
Submitted:
17 August 2024
Posted:
19 August 2024
You are already at the latest version
Abstract
Toxicity mechanisms of per- and polyfluoroalkyl substances (PFAS), a chemical class present in diverse ecosystems, are not well characterized in aquatic species. Perfluorooctanesulfonamide (PFOSA, C8H2F17NO2S) is a common precursor of perfluorooctane sulfonic acid (PFOS), a long-chain PFAS. Here, we assessed sub-lethal endpoints related to development, oxidative stress, locomotor activity, and gene expression in zebrafish embryos and larvae following continuous exposure to PFOSA beginning at 6 hours post fertilization (hpf). PFOSA decreased survival in fish treated with 1 µg/L PFOSA, however the effect was modest relative to the controls (difference of 10%). Exposure up to 10 µg/L PFOSA did not affect hatch rate, nor did it induce reactive oxygen species in 7-day-old larvae fish. However, locomotor activity of larval fish treated with 100 µg/L PFOSA was reduced relative to the solvent control. Transcripts related to oxidative stress response and apoptosis were measured and BCL2 associated X, apoptosis regulator (bax), cytochrome c, somatic (cycs), catalase (cat), superoxide dismutase 2 (sod2) were induced with high concentrations of PFOSA. Genes related to neurotoxicity were also measured and transcript levels of acetylcholinesterase (ache), elav-like RNA binding protein 3 (elavl3), growth associated protein 43 (gap43), synapsin II (syn2a), and tubulin 3 (tubb3) levels were all increased in larval fish with higher PFOSA exposure. This study contributes to our knowledge regarding the relative toxicity of PFOSA to fish and contributes data to future risk assessment strategies of these concerning, persistent environmental pollutants.
Keywords:
perfluorinated chemicals
; aquatic toxicology
; neurotoxicity
; behavior
1. Introduction
Since the late 1930s, perfluoroalkyl substances (PFAS) have been used in a variety of consumer and commercial goods, including textiles, surfactants, and food packaging materials (Sant et al., 2019). These compounds are comprised of short or long carbon chains, where short-chain PFAS are considered less harmful than long-chain PFAS [1]. According to the Danish Environmental Protection Agency, as long-chain PFAS persist in the environment due to their chemical makeup, their production usage has been gradually phased out since the early 2000s [2]. Perfluorooctanesulfonamide (PFOSA, C8H2F17NO2S) is a common precursor of perfluorooctane sulfonic acid (PFOS), a long-chain PFAS [3]. Most studies to date focus on the presence and toxicity of other commonly used PFAS, such as PFOS or perfluorooctanoic acid (PFOA) [4-6]. Currently, toxicity data regarding the precursor PFOSA are lacking.
There are a few studies that report on the environmental presence of PFOSA. Konwick et al. (2008) found that PFOSA ranged from 162-283 ng/L in the Conasauga River, Georgia, United States [7]. Additionally, other studies report that PFOSA can range from 0.09-20,000 µg/kg in surface-soil, 0.07-2500 µg/kg in subsurface-soil, 15 µg/L in surface water, and 12 µg/L in groundwater across various testing sites worldwide [8,9]. In regards to PFOSA within fish tissues, Fair et al. (2019) measured different PFAS within edible fish species from South Carolina, United States [10]. In whole fish, the average relative percent of PFOSA in mullet, spot, croaker, red drum, and seatrout was 2.04, 3.81, 3.64, 3.12, and 4.66%, respectively, whereas, in fillets, the same species had an average relative percent of 1.44, 2.03, 0.70, 2.18, 4.65, and 1.79%, respectively. PFOSA was also found to range between 0.105-16.4 ng/mL in serum across various fish species, including common carp (Cyprinus carpio), crucian carp (Carassius auratus), tilapia (Oreochromis niloticus), and leather catfish (Clarias lazera) [11]. However, the mechanism of uptake, metabolism, and toxicity of PFOSA are relatively unknown for aquatic species. One study reports that the half-life of PFOSA in rainbow trout (Oncorhynchus mykiss) is 6.0 ± 0.4 days following a 30-day dietary exposure to 10 µg/g wet weight PFOSA and a 30-day depurination period [12]. Thus, PFOSA is measurable in fish tissues and may pose a health risk to wildlife and humans.
According to studies, adverse morphological and physiological effects in aquatic organisms are potential consequences associated with the environmental presence of long-chain PFAS [13,14]. For example, studies show that PFOSA exerts cardiotoxicity in zebrafish (Danio rerio). Exposure to 0.1-100 µg/L PFOSA has been reported to reduce cardiac output, heart rate, stroke volume and reduce cardiac vasoconstriction-related genes[15]. PFOSA has also been reported to significantly increase sinus venosus and bulbus arteriosus distance at 10 and 100 µg/L [15,16]. Other studies report that exposure to PFOSA can induce hepatic and renal toxicity in zebrafish [17,18]; however, limited studies investigate the neurotoxic potential of PFOSA in developing fish. Consequently, the objectives of this study were to evaluate the neurotoxicity potential of PFOSA. To do this, we measured developmental endpoints, reactive oxygen species (ROS), locomotor behavior, and genes related to oxidative damage response, apoptosis, and neurotoxicity as indicators of central nervous system damage. We hypothesized that neurotoxicity endpoints would reflect dose response increased in PFOSA exposure, suggesting adverse effects on the nervous system.
2. Materials and Methods
2.1. Chemical Preparation
Perfluorooctanesulfonamide (PFOSA, (1,1,2,2,3,3,4,4,5,5,6,6,7,7,8,8,8-heptadecafluorooctane-1-sulfonamide Perfluorooctane sulfonamide) (CAS Number: 754-91-6, purity > 95%) was purchased from Fisher Scientific (Cat# AC459640010). Stock solutions of PFOSA were prepared in dimethyl sulfoxide (DMSO, Dimethyl Sulfoxide, CAS 67-68-5, Sigma Aldrich) and added to embryo rearing media (ERM) containing the zebrafish embryos. Recipes for ERM can be found in [19]. Exposure solutions were prepared daily to yield final nominal concentrations of 0.1, 1, 10, and 100 µg/L PFOSA with a final concentration < 0.1% v/v DMSO in experimental treatments.
2.2. Maintenance and Egg Production of Zebrafish
Adult zebrafish (AB x Tübingen, Danio rerio) were raised in a flow-through Pentair system in the Cancer-Genetics Research Center at the University of Florida as outlined previously [20,21]. The University of Florida maintains a breeding colony for research and outbreeds extraneous fish to maintain high genetic diversity. Rearing and staging of zebrafish embryos followed that described by [22]. Adult fish were fed Zeigler Zebrafish Diet ad libitum. Zebrafish at 6 months of age were selected at random from a breeding stock and placed into a shallow water breeding tank the night before embryo collection (2 males and 2 females). A divider was used to separate the males and females overnight and it was removed at 8:00 am when the facility lights turned on. Typically, three or four breeding tanks are prepared the night before to maximize eggs, and adult fish are bred once every 1 to 2 weeks. Embryos were rinsed with saline phosphate buffer (PBS) and ERM several times once they were collected and they were sorted under a light microscope in the late morning (around 4 hours post-fertilization, hpf). All experiments were conducted at pH = 7.2 ± 0.5, a conductivity value of 600 ± 100 μS/cm, light/dark cycle of 14:10 h, dissolved oxygen concentration ~80% air saturation, and temperature of 27 ± 1 °C. Institutional Animal Care and Use Committee of University of Florida approved all experiments (UFIACUC202300000140).
2.3. PFOSA Exposure Regime
Fertilized and normally developing eggs were selected at ∼6 h post-fertilization (hpf) using a dissecting microscope. Zebrafish eggs were assigned in random fashion into experimental groups [ERM, 0.1% DMSO, or one dose of 0.1, 1, 10, 100 µg/L PFOSA]. Four independent experiments were conducted using embryos that were generated from separate breeders of fish. For each experiment, there were 5 to 6 replicate glass beakers for each experimental group containing 20-30 embryos and 10 mL of embryo rearing media (ERM). Following addition of chemicals to the water, the glass beakers were placed into an incubator that was maintained at 27 ± 1 °C. Each day, data related to mortality, deformities, hatch, as well as images using an EVOS™ FL Auto Imaging System (ThermoFisher Scientific, USA) were collected. Deformity assessments included the presence of spinal lordosis and edema (yolk sack/pericardial) were noted over the duration of the exposure. Exposure solutions were prepared fresh daily from a stock stored at -20°C in glass amber vials and water was renewed every day with a 90% water change.
