
Submitted:
19 August 2024
Posted:
20 August 2024
You are already at the latest version
Abstract
The co-occurrence of different arboviruses, through (re)emergence episodes in a particular region, often exacerbates public health crises associated with these pathogens. Considering the Brazilian epidemiological panorama of arboviruses cocirculation, such as Dengue (DENV), Zika (ZIKV), and Chikungunya (CHIKV) viruses, the imminent emergence of Mayaro virus (MAYV) in urban areas, and the presence of susceptible human hosts, we looked into the coinfection and superinfection impact of CHIKV and MAYV replication in Aedes aegypti. Mosquitoes were coinfected and superinfected with CHIKV and MAYV by artificial blood-feeding, and infection (IR), dissemination (DR), transmission (TR) rates, and viral load were evaluated by qRT-PCR. Our findings suggest that, Ae. aegypti can simultaneously transmit CHIKV and MAYV in coinfection scenarios. However, in superinfection assays, the initial infection (either CHIKV or MAYV) adversely affected the transmission capacity of the subsequently acquired virus, indicating replication interference between the viruses. These results have important implications for public health management actions in the country, emphasizing complex arbovirus transmission scenarios and the associated epidemiological risks.
Keywords:
Aedes aegypti
; Alphavirus
; cocirculation
1. Introduction
Episodes of emergency and re-emergence of arthropod-borne viruses (arboviruses) are associated with the increasing contact among viruses, vectors, and human hosts [1]. This context is supported by factors such as globalization, anthropogenic changes in biomes, climate change, geographic spread of potential disease vectors, and unsanitary conditions in urban areas [2,3]. As a result, the high density of the vector population, the presence of susceptible hosts, and favorable environmental conditions result on the successful circulation, maintenance, and dissemination of multiple arboviruses in the environment [1,2,3]. In Brazil, the cocirculation of Dengue (DENV), Zika (ZIKV), and Chikungunya (CHIKV) viruses culminated in the triple epidemic registered in 2015-2016 [4]. Since then, these three viruses are found in urban areas, having humans as vertebrate hosts and Aedes aegypti as the main mosquito vector [5]. The cocirculation of arboviruses is influenced by similar biological, ecological, and socioeconomic factors, leading to an epidemiological synergy of seasonality and occurrence rates [2,6,7,8]. Recently, Mayaro virus (MAYV), an endemic arbovirus widely found in Central America and the Amazon region, has drawn attention due to its ability to invade urban areas and cause outbreaks [9,10,11], with cases reported in the North, Midwest, Southeast, and Northeast regions of Brazil [12].
One of the main factors for an arbovirus, such as MAYV, to establish itself in an urban environment is the presence of competent mosquito species for the virus [13,14,15]. In the sylvatic cycle, MAYV has non-human primates and other mammals as vertebrate hosts and Haemagoggus spp. mosquitoes as vectors [9,11]. On the other hand, entomological surveillance studies conducted in the country have detected Ae. aegypti naturally infected with MAYV in the Brazilian states of Mato Grosso [16] and Goiás [17]. Thus, the regular notification of MAYV cases near the main tropical cities, Ae. aegypti as a main urban vector, and the presence of susceptible human hosts result in a higher potential for establishing MAYV in an urban cycle [18].
From the standpoint of the vector biology, it is important to understand the dynamics between different viruses in the same individual mosquito. This phenomenon includes the coinfection (simultaneous infection of the mosquito by the viruses in a single blood meal) and superinfection (sequential infection of the mosquito by the viruses, involving more than one blood meal) mechanisms [19,20]. Both coinfection and superinfection can result in different scenarios: virus amplification; virus inhibition; competition between viruses; and neutrality [2,21]. In nature, superinfection is more likely to occur when compared to coinfection, as the mosquito becomes infected sequentially by performing a blood meal on several viremic hosts instead of a single meal on a coinfected viremic host [2,22,23]. Also, superinfection is commonly related to viral interference, referring to the inability of a virus to replicate in an organism or cell that is previously infected by another virus [24,25], which was already demonstrated between Yellow Fever virus (YFV) and DENV-2 [26], MAYV and ZIKV [21], and CHIKV and MAYV [22].
Therefore, in a setting of arboviruses cocirculation in Brazil and a high density of Ae. aegypti mosquitoes, we investigated how coinfection and superinfection impact the virus dynamics between two alphaviruses of a high epidemiological importance, CHIKV and MAYV, using an Ae. aegypti colony. The study consisted of blood-meal experiments, drawing different methods for coinfection and superinfection assays, and comparative analyses for the consequent scenarios.
2. Materials and Methods
2.1. Viral Stock
CHIKV (CHIKV BRPE408/2016) and MAYV (MAYV/BR/Sinop/H307/2015) (MH513597) stocks were gently provided by Dr. Marli Tenório (Department of Virology, LAVITE, Aggeu Magalhães Institute) and Dr. Roberta Bronzoni (Federal University of Mato Grosso, UFMT-Sinop), respectively. Viral stocks and viral titration were performed using VERO cells and plaque assays to determine plaque-forming units per mL (PFU/mL), using the protocol described in Krokovsky et al. [15].
2.2. Aedes aegypti Colony
The Ae. aegypti colony (RecL) used in the present study was derived from a natural population collected in Recife, Pernambuco, Brazil [27]. Mosquitoes were maintained under standard conditions of 26 ± 2 °C, 65–85% relative humidity, and photoperiod of 12/12 light-dark, until adults were obtained to form the groups used in each assay.
2.3. Aedes aegypti Artificial Blood-Feeding
Single infection, coinfection and superinfection assays were performed with CHIKV and MAYV. These experiments consisted of two independent artificial blood meals each one, in a BSL2 infectory (Biosafety Level 2). Seven to ten-day-old mosquitoes from the RecL colony were separated into different plastic cages containing 100 females and 20 males placed and maintained in containment cages. Twenty-four hours before blood feeding, mosquitoes were sugar-starved. MAYV and CHIKV were obtained from VERO cells collected 24 h post-inoculation at a 0.1 multiplicity of infection (MOI), while uninfected cultured cells were used in negative control groups.
For the blood meals, cell culture flasks were infected and mixed with defibrinated rabbit blood (1:1) and 3 mM ATP, then presented to the mosquitoes for up to one hour in plastic Petri dishes covered by a triple layer of parafilm paper, as previously described in Guedes et al. [28]. After the blood feeding, fully engorged females were cold anesthetized, transferred to a new cage, and maintained for 13 days post-exposure (dpe). Infected cell cultures used in each assay were collected to control the viral title offered in the blood meals (Table 1).
For single infections by MAYV or CHIKV, mosquitoes were first given an infectious blood meal with the respective virus, followed by a non-infectious blood meal six days later. In the coinfection group, both viruses were present in equal proportions in a single blood meal, followed by a non-infectious blood meal six days later. For the superinfection groups, two separate blood meals with different viruses were administered, spaced six days apart. Specifically, for CHIKV superinfection, the first blood meal contained MAYV, followed by a second blood meal containing CHIKV. Conversely, for MAYV superinfection, the initial blood meal contained CHIKV, followed by a second blood meal containing MAYV (Figure 1).
2.4. Sample Collection
The single infection, coinfection and superinfection experiments were carried out in duplicate assays. This resulted in a total number of samples for each group (midguts, carcasses, and saliva) of n=10 (5 for each replicate) for the single infected and negative control groups and n=30 samples (15 for each replicate) for the coinfected and superinfected groups at each time-point.
Midguts and carcasses were dissected from female mosquitoes collected at 6th and 12th dpe and cold anesthetized at -20 °C, washed individually in 70% alcohol and distilled water, and deposited in 1x PBS buffer (phosphate buffered saline) (Gibco). Tissues were dissected in a Petri dish containing 1x PBS with the aid of entomological pins and tweezers and visualized with a binocular stereomicroscope (LABOMED). Samples were individually transferred to a 1.5 ml DNAse/RNAse-free microtube containing 300 μl of mosquito diluent and stored at -80 °C until processing.
For saliva collection, female mosquitoes were collected at 7th and 13th dpe and cold anesthetized (-20 °C). The legs and wings were removed to stimulate salivation and the proboscis was immersed in 100 μl tips with 10 μl of 1x PBS buffer for 30 minutes. After the forced salivation, described by Gloria-Soria et al. with modifications [29], the 1x PBS buffer and the saliva were expelled into 1.5 ml DNAse/RNAse-free microtubes containing 10 μl of 1x PBS buffer and stored at -80 °C until qRT-PCR assay.
