Submitted:
16 August 2024
Posted:
20 August 2024
You are already at the latest version
Abstract
Histamine (HA), a biogenic monoamine, exerts its pleiotropic effects through four H1R–H4R histamine receptors, which are also expressed in brain tissue. Together with the projections of HA-producing neurons located within the tuberomammillary nucleus (TMN), which innervate most areas of the brain, they constitute the histaminergic system. Thus, while remaining a mediator of the inflammatory reaction and immune system function, HA also acts as a neurotransmitter and a modulator of other neurotransmitter systems in the central nervous system (CNS). Although the detailed causes are still not fully understood, neuroinflammation seems to play a crucial role in the etiopathogenesis of both neurodevelopmental and neurodegenerative (neuropsychiatric) diseases, such as autism spectrum disorders (ASD), attention-deficit/hyperactivity disorder (ADHD), Alzheimer's disease (AD) and Parkinson's disease (PD). Given the increasing prevalence/diagnosis of these disorders and their socioeconomic impact, the need to develop effective forms of therapy has focused researchers' attention on the brain's histaminergic activity and other related signaling pathways. This review presents the current state of knowledge concerning the involvement of HA and the histaminergic system within the CNS in the development of neurodevelopmental and neurodegenerative disorders.
Keywords:
histamine
; histamine receptors
; neurodevelopmental disorders
; neurodegenerative diseases
; histaminergic neurons
; tuberomammillary nucleus
; neuroinflammation
; histamine H3 receptor
; histamine H3 receptor antagonist/inverse agonist
; autism spectrum disorders
; Alzheimer’s disease
1. Introduction
Histamine (HA) is a nitrogenous compound (an imidazole ring attached to an ethylamine chain) with versatile biological effects, both local and systemic. Since its discovery in 1910 by Henry Dale and colleagues, HA has been considered a locally acting autacoid hormone closely associated with mast cells (MCs) as an inflammatory mediator released from their secretory granules, also in the course of the allergic reaction [1,2]. HA, as a participant in the immune response to foreign pathogens, is produced by basophils and MCs located in nearby connective tissue. A well-known effect of HA is an increase in the permeability of capillaries, allowing white blood cells and some proteins (e.g., glycoproteins such as immunoglobulins) to penetrate toward the pathogen in infected tissues, as well as HA-dependent itching, resulting from direct stimulation of sensory (pain) nerve endings [3,4,5]. HA also acts on endothelial cells, causing their activation and migration, which is essential for angiogenesis [6,7,8]. Much attention has recently been paid to HA in research on the physiology and pathophysiology of the gastrointestinal tract human gut microbiota due to the ability of the human gut microbiota to produce HA [9].
A breakthrough in the understanding of the role of HA in the body was the confirmation of its function as a central neurotransmitter for the brain and spinal cord [10,11,12]. In the central nervous system (CNS), HA-producing neurons are present in the tuberomammillary nucleus (TMN) of the posterior third of the hypothalamus, from which histaminergic neurons give protrusions (axons) to innervate almost all CNS regions [13,14]. Histaminergic innervation plays an important role during brain development and then in adult life, participating in homeostasis and adult neurogenesis [12,13,14,15]. This review presents the current state of knowledge concerning the involvement of HA and the histaminergic system within the CNS in the development of neurodevelopmental and neurodegenerative disorders.
2. Histamine
2.1. Histamine Biosynthesis and Metabolism
As a biogenic amine, HA is synthesized in one step from the amino acid histidine under the influence of the enzyme L-histidine decarboxylase (HDC, EC 4.1.1.22), which is the source of histaminergic signaling, via four histamine receptors (HR1–HR4).
Once formed, HA is either stored in a form bound to, e.g., heparin, primarily in the cytoplasmic secretory granules of MCs, with the possibility of rapid release upon triggering with a variety of stimuli, or undergoes immediate enzymatic degradation. HA metabolism occurs mainly through two pathways: oxidation under the influence of diamine oxidase (DAO, E.C. 1.4.3.6), leading to imidazole acetic acid (IAA), and methylation at its imidazole Nτ atom under the action of histamine N-methyltransferase (HMT, E.C. 2.1.1.8), producing tele-methylhistamine (Nτ-MH) [16]. The resulting Nτ-MH is further metabolized by monoamine oxidase B (MAO-B, EC 1.4.3.4), producing tele-methylimidazole acetic acid (t-MIAA). IAA may exist in the form of a riboside or ribotide conjugate (Figure 1).
Histamine, a biogenic monoamine, is synthesized from the amino acid histidine by decarboxylation with the participation of the enzyme L-histidine decarboxylase (HDC). HA metabolism can take place through enzymatic degradation in two ways, involving two main enzymes: diamine oxidase (DAO) and N-methyltransferase (HMT). Oxidation catalyzed by DAO leads to the formation of imidazole acetic acid (IAA) and then imidazole acetic acid ribotide (IAA-RP) via imidazole acetate phosphoribosyl transferase (marked as I.)-mediated ribosylation and imidazole acetic acid riboside (IAA-R) through ribose dephosphorylation (marked with P symbol crossed out in red) catalyzed by phosphatases and 5-ecto’-nucleotidases (marked as II.). HA methylation at its imidazole Nτ atom under the influence of HMT leads to the formation of tele-methylhistamine (Nτ-MH), which is then converted to tele-methylimidazole acetic acid (t-MIAA) in a reaction catalyzed by monoamine oxidase B (MAO-B). The temporary products during the formation of IAA and t-MIAA are aldehydes (imidazole acetaldehyde and 1-methylimidazole-4-acetaldehyde, respectively) subject to xanthine oxidase (XO) action, which are marked in a dashed border. The methylation cycle plays a role in breaking down monoamine neurotransmitters including HA. Therefore, in the human central nervous system, the vast majority of histamine is methylated, while its metabolism catalyzed by DAO is negligible [16,17].
Importantly, HA is a substrate for DAO but not for MAO because the latter enzyme converts methyl-histamine (Nτ-MH). Oxidative deamination catalyzed by MAO-B is a typical transformation for all monoaminergic neurotransmitters and neuromodulators of the CNS. HA is an evolutionarily conserved signaling molecule in the CNS of vertebrates, and virtually all CNS HA is methylated; therefore, DAO concentrations in nervous tissue are very low [16,17].
2.2. HA receptors
HA exerts its pleiotropic biological effects through interactions with four metabotropic HA receptor types designated H1R–H4R [18]. All HA receptors known thus far belong to the rhodopsin-like family of G protein-coupled receptors (GPCRs), also known as seven-(pass)-transmembrane (7TM) domain receptors [18,19]. This means that HA receptors interact with nearby heterotrimeric G proteins located within the plasma membrane in response to the conformational changes induced by ligand binding. HA receptor activation by ligands therefore promotes the exchange of guanosine diphosphate (GDP) for guanosine triphosphate (GTP) on the Gα subunit of the heterotrimer, resulting in its dissociation from Gβγ [20]. The classification of G proteins includes four families on the basis of the type of their α subunit: Gαs, Gαi, Gαq/11, and Gα12/13 [21]. Various Gα subunits that form trimers with the remaining Gβ and Gγ subunits can activate different signaling pathways [22,23]. All types of HA receptors are present in the CNS, with the predominant expression of H1R, H2R, and H3R in brain tissue [24,25]. Despite the demonstration of functional H4R expression in the CNS, confirmation of the presence of this receptor at the protein level was initially controversial owing to the lack of appropriate antibodies meeting strict criteria for G-protein-coupled receptors [25,26,27,28,29].
The basic characteristics of human H1R–H4R, their tissue distribution and their function in health and disease are presented in Table 1, while their canonical signaling pathways in the context of the main effects of action are shown in Figure 2.
5-HT – serotonin, aa – amino acids, AA – arachidonic acid, AADE – arachidonic acid-derived eicosanoids, ACh – acetylcholine, AP-1 – activator protein 1, APCs – antigen-presenting cells, cAMP – cyclic 3’,5’ adenosine monophosphate, c-fos – protooncogene (human homolog of the retroviral oncogene v-fos), cGMP – cyclic 3’,5’ guanosine monophosphate, CNS – central nervous system, DA – dopamine, DAG – diacylglycerol, DCs – dendritic cells, ECs – endothelial cells, eNOS – endothelial nitric oxide synthase, GABA – γ-amino butyric acid, GPCRs – G protein-coupled receptors, IFN-γ – interferon gamma, IL-10 – interleukin 10, IL-12 – interleukin 12, IP3 – inositol trisphosphate, MAPK – mitogen-activated protein kinase, MCs – mast cells, NA – norepinephrine, NF-κβ – nuclear factor kappa-light-chain-enhancer of activated B cells, NK cells – natural killer cells, NO – nitric oxide, PKC – protein kinase C, PLA2 – phospholipase A2, PLC – phospholipase C, PLD – phospholipase D, Th1, Th2 – Type 1 T helper and type 2 T helper cells, respectively, VEGF – vascular endothelial growth factor
* Clobenpropit, Ciproxifan, Pitolisant and ThioperamideThioperamide are antagonists/reverse agonists of H3 receptors and therefore affect not only HA synthesis and release, but also the release of other neurotransmitters, such as glutamate, ACh, DA, 5-HT or GABA.
H1R: Gαq/11 activates PLC, which signals via ❶ DAG and PKC with formation of CBM and – after recruitment of TRAF – downstream activation of NF-κβ transcriptional activity or ❷ IP3 to enhance Ca 2+ release from endoplasmic reticulum, activating eNOS to release NO; ❸ H1R can activate also PLA2, leading to the formation of AA and AADE (prostaglandins, thromboxanes, leukotrienes, and lipoxins. H2R: ❹ AC activation by Gαs leads to subsequent signaling via cAMP and PKA with phosphorylation of CREB in the nucleus; Alternatively, the PLC is activated, which triggers signaling via DAG and IP3 (❶); ❺ Gαs promotes Ca +, Na + and Cl – chanel opening, changing the permeability of the cell membrane. H3R and H4R: ❻ Gαi inhibits AC and modulate MAPK activity via interaction with ARRB2, the recruitment of which does not depend on the formation of an active G protein complex; H3R: Alternatively, activation of Gαi can inhibit K + channels, Ca 2+ channels, and the Na+/H+ transporter via AADE (❸).
AA – arachidonic acid, AADE – arachidonic acid-derived eicosanoids, ARRB2 – arrestin beta-2, also known as beta-arrestin-2, cAMP – cyclic 3’,5’ adenosine monophosphate, AC – adenylyl cyclase, CBM – CBM signalosome complex = CARD11 (the caspase recruitment domain family member 11)—BCL10 (B cell CLL/lymphoma 10)—MALT1 (mucosa associated lymphoid tissue lymphoma translocation protein 1) paracaspase, CREB – cAMP-responsive element-binding protein, DAG – diacylglycerol, HA – histamine, IP3 – inositol 1,4,5-trisphosphate, MAPK – mitogen-activated protein kinase, NF-κβ – nuclear factor kappa-light-chain-enhancer of activated B cells, P – phosphorylation, PIP2 –phosphatidylinositol 4,5-bisphosphate, PKA – protein kinase A, PKC – protein kinase C, PLA2 – phospholipase A2, PLC – phospholipase C, TRAF6 – tumor necrosis factor (TNF) receptor-associated factor 6
3. Cerebral HA and CNS Function
3.1. Importance of HA Content and Distribution in the Brain
Immunohistochemical studies have shown that the total brain HA pool consists of two compartments: the neuronal compartment, which is related to the TMN [11], and the nonneuronal compartment, the latter consisting mainly of brain-resident MCs and vascular MCs, which are closely related functionally to vascular smooth muscle cells (VSMCs) [62]. Under physiological conditions, 60–80% of cerebral HA is estimated to contain a neuronal compartment [63].
Taking into account changes in the activity of histaminergic neurons and the variable number and functional state of cerebral MCs, the HA content in brain tissue shows significant topographic differences and variability in physiological and pathological conditions, including neurodevelopment [64,65,66]. This is even more understandable if we consider the role of this biogenic amine not only in modulating the immune response and neuroinflammation but also, above all, as a neurotransmitter influencing the properties of brain neurons and synapses [59,67,68,69]. Moreover, HA appears early during mammalian brain development, and this biogenic amine is one of the first neurotransmitters to appear during neurogenesis. The role of HA in the regulation of mammalian neural stem cell growth, proliferation and differentiation has been confirmed by the fact that the highest HA concentrations are observed in the peak phase of the primary neurulation process, i.e., the formation of the neural tube from the neural plate [70,71]. Moreover, the limitation of HA synthesis in the early developmental period of the CNS is associated with underdevelopment or disruption of the corticostriatal circuitry. It has been shown that functional deficiencies of this corticostratial signaling originating in fetal life play an important role in the pathomechanisms of Tourette's syndrome and obsessive‒compulsive personality disorder (OCPD) [72,73]. Notably, in addition to HA of neuronal origin, which is the focus of most research on brain development, nonneuronal HA may play an important role. HA and other mediators released from MCs can influence microglia-mediated neuroinflammation, which can impact brain development [62,74].
The highest HA concentrations (> 3.0 pmol/mg original tissue) in the human CNS were detected in the posterior hypothalamus, which corresponds to the location of histaminergic neurons in this anatomical structure. Relatively high concentrations of HA (> 1.5 pmol/mg) were also recorded in samples collected from the anterior hypothalamus, while the lowest HA levels (approximately 0.12 pmol/mg) were detected in the cerebellum and medulla oblongata. Intermediate values (in the minimum-maximum range) of HA concentrations were found in virtually all regions/anatomical structures of the brain [63].
In patients with senile dementia of the Alzheimer's disease type (SDAT), there is an increase in the content of HA in most lobes of the cerebral cortex, primarily within the functional areas of the cerebral cortex (cortical centers: sensory, motor and association). Additionally, the increase in HA concentration compared with that in brain tissue samples collected from people whose causes of death were not related to neuropsychiatric, neurological and/or neurodegenerative diseases was also associated with structures such as the anterior and posterior regions of the hypothalamus, putamen, caudate nucleus, nucleus accumbens, thalamus, hippocampus, pons, medulla oblongata and cerebellum. Therefore, a relationship between excess HA and disorders of cognitive function, as well as neuroendocrine, neurovascular and sleep‒wake cycle functioning, is suggested [63]. However, there are also contrasting results, showing statistically significant reductions in the levels of HA and its precursor L-histidine in the frontal, temporal and occipital cortices and caudate nucleus of patients with Alzheimer's disease [74].
Elevated release and metabolism of HA has been found in the brains of chronic schizophrenia patients, which was confirmed indirectly. The increased CNS histaminergic activity was manifested by elevated levels of the HA metabolite N-tele-methylhistamine (Nτ-MH) in the cerebrospinal fluid [75].
The HA content in the CNS changes in cases of brain tissue damage. For example, increased HA levels observed in focal ischemic brain injury may play a neuroprotective role for at least 24 hours after ischemia, thereby increasing the chances of recovery from neuronal damage [76,77]. Neuronal HA is also involved in pain perception and pain hypersensitivity through the sensitization of polymodal nociceptors, resulting in increased firing rates [78].
Increased release of neuronal HA occurs after stimulation of N-methyl-D-aspartic acid or N-methyl-D-aspartate (NMDA) receptors and μ-opioid receptors, as well as dopamine D2 receptors and some serotonin receptors (5-HTRs) [10].
3.2. Histaminergic System in the Human Brain
In all mammals studied, including humans, the only source of neuronal HA in the CNS is a loose constellation of neurons within the TMN, which are scattered around the third ventricle and the mamillary body in the ventral posterior hypothalamus [11,79]. The number of neurons within the TMN whose ability to synthesize HA from L-histidine is confirmed by the expression of histidine decarboxylase (HDC) is estimated to be in the range of 60,000–125,000 and may change in various disorders (e.g., increased narcolepsy) [14,80,81,82]. The number of histaminergic neurons in the human brain is much greater than that in the mouse and rat brains, and they occupy a proportionally larger part of the posterior hypothalamus [83,84].
As is typical for arousal systems, HA axons of TMN neurons diffusely innervate almost all CNS regions, including the densest axonal projections, which are sent to the cerebral cortex (mainly to the frontal, parietal and occipital lobes), hippocampus and amygdala, ventral tegmentum, nucleus accumbens and substantia nigra, midline thalamic region, striatum, cerebellum, medulla oblongata and spinal cord [65,84,85,86]. In light of the results published by Venner et al. [87], however, the essence of the action of histaminergic TMN neurons requires further research. By assessing the influence of chronic disruption of the inhibitory neurotransmitter GABA synthesis and GABAergic transmission in TMN neurons on the sleep-wake cycle in male mice, these authors showed that these neurons “are neither necessary nor sufficient for the initiation and maintenance of arousal under baseline conditions” [87]. The reason for these findings may be that only a few TMN neurons contain the vesicular GABA transporter, which is presumably necessary to release GABA.
The axial spread of histaminergic tuberomammillary neurons is shown in Figure 3.
