
Submitted:
14 August 2024
Posted:
15 August 2024
You are already at the latest version
Abstract
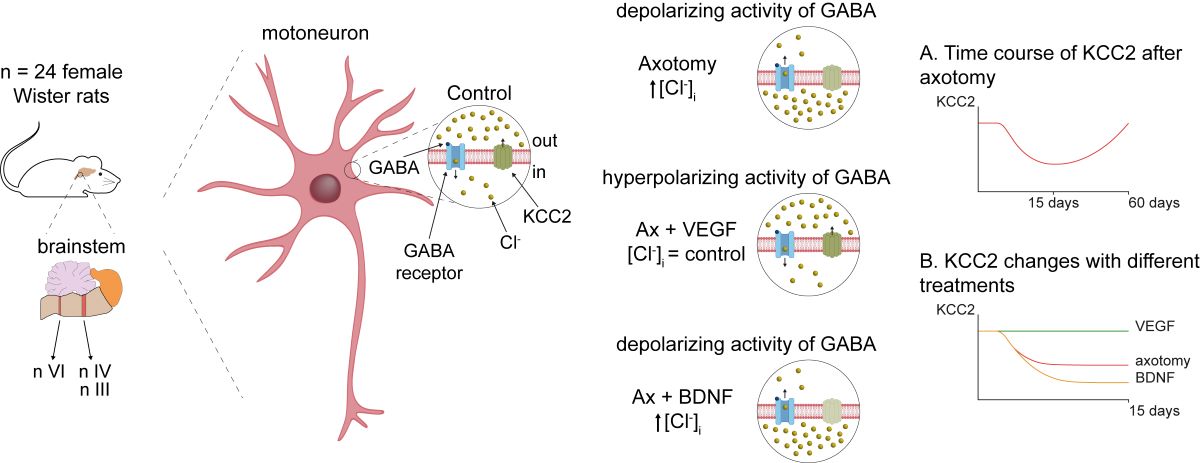
The potassium-chloride cotransporter KCC2 is the main extruder of Cl- in neurons. It plays a fundamental role in the activity of the inhibitory neurotransmitters (GABA and glycine) since low levels of KCC2 promote intracellular Cl- accumulation leading to a depolarizing activity of GABA and glycine. Downregulation of this cotransporter occurs in neurological disorders characterized by hyperexcitability, such as epilepsy, neuropathic pain and spasticity. KCC2 is also downregulated after axotomy. If muscle reinnervation is allowed, KCC2 levels recover in motoneurons. Therefore, we argued that target-derived neurotrophic factors might be involved in the regulation of KCC2 expression. For this purpose, we performed the axotomy of extraocular motoneurons by monocular enucleation of adult rats, and a pellet containing either VEGF or BDNF was chronically implanted in the orbit. Double confocal immunofluorescence of ChAT and KCC2 was carried out in brainstem sections. Axotomy led to a KCC2 decrease in the neuropil and soma of extraocular motoneurons, peaking at 15 days post-lesion, with the exception of abducens motoneuron somata. VEGF administration prevented the axotomy-induced KCC2 downregulation. By contrast, BDNF either maintained or reduced KCC2 levels following axotomy suggesting BDNF is involved in the axotomy-induced KCC2 downregulation in extraocular motoneurons. The finding that VEGF prevents KCC2 decrease opens new possibilities for the treatment of neurological disorders coursing with neuronal hyperactivity due to KCC2 downregulation.
Keywords:
oculomotor system
; cation-chloride cotransporters
; nerve injury
; choline acetyltransferase (ChAT)
; vascular endothelial growth factor
; brain-derived neurotrophic factor
; GABA depolarization
; neurological diseases
; chloride homeostasis
; NKCC1
1. Introduction
The potassium-chloride cotransporter 2 (KCC2) is the major chloride extruder in neurons, being responsible for the low intracellular concentration of this anion necessary for GABA and glycine hyperpolarizing actions [1,2,3,4]. This function is oppossed by the sodium-potassium-chloride cotransporter 1 (NKCC1), and in immature neurons predominance of NKCC1 activity over KCC2 leads to high intracellular chloride concentration, which generates the driving force for this anion to exit the cell, causing GABA and glycine membrane depolarizations [5,6,7]. During development, GABA and glycine actions switch to hyperpolarizing because up-regulation of KCC2 expression [6,8,9,10] or functional downregulation of NKCC1, as occurs in motoneurons [11,12]. KCC2 levels and activity in adult neurons are not constant and can decrease in many neurological disorders causing hyperexcitability states [13,14,15,16,17,18]. KCC2 decrease is also associated to psychiatric diseases such as schizophrenia and autism spectrum disorders [16,19,20]. In the case of motoneurons, partial removal of KCC2 membrane content after spinal cord injury results in disinhibition and spasticity [21,22,23].
Motoneurons axotomized after peripheral nerve injury are an extreme case because they stop expressing kcc2 mRNA resulting in profound removal of KCC2 in spinal [24,25,26,27] and brainstem motoneurons of the facial [28,29], hypoglossal [30] and dorsal vagal motor nuclei [31]. This causes spontaneous GABA depolarizations in motoneurons disconnected from muscle [28,30]. Recently, we extended these findings to the oculomotor system, showing that injured oculomotor and trochlear motoneurons decrease KCC2 content on their somatic membrane [32].
Motor axon reinnervation of muscle leads to KCC2 recovery in motoneurons [25,26,29,30] suggesting that muscle-derived neurotrophic factors could play important roles regulating KCC2 levels. Two potential muscle-derived neurotrophic factors are brain-derived neurotrophic factor (BDNF) and vascular endothelial growth factor (VEGF) and both have been implicated in KCC2 regulation. The actions of BDNF on KCC2 are diverse and conflicting. BDNF action on KCC2 is context- and neuron-dependent: BDNF upregulates KCC2 during postnatal development [10,33,34], but in many adult neurons, BDNF downregulates or upregulates KCC2 depending on whether they are intact or injured,respectively [21,35,36,37,38]. However, other authors have described that BDNF downregulates KCC2 after injury [39,40,41,42] and mediates the decrease in KCC2 observed in neurological diseases, such as epilepsy, neuropathic pain and spasticity [15,43,44,45]. Blocking endogenous BDNF-TrkB signaling in spinal motoneurons, either pharmacologically or genetically, does not prevent KCC2 downregulation after axotomy [25], but whether exogenous BDNF modifies KCC2 expression is unknown. Fewer studies analyzed the role of VEGF. We recently reported that VEGF administration increases KCC2 in axotomized abducens motoneurons and this correlates with increased inhibitory discharge modulation in these motoneurons. Surprisingly, abducens motoneurons are exceptional in that they do not downregulate KCC2 after axotomy [32].
The goal of the present work was to investigate the influence of VEGF and BDNF, exogenously applied, on KCC2 regulation in all three oculomotor nuclei after peripheral nerve injury. Our findings show that whereas VEGF prevented the decrease in KCC2 after axotomy, BDNF either did not affect or enhanced the KCC2 downregulation after injury.
2. Results
2.1. Time Course of Changes in KCC2 Levels after Axotomy of Extraocular Motoneurons
A time course of KCC2 expression in ChAT-identified motoneurons was performed for each nucleus studied (Figure 1). Figure 1A-E illustrates low magnification confocal images at 7, 15, 28 and 60 days post-lesion of the oculomotor nucleus in the control (Figure 1A) and the operated side (Figure 1B-E). A weak reduction in KCC2 immunolabeling could be appreciated at 7 days after injury (Figure 1B). The decrease in KCC2 labeling was much more conspicuous at 15 and 28 days (Figure 1C,D). Interestingly, 60 days after injury, the immunofluorescence appeared similar to control (Figure 1E), thereby indicating a recovery in KCC2 levels 2 months post-injury. To measure KCC2 labeling on the soma surface of motoneurons, we captured images at higher magnification, as shown in Figure 1F-J for motoneurons of the oculomotor nucleus at the same post-lesion time intervals. Oculomotor motoneurons appeared with very weak immunostaining in their somatic membrane mainly at 15 days post-axotomy (Figure 1H). KCC2 labeling recovered at longer time intervals (Figure 1I-J).
We measured KCC2 optical density in the neuropil and the somatic plasma membrane of the three nuclei and compared it with respect to control by using one-way ANOVA test followed by Fisher’s post hoc test, which revealed the following significant differences (indicated by asterisks in Figure 1K-M): OCM neuropil (F(4,10) = 26.620; p < 0.0001; Figure 1K), OCM somata (F(4,10) = 13.870; p = 0.0004; Figure 1K), TRO neuropil (F(4,10) = 11.810; p = 0.0008; Figure 1L), TRO somata (F(4,10) = 7.876; p = 0.0039; Figure 1L) and ABD neuropil (F(4,10) = 12.100; p = 0.0008; Figure 1M). KCC2 downregulation peaked at 15 days post-axotomy. At this time, the mean KCC2 optical density on the soma surface decreased to 54.59 ± 6.79% (±SD), and to 60.72 ± 13.55% in oculomotor and trochlear motoneurons, respectively. In the neuropil, optical density dropped significantly to 44.58 ± 6.93%, 66.42 ± 11.51% and 73.63 ± 10.10% in oculomotor, trochlear and abducens nuclei, respectively at 15 days post-surgery.
Axotomized abducens motoneurons did not show significant differences in KCC2 levels in their somatic membrane compared to control (p = 0.8677; Figure 1M). This result is consistent with our previous findings in cats and rats [32]. The peculiar response of axotomized abducens motoneurons contrasts markedly with other injured motoneuron pools, such as spinal motoneurons [25] as well as other cranial motoneurons (facial, hypoglossal, and dorsal vagal motor nuclei) [28,29,30,31] including the rest of extraocular motoneurons (oculomotor, trochlear) [32, present results]. Overall (except for abducens motoneuron cell bodies), the present results indicated that nerve injury induced a downregulation in KCC2, in both the dendritic (neuropil) and somatic compartments of motoneurons, with maximum downregulation occurring at 15 days post-lesion, and subsequently, KCC2 began to recover until full restoration by 60 days. The fall in KCC2 was deeper in the oculomotor, intermediate in the trochlear and least severe in the abducens nucleus.
2.2. Neuropil Changes in KCC2 7 Days after Axotomy Plus VEGF or BDNF Administration
Seven days post-lesion, faint staining of KCC2 could be appreciated in the neuropil of the oculomotor and trochlear nuclei (Figure 2D,E, respectively) compared to control (Figure 2A,B). The administration of VEGF at the time of axotomy prevented this decrease, as observed in Figure 2G for the oculomotor and in Figure 2H for the trochlear nuclei. By contrast, BDNF treatment reduced KCC2 immunofluorescence in the injured oculomotor nucleus (Figure 2J) as compared to control (Figure 2A), and axotomy states (Figure 2D). In the trochlear nucleus, however, BDNF treatment did not produce any decrease in KCC2 labeling (Figure 2K) compared to the control (Figure 2B) and axotomy states (Figure 2E). Strikingly, the abducens nucleus showed no obvious change in the intensity of immunolabeling between the different situations: control (Figure 2C), axotomy (Figure 2F), axotomy + VEGF (Figure 2I), and axotomy + BDNF (Figure 2L).
To compare the different groups, we performed a two-way ANOVA test (factors: motor nuclei and treatments). ANOVA test was followed by Fisher´s post hoc test for multiple pairwise comparisons. There were significant differences between nuclei (F(2,24) = 5.001; p = 0.0153) and between treatments (F(3,24) = 15.125; p < 0.0001). Results are represented in Figure 2M. Concerning neuropil measurements at 7 days post-axotomy, data showed a significant decay in KCC2 in the oculomotor (p = 0.0026; 80.89 ± 9.83%) and trochlear (p = 0.0024; 80.74 ± 4.92%) nuclei as compared to control (horizontal dashed line in Figure 2M, 100%). However, there was no change in KCC2 optical density in the neuropil of the abducens nucleus 7 days after injury (p = 0.689; 97.70 ± 3.43%).
VEGF treatment significantly prevented the injury-induced decrease in KCC2 that occurred in the neuropil of oculomotor and trochlear nuclei by 7 days (p = 0.0051) in both cases compared to axotomy; Figure 2M) presenting values similar to control. In marked contrast, BDNF had no effect in the trochlear nucleus versus control (p = 0.195; Figure 2M), but further reduced KCC2 levels by an additional 19.33% in the oculomotor nucleus compared to the axotomy state (p = 0.0112; Figure 2M). Finally, the abducens nucleus was the exception, as axotomy did not downregulate KCC2 levels in the neuropil 7 days after injury, and neither VEGF nor BDNF administration altered KCC2 values compared to control and axotomy.