2.4. Reactive Oxygen Species
Six hpf embryos were obtained immediately after fertilization and treated as per above for 7 days in the ERM with designated concentrations of PFOSA. Embryos were rinsed three times in ERM and assessed under a light microscope. Fertilized embryos were evenly distributed using sterile micropipettes into sterile 25 mL glass beakers containing the assigned concentration of ERM, 0.1% DMSO, 0.1, 1, 10 µg/L PFOSA (n=5 beakers of 10 fish each /treatment) in a 10 mL volume. Embryos were maintained in the controlled environment of an incubator at 27 ± 1.0°C. Media changes were conducted every 24 hours with new sterile ERM or renewed PFOSA in ERM. Following a 7-day exposure, larvae were quickly transferred from beakers into 1.7 mL microcentrifuge tubes, homogenized in 200 µL of ice-cold PBS, and centrifuged at 12,000 g for 20 min at 4° C. Following centrifugation, 20 µL of the supernatant was transferred to a black fluorescence 96-well plate and incubated for 5 minutes at room temperature. After incubation, 8.3 µL of 1 mg/mL 2′,7′-Dichlorofluorescin Diacetate (Calbiochem, Millipore Sigma, CAS 4091–99, or H2-DCFDA) dissolved in DMSO and 200 µL of PBS were added to each well. Then, the plate was incubated in the dark for 30 minutes at 37 ± 1.0 °C. Following this, excitation at 485 nm and emission at 520 nm were recorded using a Synergy™ H4 Hybrid Multi-Mode Microplate Reader. Total protein using a BCA assay (Thermo Scientific) was measured to express ROS as normalized signal intensity/(µg/mL) protein.
2.5. Visual Motor Response Test
Experiments were performed to test the dark photokinesis response in larvae. Fish were exposed continuously for 7 days with 90% daily water changes with PFOSA as described above and assessed for locomotor activity behavior at a temperature of 27 ± 1 °C and photoperiod pf 14:10 h. In each trial, zebrafish embryos at 6 hpf were randomly assigned to an experimental group of either ERM, 0.1% DMSO, or 0.1, 1, 10, or 100 µg/L PFOSA (5 beakers per treatment). Each group contained 20 zebrafish embryos and 10 mL of ERM. In mid-afternoon on the 7th day, 2 normally developed larva were selected from each replicate beaker and placed in a random fashion into a clear 96-well plate (N=10 fish per treatment/per experiment). Each well contained 200 μL of ERM. The 96-well plate was placed into DanioVision™ Observation Chamber (Noldus Information Technology, Leesburg, VA) with an infrared analog camera (25 frames/ second) to track the activities of zebrafish larvae. The assay proceeded as per our previous methods [23]. Data were analyzed independently for each trial, and total distance moved was used as an indicator of overall locomotor activity. Data were also analyzed by normalizing each of the three independent runs using a relative value = 1 for the solvent control group, and then relative data was combined into a single graph representing 30 larvae per treatment.
2.6. Real-Time PCR
Zebrafish larvae at 6 hpf were exposed to either ERM, 0.1% DMSO, 0.1, 1, or 10 µg/L PFOSA. Each beaker contained 10-15 embryos and exposure conditions were maintained as that above. Following the 7-day exposure period, larvae were pooled within a beaker, subjected to liquid nitrogen, and placed at −80 °C for RNA extraction. Extraction of RNA from larvae pools was performed using 500 mL TRIzol® Reagent (Life Technologies, Carlsbad, CA, USA) as per manufacturer’s protocol. Samples were DNase treated with DNA TURBO (Ambion). DNase treated samples were assessed for quality using the 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). The mean RIN value for RNA was >7 and concentrations ranged approximately 50-200 ng/µl sample. The cDNA synthesis was performed using ~500 ng of column purified RNA using iScript (BioRad) following the manufacturer’s protocol in a final sample volume of 15 µL. Once prepared, samples were placed into a T100™ Thermal Cycler (BioRad, USA). The cDNA was generated using the following steps: 25 °C for 5 min, 42 °C for 30 min, 85 °C for 5 min, and 4 °C for 5 min. Prior to real-time PCR, cDNA stocks were diluted 1:25 in RNAse-DNase free water. The no reverse transcriptase (NRT) controls were prepared in the same way as above without enzyme using 3 randomly selected RNA samples.
Real-time PCR was performed using the CFX Connect™ Real-Time PCR Detection System (BioRad) with SsoFast™ EvaGreen® Supermix (BioRad, Hercules, CA, USA), 200-300 nM of each forward and reverse primer, and 3.33 µL of diluted cDNA. The two-step thermal cycling parameters were as follows: initial 1-cycle Taq polymerase activation at 95 °C for 30 s, followed by 95 °C for 5 s, and 60 °C primer annealing temperature for 5 s. After 40 cycles, a dissociation curve was generated, starting at 65.0 and ending at 95.0°C, with increments of 0.5 °C every 5 s. Primers used in this study were obtained from published literature [24-33](Supplemental Table S1). Two housekeeping genes (ribosomal subunit 18, rps18, and beta actin, b-actin) were used to normalize expression levels of all target genes. Target genes included acetylcholinesterase (ache), BCL2 Associated X Apoptosis Regulator (bax), BCL2 Apoptosis Regulator (bcl2), catalase (cat), caspase 3 (caspase3), ELAV like RNA binding protein 3 (elavl3), growth associated protein 43 (gap43), glial fibrillary acidic protein (gfap), heat shock protein 70 (hsp70), mesencephalic astrocyte derived neurotrophic factor (manf), myelin basic protein (mbp), nestin (nestin), tumor protein 53 (p53), sonic hedgehog signaling molecule (shha), superoxide dismutase 1 (sod1) (Cu/Zn SOD), superoxide dismutase 2 (sod2) (Mn SOD), synapsin IIa (syn2a), and tubulin. Normalized expression was obtained for each target gene using CFX Manager™ software (v3.1) (baseline subtracted) and the Cq method was employed. The qPCR analysis included 3 NRT samples and 1 NTC sample. Negative controls indicated that RNA column purification and DNase treatment sufficiently removed gDNA. Sample sizes ranged from 4 to 6 for gene expression analysis. All primers used in the qPCR analysis amplified one product, indicated by a single melt curve.
2.7. Statistical Analysis
All data were compared to the solvent control (DMSO group). A Log-rank test (Mantel-Cox) was employed to evaluate survival data. Data for hatch time was evaluated using a One-Way ANOVA at each time point. Levels of ROS and relative mRNA levels were first log(10) transformed following a Shapiro-Wilk test for normality. Group mean differences were then tested using a One-Way ANOVA (Dunnett’s multiple comparisons test). A simple linear regression was also conducted on the gene expression data to determine whether expression varied with concentration. Because there was no difference in expression between the ERM and DMSO group, these two experimental groups were combined for the regression as a “control” or baseline group. For the VMR, distance moved for larval fish in each treatment across the three independent experiments were binned into a single graph, but each individual run is shown in Supplemental Figures. The distance moved in the DMSO group was normalized to a value of 1, and all treatments were compared relative to this group. A Kruskal-Wallis test followed by a Dunn’s multiple comparisons test was used to evaluate differences in locomotor activity, which was analyzed as discrete temporal units (light and dark sections to corresponding control group). Data are presented as mean ± S.D. Significance of difference was determined using a threshold of P < 0.05. Statistics, and graphing, was performed using GraphPad V9.3 (La Jolla, CA, USA).
3. Results
3.1. Survival, Hatch Rate, and Deformity
After zebrafish were exposed to several concentrations of PFOSA for a period of 7 days, intriguingly only fish treated with 1 µg/L PFOSA exhibited significant decreased survival [Chi square = 31.14, df = 5, P value <0.0001] (Figure 1) relative to the DMSO control group. Hatch rate was increased in fish treated with 0.1 and 10 µg/L PFOSA as all embryos were hatched at 2 dpf compared to the other treatment groups (P<0.05) (Figure 2). There was no significant deformities observed; however, the few zebrafish that did have deformities (less than 3%) had caudal tail malformations.