2.5. RNA Isolation and qRT-PCR
Collected tissues were individually homogenized as previously described in Barbosa et al. [30], and RNA extraction was performed using the TRIzol® (Invitrogen, Carlsbad, CA, USA) protocol, described in Guedes et al. [28]. The RNA extracted from the midguts, carcasses, and saliva samples were used as templates in qRT-PCR reactions carried out using the QuantiNova Kit Probe RT-PCR Kit (Qiagen, Hilden, North Rhine-Westphalia, Germany) in a final volume of 10 μl (3,5 μl of RNA) and final concentrations of 1X for QuantiNova Probe RT-PCR Master Mix, QuantiNova ROX, and QuantiNova RT Mix, and 800 nM of CHIKV primers, 1200 nM of MAYV primers, 200 nM of CHIKV probe, and 300 nM of MAYV probe. The sequence of primers and probes for CHIKV and MAYV were previously described by Lanciotti et al. [31] and Naveca et al. [32], respectively.
Reactions were performed using a QuantStudio 5 System (Applied BioSystems, Norwalk, CT, USA) under the following conditions: 45 °C for 15 minutes, 95 °C for 5 minutes, followed by 45 cycles of 95 °C for 5 seconds, and 60 °C for 45 seconds. Samples were analyzed in duplicates using negative reaction control, which corresponds to all reagents except for RNA template, negative RNA extraction control, and positive controls corresponding to a standard curve for each virus (MAYV and CHIKV), synthesized as described in Krokovsky et al. [33]. qRT-PCR results were analyzed using QuantStudio Design and Analysis Software v. 1.3.1 with automatic threshold and baseline. Samples with Cq (Cycle quantification) values ≤ 38.5 for both MAYV and CHIKV were considered positives.
2.6. Data Analyzes
The infection rate (IR = positive midguts/midguts analyzed) and dissemination rate (DR = positive carcasses/positive midguts) were calculated at 6 and 12 dpe, meanwhile the transmission rate (TR = positive saliva/saliva analyzed) was evaluated at 7 and 13 dpe. Statistical analyses were performed using GraphPad Prism 8 software (Graph Pad, San Diego, CA, USA). The Wilcoxon test was used to compare the viral load (RNA copy number/ml) between the different groups and time-points. Analyzed results were considered significant when the p-value < 0.05.
3. Results
3.1. Single Infection and Coinfection Assays
The IR for MAYV-single infected mosquitoes was 100% and 90% on the 6th and 12th dpe, respectively (Figure 2A), while the DR was 100% for both time-points (Figure 2C). Regarding the analysis of RNA copy number/ml, it ranged from 1,4 x 104 to 7,7 x 1010 on 6th dpe and 1,1 x 105 to 1 x 1011 on 12th dpe in the midguts samples (Figure 2B), while ranges from 5,9 x 106 to 2,2 x 1011 on 6th dpe and 1,1 x 107 to 5,6 x 1012 on 12th dpe (Figure 2D) were identified for carcasses (Figure 2D). CHIKV-single infected mosquitoes presented a 100% IR and DR for both time-points (Figure 3A and 3C). The RNA copy number/ml ranged from 1,5 x 1010 to 8,3 x 1012 on 6th dpe and 2,4 x 1010 to 3,6 x 1012 on 12th dpe in the midguts samples (Figure 3B), and for carcasses, ranges from 1,2 x 1012 to 1,8 x 1014 on 6th dpe and 1,4 x 1012 to 3,5 x 1013 on 12th dpe were observed (Figure 3D).
Concerning the coinfected group, CHIKV presented a higher IR (96.7% on 6th dpe and 93.3% on 12th dpe) (Figure 3A) than MAYV (93.2% on both 6 and 12 dpe) (Figure 3A), as well as a greater viral load (range from 1,2 x 107 to 2,7 x 1012 RNA copy number/ml on 6th dpe and 3,7 x 109 to 4,4 x 1012 on 12th dpe) than MAYV (range from 2,5 x 106 to 1,1 x 1010 RNA copy number/ml on 6th dpe and 1 x 107 to 4,2 x 1010 on 12th dpe) (Figure 2B and Figure 3B). Furthermore, both viruses revealed a 100% DR for both time-points analyzed (Figure 2C and Figure 3C); however, CHIKV displayed a higher viral load (range from 3,3 x 107 to 2,3 x 1013 RNA copy number/ml on 6th dpe and 1,7 x 108 to 2,7 x 1013 on 12th dpe) than MAYV (range from 1,5 x 105 to 8,9 x 1010 RNA copy number/ml on 6th dpe and 1,9 x 105 to 3 x 1011 on 12th dpe) (Figure 2D and Figure 3D).
Regarding TR, mosquitoes that were single-infected with MAYV showed positivity rates of 30% and 10% on the 7th and 13th dpe, respectively (Figure 4A). The RNA copy numbers/ml ranged from 2,8 x 105 to 2,6 x 107 on the 7th dpe and were 8,6 x 104 per ml on the 13th dpe (Figure 4B). Conversely, mosquitoes that were single-infected with CHIKV showed no positivity (Figure 4C,D). For the coinfected mosquitoes, a TR of 16.7% was observed for MAYV at both time points (Figure 4A), with RNA copy numbers/ml ranging from 3,9 x 104 to 2 x 106 on the 7th dpe and from 4,9 x 104 to 3 x 108 on the 13th dpe (Figure 4B). For CHIKV, a TR of 7.2% was noted on the 7th dpe (Figure 4C), with RNA copy numbers/ml ranging from 1.7 x 109 to 1,5 x 1010 (Figure 4D).
3.2. Superinfection Assay
CHIKV-superinfected mosquitoes presented a 70% and 93.3% IR on 6th dpe and on 12th dpe, respectively, for MAYV (Figure 5A), and an 83.3% IR on 12th dpe for CHIKV (Figure 6A). Furthermore, it revealed an 83.3% and 93.1% DR on 6th dpe and on 12th dpe, respectively, for MAYV (Figure 5C), and an 85.7% DR on 12th dpe for CHIKV (Figure 6C). Regarding the analysis of MAYV RNA copy number/ml, ranges from 5,6 x 106 to 1 x 1012 on 6th dpe and from 1x107 to 2 x 1012 on 12th dpe were found for midguts (Figure 5B), while ranges from 1 x 107 to 2,5 x 1012 on 6th dpe and 14 x 108 to 5,9 x 1013 were identified for carcasses (Figure 5D). Analysis of CHIKV RNA copy numbers/ml showed ranges from 3,8 x 108 to 1,4 x 1012 on 12th dpe for midguts (Figure 6B), and ranges from 1,3 x 107 to 1,3 x 1013 on 12th dpe were identified for carcasses (Figure 6D).
For mosquitoes superinfected with MAYV, the infection rates (IR) for CHIKV were 73.3% on the 6th dpe and 76.7% on the 12th dpe (Figure 6A). For MAYV, the IR was 7.3% on the 12th dpe (Figure 5A). The DR for CHIKV was 95.5% on the 6th dpe and 100% on the 12th dpe (Figure 6C), while the DR for MAYV was 71.4% on the 12th dpe (Figure 5C). Analysis of CHIKV RNA copy numbers/ml showed ranges from 2,4 x 106 to 5,4 x 1011 on the 6th dpe and from 4 x 105 to 3,1 x 1012 on the 12th dpe for midguts (Figure 6B). For carcasses, the ranges were from 3,6 x 106 to 9 x 1012 on the 6th dpe and from 3,3 x 107 to 5.7 x 1012 on the 12th dpe (Figure 6D). Regarding MAYV RNA copy numbers/ml, ranges from 4 x 107 to 2,3 x 1012 were observed on the 12th dpe for midguts (Figure 5B) and from 1,7 x 108 to 2,7 x 1013 for carcasses (Figure 5D).
Concerning the TR in the CHIKV-superinfection assay, MAYV was detected in 6.7% of the saliva on the 7th dpe (Figure 7A) (range from 2,5 x 109 to 1,8 x 1011 RNA copy number/ml) (Figure 7B), while MAYV-superinfected mosquitoes showed a 3.3% positivity for CHIKV on 13th dpe (Figure 7C) (3,9 x 1010 RNA copy number/ml) (Figure 7D).
4. Discussion
The current epidemiological landscape on a global scale has been characterized by numerous cases of arbovirus (re)emergence, accompanied by their rapid geographical dissemination [1]. This phenomenon has prompted the simultaneous circulation of diverse pathogens within particular regions [34]. In Brazil, alongside DENV, ZIKV, CHIKV, and YFV, Oropouche virus (OROV) has recently emerged as an increasingly notified virus, surpassing four thousand cases [35]. In this context, considering the potential emergence of MAYV in urban regions of Brazil, a country with the highest reported cases of MAYV human infections [9,10,11,12,18], and the favorable conditions for the simultaneous circulation of arboviruses- including the presence of competent vectors and susceptible vertebrate hosts [5]- it becomes imperative to comprehend the relationship among various viruses within the same vector [22]. This area of investigation remains full of gaps in the scientific literature, underscoring its critical significance [21]. Confronted with this scenario, we investigated the coinfection and superinfection consequences on the replication of CHIKV and MAYV in Ae. aegypti mosquitoes.