Comparisons between nuclei within the same treatment yielded two significant differences. First, at 7 days post-axotomy, KCC2 optical density in the neuropil of the abducens nucleus was similar to control and statistically higher than in the oculomotor (p = 0.0069) and trochlear (p = 0.0065) nuclei (Figure 2M). Second, in the axotomy + BDNF situation, the oculomotor nucleus presented a significant decrease compared to trochlear (p = 0.0005) and abducens nuclei (p = 0.0005), and in turn these two latter nuclei did not differ from control (Figure 2M).
2.3. Effects of Axotomy and VEGF or BDNF Administration in KCC2 Levels on the Soma Surface of the Motoneurons at 7 Days Post-Lesion
KCC2 immunofluorescence in control motoneurons completely outlined the perimeter of motoneurons (Figure 3A-C). Axotomy reduced the intensity of perisomatic KCC2 immunofluorescence in trochlear motoneurons (Figure 3E), while axotomized oculomotor and abducens motoneurons did not appear to reduce their labeling (Figure 3D,F, respectively). Exogenous VEGF application (Figure 3G-I) maintained KCC2 levels in trochlear motoneurons similar to control values (Figure 3H). However, images obtained in the axotomy + BDNF situation revealed weak staining of KCC2 delineating the plasma membrane of oculomotor (Figure 3J) and trochlear (Figure 3K) motoneurons. By contrast, abducens motoneurons (Figure 3L) showed a similar labeling compared to control (Figure 3C).
A two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons showed significant differences within experimental situations (F(3,24) = 10.388; p = 0.0001) and nuclei (F(2,24) = 6.246; p = 0.0065).
At this time point, axotomy was not followed by a reduction in perisomatic KCC2 optical density in comparison to control in oculomotor motoneurons (Figure 3M; p = 0.1607). The administration of VEGF also rendered values similar to control (p = 0.9562). Strikingly, axotomy plus BDNF treatment did reduce KCC2 optical density in oculomotor motoneurons to 66.1 ± 17.25% as compared to control (p < 0.0001; Figure 3M).
In trochlear motoneurons, perisomatic KCC2 optical density decreased significantly (p = 0.0017) to 79.62 ± 2.14% (Figure 3M) at 7 days post-lesion compared to control. Interestingly, VEGF supply to axotomized trochlear motoneurons significantly (p = 0.0365) prevented this downregulation, yielding a result of 92.37 ± 7.95%, which was similar to control (p = 0.1973). By contrast, BDNF treatment to axotomized trochlear motoneurons produced a value (79.68 ± 10.84%) which was similar to axotomy alone (p = 0.9919) (Figure 3M). In abducens motoneurons 7 days after injury, perisomatic KCC2 optical density was similar (p > 0.05) between the three experimental conditions (axotomy, axotomy + VEGF and axotomy + BDNF) as well as with control (Figure 3M).
Comparison between nuclei within the same treatment revealed that trochlear motoneurons were the only ones that downregulated their perisomatic KCC2 values 7 days after axotomy in contrast to oculomotor (p < 0.0001) and abducens (p = 0.0017) motoneurons (Figure 3M). In the three motoneuronal types, the axotomy + VEGF situation yielded similar responses, that is, KCC2 data were similar (p > 0.05) to their respective controls (Figure 3M). Regarding the axotomy + BDNF situation, oculomotor and trochlear motoneurons showed significantly lower perisomatic KCC2 than abducens motoneurons (p < 0.0001 and p = 0.0009, respectively), which showed no change compared to control. In turn, oculomotor motoneurons showed a KCC2 downregulation which was significantly greater than that of trochlear motoneurons (p = 0.0265).
2.4. Changes in KCC2 Levels after Axotomy and VEGF or BDNF Administration in the Neuropil of Extraocular Motor Nuclei at 15 Days Post-Lesion
According to the time course of KCC2 changes after axotomy (Figure 1), KCC2 optical density reached its minimum at 15 days. The three motor nuclei showed a similar response to the different experimental situations by 15 days (Figure 4A-L). Thus, the neuropil of the three nuclei (Figure 4D,F) showed a remarkable descent in the intensity of KCC2 immunolabeling 15 days after injury compared to control (Figure 4A-C). In the axotomy + VEGF situation (Figure 4G-I), KCC2 immunofluorescence showed no change with respect to control in the oculomotor (Figure 4A,G), trochlear (Figure 4B,H), and abducens (Figure 4C,I) nuclei. It was notorious that VEGF administration (Figure 4G-I) prevented the decay in ChAT expression that occurs in axotomized motoneurons (Figure 4D-F), as we have previously described [46]. Finally, BDNF + axotomy situation showed a decrease in KCC2 (Figure 4J-L) in all nuclei that resembled the axotomy state (Figure 4D-F).
A two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons was carried out. Significant differences between experimental situations (F(3,24) = 88.406, p < 0.0001) and nuclei (F(2,24) = 19.829, p < 0.0001) were found. Axotomy reduced drastically KCC2 optical density in the three nuclei neuropil (values states above relative to Figure 1; p < 0.0001) with respect to controls (horizontal dashed line in Figure 4M).
The administration of VEGF prevented the axotomy-induced decay in KCC2 (oculomotor 96.40 ± 6.04%, p = 0.5256; trochlear 107.78 ± 4.13%, p = 0.1767; abducens 106.48 ± 10.63%, p = 0.2577). Optical density after VEGF treatment was also significantly higher (p < 0.0001) than axotomy in the three nuclei. By contrast, BDNF administration produced a mean KCC2 optical density (oculomotor 52.09 ± 3.13%, trochlear 77.44 ± 4.99%, and abducens 76.06 ± 8.86%) that was significantly lower than control (p < 0.0001, p = 0.0005, p = 0.0003, respectively) and similar to the axotomy group (p = 0.1916, p = 0.0602, p = 0.6679, respectively). Data obtained with BDNF were also significantly lower (p < 0.0001 in the three groups) than those with VEGF (Figure 4M).
Figure 4.
Effects of axotomy alone or together with administration of VEGF or BDNF on the optical density of KCC2 in the neuropil of extraocular motor nuclei 15 days after axotomy. (A-L) Confocal images showing KCC2 immunostaining (green) in the neuropil of the oculomotor (OCM; A,D,G,J), trochlear (TRO; B,E,H,K) and abducens (ABD; C,F,I,L) nuclei, in control (A,B,C), and 15 days after axotomy (D,E,F) or axotomy (Ax) + VEGF (G,H,I) or BDNF (J,K,L). ChAT immunofluorescence (red) was used to label the motoneurons. The dashed white lines in (C,F,I,L) delimit the genu of the facial nerve. (M) Bar chart of KCC2 optical density (OD) measurements in the neuropil of oculomotor, trochlear, and abducens nuclei 15 days after axotomy and after axotomy + VEGF or BDNF treatment. Data are represented as percentages relative (Re) to the control side (100%, dashed horizontal line). A two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons was used to detect significant differences between groups. The following symbols were used to indicate significant differences between treatments within the same motor nucleus: *, significant difference (p < 0.05) compared to control; §, significant difference (p < 0.05) with respect to axotomy + VEGF. Hashtags were used to indicate significant differences (#, p < 0.05) between the three extraocular motor nuclei within the same treatment. Data correspond to mean ± SD. The number of animals per group was n = 3 (black dots). Scale bar is 75 µm in (L) for (A-L).
Figure 4.
Effects of axotomy alone or together with administration of VEGF or BDNF on the optical density of KCC2 in the neuropil of extraocular motor nuclei 15 days after axotomy. (A-L) Confocal images showing KCC2 immunostaining (green) in the neuropil of the oculomotor (OCM; A,D,G,J), trochlear (TRO; B,E,H,K) and abducens (ABD; C,F,I,L) nuclei, in control (A,B,C), and 15 days after axotomy (D,E,F) or axotomy (Ax) + VEGF (G,H,I) or BDNF (J,K,L). ChAT immunofluorescence (red) was used to label the motoneurons. The dashed white lines in (C,F,I,L) delimit the genu of the facial nerve. (M) Bar chart of KCC2 optical density (OD) measurements in the neuropil of oculomotor, trochlear, and abducens nuclei 15 days after axotomy and after axotomy + VEGF or BDNF treatment. Data are represented as percentages relative (Re) to the control side (100%, dashed horizontal line). A two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons was used to detect significant differences between groups. The following symbols were used to indicate significant differences between treatments within the same motor nucleus: *, significant difference (p < 0.05) compared to control; §, significant difference (p < 0.05) with respect to axotomy + VEGF. Hashtags were used to indicate significant differences (#, p < 0.05) between the three extraocular motor nuclei within the same treatment. Data correspond to mean ± SD. The number of animals per group was n = 3 (black dots). Scale bar is 75 µm in (L) for (A-L).
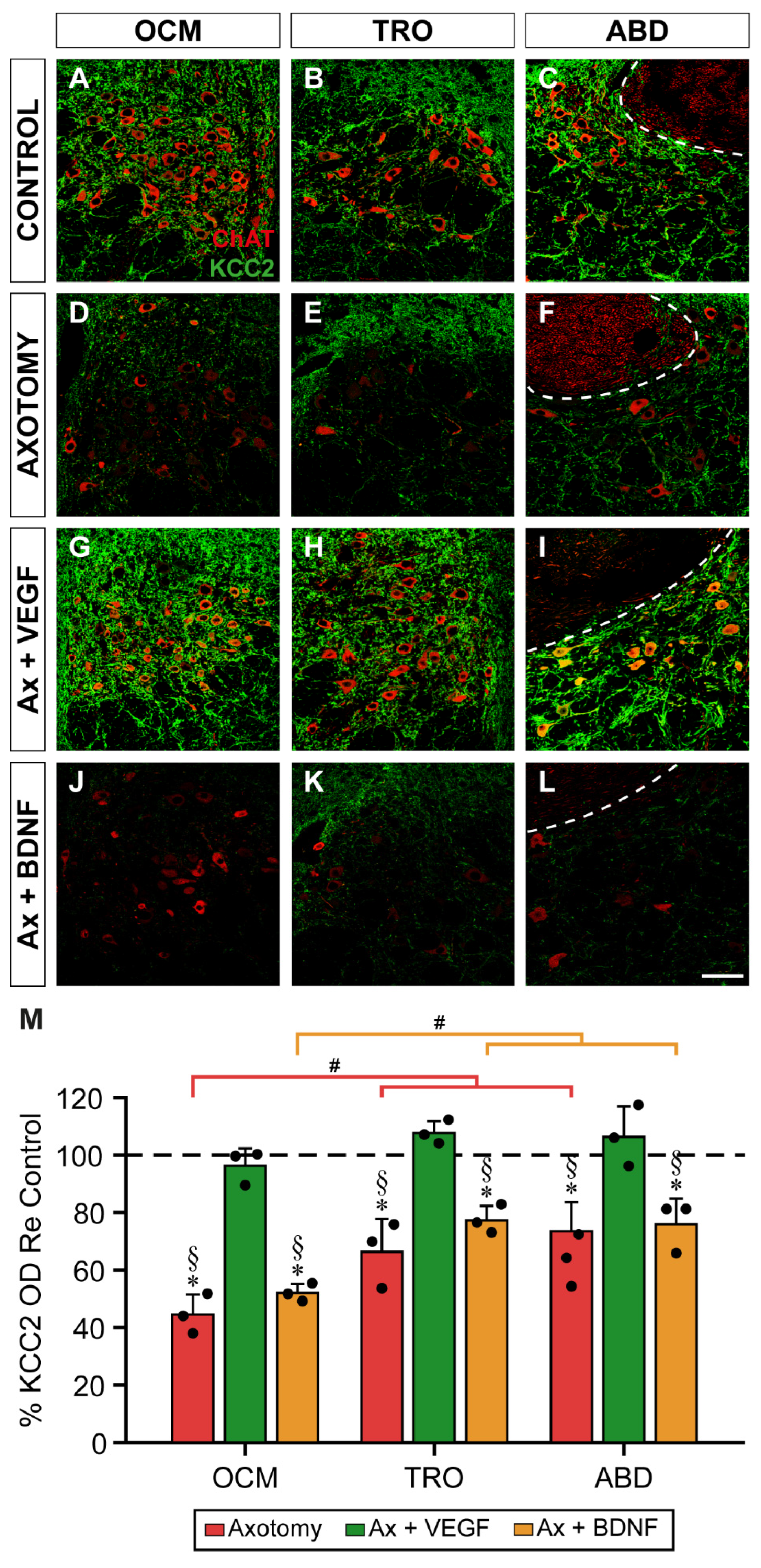
Comparisons between nuclei within the same experimental situation revealed that the oculomotor nucleus showed a mean KCC2 optical density after axotomy significantly lower than that obtained in the trochlear and abducens nuclei (p = 0.0007 and p < 0.0001, respectively). In turn, the latter two nuclei did not differ between them (i.e., trochlear and abducens, p = 0.2092) (Figure 4M). A similar finding was obtained when comparisons between nuclei were performed in the axotomy + BDNF situation. Thus, in the oculomotor nucleus, the decay in KCC2 was significantly greater than that obtained in the trochlear and abducens nuclei (p = 0.0001 and p = 0.0003, respectively), the latter two nuclei being similar to each other (i.e., trochlear and abducens, p = 0.8066) (Figure 4M).
