3). Using the PRISM algorithm, a total of 11 clusters were identified. Ten of these clusters overlapped with the AntiSMASH results, but an additional polyketide cluster, consisting of beta-ketoacyl-ACP synthase II, acyl carrier protein, ACP S-malonyltransferase, etc., was identified in cluster 10 (Figure S3 and Table S4). Based on the BGCs identified through the three approaches, we were ultimately able to identify 22 BGCs, including 19 from antiSMASH and an additional one each from BAGEL 4 and PRISM 4.4.5.
3.3. Comparative Characterization of the Butorosin BGCs
Enzymatic studies of the butirosin biosynthetic pathway have led to remarkable scientific advances in Niallia circulans, with BGCs for N. circulans SANK 72073, N. circulans ATCC 21557, and N. circulans ATCC 21558 [4, 25, 26] identified in the NCBI database. First, we obtained a large gene cluster from BtrA to BtrZ based on the butirosin BGC of N. circulans SANK 72073 and the BGC of N. circulans ATCC 21558, extended by genomic walking. We then identified additional genes in the upstream and downstream regions of the genome of B. vitellinus NBRC 13296, in addition to the tentative butirosin BGCs in region 12 analyzed by antiSMASH. As shown in Figure 2 and Table S5, B. vitellinus NBRC 13296 and the butirosin BGC from N. circulans share 67.81% to 92.87% high homology. Interestingly, during a BLASTP search for region 12 derived by antiSMASH, putative butirosine BGCs were identified in the type strain P. chitinolyticus KCCM 41400 as well as in P. chitinolyticus NRRL B-23119, P. chitinolyticus NRRL B-23120, P. chitinolyticus B-14908, P. chitinolyticus YSY-3.1, P. chitinolyticus JMW06, Paenibacillus sp. GbtcB18, Paenibacillus sp. HGH0039, Paenibacillus sp. MZ04-78.2. Next, we analyzed the genetic similarities and differences between B. vitellinus NBRC 13296, N, circulans, and the putative butirosin BGCs identified in Paenibacillus sp. The results showed that all BGCs had the following genes in common: BtrT, BtrS, BtrN, BtrM, BtrL, BtrA, BtrB, BtrC, BtrD, BtrD, BtrE, BtrF, BtrG, BtrH, BtrI, BtrI, BtrJ, BtrK, BtrO, BtrP, BtrV were located in the same transcriptional orientation, followed by BtrQ, BtrW, BtrX, BtrY, and BtrZ genes upstream of these core regions in the same transcriptional orientation and consecutive order. In contrast, the enzymes BtrT, BtrR2, BtrU, and BtrR1 are commonly found downstream of the core region, but beyond that, the genes are arranged in a strain-specific manner. Specifically, B. vitellinus NBRC 13296, P. chitinolyticus KCCM 41400, P. chitinolyticus NRRL B-23119, P. chitinolyticus NRRL B-23120, and Paenibacillus sp. GbtcB18 B18 show the same genetic organization from BtrT2 to BtrT7, while P. chitinolyticus YSY-3.1 is similar only up to BtrT2, and the remaining strains lack some of the BtrT3 to BtrT7 genes. Furthermore, Paenibacillus sp. MZ04-78.2 contained five completely unrelated ORFs instead of the BtrT, BtrR2, BtrU, and BtrR1 genes. Finally, we sought to deduce the minimal unit of the butirosin BGC. Many classes of secondary metabolite BGCs, such as aminoglycosides, type I polyketides, type II polyketides, indolocarbazoles, and enediynes, are often accompanied by regulatory and resistance genes [27-31]. Therefore, it is likely that these genes are also present in the butirosin BGC. Based on these findings, it is highly likely that the regulatory genes BtrR1 and BurR2 are included in the butirosin BGC, as are all or part of the transporter genes BtrT2, BtrW, BtrX, and BtrY, which may act as resistance genes. However, further studies, such as heterologous expression or enzymatic approaches, are required.