In coinfection scenarios, both CHIKV and MAYV replicate similarly to single infections, indicating minimal interference in the transmissibility of the viruses and, consequently, in vector competence. Previous studies of vector competence involving Ae. aegypti have shown this mosquito has the ability to become infected and transmit multiple viruses during a single blood meal, such as combinations of DENV, ZIKV, and CHIKV [36,37], ZIKV-MAYV [21], and, as observed in this study, CHIKV-MAYV [22], supporting the hypothesis that coinfection event does not significantly interfere with the amplification of the viruses [2,37,38]. On the other hand, we noted interference in the transmission of the viruses during the second blood meal in the superinfection experiments by CHIKV and MAYV. This shows the occurrence of a potential inhibition of viral replication. The group superinfected with CHIKV displayed a predominance of MAYV (first blood meal), as only MAYV was detected in saliva collected from the 7th dpe. Conversely, the group superinfected with MAYV displayed a prevalence of CHIKV (first blood meal), as supported by TR results in which only CHIKV was detected in samples collected on the 13th dpe. Previous studies have also found similar results. Kantor et al. [22] found interference in the transmission between MAYV and CHIKV in Ae. aegypti mosquitoes superinfected by CHIKV, similar to our findings. However, they did not find interference in the reverse scenario (mosquitoes superinfected by MAYV). In contrast, our study identified interference in mosquitoes superinfected by MAYV as well. Additionally, Abrao & Da Fonseca [26] reported, through in vitro superinfection studies, a reduced viral replication of Yellow Fever virus (YFV) in C6/36 cells previously infected with DENV-2 when compared to single infections by YFV [26].
Previous studies have discussed factors that may be associated with interference, inhibition, or even exclusion of viral replication in a superinfection scenario [26]. One key factor is the activation of the vector host's immune system by the primary viral infection, which can make it difficult for a subsequently acquired arbovirus to replicate [2,26]. Additionally, other factors could be involved, such as the vector's microbiome competition, a decrease in cellular resources required for the replication of the secondarily acquired virus, reduced binding of the virion to cell surface receptors during the second infection, and inhibition of the nucleocapsid uncoating related to the second infection [25,39,40]. However, this phenomenon remains somewhat ambiguous, as certain studies have indicated no interference in cases of superinfection. For instance, Kuwata et al. [41] demonstrated in vitro that a pre-existing infection with CxFV (an insect-specific flavivirus initially isolated from Culex spp.) did not interfere with Japanese Encephalitis virus (JEV) or DENV infection at the cellular (CTR cells) level. Similarly, Kent et al. [42] reported that prior exposure to CxFV had no noticeable effect on West Nile virus (WNV) replication in experiments conducted with C6/36 cells.
The potential for viral interference in superinfection scenarios has been extensively investigated, particularly in the context of insect-specific viruses (ISVs) [24]. These viruses can replicate exclusively in insect cells and can directly impact the infection, dissemination, and transmission of arboviruses, as demonstrated between Nhumirim virus (NHUV) and WNV in Culex quinquefasciatus [43], and Eilat virus (EILV) and CHIKV in Ae. aegypti [40]. These studies support the exploration of innovative methods for vector control [22,24,39,41]. From a different perspective, viral interference between mammalian pathogenic arboviruses may, to some extent, provide explanations for the infrequent or nonexistent occurrence of certain viruses in regions with a high prevalence of others [24,26]. Abrao & Da Fonseca [26] previously proposed this concept to account for the limited occurrence of YFV in areas with substantial DENV circulation. However, this hypothesis remains speculative, as it requires not only in vitro analyses but also in vivo and field validations for confirmation. Regarding our results, we observed interference in both pathways MAYV-superinfection and CHIKV-superinfection, which allows us to discuss a possible variable that contributes to the absence of MAYV emergence in Brazilian urban areas, characterized by a high circulation of CHIKV and a high density of Ae. aegypti [5], which, in turn, demonstrated to be competent for MAYV [15,44]. Similarly, the interference caused by an initial MAYV infection in mosquitoes might contribute to the comparatively low detection rates of CHIKV in the northern regions of Brazil [45]. This phenomenon is compounded by the fact that CHIKV has already adapted to urban environments, undergoing direct amplification cycles in humans [46], and only emerged in Brazil in 2014 [47], whereas MAYV has been circulating in the northern region since the 20th century [48].
It is important to note potential biases associated with the forced salivation technique, used in the present study, such as its non-standardization, even though it is considered the gold method for assessing transmission rates in vector competence studies without resorting to an animal model, and the non-quantification of saliva expulsion [49]. Given this context, it is crucial to conduct vector competence studies using animal models. Such studies should observe histopathologic effects and quantify viral presence in various tissues, as exemplified by Krokovsky et al. [15]. In our study, we observed low numbers of infected saliva across all tested groups, which implies the existence of a robust barrier within the mosquito's salivary gland against CHIKV and MAYV, despite a viral infection disseminated in the organism [38,50,51]. Low TR have also been observed in prior studies involving CHIKV and MAYV within Ae. aegypti. For instance, Göertz et al. [38] reported a TR of 21.2% at 14 dpe for mosquitoes single infected by CHIKV, using initial titers with log 107 TCID50/ml. In the same study, the TR was 14.6% for CHIKV in mosquitoes coinfected with ZIKV (72.9%), with an initial titer of log 107 TCID50/ml for both viruses [38]. Another study by Wiggins et al. [52] found TR of 10% and 23% at 6 and 12 dpe, respectively, for mosquitoes single infected by MAYV, using an initial viral titer with a log of 1010 PFU/ml.
Our findings indicate that in a coinfection scenario, both MAYV and CHIKV can replicate in Ae. aegypti without adversely affecting viral replication and, consequently, vector competence for both viruses. However, superinfection events by MAYV or CHIKV can disrupt the transmission of the secondary acquired virus. This highlights a notable epidemiological concern, particularly in Brazilian urban areas where multiple viruses, including DENV, ZIKV, CHIKV, YFV, and OROV, circulate, underscoring the critical importance of multiplex diagnostic methods for simultaneous detection of these co-circulating viruses. Therefore, these results bear significant consequences for shaping public health strategies in the region, emphasizing the intricate nature of arbovirus transmission scenarios and the related epidemiological risks.
5. Conclusions
Our study highlights the capability of MAYV and CHIKV to replicate in Ae. aegypti during coinfection scenarios, however, with observed interference in viral transmission during superinfection by MAYV or CHIKV events. Therefore, we underscore the importance of considering coinfection and superinfection dynamics in studies related to virus-vector interactions, entomological surveillance programs, and risk assessments in regions susceptible to arbovirus epidemics. The implications of coinfection and superinfection within the fields of epidemiology, pathogenesis, and viral evolution, both in mammalian hosts and mosquito vectors, remain underexplored. Future studies could delve into investigating the impact of these phenomena on the diversity of the viral genome and subsequent changes in the pathogenicity of diseases associated with these arbovirus infections.