2.5. KCC2 Optical Density in Motoneuron Plasma Membrane 15 Days after Axotomy and VEGF or BDNF Treatment
The immunofluorescence against KCC2 15 days after injury revealed that oculomotor (Figure 5D) and trochlear (Figure 5E) motoneurons exhibited a marked decrease in staining compared to their respective control (Figure 5 A,B). As we have previously reported [32], abducens motoneurons, strikingly, did not show a downregulation of KCC2 in their somatic surface 15 days after axotomy (Figure 5F, compared to control in Figure 5C), when changes were more drastic (Figure 1).
In the axotomy + VEGF situation, all motoneurons showed an intense perisomatic labeling (Figure 5G-I). However, BDNF administration did not show any change compared to axotomy in oculomotor (Figure 5J) and trochlear (Figure 5K) motoneurons. An interesting finding was that treatment with BDNF for 15 days markedly reduced KCC2 around the cell body perimeter of axotomized abducens motoneurons Figure 5L.
A two-way ANOVA test was used followed by Fisher´s post hoc test for multiple pairwise comparisons revealing significant differences between treatments (F(3,24) = 30.757, p < 0.0001), but not between nuclei (F(2,24) = 3.181, p = 0.0595).
In oculomotor motoneurons, the decrease in perisomatic KCC2 15 days post-lesion reached a mean of 54.59 ± 6.8% (Figure 5M), a value which was significantly lower than control (horizontal dashed line in Figure 5M, 100%; p < 0.0001). When VEGF was administered to axotomized oculomotor motoneurons, the mean optical density of KCC2 was 98.97 ± 4.82%, which was similar to control (p = 0.9108) and statistically higher than axotomy (p < 0.0001). By contrast, axotomized oculomotor motoneurons treated for 15 days with BDNF showed a mean perisomatic KCC2 optical density of 56.47 ± 8.97%, a value significantly lower than control (p < 0.0001), but similar to axotomy (p = 0.8387).
Similar findings were obtained in trochlear motoneurons. Fifteen days after axotomy, there was a significant (p = 0.0002) decrease of KCC2 optical density on the soma surface of trochlear motoneurons (60.72 ± 13.55%) compared to control (Figure 5M). VEGF administration to axotomized trochlear motoneurons for 15 days prevented the injury-induced downregulation of KCC2 reaching a mean optical density of 105.52 ± 16.08%, which was similar to control (p = 0.5499), and significantly higher than axotomy (p < 0.0001). By contrast, BDNF delivery produced a mean KCC2 value of 78.80 ± 16.97%, which did not differ from axotomy (p = 0.0585) but was significantly lower than control (p = 0.0286) and axotomy + VEGF (p = 0.0072) (Figure 5M). Therefore, in oculomotor and trochlear motoneurons, VEGF prevented the axotomy-induced KCC2 downregulation, while BDNF maintained the axotomy state.
Figure 5.
Analysis of the intensity of KCC2 immunofluorescence on the soma surface of extraocular motoneuron 15 days after axotomy, and VEGF or BDNF administration. (A-L) Confocal images of oculomotor (OCM; A,D,G,J), trochlear (TRO; B,E,H,K), and abducens (ABD; C,F,I,L) motoneurons (identified by ChAT immunostaining, in red, inserts in each panel) showing perisomatic immunofluorescence to KCC2 (green). Images correspond to control (A,B,C), and 15 days after axotomy alone (D,E,F) or axotomy (Ax) + VEGF (G,H,I) or BDNF (J,K,L) administration. Asterisks in (D,E,J,K,L) indicate motoneurons with very low levels of KCC2 immunostaining around their plasma membrane. (M) Bar chart showing KCC2 optical density (OD) on the soma surface of oculomotor, trochlear, and abducens motoneuron cell bodies. Data are expressed as percentages relative (Re) to the control side (100%, dashed horizontal line). A two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons was used to detect significant differences between groups. To indicate significant differences between experimental situations within the same motor nucleus we used the following symbols: *, significant difference (p < 0.05) compared to control; §, significant difference (p < 0.05) with the axotomy + VEGF situation; †, significant difference (p < 0.05) relative to the axotomy + BDNF situation. Hashtags illustrate significant differences (#, p < 0.05) between the three motoneuronal types within the same experimental situation. Data correspond to mean ± SD. The number of animals per group was n = 3 (black dots). Scale bars are 5 µm in (L) for (A-L), and 5 µm in the insert in (L) for all inserts.
Figure 5.
Analysis of the intensity of KCC2 immunofluorescence on the soma surface of extraocular motoneuron 15 days after axotomy, and VEGF or BDNF administration. (A-L) Confocal images of oculomotor (OCM; A,D,G,J), trochlear (TRO; B,E,H,K), and abducens (ABD; C,F,I,L) motoneurons (identified by ChAT immunostaining, in red, inserts in each panel) showing perisomatic immunofluorescence to KCC2 (green). Images correspond to control (A,B,C), and 15 days after axotomy alone (D,E,F) or axotomy (Ax) + VEGF (G,H,I) or BDNF (J,K,L) administration. Asterisks in (D,E,J,K,L) indicate motoneurons with very low levels of KCC2 immunostaining around their plasma membrane. (M) Bar chart showing KCC2 optical density (OD) on the soma surface of oculomotor, trochlear, and abducens motoneuron cell bodies. Data are expressed as percentages relative (Re) to the control side (100%, dashed horizontal line). A two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons was used to detect significant differences between groups. To indicate significant differences between experimental situations within the same motor nucleus we used the following symbols: *, significant difference (p < 0.05) compared to control; §, significant difference (p < 0.05) with the axotomy + VEGF situation; †, significant difference (p < 0.05) relative to the axotomy + BDNF situation. Hashtags illustrate significant differences (#, p < 0.05) between the three motoneuronal types within the same experimental situation. Data correspond to mean ± SD. The number of animals per group was n = 3 (black dots). Scale bars are 5 µm in (L) for (A-L), and 5 µm in the insert in (L) for all inserts.
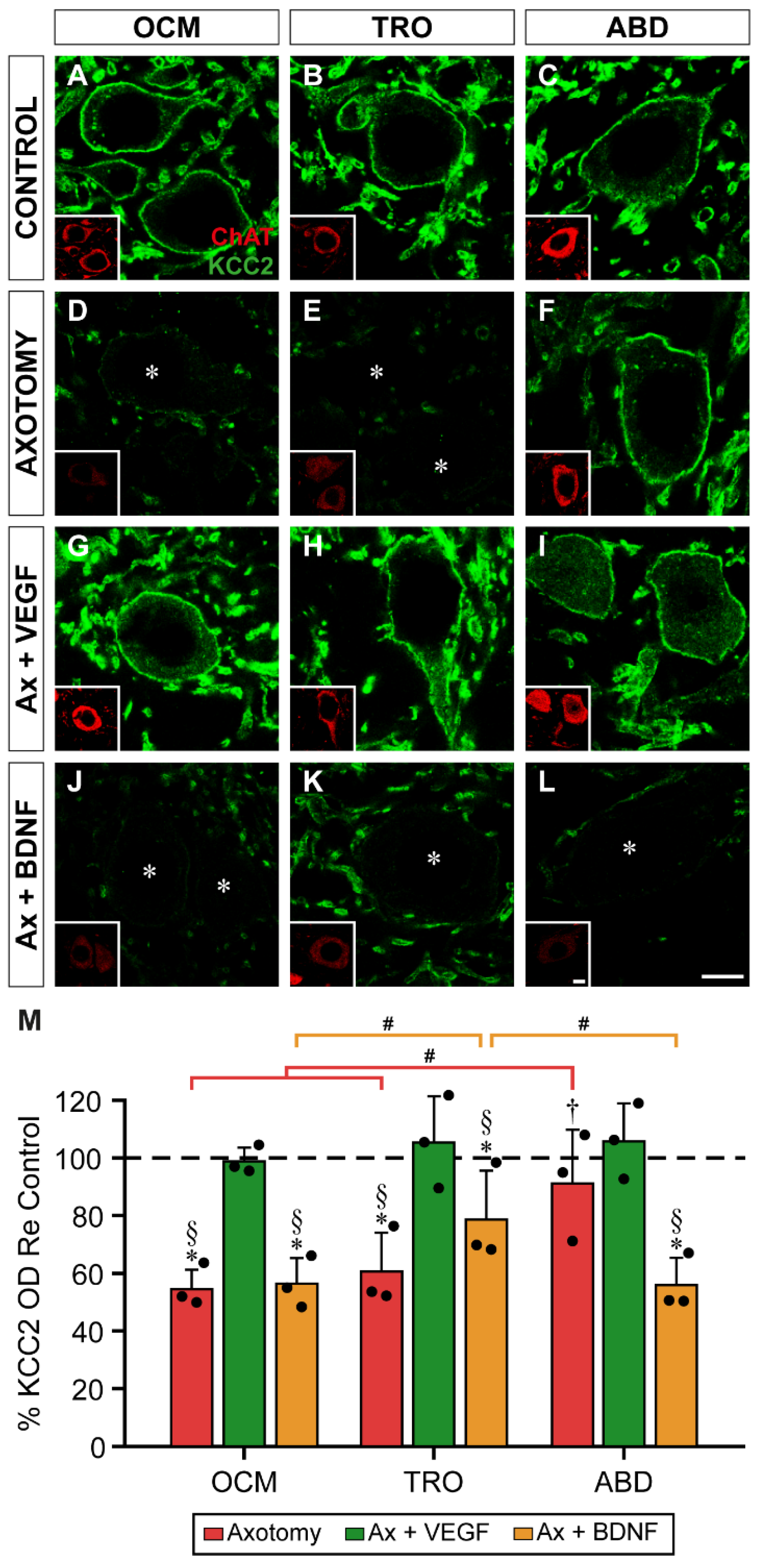
Abducens motoneurons showed a differential response to axotomy and neurotrophic factor administration. First, axotomy did not downregulate KCC2 in the somatic plasma membrane of these motoneurons at 15 days, presenting a mean optical density of 91.33 ± 18.64%, a value which was similar to control (p = 0.3502). The administration of VEGF yielded a value (105.93 ± 13.13%) similar to both control (p = 0.5211) and axotomy (p = 0.1218). The most outstanding finding in abducens motoneurons was that the administration of BDNF to injured motoneurons decreased KCC2 optical density on the soma surface of these motoneurons. Thus, abducens motoneurons in the axotomy + BDNF situation showed a value of 56.0 ± 9.54%, which was significantly lower than control (p < 0.0001), axotomy (p = 0.0007) and axotomy + VEGF (p < 0.0001) (Figure 5M).
When comparisons were performed between nuclei within the same treatment, pairwise comparisons by Fisher´s test revealed the following differences. First, perisomatic KCC2 optical density 15 days after lesion was significantly higher in abducens motoneurons than in oculomotor (p = 0.0005) and trochlear (p = 0.0026) motoneurons. Second, in the axotomy + BDNF situation, the value obtained in trochlear motoneurons was significantly higher than in oculomotor (p = 0.0218) and abducens (p = 0.0194) motoneurons (Figure 5M).
