
Submitted:
09 September 2024
Posted:
11 September 2024
You are already at the latest version
Abstract
The ECM is composed of a considerable number of biochemically and structurally diverse constituents. ECM is a highly dynamic system that constantly receives and sends biological, chemical and mechanical signals. Several studies suggest that mechanical signals derived from the extracellular microenvironment regulate skin regeneration and wound healing. We evaluated effects of a mixture of biomolecules extracted and purified from bovine colostrum on restoring the mechanical properties of ECM. Test measuring collagen contraction showed a significant difference in contraction activation in samples treated with the 2% colostrum derivative mixture compared to the control. The analysis of the supernatant showed an inhibition of metalloproteinase-2 expression and an increase in collagen secretion by fibroblasts in treatment samples. Our hypothesis is that the molecules extracted and purified from bovine colostrum restore the ECM environment qualitative and quantitative characteristics, thus guaranteeing through a mechanical action the restoration of the wound due to the transduction of the signal activated by the integrins.
Keywords:
mechanical signals
; wound healing
; collagen
; integrin
; mechanotransducer
; ELISA test
1. Introduction
The cells within tissues are inserted in a highly structured microenvironment and are very sensitive to the geometric and mechanical constraints of the latter [1]. The cellular microenvironment, formed by the extracellular matrix (ECM) and neighboring cells, influences not only cellular architecture and mechanics, but also the polarity and functions of the cell [1,2]. Mechanical properties of the ECM depend on a complex protein network that forms a fibrous 3D scaffold whose structural components are collagen fibers, proteoglycans, and glycosaminoglycans [3]. Cells adhere to this network and act as a reservoir for nonstructural components such as growth factors, cytokines and proteolytic enzymes [4]. ECM proteins, by moving under the influence of forces, can act as mechanotransducers by exposing specific sites and growth factors [5]. The ECM is a dynamic structure, constantly undergoing a remodeling process, an important mechanism whereby cell differentiation can be regulated wound repair. In physiological conditions, the balance between processes of destruction and regeneration of the constituents of the extracellular matrix is regulated by specific tissue inhibitors of metalloproteinases (MMPs) [7]. An alteration of this balance, as in the case of wounds, is related to a catalytic hyperactivity of MMPs that play a critical role in wound healing. Their main function is degradation with removal of the damaged ECM during the inflammatory phase. The presence of these enzymes is necessary for effective wound healing, but they can play a harmful role at high concentrations, causing excessive tissue degradation and slow wound healing [7].
Knowledge of the cellular response to mechanical stimuli coming from the cellular microenvironment can be fundamental for applications in regenerative medicine to de-sign new and more effective scaffolds or biomaterials [8].
Scaffolds should mimic the properties of the target tissue [9], ensure the restoring of anisotropy (one of the fundamental characteristics of most tissues) [10,11,12] and through mechanotransduction activate integrins and induce the cascade of growth factors (TGF-beta 1, CTGF, IGF-1, etc.) necessaries, among other things, to produce new collagen by the fibroblast [13]. The response of integrins to force involves three mechanochemical steps. Integrins bind to ECM molecules transmitting forces into the cell that are converted into biochemical signals (mechanotransduction). Finally, integrins connect to the cytoskeleton to transmit forces throughout the cell and strengthen adhesions to resist forces. Mechanical signals that are transmitted through structural components of the cytoskeleton play a key role in events that regulate cell migration, polarity, and proliferation [14].