Author Contributions
Conceptualization, M.H.S.P., D.R.D.G., L.K. and M.E.S.P.O; methodology, M.H.S.P., D.R.D.G., L.K. and M.E.S.P.O; formal analysis, M. J. B. C. and M.E.S.P.O; writing—original draft preparation, M.E.S.P.O; writing—review and editing, M.H.S.P., D.R.D.G., C.F.J.A., L.K. and M.E.S.P.O; supervision, M.H.S.P., C.F.J.A. and D.R.D.G.; project administration, M.H.S.P. and C.F.J.A.; funding acquisition, M.H.S.P., and C.F.J.A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially supported by the Fundação de Amparo à Pesquisa do Estado de Pernambuco (FACEPE): APQ-0230-2.02/20 to M.H.S.P. M.H.S.P. and C.F.J.A are supported by a productivity fellowship from the Brazilian National Council for Research and Development (CNPq) (09001/2019-6) and Fundação de Amparo à Pesquisa do Estado de Pernambuco (FACEPE) (APQ-1470-2.13/24).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Acknowledgments
We are grateful to the Instituto Aggeu Magalhães—Fundação Oswaldo Cruz (IAM/FIOCRUZ), Programa de Pós-Graduação em Biociências e Biotecnologia em Saúde. We would also like to thank the team at the Entomology Department at the Aggeu Magalhães Institute (AMI) for providing the mosquitoes. Also, we would like to thank the Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE) for the financing of the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weaver, S.C.; Reisen, W.K. Present and Future Arboviral Threats. Antiviral Res. 2010, 85, 328–345. [Google Scholar] [CrossRef]
- Vogels, C.B.F.; Rückert, C.; Cavany, S.M.; Perkins, T.A.; Ebel, G.D.; Grubaugh, N.D. Arbovirus Coinfection and Co-Transmission: A Neglected Public Health Concern? PLOS Biol. 2019, 17, e3000130. [Google Scholar] [CrossRef]
- Almeida, L.S.; Cota, A.L.S.; Rodrigues, D.F. Sanitation, Arboviruses, and Environmental Determinants of Disease: impacts on urban health. Cienc. Saude Coletiva 2020, 25, 3857–3868. [Google Scholar] [CrossRef]
- Santos, L.L.M.; de Aquino, E.C.; Fernandes, S.M.; Ternes, Y.M.F.; Feres, V.C. de R. Dengue, Chikungunya, and Zika Virus Infections in Latin America and the Caribbean: A Systematic Review. Rev. Panam. Salud Pública 2023, 47, e34. [Google Scholar] [CrossRef]
- Whiteman, A.; Loaiza, J.R.; Yee, D.A.; Poh, K.C.; Watkins, A.S.; Lucas, K.J.; Rapp, T.J.; Kline, L.; Ahmed, A.; Chen, S.; et al. Do Socioeconomic Factors Drive Aedes Mosquito Vectors and Their Arboviral Diseases? A Systematic Review of Dengue, Chikungunya, Yellow Fever, and Zika Virus. One Health Amst. Neth. 2020, 11, 100188. [Google Scholar] [CrossRef]
- Albuquerque, M. de F.; Souza, W.; Araújo, T.; Braga, C.; Filho, D.; Ximenes, R.; Filho, D.; Brito, C.; Valongueiro, S.; Melo, A.; et al. Epidemia de Microcefalia e Vírus Zika: A Construção Do Conhecimento Em Epidemiologia. Cad. Saúde Pública 2018, 34. [Google Scholar] [CrossRef] [PubMed]
- Souza, W.V. de; Albuquerque, M. de F.P.M. de; Vazquez, E.; Bezerra, L.C.A.; Mendes, A. da C.G.; Lyra, T.M.; Araujo, T.V.B. de; Oliveira, A.L.S. de; Braga, M.C.; Ximenes, R.A. de A.; et al. Microcephaly Epidemic Related to the Zika Virus and Living Conditions in Recife, Northeast Brazil. BMC Public Health 2018, 18, 130. [Google Scholar] [CrossRef]
- Weaver, S.C. Prediction and Prevention of Urban Arbovirus Epidemics: A Challenge for the Global Virology Community. Antiviral Res. 2018, 156, 80–84. [Google Scholar] [CrossRef]
- Acosta-Ampudia, Y.; Monsalve, D.M.; Rodríguez, Y.; Pacheco, Y.; Anaya, J.-M.; Ramírez-Santana, C. Mayaro: An Emerging Viral Threat? Emerg. Microbes Infect. 2018, 7, 163. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Luis, M.A.; del Valle-Mendoza, J.; Sandoval, I.; Silva-Caso, W.; Mazulis, F.; Carrillo-Ng, H.; Tarazona-Castro, Y.; Martins-Luna, J.; Aquino-Ortega, R.; Peña-Tuesta, I.; et al. A Silent Public Health Threat: Emergence of Mayaro Virus and Co-Infection with Dengue in Peru. BMC Res. Notes 2021, 14, 29. [Google Scholar] [CrossRef] [PubMed]
- Caicedo, E.-Y.; Charniga, K.; Rueda, A.; Dorigatti, I.; Mendez, Y.; Hamlet, A.; Carrera, J.-P.; Cucunubá, Z.M. The Epidemiology of Mayaro Virus in the Americas: A Systematic Review and Key Parameter Estimates for Outbreak Modelling. PLoS Negl. Trop. Dis. 2021, 15, e0009418. [Google Scholar] [CrossRef] [PubMed]
- Lima, W.G.; Pereira, R.S.; da Cruz Nizer, W.S.; Brito, J.C.M.; Godói, I.P.; Cardoso, V.N.; Fernandes, S.O.A.; Ferreira, J.M.S. Rate of Exposure to Mayaro Virus (MAYV) in Brazil between 1955 and 2018: A Systematic Review and Meta-Analysis. Arch. Virol. 2021, 166, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Parida, M.; Dash, P.K. Impact of Transmission Cycles and Vector Competence on Global Expansion and Emergence of Arboviruses. Rev. Med. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kain, M.P.; Skinner, E.B.; Athni, T.S.; Ramirez, A.L.; Mordecai, E.A.; van den Hurk, A.F. Not All Mosquitoes Are Created Equal: Incriminating Mosquitoes as Vectors of Arboviruses; Infectious Diseases, 2022.
- Krokovsky, L.; Lins, C.R.B.; Guedes, D.R.D.; Wallau, G. da L.; Ayres, C.F.J.; Paiva, M.H.S. Dynamic of Mayaro Virus Transmission in Aedes Aegypti, Culex Quinquefasciatus Mosquitoes, and a Mice Model. Viruses 2023, 15, 799. [Google Scholar] [CrossRef]
- Serra, O.P.; Cardoso, B.F.; Ribeiro, A.L.M.; Santos, F.A.L. dos; Slhessarenko, R.D. Mayaro Virus and Dengue Virus 1 and 4 Natural Infection in Culicids from Cuiabá, State of Mato Grosso, Brazil. Mem Inst Oswaldo Cruz 2016, 20–29. [Google Scholar] [CrossRef]
- de Curcio, J.S.; Salem-Izacc, S.M.; Pereira Neto, L.M.; Nunes, E.B.; Anunciação, C.E.; Silveira-Lacerda, E. de P. Detection of Mayaro Virus in Aedes Aegypti Mosquitoes Circulating in Goiânia-Goiás-Brazil. Microbes Infect. 2022, 24, 104948. [Google Scholar] [CrossRef]
- Esposito, D.L.A.; Fonseca, B.A.L. da. Will Mayaro Virus Be Responsible for the next Outbreak of an Arthropod-Borne Virus in Brazil? Braz. J. Infect. Dis. 2017, 21, 540–544. [Google Scholar] [CrossRef]
- Miralles, R.; Ferrer, R.; Solé, R.V.; Moya, A.; Elena, S.F. Multiple Infection Dynamics Has Pronounced Effects on the Fitness of RNA Viruses. J. Evol. Biol. 2001, 14, 654–662. [Google Scholar] [CrossRef]
- Muturi, E.J.; Bara, J. Sindbis Virus Interferes with Dengue 4 Virus Replication and Its Potential Transmission by Aedes Albopictus. Parasit. Vectors 2015, 8, 65. [Google Scholar] [CrossRef]
- Brustolin, M.; Pujhari, S.; Terradas, G.; Werling, K.; Asad, S.; Metz, H.C.; Henderson, C.A.; Kim, D.; Rasgon, J.L. In Vitro and In Vivo Coinfection and Superinfection Dynamics of Mayaro and Zika Viruses in Mosquito and Vertebrate Backgrounds. J. Virol. 2023, 0, e01778–22. [Google Scholar] [CrossRef]
- Kantor, A.M.; Lin, J.; Wang, A.; Thompson, D.C.; Franz, A.W.E. Infection Pattern of Mayaro Virus in Aedes Aegypti (Diptera: Culicidae) and Transmission Potential of the Virus in Mixed Infections With Chikungunya Virus. J. Med. Entomol. 2019, 56, 832–843. [Google Scholar] [CrossRef]
- Leroy, E.M.; Nkoghe, D.; Ollomo, B.; Nze-Nkogue, C.; Becquart, P.; Grard, G.; Pourrut, X.; Charrel, R.; Moureau, G.; Ndjoyi-Mbiguino, A.; et al. Concurrent Chikungunya and Dengue Virus Infections during Simultaneous Outbreaks, Gabon, 2007. Emerg. Infect. Dis. 2009, 15, 591–593. [Google Scholar] [CrossRef] [PubMed]