4. Conclusions
Over the past three decades, the 2-deoxystreptamine (2-DOS) amonoglycoside antibiotic butirosin has been intensively studied in N. circulans to develop semisynthetic antibiotics using the (2S)-4-amino-2-hydroxybutyrate (AHBA) side chain, and nearly all biosynthetic enzymes and BGCs have been characterized. The present study was initiated by the identification of the whole genome of another butirosin-producer B. vitellinus NBRC 13296, to pursue a new approach to the butirosin biosynthetic pathway. Through complete genome analysis of B. vitellinus NBRC 13296, we compared its dDDH and ANI values with P. chitinolyticus, the most closely related species based on 16SrRNA phylogenetic analysis, including N. circulans. Based on these genome-based comparative analyses, we propose to reclassify B. vitellinus NBRC 13296 to P. chitinolyticus. In this study, genome mining using AntiSMASH, BAGEL 4, and PRISM programs revealed 22 gene clusters encoding the biosynthesis of diverse secondary metabolites in the genome of B. vitellinus NBRC 13296. Finally, we comparatively characterized butirosin BGCs from N. circulans and P. chitinolyticus, revealing the minimal region of the butirosin BGC from BtrA to BtrZ and highlighting the need for continued study of butirosin biosynthesis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, K.A.H. and C.G.H.; methodology, K.A.H; bioinformatic analyses, K.A.H.; writing—original draft preparation, C.G.H.; writing—review and editing, C.G.H.; supervision, K.H.B. S.Y.K., and C.G.H.; project administration, W.J.C.; funding acquisition, W.J.C. All the authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the National Institute of Biological Resources (NIBR) funded by the Ministry of Environment (MOE) of the Republic of Korea (NIBR202402105).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The whole-genome of B. vitellinus NBRC 13296 has been deposited at the NCBI genome database under the accession number CP*****. The assembly reported in the paper is associated with NCBI BioProject: ******** and BioSample: *******. The authors confirm that all the data needed to support the study is represented within the article and supplementary files.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An Overview. Cold Spring Harb Perspect Med. 2016, 6, a027029. [Google Scholar] [CrossRef] [PubMed]
- Serio, A.W.; Keepers, T.; Andrews, L.; Krause, K.M. Aminoglycoside Revival: Review of a Historically Important Class of Antimicrobials Undergoing Rejuvenation. EcoSal Plus. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Park, S.R.; Park, J.W.; Ban, Y.H.; . Sohng, J.K.; Yoon, Y.J. 2-Deoxystreptamine-containing aminoglycoside antibiotics: recent advances in the characterization and manipulation of their biosynthetic pathways. Nat. Prod. Rep. 2013, 30, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Kudo, F.; Numakura, M.; Tamegai, H.; Yamamoto, H.; Eguchi, T.; Kakinuma, K. Extended sequence and functional analysis of the butirosin biosynthetic gene cluster in Bacillus circulans SANK 72073. J. Antibiot (Tokyo), 2005, 58, 373-379. [CrossRef]
- Li, Y.; Llewellyn, N.M.; Giri, R.; Huang, F.; Spencer, J.B. Biosynthesis of the unique amino acid side chain of butirosin: possible protective-group chemistry in an acyl carrier protein-mediated pathway. Chem. Biol. 2005, 12, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, Q.; Deng, Z. Parallel pathways in the biosynthesis of aminoglycoside antibiotics. F1000Res. 2017, 6, F1000 Faculty Rev-723. [CrossRef]
- Arenas, L.A.R.; de Paiva, F.C.F.; Rossini. N. de O.; Li, Y.; Spencer J.; Leadlay, P.; Dias. M.V.B. Crystal structure of BtrK, a decarboxylase involved in the (S)-4-amino-2-hydroxybutyrate (AHBA) formation during butirosin biosynthesis. J. Mol. Struct. 2022, 1267, 133576. [Google Scholar] [CrossRef]
- Dion, H.W.; Woo, P.W.; Willmer, N.E.; Kern, D.L.; Onaga, J.; Fusari, S.A. Butirosin, a new aminoglycosidic antibiotic complex: isolation and characterization. Antimicrob. Agents. Chemother. 1972, 2, 84–88. [Google Scholar] [CrossRef]