3.2. Reactive Oxygen Species
We assessed the effects of PFOSA on ROS induction in larval zebrafish at 7 dpf. There was no significant effect of PFOSA on ROS levels (F (4, 19) = 0.14, P=0.96) (Figure 3).
3.3. Visual Motor Response Test
The DanioVision™ was used to assess the Visual Motor Response. Three independent trials were conducted, and data for relative distance travelled (log transformed and relative to the DMSO control standardized to a value of 1) was combined for each experiment. Zebrafish larvae exposed to 100 µg/L exhibited reduced activity in Dark Period 2 only ( Number of groups = 6, Kruskal-Wallis statistic = 13.60, P value = 0.018) (Figure 4, panel C). Individual VMR trials can be found in Supplementary Figures, Supplemental Figure S1.
3.4. Expression Analysis of Transcripts
The effect of PFOSA on mRNA steady state levels were measured in larval fish. Regression analysis revealed that the apoptosis-related transcript (bax) on oxidative stress-related transcripts (cat, sod2) increased with increasing concentrations of PFOSA and the statistical information and R2 of the linear regressions are shown in Figure 5. Bax mRNA levels were elevated in zebrafish from the 1 µg/L PFOSA treatment compared to the DMSO control group (F (4, 21) = 4.160, P=0.0123) (Figure 6A).
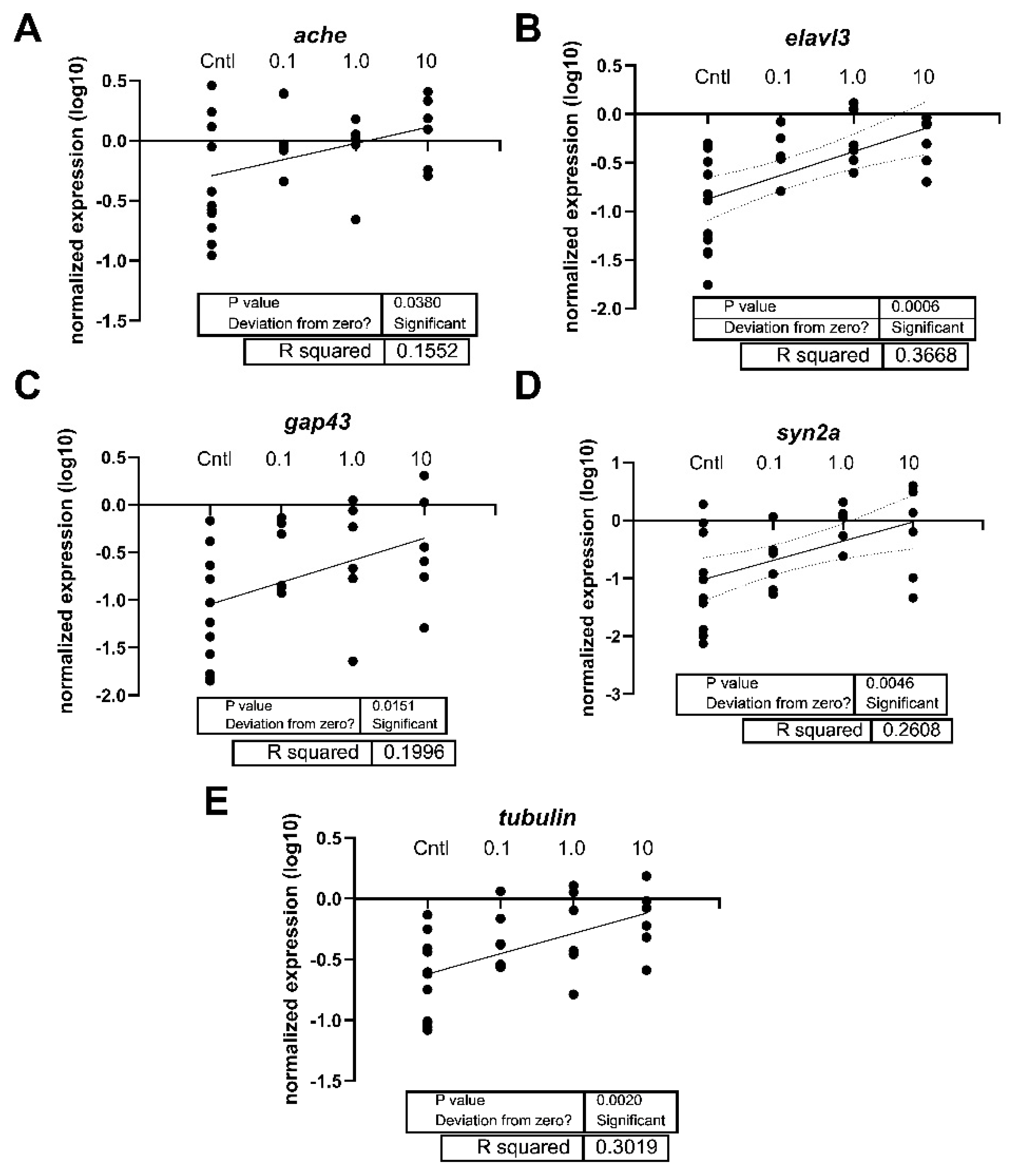
Regarding neurotoxicity-related genes, transcript levels of elavl3 were significantly elevated in zebrafish treated with 1 and 10 µg/L PFOSA (F (4, 23) = 4.802, P=0.0058) (Figure 6B). No other transcript was significantly different when comparing group means to the mean of the DMSO control (P>0.05); however, regression analysis revealed that many neurotoxicity transcripts showed a concentration independent response with PFOSA, and several transcripts were increased with higher exposure concentrations (Figure 7). The statistical information and R2 of the simple linear regressions are provided in the figures.
4. Discussion
PFOSA is a synthetic compound used to produce non-stick, waterproof, and stain-repellent coatings [34]. PFOSA is a precursor to PFOS, a PFAS that is regulated by the United States Environmental Protection Agency due to its known toxicity towards various organisms. It is important to recognize that not all PFOSA breaks down into PFOS. For example, Galatius et al. (2013) studied three top predators in the Danish North Sea and found evidence that the ability to metabolize PFOSA to PFOS was variable [35]. There remains a lack of focus on PFOSA despite its prevalence alongside PFOS. To address this knowledge gap, we investigated the effects of waterborne PFOSA exposure on developmental and behavior endpoints in zebrafish. Additionally, we measured transcripts associated with oxidative stress and neurotoxicity to account for potential mechanisms underlying locomotor activity alterations.
We observed that PFOSA affected the survival of fish treated with 1 µg/L (10% decline at this concentration relative to the DMSO control). Dasgupta et al. (2020) exposed zebrafish embryos to 0.78-50 µM (389-24,900 µg/L) PFOSA for up to 72 hpf and 100% mortality was observed in all treated embryos by 72 hpf [17]. Regarding abnormalities, we observed only a few deformities (less than 2-3 percent) across all treatment groups; several studies examining toxicity effects of PFOSA report significant deformities, which are likely due to much higher treatment concentrations. It was also reported that all treated embryos exhibiting concentration- and duration-dependent abnormalities, as well as developmental delays, at 24 hpf. For instance, embryos exposed to PFOSA starting at 0.75 hpf exhibited stronger concentration-dependent delays in epiboly compared to embryos exposed at 4 or 5 hpf. Truong et al. (2022) exposed dechorionated zebrafish embryos to 0.015-100 µM (7.4-49,900 µg/L) of various PFAS, including PFOSA from 6 to 120 hpf [36]. PFAS were ranked on potency based on morphological effects (i.e., pericardial and yolk sac edema, brain and eye malformation) in which PFOSA was ranked second highest. Various studies also report cardiac abnormalities, including heart elongation and reduced cardiac output, heart rate, and stroke volume, in zebrafish exposed from 0.01-100 µg/L [15,16]. In our case with survival, 1 µg/L may have been too low a concentration to sufficiently activate defense mechanisms to PFOSA exposure, leading to lower overall survival in developing fish while higher concentrations of PFOSA may elicit a stronger defense response to mitigate toxicity, leading to higher survival. Such dose-dependent responses have been observed for other chemical exposures in zebrafish [37,38]. This hypothesis is supported by the increase in anti-oxidant defense enzymes with higher concentrations of PFOSA. Nevertheless, survival remained relatively high for fish up to the 100 µg/L exposure, suggesting that PFOSA, spanning environmental concentrations, is not overtly toxic up to 100 µg/L.