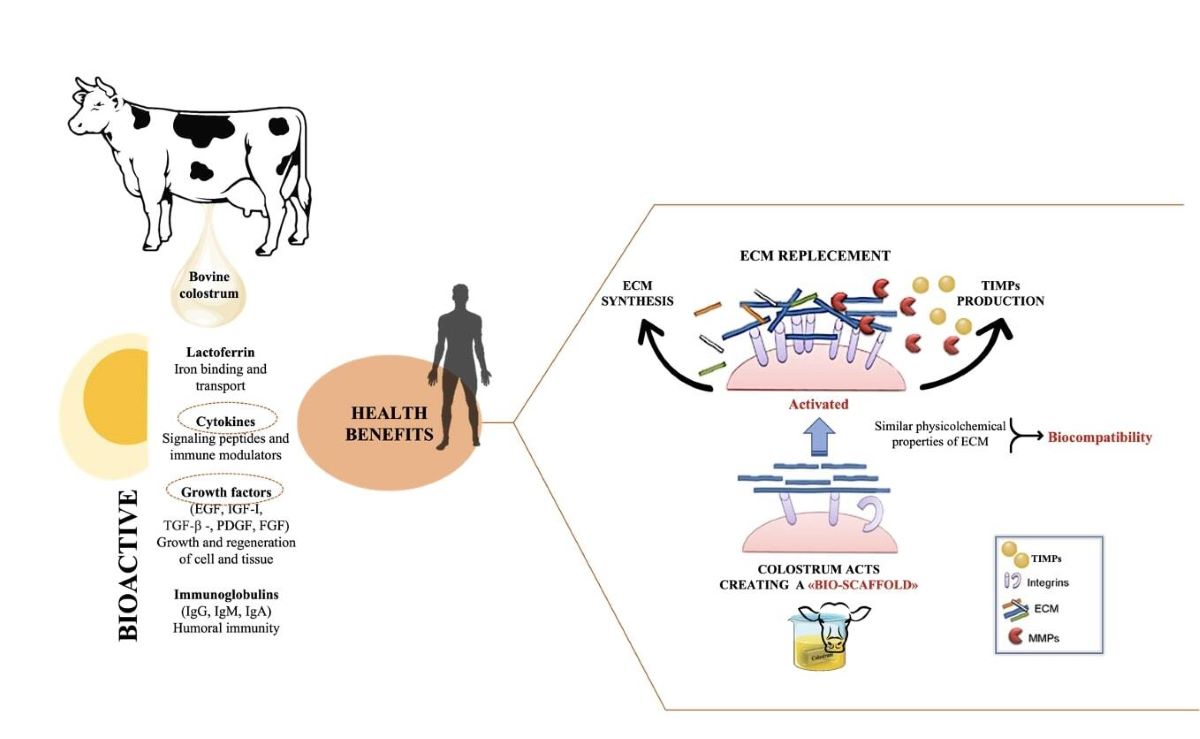
The aim of our research is to demonstrate that bioactive factors purified from bovine colostrum are like molecules normally present in the extracellular matrix and that if supplied to stressed tissues (such as in chronic wounds), they can provide the scaffolding necessary to activate the signal transduction mechanisms that activate the response of integrins. Bovine colostrum is a nutrient milk secretion containing bioactive compounds that support calf nutrition and immune development. Colostrum is profuse in bioactive compounds like immunoglobulins, growth factors, lysozymes, lactoferrin, lactoperoxidase, but also possesses elevated levels of fats, proteins, minerals and vitamins. Bovine colostrum is also rich in extracellular nanovesicles, such as exosomes, which protect bioactive components from degradation [15]. These bioactive molecules are also components of the ECM, and their mechanical role is fundamental in its remodeling [16,17,18]. In order to demonstrate our hypothesis, we carried out also a test to evaluate collagen contraction and measured the expression of collagen production and metalloproteinases-2, crucial elements for wound healing [7,19,20].
2. Materials and Methods
2.1. Colostrum Derivative Mixture Preparation
The colostrum derivative mixture (CDM) preparation was processed according to the procedure described by [21]. Bovine colostrum was collected from Holstein cows from 1 up to 6 h after parturition. This methodology, through micro and nano filtration, allows the elimination of casein, fats and other non-functional macromolecules including bovine immunoglobulins from the colostrum, and essentially isolates growth factors and cytokines identified with ELISA tests [21]. In all experiments, we used 2% colostrum derivative mixture because, our unpublished data, we have seen that this corresponds to concentration growth factors and cytokines normally present in the extracellular matrix.
2.2. Cell Lines
The Human Dermal Fibroblasts (HDF) (106-05A, Merck) were grown in Dulbecco’s Modified Eagle Medium (DMEM). Medium were supplemented with 10% fetal bovine serum. The medium was replaced every 2 days, starting when the cells reached 80% confluence. Cultures were maintained at 37°C in a humidified environment containing 5% CO2.
2.3. Collagen Contraction
To assess matrix contraction free-floating collagen lattice models were used according to [19]. The lattices were detached before addition of test substances. The reduction of the lattice area due to contraction was evaluated at 24 hours intervals for up to 96 hours.
2.4. Collagen Production
Matrix synthesis was determined over 7 days in the presence or absence of colostrum derivative mixture 2%. Secreted C-terminal propeptide of collagen type I (CICP) was measured in culture supernatant using an enzyme immunoassay kit (Metra Biosystem, Quidel Corporation).
2.5. Determination of MMP-2 Concentration
MMP-2 expression, constitutively secreted from fibroblasts, was determined with a commercially available quantitative ELISA test (Amersham Pharmacia Biotech). All procedures were performed according to the instructions of the manufacturer.
2.6. Statistical Analysis
Data were analyzed using the GraphPad Prism 5.0 program (GraphPad Software, La Jolla, CA, USA), using the analysis of variance (ANOVA) test and Tukey’s test. A p value ≤ 0.05 was used to identify statistically significant differences. Parametric correlation was calculated using Pearson correlation coefficient.