The specificity of signaling in the histaminergic system of the brain results from the fact that H3Rs are expressed within the neuron body, dendrites and axons, as well as on the axons of other nerve cells, whereas H1Rs and H2Rs are expressed on nonneuronal target cells. H3R stimulation induces a negative feedback loop, a type of self-regulating system leading to a reduction in the synthesis and release of HA, as well as the release of other neurotransmitters, such as ACh, NA and glutamate [65,80,88,89,90].
The functional expression of H4Rs in brain neural tissue is still insufficiently documented or nonexistent. However, H4Rs are expressed in the brain by nonneuronal cells (e.g., macrophages, MCs, and T lymphocytes), the number of which may change during various periods of life and under various pathological conditions [91,92,93].
Brain HA has been shown to be a mediator of arousal, and activation of TMN neurons promotes wakefulness, whereas TMN neurons fire little during the nonrapid eye movement (NREM) phase of sleep and are usually silent in rapid eye movement (REM) sleep [11,57,94]. The histaminergic system has also been shown to play a fundamental role in cognitive processes (e.g., learning) and the consolidation, retrieval and expression of memory [95,96]. The results of the latest research suggest that the formation or retrieval of recognition memory requires an appropriate level of tonic activity in TMN neurons. This finding suggests that the enhancement of cerebral histaminergic signaling may promote the recovery of seemingly lost memories [97].
Electrographically documented induction of arousal is usually accompanied by disparate behaviors, including those aimed at maintaining energy homeostasis [98]. The brain's histaminergic system modulates energy expenditure and caloric intake, also influencing thermoregulation and the circadian rhythm [23,98,99,100,101].
The fact that the H1R-H4R receptor system affects many different vital functions of the body may result not only from the variable expression of appropriate HA receptors on target cells but also from the existence of functionally specialized subpopulations of neurons within the TMN, which also show differences in terms of electrophysiology [14,79,102].
Two different functions of cerebral HA should be distinguished: neuromodulatory and classical neurotransmitter [10]. The neuromodulatory function of the histaminergic system results from the fact that most of the HA released by tuberomammillary cells does not enter the synaptic clefts but stimulates target cells via the wide diffusion pathway. A "slow" transmission mechanism is then used; after HA binds to the receptor, intracellular synthesis of the second messenger is necessary. This finding is consistent with the characteristics of histamine metabotropic receptors [103]. The "fast" effect of HA as a neurotransmitter involves its diffusion across the synaptic cleft and the activation of the H3 receptor located in the postsynaptic membrane, with a subsequent change in the functional state (opening or closing) of the controlled ion channel and a change in the permeability of the cell membrane. For example, electrical stimulation of the TMN has been shown to lead to the appearance of fast excitatory postsynaptic potentials in phasically firing neurons of the supraoptic nucleus [104]. However, owing to the significant technical difficulties associated with in vivo hemodynamic measurements, the basic neurochemical aspects of the function of HA as a neurotransmitter remain undetermined [105,106].
The brain's histaminergic system is not isolated from other neurotransmission systems, and mutual interaction patterns may differ depending on the CNS region. Stimulation of the hetero H3 receptor inhibits the release of ACh, 5-HT, DA, and NA, whereas activation of H1R and H2R may stimulate some systems related to these neurotransmitters. The interaction of locally elevated concentrations of some of these neurotransmitters (e.g., 5-HT, DA) with their own receptors may, in turn, contribute to increased HA release [10,11,107].
Given the neuroanatomical specificity of the histaminergic system of the brain, its physiological role is related to its ability to increase neuronal excitability in virtually the entire CNS [108].
4. Histaminergic Dysfunction in the Developing and Adult CNS
Disturbances in HA signaling and subsequent abnormalities related to other neurotransmitters may occur both during the developmental stage of the CNS and in adulthood [109,110,111,112]. In the first case, neurodevelopmental defects usually occur, and in the second case, clinical symptoms of neurodegenerative diseases appear. However, neurodegenerative changes may be initiated at an early stage of CNS development, which is why developmental motifs are observed during the progressive and selective loss of neuronal function in adulthood [113]. In other words, some neurodegenerative diseases may be caused, or at least predefined, by disorders that arise during neurodevelopment and probably sensitize nerve cells to become susceptible to degeneration later in life [114]. Interestingly, the pathogenetic molecular mechanisms (including some key causative genes) underlying neurodevelopmental and neurodegenerative disorders overlap. These findings indicate that mitochondrial dysfunction, defects in ribonuclease protein processing, disorders of protein aggregation, synaptic plasticity, neuronal cell morphology and the proliferative capacity of stem cells are, to varying degrees, similar in neurodegenerative and neurodevelopmental diseases [113,115].
Notably, neurogenesis not only occurs during normal prenatal and early postnatal development but also takes place in restricted brain regions (the olfactory bulb, the hypothalamus, and the hippocampal dentate gyrus) in adult mammals, including humans [116,117,118]. The histaminergic system of the brain may therefore be involved in the process, which is distinct from prenatal neurogenesis, in which neurons are generated from neural stem cells in the adult.
4.1. The Role of HA and Neuroinflammation during the Development of the CNS
HA, as a modulator of the properties of neurons and synapses in the brain involved in neurogenesis and gliogenesis, plays an important role at every stage of CNS development [66]. For example, in the human substantia nigra, synapses appear from the 12th week of pregnancy and mature in the 16th week [119,120]. On the basis of studies of the rodent brain, HA has been established as one of the earliest neuromodulators of embryonic development [121]. Temporary neuronal sources of HA in the early stages of CNS development, before the generation of histaminergic neurons within the TMN, have been found in various locations, including the rhombencephalon, mesencephalon, and some regions of the diencephalon [122]. This transient embryonic histaminergic system includes, among others, serotonergic neurons from the raphe nucleus that coexpress HA, thalamic neurons and ependymal cells lining the ventricles. A characteristic feature of this system is an extensive network of histaminergic fibers in various areas of the developing brain [123]. Additionally, the HA pool is supplemented by MCs, which occur in two locations, namely, the pia and the brain parenchyma [124]. Importantly, the expression of H1R, H2R and H3R in neural stem cells (NSCs) was confirmed in vitro [125]. Consistently, H1R activation is accompanied by the promotion of NSC differentiation toward neurons, whereas H2R activation enhances the proliferation of cortical neuroepithelial cells in the presence of basic fibroblast growth factor (bFGF) [123,126].
The relationship between HA and neuroinflammation is extremely complex, especially in the case of neurogenesis [66]. First, depending on the cytokine profile, HA can locally mediate a pro- or anti-inflammatory response. Different HA concentrations may be accompanied by shifts in the cytokine composition, with a predominance of proinflammatory [e.g., interleukin 1 beta (IL-1β), IL-33, and tumor necrosis factor alpha (TNFα)] or anti-inflammatory [e.g., transforming growth factor beta (TGFβ)] cytokines [62,127,128,129,130,131]. The source of HA may define the composition of cytokines in its environment and thus determine the neuroprotective or neurotoxic properties of this monoamine. Neuronal HA is stored in the cell body and, owing to the action of the vesicular monoamine transporter (VMAT2), in vesicles within the axonal varicosities [65,132,133]. Therefore, neuronal HA operates in a different environment than HA released after MC degranulation [134,135,136,137]. Moreover, as mentioned earlier, changes in the expression of histamine receptors and their functional state have activating (H1R, H2R) or inhibitory (presynaptic H3R) effects on both the histaminergic system and other systems via different neurotransmitters [138,139]. Therefore, in certain developmental stages of the CNS, neuroinflammation may occur with HA deficiency and, in other stages, with HA excess. Interestingly, while a properly functioning blood‒brain barrier (BBB) prevents the passage of somatic (blood) HA into the adult CNS, during development, the BBB is permeable to HA [123].
The local concentration of HA may be influenced by its reuptake by both astrocytes and neurons. Both types of cells have HA transport systems related to organic cation transporter 3 (OCT3) and plasma membrane monoamine transporter (PMAT) [140,141]. In addition to their important role in regulating extraneuronal HA clearance, astrocytes are also responsible for key phenomena related to neurogenesis, such as the survival/migration of neurons, the direction of their axon growth, participation in synapse formation/maturation/elimination (pruning) and neural circuit development [142].
The spectrum of disorders and diseases arising during CNS development in which a significant role in its pathomechanism is played by neuroinflammation with abnormal histaminergic activity is constantly being supplemented [59,66,118]. These disorders can be classified into two large groups:
- Neurodevelopmental disorders, including intellectual disability (intellectual development disorder), communication disorders, autism spectrum disorders (ASD), attention-deficit/hyperactivity disorder (ADHD) – the most prevalent neurodevelopmental disorder worldwide, neurodevelopmental motor disorders (e.g., Tourette’s syndrome), specific learning disorders and schizophrenia [143,144].
- Neuropsychiatric (neurodegenerative) disorders, including Parkinson's disease (PD), Alzheimer's disease (AD), Huntington's disease (HD), depression, and narcolepsy [57,145,146].
The above disorders adversely affect the functioning of the affected people, impairing to varying degrees one or more spheres of functioning (e.g., personal, social, academic, occupational), depending on the type of deficits caused. These disorders are commonly comorbid conditions, so a person affected by one of these disorders will usually meet the criteria for another disorder within the above groups [147,148]. Neurodevelopmental disorders generally appear in early childhood and can persist into adulthood. The clinical symptoms of neurodevelopmental disorders are largely the result of the formation of abnormal key neuronal circuits at the early stages of development of the nervous system, the effects of which last throughout life [149]. However, a detailed elucidation of how the dysregulation of HA itself impairs the formation/function of these neuronal circuits requires further investigation [11].
The current state of knowledge concerning the involvement of histamine signaling and neuroinflammation in the etiopathogenesis of neurodevelopmental disorders is presented in Table 2. A similar summary of neuropsychiatric diseases, which most often manifest themselves clinically after the completion of CNS development, is presented in the next subsection about neurodegenerative diseases (see Table 3).
4.2. The Role of HA and Neuroinflammation in Neuropsychiatric (Neurodegenerative) Disorders
An inflammatory background accompanies almost all neurodegenerative diseases in the CNS, although the specificity of this inflammation may differ significantly [170,171,172]. Moreover, many neurodegenerative diseases do not occur in isolated form but cooccur (overlap) with other disorders, including neurodevelopmental disorders [173,174]. This is due to, among other factors, the heterogeneity of neurodegenerative diseases, genotype differences, the influence of epigenetic factors, changes related to brain aging and the co-occurrence of common diseases [175,176]. The co-occurrence of various diseases within the CNS, including the overlap of neurodegenerative disorders, may explain the fact that the degree of neurodegenerative changes indicating neuronal network disintegration in brain tissue does not directly translate into the parameters of patients' mental assessment [177,178,179].
Neuropsychiatric disorders are easily associated with the improper functioning of histaminergic transmission pathways in the brain because HA, either directly or through other neurotransmitter systems, plays an important role in the maintenance of wakefulness, appetite regulation, mood, cognition (including learning and memory) and arousal [57]. The intensity of the inflammatory response may be crucial because hematopoietic and immune cells that flow into the CNS, such as mast cells (MCs), basophils, eosinophils, antigen-presenting cells (APCs) and CD4(+) T cells, express H4Rs [59,180]. Interestingly, under conditions of increased H4R expression in a specific location in the CNS, HA may either intensify neuroinflammation, contributing to a vicious cycle, or have anti-inflammatory effects [66,181]. Under these conditions, the direction of HA action is determined by the presence of a pro- or anti-inflammatory (damaging or neuroprotective) cytokine profile in the immediate environment.
Despite the diverse, often not fully understood etiology of neurodegenerative diseases, such as Parkinson's disease (PD), Alzheimer's disease (AD), Huntington's disease (HD) depression and narcolepsy, their common denominator is the presence of neuroinflammation [171,182,183]. Aberrant activation of apoptosis (programmed cell death) pathways is a common feature in these diseases that may result in unwanted loss of neuronal cells and function in a specific region of the brain [184]. Typically, neurodegenerative diseases involve the formation and intracellular deposition of protein aggregates [e.g., α-synuclein aggregations in the form of Lewy bodies (LBs) in PD patients and other α-synucleinopathies, such as dementia with LBs, multiple system atrophy, pure autonomic failure or rapid eye movement (REM) sleep behavior disorder] due to protein misfolding or a lack of degradation, as well as excessive formation of reactive oxygen species (ROS) and reactive nitrogen species (RNS) promoting oxidative stress, endoplasmic reticulum stress (ERS), mitochondrial dysfunction, lysosomal dysfunction, alterations in calcium homeostasis and aberrant adult neurogenesis [185,186,187].
The activation of glial cells (oligodendrocytes, microglia and astrocytes) during the inflammatory response plays a key role in neuroinflammation, which is also due to the involvement of astrocytes in the formation and maintenance of the integrity of the BBB [88,188,189,190].
Naturally, it is important to determine whether and to what extent disorders of the histaminergic system in the CNS, and in particular neurodegenerative diseases, contribute to neuroinflammation or are rather the result of neurodegeneration. This is not obvious, although attempts to influence individual histamine receptors with agonists, antagonists and inverse agonists provide valuable information (see Chapter 5). When the involvement of the brain's histaminergic system in the pathomechanism of neurodegenerative diseases is analyzed, the HA content in nervous tissue and the level of expression of histamine receptors of a specific type in the CNS should be considered. The role of HA and histamine receptor signaling in selected representative neurodegenerative diseases in the context of neuroinflammation is summarized in Table 3.
5. Brain HA Signaling as a Therapeutic Target in Neurodevelopmental and Neurodegenerative Disorders
The complex level of interaction of histaminergic neurons with many other neurotransmitter systems makes the possibility of influencing histamine receptors within the CNS a potential therapeutic target [85,88,211]. This also applies to congenital and acquired diseases of the nervous system, the etiopathogenesis of which is still unclear, and disorders whose essence is only indirectly related to HA [57,145,212]. In such cases, treatment may only address the symptoms, not taking into account the causes of the disease (not specific), but not without effectiveness resulting from the interconnection of HA and other signaling pathways during the regulation of neuroendocrine, cardiovascular and mental functions (e.g., arousal), including brain blood flow (vascular dynamics), BBB integrity, sleep and wakefulness, memory, cognition, learning ability, and eating and drinking behaviors [15,64,66,192,213]. Importantly, the expression of all four histamine receptors (H1R–H4R) has been demonstrated in brain tissue, but the functional expression of H4Rs has not been sufficiently documented and is rather a result of the presence of incoming immune and hematopoietic cells (e.g., MCs, T cells, and macrophages) in the CNS [25,214]. Therefore, in neurodevelopmental and neurodegenerative diseases, which are inevitably accompanied by neuroinflammation of varying severity, the role of H4Rs may be particularly important [59,215]. Simply reducing neuroinflammation may reduce brain H4R expression in these cases and thus disrupt the vicious cycle in which H4Rs modulate eosinophil chemotaxis and the selective recruitment of MCs, resulting in the amplification of HA-mediated immune responses and ultimately leading to chronic inflammation [88,216].
Modulation of the effects of HA on other brain neurotransmitter systems (e.g., DA, ACh, 5-HT, NE, GABA, glutamate, and orexin) can also be achieved by affecting H1Rs and H2Rs located on neuronal somata and dendrites [11,217,218,219]. However, the fundamental problem with the therapeutic use of H1R and H2R agonists and antagonists is that their expression is not limited to the CNS, thus causing systemic side effects [29]. This problem practically does not occur with H3Rs, whose expression is restricted to the CNS. Moreover, H3R functions not only as an autoreceptor but also as an inhibitory heteroreceptor expressed within the axon varicosities, including glutamatergic and aminergic neurons [220]. This is central to the role of H3Rs in the CNS. As presynaptic autoreceptors, H3Rs inhibit the production and release of HA from TMN neurons, and acting as presynaptic heteroreceptors may also inhibit the release of other neurotransmitters (mentioned above). Interestingly, the vast majority of H3Rs are located in the postsynaptic space and on cells outside the neuronal histaminergic system. It is much more difficult to predict or interpret the effects of excitation/inhibition of postsynaptic H3Rs on the levels of HA and HA-related neurotransmitters, especially with respect to specific clinical aspects [106,221,222]. Therefore, despite the expression of all four known histamine receptors in the CNS, in recent years, only H3Rs have become the target of numerous therapeutic trials of potential novel drugs, including those developed for neurodevelopmental and neurodegenerative diseases [223,224,225].
This particular situation, when, on the basis of current knowledge about the participation of the histaminergic system and individual histamine receptors in the etiopathogenesis of brain diseases, H3R is almost exclusively selected as a direct therapeutic target and secondarily affects other histamine receptors important in a given disease, is reflected in Figure 4.