The amount of ROS is often indicative of the amount of oxidative stress in cells, and an excess of ROS can contribute to damage at the molecular level. Limited studies in literature examine the impact PFOSA has on organisms. In our study, we did not observe any increase in ROS in zebrafish treated with PFOSA; however, there was a concentration dependent increase in both cat and sod2 expression. Mitochondrial dysfunction has been thought to contribute to the progression of neurodegenerative disorders and the presence of ROS is one clear indicator of dysfunction as antioxidant systems are implemented to counteract oxidative stress. This supports our oxidative stress-related gene responses (PFOSA-induced increase in cat and sod2 mRNA levels) which could have mitigated any change in ROS levels in the larval zebrafish. Similar results have also been observed in rodent models where oxidative stress-related genes, like cat, were significantly increased to counteract damage by PFOA-induced lipid peroxidation in mouse brain and liver tissues [39]. ROS can also trigger apoptosis to mediate inflammation. Though PFOSA was found to only significantly upregulate two apoptosis-related genes (bax and cycs) in our study, another study reports increased apoptotic cells in the brain and upregulated bcl-2, caspase3, and p53 zebrafish exposed to PFOS (Mahapatra et al., 2023). Bax, is a pro-apoptotic factor in the Bcl-2 family, signaling mitochondria and cell death while cytochrome c is an intrinsic apoptotic signal activating downstream caspase enzymes. Other studies investigating PFOSA report mixed results for antioxidant gene expression and proteins. Olufsen and Arukwe (2015) exposed Atlantic salmon (Salmo salar) hepatocytes to 25 or 50 µM (12,400-24,900 µg/L) PFOSA for 24 or 48 hours and analyzed catalase (cat), glutathione peroxidase (gpx), glucocorticoid receptor (gr), and glutathione S-transferase (gst) mRNA levels, which were not significantly impacted [40]. Another study also exposed Atlantic salmon hepatocytes to 2, 20, or 50 µM (998, 9,900, or 24,900 µg/L) PFOSA for 12 or 24 hours [3]. No significant changes to gpx mRNA levels were found, but cat mRNA levels were significantly increased by 20 and 50 µM PFOSA following 24 hours of exposure, suggesting that antioxidant defense mechanisms were activated. Differences among studies may occur due to the type of model used to investigate PFOSA toxicity (e.g. cells versus larvae). Taken together, there is evidence that PFOSA initiates an antioxidant defense and any elevation in ROS may lead to higher levels of apoptosis in larval zebrafish.
We hypothesized that PFOSA would induce neurotoxicity in the form of behavioral changes and altered expression of genes related to neurotoxicity. Indeed, we observed hypoactivity at 100 µg/L which corresponded to the highest expression levels of several neurotoxicity biomarkers. These responses indicate some form of neurological impairment [41]. Chemical neurotoxins cause damage to, or death of, cells in the nervous system, disrupting neuronal pathways linked to neurodegenerative illnesses and other neurodevelopmental issues (i.e., Parkinson’s disease and schizophrenia). For instance, zebrafish exposed to PFOS had transcriptome changes linked to disturbance of the neuromuscular system [42] and zebrafish exposed to perfluorononanoic acid (PFNA) showed evidence of altered neuroinflammatory pathways [43]. To our knowledge, this is the first study to examine neurotoxicity mechanism in zebrafish exposed to PFOSA. We observed hypoactivity effects on locomotor activity in larval zebrafish, suggesting neurotoxicity or motor deficits with PFOSA exposure. Fish treated with 100 µg/L PFOSA showed reduced activity in Dark Period 2 of the combined VMR. Truong et al. (2022) exposed dechorionated zebrafish embryos to 0.015-100 µM PFOSA from 6 to 120 hpf and found that PFOSA induced both a refractory and an excitatory phase of hyperactivity [36]. Consistent with our observation, after exposing zebrafish embryos to 1 or 10 µg/L PFOSA for 120 hours, Liu et al. (2022) observed reductions in total distance moved, average swimming velocity, and maximum acceleration in fish treated with 1 µg/L PFOSA [16]. Our results also revealed that PFOSA alters the expression of neurotoxicity-related genes, as notable effects were observed in elavl3, and positive associations were detected between PFOSA concentration and expression levels (e.g. ache, elavl3, gap43, syn2a, and tubb3). Elavl3 is expressed in different nervous system structures and is known to regulate neurogenesis [44]. Additionally, ache is involved in cholinergic functioning and syn2a is involved in dopamine and serotonin release. PFAS exposure has previously been shown to alter these transcripts; PFOS, the metabolic product of PFOSA, was reported to decrease ache expression [45] and perfluorododecanoate (PFDoA) decreased mRNA levels of elavl3, gap43, and syn2a [46]. Here we report an elevation in the expression of elavl3 and many other neurotoxic-related transcripts with PFOSA exposure, and this may reflect a compensatory response to impaired neurogenesis and neurotransmitter release. Conversely, different PFAS may elicit unique responses in the CNS in relation to gene expression patterns. Regardless, there is evidence from the molecular response that PFOSA alters genes related to neuronal integrity and structure, suggesting the potential for neurotoxicity in developing larval fish. Thus, early developmental exposures to PFOSA may have long lasting detrimental effects on the adult brain and this should be further investigated.
5. Conclusions
In summary, literature regarding PFOSA is currently lacking and there is little information on PFOSA’s environmental presence and accumulation in aquatic species. PFOSA did not significantly impact the prevalence of malformations or reactive oxygen species generated in larval fish; however, PFOSA did affect locomotor activity and transcripts related to oxidative damage response, apoptosis, and neurotoxicity. Further mechanistic studies in zebrafish are warranted to further address PFOSA neurotoxicity in the CNS. This study contributes to our knowledge regarding the relative toxicity of PFAS on fish to assist future risk assessment methodologies of these concerning, persistent environmental pollutants.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Independent VMR experiments; Table S1: Primers used for qPCR analysis.
Author Contributions
Conceptualization, C.J.M.; Investigation, A.S., C.D.E., C.E., D.S., E.I., E.M.A.V., I.K., L.A., M.K., M.R., and N.D.; writing—original draft preparation, E.I.; writing—review and editing, E.I. and C.J.M.; supervision, C.J.M; funding acquisition, C.J.M.,
Funding
This research was funded by the University of Florida and the College of Veterinary Medicine (UF IACUC202300000140). E.M.A.V was supported by FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo). I.K. would like to thank the Coordination for the Improvement of Higher Education Personnel (CAPES, Brazil) for scholarship support.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board Institutional Animal Care and Use Committee of University of Florida
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available upon request.
Acknowledgments
We thank Kari Buchanan and the Animal Care Services at the University of Florida for zebrafish husbandry and technical support.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Solan, M.E.; Lavado, R. The use of in vitro methods in assessing human health risks associated with short-chain perfluoroalkyl and polyfluoroalkyl substances (PFAS). Journal of Applied Toxicology 2022, 42, 1298–1309. [Google Scholar] [CrossRef]
- Agency, T.D.E.P. Short-chain Polyfluoroalkyl Substances (PFAS). 2015.