3. Discussion
The present results show that axotomy induced a general decrease in KCC2 levels in extraocular motoneurons, both at the soma surface and in the neuropil (dendritic compartment). This was particularly evident at 15 days after injury. The most remarkable finding of the present work was the ability of VEGF to prevent the decrease in KCC2 induced by injury. In contrast, BDNF administration maintained the low levels of KCC2 induced by axotomy or further reduced KCC2 below axotomy levels (i.e., oculomotor neuropil, 7 days) or, in those cases in which axotomy did not downregulate KCC2, then BDNF did induce a significant KCC2 decline (i.e., abducens soma, 15 days). The fact that BDNF clearly induced the downregulation of KCC2 in some particular groups of the present study opens the possibility that this factor might be mediating the decrease in KCC2 mostly observed in axotomized extraocular motoneurons. However, this hypothesis requires further investigation to be contrasted. In addition, the present work shows a noticeable difference in the effects obtained after the administration of VEGF or BDNF. Whereas VEGF prevented KCC2 downregulation, BDNF either did not change or induced a reduction in the levels of KCC2 in axotomized extraocular motoneurons (Figure 6).
3.1. VEGF Prevented KCC2 Downregulation in Axotomized Extraocular Motoneurons
The most outstanding result of the present work was that VEGF administration prevented KCC2 decrease induced by axotomy in extraocular motoneurons. This effect occurred in both the neuropil and soma, and was more evident at 15 days than at 7 days. However, unlike oculomotor and trochlear motoneurons, abducens motoneurons did not show axotomy-induced KCC2 depletion, as we have previously reported in cats and rats [32]. This controversial finding might be due to the different embryonic origin of abducens motoneurons [46] or may reflect different neurotrophic requirements of motoneurons or different sources of neurotrophic factors [32].
It is interesting to note that, in the cat, VEGF upregulates perisomatic KCC2 in axotomized abducens motoneurons above control values [32]. We have also shown that this KCC2 upregulation is accompanied by an increase in the inhibitory eye-related signals in the discharge of abducens motoneurons recorded under alert conditions [32]. These physiological data are in agreement with a larger chloride extrusion and therefore an enhancement of synaptic inhibitory inputs. In the present results, axotomized abducens motoneurons treated with VEGF also showed a small increase above control levels at 15 days, but this did not reach significance. This might suggest a differential response of rat versus cat abducens motoneurons to VEGF. In consequence, VEGF in the rat likely restored the normal excitatory/inhibitory synaptic balance in abducens and also other injured extraocular motoneurons.
We have also previously demonstrated that VEGF is an essential neurotrophic factor for extraocular motoneurons. The alterations in firing and synaptic properties induced by axotomy in cat abducens motoneurons are fully restored following the exogenous administration of VEGF [47]. Moreover, the retrograde intramuscular delivery of a neutralizing VEGF antibody to intact, uninjured abducens motoneurons produces severe changes in their discharge characteristics and synaptic inputs that resemble the axotomy situation [48]. In this context, it is interesting to consider that the deficit of VEGF in mutant mice VEGFδ/δ leads to motor weakness and muscle atrophy produced by the degeneration of spinal motoneurons in a model that resembles amyotrophic lateral sclerosis [49]. All these data point to VEGF as a crucial neurotrophic factor for motoneurons [50]. Future research would be needed to elucidate whether VEGF delivery could also rescue the injury-induced downregulation of KCC2 in spinal motoneurons.
The mechanism by which VEGF exerts its neuroprotective action has been described previously [51] in spinal motoneurons exposed to excitotoxicity. VEGF acting on its receptor VEGFR-2 activates the pathway of PI3-K/Akt and MEK/ERK [52]. Interestingly, these signaling mechanisms are also used by BDNF when this factor acts upregulating KCC2 (see below) [37,43].
3.2. Effects of BDNF Administration in KCC2 Levels of Axotomized Extraocular Motoneurons
The administration of BDNF to axotomized motoneurons produced diverse responses depending on the motoneuronal type and time post-lesion. Overall (particularly at 15 days after injury), it was observed that, in those cases in which KCC2 was downregulated by axotomy, the supply of BDNF did not produce any effect, that is, BDNF maintained the low KCC2 levels induced by axotomy (except in the oculomotor neuropil at 7 days post-lesion where BDNF delivery further reduced KCC2 levels). By contrast, when KCC2 was not modified by axotomy (perisomatic KCC2 in oculomotor motoneurons at 7 days and abducens motoneurons at 7 and 15 days), then the response to BDNF administration was remarkably a reduction in KCC2 levels. These data might suggest that BDNF participated in the post-injury downregulation of KCC2 in extraocular motoneurons, in agreement with previous literature. A large body of evidence has previously shown that BDNF downregulates KCC2 in different neuronal types and lesion models [42,44]. For instance, hippocampal neurons exposed to seizures induced by kindling in vivo, or the addition of BDNF in vitro from intact tissue, experience a downregulation in KCC2 mRNA and protein which is mediated by TrkB signaling [43,53]. In a mouse model of viral-induced epilepsy, BDNF levels in the hippocampus increase, which leads to a decrease in KCC2 followed by hyperexcitability and seizures [54]. BDNF-TrkB signaling is also involved in the downregulation of KCC2 in epileptic patients [17]. Similarly, spinal dorsal horn neurons experience a decrease in KCC2 after peripheral nerve injury or injection of formalin (leading to hyperalgesia) which is mediated by BDNF. Administration of inhibitors of BDNF or its signaling receptor TrkB prevents KCC2 downregulation and reduces pain [39,40,41,45]. Even the administration of BDNF intrathecally in uninjured animals evokes allodynia and decreases KCC2 expression [41]. In this context, it is worth mentioning that in some neurological diseases, such as epilepsy, neuropathic pain and spinal cord injury, a downregulation in KCC2 has also been shown to be mediated by BDNF, which underlies the characteristic hyperexcitability of these diseases due to the depolarizing activity of GABA [17,18,42,43,53]. Accordingly, selective antagonists of TrkB receptors have been suggested as likely therapeutic agents for the treatment of these neurological disorders [14,15,16,55].
However, previous reports have also shown opposite results indicating that BDNF upregulates KCC2 in lesion models such as epilepsy [36], spinal cord injury [37,56], or axotomized corticospinal neurons [35]. Some authors have postulated that BDNF decreases KCC2 in intact adult neurons, while increases KCC2 in injured adult neurons, as occurs during development [33,57], considering that neurons would return to an “immature” or “dedifferentiated” state [21,37,58].
Although facial [28,29], hypoglossal [30], vagal [31] and spinal [25,27] motoneurons experience a downregulation in KCC2 after axotomy, the regulation of KCC2 by BDNF has been evaluated only in spinal motoneurons. Thus, it has been demonstrated that KCC2 downregulation in axotomized spinal motoneurons is not altered by deletion of bdnf specifically from microglia or motoneurons, or pharmaco-genetic block of TrkB [25]. However, only one time-point coincident with maximal KCC2 depletion was examined. Our results suggest that exogenous BDNF accelerates this process, may be by promoting KCC2 removal from the membrane in the context of lack of gene expression.
The fact that BDNF can induce either downregulation or upregulation of KCC2 under different situations likely depends on the particular signaling pathway activated. BDNF-TrkB downstream cascades involve both Shc/FRS-2 and PLCγ. Downregulation of KCC2 occurs when the Shc pathway is activated and PLCγ is present. Upregulation of KCC2 is triggered by the Shc pathway when PLCγ is absent [37,43,44,58]. The exact pathways triggered in different types of axotomized motoneurons are currently unknown.
In conclusion, we have found that VEGF or BDNF administration acts differentially on axotomized extraocular motoneurons. VEGF administration prevented the decrease in KCC2 induced by axotomy. By contrast, BDNF supply, in general, maintained or reduced the low KCC2 levels induced by axotomy. Moreover, BDNF downregulated KCC2 whenever axotomy did not reduce the levels of this cotransporter (e.g., abducens motoneuron cell bodies, 15 days post-lesion). The finding that VEGF prevented injury-induced decrease in KCC2 has great clinical relevance, as it could act as a potential therapeutic agent for the treatment of neurological diseases characterized by downregulation of KCC2.
4. Materials and Methods
Adult Wistar rats were obtained from the animal house of University of Sevilla. All experimental procedures were performed following the guidelines of the European Union (2010/63/EU) and the Spanish legislation (R.D. 53/2013, BOE 34/11370-421), and were ethically approved (04/07/2022/098) for the refinement of procedures and the reduction of the number of animals.
Animals and Surgical Procedures
A total of n = 24 female Wistar rats (2-3 months old) were used in the present work. The experimental groups included: lesioned animals treated with vehicle and sacrificed at 7, 15, 28 or 60 days (axotomy alone; 12 animals), and lesioned animals treated with either VEGF or BDNF sacrificed at 7 or 15 days (axotomy + treatment; 12 animals). The number of animals used in each group was n = 3. Surgery was performed under deep anesthesia (ketamine, 80 mg/kg and xylazine, 10 mg/kg, i.p.) and consisted in the enucleation of the left eye to induce the axotomy of the motoneurons innervating the extraocular muscles on that side. The procedure for enucleation has been described in detail previously [59,60,61,62]. Briefly, the left eyeball was surgically excised, including all extraocular muscles. The orbit was then filled with an absorbable gelatin sponge pellet (Gelfoam, Pfizer, Belgium) soaked in vehicle [(20 µl of phosphate-buffer saline (PBS) pH 7.4, containing 0.1% bovine serum albumin (BSA)], or VEGF (1 µg of VEGF dissolved in 20 µl of PBS with 0.1% BSA), or BDNF (1 µg of BDNF dissolved in 20 µl of PBS with 0.1% BSA). These doses were selected according to our previous findings in electrophysiological experiments [47,63]. The recombinant rat VEGF164 amino acid form or the recombinant human BDNF (both from R&D Systems, Minneapolis, MN, USA) were applied to injured animals, which were allowed to survive for 7 or 15 days. The groups prepared for a survival of 15 days received a second dose of factors by 7 days. A time course of KCC2 changes was studied at 7, 15, 28 and 60 days post-lesion. These animals received the same surgery as described above but received only vehicle. After the implant, eyelids were sutured. Animals received postoperative care daily and were treated with anti-inflammatories and analgesics (meloxicam, 2 mg/kg, s.c.), and antibiotics (amoxicillin, 10 mg/kg, i.m.) as needed.
At the scheduled end-points the animals were sacrificed by an overdose of sodium pentobarbital (100 mg/kg, i.p.) and the vascular system perfused transcardially with 0.9% saline followed by 4% paraformaldehyde in PB. The brainstem was extracted, postfixed for 2 hours, and cryoprotected by immersion in 30% sucrose solution in PBS. The brainstem was then cut into 30 µm-thick coronal sections on a cryostat (Leica CM 1850, Wetzlar, Germany).
Extraocular motoneurons are located in three brainstem nuclei: the oculomotor (III), the trochlear (IV) and the abducens (VI) nucleus. The oculomotor nucleus contains four motoneuronal pools: the motoneurons innervating the inferior rectus, the medial rectus and the inferior oblique muscles, which project ipsilaterally, and the superior rectus subdivision, which innervates the muscle contralaterally. Since superior rectus motoneurons are located caudally in the rat oculomotor complex, we selected sections from the rostral half of the nucleus. The trochlear nucleus innervates the superior oblique muscle contralaterally, and the abducens nucleus contains the motoneurons innervating the lateral rectus muscle ipsilaterally [64].
Immunofluorescence and Quantification
Free-floating brainstem sections were processed sequentially for double immunofluorescence. The primary antibodies used were: goat polyclonal anti-ChAT, a marker for motoneurons (1:200; Merck-Millipore, Darmstadt, Germany), and rabbit polyclonal antibody against the rat KCC2 cotransporter (1:250; Millipore) in a region shared by KCC2a and KCC2b isoforms [65,66], thus the antibody recognized both isoforms. Secondary antibodies were obtained from Jackson ImmunoResearch (1:50; West Baltimore Pike, West Grove, PA, USA) and were the following: i) donkey anti-goat IgG coupled to TRITC (for ChAT detection), and ii) donkey anti-rabbit IgG coupled to FITC (for KCC2 detection).
After immunofluorescence, sections were visualized using a confocal laser scanning microscope (Zeiss LSM 7 DUO; Oberkochen, Germany). Images were captured at the level of the three extraocular motor nuclei using X40 and X63 objectives, in order to analyze KCC2 optical density at the neuropil level (area devoid of cells, mostly the dendritic compartment) and at the somatic level, respectively. Acquisition parameters were adjusted to maximize the dynamic range of the images. Series of focal planes of 0.75 µm virtual thickness were captured through the regions of interest. Z-stacks were generated by overlapping 4 focal planes of the same histological section using Zeiss imaging software ZEN. Confocal images were analyzed using the ImageJ program (NIH, Bethesda, MD, USA).