All histamine H1R–H4R receptors are involved to varying degrees in the regulation of many brain processes occurring in physiological and disease states, including neurodevelopmental disorders and neurodegenerative diseases. Taking into account the neuroinflammation that accompanies these diseases, H4R was included, the functional expression of which is not sufficiently confirmed in nervous tissue, but appears to be a derivative of the influx of immune and hematopoietic cells (e.g., MCs, T cells, macrophages) into the CNS. Due to its pre- and postsynaptic expression, H3R plays a key role in the histaminergic system, regulating the release of HA and, acting on the presynaptic heteroreceptors, modulates the release of several other neurotransmitters (e.g., 5-HT, dopamine, norepinephrine, ACh, GABA and glutamate). Furthermore, the presence of H3R confined to the CNS means that systemic symptoms that constitute side effects of H3R-acting therapy do not occur or are significantly reduced compared to direct effects on H1R and H2R. Therefore, in experimental therapeutic applications, the use of H3R antagonists/inverse agonists or agonists is seen as promising. Green arrows for neurodevelopmental disorders and black for neurodegenerative diseases indicate effects across the entire brain tissue, not a specific region.
AD – Alzheimer’s disease; ADHD – attention-deficit/hyperactivity disorder; ASD – autism spectrum disorder; HD – Huntington’s disease; ID – intellectual disability; MDD – major depressive disorder; NMD – neurodevelopmental motor disorders; NSD – Narcolepsy spectrum disorder; PD – Parkinson’s disease; Schiz – schizophrenia; D1-H3R – dopamine D1-histamine H3 receptor heteromer; ↓H1R, ↓H2R – decreased expression of H1R and H2R, respectively; ↑H3R – increased expression of H3R; ? – contradictory results
A detailed presentation of the research results on the use of numerous H3R-active substances, primarily antagonists/inverse agonists, in the context of their use in neurodevelopmental disorders and neurodegenerative diseases is beyond the scope of this review. This topic has been extensively discussed and summarized in recent years, as in the review papers of Thomas et al. [88], Ghamari et al. [226] and Szczepańska et al. [227]. Overall, the results appear to be divergent and inconclusive, ranging from promising to unsupportive of the effectiveness of a given treatment, reflecting the complexity and heterogeneity of the nervous system disorders in which these experimental therapies are used [59,228,229]. However, these studies are undoubtedly worth continuing while clarifying the indications for a specific method of treatment for a given neuropsychiatric disorder.
6. Concluding Remarks
The pleiotropic effects of HA are mediated by its binding to four types of membrane receptors, H1R–H4R, which are expressed in different cell types, including those occurring in the CNS. Apart from the developmental period and pathological conditions leading to disruption of the integrity of the BBB and the influx of immune and hematopoietic cells, the primary source of HA in brain tissue is histaminergic neurons located within the TMN, which send fibers to all major brain regions. The existence of the histaminergic network makes HA a conserved modulator in mammalian brains that is critically involved in many physiological functions through mutual interactions and the regulation of the activity of other neurotransmitters in the CNS, such as 5-HT, DA, NE, ACh, GABA and glutamate. The direct or indirect involvement of histaminergic neurons in the regulation of basic homeostatic functions of the body, as well as arousal, circadian and feeding rhythms (appetite), behavioral tasks and higher nervous activities such as cognition, inference, prediction, memory, association, consideration, speech production and understanding, and spatial orientation, has been proven.
H3Rs play a special role in modulating the activity of the central histaminergic system and other neurotransmitter systems associated with it, which results from their pre- and postsynaptic expression and autoreceptor and heteroreceptor functions. By influencing HA production and release, H3R signaling simultaneously affects H1R and H2R transmission and, in the case of neuroinflammation, H4R. Variable neuroinflammation, often accompanied by local changes in HA concentrations in brain tissue, is a constant symptom of neurodevelopmental and neurodegenerative diseases. Regardless of the etiopathogenesis of these nervous system disorders, which are usually not fully understood, there are constant attempts to improve the clinical condition by influencing HA receptors.
With few exceptions, H3R in recent years has almost exclusively become the target of numerous therapeutic trials for potentially novel drugs, including those developed for neurodevelopmental and neurodegenerative diseases. However, the results of many therapeutic concepts targeting the brain's histaminergic system have proven uncertain or controversial, as the translation of the results of functional and behavioral studies conducted on animals to human conditions is fraught with uncertainty. Moreover, in clinical practice, it is extremely rare to encounter individual neurological diseases in isolated ("pure") form without overlapping comorbidities. The previously mentioned dual role of HA as a neurotransmitter and mediator of the inflammatory reaction, the final neurodestructive or neuroprotective effect of which depends on the recruitment of a specific cytokine profile, also makes it difficult to clearly interpret and determine the effectiveness of a given therapy. The heterogeneity of both individual subpopulations of neurons in the TMN and the histamine receptors themselves within histaminergic neurons should also be taken into account.
The above difficulties may motivate further research aimed at developing new, detailed diagnostic methods for neurodevelopmental and neurodegenerative diseases, after which it will be possible to verify/clarify therapeutic goals related to the influence on the brain’s histaminergic system.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created. Instead, the data are quoted from the available cited literature.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
- 5-HT – serotonin
- 5-HTRs – serotonin receptors
- aa – amino acids
- AA – arachidonic acid
- AADE – arachidonic acid-derived eicosanoids
- AC – adenylyl cyclase
- ACh – acetylcholine
- AD – Alzheimer’s disease
- ADHD – attention-deficit/hyperactivity disorder
- AP-1 – activator protein 1
- APCs – antigen-presenting cells
- ARRB2 – arrestin beta-2, also known as beta-arrestin-2
- ASD – autism spectrum disorders
- BBB – blood-brain barrier
- bFGF – basic fibroblast growth factor
- cAMP – cyclic 3’,5’ adenosine monophosphate
- CBM – CBM signalosome complex = CARD11 (the caspase recruitment domain family member 11)—BCL10 (B cell CLL/lymphoma 10)—MALT1 (mucosa associated lymphoid tissue lymphoma translocation protein 1) paracaspase,
- c-fos – protooncogene (human homolog of the retroviral oncogene v-fos)
- cGMP – cyclic guanosine monophosphate
- CNS – central nervous system
- CREB – cAMP-responsive element-binding protein
- CSF – cerebrospinal fluid
- DA – dopamine
- DAG – diacylglycerol
- DAO – diamine oxidase
- DCs – dendritic cells
- ECL cells – enterochromaffin-like cells
- ECs – endothelial cells
- eNOS – endothelial nitric oxide synthase
- ERS – endoplasmic reticulum stress
- GABA – γ-amino butyric acid
- GDP – guanosine diphosphate
- GLU – glutamate
- GTP – guanosine triphosphate
- GPCRs – G protein-coupled receptors, also known as the seven-(pass)-transmembrane (7TM) domain receptors
- H1R, H2R, H3R, H4R – four histamine receptors, respectively
- HA – histamine
- HD – Huntington's disease
- HDC – enzyme L-histidine decarboxylase
- HMT – histamine N-methyltransferase, also known as HNMT
- IAA – imidazole acetic acid
- IAA-RP – imidazole acetic acid ribotide
- ID – intellectual disability
- IFG – inferior frontal gyrus
- IFN-γ – interferon gamma
- IL-1β, IL-10, IL-12, IL-33 – interleukins: 1 beta, 10, 12 and 33, respectively
- IP3 – inositol 1,4,5-trisphosphate
- KD – dissociation constant
- LBs – Lewy bodies
- MAO-B – monoamine oxidase B
- MAPK – mitogen-activated protein kinase
- MCH – melanin-concentrating hormone
- MCs – mast cells
- MDD – major depressive disorder
- NA – norepinephrine
- NMD – neurodevelopmental motor disorders
- NSCs – neural stem cells
- NSD – narcolepsy spectrum disorder
- NF-κβ – nuclear factor kappa-light-chain-enhancer of activated B cells
- NK cells – natural killer cells
- NMDA – N-methyl-D-aspartate
- NO – nitric oxide
- NREM – non-rapid eye movement phase of sleep
- Nτ-MH – tele-methylhistamine
- OCPD – obsessive-compulsive personality disorder
- OCT3 – organic cation transporter 3
- PD – Parkinson’s disease
- PFC – prefrontal cortex
- PIP2 –phosphatidylinositol 4,5-bisphosphate
- PKA – protein kinase A
- PKC – protein kinase C
- PLA2 – phospholipase A2
- PLC – phospholipase C
- PLD – phospholipase D
- PMAT – plasma membrane monoamine transporter
- REM – rapid eye movement phase of sleep
- ROS – reactive oxygen species
- Schiz – schizophrenia
- SDAT – senile dementia of the Alzheimer's disease type
- SNP – single nucleotide polymorphism
- TGFβ – transforming growth factor beta
- Th1, Th2 – Type 1 T helper and type 2 T helper cells, respectively
- t-MIAA – tele-methylimidazole acetic acid
- TMN – tuberomammillary nucleus
- TNFα – tumor necrosis factor alpha
- TRAF6 – tumor necrosis factor (TNF) receptor-associated factor 6
- VEGF – vascular endothelial growth factor
- VMAT2 – vesicular monoamine transporter
- VSMCs – vascular smooth muscle cells
- XO – xanthine oxidase
References
- Tiligada E, Ennis M. Histamine pharmacology: from Sir Henry Dale to the 21st century. Br J Pharmacol. 2020, 177, 469–489. [Google Scholar] [CrossRef] [PubMed]
- Church, MK. Allergy, Histamine and Antihistamines. Handb Exp Pharmacol. 2017, 241, 321–331. [Google Scholar] [CrossRef]
- Ashina K, Tsubosaka Y, Nakamura T, Omori K, Kobayashi K, Hori M, Ozaki H, Murata T. Histamine Induces Vascular Hyperpermeability by Increasing Blood Flow and Endothelial Barrier Disruption In Vivo. PLoS One. 2015, 10, e0132367. [Google Scholar] [CrossRef]
- Mikelis CM, Simaan M, Ando K, Fukuhara S, Sakurai A, Amornphimoltham P, Masedunskas A, Weigert R, Chavakis T, Adams RH, Offermanns S, Mochizuki N, Zheng Y, Gutkind JS. RhoA and ROCK mediate histamine-induced vascular leakage and anaphylactic shock. Nat Commun. 2015, 6, 6725. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik R, Shaharyar MA, Sarkar A, Mandal A, Anand K, Shabana H, Mitra A, Karmakar S. Immunopathogenesis of urticaria: a clinical perspective on histamine and cytokine involvement. Inflamm Res. 2024, 73, 877–896. [Google Scholar] [CrossRef] [PubMed]
- Sörbo J, Jakobsson A, Norrby K. Mast-cell histamine is angiogenic through receptors for histamine1 and histamine2. Int J Exp Pathol. 1994, 75, 43–50. [Google Scholar]
- Szewczyk G, Pyzlak M, Klimkiewicz J, Smiertka W, Miedzińska-Maciejewska M, Szukiewicz D. Mast cells and histamine: do they influence placental vascular network and development in preeclampsia? Mediators Inflamm. 2012, 2012, 307189. [Google Scholar] [CrossRef]
- Chen J, Liu G, Wang X, Hong H, Li T, Li L, Wang H, Xie J, Li B, Li T, Lu D, Zhang Y, Zhao H, Yao C, Wen K, Li T, Chen J, Wu S, He K, Zhang WN, Zhao J, Wang N, Han Q, Xia Q, Qi J, Chen J, Zhou T, Man J, Zhang XM, Li AL, Pan X. Glioblastoma stem cell-specific histamine secretion drives pro-angiogenic tumor microenvironment remodeling. Cell Stem Cell. 2022, 29, 1531–1546. [Google Scholar] [CrossRef]
- Smolinska S, Winiarska E, Globinska A, Jutel M. Histamine: A Mediator of Intestinal Disorders-A Review. Metabolites 2022, 12, 895. [Google Scholar] [CrossRef]
- Hough LB. Histamine Actions in the Central Nervous System. In: Siegel GJ, Agranoff BW, Albers RW, et al., editors. Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 6th edition. Philadelphia: Lippincott-Raven; 1999. Available from: https://www.ncbi.nlm.nih.gov/books/NBK28245/.
- Scammell TE, Jackson AC, Franks NP, Wisden W, Dauvilliers Y. Histamine: neural circuits and new medications. Sleep 2019, 42, zsy183. [Google Scholar] [CrossRef]
- Arrigoni E, Fuller PM. The Role of the Central Histaminergic System in Behavioral State Control. Curr Top Behav Neurosci. 2022, 59, 447–468. [Google Scholar] [CrossRef]
- Hough LB. Histaminergic Cells of the Central Nervous System: Anatomy and Morphology. In: Siegel GJ, Agranoff BW, Albers RW, et al., editors. Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 6th edition. Philadelphia: Lippincott-Raven; 1999. Available from: https://www.ncbi.nlm.nih.gov/books/NBK28098/.
- Blandina P, Munari L, Provensi G, Passani MB. Histamine neurons in the tuberomamillary nucleus: a whole center or distinct subpopulations? Front Syst Neurosci. 2012, 6, 33. [Google Scholar] [CrossRef]
- Yoshikawa T, Nakamura T, Yanai K. Histaminergic neurons in the tuberomammillary nucleus as a control centre for wakefulness. Br J Pharmacol. 2021, 178, 750–769. [Google Scholar] [CrossRef] [PubMed]
- Hough LB. Dynamics of Histamine in the Brain. In: Siegel GJ, Agranoff BW, Albers RW, et al., editors. Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 6th edition. Philadelphia: Lippincott-Raven; 1999. Available from: https://www.ncbi.nlm.nih.gov/books/NBK27916/.