- Blitvich, B.J.; Firth, A.E. Insect-Specific Flaviviruses: A Systematic Review of Their Discovery, Host Range, Mode of Transmission, Superinfection Exclusion Potential and Genomic Organization. Viruses 2015, 7, 1927–1959. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.E.; Wan, K.; Bose, H.R. Homologous Interference Induced by Sindbis Virus. J. Virol. 1974, 14, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Abrao, E.P.; Lopes da Fonseca, B.A. Infection of Mosquito Cells (C6/36) by Dengue-2 Virus Interferes with Subsequent Infection by Yellow Fever Virus. Vector-Borne Zoonotic Diseases. 2016, 16, 124. [Google Scholar] [CrossRef] [PubMed]
- Melo-Santos, M. a. V.; Varjal-Melo, J.J.M.; Araújo, A.P.; Gomes, T.C.S.; Paiva, M.H.S.; Regis, L.N.; Furtado, A.F.; Magalhaes, T.; Macoris, M.L.G.; Andrighetti, M.T.M.; et al. Resistance to the Organophosphate Temephos: Mechanisms, Evolution and Reversion in an Aedes Aegypti Laboratory Strain from Brazil. Acta Trop. 2010, 113, 180–189. [Google Scholar] [CrossRef]
- Guedes, D.R.; Paiva, M.H.; Donato, M.M.; Barbosa, P.P.; Krokovsky, L.; Rocha, S.W. dos S.; Saraiva, K.L.; Crespo, M.M.; Rezende, T.M.; Wallau, G.L.; et al. Zika Virus Replication in the Mosquito Culex Quinquefasciatus in Brazil. Emerg. Microbes Infect. 2017, 6, 1–11. [Google Scholar] [CrossRef]
- Gloria-Soria, A.; Payne, A.F.; Bialosuknia, S.M.; Stout, J.; Mathias, N.; Eastwood, G.; Ciota, A.T.; Kramer, L.D.; Armstrong, P.M. Vector Competence of Aedes Albopictus Populations from the Northeastern United States for Chikungunya, Dengue, and Zika Viruses. Am. J. Trop. Med. Hyg. 2021, 104, 1123–1130. [Google Scholar] [CrossRef]
- Barbosa, P.P.; Guedes, D.R.D.; Melo-Santos, M. a. V.; Cordeiro, M.T.; Acioli, R.V.; Batista, C. a. V.; Gonçalves, L.S.M.; Souza, M.F.M.; Araújo, Y.V.; Magalhães, F.J.R.; et al. Vector Surveillance for Dengue Virus Detection in the Archipelago of Fernando de Noronha, Brazil. J. Med. Entomol. 2016, 53, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.; Kosoy, O.; Laven, J.; Panella, A.J.; Velez, J.; Lambert, A.; Campbell, G. Chikungunya Virus in US Travelers Returning from India, 2006. Emerg. Infect. Dis. 2007. [Google Scholar] [CrossRef]
- Naveca, F.G.; do Nascimento, V.A.; de Souza, V.C.; Nunes, B.T.D.; Rodrigues, D.S.G.; Vasconcelos, P.F. da C. Multiplexed Reverse Transcription Real-Time Polymerase Chain Reaction for Simultaneous Detection of Mayaro, Oropouche, and Oropouche-like Viruses. Mem. Inst. Oswaldo Cruz 2017, 112, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Krokovsky, L.; Paiva, M.H.S.; Guedes, D.R.D.; Barbosa, R.M.R.; de Oliveira, A.L.S.; Anastácio, D.B.; Pontes, C.R.; Ayres, C.F.J. Arbovirus Surveillance in Field-Collected Mosquitoes From Pernambuco-Brazil, During the Triple Dengue, Zika and Chikungunya Outbreak of 2015-2017. Front. Trop. Dis. 2022, 3. [Google Scholar] [CrossRef]
- Ciota, A.T. The Role of Co-Infection and Swarm Dynamics in Arbovirus Transmission. Virus Res. 2019, 265, 88–93. [Google Scholar] [CrossRef]
- Fujita, D.M.; Salvador, F.S.; da Silva Nali, L.H.; de Andrade Júnior, H.F. Oropouche in Brazil in 2024. J. Travel Med. 2024, taae075. [Google Scholar] [CrossRef]
- Nuckols, J.T.; Huang, Y.-J.S.; Higgs, S.; Miller, A.L.; Pyles, R.B.; Spratt, H.M.; Horne, K.M.; Vanlandingham, D.L. Evaluation of Simultaneous Transmission of Chikungunya Virus and Dengue Virus Type 2 in Infected Aedes Aegypti and Aedes Albopictus (Diptera: Culicidae). J. Med. Entomol. 2015, 52, 447–451. [Google Scholar] [CrossRef]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of Simultaneous Exposure to Arboviruses on Infection and Transmission by Aedes Aegypti Mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef] [PubMed]
- Göertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito Co-Infection with Zika and Chikungunya Virus Allows Simultaneous Transmission without Affecting Vector Competence of Aedes Aegypti. PLoS Negl. Trop. Dis. 2017, 11, e0005654. [Google Scholar] [CrossRef]
- Viglietta, M.; Bellone, R.; Blisnick, A.A.; Failloux, A.-B. Vector Specificity of Arbovirus Transmission. Front. Microbiol. 2021, 12, 773211. [Google Scholar] [CrossRef]
- Nasar, F.; Erasmus, J.H.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. Eilat Virus Induces Both Homologous and Heterologous Interference. Virology 2015, 484, 51–58. [Google Scholar] [CrossRef]
- Kuwata, R.; Isawa, H.; Hoshino, K.; Sasaki, T.; Kobayashi, M.; Maeda, K.; Sawabe, K. Analysis of Mosquito-Borne Flavivirus Superinfection in Culex Tritaeniorhynchus (Diptera: Culicidae) Cells Persistently Infected with Culex Flavivirus (Flaviviridae). J. Med. Entomol. 2015, 52, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Kent, R.J.; Crabtree, M.B.; Miller, B.R. Transmission of West Nile Virus by Culex Quinquefasciatus Say Infected with Culex Flavivirus Izabal. PLoS Negl. Trop. Dis. 2010, 4, e671. [Google Scholar] [CrossRef]
- Goenaga, S.; Kenney, J.L.; Duggal, N.K.; Delorey, M.; Ebel, G.D.; Zhang, B.; Levis, S.C.; Enria, D.A.; Brault, A.C. Potential for Co-Infection of a Mosquito-Specific Flavivirus, Nhumirim Virus, to Block West Nile Virus Transmission in Mosquitoes. Viruses 2015, 7, 5801–5812. [Google Scholar] [CrossRef]
- Long, K.; Ziegler, S.; Thangamani, S.; Hausser, N.; Kochel, T.; Higgs, S.; Tesh, R. Experimental Transmission of Mayaro Virus by Aedes Aegypti. Am. J. Trop. Med. Hyg. 2011, 85, 750–757. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, N.R.; Costa, A.R.M.; Feitosa, C.A.; Loth, T.P.; Klingelfus, A. A EVOLUÇÃO DE CASOS DE ARBOVIROSES DENGUE, CHIKUNGUNYA E ZIKA VÍRUS NO BRASIL ENTRE 2018 E 2020. Braz. J. Infect. Dis. 2022, 26, 101956. [Google Scholar] [CrossRef]
- Weaver, S.C.; Forrester, N.L.; Liu, J.; Vasilakis, N. Population Bottlenecks and Founder Effects: Implications for Mosquito-Borne Arboviral Emergence. Nat. Rev. Microbiol. 2021, 19, 184–195. [Google Scholar] [CrossRef]
- Cunha, M.S.; Costa, P.A.G.; Correa, I.A.; de Souza, M.R.M.; Calil, P.T.; da Silva, G.P.D.; Costa, S.M.; Fonseca, V.W.P.; da Costa, L.J. Chikungunya Virus: An Emergent Arbovirus to the South American Continent and a Continuous Threat to the World. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Causey, O.R.; Maroja, O.M. Mayaro Virus: A New Human Disease Agent. III. Investigation of an Epidemic of Acute Febrile Illness on the River Guama in Pará, Brazil, and Isolation of Mayaro Virus as Causative Agent. Am. J. Trop. Med. Hyg. 1957, 6, 1017–1023. [Google Scholar] [PubMed]
- Gloria-Soria, A.; Brackney, D.E.; Armstrong, P.M. Saliva Collection via Capillary Method May Underestimate Arboviral Transmission by Mosquitoes. Parasit. Vectors 2022, 15, 103. [Google Scholar] [CrossRef]
- Franz, A.W.E.; Sanchez-Vargas, I.; Adelman, Z.N.; Blair, C.D.; Beaty, B.J.; James, A.A.; Olson, K.E. Engineering RNA Interference-Based Resistance to Dengue Virus Type 2 in Genetically Modified Aedes Aegypti. Proc. Natl. Acad. Sci. 2006, 103, 4198–4203. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Dong, S.; Dizaji, N.B.; Rutkowski, N.; Pohlenz, T.; Myles, K.; Dimopoulos, G. The Aedes Aegypti siRNA Pathway Mediates Broad-Spectrum Defense against Human Pathogenic Viruses and Modulates Antibacterial and Antifungal Defenses. PLOS Biol. 2022, 20, e3001668. [Google Scholar] [CrossRef]
- Wiggins, K.; Eastmond, B.; Alto, B.W. Transmission Potential of Mayaro Virus in Florida Aedes Aegypti and Aedes Albopictus Mosquitoes. Med. Vet. Entomol. 2018, 32, 436–442. [Google Scholar] [CrossRef]
Figure 1.