- Wågbø, A.M.; Cangialosi, M.V.; Cicero, N.; Letcher, R.J.; Arukwe, A. Perfluorooctane sulfonamide-mediated modulation of hepatocellular lipid homeostasis and oxidative stress responses in Atlantic salmon hepatocytes. Chemical research in Toxicology 2012, 25, 1253–1264. [Google Scholar] [CrossRef]
- Gust, K.A.; Mylroie, J.E.; Kimble, A.N.; Wilbanks, M.S.; Steward, C.S.; Chapman, K.A.; Jensen, K.M.; Kennedy, A.J.; Krupa, P.M.; Waisner, S.A. Survival, Growth, and Reproduction Responses in a Three-Generation Exposure of the Zebrafish (Danio rerio) to Perfluorooctane Sulfonate. Environmental Toxicology and Chemistry 2024, 43, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Wu, P.; Wang, L.; Li, Q.; Li, X.; Luo, Y. Toxicity of per-and polyfluoroalkyl substances to aquatic vertebrates. Frontiers in Environmental Science 2023, 11, 1101100. [Google Scholar] [CrossRef]
- Razak, M.R.; Aris, A.Z.; Zainuddin, A.H.; Yusoff, F.M.; Yusof, Z.N.B.; Kim, S.D.; Kim, K.W. Acute toxicity and risk assessment of perfluorooctanoic acid (PFOA) and perfluorooctanesulfonate (PFOS) in tropical cladocerans Moina micrura. Chemosphere 2023, 313, 137377. [Google Scholar] [CrossRef]
- Konwick, B.J.; Tomy, G.T.; Ismail, N.; Peterson, J.T.; Fauver, R.J.; Higginbotham, D.; Fisk, A.T. Concentrations and patterns of perfluoroalkyl acids in Georgia, USA surface waters near and distant to a major use source. Environmental Toxicology and Chemistry: An International Journal 2008, 27, 2011–2018. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H.; Long, G.C.; Porter, R.C.; Anderson, J.K. Occurrence of select perfluoroalkyl substances at US Air Force aqueous film-forming foam release sites other than fire training areas: field validation of critical fate and transport properties. In Perfluoroalkyl Substances in the Environment, CRC Press: 2018; pp. 353–372. [CrossRef]
- Brusseau, M.L.; Anderson, R.H.; Guo, B. PFAS concentrations in soils: Background levels versus contaminated sites. Science of the Total environment 2020, 740, 140017. [Google Scholar] [CrossRef] [PubMed]
- Fair, P.A.; Wolf, B.; White, N.D.; Arnott, S.A.; Kannan, K.; Karthikraj, R.; Vena, J.E. Perfluoroalkyl substances (PFASs) in edible fish species from Charleston Harbor and tributaries, South Carolina, United States: Exposure and risk assessment. Environmental research 2019, 171, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yeung, L.W.Y.; Xu, M.; Taniyasu, S.; Lam, P.K.; Yamashita, N.; Dai, J. Perfluorooctane sulfonate (PFOS) and other fluorochemicals in fish blood collected near the outfall of wastewater treatment plant (WWTP) in Beijing. Environmental Pollution 2008, 156, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Brandsma, S.H.; Smithwick, M.; Solomon, K.; Small, J.; de Boer, J.; Muir, D.C. Dietary exposure of rainbow trout to 8: 2 and 10: 2 fluorotelomer alcohols and perfluorooctanesulfonamide: Uptake, transformation and elimination. Chemosphere 2011, 82, 253–258. [Google Scholar] [CrossRef]
- Hagenaars, A.; Vergauwen, L.; De Coen, W.; Knapen, D. Structure–activity relationship assessment of four perfluorinated chemicals using a prolonged zebrafish early life stage test. Chemosphere 2011, 82, 764–772. [Google Scholar] [CrossRef]
- Wasel, O.; Thompson, K.M.; Freeman, J.L. Assessment of unique behavioral, morphological, and molecular alterations in the comparative developmental toxicity profiles of PFOA, PFHxA, and PFBA using the zebrafish model system. Environment international 2022, 170, 107642. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qiu, W.; Yang, X.; Chen, F.; Chen, J.; Tang, L.; Zhong, H.; Magnuson, J.T.; Zheng, C.; Xu, E.G. Perfluorooctane sulfonamide (PFOSA) induces cardiotoxicity via aryl hydrocarbon receptor activation in zebrafish. Environmental science & technology 2022, 56, 8438–8448. [Google Scholar] [CrossRef]
- Liu, X.; Liu, S.; Qiu, W.; Magnuson, J.T.; Liu, Z.; Yang, G.; Chen, H.; Li, Y.; Xu, X.; Zheng, C. Cardiotoxicity of PFOA, PFOS, and PFOSA in early life stage zebrafish: Molecular changes to behavioral-level response. Sustainable Horizons 2022, 3, 100027. [Google Scholar] [CrossRef]
- Dasgupta, S.; Reddam, A.; Liu, Z.; Liu, J.; Volz, D.C. High-content screening in zebrafish identifies perfluorooctanesulfonamide as a potent developmental toxicant. Environmental Pollution 2020, 256, 113550. [Google Scholar] [CrossRef] [PubMed]
- Xuan, R.; Qiu, X.; Wang, J.; Liu, S.; Magnuson, J.T.; Xu, B.; Qiu, W.; Zheng, C. Hepatotoxic response of perfluorooctane sulfonamide (PFOSA) in early life stage zebrafish (Danio rerio) is greater than perfluorooctane sulfonate (PFOS). Journal of Hazardous Materials 2024, 461, 132552. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book; A guide for the laboratory use of zebrafish (Danio rerio). (No Title) 2007.
- Cao, F.; Souders II, C.L.; Li, P.; Pang, S.; Liang, X.; Qiu, L.; Martyniuk, C.J. Developmental neurotoxicity of maneb: Notochord defects, mitochondrial dysfunction and hypoactivity in zebrafish (Danio rerio) embryos and larvae. Ecotoxicology and Environmental Safety 2019, 170, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rodriguez, V.; Souders II, C.L.; Tischuk, C.; Martyniuk, C.J. Tebuconazole reduces basal oxidative respiration and promotes anxiolytic responses and hypoactivity in early-staged zebrafish (Danio rerio). Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2019, 217, 87–97. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Developmental dynamics 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Adamovsky, O.; Souders II, C.L.; Martyniuk, C.J. Biological effects of the benzotriazole ultraviolet stabilizers UV-234 and UV-320 in early-staged zebrafish (Danio rerio). Environmental pollution 2019, 245, 272–281. [Google Scholar] [CrossRef]
- Dong, M.; Wang, J.; Liu, Y.; He, Q.; Sun, H.; Xu, Z.; Hong, H.; Lin, H.; Gao, P. 3-bromocarbazole-induced developmental neurotoxicity and effect mechanisms in zebrafish. ACS ES&T Water 2023, 3, 2471–2480. [Google Scholar] [CrossRef]
- Fang, C.; Di, S.; Yu, Y.; Qi, P.; Wang, X.; Jin, Y. 6PPD induced cardiac dysfunction in zebrafish associated with mitochondrial damage and inhibition of autophagy processes. Journal of Hazardous Materials 2024, 471, 134357. [Google Scholar] [CrossRef]
- Guo, Y.; Fu, Y.; Sun, W. 50 Hz Magnetic Field Exposure Inhibited Spontaneous Movement of Zebrafish Larvae through ROS-Mediated syn2a Expression. International Journal of Molecular Sciences 2023, 24, 7576. [Google Scholar] [CrossRef]
- Hahn, M.E.; McArthur, A.G.; Karchner, S.I.; Franks, D.G.; Jenny, M.J.; Timme-Laragy, A.R.; Stegeman, J.J.; Woodin, B.R.; Cipriano, M.J.; Linney, E. The transcriptional response to oxidative stress during vertebrate development: effects of tert-butylhydroquinone and 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin. PloS one 2014, 9, e113158. [Google Scholar]
- Jiang, F.; Liu, J.; Zeng, X.; Yu, L.; Liu, C.; Wang, J. Tris (2-butoxyethyl) phosphate affects motor behavior and axonal growth in zebrafish (Danio rerio) larvae. Aquatic Toxicology 2018, 198, 215–223. [Google Scholar] [CrossRef] [PubMed]
- McCurley, A.T.; Callard, G.V. Characterization of housekeeping genes in zebrafish: male-female differences and effects of tissue type, developmental stage and chemical treatment. BMC molecular biology 2008, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Mukherjee, S.; Chattopadhyay, A.; Bhattacharya, S. Low dose of arsenic trioxide triggers oxidative stress in zebrafish brain: expression of antioxidant genes. Ecotoxicology and environmental safety 2014, 107, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Souders II, C.L.; Zhao, Y.H.; Martyniuk, C.J. Paraquat affects mitochondrial bioenergetics, dopamine system expression, and locomotor activity in zebrafish (Danio rerio). Chemosphere 2018, 191, 106–117. [Google Scholar] [CrossRef]
- Wu, Q.; Yan, W.; Liu, C.; Li, L.; Yu, L.; Zhao, S.; Li, G. Microcystin-LR exposure induces developmental neurotoxicity in zebrafish embryo. Environmental pollution 2016, 213, 793–800. [Google Scholar] [CrossRef]
- Yang, Q.; Deng, P.; Xing, D.; Liu, H.; Shi, F.; Hu, L.; Zou, X.; Nie, H.; Zuo, J.; Zhuang, Z. Developmental neurotoxicity of difenoconazole in zebrafish embryos. Toxics 2023, 11, 353. [Google Scholar] [CrossRef]
- EWG. Perfluorooctanesulfonamide (PFOSA). Availabe online: https://www.ewg.org/tapwater/contaminant.php?contamcode=E314#:~:text=Perfluorooctanesulfonamide%20(PFOSA)%20is%20a%20member,read%20more%20about%20perfluorinated%20chemicals. (accessed on.