- Maršavelski A, Mavri J, Vianello R, Stare J. Why Monoamine Oxidase B Preferably Metabolizes N-Methylhistamine over Histamine: Evidence from the Multiscale Simulation of the Rate-Limiting Step. Int J Mol Sci. 2022, 23, 1910. [Google Scholar] [CrossRef]
- Thangam EB, Jemima EA, Singh H, Baig MS, Khan M, Mathias CB, Church MK, Saluja R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed]
- Parsons ME, Ganellin CR. Histamine and its receptors. Br J Pharmacol. 2006, 147 (Suppl 1), S127–35. [Google Scholar] [CrossRef]
- Weis WI, Kobilka BK. The Molecular Basis of G Protein-Coupled Receptor Activation. Annu Rev Biochem. 2018, 87, 897–919. [Google Scholar] [CrossRef] [PubMed]
- Kamato D, Thach L, Bernard R, Chan V, Zheng W, Kaur H, Brimble M, Osman N, Little PJ. Structure, Function, Pharmacology, and Therapeutic Potential of the G Protein, Gα/q,11. Front Cardiovasc Med. 2015, 2, 14. [Google Scholar] [CrossRef]
- Pfleger J, Gresham K, Koch WJ. G protein-coupled receptor kinases as therapeutic targets in the heart. Nat Rev Cardiol. 2019, 16, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Khouma A, Moeini MM, Plamondon J, Richard D, Caron A, Michael NJ. Histaminergic regulation of food intake. Front Endocrinol (Lausanne). 2023, 14, 1202089. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Mir MI, Pollard H, Moreau J, Arrang JM, Ruat M, Traiffort E, Schwartz JC, Palacios JM. Three histamine receptors (H1, H2 and H3) visualized in the brain of human and non-human primates. Brain Res. 1990, 526, 322–7. [Google Scholar] [CrossRef]
- Connelly WM, Shenton FC, Lethbridge N, Leurs R, Waldvogel HJ, Faull RL, Lees G, Chazot PL. The histamine H4 receptor is functionally expressed on neurons in the mammalian CNS. Br J Pharmacol. 2009, 157, 55–63. [Google Scholar] [CrossRef]
- Galeotti N, Sanna MD, Ghelardini C. Pleiotropic effect of histamine H4 receptor modulation in the central nervous system. Neuropharmacology 2013, 71, 141–7. [Google Scholar] [CrossRef] [PubMed]
- Schneider EH, Seifert R. The histamine H4-receptor and the central and peripheral nervous system: A critical analysis of the literature. Neuropharmacology. 2016, 106, 116–28. [Google Scholar] [CrossRef] [PubMed]
- Sanna MD, Ghelardini C, Thurmond RL, Masini E, Galeotti N. Behavioural phenotype of histamine H4 receptor knockout mice: Focus on central neuronal functions. Neuropharmacology. 2017, 114, 48–57. [Google Scholar] [CrossRef]
- Panula, P. Histamine receptors, agonists, and antagonists in health and disease. Handb Clin Neurol. 2021, 180, 377–387. [Google Scholar] [CrossRef]
- Panula P, Chazot PL, Cowart M, Gutzmer R, Leurs R, Liu WL, Stark H, Thurmond RL, Haas HL. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol Rev. 2015, 67, 601–55. [Google Scholar] [CrossRef]
- De Backer MD, Loonen I, Verhasselt P, Neefs JM, Luyten WH. Structure of the human histamine H1 receptor gene. Biochem J. 1998, 335 (Pt 3), 663–70. [Google Scholar] [CrossRef]
- Smit MJ, Hoffmann M, Timmerman H, Leurs R. Molecular properties and signalling pathways of the histamine H1 receptor. Clin Exp Allergy. 1999, 29, 19–28. [Google Scholar] [CrossRef]
- Seifert R, Strasser A, Schneider EH, Neumann D, Dove S, Buschauer A. Molecular and cellular analysis of human histamine receptor subtypes. Trends Pharmacol Sci. 2013, 34, 33–58. [Google Scholar] [CrossRef]
- Conrad M, Söldner CA, Miao Y, Sticht H. Agonist Binding and G Protein Coupling in Histamine H2 Receptor: A Molecular Dynamics Study. Int J Mol Sci. 2020, 21, 6693. [Google Scholar] [CrossRef]
- Ishola AA, Joshi T, Abdulai SI, Tijjani H, Pundir H, Chandra S. Molecular basis for the repurposing of histamine H2-receptor antagonist to treat COVID-19. J Biomol Struct Dyn. 2022, 40, 5785–5802. [Google Scholar] [CrossRef] [PubMed]
- Höring C, Conrad M, Söldner CA, Wang J, Sticht H, Strasser A, Miao Y. Specific Engineered G Protein Coupling to Histamine Receptors Revealed from Cellular Assay Experiments and Accelerated Molecular Dynamics Simulations. Int J Mol Sci. 2021, 22, 10047. [Google Scholar] [CrossRef]
- Wiedemann P, Bönisch H, Oerters F, Brüss M. Structure of the human histamine H3 receptor gene (HRH3) and identification of naturally occurring variations. J Neural Transm (Vienna). 2002, 109, 443–53. [Google Scholar] [CrossRef] [PubMed]
- Lai X, Ye L, Liao Y, Jin L, Ma Q, Lu B, Sun Y, Shi Y, Zhou N. Agonist-induced activation of histamine H3 receptor signals to extracellular signal-regulated kinases 1 and 2 through PKC-, PLD-, and EGFR-dependent mechanisms. J Neurochem. 2016, 137, 200–15. [Google Scholar] [CrossRef] [PubMed]
- Rahman SN, McNaught-Flores DA, Huppelschoten Y, da Costa Pereira D, Christopoulos A, Leurs R, Langmead CJ. Structural and Molecular Determinants for Isoform Bias at Human Histamine H3 Receptor Isoforms. ACS Chem Neurosci. 2023, 14, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Im D, Kishikawa JI, Shiimura Y, Hisano H, Ito A, Fujita-Fujiharu Y, Sugita Y, Noda T, Kato T, Asada H, Iwata S. Structural insights into the agonists binding and receptor selectivity of human histamine H4 receptor. Nat Commun. 2023, 14, 6538. [Google Scholar] [CrossRef] [PubMed]
- Nijmeijer S, de Graaf C, Leurs R, Vischer HF. Molecular pharmacology of histamine H4 receptors. Front Biosci (Landmark Ed). 2012, 17, 2089–106. [Google Scholar] [CrossRef] [PubMed]
- Xia R, Shi S, Xu Z, Vischer HF, Windhorst AD, Qian Y, Duan Y, Liang J, Chen K, Zhang A, Guo C, Leurs R, He Y. Structural basis of ligand recognition and design of antihistamines targeting histamine H4 receptor. Nat Commun. 2024, 15, 2493. [Google Scholar] [CrossRef] [PubMed]
- Mehta P, Miszta P, Filipek S. Molecular Modeling of Histamine Receptors-Recent Advances in Drug Discovery. Molecules. 2021, 26, 1778. [Google Scholar] [CrossRef]
- Bongers G, de Esch I, Leurs R. Molecular pharmacology of the four histamine receptors. Adv Exp Med Biol. 2010, 709, 11–9. [Google Scholar] [CrossRef]
- Gao M, Dekker ME, Leurs R, Vischer HF. Pharmacological characterization of seven human histamine H3 receptor isoforms. Eur J Pharmacol. 2024, 968, 176450. [Google Scholar] [CrossRef] [PubMed]
- Mocking T, Bosma R, Rahman SN, Verweij EWE, McNaught-Flores DA, Vischer HF and Leurs R. Molecular Aspects of Histamine Receptors. In: Blandina P, Passani M. (eds) Histamine Receptors. The Receptors, 2016, vol 28. Humana, Cham. [CrossRef]
- Beyer L, Kabatas AS, Mommert S, Stark H, Werfel T, Gutzmer R, Schaper-Gerhardt K. Histamine Activates Human Eosinophils via H2R and H4R Predominantly in Atopic Dermatitis Patients. Int J Mol Sci. 2022, 23, 10294. [Google Scholar] [CrossRef]
- Hatipoglu OF, Nishinaka T, Nishibori M, Watanabe M, Toyomura T, Mori S, Yaykasli KO, Wake H, Takahashi H. Histamine promotes angiogenesis through a histamine H1 receptor-PKC-VEGF-mediated pathway in human endothelial cells. J Pharmacol Sci. 2023, 151, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Moya-García AA, Pino-Ángeles A, Sánchez-Jiménez F, Urdiales JL, Medina MÁ. Histamine, Metabolic Remodelling and Angiogenesis: A Systems Level Approach. Biomolecules 2021, 11, 415. [Google Scholar] [CrossRef]
- Um MY, Yoon M, Kim M, Jung J, Kim S, Kim DO, Cho S. Curcuminoids, a major turmeric component, have a sleep-enhancing effect by targeting the histamine H1 receptor. Food Funct. 2022, 13, 12697–12706. [Google Scholar] [CrossRef]
- Yuan H, Silberstein SD. Histamine and Migraine. Headache. 2018, 58, 184–193. [Google Scholar] [CrossRef]
- Szukiewicz D, Pyzlak M, Stangret A, Rongies W, Maslinska D. Decrease in expression of histamine H2 receptors by human amniotic epithelial cells during differentiation into pancreatic beta-like cells. Inflamm Res. 2010, 59 Suppl 2, S205–7. [Google Scholar] [CrossRef]
- Szukiewicz D, Szewczyk G, Mittal TK, Rongies W, Maslinski S. Involvement of histamine and histamine H2 receptors in nicotinamide-induced differentiation of human amniotic epithelial cells into insulin-producing cells. Inflamm Res. 2010, 59 Suppl 2, S209–11. [CrossRef]
- Ichikawa T, Hotta K, Ishihara K. Second-generation histamine H(2)-receptor antagonists with gastric mucosal defensive properties. Mini Rev Med Chem. 2009, 9, 581–9. [Google Scholar] [CrossRef]
- Arumuham A, Nour MM, Veronese M, Onwordi EC, Rabiner EA, Howes OD. The histamine system and cognitive function: An in vivo H3 receptor PET imaging study in healthy volunteers and patients with schizophrenia. J Psychopharmacol. 2023, 37, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Honkisz-Orzechowska E, Popiołek-Barczyk K, Linart Z, Filipek-Gorzała J, Rudnicka A, Siwek A, Werner T, Stark H, Chwastek J, Starowicz K, Kieć-Kononowicz K, Łażewska D. Anti-inflammatory effects of new human histamine H3 receptor ligands with flavonoid structure on BV-2 neuroinflammation. Inflamm Res. 2023, 72, 181–194. [Google Scholar] [CrossRef]
- Cheng L, Liu J, Chen Z. The Histaminergic System in Neuropsychiatric Disorders. Biomolecules 2021, 11, 1345. [Google Scholar] [CrossRef]
- Zhang XY, Peng SY, Shen LP, Zhuang QX, Li B, Xie ST, Li QX, Shi MR, Ma TY, Zhang Q, Wang JJ, Zhu JN. Targeting presynaptic H3 heteroreceptor in nucleus accumbens to improve anxiety and obsessive-compulsive-like behaviors. Proc Natl Acad Sci U S A. 2020, 117, 32155–32164. [Google Scholar] [CrossRef]
- Zhou Z, An Q, Zhang W, Li Y, Zhang Q, Yan H. Histamine and receptors in neuroinflammation: Their roles on neurodegenerative diseases. Behav Brain Res. 2024, 465, 114964. [Google Scholar] [CrossRef]
- Grzybowska-Kowalczyk A, Maslinska D, Wojciechowska M, Szukiewicz D, Wojtecka-Lukasik E, Paradowska A, Maldyk P, Maslinski S. Expression of histamine H4 receptor in human osteoarthritic synovial tissue. Inflamm Res. 2008, 57 Suppl 1, S63–4. [CrossRef]
- Schirmer B, Neumann D. The Function of the Histamine H4 Receptor in Inflammatory and Inflammation-Associated Diseases of the Gut. Int J Mol Sci. 2021, 22, 6116. [Google Scholar] [CrossRef]
- Silver R, Silverman AJ, Vitković L, Lederhendler II. Mast cells in the brain: evidence and functional significance. Trends Neurosci. 1996, 19, 25–31. [Google Scholar] [CrossRef]
- Cacabelos R, Yamatodani A, Niigawa H, Hariguchi S, Tada K, Nishimura T, Wada H, Brandeis L, Pearson J. Brain histamine in Alzheimer's disease. Methods Find Exp Clin Pharmacol. 1989, 11, 353–60. [Google Scholar]
- Passani MB, Panula P, Lin JS. Histamine in the brain. Front Syst Neurosci. 2014, 8, 64. [Google Scholar] [CrossRef]
- Haas HL, Sergeeva OA, Selbach O. Histamine in the nervous system. Physiol Rev. 2008, 88, 1183–241. [Google Scholar] [CrossRef] [PubMed]
- Carthy E, Ellender T. Histamine, Neuroinflammation and Neurodevelopment: A Review. Front Neurosci. 2021, 15, 680214. [Google Scholar] [CrossRef] [PubMed]
- Bernardino, L. Histamine in the Crosstalk Between Innate Immune Cells and Neurons: Relevance for Brain Homeostasis and Disease. Curr Top Behav Neurosci. 2022, 59, 261–288. [Google Scholar] [CrossRef] [PubMed]
- Han S, Márquez-Gómez R, Woodman M, Ellender T. Histaminergic Control of Corticostriatal Synaptic Plasticity during Early Postnatal Development. J Neurosci. 2020, 40, 6557–6571. [Google Scholar] [CrossRef] [PubMed]
- Nuutinen S, Panula P. Histamine in neurotransmission and brain diseases. Adv Exp Med Biol. 2010, 709, 95–107. [Google Scholar] [CrossRef]
- Panula P, Sundvik M, Karlstedt K. Developmental roles of brain histamine. Trends Neurosci. 2014, 37, 159–68. [Google Scholar] [CrossRef]
- Rodríguez-Martínez G, Velasco I, García-López G, Solís KH, Flores-Herrera H, Díaz NF, Molina-Hernández A. Histamine is required during neural stem cell proliferation to increase neuron differentiation. Neuroscience. 2012, 216, 10–7. [Google Scholar] [CrossRef] [PubMed]
- Lamanna J, Ferro M, Spadini S, Racchetti G, Malgaroli A. The Dysfunctional Mechanisms Throwing Tics: Structural and Functional Changes in Tourette Syndrome. Behav Sci (Basel). 2023, 13, 668. [Google Scholar] [CrossRef]
- Calzà J, Gürsel DA, Schmitz-Koep B, Bremer B, Reinholz L, Berberich G, Koch K. Altered Cortico-Striatal Functional Connectivity During Resting State in Obsessive-Compulsive Disorder. Front Psychiatry. 2019, 10, 319. [Google Scholar] [CrossRef]
- Mazurkiewicz-Kwilecki IM, Nsonwah S. Changes in the regional brain histamine and histidine levels in postmortem brains of Alzheimer patients. Can J Physiol Pharmacol. 1989, 67, 75–8. [Google Scholar] [CrossRef]
- Prell GD, Green JP, Kaufmann CA, Khandelwal JK, Morrishow AM, Kirch DG, Linnoila M, Wyatt RJ. Histamine metabolites in cerebrospinal fluid of patients with chronic schizophrenia: their relationships to levels of other aminergic transmitters and ratings of symptoms. Schizophr Res. 1995, 14, 93–104. [Google Scholar] [CrossRef]
- Bravo TP, Matchett GA, Jadhav V, Martin RD, Jourdain A, Colohan A, Zhang JH, Tang J. Role of histamine in brain protection in surgical brain injury in mice. Brain Res. 2008, 1205, 100–7. [Google Scholar] [CrossRef] [PubMed]
- Adachi, N. Cerebral ischemia and brain histamine. Brain Res Brain Res Rev. 2005, 50, 275–86. [Google Scholar] [CrossRef] [PubMed]
- Obara I, Telezhkin V, Alrashdi I, Chazot PL. Histamine, histamine receptors, and neuropathic pain relief. Br J Pharmacol. 2020, 177, 580–599. [CrossRef]
- Fujita A, Bonnavion P, Wilson MH, Mickelsen LE, Bloit J, de Lecea L, Jackson AC. Hypothalamic Tuberomammillary Nucleus Neurons: Electrophysiological Diversity and Essential Role in Arousal Stability. J Neurosci. 2017, 37, 9574–9592. [Google Scholar] [CrossRef]
- Haas H, Panula P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat Rev Neurosci. 2003, 4, 121–30. [Google Scholar] [CrossRef] [PubMed]
- Valko PO, Gavrilov YV, Yamamoto M, Reddy H, Haybaeck J, Mignot E, Baumann CR, Scammell TE. Increase of histaminergic tuberomammillary neurons in narcolepsy. Ann Neurol. 2013, 74, 794–804. [Google Scholar] [CrossRef] [PubMed]