Groups of mosquitoes analyzed in the single infection, coinfection and superinfection experiments: (A) Single infection with MAYV or CHIKV. (B) Coinfection with MAYV and CHIKV. (C) Superinfection with CHIKV. (D) Superinfection with MAYV.
Figure 1.
Groups of mosquitoes analyzed in the single infection, coinfection and superinfection experiments: (A) Single infection with MAYV or CHIKV. (B) Coinfection with MAYV and CHIKV. (C) Superinfection with CHIKV. (D) Superinfection with MAYV.
Figure 2.
Infection and Dissemination rates (%) for MAYV and number of MAYV RNA copies/ml in the single infection and coinfection assays. (A) Infection rate (IR); (B) Viral RNA copy number/ml in positive midguts; (C) Dissemination rate (DR); (D) Viral RNA copy number/ml in positive carcasses. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 samples of each tissue (midgut and carcass) for MAYV single-infected group and n=30 for MAYV and CHIKV coinfected group.
Figure 2.
Infection and Dissemination rates (%) for MAYV and number of MAYV RNA copies/ml in the single infection and coinfection assays. (A) Infection rate (IR); (B) Viral RNA copy number/ml in positive midguts; (C) Dissemination rate (DR); (D) Viral RNA copy number/ml in positive carcasses. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 samples of each tissue (midgut and carcass) for MAYV single-infected group and n=30 for MAYV and CHIKV coinfected group.
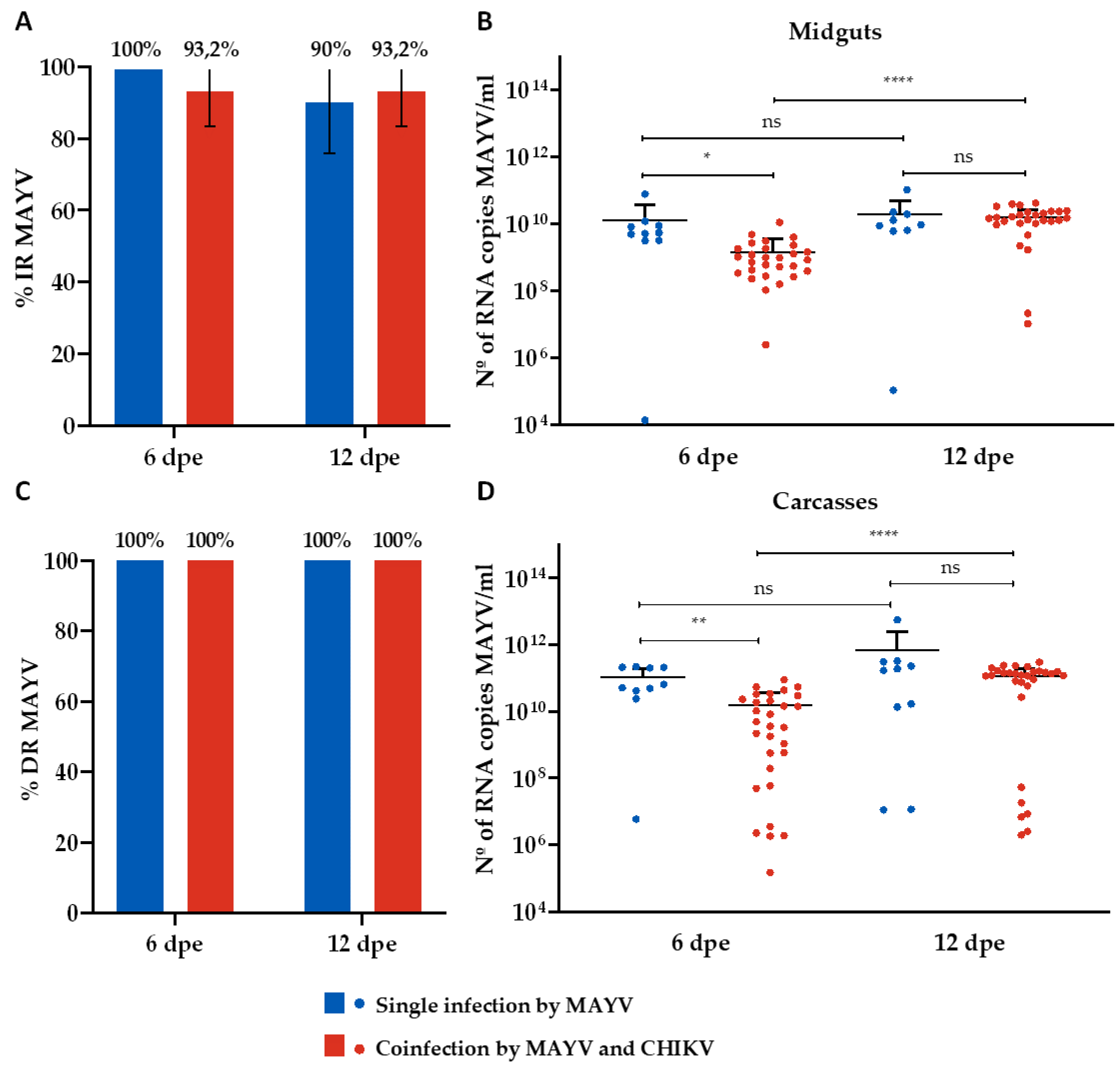
Figure 3.
Infection and Dissemination rates (%) for CHIKV and number of CHIKV RNA copies/ml in the single infection and coinfection assays. (A) Infection rate (IR); (B) Viral RNA copy number/ml in positive midguts; (C) Dissemination rate (DR); (D) Viral RNA copy number/ml in positive carcasses. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 samples of each tissue (midgut and carcass) for CHIKV single-infected group and n=30 for MAYV and CHIKV coinfected group.
Figure 3.
Infection and Dissemination rates (%) for CHIKV and number of CHIKV RNA copies/ml in the single infection and coinfection assays. (A) Infection rate (IR); (B) Viral RNA copy number/ml in positive midguts; (C) Dissemination rate (DR); (D) Viral RNA copy number/ml in positive carcasses. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 samples of each tissue (midgut and carcass) for CHIKV single-infected group and n=30 for MAYV and CHIKV coinfected group.