3. Results and Discussions
Tissues are not only made up of cells, but a significant part of their volume is formed by the extracellular space, occupied by an intricate network of macromolecules, whose three-dimensional organization is represented by the extracellular matrix. Biochemical analysis of the ECM reveals that it is composed of several proteins and polysaccharides, which aggregate in a compactly organized network connected to the surface of the cells that produced it and the surrounding ones [6,16,22]. ECM forms the bio-scaffold of our body and adapts to the environmental variables. The biomechanical properties of ECM are negatively affected by aging, but also during wound healing or restore skin in burning injuries [23]. Many computational models and experimental studies have revealed the important effects of cell-generated mechanical forces, forces acting upon cells, and mechanical characteristics of the extracellular matrix on cell morphology and function [23]. The microenvironment surrounding cells play a large role in directing cell behavior and the mechanical functions are important for both understanding cell behavior and excellent scaffolds designed. Cell can actively sense the mechanical properties of their surroundings by exerting contractile force, which can transmit between cell–matrix or cell–cell. Cells produce and can modify the organization of this ECM, which can vary widely in both composition and mechanical proprieties. Thus, these mechanical properties are a direct result of cellular activity, leading to the principle of dynamic reciprocity between the cell and its environment. But on cells can act mechanical forces exerted from ECM if as tissues are deformed, such as in wounds [24,25,26]. Our hypothesis is that the molecules extracted from colostrum (Table 1), identified with ELISA test according to [21], restore chemical/physical ECM environment, thus guaranteeing through a mechanical action the restoration of the wound due to the transduction of the signal activated by the integrins. Our hypothesis is supported by the results obtained with collagen contraction test that demonstrated exposure of fibroblast-populated collagen lattices to concentrations of colostrum derivative mixture significantly has activated contraction by day 7, compared with the control (Figure 1). These results are of great interest especially if correlated to the simultaneous results of ELISA test made during collagen contraction. Tests demonstrated that fibroblasts actively have secreted CICP during lattice contraction up to 7 days (Figure 2) and inhibited MMP-2 expression (Figure 3) in the samples treated with the 2% colostrum derivative mixture.
Statistical analysis using Pearson correlation coefficient, confirmed a significant positive linear correlation between collagen contraction and collagen production during collagen contraction from fibroblasts (r = 0.98, p < 0.001) and a significant negative linear correlation between collagen contraction and proenzyme MMP-2 production during collagen contraction (r = 0.99, p < 0.001) (Figure 4).
Contraction and remodeling of the extracellular matrix are essential processes during wound healing [27,28]. At the center of these two phenomena are fibroblasts, which not only produce and secrete extracellular matrix proteins but can also reorganize them through mechanical interactions [29,30]. Collagen is the predominant structural protein in the ECM providing not only tensile strength but also play a role in cells adhesion and migration [31]. Matrix metalloproteinases (MMPs) are a family of proteolytic enzymes capable of degrading various structural components of the matrix [32,33,34,35,36,37]. They therefore play a fundamental role in various physiological and pathological processes including wound healing [17,28]. It is now known that the biomechanical properties of the ECM contribute to physiological development of cells and tissues [24].
The elasticity of the ECM allows to perceive external forces and, therefore, provides an important environmental input signal that determines the cell’s response. Indeed, the adhesion complex, which consists of integrins and a multicomplex of adapters and signaling proteins, can be considered as a mechanosensor that connects the actomyosin cytoskeleton with the ECM [38]. Many of the adhesion components undergo conformational changes that determine functional consequences in the response to applied force. Cytoskeleton together with nuclear matrice, nuclear envelope and chromatin, they constitute a mechano-sensing system that determines how cells react to forces transmitted by the ECM [38,39,40,41,42]. Many of the focal adhesion components undergo conformational changes that determine functional consequences in the response to applied force. Together with the cytoskeleton and nuclear matrices, nuclear envelope, and chromatin, they constitute a sophisticated mechanosensing machinery that determines how cells react to forces transmitted by the ECM, which regulates various essential cellular behaviors, including cell fate determination, differentiation, and cell function [38,39,40,41,42]. The different properties of the ECM are not independent of each other but influence each other. Therefore, when the ECM increases in stiffness, as, for example, in pathological conditions, its biomechanical properties change, and cells respond by exerting markedly different types of force. Furthermore, the stiffening of the matrix also determines a change in the other physical properties of the ECM and, consequently, directly modifies the cellular ability to migrate. ECM, constantly undergoing restructuring in different tissues, is highly dynamic [42]. ECM dynamics can arise from changes in the amount or composition of ECM, due to altered synthesis or degradation of one or more components. Alternatively, ECM dynamics may show no changes in the composition of its components, but instead involve only the transformation of its components, into spatially organized structures, thanks to covalent and non-covalent bonds [6,22].
Finally, one of the most important features of cell-ECM interactions is reciprocity [43]. Cells constantly make, degrade or rearrange components of the ECM to modify it one or more properties; but, since the ECM regulates the behavior of cells, any change in it, as result of cellular activities, will in turn influence adjacent cells and modify their behaviors. This feedback regulatory mechanism between cells and the ECM allows cells and tissues to rapidly adapt to the changes of environment [6].
4. Conclusions
Wounds heal through coordinated action of fibroblast-mediated extracellular matrix (ECM) deposition, ECM remodeling, and wound contraction. For these processes, are fundamental the mechanical signals between ECM and cells. In this paper, we demonstrated that biomolecules extracted and purified of bovine colostrum can restore chemical/physical ECM environment, reestablish the mechanical properties of ECM and the anisotropy characteristics of the damaged tissue. This process activates transduction of the signal by the integrins and reactivates the ability of the fibroblast to synthesize new collagen, inducing the autologous mechanisms of repair and remodeling of damaged connective tissue. Fibroblasts, in fact, can be able to generate tensile forces as well as receive them; these contraction forces of the fibroblast are indispensable for wound resolution processes.