- Galatius, A.; Bossi, R.; Sonne, C.; Rigét, F.F.; Kinze, C.C.; Lockyer, C.; Teilmann, J.; Dietz, R. PFAS profiles in three North Sea top predators: metabolic differences among species? Environmental Science and Pollution Research 2013, 20, 8013–8020. [Google Scholar] [CrossRef] [PubMed]
- Truong, L.; Rericha, Y.; Thunga, P.; Marvel, S.; Wallis, D.; Simonich, M.T.; Field, J.A.; Cao, D.; Reif, D.M.; Tanguay, R.L. Systematic developmental toxicity assessment of a structurally diverse library of PFAS in zebrafish. Journal of hazardous materials 2022, 431, 128615. [Google Scholar] [CrossRef]
- English, C.D.; Kazi, K.J.; Konig, I.; Ivantsova, E.; Souders II, C.L.; Martyniuk, C.J. Exposure to the antineoplastic ifosfamide alters molecular pathways related to cardiovascular function, increases heart rate, and induces hyperactivity in zebrafish (Danio rerio). Environmental Toxicology and Pharmacology 2024, 107, 104427. [Google Scholar] [PubMed]
- Ivantsova, E.; Huang, M.; Wengrovitz, A.S.; Souders II, C.L.; Martyniuk, C.J. Molecular and behavioral assessment in larval zebrafish (Danio rerio) following exposure to environmentally relevant levels of the antineoplastic cyclophosphamide. Environmental Toxicology and Pharmacology 2022, 90, 103809. [Google Scholar]
- Endirlik, B.Ü.; Eken, A.; Canpınar, H.; Öztürk, F.; Gürbay, A. Perfluorooctanoic acid affects mouse brain and liver tissue through oxidative stress. Arhiv za higijenu rada i toksikologiju 2022, 73, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Olufsen, M.; Arukwe, A. Endocrine, biotransformation, and oxidative stress responses in salmon hepatocytes exposed to chemically induced hypoxia and perfluorooctane sulfonamide (PFOSA), given singly or in combination. Environmental Science and Pollution Research 2015, 22, 17350–17366. [Google Scholar] [CrossRef] [PubMed]
- Selderslaghs, I.W.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: a new method to assess developmental neurotoxicity. Neurotoxicology and teratology 2010, 32, 460–471. [Google Scholar] [CrossRef]
- Wang, X.; Shi, X.; Zheng, S.; Zhang, Q.; Peng, J.; Tan, W.; Wu, K. Perfluorooctane sulfonic acid (PFOS) exposures interfere with behaviors and transcription of genes on nervous and muscle system in zebrafish embryos. Science of The Total Environment 2022, 848, 157816. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Qiu, W.; Li, R.; Chen, B.; Wu, X.; Magnuson, J.T.; Xu, B.; Luo, S.; Xu, E.G.; Zheng, C. Perfluorononanoic acid induces neurotoxicity via synaptogenesis signaling in zebrafish. Environmental Science & Technology 2023, 57, 3783–3793. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kakumoto, K.; Yoshida, T.; Kuwako, K.I.; Miyazaki, T.; Yamaguchi, J.; Konno, A.; Hata, J.; Uchiyama, Y.; Hirai, H.; et al. Elavl3 is essential for the maintenance of Purkinje neuron axons. Sci Rep 2018, 8, 2722. [Google Scholar] [CrossRef] [PubMed]
- Starnes, H.M.; Rock, K.D.; Jackson, T.W.; Belcher, S.M. A critical review and meta-analysis of impacts of per-and polyfluorinated substances on the brain and behavior. Frontiers in Toxicology 2022, 4, 881584. [Google Scholar] [CrossRef] [PubMed]
- Foguth, R.; Sepúlveda, M.S.; Cannon, J. Per-and polyfluoroalkyl substances (PFAS) neurotoxicity in sentinel and non-traditional laboratory model systems: Potential utility in predicting adverse outcomes in human health. Toxics 2020, 8, 42. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Total percent of surviving zebrafish embryos/larvae exposed to ERM, 0.1% DMSO, 0.1, 1, 10, or 100 µg/L PFOSA over 7 days. .
Figure 1.
Total percent of surviving zebrafish embryos/larvae exposed to ERM, 0.1% DMSO, 0.1, 1, 10, or 100 µg/L PFOSA over 7 days. .
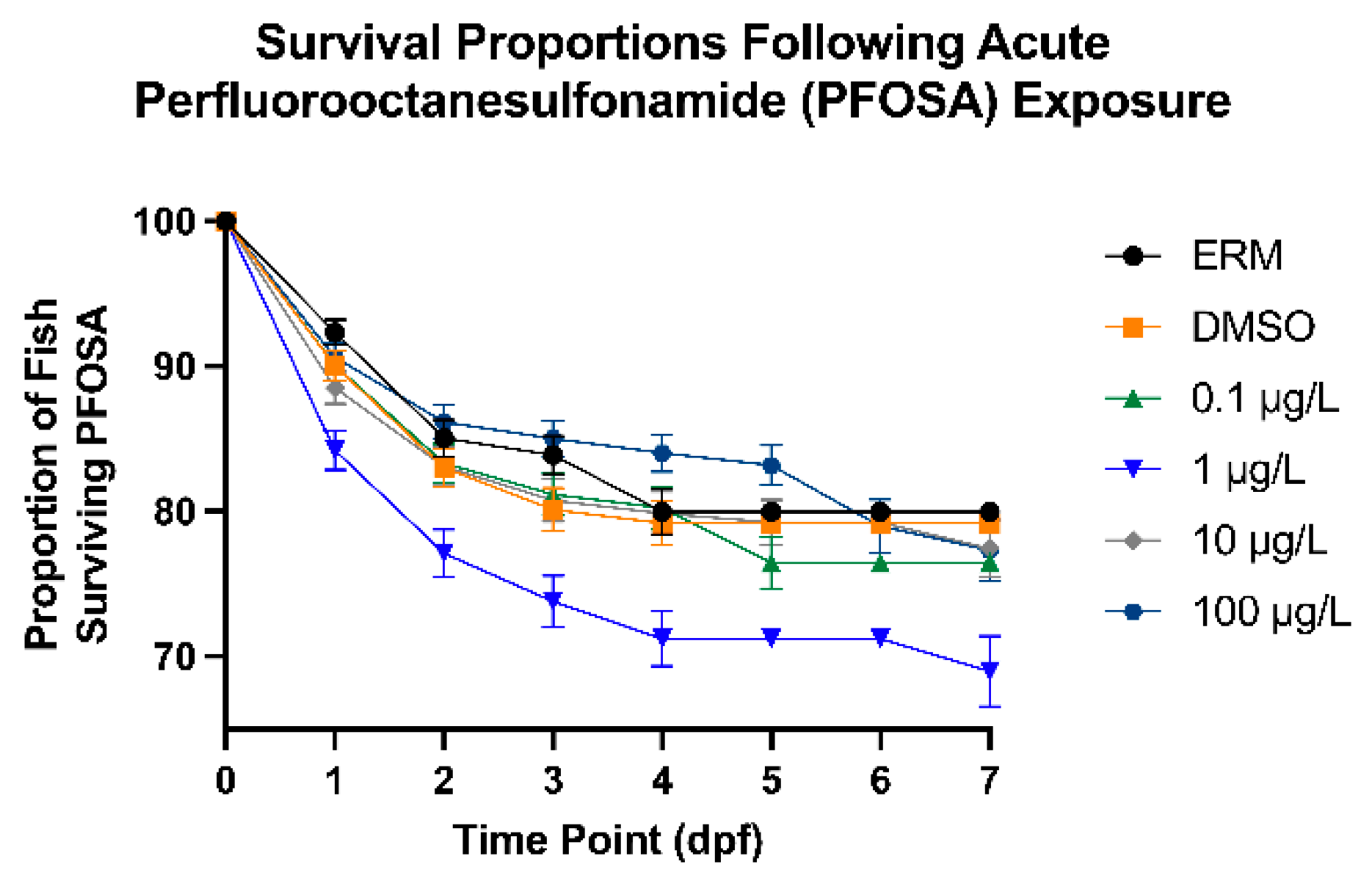
Figure 2.