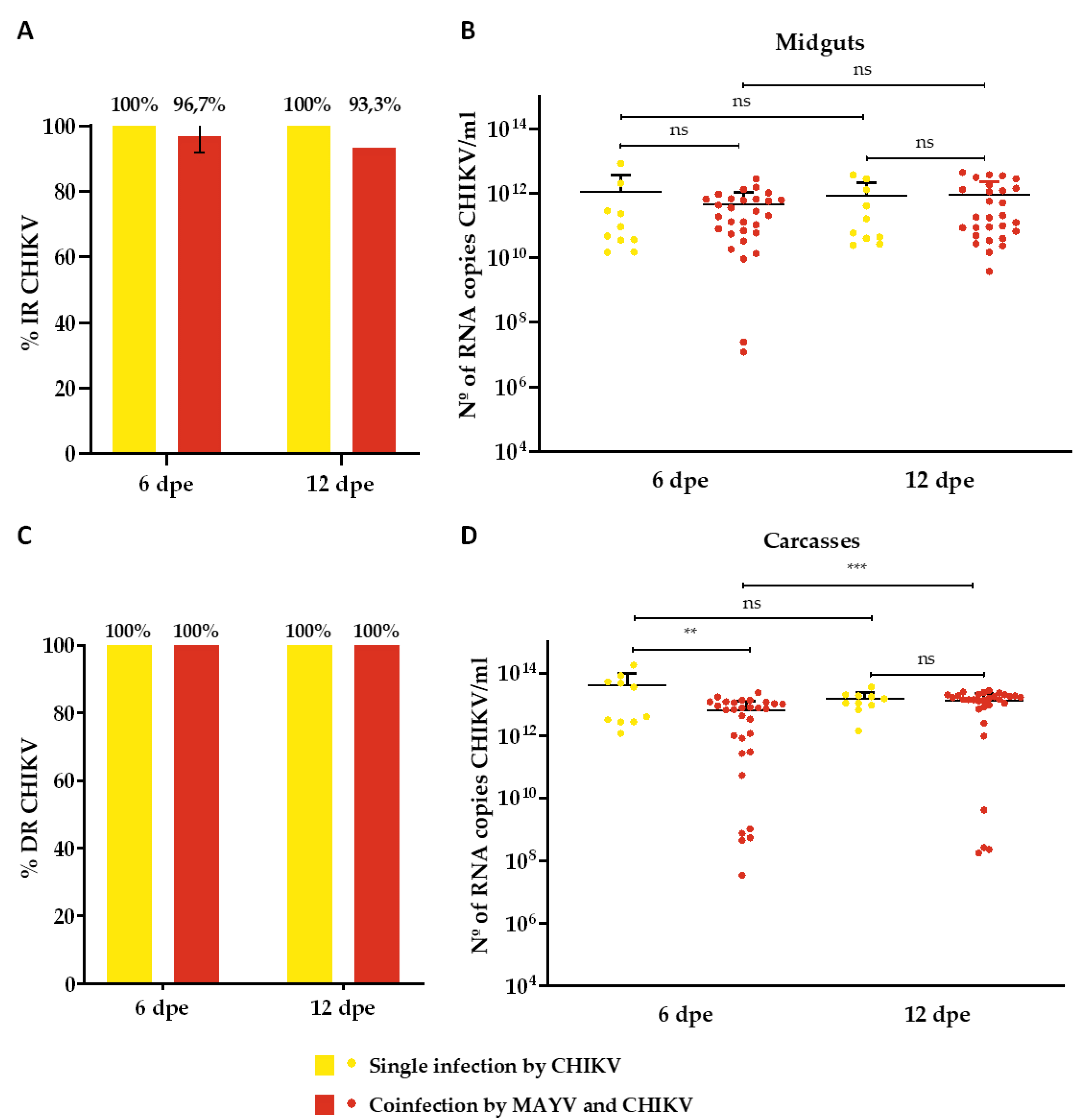
Figure 4.
Transmission rate (%) for MAYV and CHIKV and number of MAYV and CHIKV RNA copies/ml in the coinfection assay. (A) Transmission rate (TR) for MAYV; (B) MAYV RNA copy number/ml in positive saliva; (C) Transmission rate (TR) for CHIKV; (D) CHIKV RNA copy number/ml in positive saliva. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 saliva for MAYV and CHIKV single-infected groups and n=30 for MAYV and CHIKV coinfected group.
Figure 4.
Transmission rate (%) for MAYV and CHIKV and number of MAYV and CHIKV RNA copies/ml in the coinfection assay. (A) Transmission rate (TR) for MAYV; (B) MAYV RNA copy number/ml in positive saliva; (C) Transmission rate (TR) for CHIKV; (D) CHIKV RNA copy number/ml in positive saliva. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 saliva for MAYV and CHIKV single-infected groups and n=30 for MAYV and CHIKV coinfected group.
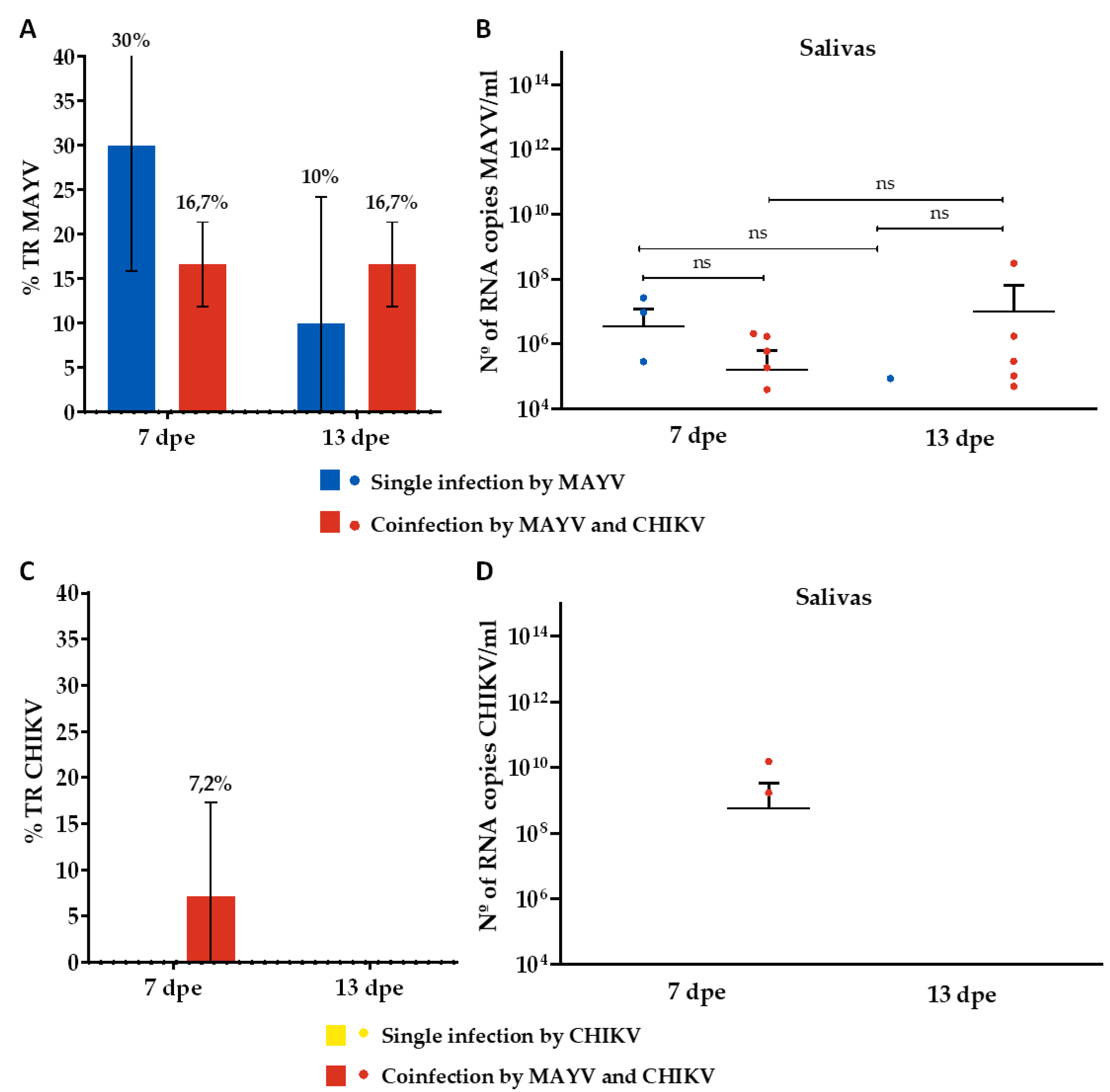
Figure 5.
Infection and Dissemination rates (%) for MAYV and number of MAYV RNA copies/ml in the superinfection assay. (A) Infection rate (IR); (B) Viral RNA copy number/ml in positive midguts; (C) Dissemination rate (DR); (D) Viral RNA copy number/ml in positive carcasses. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 samples of each tissue (midgut and carcass) for MAYV single-infected group and n=30 for MAYV or CHIKV superinfected groups.
Figure 5.
Infection and Dissemination rates (%) for MAYV and number of MAYV RNA copies/ml in the superinfection assay. (A) Infection rate (IR); (B) Viral RNA copy number/ml in positive midguts; (C) Dissemination rate (DR); (D) Viral RNA copy number/ml in positive carcasses. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 samples of each tissue (midgut and carcass) for MAYV single-infected group and n=30 for MAYV or CHIKV superinfected groups.