- Nakahama, K.; Shirafuji, H.; Nogami, I.; Kida, M.; Yoneda, M. Butirosin 3′-Phosphotransferase from Bacillus vitellinus, a Butirosin-producing Organism, Agric. Biol. Chem. 1977, 41, 2437–2445. [Google Scholar] [CrossRef]
- Espinosa, E.; Bautista, R.; Larrosa, R.; Plata, O. Advancements in long-read genome sequencing technologies and algorithms. Genomics. 2024, 116, 110842. [Google Scholar] [CrossRef]
- Logsdon, G.A.; Vollger, M.R.; Eichler, E.E. Long-read human genome sequencing and its applications. Nat. Rev. Genet. 2020, 21, 597–614. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 7.0: new and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic. Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef] [PubMed]
- Terlouw, B.R.; Blin, K.; Navarro-Muñoz, JC.; . Avalon, N.E.; Chevrette, M.G.; Egbert, S.; Lee, S.; Meijer, D.; Recchia, M.J.J.; Reitz, Z.L.; et al. MIBiG 3.0: a community-driven. effort to annotate experimentally validated biosynthetic gene clusters. Nucleic. Acids Res. 2023, 51, D603–D610. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.; Silva, N.D.; Otto, T.D.; Parkhill, J.; Keane, J.A.; Harris, S.R. Circlator: automated circularization of genome assemblies using long sequencing reads. Genome Biol. 2015, 16, 294. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic. Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Meier-Kolthoff, JP.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS.and LPSN: a database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic. Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Chalita, M.; Kim, Y.O.; Park, S.; Oh, H.S.; Cho, J.H.; Moon, J.; Baek, N.; Moon, C.; Lee, K.; Yang, J.; Nam, G.G.; Jung, Y.; Na, S.I.; Bailey, M.J.; Chun, J. EzBioCloud: a genome-driven database and platform for microbiome identification and discovery. Int. J. Syst. Evol. Microbiol. 2024, 74, 006421. [Google Scholar] [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: a user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic. Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef] [PubMed]
- Skinnider, M.A.; Johnston, C.W.; Gunabalasingam, M.; Merwin, N.J.; Kieliszek, A.M.; MacLellan, R.J.; Li, H.; Ranieri, M.R.M.; Webster, A.L.H.; Cao, M.P.T.; Pfeifle, A.; Spencer, N.; To, Q.H.; Wallace, D.P.; Dejong, C.A.; Magarvey, N.A. Comprehensive prediction of secondary metabolite structure and biological activity from microbial genome sequences. Nat. Commun. 2020, 11, 6058. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H, Chun, J. Introducing EzBioCloud: a taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. U S A. 2009, 106, 9126–9131. [Google Scholar] [CrossRef] [PubMed]
- Kudo, F.; Mori,, A. ; Koide M.; Yajima, R.; Takeishi, R.; Miyanaga, A.; Eguchi, T. One-pot enzymatic synthesis of 2-deoxy-scyllo-inosose from d-glucose and polyphosphate. Biosci. Biotechnol. Biochem. 2021, 85, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Wehmeier, U.F.; Piepersberg,W. Enzymology of aminoglycoside biosynthesis-deduction from gene clusters. Methods Enzymol. 2009, 459, 459–491. [Google Scholar] [CrossRef]
- Gao, Q.; Thorson, J.S. The biosynthetic genes encoding for the production of the dynemicin enediyne core in Micromonospora chersina ATCC53710. FEMS Microbiol. Lett. 2008, 282, 105–114. [Google Scholar] [CrossRef]
- Bililign, T.; Hyun,C. G.; Williams, J.S.; Czisny, A.M.; Thorson,.JS. The hedamycin locus implicates a novel aromatic PKS priming mechanism. Chem. Biol. 2004, 11, 959–969. [Google Scholar] [CrossRef]
- Ahlert, J.; Shepard, E.; Lomovskaya, N.; Zazopoulos, E.; Staffa, A.; Bachmann, B.O.; Huang, K.; Fonstein, L.; Czisny, A.; Whitwam, R.E.; Farnet, C.M.; Thorson, J.S. The calicheamicin gene cluster and its iterative type I enediyne PKS. Science. 2002, 297, 1173–1176. [Google Scholar] [CrossRef]
- Hyun, C.G.; Bililign, T.; Liao, J.; Thorson, S. The biosynthesis of indolocarbazoles in a heterologous E. coli host. Chembiochem. 2003, 4, 114–117. [Google Scholar] [CrossRef]
- Hyun, C.G.; Kim, S.S.; Sohng, J.K.; Hahn, J.; Kim, J.; Suh, J. An efficient approach for cloning the dNDP-glucose synthase gene from actinomycetes and its application in Streptomyces spectabilis, a spectinomycin producer. FEMS Microbiol. Lett. 2000, 183, 183–189. [Google Scholar] [CrossRef]
Figure 1.