To quantify the intensity of KCC2 immunofluorescence on the motoneuron surface (i.e., perisomatic optical density), focal planes were selected at the mid-nuclear level. In each section, background measurements were taken from a 25 µm2 square region of the neuropil in the same optical plane, next to the motoneurons and lacking any somatic or dendritic labeling. The average KCC2 optical density for each cell was measured on the perimeter of the motoneuronal somata after background correction. KCC2 immunofluorescence was measured on both the control and the operated sides. Subsequently, KCC2 optical density on the somatic surface of lesioned motoneurons was normalized to that of control motoneurons from the same histological section and expressed as a percentage. The number of motoneurons analyzed per animal varied between 135 and 252, with a mean of 203.22 ± 35.41 (SD).
To analyze optical density in the neuropil, the intensity of KCC2 immunofluorescence was measured in 20 x 20 µm regions of interest (ROIs) in each extraocular motor nucleus, identified by the presence of ChAT-immunoreactive motoneurons, but avoiding cell profiles. Background correction was carried out measuring neuropil areas devoid of labeled cell profiles for each image. Thereafter, data were normalized to the control side of the same section and expressed as percentages. The number of ROIs analyzed per animal varied between 360 and 480, with a mean of 391.11 ± 40.13 (SD).
Statistics
Statistical comparisons were performed by one-way or two-way ANOVA tests using the program GraphPad Prism 10.1.2 (GraphPad Software, Dotmatics, Boston, MA, USA), at an overall significance level of p < 0.05. Data were expressed as mean ± SD.
Author Contributions
A.M.P., R.R.C. and F.J.A. designed the experiments; J.C.-L., R.G.H. and G.C.-R. performed research and analyzed data; J.C.-L., R.G.H. and P.M.C. elaborated figures and graphs; A.M.P. and R.R.C. supervised analysis; R.R.C. and A.M.P. wrote the paper; F.J.A. revised the manuscript.
Funding
This work has been funded by the I+D+i project PID2021-124300NBI00 of MICIU/AEI/10.13039/501100011033/ and by FEDER A way of making Europe (A.M.P.). Research materials were also provided by project P20_00529 Consejería de Transformación Económica Industria y Conocimiento, Junta de Andalucía-FEDER. R.G.H. is a “Ramón y Cajal” fellow supported by RYC2021-033058-I, funded by MCIN, AEI, and European Union “Next Generation EU/Recovery, Transformation and Resilience Plan”. P.M.C. is a Margarita Salas fellow. J.C.-L. is a fellowship holder of Universidad de Sevilla.
Institutional Review Board Statement
All experimental procedures were performed in adult rats following the guidelines of the European Union (2010/63/EU) and the Spanish legislation (R.D. 53/2013, BOE 34/11370-421). Special care was taken to refine procedures and reduce the number of animals and their suffering. Experiments were approved by the Animal Ethics Committee of the University of Sevilla and the local Goverment Junta de Andalucía (04/07/2022/098).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Akita, T.; Fukuda, A. Intracellular Cl- dysregulation causing and caused by pathogenic neuronal activity. Pflugers Arch. 2020, 472, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, M.A.; Uvarov, P.; Mavrovic, M.; Poncer, J.C.; Kaila, K. The multifaceted roles of KCC2 in cortical development. Trends Neurosci. 2021, 44, 378–392. [Google Scholar] [CrossRef]
- Pressey, J.C.; de Saint-Rome, M.; Raveendran, V.A.; Woodin, M.A. Chloride transporters controlling neuronal excitability. Physiol. Rev. 2023, 103, 1095–1135. [Google Scholar] [CrossRef]
- Kim, H.R.; Martina, M. Bidirectional Regulation of GABAA Reversal Potential in the Adult Brain: Physiological and Pathological Implications. Life (Basel) 2024, 14, 143. [Google Scholar] [CrossRef]
- Rivera, C.; Voipio, J.; Payne, J.A.; Ruusuvuori, E.; Lahtinen, H.; Lamsa, K.; Pirvola, U.; Saarma, M.; Kaila, K. The K+/Cl- co-transporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature 1999, 397, 251–255. [Google Scholar] [CrossRef]
- Peerboom, C.; Wierenga, C.J. The postnatal GABA shift: A developmental perspective. Neurosci. Biobehav. Rev. 2021, 124, 179–192. [Google Scholar] [CrossRef]
- Shimizu-Okabe, C.; Kobayashi, S.; Kim, J.; Kosaka, Y.; Sunagawa, M.; Okabe, A.; Takayama, C. Developmental formation of the GABAergic and glycinergic networks in the mouse spinal cord. Int. J. Mol. Sci. 2022, 23, 834. [Google Scholar] [CrossRef]
- Raol, Y.H.; Joksimovic, S.M.; Sampath, D.; Matter, B.A.; Lam, P.M.; Kompella, U.B.; Todorovic, S.M.; González, M.I. The role of KCC2 in hyperexcitability of the neonatal brain. Neurosci. Lett. 2020, 738, 135324. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y.; Cherubini, E. The GABA polarity shift and bumetanide treatment: Making sense requires unbiased and undogmatic analysis. Cells 2022, 11, 396. [Google Scholar] [CrossRef]
- Kalemaki, K.; Velli, A.; Christodoulou, O.; Denaxa, M.; Karagogeos, D.; Sidiropoulou, K. The developmental changes in intrinsic and synaptic properties of prefrontal neurons enhance local network activity from the second to the third postnatal weeks in mice. Cereb. Cortex 2022, 32, 3633–3650. [Google Scholar] [CrossRef]
- Delpy, A.; Allain, A.E.; Meyrand, P.; Branchereau, P. NKCC1 cotransporter inactivation underlies embryonic development of chloride-mediated inhibition in mouse spinal motoneuron. J. Physiol. 2008, 586, 1059–1075. [Google Scholar] [CrossRef]
- Branchereau, P.; Cattaert, D. Chloride homeostasis in developing motoneurons. Adv. Neurobiol. 2022, 28, 45–61. [Google Scholar] [CrossRef]
- Coull, J.A.; Boudreau, D.; Bachand, K.; Prescott, S.A.; Nault, F.; Sík, A.; De Koninck, P.; De Koninck, Y. Trans-synaptic shift in anion gradient in spinal lamina I neurons as a mechanism of neuropathic pain. Nature 2003, 424, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Tillman, L.; Zhang, J. Crossing the chloride channel: The current and potential therapeutic value of the neuronal K+-Cl- cotransporter KCC2. Biomed. Res. Int. 2019, 2019, 8941046. [Google Scholar] [CrossRef]
- Belperio, G.; Corso, C.; Duarte, C.B.; Mele, M. Molecular mechanisms of epilepsy: The role of the chloride transporter KCC2. J. Mol. Neurosci. 2022, 72, 1500–1515. [Google Scholar] [CrossRef]
- Lam, P.; Newland, J.; Faull, R.L.M.; Kwakowsky, A. Cation-chloride cotransporters KCC2 and NKCC1 as therapeutic targets in neurological and neuropsychiatric disorders. Molecules 2023, 28, 1344. [Google Scholar] [CrossRef]
- McMoneagle, E.; Zhou, J.; Zhang, S.; Huang, W.; Josiah, S.S.; Ding, K.; Wang, Y.; Zhang, J. Neuronal K+-Cl- cotransporter KCC2 as a promising drug target for epilepsy treatment. Acta Pharmacol. Sin. 2024, 45, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.A.; Pereira-Figueiredo, D.; Borges-Martins, V.P.; Kubrusly, R.C.; Calaza, K.C. GABAergic system and chloride cotransporters as potential therapeutic targets to mitigate cell death in ischemia. J. Neurosci. Res. 2024, 102, e25355. [Google Scholar] [CrossRef]
- Ferrari, D.C.; Ben-Ar, Y. Disruptions in chloride transporter activity in autism spectrum disorders. In Neuronal Chloride Transporters in Health and Disease; Tang, X., Ed.; Elsevier: London, UK, 2020; pp. 549–568. [Google Scholar]
- Mohaghghegh, H.; Ananloo, E.S.; Hadjighasemm, M.; Karimipour, M.; Hashemizadeh, S.; Abhari, S.A.A. NKCC1 to KCC2 mRNA Ratio in Schizophrenia and Its Psychopathology: a Case Control Study. J. Mol. Neurosci. 2022, 72, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Boulenguez, P.; Liabeuf, S.; Bos, R.; Bras, H.; Jean-Xavier, C.; Brocard, C.; Stil, A.; Darbon, P.; Cattaert, D.; Delpire, E.; Marsala, M.; Vinay, L. Down-regulation of the potassium-chloride cotransporter KCC2 contributes to spasticity after spinal cord injury. Nat. Med. 2010, 16, 302–307. [Google Scholar] [CrossRef]
- Bos, R.; Sadlaoud, K.; Boulenguez, P.; Buttigieg, D.; Liabeuf, S.; Brocard, C.; Haase, G.; Bras, H.; Vinay, L. Activation of 5-HT2A receptors upregulates the function of the neuronal K-Cl cotransporter KCC2. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Bilchak, J.N.; Yeakle, K.; Caron, G.; Malloy, D.; Côté, M.P. Enhancing KCC2 activity decreases hyperreflexia and spasticity after chronic spinal cord injury. Exp. Neurol. 2021, 338, 113605. [Google Scholar] [CrossRef]
- Mòdol, L.; Mancuso, R.; Alé, A.; Francos-Quijorna, I.; Navarro, X. Differential effects on KCC2 expression and spasticity of ALS and traumatic injuries to motoneurons. Front. Cell. Neurosci. 2014, 8, 7. [Google Scholar] [CrossRef]
- Akhter, E.T.; Griffith, R.W.; English, A.W.; Alvarez, F.J. Removal of the potassium chloride co-transporter from the somatodendritic membrane of axotomized motoneurons is independent of BDNF/TrkB signaling but is controlled by neuromuscular innervation. eNeuro 2019, 6, ENEURO.0172‐19.2019. [Google Scholar] [CrossRef] [PubMed]
- Yafuso, T.; Kosaka, Y.; Shimizu-Okabe, C.; Okura, N.; Kobayashi, S.; Kim, J.; Matsuda, K.; Kinjo, D.; Okabe, A.; Takayama, C. Slow progression of sciatic nerve degeneration and regeneration after loose ligation through microglial activation and decreased KCC2 levels in the mouse spinal cord ventral horn. Neurosci. Res. 2022, 177, 52–63. [Google Scholar] [CrossRef]
- Cheung, D.L.; Toda, T.; Narushima, M.; Eto, K.; Takayama, C.; Ooba, T.; Wake, H.; Moorhouse, A.J.; Nabekura, J. KCC2 downregulation after sciatic nerve injury enhances motor function recovery. Sci. Rep. 2023, 13, 7871. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, H.; Ohno, K.; Yamada, J.; Ikeda, M.; Okabe, A.; Sato, K.; Hashimoto, K.; Fukuda, A.J. Induction of NMDA and GABAA receptor-mediated Ca2+ oscillations with KCC2 mRNA downregulation in injured facial motoneurons. J. Neurophysiol. 2003, 89, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kobayashi, S.; Shimizu-Okabe, C.; Okabe, A.; Moon, C.; Shin, T.; Takayama, C. Changes in the expression and localization of signaling molecules in mouse facial motor neurons during regeneration of facial nerves. J. Chem. Neuroanat. 2018, 88, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Tatetsu, M.; Kim, J.; Kina, S.; Sunakawa, H.; Takayama, C. GABA/glycine signaling during degeneration and regeneration of mouse hypoglossal nerves. Brain Res. 2012, 1446, 22–33. [Google Scholar] [CrossRef]