Total percent of hatched zebrafish embryos/larvae exposed to ERM, 0.1% DMSO, 0.1, 1, 10, or 100 µg/L PFOSA over 4 days.
Figure 2.
Total percent of hatched zebrafish embryos/larvae exposed to ERM, 0.1% DMSO, 0.1, 1, 10, or 100 µg/L PFOSA over 4 days.
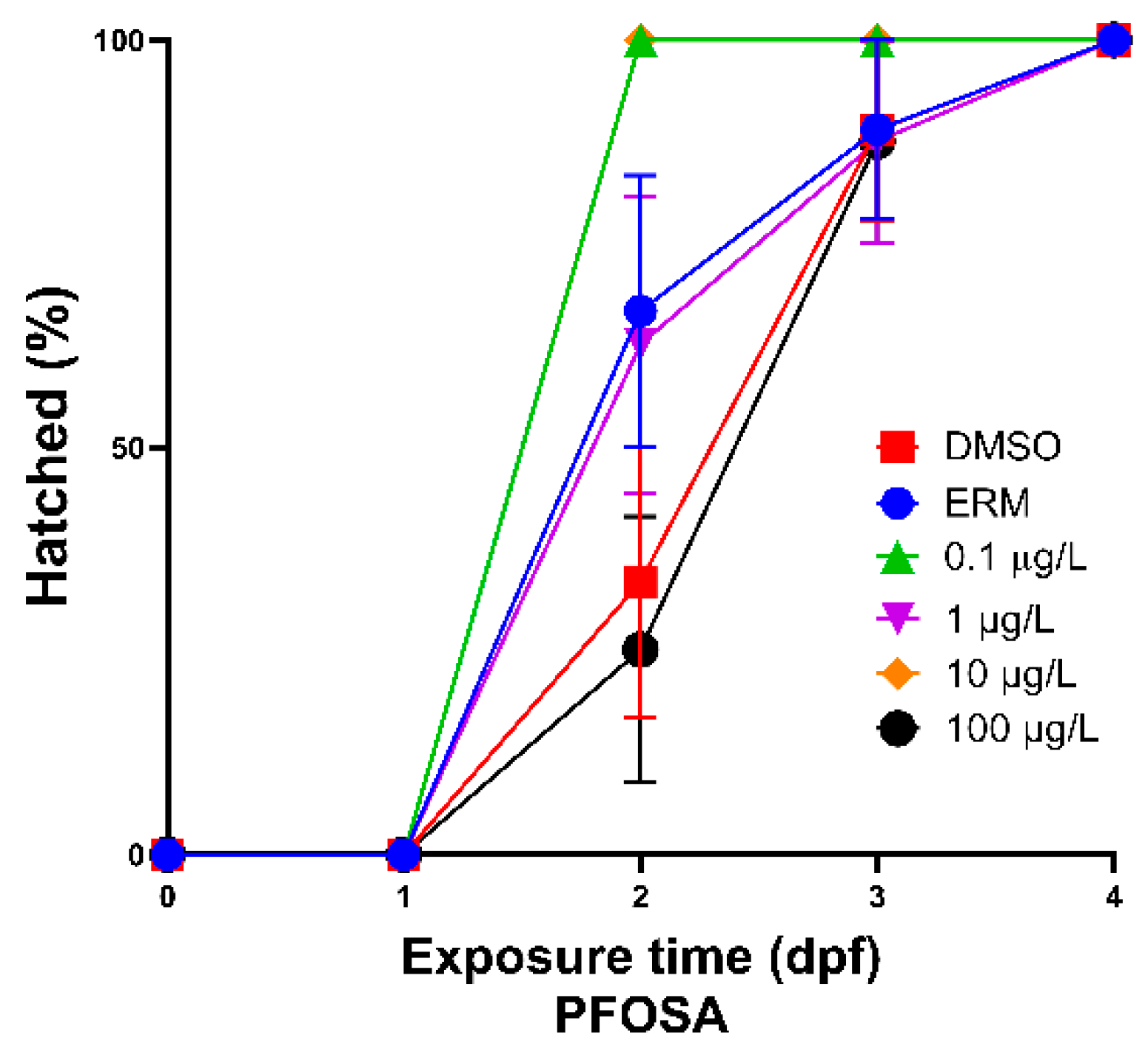
Figure 3.
Normalized reactive oxygen species (to µg/mL media/protein) present in zebrafish at 7 dpf. Each circle represents a biological replicate, the horizontal line indicates the mean (± S.D.), and ns = not significant (One-Way ANOVA, Dunnett’s multiple comparisons test, n=5/treatment).
Figure 3.
Normalized reactive oxygen species (to µg/mL media/protein) present in zebrafish at 7 dpf. Each circle represents a biological replicate, the horizontal line indicates the mean (± S.D.), and ns = not significant (One-Way ANOVA, Dunnett’s multiple comparisons test, n=5/treatment).
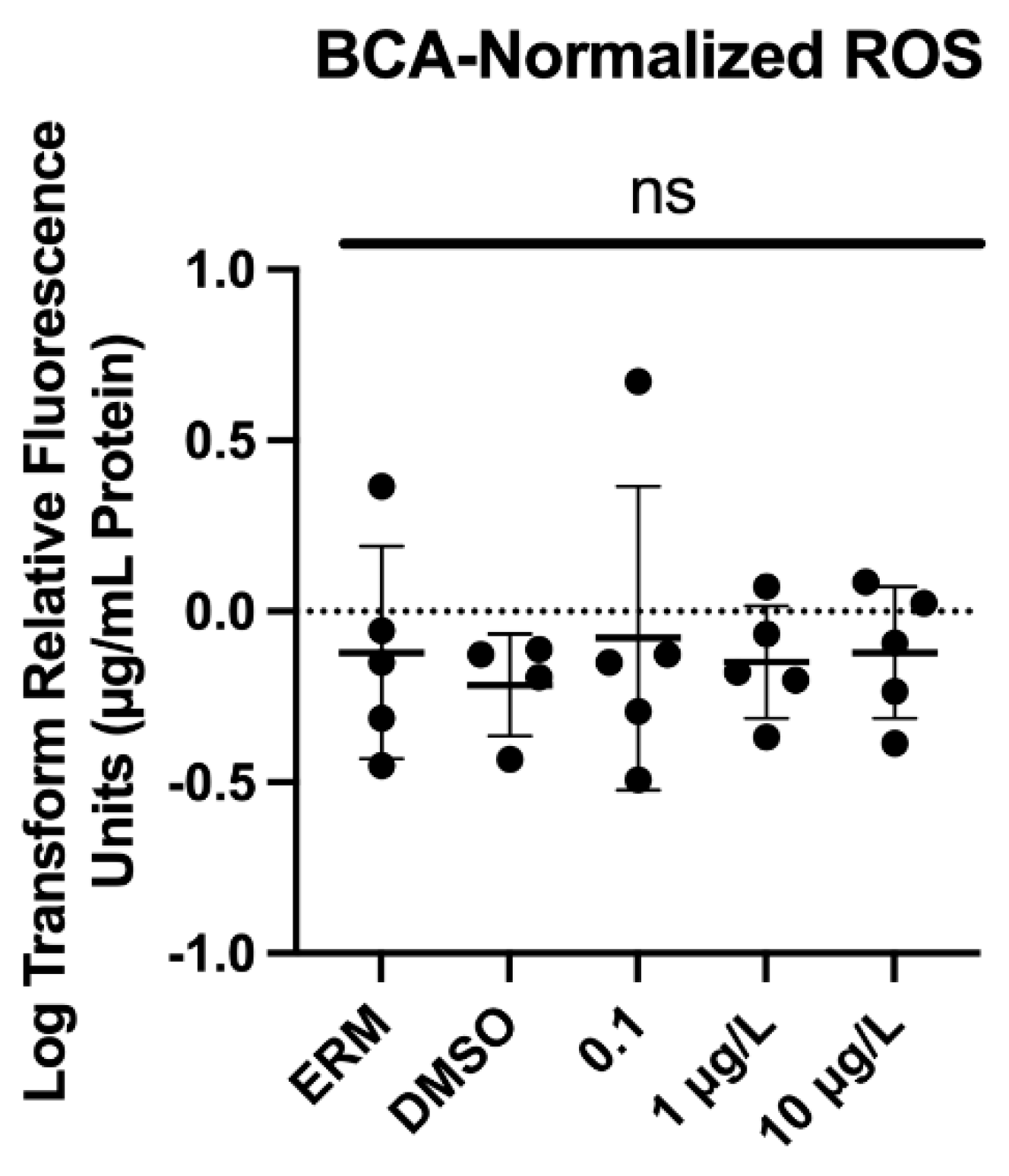
Figure 4.