Author Contributions
Conceptualization, M.V.B.; methodology, M.V.B..; software, G.F.; formal analysis, F.C., G.G., G.I., and S.P.; investigation, F.C., G.G., G.I., and S.P.; data curation, M.V.B.; writing—original draft preparation, M.V.B.; writing—review and editing, G.F. and M.V.B.; supervision, M.V.B.; funding acquisition, M.V.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
G.F. thanks the Ph.D. program FSE Notice 1/2021. F.C., G.G., G.I, S.P. thanks the Ph.D. program PNRR—D.M. 117/2023.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mierke, C.T. Bidirectional Mechanical Response Between Cells and Their Microenvironment. Front Phys 2021, 9, 749830. [Google Scholar] [CrossRef]
- Xie, N.; Xiao, C.; Shu, Q.; Cheng, B.; Wang, Z.; Xue, R.; Wen, Z.; Wang, J.; Shi, H.; Fan, D.; Liu, N.; Xu, F. Cell response to mechanical microenvironment cues via Rho signaling: From mechanobiology to mechanomedicine. Acta Biomater 2023, 159, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Jansen KA, Donato DM, Balcioglu HE, Schmidt T, Danen EH, Koenderink GH. A guide to mechanobiology: Where biology and physics meet. Biochim Biophys Acta. 2015, 1853, 3043–3052. [Google Scholar] [CrossRef]
- Boyd, D.F.; Thomas, P.G. Towards integrating extracellular matrix and immunological pathways. Cytokine 2017, 98, 79–86. [Google Scholar] [CrossRef]
- Di, X.; Gao, X.; Peng, L.; Ai, J.; Jin, X.; Qi, S.; Li, H.; Wang, K.; Luo, D. Cellular mechanotransduction in health and diseases: from molecular mechanism to therapeutic targets. Sig Transduct Target Ther 2023, 8, 282. [Google Scholar]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb Perspect Biol 2011, 3, a005058. [Google Scholar] [CrossRef] [PubMed]
- Caley, M.P.; Martins, V.L.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv Wound Care (New Rochelle) 2015, 4, 225–234. [Google Scholar] [CrossRef]
- Zhao, X.; Li, Q.; Guo, Z.; Li, Z. Constructing a cell microenvironment with biomaterial scaffolds for stem cell therapy. Stem Cell Res Ther 2021, 12, 583. [Google Scholar] [CrossRef]
- Kozaniti, F.K.; Manara, A.E.; Kostopoulos, V.; Mallis, P.; Michalopoulos, E.; Polyzos, D.; Deligianni, D.D.; Portan, D.V. Computational and Experimental Investigation of the Combined Effect of Various 3D Scaffolds and Bioreactor Stimulation on Human Cells’ Feedback. Appl Biosci 2023, 2, 249–277. [Google Scholar] [CrossRef]
- Caliari, S.R.; Harley, B.A. The effect of anisotropic collagen-GAG scaffolds and growth factor supplementation on tendon cell recruitment, alignment, and metabolic activity. Biomaterials 2011, 32, 5330–5340. [Google Scholar] [CrossRef]
- Harmansa, S.; Erlich, A.; Eloy, C.; Zurlo, G.; Lecuit, T. Growth anisotropy of the extracellular matrix shapes a developing organ. Nat Commun 2023, 14, 1220. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.; Vyas, V.; Dhara, S.; Chowdhury, A.; Barui, A. Anisotropy Properties of Tissues: A Basis for Fabrication of Biomimetic Anisotropic Scaffolds for Tissue Engineering. J Bionic Eng 2019, 16, 842–868. [Google Scholar] [CrossRef]
- Reid, J.A.; Dwyer, K.D.; Schmitt, P.R.; Soepriatna, A.H.; Coulombe, K.L.; Callanan, A. Architected fibrous scaffolds for engineering anisotropic tissues. Biofabrication 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Ross, T.D.; Coon, B.G.; Yun, S.; Baeyens, N.; Tanaka, K.; Ouyang, M.; Schwartz, M.A. Integrins in mechanotransduction. Curr Opin Cell Biol 2013, 25, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.; Atalla, H.; Karrow, N.; Mallard, B.A. The bioactivity of colostrum and milk exosomes of high, average, and low immune responder cows on human intestinal epithelial cells. J Dairy Sci 2021, 104, 2499–2510. [Google Scholar] [CrossRef]
- Yue, B. Biology of the extracellular matrix: an overview. J Glaucoma 2014, 23(8 Suppl 1), S20-3. [Google Scholar] [CrossRef]
- Diller, R.B.; Tabor, A.J. The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review. Biomimetics (Basel) 2022, 7, 87. [Google Scholar] [CrossRef]
- Kuehlmann, B.; Bonham, C.A.; Zucal, I.; Prantl, L.; Gurtner, G.C. Mechanotransduction in Wound Healing and Fibrosis. J Clin Med 2020, 9, 1423. [Google Scholar] [CrossRef]
- Daniels, J.T.; Cambrey, A.D.; Occleston, N.L.; Garrett, Q.; Tarnuzzer, R.W.; Schultz, G.S.; Khaw, P.T. Matrix metalloproteinase inhibition modulates fibroblast-mediated matrix contraction and collagen production in vitro. Invest Ophthalmol Vis Sci 2003, 44, 1104–1110. [Google Scholar] [CrossRef]
- Singh, D.; Rai, V.; Agrawal, D.K. Regulation of Collagen I and Collagen III in Tissue Injury and Regeneration. Cardiol Cardiovasc Med 2023, 7, 5–16. [Google Scholar] [CrossRef]
- Sacerdote, P.; Mussano, F.; Franchi, S.; Panerai, A.E.; Bussolati, G.; Carossa, S.; Bartorelli, A.; Bussolati, B. Biological components in a standardized derivative of bovine colostrum. J Dairy Sci 2013, 96, 1745–1754. [Google Scholar] [CrossRef]