- John J, Thannickal TC, McGregor R, Ramanathan L, Ohtsu H, Nishino S, Sakai N, Yamanaka A, Stone C, Cornford M, Siegel JM. Greatly increased numbers of histamine cells in human narcolepsy with cataplexy. Ann Neurol. 2013, 74, 786–93. [Google Scholar] [CrossRef] [PubMed]
- Airaksinen MS, Paetau A, Paljärvi L, Reinikainen K, Riekkinen P, Suomalainen R, Panula P. Histamine neurons in human hypothalamus: anatomy in normal and Alzheimer diseased brains. Neuroscience 1991, 44, 465–81. [Google Scholar] [CrossRef] [PubMed]
- Lin W, Xu L, Zheng Y, An S, Zhao M, Hu W, Li M, Dong H, Li A, Li Y, Gong H, Pan G, Wang Y, Luo Q, Chen Z. Whole-brain mapping of histaminergic projections in mouse brain. Proc Natl Acad Sci U S A. 2023, 120, e2216231120. [Google Scholar] [CrossRef]
- Satpati A, Neylan T, Grinberg LT. Histaminergic neurotransmission in aging and Alzheimer's disease: A review of therapeutic opportunities and gaps. Alzheimers Dement (N Y). 2023, 9, e12379. [Google Scholar] [CrossRef] [PubMed]
- Moriwaki C, Chiba S, Wei H, Aosa T, Kitamura H, Ina K, Shibata H, Fujikura Y. Distribution of histaminergic neuronal cluster in the rat and mouse hypothalamus. J Chem Neuroanat. 2015, 68, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Venner A, Mochizuki T, De Luca R, Anaclet C, Scammell TE, Saper CB, Arrigoni E, Fuller PM. Reassessing the Role of Histaminergic Tuberomammillary Neurons in Arousal Control. J Neurosci. 2019, 39, 8929–8939. [Google Scholar] [CrossRef] [PubMed]
- Thomas SD, Abdalla S, Eissa N, Akour A, Jha NK, Ojha S, Sadek B. Targeting Microglia in Neuroinflammation: H3 Receptor Antagonists as a Novel Therapeutic Approach for Alzheimer’s Disease, Parkinson’s Disease, and Autism Spectrum Disorder. Pharmaceuticals 2024, 17, 831. [Google Scholar] [CrossRef] [PubMed]
- Molderings GJ, Weissenborn G, Schlicker E, Likungu J, Göthert M. Inhibition of noradrenaline release from the sympathetic nerves of the human saphenous vein by presynaptic histamine H3 receptors. Naunyn Schmiedebergs Arch Pharmacol. 1992, 346, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Brown RE, Haas HL. On the mechanism of histaminergic inhibition of glutamate release in the rat dentate gyrus. J Physiol. 1999, 515 (Pt 3), 777–86. [CrossRef] [PubMed]
- Haas HL, Panula P. Histamine receptors. Neuropharmacology. 2016, 106, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Zhu J, Qu C, Lu X, Zhang S. Activation of microglia by histamine and substance P. Cell Physiol Biochem. 2014, 34, 768–80. [Google Scholar] [CrossRef] [PubMed]
- Mehta P, Miszta P, Rzodkiewicz P, Michalak O, Krzeczyński P, Filipek S. Enigmatic Histamine Receptor H4 for Potential Treatment of Multiple Inflammatory, Autoimmune, and Related Diseases. Life 2020, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- John J, Wu MF, Boehmer LN, Siegel JM. Cataplexy-active neurons in the hypothalamus: implications for the role of histamine in sleep and waking behavior. Neuron. 2004, 42, 619–34. [Google Scholar] [CrossRef] [PubMed]
- Provensi G, Costa A, Izquierdo I, Blandina P, Passani MB. Brain histamine modulates recognition memory: possible implications in major cognitive disorders. Br J Pharmacol. 2020, 177, 539–556. [Google Scholar] [CrossRef]
- Nomura H, Shimizume R, Ikegaya Y. Histamine: A Key Neuromodulator of Memory Consolidation and Retrieval. Curr Top Behav Neurosci. 2022, 59, 329–353. [Google Scholar] [CrossRef]
- Costa A, Ducourneau E, Curti L, Masi A, Mannaioni G, Hardt L, Biyong EF, Potier M, Blandina P, Trifilieff P, Provensi G, Ferreira G, Passani MB. Chemogenetic activation or inhibition of histaminergic neurons bidirectionally modulates recognition memory formation and retrieval in male and female mice. Sci Rep. 2024, 14, 11283. [Google Scholar] [CrossRef] [PubMed]
- Tabarean, IV. Histamine receptor signaling in energy homeostasis. Neuropharmacology. 2016, 106, 13–9. [Google Scholar] [CrossRef]
- Sakata T, Yoshimatsu H, Kurokawa M. Hypothalamic neuronal histamine: implications of its homeostatic control of energy metabolism. Nutrition. 1997, 13, 403–11. [Google Scholar] [CrossRef]
- Provensi G, Blandina P, Passani MB. The histaminergic system as a target for the prevention of obesity and metabolic syndrome. Neuropharmacology. 2016, 106, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Xu L, Lin W, Zheng Y, Wang Y, Chen Z. The Diverse Network of Brain Histamine in Feeding: Dissect its Functions in a Circuit-Specific Way. Curr Neuropharmacol. 2024, 22, 241–259. [Google Scholar] [CrossRef]
- Michael NJ, Zigman JM, Williams KW, Elmquist JK. Electrophysiological Properties of Genetically Identified Histaminergic Neurons. Neuroscience. 2020, 444, 183–195. [Google Scholar] [CrossRef]
- Kuno M. Metabotropic receptors mediate slow synaptic responses. In: The Synapse: Function, Plasticity, and Neurotrophism, Oxford, 1994; online edn, Oxford Academic, 22 Mar. 2012), pp. 68–82. [CrossRef]
- Hatton GI, Yang QZ. Synaptically released histamine increases dye coupling among vasopressinergic neurons of the supraoptic nucleus: mediation by H1 receptors and cyclic nucleotides. J Neurosci. 1996, 16, 123–9. [Google Scholar] [CrossRef]
- Berger SN, Baumberger B, Samaranayake S, Hersey M, Mena S, Bain I, Duncan W, Reed MC, Nijhout HF, Best J, Hashemi P. An In Vivo Definition of Brain Histamine Dynamics Reveals Critical Neuromodulatory Roles for This Elusive Messenger. Int J Mol Sci. 2022, 23, 14862. [Google Scholar] [CrossRef]
- Zheng Y, Fan L, Fang Z, Liu Z, Chen J, Zhang X, Wang Y, Zhang Y, Jiang L, Chen Z, Hu W. Postsynaptic histamine H3 receptors in ventral basal forebrain cholinergic neurons modulate contextual fear memory. Cell Rep. 2023, 42, 113073. [Google Scholar] [CrossRef] [PubMed]
- Flik G, Folgering JH, Cremers TI, Westerink BH, Dremencov E. Interaction Between Brain Histamine and Serotonin, Norepinephrine, and Dopamine Systems: In Vivo Microdialysis and Electrophysiology Study. J Mol Neurosci. 2015, 56, 320–8. [Google Scholar] [CrossRef]
- Wada H, Inagaki N, Yamatodani A, Watanabe T. Is the histaminergic neuron system a regulatory center for whole-brain activity? Trends Neurosci. 1991, 14, 415–8. [Google Scholar] [CrossRef] [PubMed]
- Nimgampalle M, Chakravarthy H, Sharma S, Shree S, Bhat AR, Pradeepkiran JA, Devanathan V. Neurotransmitter systems in the etiology of major neurological disorders: Emerging insights and therapeutic implications. Ageing Res Rev. 2023, 89, 101994. [Google Scholar] [CrossRef]
- Saad AK, Akour A, Mahboob A, AbuRuz S, Sadek B. Role of Brain Modulators in Neurodevelopment: Focus on Autism Spectrum Disorder and Associated Comorbidities. Pharmaceuticals (Basel). 2022, 15, 612. [Google Scholar] [CrossRef]
- Davis SE, Cirincione AB, Jimenez-Torres AC, Zhu J. The Impact of Neurotransmitters on the Neurobiology of Neurodegenerative Diseases. Int J Mol Sci. 2023, 24, 15340. [Google Scholar] [CrossRef]
- Teleanu RI, Niculescu AG, Roza E, Vladâcenco O, Grumezescu AM, Teleanu DM. Neurotransmitters-Key Factors in Neurological and Neurodegenerative Disorders of the Central Nervous System. Int J Mol Sci. 2022, 23, 5954. [Google Scholar] [CrossRef]
- Grice SJ, Liu JL. Neurodevelopmental defects as a primer of neurodegeneration: lessons from spinal muscular atrophy and Huntington's disease. Neural Regen Res. 2023, 18, 1952–1953. [Google Scholar] [CrossRef]
- Hickman RA, O'Shea SA, Mehler MF, Chung WK. Neurogenetic disorders across the lifespan: from aberrant development to degeneration. Nat Rev Neurol. 2022, 18, 117–124. [Google Scholar] [CrossRef]
- Schor NF, Bianchi DW. Neurodevelopmental Clues to Neurodegeneration. Pediatr Neurol. 2021, 123, 67–76. [Google Scholar] [CrossRef]
- Fares J, Bou Diab Z, Nabha S, Fares Y. Neurogenesis in the adult hippocampus: history, regulation, and prospective roles. Int J Neurosci. 2019, 129, 598–611. [Google Scholar] [CrossRef] [PubMed]
- Abbott LC, Nigussie F. Adult neurogenesis in the mammalian dentate gyrus. Anat Histol Embryol. 2020, 49, 3–16. [Google Scholar] [CrossRef]
- Saraiva C, Barata-Antunes S, Santos T, Ferreiro E, Cristóvão AC, Serra-Almeida C, Ferreira R, Bernardino L. Histamine modulates hippocampal inflammation and neurogenesis in adult mice. Sci Rep. 2019, 9, 8384. [Google Scholar] [CrossRef]
- Sailaja K, Gopinath G. Ultrastructure of developing substantia nigra in humans. Int J Dev Neurosci. 1996, 14, 761–70. [Google Scholar] [CrossRef]
- Mottahedin A, Ardalan M, Chumak T, Riebe I, Ek J, Mallard C. Effect of Neuroinflammation on Synaptic Organization and Function in the Developing Brain: Implications for Neurodevelopmental and Neurodegenerative Disorders. Front Cell Neurosci. 2017, 11, 190. [Google Scholar] [CrossRef] [PubMed]
- Vanhala A, Yamatodani A, Panula P. Distribution of histamine-, 5-hydroxytryptamine-, and tyrosine hydroxylase-immunoreactive neurons and nerve fibers in developing rat brain. J Comp Neurol. 1994, 347, 101–14. [Google Scholar] [CrossRef]
- Auvinen S, Panula P. Development of histamine-immunoreactive neurons in the rat brain. J Comp Neurol. 1988, 276, 289–303. [Google Scholar] [CrossRef]
- Molina-Hernández A, Díaz NF, Arias-Montaño JA. Histamine in brain development. J Neurochem. 2012, 122, 872–82. [Google Scholar] [CrossRef]
- Khalil M, Ronda J, Weintraub M, Jain K, Silver R, Silverman AJ. Brain mast cell relationship to neurovasculature during development. Brain Res. 2007, 1171, 18–29. [Google Scholar] [CrossRef]
- Molina-Hernández A, Velasco I. Histamine induces neural stem cell proliferation and neuronal differentiation by activation of distinct histamine receptors. J Neurochem. 2008, 106, 706–17. [Google Scholar] [CrossRef] [PubMed]
- Liao R, Chen Y, Cheng L, Fan L, Chen H, Wan Y, You Y, Zheng Y, Jiang L, Chen Z, Zhang X, Hu W. Histamine H1 Receptors in Neural Stem Cells Are Required for the Promotion of Neurogenesis Conferred by H3 Receptor Antagonism following Traumatic Brain Injury. Stem Cell Reports. 2019, 12, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Johnson D, Krenger W. Interactions of mast cells with the nervous system--recent advances. Neurochem Res. 1992, 17, 939–51. [Google Scholar] [CrossRef] [PubMed]
- Skaper SD, Facci L, Kee WJ, Strijbos PJ. Potentiation by histamine of synaptically mediated excitotoxicity in cultured hippocampal neurones: a possible role for mast cells. J Neurochem. 2001, 76, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Dong H, Zhang W, Zeng X, Hu G, Zhang H, He S, Zhang S. Histamine induces upregulated expression of histamine receptors and increases release of inflammatory mediators from microglia. Mol Neurobiol. 2014, 49, 1487–500. [Google Scholar] [CrossRef] [PubMed]
- Xu J, Zhang X, Qian Q, Wang Y, Dong H, Li N, Qian Y, Jin W. Histamine upregulates the expression of histamine receptors and increases the neuroprotective effect of astrocytes. J Neuroinflammation. 2018, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Zhang W, Zhang X, Zhang Y, Qu C, Zhou X, Zhang S. Histamine Induces Microglia Activation and the Release of Proinflammatory Mediators in Rat Brain Via H1R or H4R. J Neuroimmune Pharmacol. 2020, 15, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Puttonen HAJ, Semenova S, Sundvik M, Panula P. Storage of neural histamine and histaminergic neurotransmission is VMAT2 dependent in the zebrafish. Sci Rep. 2017, 7, 3060. [Google Scholar] [CrossRef] [PubMed]
- Yanai K, Tashiro M. The physiological and pathophysiological roles of neuronal histamine: an insight from human positron emission tomography studies. Pharmacol Ther. 2007, 113, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Moon TC, Befus AD, Kulka M. Mast cell mediators: their differential release and the secretory pathways involved. Front Immunol. 2014, 5, 569. [Google Scholar] [CrossRef]
- Baranoglu Kilinc Y, Dilek M, Kilinc E, Torun IE, Saylan A, Erdogan Duzcu S. Capsaicin attenuates excitotoxic-induced neonatal brain injury and brain mast cell-mediated neuroinflammation in newborn rats. Chem Biol Interact. 2023, 376, 110450. [Google Scholar] [CrossRef]
- Conti P, Ronconi G, Lauritano D, Mastrangelo F, Caraffa A, Gallenga CE, Frydas I, Kritas SK, Carinci F, Gaudelli F, Annicchiarico C, D'Ovidio C. Impact of TNF and IL-33 Cytokines on Mast Cells in Neuroinflammation. Int J Mol Sci. 2024, 25, 3248. [Google Scholar] [CrossRef]
- Conti P, Lauritano D, Caraffa A, Gallenga CE, Kritas SK, Ronconi G, Martinotti S. Microglia and mast cells generate proinflammatory cytokines in the brain and worsen inflammatory state: Suppressor effect of IL-37. Eur J Pharmacol. 2020, 875, 173035. [Google Scholar] [CrossRef] [PubMed]
- Ellenbroek, BA. Histamine H₃ receptors, the complex interaction with dopamine and its implications for addiction. Br J Pharmacol. 2013, 170, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Varaschin RK, Osterstock G, Ducrot C, Leino S, Bourque MJ, Prado MAM, Prado VF, Salminen O, Rannanpää Née Nuutinen S, Trudeau LE. Histamine H3 Receptors Decrease Dopamine Release in the Ventral Striatum by Reducing the Activity of Striatal Cholinergic Interneurons. Neuroscience. 2018, 376, 188–203. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa T, Naganuma F, Iida T, Nakamura T, Harada R, Mohsen AS, Kasajima A, Sasano H, Yanai K. Molecular mechanism of histamine clearance by primary human astrocytes. Glia. 2013, 61, 905–16. [Google Scholar] [CrossRef]
- Naganuma F, Yoshikawa T. Organic Cation Transporters in Brain Histamine Clearance: Physiological and Psychiatric Implications. Handb Exp Pharmacol. 2021, 266, 169–185. [Google Scholar] [CrossRef]
- Chung WS, Allen NJ, Eroglu C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb Perspect Biol. 2015, 7, a020370. [Google Scholar] [CrossRef]
- Diagnostic and statistical manual of mental disorders: DSM-5 [5th ed., text revision (DSM-5-TR (TM))]. Washington: American Psychiatric Association. 2022. ISBN 978-0-89042-554-1.