- Nabekura, J.; Ueno, T.; Okabe, A.; Furuta, A.; Iwaki, T.; Shimizu-Okabe, C.; Fukuda, A.; Akaike, N. Reduction of KCC2 expression and GABAA receptor-mediated excitation after in vivo axonal injury. J. Neurosci. 2002, 22, 4412–4417. [Google Scholar] [CrossRef]
- Calvo, P.M.; de la Cruz, R.R.; Pastor, A.M.; Alvarez, F.J. Preservation of KCC2 expression in axotomized abducens motoneurons and its enhancement by VEGF. Brain Struct. Funct. 2023, 228, 967–984. [Google Scholar] [CrossRef] [PubMed]
- Aguado, F.; Carmona, M.A.; Pozas, E.; Aguiló, A.; Martínez-Guijarro, F.J.; Alcantara, S.; Borrell, V.; Yuste, R.; Ibañez, C.F.; Soriano, E. BDNF regulates spontaneous correlated activity at early developmental stages by increasing synaptogenesis and expression of the K+/Cl- co-transporter KCC2. Development 2003, 130, 1267–1280. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Uvarov, P.; Soni, S.; Thomas-Crusells, J.; Airaksinen, M.S.; Rivera, C. Early growth response 4 mediates BDNF induction of potassium chloride cotransporter 2 transcription. J. Neurosci. 2011, 31, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Shulga, A.; Thomas-Crusells, J.; Sigl, T.; Blaesse, A.; Mestres, P.; Meyer, M.; Yan, Q.; Kaila, K.; Saarma, M.; Rivera, C.; Giehl, K.M. Posttraumatic GABA(A)-mediated [Ca2+]i increase is essential for the induction of brain-derived neurotrophic factor-dependent survival of mature central neurons. J. Neurosci. 2008, 28, 6996–7005. [Google Scholar] [CrossRef] [PubMed]
- Eftekhari, S.; Mehrabi, S.; Soleimani, M.; Hassanzadeh, G.; Shahrokhi, A.; Mostafavi, H.; Hayat, P.; Barati, M.; Mehdizadeh, H.; Rahmanzadeh, R.; Hadjighassem, M.R.; Joghataei, M.T. BDNF modifies hippocampal KCC2 and NKCC1 expression in a temporal lobe epilepsy model. Acta Neurobiol. Exp. 2014, 74, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Lee-Hotta, S.; Uchiyama, Y.; Kametaka, S. Role of the BDNF-TrkB pathway in KCC2 regulation and rehabilitation following neuronal injury: A mini review. Neurochem. Int. 2019, 128, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Beverungen, H.; Klaszky, S.C.; Klaszky, M.; Côté, M.P. Rehabilitation decreases spasticity by restoring chloride homeostasis through the brain-derived neurotrophic factor-KCC2 pathway after spinal cord injury. J. Neurotrauma. 2020, 37, 846–859. [Google Scholar] [CrossRef]
- Miletic, G.; Miletic, V. Loose ligation of the sciatic nerve is associated with TrkB receptor-dependent decreases in KCC2 protein levels in the ipsilateral spinal dorsal horn. Pain 2008, 137, 532–539. [Google Scholar] [CrossRef]
- Tsuruga, K.; Hashimoto, T.; Kato, R.; Kato, R.; Uchida, Y.; Hase, T.; Morimoto, Y. Plantar injection of formalin in rats reduces the expression of a potassium chroride cotransporter KCC2 in the spinal cord and a kinase inhibitor suppresses this reduction. Biomed. Res. 2016, 37, 243–249. [Google Scholar] [CrossRef]
- Ma, J.J.; Zhang, T.Y.; Diao, X.T.; Yao, L.; Li, Y.X.; Suo, Z.W.; Yang, X.; Hu, X.D.; Liu, Y.N. BDNF modulated KCC2 ubiquitylation in spinal cord dorsal horn of mice. Eur. J. Pharmacol. 2021, 906, 174205. [Google Scholar] [CrossRef]
- Hu, Z.; Yu, X.; Chen, P.; Jin, K.; Zhou, J.; Wang, G.; Yu, J.; Wu, T.; Wang, Y.; Lin, F.; Zhang, T.; Wang, Y.; Zhao, X. BDNF-TrkB signaling pathway-mediated microglial activation induces neuronal KCC2 downregulation contributing to dynamic allodynia following spared nerve injury. Mol. Pain 2023, 19, 17448069231185439. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.; Voipio, J.; Thomas-Crusells, J.; Li, H.; Emri, Z.; Sipilä, S.; Payne, J.A.; Minichiello, L.; Saarma, M.; Kaila, K. Mechanism of activity-dependent downregulation of the neuron-specific K-Cl cotransporter KCC2. J. Neurosci. 2004, 24, 4683–4691. [Google Scholar] [CrossRef]
- Hudson, K.E.; Grau, J.W. Ionic plasticity: Common mechanistic underpinnings of pathology in spinal cord injury and the brain. Cells 2022, 11, 2910. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, F.; Wang, L.; Xu, Y.; Lv, L.; Duan, W.; Bai, R.; Meng, Z.; Shao, X. Involvement of the BDNF-TrkB-KCC2 pathway in neuropathic pain after brachial plexus avulsion. Brain Behav. 2022, 12, e2464. [Google Scholar] [CrossRef]
- Fritzsch, B. Evolution and development of extraocular motor neurons, nerves and muscles in vertebrates. Ann. Anat. 2024, 253, 152225. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.M.; de la Cruz, R.R.; Pastor, A.M. Synaptic loss and firing alterations in axotomized motoneurons are restored by vascular endothelial growth factor (VEGF) and VEGF-B. Exp. Neurol. 2018, 304, 67–81. [Google Scholar] [CrossRef]
- Calvo, P.M.; Hernández, R.G.; de la Cruz, R.R.; Pastor, A.M. VEGF is an essential retrograde trophic factor for motoneurons. Proc. Natl. Acad. Sci. U.S.A. 2022, 119, e2202912119. [Google Scholar] [CrossRef] [PubMed]
- Oosthuyse, B.; et al. Deletion of the hypoxia-response element in the vascular endothelial growth factor promoter causes motor neuron degeneration. Nat. Genet. 2001, 28, 131–138. [Google Scholar] [CrossRef]
- Calvo, P.M.; Hernández, R.G.; Pastor, A.M.; de la Cruz, R.R. VEGF and neuronal survival. Neuroscientist 2024, 30, 71–86. [Google Scholar] [CrossRef]
- Tovar-y-Romo, L.B.; Tapia, R. VEGF protects spinal motor neurons against chronic excitotoxic degeneration in vivo by activation of PI3-K pathway and inhibition of p38MAPK. J. Neurochem. 2010, 115, 1090–1101. [Google Scholar] [CrossRef]
- Lladó, J.; Tolosa, L.; Olmos, G. Cellular and molecular mechanisms involved in the neuroprotective effects of VEGF on motoneurons. Front. Cell. Neurosci. 2013, 7, 181. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.; Li, H.; Thomas-Crusells, J.; Lahtinen, H.; Viitanen, T.; Nanobashvili, A.; Kokaia, Z.; Airaksinen, M.S.; Voipio, J.; Kaila, K.; Saarma, M. BDNF-induced TrkB activation down-regulates the K+-Cl- cotransporter KCC2 and impairs neuronal Cl- extrusion. J. Cell. Biol. 2002, 159, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.C.; Thompson, E.G.; Sontheimer, H. Brain-derived neurotrophic factor inhibits the function of cation-chloride cotransporter in a mouse model of viral infection-induced epilepsy. Front. Cell Dev. Biol. 2022, 10, 961292. [Google Scholar] [CrossRef]
- Carter, B.M.; Sullivan, B.J.; Landers, J.R.; Kadam, S.D. Dose-dependent reversal of KCC2 hypofunction and phenobarbital-resistant neonatal seizures by ANA12. Sci. Rep. 2018, 8, 11987. [Google Scholar] [CrossRef]
- Tashiro, S.; Shinozaki, M.; Mukaino, M.; Renault-Mihara, F.; Toyama., Y.; Liu, M.; Nakamura, M.; Okano, H. BDNF induced by treadmill training contributes to the suppression of spasticity and allodynia after spinal cord injury via upregulation of KCC2. Neurorehabil. Neural Repair 2015, 29, 677–689. [Google Scholar] [CrossRef]
- Carmona, M.A.; Pozas, E.; Martínez, A.; Espinosa-Parrilla, J.F.; Soriano, E.; Aguado, F. Age-dependent spontaneous hyperexcitability and impairment of GABAergic function in the hippocampus of mice lacking trkB. Cereb. Cortex 2006, 16, 47–63. [Google Scholar] [CrossRef]
- Huang, Y.J.; Lee, K.H.; Grau, J.W. Complete spinal cord injury (SCI) transforms how brain derived neurotrophic factor (BDNF) affects nociceptive sensitization. Exp. Neurol. 2017, 288, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Morcuende, S.; Benítez-Temiño, B.; Pecero, M.L.; Pastor, A.M.; de la Cruz, R.R. Abducens internuclear neurons depend on their target motoneurons for survival during early postnatal development. Exp. Neurol. 2005, 195, 244–256. [Google Scholar] [CrossRef]
- Morcuende, S.; Matarredona, E.R.; Benítez-Temiño, B.; Muñoz-Hernández, R.; Pastor, A.M.; de la Cruz, R.R. Differential regulation of the expression of neurotrophin receptors in rat extraocular motoneurons after lesion. J. Comp. Neurol. 2011, 519, 2335–2352. [Google Scholar] [CrossRef]
- Morcuende, S.; Muñoz-Hernández, R.; Benítez-Temiño, B.; Pastor, A.M.; de la Cruz, R.R. Neuroprotective effects of NGF, BDNF, NT-3 and GDNF on axotomized extraocular motoneurons in neonatal rats. Neuroscience 2013, 250, 31–48. [Google Scholar] [CrossRef]
- Acosta, L.; Morcuende, S.; Silva-Hucha, S.; Pastor, A.M.; de la Cruz, R.R. Vascular endothelial growth factor (VEGF) prevents the downregulation of the cholinergic phenotype in axotomized motoneurons of the adult rat. Front. Mol. Neurosci. 2018, 11, 241. [Google Scholar] [CrossRef] [PubMed]
- Davis-López de Carrizosa, M.A.; Morado-Díaz, C.J.; Tena, J.J.; Benítez-Temiño, B.; Pecero, M.L.; Morcuende, S.R.; de la Cruz, R.R.; Pastor, A.M. Complementary actions of BDNF and neurotrophin-3 on the firing patterns and synaptic composition of motoneurons. J. Neurosci. 2009, 29, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Evinger, C. Extraocular motor nuclei: location, morphology and afferents. In Neuroanatomy of the Oculomotor System; Büttner-Ennever, J.A., Ed.; Elsevier: Amsterdam, Netherlands, 1988; pp. 81–117. [Google Scholar]
- Uvarov, P.; Ludwig, A.; Markkanen, M.; Pruunsild, P.; Kaila, K.; Delpire, E.; Timmusk, T.; Rivera, C.; Airaksinen, M.S. A novel N-terminal isoform of the neuron-specific K-Cl cotransporter KCC2. J. Biol. Chem. 2007, 282, 30570–30576. [Google Scholar] [CrossRef] [PubMed]
- Markkanen, M.; Karhunen, T.; Llano, O.; Ludwig., A.; Rivera, C.; Uvarov, P.; Airaksinen, M.S. Distribution of neuronal KCC2a and KCC2b isoforms in mouse CNS. J. Comp. Neurol. 2014, 522, 1897–1914. [Google Scholar] [CrossRef]
Figure 1.
Time course of changes in KCC2 levels after axotomy of extraocular motoneurons. (A-E) Low-magnification confocal images of the oculomotor nucleus (OCM) in control (A), and 7 days (7d, B), 15 days (C), 28 days (D) or 60 days (E) after axotomy (Ax). (F-J) Same as (A-E) but for images at higher magnification to illustrate KCC2 around the soma surface of motoneurons. For (A-J), double immunofluorescence for ChAT (red) and KCC2 (green) is shown. Asterisks in (H) point to three motoneurons whose KCC2 staining was extremely weak. (K-M) Quantification of KCC2 optical density (OD) in relation (Re) to control (in percentages) at different time intervals after axotomy in the three extraocular motor nuclei: oculomotor (K), trochlear (TRO, L) and abducens (ABD, M). Blue lines represent KCC2 measurements in the neuropil, and red lines are measurements at the somatic membrane. * indicates significant difference (p < 0.05) with respect to control (C) (one-way ANOVA test, followed by Fisher´s post hoc test). Data represent mean ± SD. The number of animals per group was n = 3. Scale bars are 75 µm in (E) for (A-E), 5 µm in (J) for (F-J), and 5 µm in insert (J) for all inserts.
Figure 1.