- Hu, M.; Ling, Z.; Ren, X. Extracellular matrix dynamics: tracking in biological systems and their implications. J Biol Eng 2022, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Franchi, M.; Piperigkou, Z.; Mastronikolis, N.S.; Karamanos, N. Extracellular matrix biomechanical roles and adaptation in health and disease. FEBS J 2024, 291, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Holle, A.W.; Young, J.L.; Van Vliet, K.J.; Kamm, R.D.; Discher, D.; Janmey, P.; Spatz, J.P.; Saif, T. Cell-Extracellular Matrix Mechanobiology: Forceful Tools and Emerging Needs for Basic and Translational Research. Nano Lett 2018, 18, 1–8. [Google Scholar] [CrossRef]
- Arnold, M.; Cavalcanti-Adam, E.A.; Glass, R.; Blümmel, J.; Eck, W.; Kantlehner, M.; Kessler, H.; Spatz, J.P. Activation of integrin function by nanopatterned adhesive interfaces. Chemphyschem 2004, 5, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Boudreau, A.; Bissell, M.J. Tissue architecture and function: dynamic reciprocity via extra- and intra-cellular matrices. Cancer Metastasis Rev 2009, 28, 167–176. [Google Scholar] [CrossRef]
- Olczyk, P.; Mencner, Ł.; Komosinska-Vassev, K. The role of the extracellular matrix components in cutaneous wound healing. Biomed Res Int 2014, 2014, 747584. [Google Scholar] [CrossRef]
- Potekaev, N.N.; Borzykh, O.B.; Medvedev, G.V.; Pushkin, D.V.; Petrova, M.M.; Petrov, A.V.; Dmitrenko, D.V.; Karpova, E.I.; Demina, O.M.; Shnayder, N.A. The Role of Extracellular Matrix in Skin Wound Healing. J Clin Med 2021, 10, 5947. [Google Scholar] [CrossRef]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv Wound Care (New Rochelle) 2016, 5, 119–136. [Google Scholar] [CrossRef]
- Kular, J.K.; Basu, S.; Sharma, R.I. The extracellular matrix: Structure, composition, age-related differences, tools for analysis and applications for tissue engineering. J Tissue Eng 2014, 5, 2041731414557112. [Google Scholar] [CrossRef]
- Sun, B. The mechanics of fibrillar collagen extracellular matrix. Cell Rep Phys Sci 2021, 2, 100515. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.E.; Parks, W.C. Metalloproteinases and their inhibitors: regulators of wound healing. Int J Biochem Cell Biol 2008, 40, 1334–1347. [Google Scholar] [CrossRef] [PubMed]
- Lukes, A.; Mun-Bryce, S.; Lukes, M.; Rosenberg, G.A. Extracellular matrix degradation by metalloproteinases and central nervous system diseases. Mol Neurobiol 1999, 19, 267–284. [Google Scholar] [CrossRef]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuña, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int J Mol Sci 2020, 21, 9739. [Google Scholar] [CrossRef]
- Burbridge, M.F.; Cogé, F.; Galizzi, J.P.; Boutin, J.A.; West, D.C.; Tucker, G.C. The role of the matrix metalloproteinases during in vitro vessel formation. Angiogenesis 2002, 5, 215–226. [Google Scholar] [CrossRef]
- Lombard, C.; Saulnier, J.; Wallach, J. Assays of matrix metalloproteinases (MMPs) activities: a review. Biochimie 2005, 87, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Agren, M.S.; Mirastschijski, U.; Karlsmark, T.; Saarialho-Kere, U.K. Topical synthetic inhibitor of matrix metalloproteinases delays epidermal regeneration of human wounds. Exp Dermatol 2001, 10, 337–348. [Google Scholar] [CrossRef]
- Wong, V.W.; Akaishi, S.; Longaker, M.T.; Gurtner, G.C. Pushing back: wound mechanotransduction in repair and regeneration. J Invest Dermatol 2011, 131, 2186–2196. [Google Scholar] [CrossRef] [PubMed]
- DuFort, C.C.; Paszek, M.J.; Weaver, V.M. Balancing forces: architectural control of mechanotransduction. Nat Rev Mol Cell Biol 2011, 12, 308–319. [Google Scholar] [CrossRef]
- Ricca, B.L.; Venugopalan, G.; Fletcher, D.A. To pull or be pulled: parsing the multiple modes of mechanotransduction. Curr Opin Cell Biol 2013, 25, 558–564. [Google Scholar] [CrossRef]
- Saraswathibhatla, A.; Indana, D.; Chaudhuri, O. Cell-extracellular matrix mechanotransduction in 3D. Nat Rev Mol Cell Biol 2023, 24, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Wei, X.; Kang, H.; Jiang, H.; Chu, Z.; Lin, Y.; Hou, Y.; Wei, Q. Static and Dynamic: Evolving Biomaterial Mechanical Properties to Control Cellular Mechanotransduction. Adv Sci (Weinh) 2023, 10, e2204594. [Google Scholar] [CrossRef] [PubMed]
- Urciuolo, F.; Imparato, G.; Netti, P.A. In vitro strategies for mimicking dynamic cell–ECM reciprocity in 3D culture models. Front Bioeng Biotechnol 2023, 11, 1197075. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Graphical representation of collagen contraction. A. Collagen gel contraction over 96 h at 24 h time intervals. B. Fold change in collagen gel contraction at 96 h. Asterisk denotes the degree of significance between results: *p < 0.01.