The distance moved in each of the light and dark zones (10-minute bins) of 7-day zebrafish larvae exposed to 0.1% DMSO, ERM, 0.1, 1, 10, or 100 µg/L PFOSA. Graphs are the combined output from three independent VMR runs. Mean values are depicted by the columns in each dark-light phase (mean ± S.D.) (Kruskal-Wallis test followed by a multiple comparisons test, n=8-12 fish/treatment/run). Asterisk indicates **P<0.01. .
Figure 4.
The distance moved in each of the light and dark zones (10-minute bins) of 7-day zebrafish larvae exposed to 0.1% DMSO, ERM, 0.1, 1, 10, or 100 µg/L PFOSA. Graphs are the combined output from three independent VMR runs. Mean values are depicted by the columns in each dark-light phase (mean ± S.D.) (Kruskal-Wallis test followed by a multiple comparisons test, n=8-12 fish/treatment/run). Asterisk indicates **P<0.01. .
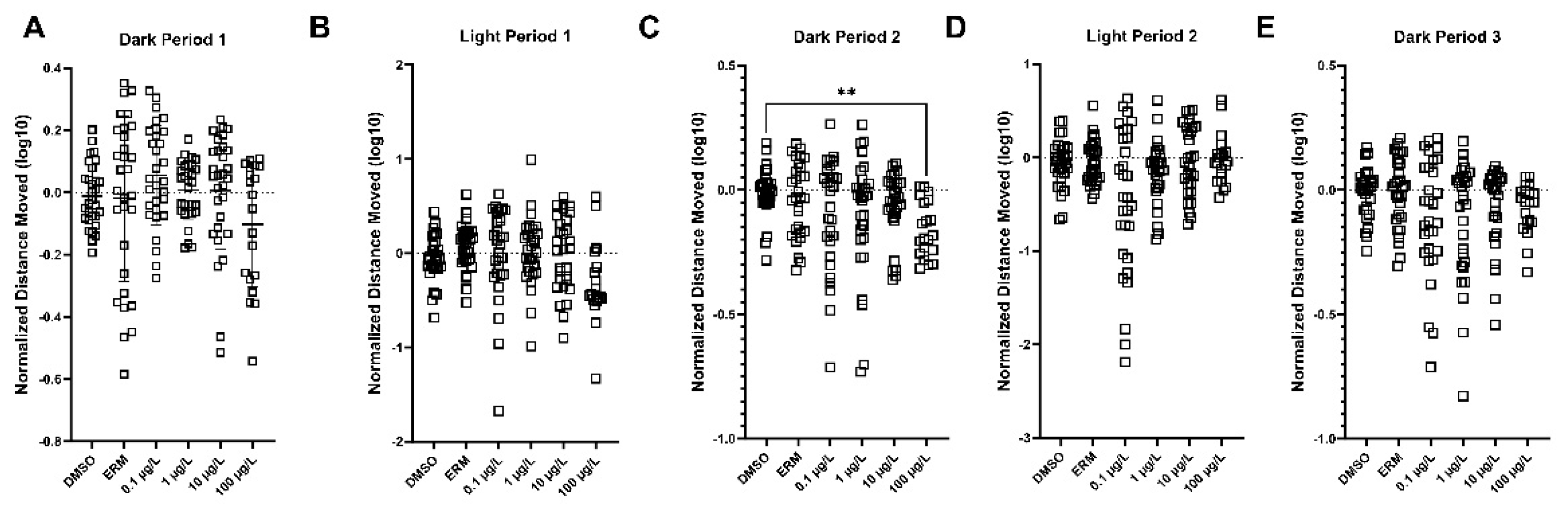
Figure 5.
Linear regression of the expression levels of (A) bax, (B) cat, and (C) sod2 in 7-day old larval zebrafish exposed to either 0.1% DMSO, ERM, 0.1, 1, or 10 µg/L PFOSA. The DMSO and ERM group were combined to yield the control (Cntl) group. Each circle is a biological replicate (n=4 to 6).
Figure 5.
Linear regression of the expression levels of (A) bax, (B) cat, and (C) sod2 in 7-day old larval zebrafish exposed to either 0.1% DMSO, ERM, 0.1, 1, or 10 µg/L PFOSA. The DMSO and ERM group were combined to yield the control (Cntl) group. Each circle is a biological replicate (n=4 to 6).
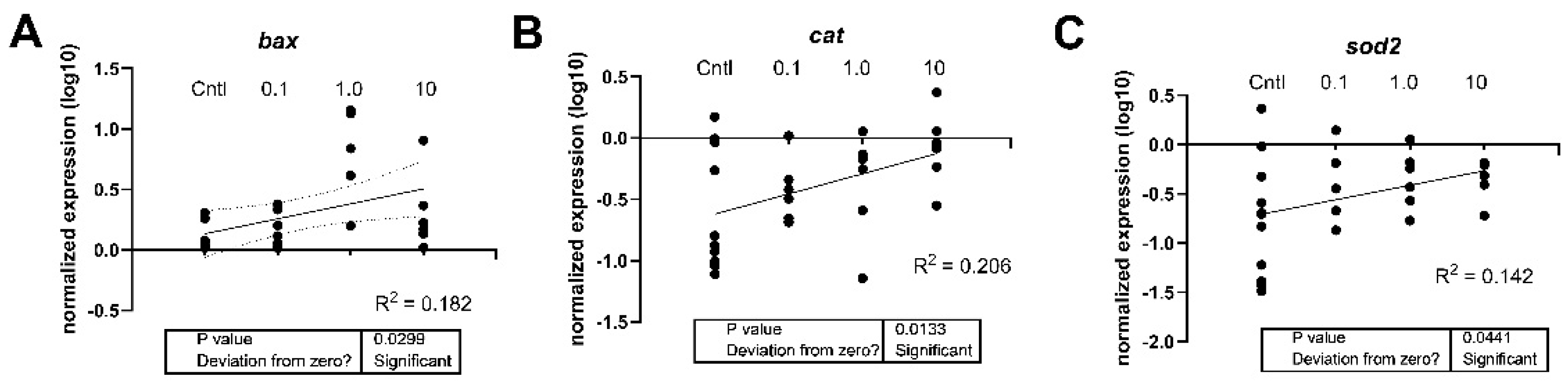
Figure 6.
The expression levels of (A) bax and (B) elavl3 in 7-day old larval zebrafish exposed to 0.1% DMSO, ERM, 0.1, 1, or 10 µg/L PFOSA. Each circle is a biological replicate, and the horizontal line indicates the mean (± S.D.) (One-Way ANOVA, Dunnett’s multiple comparisons test, n=4 to 6). Asterisk indicates *P<0.05 or **P<0.01. .
Figure 6.
The expression levels of (A) bax and (B) elavl3 in 7-day old larval zebrafish exposed to 0.1% DMSO, ERM, 0.1, 1, or 10 µg/L PFOSA. Each circle is a biological replicate, and the horizontal line indicates the mean (± S.D.) (One-Way ANOVA, Dunnett’s multiple comparisons test, n=4 to 6). Asterisk indicates *P<0.05 or **P<0.01. .

Figure 7.
Linear regression of the expression levels of (A) ache, (B) elavl3, (C) gap43, (D) syn2a, and (E) tubulin (tubb3) in 7-day old larval zebrafish exposed to either 0.1% DMSO, ERM, 0.1, 1, or 10 µg/L PFOSA. The DMSO and ERM group were combined to yield the control (Cntl) group. Each circle is a biological replicate (n=4 to 6).
Figure 7.
Linear regression of the expression levels of (A) ache, (B) elavl3, (C) gap43, (D) syn2a, and (E) tubulin (tubb3) in 7-day old larval zebrafish exposed to either 0.1% DMSO, ERM, 0.1, 1, or 10 µg/L PFOSA. The DMSO and ERM group were combined to yield the control (Cntl) group. Each circle is a biological replicate (n=4 to 6).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