Time course of changes in KCC2 levels after axotomy of extraocular motoneurons. (A-E) Low-magnification confocal images of the oculomotor nucleus (OCM) in control (A), and 7 days (7d, B), 15 days (C), 28 days (D) or 60 days (E) after axotomy (Ax). (F-J) Same as (A-E) but for images at higher magnification to illustrate KCC2 around the soma surface of motoneurons. For (A-J), double immunofluorescence for ChAT (red) and KCC2 (green) is shown. Asterisks in (H) point to three motoneurons whose KCC2 staining was extremely weak. (K-M) Quantification of KCC2 optical density (OD) in relation (Re) to control (in percentages) at different time intervals after axotomy in the three extraocular motor nuclei: oculomotor (K), trochlear (TRO, L) and abducens (ABD, M). Blue lines represent KCC2 measurements in the neuropil, and red lines are measurements at the somatic membrane. * indicates significant difference (p < 0.05) with respect to control (C) (one-way ANOVA test, followed by Fisher´s post hoc test). Data represent mean ± SD. The number of animals per group was n = 3. Scale bars are 75 µm in (E) for (A-E), 5 µm in (J) for (F-J), and 5 µm in insert (J) for all inserts.
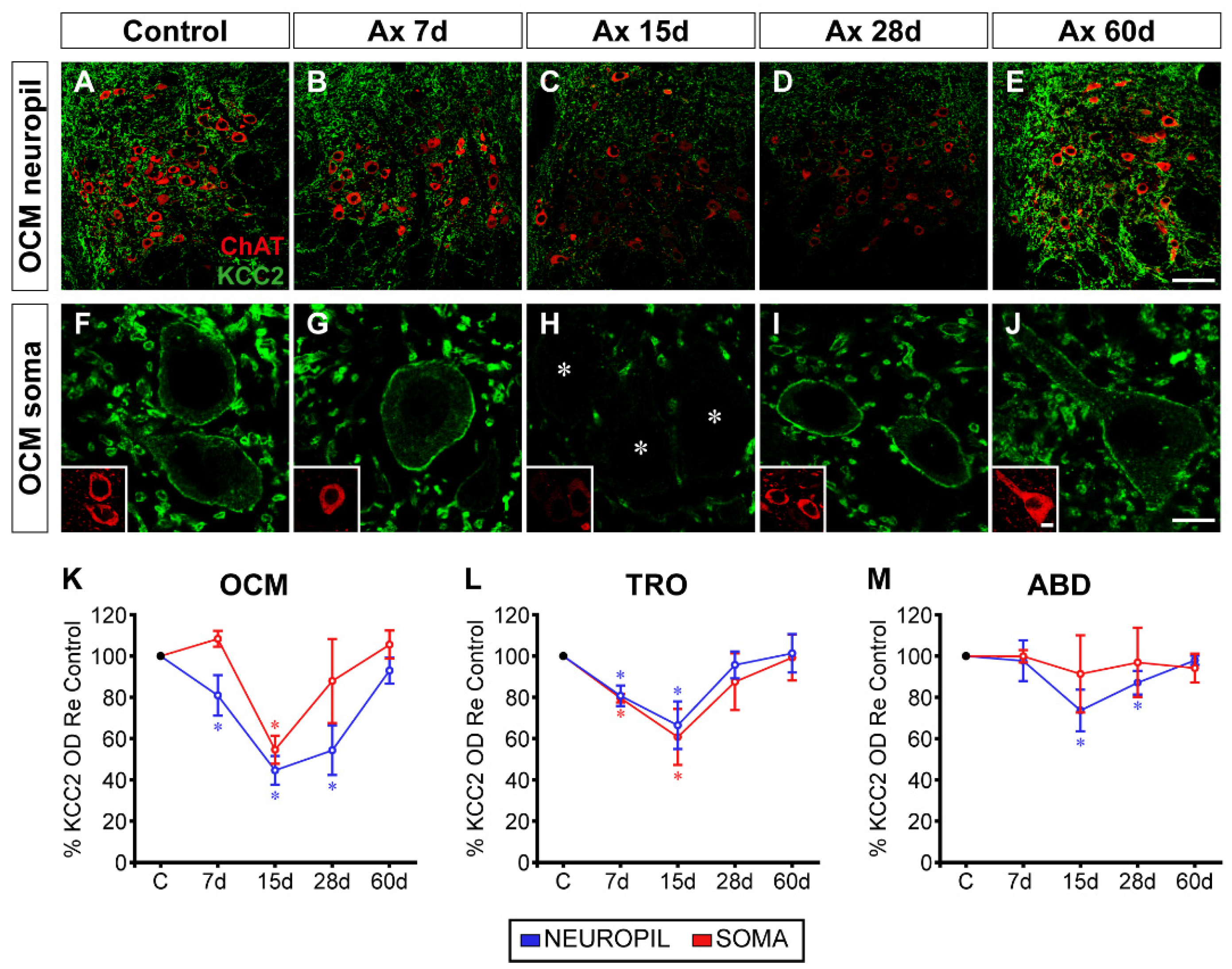
Figure 2.
KCC2 immunofluorescence in the neuropil of extraocular motor nuclei 7 days after axotomy and axotomy with VEGF or BDNF treatment. Confocal images showing KCC2 immunofluorescence (green) in the oculomotor (OCM; A,D,G,J), trochlear (TRO; B,E,H,K) and abducens (ABD; C,F,I,L) nuclei. Extraocular motoneurons were identified by ChAT (red). KCC2 immunofluorescence is shown for control (A,B,C), 7 days after axotomy (D,E,F), and 7 days after axotomy (Ax) + VEGF (G,H,I) or axotomy + BDNF (J,K,L). The dashed white lines in C, F, I, L delimit the genu of the facial nerve. (M) Bar chart illustrating KCC2 optical density (OD) in the neuropil of the oculomotor, trochlear, and abducens nuclei 7 days after axotomy and axotomy with VEGF or BDNF delivery. Data are expressed as percentages relative (Re) to the control side (100%, dashed horizontal line). The following symbols were used to indicate significant differences between treatments within the same motor nucleus: *, significant difference (p < 0.05) compared to control; §, significant difference (p < 0.05) with respect to axotomy + VEGF; †, significant difference (p < 0.05) with respect to the axotomy + BDNF group. Hashtags were used to indicate significant differences (#, p < 0.05) between the three extraocular motor nuclei within the same treatment (two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons). Data correspond to mean ± SD. The number of animals per group was n = 3 (black dots). Scale bar is 75 µm in (L) for (A-L).
Figure 2.
KCC2 immunofluorescence in the neuropil of extraocular motor nuclei 7 days after axotomy and axotomy with VEGF or BDNF treatment. Confocal images showing KCC2 immunofluorescence (green) in the oculomotor (OCM; A,D,G,J), trochlear (TRO; B,E,H,K) and abducens (ABD; C,F,I,L) nuclei. Extraocular motoneurons were identified by ChAT (red). KCC2 immunofluorescence is shown for control (A,B,C), 7 days after axotomy (D,E,F), and 7 days after axotomy (Ax) + VEGF (G,H,I) or axotomy + BDNF (J,K,L). The dashed white lines in C, F, I, L delimit the genu of the facial nerve. (M) Bar chart illustrating KCC2 optical density (OD) in the neuropil of the oculomotor, trochlear, and abducens nuclei 7 days after axotomy and axotomy with VEGF or BDNF delivery. Data are expressed as percentages relative (Re) to the control side (100%, dashed horizontal line). The following symbols were used to indicate significant differences between treatments within the same motor nucleus: *, significant difference (p < 0.05) compared to control; §, significant difference (p < 0.05) with respect to axotomy + VEGF; †, significant difference (p < 0.05) with respect to the axotomy + BDNF group. Hashtags were used to indicate significant differences (#, p < 0.05) between the three extraocular motor nuclei within the same treatment (two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons). Data correspond to mean ± SD. The number of animals per group was n = 3 (black dots). Scale bar is 75 µm in (L) for (A-L).
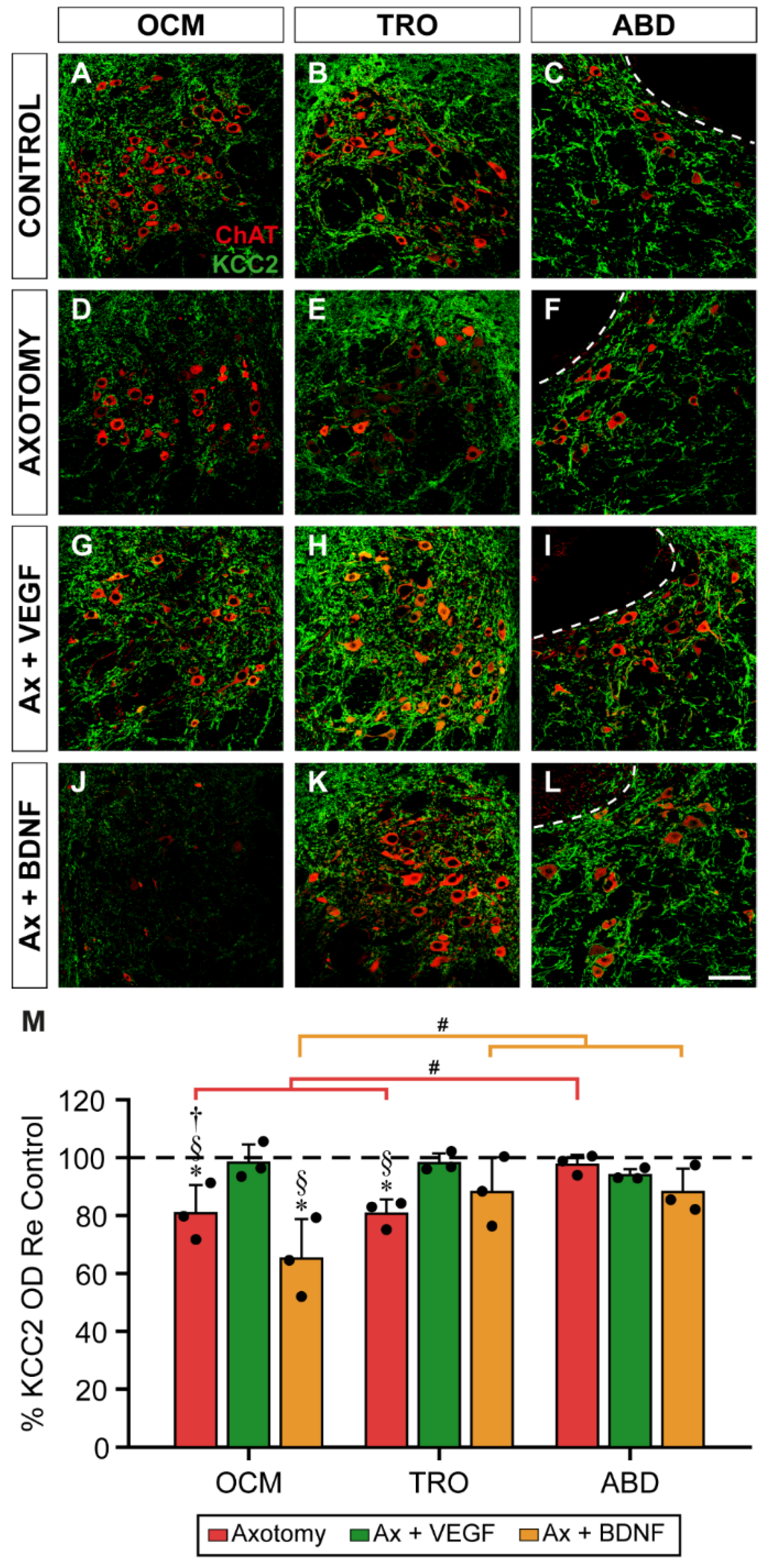
Figure 3.
KCC2 immunofluorescence in the somatic membrane of extraocular motoneurons 7 days after axotomy, and axotomy plus VEGF or BDNF. (A-L) Confocal images of oculomotor (OCM; A,D,G,J), trochlear (TRO; B,E,H,K), and abducens (ABD; C,F,I,L) motoneurons after immunostaining against KCC2 (green), in control (A,B,C), 7 days after axotomy (D,E,F) and 7 days after axotomy (Ax) + VEGF (G,H,I) or axotomy + BDNF (J,K,L). Inserts in each panel illustrate the same motoneuron/s identified by ChAT (red) immunoreactivity. Asterisks in (E,J,K) point to motoneurons whose KCC2 perisomatic staining was very faint. (M) Bar chart showing KCC2 optical density (OD) in the soma surface of extraocular motoneurons normalized with respect to (Re) the control side (100%, dashed horizontal line) and expressed as percentages, in the following situations: axotomy, axotomy + VEGF, or axotomy + BDNF. To illustrate significant differences between treatments within the same motor nucleus, the following symbols were used: *, significant difference (p < 0.05) versus control; §, significant difference (p < 0.05) versus axotomy + VEGF; †, significant difference (p < 0.05) versus axotomy + BDNF. Hashtags were used to mark significant differences (#, p < 0.05) between the three types of extraocular motoneurons within the same treatment (two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons). Data correspond to mean ± SD. The number of animals per group was n = 3 (black dots). Scale bars are 5 µm in (L) for (A-L) panels, and 5 µm in the insert in (L) for all inserts.