Figure 1.
Graphical representation of collagen contraction. A. Collagen gel contraction over 96 h at 24 h time intervals. B. Fold change in collagen gel contraction at 96 h. Asterisk denotes the degree of significance between results: *p < 0.01.
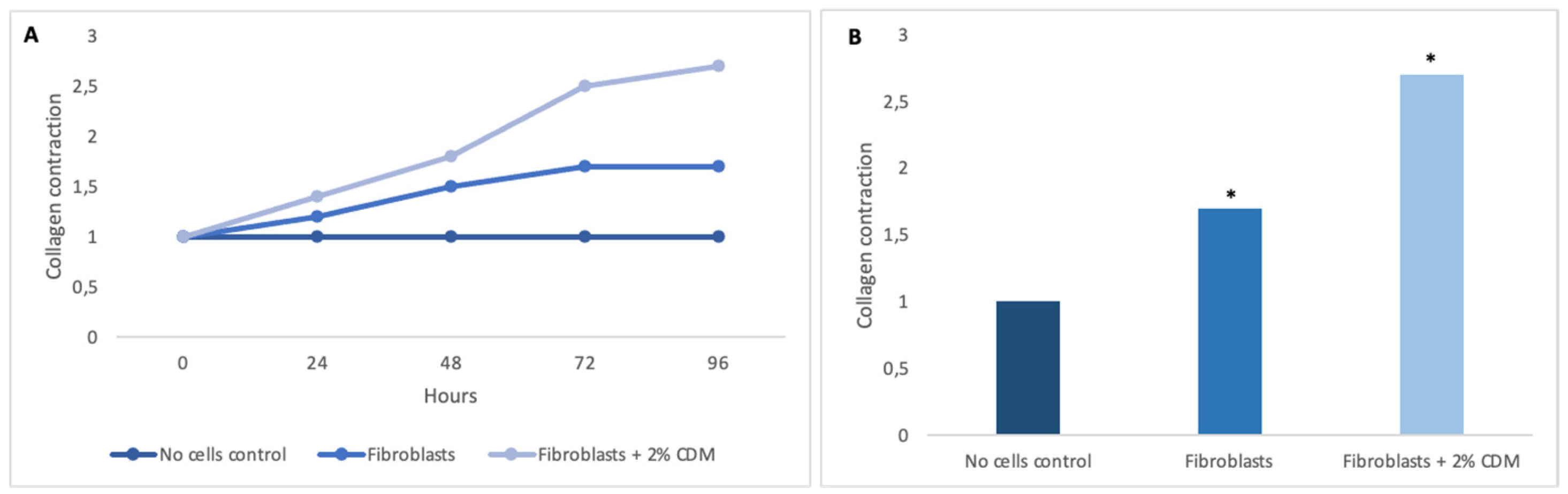
Figure 2.
Graphical representation of collagen production during collagen contraction. Fibroblasts actively has secreted CICP during lattice contraction up to 7 days. In control samples, secretion of CICP has increased up to 3 days and after was reduced. Asterisk denotes the degree of significance between results: **p < 0.001.
Figure 2.
Graphical representation of collagen production during collagen contraction. Fibroblasts actively has secreted CICP during lattice contraction up to 7 days. In control samples, secretion of CICP has increased up to 3 days and after was reduced. Asterisk denotes the degree of significance between results: **p < 0.001.