- Gupta S, Kulhara P. What is schizophrenia: A neurodevelopmental or neurodegenerative disorder or a combination of both? A critical analysis. Indian J Psychiatry. 2010, 52, 21–7. [Google Scholar] [CrossRef]
- Shan L, Fronczek R, Lammers GJ, Swaab DF. The tuberomamillary nucleus in neuropsychiatric disorders. Handb Clin Neurol. 2021, 180, 389–400. [Google Scholar] [CrossRef]
- Shan L, Bao AM, Swaab DF. The human histaminergic system in neuropsychiatric disorders. Trends Neurosci. 2015, 38, 167–77. [Google Scholar] [CrossRef] [PubMed]
- Cristino AS, Williams SM, Hawi Z, An JY, Bellgrove MA, Schwartz CE, Costa Lda F, Claudianos C. Neurodevelopmental and neuropsychiatric disorders represent an interconnected molecular system. Mol Psychiatry. 2014, 19, 294–301. [Google Scholar] [CrossRef]
- Gudmundsson OO, Walters GB, Ingason A, Johansson S, Zayats T, Athanasiu L, Sonderby IE, Gustafsson O, Nawaz MS, Jonsson GF, Jonsson L, Knappskog PM, Ingvarsdottir E, Davidsdottir K, Djurovic S, Knudsen GPS, Askeland RB, Haraldsdottir GS, Baldursson G, Magnusson P, Sigurdsson E, Gudbjartsson DF, Stefansson H, Andreassen OA, Haavik J, Reichborn-Kjennerud T, Stefansson K. Attention-deficit hyperactivity disorder shares copy number variant risk with schizophrenia and autism spectrum disorder. Transl Psychiatry. 2019; 9, 258. [Google Scholar] [CrossRef]
- Ruediger T, Bolz J. Neurotransmitters and the development of neuronal circuits. Adv Exp Med Biol. 2007, 621, 104–15. [Google Scholar] [CrossRef]
- Di Marco B, Bonaccorso CM, Aloisi E, D'Antoni S, Catania MV. Neuro-Inflammatory Mechanisms in Developmental Disorders Associated with Intellectual Disability and Autism Spectrum Disorder: A Neuro- Immune Perspective. CNS Neurol Disord Drug Targets 2016, 15, 448–63. [Google Scholar] [CrossRef]
- Verhoeven WMA, Egger JIM, Janssen PKC, van Haeringen A. Adult male patient with severe intellectual disability caused by a homozygous mutation in the HNMT gene. BMJ Case Rep. 2020, 13, e235972. [Google Scholar] [CrossRef] [PubMed]
- Griswold AJ, Ma D, Cukier HN, Nations LD, Schmidt MA, Chung RH, Jaworski JM, Salyakina D, Konidari I, Whitehead PL, Wright HH, Abramson RK, Williams SM, Menon R, Martin ER, Haines JL, Gilbert JR, Cuccaro ML, Pericak-Vance MA. Evaluation of copy number variations reveals novel candidate genes in autism spectrum disorder-associated pathways. Hum Mol Genet. 2012, 21, 3513–23. [Google Scholar] [CrossRef] [PubMed]
- Wright C, Shin JH, Rajpurohit A, Deep-Soboslay A, Collado-Torres L, Brandon NJ, Hyde TM, Kleinman JE, Jaffe AE, Cross AJ, Weinberger DR. Altered expression of histamine signaling genes in autism spectrum disorder. Transl Psychiatry. 2017, 7, e1126. [Google Scholar] [CrossRef] [PubMed]
- Eissa N, Sadeq A, Sasse A, Sadek B. Role of Neuroinflammation in Autism Spectrum Disorder and the Emergence of Brain Histaminergic System. Lessons Also for BPSD? Front Pharmacol. 2020, 11, 886. [Google Scholar] [CrossRef]
- Abruzzo PM, Matté A, Bolotta A, Federti E, Ghezzo A, Guarnieri T, Marini M, Posar A, Siciliano A, De Franceschi L, Visconti P. Plasma peroxiredoxin changes and inflammatory cytokines support the involvement of neuro-inflammation and oxidative stress in Autism Spectrum Disorder. J Transl Med. 2019, 17, 332. [Google Scholar] [CrossRef]
- Blasco-Fontecilla, H. Is Histamine and Not Acetylcholine the Missing Link between ADHD and Allergies? Speer Allergic Tension Fatigue Syndrome Re-Visited. J Clin Med. 2023, 12, 5350. [Google Scholar] [CrossRef]
- Yoshikawa T, Nakamura T, Yanai K. Histamine N-Methyltransferase in the Brain. Int J Mol Sci. 2019, 20, 737. [Google Scholar] [CrossRef]
- Blasco-Fontecilla H, Bella-Fernández M, Wang P, Martin-Moratinos M, Li C. Prevalence and Clinical Picture of Diamine Oxidase Gene Variants in Children and Adolescents with Attention Deficit Hyperactivity Disorder: A Pilot Study. J Clin Med. 2024, 13, 1659. [Google Scholar] [CrossRef]
- Ercan-Sencicek AG, Stillman AA, Ghosh AK, Bilguvar K, O'Roak BJ, Mason CE, Abbott T, Gupta A, King RA, Pauls DL, Tischfield JA, Heiman GA, Singer HS, Gilbert DL, Hoekstra PJ, Morgan TM, Loring E, Yasuno K, Fernandez T, Sanders S, Louvi A, Cho JH, Mane S, Colangelo CM, Biederer T, Lifton RP, Gunel M, State MW. L-histidine decarboxylase and Tourette's syndrome. N Engl J Med. 2010, 362, 1901–8. [Google Scholar] [CrossRef]
- Pittenger, C. The histidine decarboxylase model of tic pathophysiology: a new focus on the histamine H3 receptor. Br J Pharmacol. 2020, 177, 570–579. [Google Scholar] [CrossRef]
- Baldan LC, Williams KA, Gallezot JD, Pogorelov V, Rapanelli M, Crowley M, Anderson GM, Loring E, Gorczyca R, Billingslea E, Wasylink S, Panza KE, Ercan-Sencicek AG, Krusong K, Leventhal BL, Ohtsu H, Bloch MH, Hughes ZA, Krystal JH, Mayes L, de Araujo I, Ding YS, State MW, Pittenger C. Histidine decarboxylase deficiency causes tourette syndrome: parallel findings in humans and mice. Neuron 2014, 81, 77–90. [Google Scholar] [CrossRef]
- Xu L, Zhang C, Zhong M, Che F, Guan C, Zheng X, Liu S. Role of histidine decarboxylase gene in the pathogenesis of Tourette syndrome. Brain Behav. 2022, 12, e2511. [Google Scholar] [CrossRef] [PubMed]
- Zhongling K, Yanhui C, Guofeng C, Yanyan L. Neuroinflammation in a Rat Model of Tourette Syndrome. Front Behav Neurosci. 2022, 16, 710116. [Google Scholar] [CrossRef]
- Wang X, Liu X, Chen L, Zhang X. The inflammatory injury in the striatal microglia-dopaminergic-neuron crosstalk involved in Tourette syndrome development. Front Immunol. 2023, 14, 1178113. [Google Scholar] [CrossRef]
- Ito, C. The role of the central histaminergic system on schizophrenia. Drug News Perspect. 2004, 17, 383–7. [Google Scholar] [CrossRef] [PubMed]
- Arrang, JM. Histamine and schizophrenia. Int Rev Neurobiol. 2007, 78, 247–87. [Google Scholar] [CrossRef]
- Vallée, A. Neuroinflammation in Schizophrenia: The Key Role of the WNT/β-Catenin Pathway. Int J Mol Sci. 2022, 23, 2810. [Google Scholar] [CrossRef]
- Murphy CE, Walker AK, Weickert CS. Neuroinflammation in schizophrenia: the role of nuclear factor kappa B. Transl Psychiatry. 2021, 11, 528. [Google Scholar] [CrossRef] [PubMed]
- Ma Q, Jiang L, Chen H, An D, Ping Y, Wang Y, Dai H, Zhang X, Wang Y, Chen Z, Hu W. Histamine H2 receptor deficit in glutamatergic neurons contributes to the pathogenesis of schizophrenia. Proc Natl Acad Sci U S A. 2023, 120, e2207003120. [Google Scholar] [CrossRef]
- Behl T, Kaur G, Sehgal A, Bhardwaj S, Singh S, Buhas C, Judea-Pusta C, Uivarosan D, Munteanu MA, Bungau S. Multifaceted Role of Matrix Metalloproteinases in Neurodegenerative Diseases: Pathophysiological and Therapeutic Perspectives. Int J Mol Sci. 2021, 22, 1413. [Google Scholar] [CrossRef]
- Giri PM, Banerjee A, Ghosal A, Layek B. Neuroinflammation in Neurodegenerative Disorders: Current Knowledge and Therapeutic Implications. Int J Mol Sci. 2024, 25, 3995. [Google Scholar] [CrossRef]
- Singh K, Sethi P, Datta S, Chaudhary JS, Kumar S, Jain D, Gupta JK, Kumar S, Guru A, Panda SP. Advances in gene therapy approaches targeting neuro-inflammation in neurodegenerative diseases. Ageing Res Rev. 2024, 98, 102321. [Google Scholar] [CrossRef]
- Armstrong RA, Lantos PL, Cairns NJ. Overlap between neurodegenerative disorders. Neuropathology. 2005, 25, 111–24. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, RA. On the 'classification' of neurodegenerative disorders: discrete entities, overlap or continuum? Folia Neuropathol. 2012, 50, 201–8. [Google Scholar] [CrossRef]
- De Marchi F, Munitic I, Vidatic L, Papić E, Rački V, Nimac J, Jurak I, Novotni G, Rogelj B, Vuletic V, Liscic RM, Cannon JR, Buratti E, Mazzini L, Hecimovic S. Overlapping Neuroimmune Mechanisms and Therapeutic Targets in Neurodegenerative Disorders. Biomedicines. 2023, 11, 2793. [Google Scholar] [CrossRef]
- Herman FJ, Simkovic S, Pasinetti GM. Neuroimmune nexus of depression and dementia: Shared mechanisms and therapeutic targets. Br J Pharmacol. 2019, 176, 3558–3584. [Google Scholar] [CrossRef]
- Ahmed RM, Devenney EM, Irish M, Ittner A, Naismith S, Ittner LM, Rohrer JD, Halliday GM, Eisen A, Hodges JR, Kiernan MC. Neuronal network disintegration: common pathways linking neurodegenerative diseases. J Neurol Neurosurg Psychiatry. 2016, 87, 1234–1241. [Google Scholar] [CrossRef]
- Kwakowsky A, Prasad AA, Peña-Ortega F, Lim SAO. Editorial: Neuronal network dysfunction in neurodegenerative disorders. Front Neurosci. 2023, 17, 1151156. [Google Scholar] [CrossRef]
- Taslim S, Shadmani S, Saleem AR, Kumar A, Brahma F, Blank N, Bashir MA, Ansari D, Kumari K, Tanveer M, Varrassi G, Kumar S, Raj A. Neuropsychiatric Disorders: Bridging the Gap Between Neurology and Psychiatry. Cureus. 2024, 16, e51655. [Google Scholar] [CrossRef]
- Aldossari AA, Assiri MA, Ansari MA, Nadeem A, Attia SM, Bakheet SA, Albekairi TH, Alomar HA, Al-Mazroua HA, Almanaa TN, Al-Hamamah MA, Alwetaid MY, Ahmad SF. Histamine H4 Receptor Antagonist Ameliorates the Progression of Experimental Autoimmune Encephalomyelitis via Regulation of T-Cell Imbalance. Int J Mol Sci. 2023, 24, 15273. [Google Scholar] [CrossRef]
- Lenz KM, Pickett LA, Wright CL, Davis KT, Joshi A, McCarthy MM. Mast Cells in the Developing Brain Determine Adult Sexual Behavior. J Neurosci. 2018, 38, 8044–8059. [Google Scholar] [CrossRef] [PubMed]
- Zhang W, Xiao D, Mao Q, Xia H. Role of neuroinflammation in neurodegeneration development. Signal Transduct Target Ther. 2023, 8, 267. [Google Scholar] [CrossRef]
- Owen JE, Veasey SC. Impact of sleep disturbances on neurodegeneration: Insight from studies in animal models. Neurobiol Dis. 2020, 139, 104820. [Google Scholar] [CrossRef]
- Dailah, HG. Potential of Therapeutic Small Molecules in Apoptosis Regulation in the Treatment of Neurodegenerative Diseases: An Updated Review. Molecules. 2022, 27, 7207. [Google Scholar] [CrossRef]
- Calabresi P, Mechelli A, Natale G, Volpicelli-Daley L, Di Lazzaro G, Ghiglieri V. Alpha-synuclein in Parkinson's disease and other synucleinopathies: from overt neurodegeneration back to early synaptic dysfunction. Cell Death Dis. 2023, 14, 176. [Google Scholar] [CrossRef]
- Troncoso-Escudero P, Parra A, Nassif M, Vidal RL. Outside in: Unraveling the Role of Neuroinflammation in the Progression of Parkinson's Disease. Front Neurol. 2018, 9, 860. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, KA. Basic mechanisms of neurodegeneration: a critical update. J Cell Mol Med. 2010, 14, 457–87. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni F, Quintana FJ. The Role of Astrocytes in CNS Inflammation. Trends Immunol. 2020, 41, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Shao F, Wang X, Wu H, Wu Q, Zhang J. Microglia and Neuroinflammation: Crucial Pathological Mechanisms in Traumatic Brain Injury-Induced Neurodegeneration. Front Aging Neurosci. 2022, 14, 825086. [Google Scholar] [CrossRef]
- Muzio L, Viotti A, Martino G. Microglia in Neuroinflammation and Neurodegeneration: From Understanding to Therapy. Front Neurosci. 2021, 15, 742065. [Google Scholar] [CrossRef]
- Shan L, Swaab DF, Bao AM. Neuronal histaminergic system in aging and age-related neurodegenerative disorders. Exp Gerontol. 2013, 48, 603–7. [Google Scholar] [CrossRef] [PubMed]
- Shan L, Swaab DF. Changes in Histaminergic System in Neuropsychiatric Disorders and the Potential Treatment Consequences. Curr Neuropharmacol. 2022, 20, 403–411. [Google Scholar] [CrossRef]
- Whalen TC, Gittis AH. Histamine and deep brain stimulation: the pharmacology of regularizing a brain. J Clin Invest. 2018, 128, 5201–5202. [Google Scholar] [CrossRef]
- Grotemeyer A, McFleder RL, Wu J, Wischhusen J, Ip CW. Neuroinflammation in Parkinson's Disease - Putative Pathomechanisms and Targets for Disease-Modification. Front Immunol. 2022, 13, 878771. [Google Scholar] [CrossRef]
- Shan L, Liu CQ, Balesar R, Hofman MA, Bao AM, Swaab DF. Neuronal histamine production remains unaltered in Parkinson's disease despite the accumulation of Lewy bodies and Lewy neurites in the tuberomamillary nucleus. Neurobiol Aging. 2012, 33, 1343–4. [Google Scholar] [CrossRef]
- Abdalla S, Eissa N, Jayaprakash P, Beiram R, Kuder KJ, Łażewska D, Kieć-Kononowicz K, Sadek B. The Potent and Selective Histamine H3 Receptor Antagonist E169 Counteracts Cognitive Deficits and Mitigates Disturbances in the PI3K/AKT/GSK-3β Signaling Pathway in MK801-Induced Amnesia in Mice. Int J Mol Sci. 2023, 24, 12719. [Google Scholar] [CrossRef]
- Naddafi F, Mirshafiey A. The neglected role of histamine in Alzheimer's disease. Am J Alzheimers Dis Other Demen. 2013, 28, 327–36. [Google Scholar] [CrossRef] [PubMed]
- van Wamelen DJ, Shan L, Aziz NA, Anink JJ, Bao AM, Roos RA, Swaab DF. Functional increase of brain histaminergic signaling in Huntington's disease. Brain Pathol. 2011, 21, 419–27. [Google Scholar] [CrossRef]
- Moreno-Delgado D, Puigdellívol M, Moreno E, Rodríguez-Ruiz M, Botta J, Gasperini P, Chiarlone A, Howell LA, Scarselli M, Casadó V, Cortés A, Ferré S, Guzmán M, Lluís C, Alberch J, Canela EI, Ginés S, McCormick PJ. Modulation of dopamine D1 receptors via histamine H3 receptors is a novel therapeutic target for Huntington's disease. Elife. 2020, 9, e51093. [Google Scholar] [CrossRef]
- Rapanelli, M. The magnificent two: histamine and the H3 receptor as key modulators of striatal circuitry. Prog Neuropsychopharmacol Biol Psychiatry. 2017, 73, 36–40. [Google Scholar] [CrossRef]
- Jia Q, Li S, Li XJ, Yin P. Neuroinflammation in Huntington's disease: From animal models to clinical therapeutics. Front Immunol. 2022, 13, 1088124. [Google Scholar] [CrossRef]
- Angeles-López QD, González-Espinosa C, Pérez-Severiano F. Mast cells and histamine are involved in the neuronal damage observed in a quinolinic acid-induced model of Huntington's disease. J Neurochem. 2022, 160, 256–270. [Google Scholar] [CrossRef] [PubMed]
- Hersey M, Hashemi P, Reagan LP. Integrating the monoamine and cytokine hypotheses of depression: Is histamine the missing link? Eur J Neurosci. 2022, 55, 2895–2911. [Google Scholar] [CrossRef]
- Qian H, Shu C, Xiao L, Wang G. Histamine and histamine receptors: Roles in major depressive disorder. Front Psychiatry. 2022, 13, 825591. [Google Scholar] [CrossRef] [PubMed]
- Kumar A, Dogra S, Sona C, Umrao D, Rashid M, Singh SK, Wahajuddin M, Yadav PN. Chronic histamine 3 receptor antagonism alleviates depression like conditions in mice via modulation of brain-derived neurotrophic factor and hypothalamus-pituitary adrenal axis. Psychoneuroendocrinology. 2019, 101, 128–137. [Google Scholar] [CrossRef]
- Alhusaini M, Eissa N, Saad AK, Beiram R, Sadek B. Revisiting Preclinical Observations of Several Histamine H3 Receptor Antagonists/Inverse Agonists in Cognitive Impairment, Anxiety, Depression, and Sleep-Wake Cycle Disorder. Front Pharmacol. 2022, 13, 861094. [Google Scholar] [CrossRef] [PubMed]
- Dauvilliers Y, Delallée N, Jaussent I, Scholz S, Bayard S, Croyal M, Schwartz JC, Robert P. Normal cerebrospinal fluid histamine and tele-methylhistamine levels in hypersomnia conditions. Sleep. 2012, 35, 1359–66. [Google Scholar] [CrossRef]
- Valizadeh P, Momtazmanesh S, Plazzi G, Rezaei N. Connecting the dots: An updated review of the role of autoimmunity in narcolepsy and emerging immunotherapeutic approaches. Sleep Med. 2024, 113, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Shan L, Dauvilliers Y, Siegel JM. Interactions of the histamine and hypocretin systems in CNS disorders. Nat Rev Neurol. 2015, 11, 401–13. [Google Scholar] [CrossRef]
- Barateau L, Krache A, Da Costa A, Lecendreux M, Debs R, Chenini S, Arlicot N, Vourc'h P, Evangelista E, Alonso M, Salabert AS, Silva S, Béziat S, Jaussent I, Mariano-Goulart D, Payoux P, Dauvilliers Y. Microglia Density and Its Association With Disease Duration, Severity, and Orexin Levels in Patients With Narcolepsy Type 1. Neurology. 2024, 102, e209326. [Google Scholar] [CrossRef]
- Passani MB, Blandina P. Histamine receptors in the CNS as targets for therapeutic intervention. Trends Pharmacol Sci. 2011, 32, 242–9. [Google Scholar] [CrossRef]
- Baronio D, Gonchoroski T, Castro K, Zanatta G, Gottfried C, Riesgo R. Histaminergic system in brain disorders: lessons from the translational approach and future perspectives. Ann Gen Psychiatry. 2014, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Yang L, Wang Y, Chen Z. Central histaminergic signalling, neural excitability and epilepsy. Br J Pharmacol. 2022, 179, 3–22. [Google Scholar] [CrossRef] [PubMed]
- del Rio R, Noubade R, Saligrama N, Wall EH, Krementsov DN, Poynter ME, Zachary JF, Thurmond RL, Teuscher C. Histamine H4 receptor optimizes T regulatory cell frequency and facilitates anti-inflammatory responses within the central nervous system. J Immunol. 2012, 188, 541–7. [Google Scholar] [CrossRef]
- Liberman AC, Trias E, da Silva Chagas L, Trindade P, Dos Santos Pereira M, Refojo D, Hedin-Pereira C, Serfaty CA. Neuroimmune and Inflammatory Signals in Complex Disorders of the Central Nervous System. Neuroimmunomodulation. 2018, 25, 246–270. [Google Scholar] [CrossRef]
- Zampeli E, Tiligada E. The role of histamine H4 receptor in immune and inflammatory disorders. Br J Pharmacol. 2009, 157, 24–33. [Google Scholar] [CrossRef]
- Panula P, Nuutinen S. The histaminergic network in the brain: basic organization and role in disease. Nat Rev Neurosci. 2013, 14, 472–87. [Google Scholar] [CrossRef]
- Sundvik M, Panula P. Interactions of the orexin/hypocretin neurones and the histaminergic system. Acta Physiol (Oxf). 2015, 213, 321–33. [Google Scholar] [CrossRef]
- Kou E, Zhang X, Dong B, Wang B, Zhu Y. Combination of H1 and H2 Histamine Receptor Antagonists: Current Knowledge and Perspectives of a Classic Treatment Strategy. Life (Basel). 2024, 14, 164. [Google Scholar] [CrossRef]
- Schlicker E, Kathmann M. Role of the Histamine H3 Receptor in the Central Nervous System. Handb Exp Pharmacol. 2017, 241, 277–299. [Google Scholar] [CrossRef]
- Feuerstein, TJ. Presynaptic receptors for dopamine, histamine, and serotonin. Handb Exp Pharmacol. 2008, 289–338. [Google Scholar] [CrossRef]
- Ellenbroek BA, Ghiabi B. The other side of the histamine H3 receptor. Trends Neurosci 2014, 37, 191–9. [Google Scholar] [CrossRef]
- Harwell V, Fasinu PS. Pitolisant and Other Histamine-3 Receptor Antagonists-An Update on Therapeutic Potentials and Clinical Prospects. Medicines (Basel). 2020, 7, 55. [Google Scholar] [CrossRef]
- Sadek B, Saad A, Sadeq A, Jalal F, Stark H. Histamine H3 receptor as a potential target for cognitive symptoms in neuropsychiatric diseases. Behav Brain Res. 2016, 312, 415–30. [Google Scholar] [CrossRef] [PubMed]
- Fang Z, Chen J, Zheng Y, Chen Z. Targeting Histamine and Histamine Receptors for Memory Regulation: An Emotional Perspective. Curr Neuropharmacol. 2024, 22, 1846–1869. [Google Scholar] [CrossRef] [PubMed]
- Ghamari N, Zarei O, Arias-Montaño JA, Reiner D, Dastmalchi S, Stark H, Hamzeh-Mivehroud M. Histamine H3 receptor antagonists/inverse agonists: Where do they go? Pharmacol Ther. 2019, 200, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Szczepańska K, Kuder KJ, Kieć-Kononowicz K. Dual-targeting Approach on Histamine H3 and Sigma-1 Receptor Ligands as Promising Pharmacological Tools in the Treatment of CNS-linked Disorders. Curr Med Chem. 2021, 28, 2974–2995. [Google Scholar] [CrossRef]
- Flik G. The histaminergic system and its interaction with relevant central neurotransmitter systems. [Thesis fully internal (DIV), University of Groningen]. University of Groningen 2022. [CrossRef]
- Adamu A, Li S, Gao F, Xue G. The role of neuroinflammation in neurodegenerative diseases: current understanding and future therapeutic targets. Front Aging Neurosci. 2024, 16, 1347987. [Google Scholar] [CrossRef]
Figure 1.