Neighbor-joining phylogenetic tree based on the complete 16S rRNA gene sequence extracted from the assembled genome. The relationship between closely related species of type cultures from genus Paenibacillus are demonstrated. Niallia circulans ATCC 4513 was used as an out group. The analysis, performed using MEGA 11, demonstrates that P. chitinolyticus HSCC 596T is its closest neighbor.
Figure 1.
Neighbor-joining phylogenetic tree based on the complete 16S rRNA gene sequence extracted from the assembled genome. The relationship between closely related species of type cultures from genus Paenibacillus are demonstrated. Niallia circulans ATCC 4513 was used as an out group. The analysis, performed using MEGA 11, demonstrates that P. chitinolyticus HSCC 596T is its closest neighbor.

Figure 2.
Comparison of butirosin BGCs encoded in the genomes of Bacillus vitellinus NBRC 13296, Paenibacillus chitinolyticus KCCM 41400T (SAMN08222605), P. chitinolyticus NRRL B-23119 (SAMN27675096), P. chitinolyticus NRRL B-23120 (SAMN27675097), P. chitinolyticus YSY-3.1 (SAMD00444452), P. chitinolyticus JMW06 (SAMN19998407), P. chitinolyticus B-14908 (SAMN33770086), Paenibacillus sp. GbtcB18 (SAMN18679176), Paenibacillus sp. MZ04-78.2 (SAMN28689874), and Paenibacillus sp. HGH0039 (SAMN02596731). The individually reported N. circulans ATCC 21557 (AJ781030), N. circulans ATCC 21558 (LC571042.1), and N. circulans SANK 72073 (AB097196) are also compared.
Figure 2.
Comparison of butirosin BGCs encoded in the genomes of Bacillus vitellinus NBRC 13296, Paenibacillus chitinolyticus KCCM 41400T (SAMN08222605), P. chitinolyticus NRRL B-23119 (SAMN27675096), P. chitinolyticus NRRL B-23120 (SAMN27675097), P. chitinolyticus YSY-3.1 (SAMD00444452), P. chitinolyticus JMW06 (SAMN19998407), P. chitinolyticus B-14908 (SAMN33770086), Paenibacillus sp. GbtcB18 (SAMN18679176), Paenibacillus sp. MZ04-78.2 (SAMN28689874), and Paenibacillus sp. HGH0039 (SAMN02596731). The individually reported N. circulans ATCC 21557 (AJ781030), N. circulans ATCC 21558 (LC571042.1), and N. circulans SANK 72073 (AB097196) are also compared.