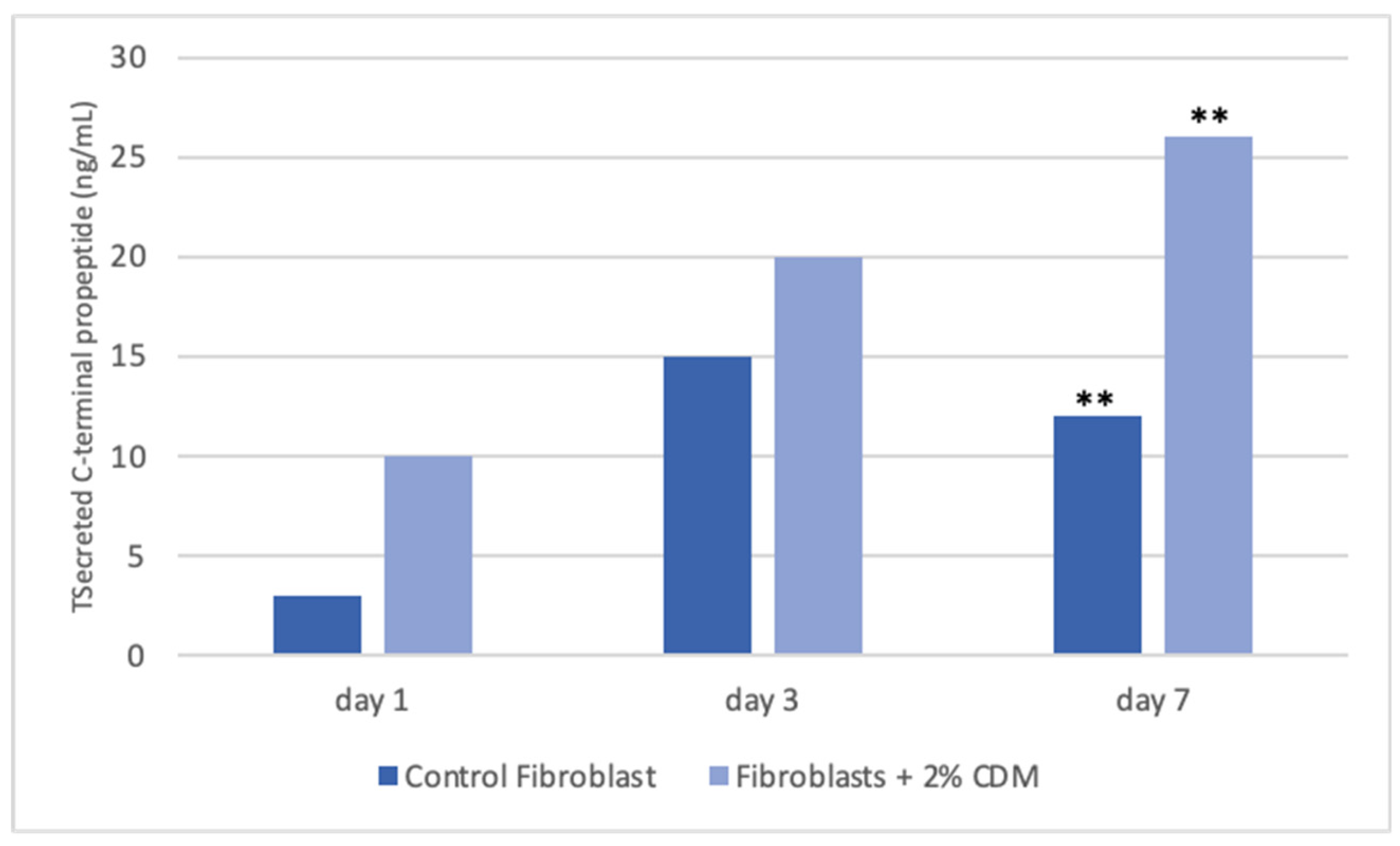
Figure 3.
Graphical representation of proenzyme MMP-2 production during collagen contraction. Asterisk denotes the degree of significance between results: ***p < 0.0001.
Figure 3.
Graphical representation of proenzyme MMP-2 production during collagen contraction. Asterisk denotes the degree of significance between results: ***p < 0.0001.
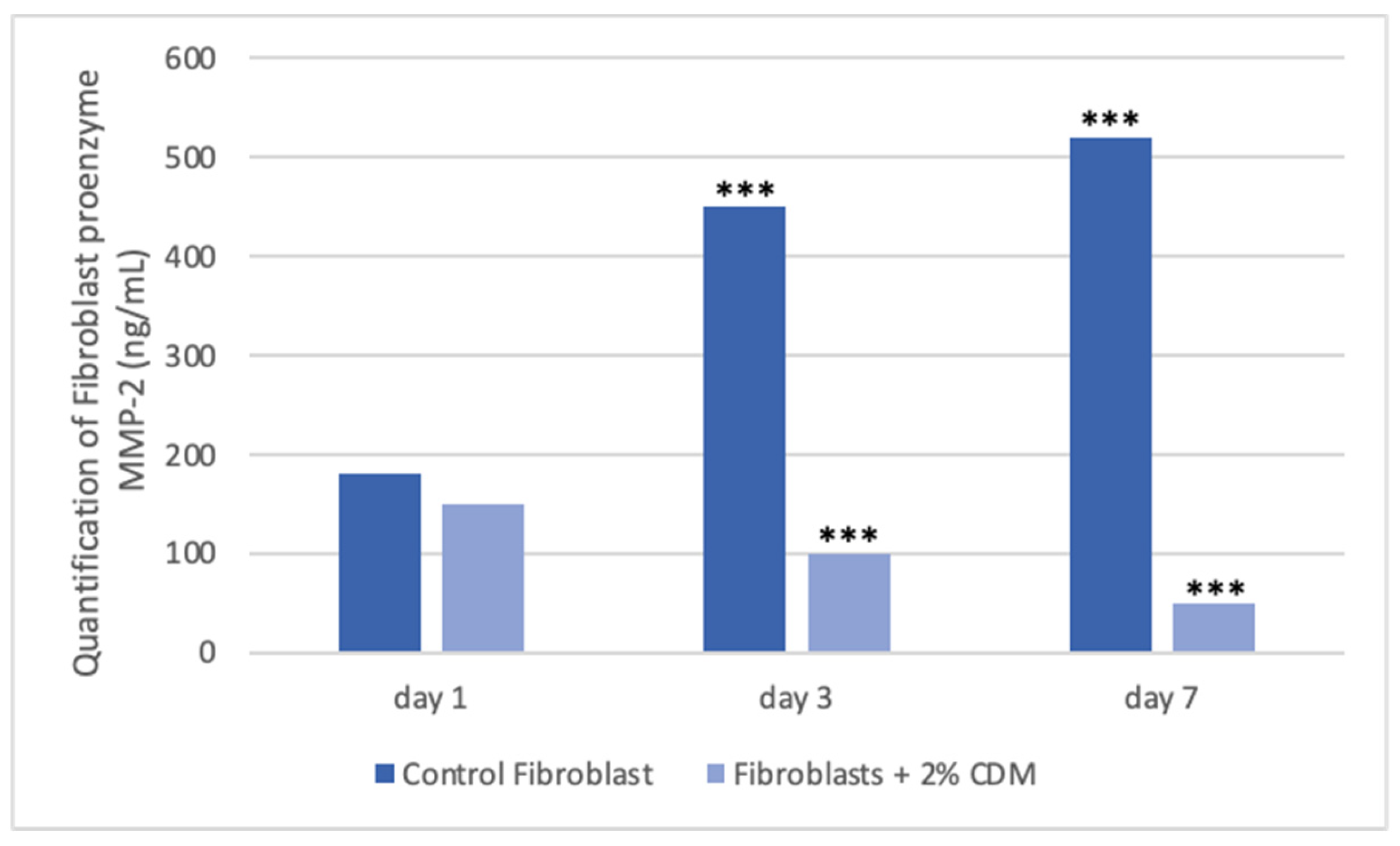
Figure 4.