Histamine biosynthesis and metabolism.
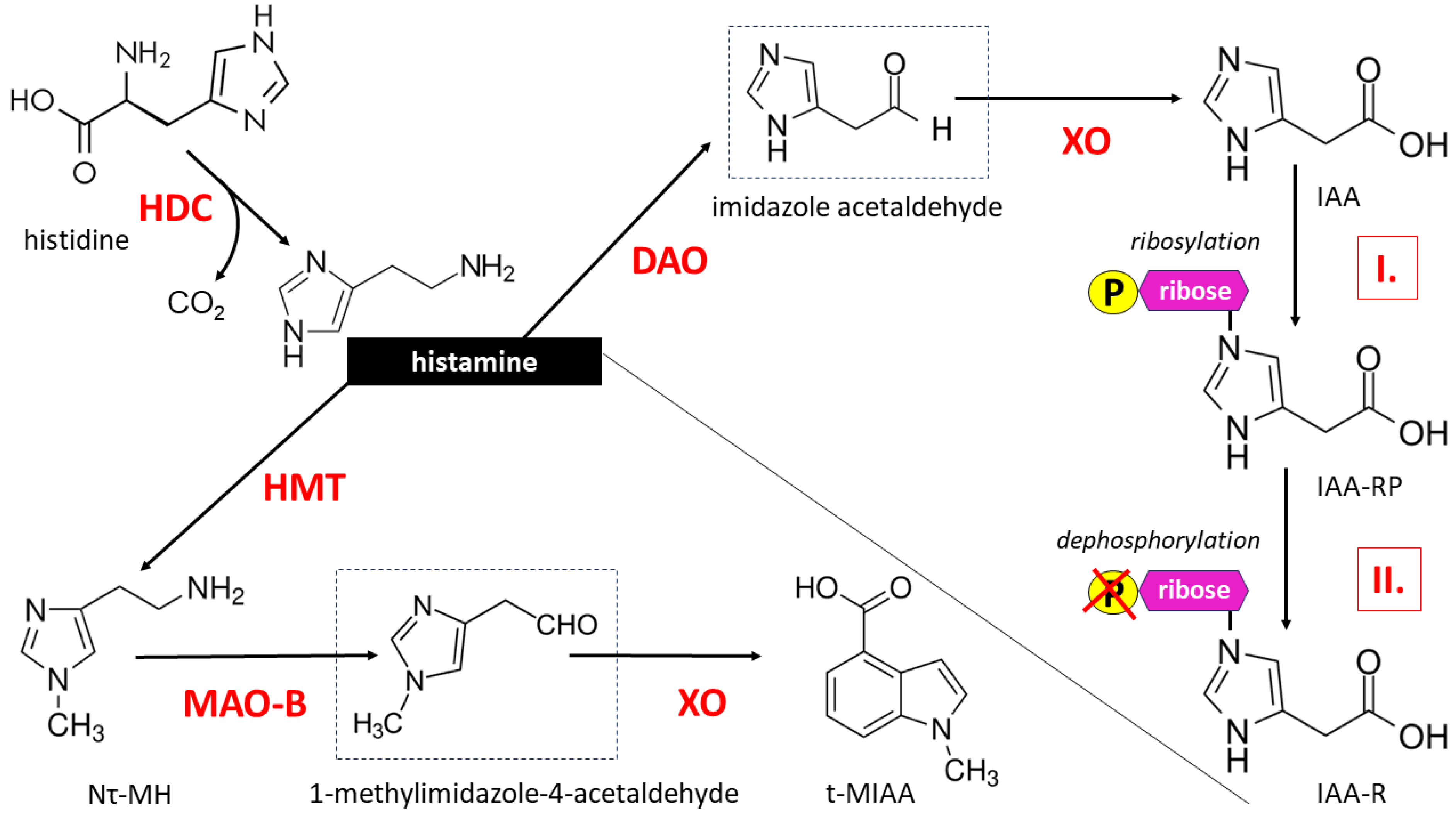
Figure 2.
Canonical signaling pathways related to histamine H1R– H4R receptors underlying the most typical manifestations of their activity, such as allergic inflammation, gastric acid secretion, neurotransmission and immunomodulation, respectively [19,31,33,37, 39,40,43]. Due to the polymorphism of the H1R - H4R receptor genes, various alternative pathways are described, which are omitted from the figure to maintain its readability. Activation of signaling pathways for all histamine receptors is accompanied by an increase (↑) in Ca 2+ concentration in the cytoplasm, while cAMP concentration increases (↑) after H1R and H2R stimulation and decreases (↓) after H3R and H4R stimulation.
Figure 2.
Canonical signaling pathways related to histamine H1R– H4R receptors underlying the most typical manifestations of their activity, such as allergic inflammation, gastric acid secretion, neurotransmission and immunomodulation, respectively [19,31,33,37, 39,40,43]. Due to the polymorphism of the H1R - H4R receptor genes, various alternative pathways are described, which are omitted from the figure to maintain its readability. Activation of signaling pathways for all histamine receptors is accompanied by an increase (↑) in Ca 2+ concentration in the cytoplasm, while cAMP concentration increases (↑) after H1R and H2R stimulation and decreases (↓) after H3R and H4R stimulation.

Figure 3.
Neuroanatomy of histaminergic transmission pathways in the human brain (inspired by Haas and Panula [80], modified from Haas, Sergeeva and Selbach [65]).
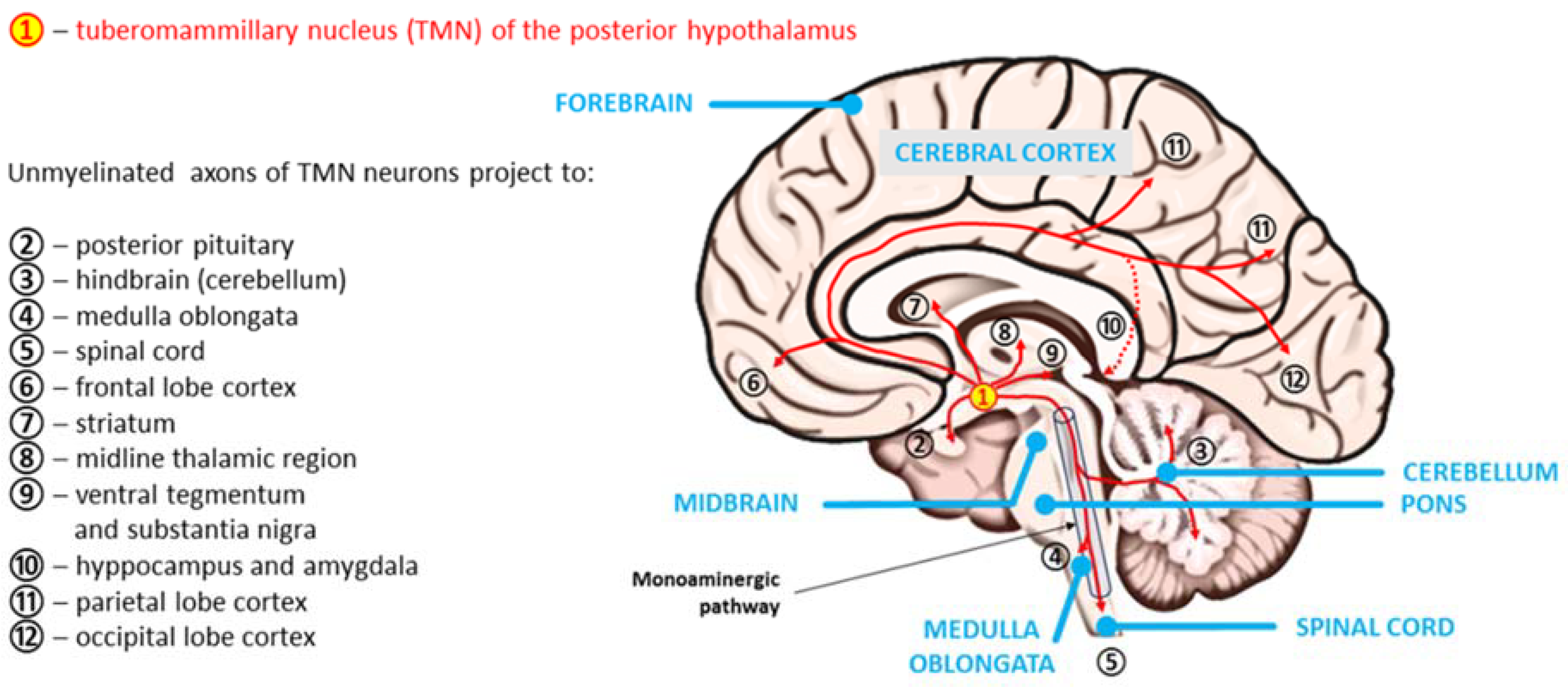
Figure 4.
Therapeutic approach taking into account the involvement of the CNS histaminergic system in the pathomechanism of neurodevelopmental disorders (marked in green) and neurodegenerative diseases (marked in brown): histamine H3 receptor (H3R) as the main target. Other major histamine receptors have also been marked in individual disorders [see Table 2. and Table 3.].
Figure 4.
Therapeutic approach taking into account the involvement of the CNS histaminergic system in the pathomechanism of neurodevelopmental disorders (marked in green) and neurodegenerative diseases (marked in brown): histamine H3 receptor (H3R) as the main target. Other major histamine receptors have also been marked in individual disorders [see Table 2. and Table 3.].
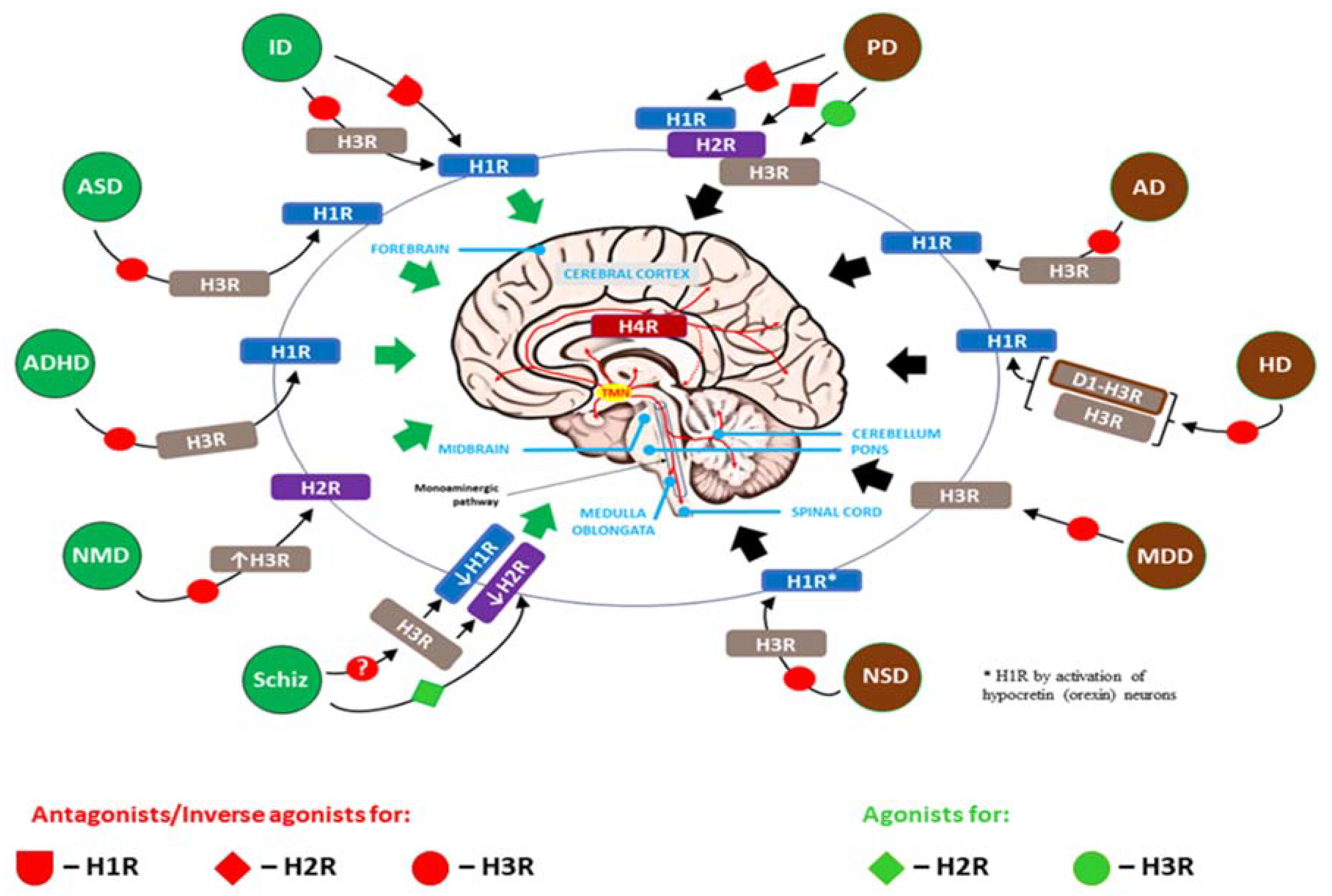

Table 1.