Figure 3.
KCC2 immunofluorescence in the somatic membrane of extraocular motoneurons 7 days after axotomy, and axotomy plus VEGF or BDNF. (A-L) Confocal images of oculomotor (OCM; A,D,G,J), trochlear (TRO; B,E,H,K), and abducens (ABD; C,F,I,L) motoneurons after immunostaining against KCC2 (green), in control (A,B,C), 7 days after axotomy (D,E,F) and 7 days after axotomy (Ax) + VEGF (G,H,I) or axotomy + BDNF (J,K,L). Inserts in each panel illustrate the same motoneuron/s identified by ChAT (red) immunoreactivity. Asterisks in (E,J,K) point to motoneurons whose KCC2 perisomatic staining was very faint. (M) Bar chart showing KCC2 optical density (OD) in the soma surface of extraocular motoneurons normalized with respect to (Re) the control side (100%, dashed horizontal line) and expressed as percentages, in the following situations: axotomy, axotomy + VEGF, or axotomy + BDNF. To illustrate significant differences between treatments within the same motor nucleus, the following symbols were used: *, significant difference (p < 0.05) versus control; §, significant difference (p < 0.05) versus axotomy + VEGF; †, significant difference (p < 0.05) versus axotomy + BDNF. Hashtags were used to mark significant differences (#, p < 0.05) between the three types of extraocular motoneurons within the same treatment (two-way ANOVA test followed by Fisher´s post hoc test for multiple comparisons). Data correspond to mean ± SD. The number of animals per group was n = 3 (black dots). Scale bars are 5 µm in (L) for (A-L) panels, and 5 µm in the insert in (L) for all inserts.
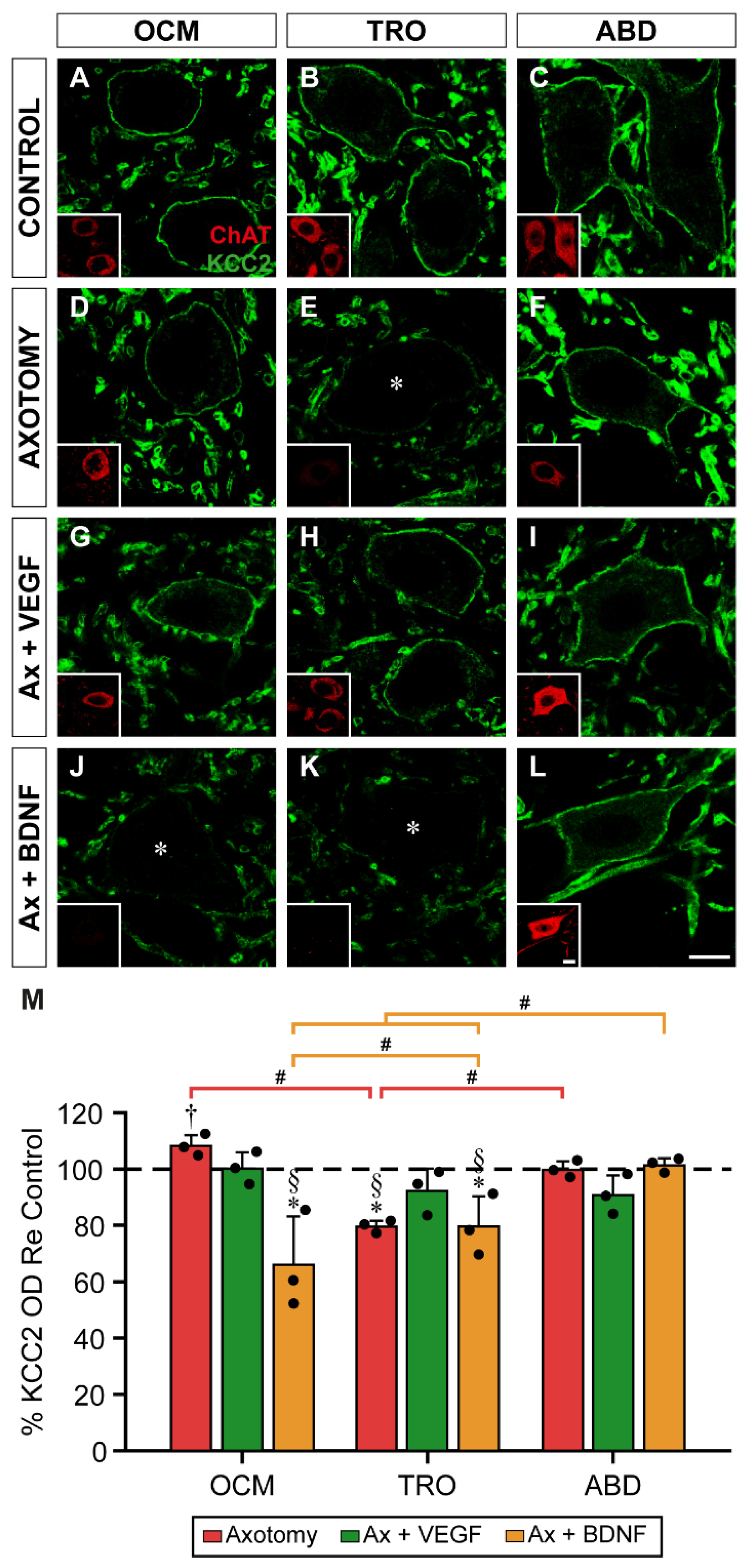
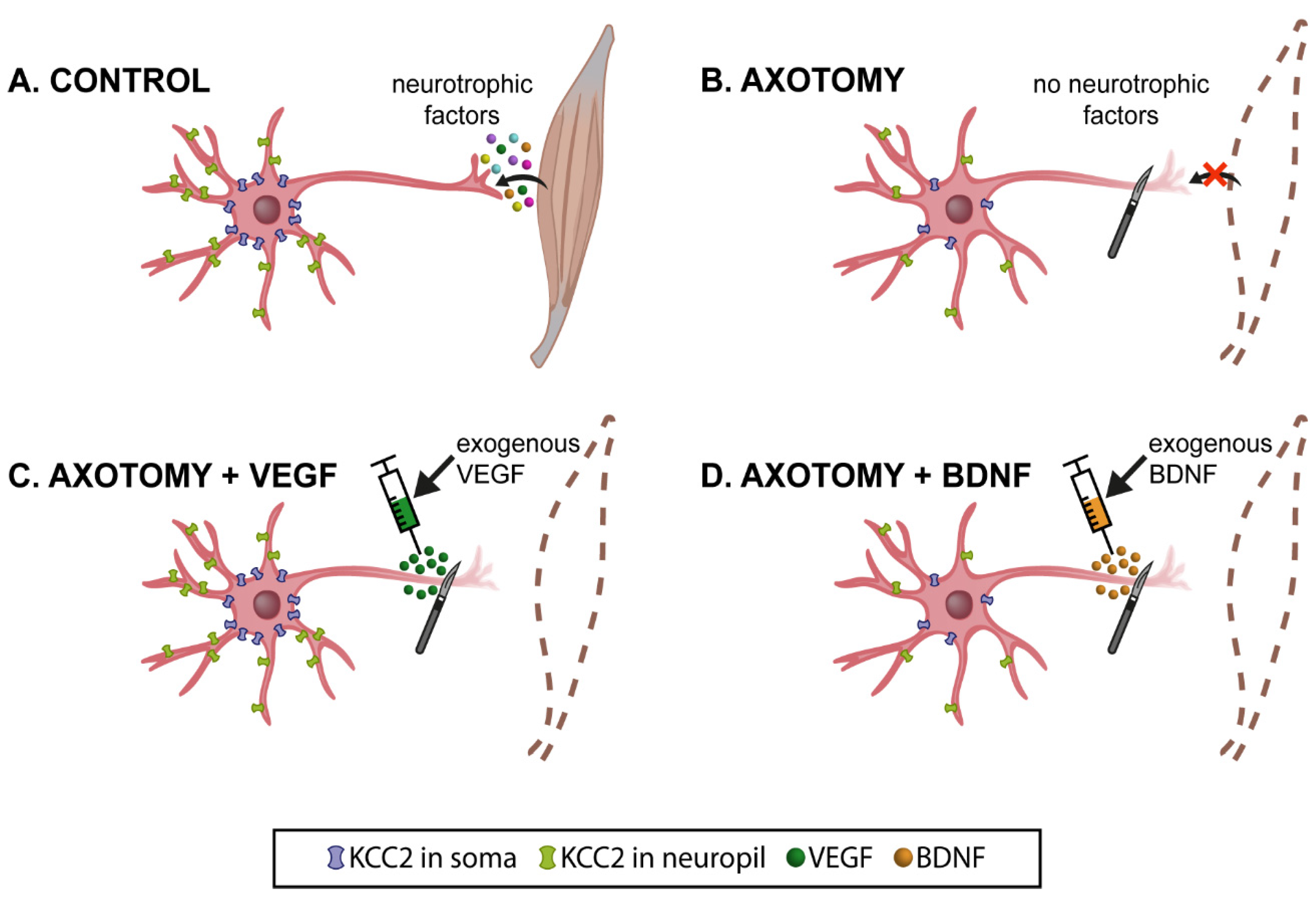
Figure 6.
Schematic illustration of the present results at 15 days post-lesion (when KCC2 levels decreased to a minimum). (A) Control extraocular motoneurons (oculomotor, trochlear and abducens) contained the cotransporter KCC2 in the plasma membrane of their dendrites (neuropil; in green) and cell bodies (in blue). These motoneurons receive neurotrophic factors retrogradely from their target muscle. (B) After axotomy, motoneurons were deprived of neurotrophic retrograde supply. Muscles were also removed (dashed brown lines). Axotomy induced a remarkable decay in the levels of KCC2 in the neuropil and soma surface of motoneurons. The only exception was abducens motoneurons at the somatic surface level. (C) VEGF administration to axotomized motoneurons prevented the KCC2 decrease induced by lesion, at the level of the neuropil and soma surface. (D) By contrast, BDNF administration to axotomized motoneurons did not modify the injury-induced KCC2 low levels observed in the neuropil and soma surface. However, it should be emphasized that, in the particular case of the somatic membrane of abducens motoneurons, whose KCC2 levels did not change after injury, BDNF caused a significant downregulation of this cotransporter. Taken together, our results suggest that VEGF is an important neurotrophic factor involved in maintaining adequate levels of KCC2 along the somatodendritic membrane of these motoneurons, while BDNF might play a relevant role mediating the downregulation of KCC2 after injury.
Figure 6.
Schematic illustration of the present results at 15 days post-lesion (when KCC2 levels decreased to a minimum). (A) Control extraocular motoneurons (oculomotor, trochlear and abducens) contained the cotransporter KCC2 in the plasma membrane of their dendrites (neuropil; in green) and cell bodies (in blue). These motoneurons receive neurotrophic factors retrogradely from their target muscle. (B) After axotomy, motoneurons were deprived of neurotrophic retrograde supply. Muscles were also removed (dashed brown lines). Axotomy induced a remarkable decay in the levels of KCC2 in the neuropil and soma surface of motoneurons. The only exception was abducens motoneurons at the somatic surface level. (C) VEGF administration to axotomized motoneurons prevented the KCC2 decrease induced by lesion, at the level of the neuropil and soma surface. (D) By contrast, BDNF administration to axotomized motoneurons did not modify the injury-induced KCC2 low levels observed in the neuropil and soma surface. However, it should be emphasized that, in the particular case of the somatic membrane of abducens motoneurons, whose KCC2 levels did not change after injury, BDNF caused a significant downregulation of this cotransporter. Taken together, our results suggest that VEGF is an important neurotrophic factor involved in maintaining adequate levels of KCC2 along the somatodendritic membrane of these motoneurons, while BDNF might play a relevant role mediating the downregulation of KCC2 after injury.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