Scatter Plot. A, strong positive linear correlation and B, strong negative linear correlation, indicated by Pearson coefficient values (r).
Figure 4.
Scatter Plot. A, strong positive linear correlation and B, strong negative linear correlation, indicated by Pearson coefficient values (r).
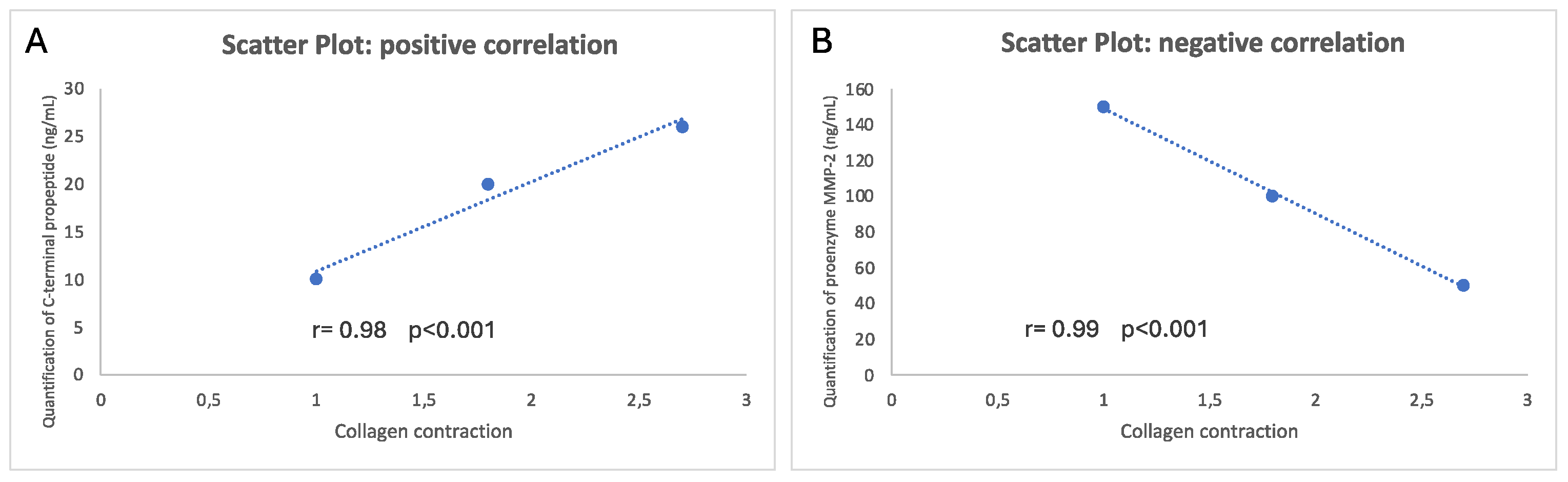

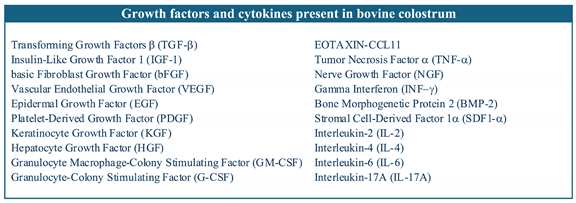
Table 1.
Growth factors and cytokines present in bovine colostrum identified with Elisa test.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