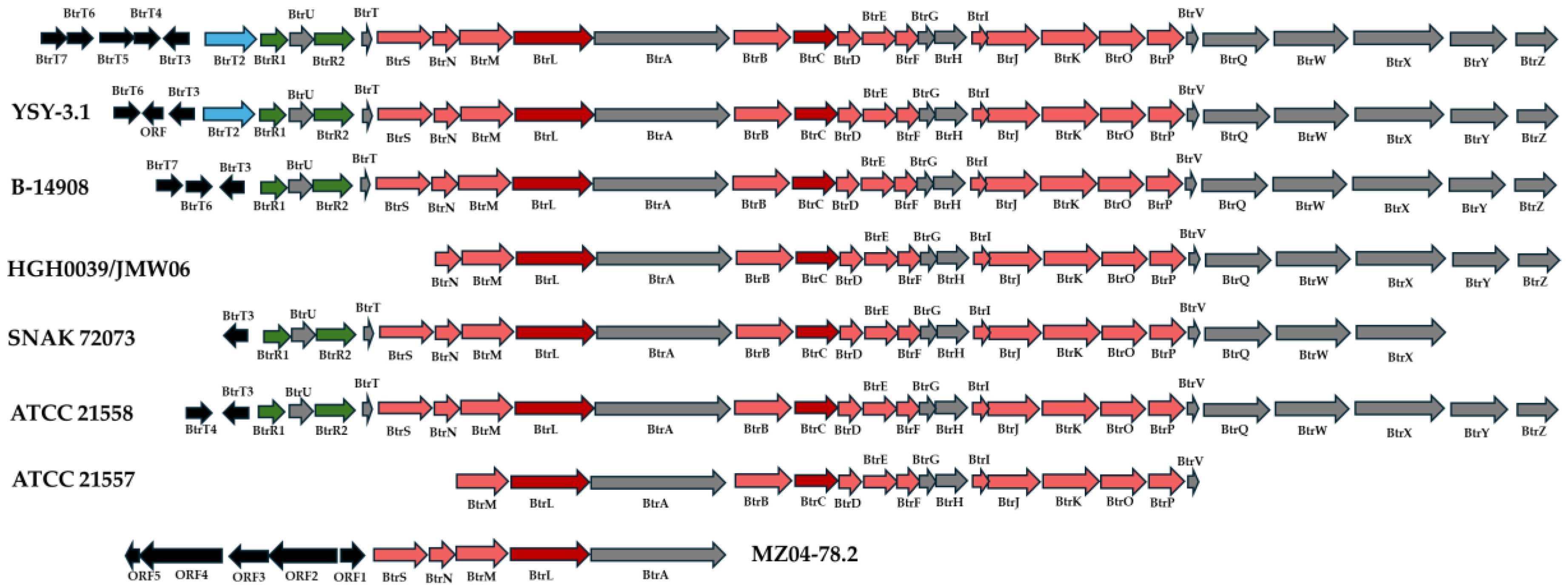

Table 1.
Putative gene clusters coding for secondary metabolites in B. vitellinus NBRC 13296.
| Region | Type | From | To | Most similar known cluster | Similarity |
|---|---|---|---|---|---|
| Region 1 | NRPS, LAP | 305619 | 356240 | ||
| Region 2 | RRE-containing | 747520 | 767798 | ||
| Region 3 | RiPP-like | 851869 | 864055 | ||
| Region 4 | Crocagin, HR-T2PKS | 945752 | 1014754 | ||
| Region 5 | TransAT-PKS, NRPS | 2461872 | 2549008 | ||
| Region 6 | TransAT-PKS, NRPS | 2563881 | 2640650 | Pelgipeptin | 25% |
| Region 7 | Cyclic-lactone-autoinducer | 2734390 | 2754992 | ||
| Region 8 | Terpene | 2821778 | 2843727 | ||
| Region 9 | Betalactone | 3032661 | 3062918 | ||
| Region 10 | TransAT-PKS, NRPS | 3175761 | 3243813 | ||
| Region 11 | Phosphonate | 3441175 | 3454343 | ||
| Region 12 | Amglyccycl | 3489373 | 3517357 | Butirosin A/B | 84% |
| Region 13 | NRPS | 3540249 | 3611046 | Octapeptin C4 | 29% |
| Region 14 | NRP-metallophore, NRPS | 3614412 | 3666638 | Bacillibactin | 100% |
| Region 15 | Opine-like-metallophore | 3687196 | 3729044 | Bacillopaline | 100% |
| Region 16 | T3PKS | 3837645 | 3878793 | ||
| Region 17 | Proteusin | 5176800 | 5197111 | ||
| Region 18 | TransAT-PKS-like, NRPS | 5276428 | 5336092 |
* “similarity” refers to the ratio of homologous genes in the query and hit clusters. As defined by antiSMASH, homologous genes were selected based on high sequence identity (>30%) and short BLAST alignments (>25%).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