Molecular characteristics [30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45] and biological properties [18,19,27,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61] of the histamine (HA) receptor subtypes (H1R–H4R) in humans.
| Molecular Characteristics and Biological Properties | H1R | H2R | H3R | H4R |
|---|---|---|---|---|
| Chromosomal location of the gene | 3p25.3 | 5q35.2 | 20q13.33 | 18q11.2 |
| Receptor proteins, molecular weight (MW) | 487 aa, 55.78 kDa | 358-359 aa, 40.1–44.5 kDa (2 isoforms) | 445 aa (full-length), 36.4–49 kDa (≥ 20 possible mRNA isoforms) | 390 aa, 34.5–44.5 kDa (2 isoforms) |
| Gene structure | No introns | No introns | Three introns | Two introns |
| Class of receptor | Rhodopsin-like GPCRs (metabotropic), seven transmembrane domain (7TM) receptors | |||
| Binding affinity | Low (2.5 x 10 –5 M) | Low (7.9 x 10 –6 M) | High (6.3 x 10 –9 M) | High (7.9 x 10 –9 M) |
| Coupling subunit of the G protein complex | Gαq/11 | Gαs | Gαi | Gαi |
| The equilibrium dissociation constant (KD) | ~ 10 μmol/L | ~ 30 μmol/L | ~ 10 nmol/L | ~ 20-40 nmol/L |
| Selective agonists | Histaprodiphen | Amthamine, Dimaprit, Impromidine | Imetit, Immepip, α-Methylhistamine | Imetit *, Immepip *, 4-methylhistamine, Clobenopropit (partial agonist) |
| Antagonists (incl. inverse agonists) | Mepyramine, Cetirizine, Chlorpheniramine, Clemastine, Diphenhydramine, Pyrilamine, Triploidine, | Cimetidine, Famotidine, Nizatidine, Ranitidine | Clobenpropit, Ciproxifan, Pitolisant, Thioperamide * | 2-aminopyrimidine Thioperamide, VUF-6002, JNJ-10191584, Toreforant (JNJ-38518168), JNJ-7777120, CZC-13788, PF-2988403, A-940894, A-987306 |
| Tissue distribution | Widespread expression in many cells and tissues, including smooth muscles (e.g., in the respiratory tract and vessels), vascular ECs cells, the heart, CNS, adrenal medulla | Widespread expression in various cells and tissues, such as gastric mucosa parietal cells, smooth mucle (e.g., airways, uterus, vessels), the heart, immune cells (e.g., B cells, T cells, DCs), CNS, skin, genitourinary system, endocrine and exocrine glands | High expression in the CNS (histaminergic neurons) and neuroendocrine tissues, including enterochromaffin-like (ECL) cells in the gastric mucosa and adrenal cortex; to a lesser extent epression in the peripheral nervous system; low expression elsewhere | High expression in bone marrow, peripheral hematopoietic cells and immune cells, including MCs, DCs, T cells, NK cells, monocytes and eosinophils; low expression elsewhere |
| Transmitting intracellular signals/effector molecules (see also Figure 2.) | Mobilization of intracellular Ca 2+ levels: Gαq/11 activates PLC, which signals via DAG and PKC or IP3 to enhance Ca 2+ release from endoplasmic reticulum, activating eNOS to release NO; H1R can activate also PLA2, leading to the formation of AA and AADE. ↑ cGMP, cAMP accumulation (via Gβγ subunits), ↑ Ca 2+, ↑ NF-κβ, ↑ PLA2, ↑ PLD, ↑ AA, ↑ AADE |
Once AC activation is initiated by Gαs, subsequent signaling occurs via cAMP and PKA; Alternatively, via Gs, the H2R can activate PLC inducing DAG and IP3 pathways; Gαs promotes Ca2+, Na+ and Cl– chanel opening, changing the permeability of the cell membrane ↑ Ca 2+, ↑ c-fos , ↑ cAMP, ↑ PLC |
Activated Gαi inhibits AC and modulate MAPK activity via interaction with β-arrestin 2, the recruitment of which does not depend on the formation of an active G protein complex; Alternatively, activation of Gαi can inhibit K⁺ channels, Ca²⁺ channels, and the Na⁺/H⁺ transporter via arachidonic acid metabolites; ↓ cAMP, ↑ Ca 2+, ↑ MAPK and ↑ MAPK phosphorylation |
Recruitment of Gαi activates PLC and inhibits membrane-bound AC activity. As a result, Ca²⁺ is mobilized from intracellular stores, while cytosolic cAMP is diminished; Independently of G protein complex activation, β-arrestin 2 is recruited to the agonist-bound H4R, initiating MAPK cascade activation with subsequent receptor desensitization and internalization; ↓ cAMP, ↑ Ca 2+, ↑ MAPK and ↑ MAPK phosphorylation |
| Actions mediated | - Allergic disease: ↑ APCs capacity, degranulation of MCs and basophils (release of HA and other mediators), ↓ humoral immunity and ↑ Th1 priming, ↑ Th1, ↑ IFN-γ production, ↑ cellular adhesion molecule expression, and ↑ chemotaxis of eosinophils and neutrophils, ↑ vascular permeability, hypotension; - Smooth muscle contraction (airways, intestine, uterus) resulting in bronchoconstriction, vasodilation and uterine contraction, respectively; - Nociception (↑ pruritus, ↑ burning, ↑ pain); - Thermoregulation; - Regulation of the sleep-wake cycle, food intake, locomotion, emotions (aggressive behavior), memory and learning; - Negative dromotropic and negative atrial inotropic cardiac stimulation; - pro-angiogenic activity through H1R-PKC-VEGF-mediated pathway in EC. |
- Immune system: Suppression of immune cells (development of tolerance), including stimulation of suppressor T cells (Treg), ↓ neutrophil and basophil chemotaxis, ↓ neutrophil and basophil activation, ↓proliferation of lymphocytes, ↓ activity of NK cells; suppression of Th2 cells and cytokines; induction of IL-10 and suppression of IL-12 by DCs; ↑ humoral immunity; Indirect role in allergic response, autoimmunity, malignancy, and graft rejection; - Changes in H2R expression accompany the process of cell differentiation - ↑ Gastric acid secretion (↑ risk of gastric ulcer): - Inflammatory response: ↑ vascular permeability, vasodialtion (↑ risk of hypotension), flushing, headache; - Positive atrial chronotropic (↑ risk of tachycardia) and positive ventricular inotropic cardiac stimulation; - Bronchodilatation and ↑ mucus production (airways); - Uterus: relaxation. |
- Neurotransmission: Presynaptic autoreceptor function – inhibition of HA release (↓ histaminergic neurons activity), as well as presynaptic heteroreceptor activity in non-histaminergic neurons – inhibition of the release of DA, 5-HT, NA (↓ sympathetic tone), ACh, GABA and glutamate; - Sympathoinhibitory action, in vivo, leads to reduced vasoconstriction, thus may promote a vasodilatory effect; - Involvement in the pathophysiology of neuroinflammation through local neuron-MCs loops; proinflammatory effect through ↑ APCs activity; - Modulation of executive functions, involvement in cognitive impairment. |
- Immunomodulation: Involvement in DC activation and T cell differentiation; induction of proinflammatory AP-1 in Th2 cells and monocyte-derived DCs with simultaneous reduced production of the Th1-associated cytokine IL-12 and chemokine CXCL10 (IP10) in monocyte-derived DCs; modulation of eosinophil migration and selective recruitment of MCs leading to amplification of HA-mediated immune responses and subsequent chronic inflammation. |
Table 2.
The role of histamine (HA) signaling and neuroinflammation in the etiopathogenesis of neurodevelopmental disorders.
Table 2.
The role of histamine (HA) signaling and neuroinflammation in the etiopathogenesis of neurodevelopmental disorders.
| Type/Name of disorder | HA level in the CNS (incl. controversies)/possible causes or accompanying circumstances | Major HA receptor involved* | Neuroinflammation (regardless of HA)/ typical clinical symptoms in brief | References |
|---|---|---|---|---|
| Intellectual disability (intellectual development disorder) | Increased, due to a homozygous mutation in the histamine N-methyltransferase (HNMT) enzyme gene. HNMT is responsible for the degradation of intracellular HA. | H1R | Present / Previously called “mental retardation”; affected children start crawling, walking or talking later than other children and have trouble with learning (the main symptom is learning slowly), communicating, thinking rationally, making judgments, and planning | Di Marco et al., 2016 [150]; Verhoeven et al., 2020 [151] |
| Autism spectrum disorders (ASD) | Normal. This suggests that the response to HA is changed rather than its secretion or production in ASD. Different expression of the gene set of HNMT, H1R, H2R and H3R may explain the different effects of HS in ASD. | H1R, H3R | Present / Symptoms of this complex condition generally appear in the first 2 years of life and include persistent challenges with social communication (e.g., decreased sharing of interests with others, difficulty assessing emotions, lack of abstract thinking), restricted interests (inflexibility of behavior, extreme difficulty coping with change) and repetitive behavior (stereotypical movements such as hand flapping, rocking, spinning); the limitation of normal functioning due to these disorders shows significant differences between individuals with autism. | Di Marco et al., 2016 [150]; Verhoeven et al., 2020 [151]; Griswold et al., 2012 [152]; Wright et al., 2017 [153]; Eissa N et al., 2020 [154]; Abruzzo et al., 2019 [155] |
| Attention-deficit/ hyperactivity disorder (ADHD) | Increased, due to decreased activity of the diamine oxidase (DAO), the key enzyme responsible for extracellular HA degradation. Some single nucleotide polymorphism (SNP) variants of the AOC1 gene associated with DAO deficiency or decreased DAO functionality may be involved in the pathomechanism of ADHD. The increase in brain HA may also result from SNP of the HNMT, encoding the main enzyme involved in intracellular metabolism of HA. It was demonstrated that the “T” allele at rs11558538 is associated with decreased HNMT activity. |
H1R, H3R | Present / Executive dysfunction that can be categorized into 2 types of behavioral problems: inattentiveness (difficulty concentrating and focusing) and hyperactivity and impulsiveness | Blasco-Fontecilla 2023 [156]; Yoshikawa et al., 2019 [157]; Blasco-Fontecilla et al., 2024 [158] |
| Neurodevelopmental motor disorders (e.g., Tourette’s syndrome) | Decreased, due to a rare nonsense mutation, HDC W317X, in the exon 9 of the L-histidine decarboxylase (HDC) gene. This hypomorphic mutation in the HDC gene is a rare but high penetrance genetic cause of TS. Subsequent deficiency of HA disrupts dopamine modulation of basal ganglia, because HA reduces dopamine levels in the stratum. | H2R and up-regulated H3R in the striatum | Present / Symptoms of motor disorders include tremors, jerks, twitches, spasms, contractions, or gait problems; these disorders can significantly limit intentional movements and cause an excess of involuntary movements, including tics (fast, repetitive muscle movements that result in sudden and difficult to control body jolts or sounds) | Ercan-Sencicek et al., 2010 [159]; Pittenger 2020 [160]; Baldan et al., [161]; Xu et al., 2022 [162]; Zhongling et al., 2022 [163]; Wang et al., 2023 [164] |
| Schizophrenia | Increased levels of N-tele-methylhistamine (Nτ-MH), a major brain metabolite of HA in the cerebrospinal fluid (CSF) | Reduced efficiency and/or decreased number of H1R binding sites, H2R (including deficit in glutamatergic neurons), H3R | Present, particularly in the dorsolateral prefrontal cortex (PFC) / A chronic mental health condition that causes a range of different psychological symptoms including reoccurring episodes of psychosis that are correlated with a general misperception of reality. The active form of schizophrenia is characterized by: delusions, hallucinations, disorganized speech, trouble with thinking and lack of motivation. | Ito 2004 [165]; Arrang 2007 [166]; Vallée 2022 [167]; Murphy et al., 2021 [168]; Ma et al., 2023 [169] |
* In the case of neuroinflammation, the involvement of H4R, which is expressed by increased amounts of hematopoietic and immune cells (i.e., mast cells, basophils, eosinophils, dendritic cells and CD4(+) T cells) flowing into the CNS, should be taken into account.
Table 3.
The role of histamine (HA) signaling and neuroinflammation in the etiopathogenesis of neuropsychiatric (neurodegenerative) disorders.
Table 3.
The role of histamine (HA) signaling and neuroinflammation in the etiopathogenesis of neuropsychiatric (neurodegenerative) disorders.
| Type/Name of disorder | HA level in the CNS (incl. controversies)/possible causes or accompanying circumstances | Major HA receptor involved* | Neuroinflammation (regardless of HA)/typical clinical symptoms in brief | References |
|---|---|---|---|---|
| Parkinson's disease (PD) | Unaltered or increased. Expression of HDC-mRNA in the TMN remains undisturbed and the enzymatic activity of HDC preserved within normal range. Moreover, the concentration of the main metabolite of HA, t-MeHA, remains unchanged in the CSF. However, abnormally high HA concentration in the basal ganglia of the brains of PD patients has been shown postmortem in other studies. |
H1R,H2R, H3R | Present / Motor symptoms include: tremor (particularly when at rest), slowness of movement (bradykinesia), rigidity (stiffness), posture and balance problems (frequent falls), involuntary movements (dyskinesia) and muscle spasms (dystonia), speech changes (weak, hoarse, nasal or monotonous voice, imprecise articulation, slow or fast speech, difficulty starting to speak, problems with accentuation or rhythm); non-motor symptoms include: anxiety and depression, stress, apathy, compulsive or impulsive behavior, dementia, changes in sleep | Cheng et al., 2021 [57]; Nuutinen and Panula, 2010 [69]; Shan et al., 2013 [191]; Shan and Swaab, 2022 [192]; Whalen and Gittis, 2018 [193]; Grotemeyer et al., [194] |
| Alzheimer's disease (AD) | Unaltered or decreased. Despite of the significant loss of histaminergic neurons in AD, expression of HDC-mRNA in the TMN remains unaltered. The decline in H1R binding may have a role in the cognitive deficits of patients with AD. Many studies have also demonstrated the involvement of H3R, whose loss of integrity and/or increased expression may locally reduce HA concentration in the CNS. |
H1R, H3R | Present / The typical and common early symptom is difficulty in remembering recent events; the symptoms of dementia develop gradually over many years and eventually become more severe; in the early stages of the disease, decline in non-memory aspects of cognition, such as finding the right word, trouble understanding visual images and spatial relationships, and impaired reasoning or judgment may be observed; in the advanced-stage Alzheimer’s, patients are diagnosed with severe impairment/loss of speaking ability, disorientation (including easily getting lost), mood swings, loss of motivation, self-neglect, and behavioral issues, including lack of response to stimuli from the environment; Alzheimer's disease is the most common type of dementia. | Zhou et al., 2024 [59]; Nuutinen and Panula, 2010 [69]; Shan et al., 2013 [191]; Shan and Swaab, 2022 [192]; Shan et al., 2012 [195]; Abdalla et al., 2023 [196]; Naddafi and Mirshafiey, 2013 [197] |
| Huntington's disease (HD) | Normal to increased. TMN volume and number of TMN neurons remain within the normal range, whereas the levels of HDC mRNA, HMT activity within the inferior frontal gyrus (IFG), H1R and H3R mRNA levels are increased. These findings indicate a functional increase in brain histaminergic signaling in HD that is linked to aberrant striatal function. | H3R, especially a hybrid dopamine-histamine receptor (D1R–H3R, heteromer), H1R | Present / Choreiform movements (jerking or writhing movements), psychiatric problems [depression, irritability/aggression, psychosis, executive dysfunction (e.g., obsessive-compulsive behaviors, apathy)], cognitive decline and dementia (the commonest genetic dementia with autosomal dominant inheritance). | van Wamelen et al., 2011 [198]; Moreno-Delgado et al., 2020 [199]; Rapanelli 2017 [200]; Jia Q et al., 2022 [201]; Angeles-López et al., 2022 [202] |
| Depression (also called major depressive disorder or clinical depression) | Decreased. A decrease in the HA level, especially related to the activation of H3R. Melanin-concentrating hormone (MCH) neurons express H3R through which HA directly inhibits release of MCH, an orexigenic peptide, with confirmed depressogenic effects. Moreover, other neural circuits may be influenced by lowered level of HA via H3R signaling, due to the crosstalk of HA-H3R signaling pathways with other depression-related neurotransmitters, such as 5-HT, DA and GLU. | H3R can interact with other depression-related transmitters (including 5-HT, DA, GLU, and MCH); thus, histamine may participate in the occurrence of depression through other neural circuits. | Present / A persistent feeling of sadness, tearfulness, emptiness or hopelessness with loss of interest or pleasure in most or all normal activities (e.g., sex, hobbies sports); angry outbursts, irritability or frustration, even over small matters; sleep disturbances (both insomnia or excessive sleepiness); weight loss or weight gain related to lack of appetite or overeating, respectively; loss of concentration, difficulty making decisions, slowness; feeling anxious, thinking about impending death or having suicidal thoughts; somatic ailments caused by a lower pain threshold. | Hersey et al., 2022 [203]; Qian et al., 2022 [204]; Kumar et al., 2019 [205]; Alhusaini et al., 2022 [206] |
| Narcolepsy [also called narcolepsy spectrum disorder (NSD)] | Increased together with the increase in the number of HA neurons in narcolepsy type 1 (narcolepsy with cataplexy) or unchanged to decreased, despite of increased number of HDC-positive neurons. In cases of elevated HA concentration, an ineffective compensatory mechanism in response to hypocretin (orexin) neurons may be suspected. |
H1R by activation of hypocretin (orexin) neurons | Present / A chronic sleep disorder characterized by sudden, excessive and uncontrollable daytime sleepiness, catalepsy (a sudden loss of control of muscle tension ranging from slight weakness to total collapse), sleep paralysis and hypnagogic (occurring immediately before falling asleep) hallucinations. | Cheng et al., 2021 [57]; Valko et al., 2013 [81]; John et al., 2013 [82]; Dauvilliers et al., 2012 [207]; Valizadeh P et al., 2024 [208]; Shan et al., 2015 [209]; Barateau et al., 2024 [210]; |
* In the case of neuroinflammation, the involvement of H4R, which is expressed by increased amounts of hematopoietic and immune cells (i.e., mast cells, basophils, eosinophils, dendritic cells and CD4(+) T cells) flowing into the CNS, should be taken into account.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