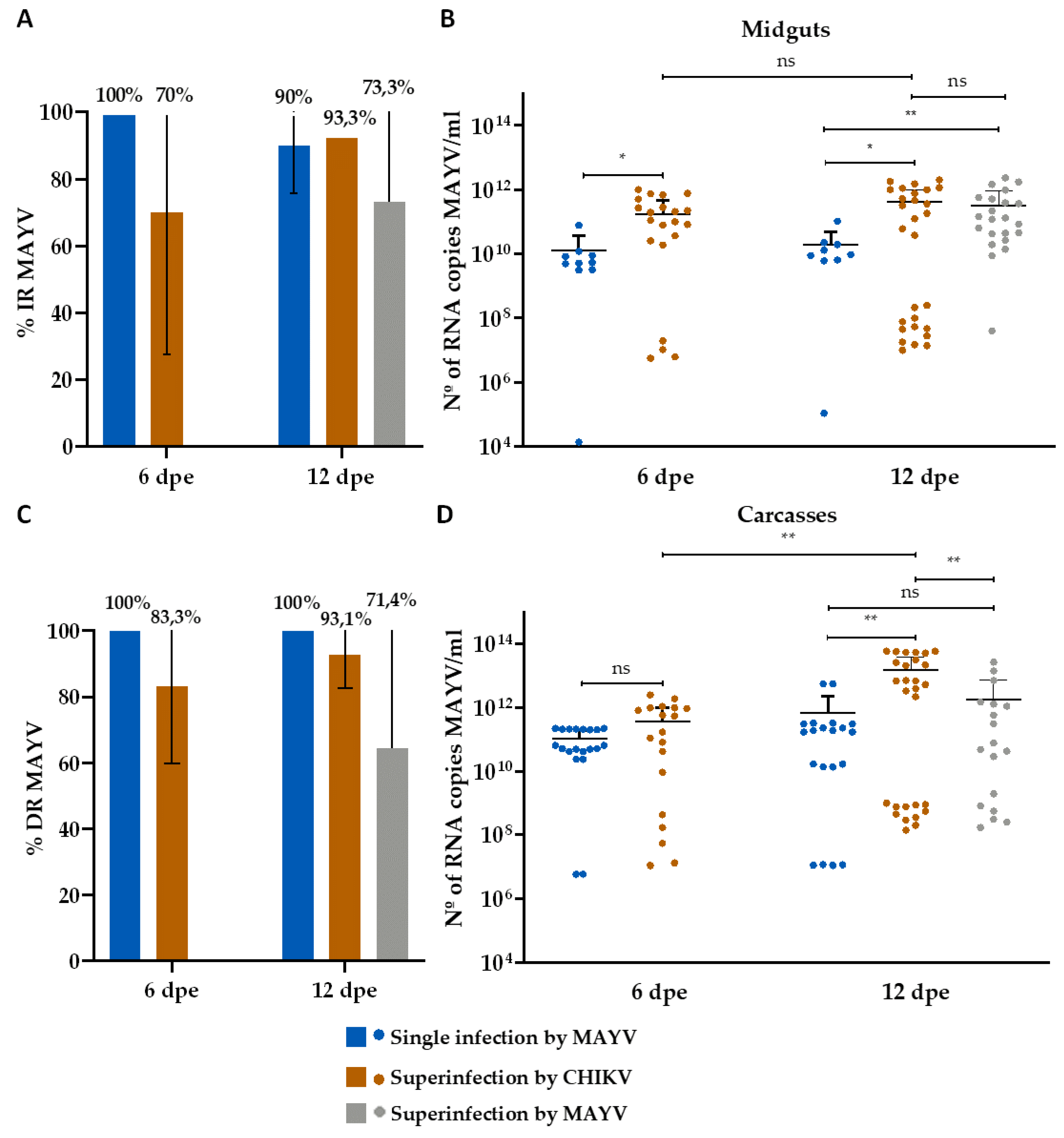
Figure 6.
Infection and Dissemination rates (%) for CHIKV and number of CHIKV RNA copies/ml in the superinfection assay. (A) Infection rate (IR); (B) Viral RNA copy number/ml in positive midguts; (C) Dissemination rate (DR); (D) Viral RNA copy number/ml in positive carcasses. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 samples of each tissue (midgut and carcass) for CHIKV single-infected group and n=30 for MAYV or CHIKV superinfected groups.
Figure 6.
Infection and Dissemination rates (%) for CHIKV and number of CHIKV RNA copies/ml in the superinfection assay. (A) Infection rate (IR); (B) Viral RNA copy number/ml in positive midguts; (C) Dissemination rate (DR); (D) Viral RNA copy number/ml in positive carcasses. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 samples of each tissue (midgut and carcass) for CHIKV single-infected group and n=30 for MAYV or CHIKV superinfected groups.
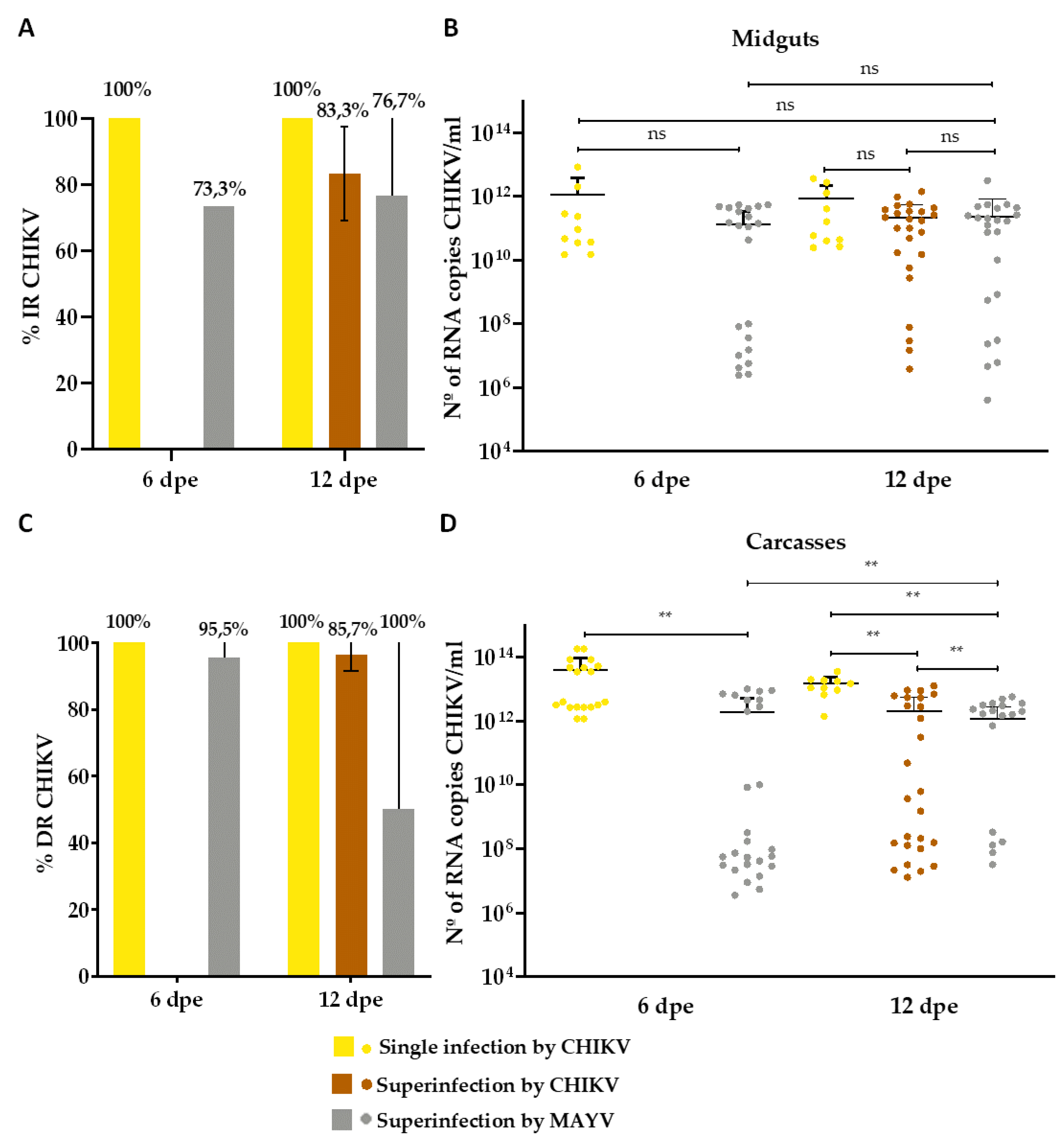
Figure 7.
Transmission rate (%) for MAYV and CHIKV and number of MAYV and CHIKV RNA copies/ml in the superinfection assay. (A) Transmission rate (TR) for MAYV; (B) MAYV RNA copy number/ml in positive saliva; (C) Transmission rate (TR) for CHIKV; (D) CHIKV RNA copy number/ml in positive saliva. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 saliva for MAYV and CHIKV single-infected groups and n=30 for MAYV or CHIKV superinfected groups.
Figure 7.
Transmission rate (%) for MAYV and CHIKV and number of MAYV and CHIKV RNA copies/ml in the superinfection assay. (A) Transmission rate (TR) for MAYV; (B) MAYV RNA copy number/ml in positive saliva; (C) Transmission rate (TR) for CHIKV; (D) CHIKV RNA copy number/ml in positive saliva. Dpe- days post-exposure; * p ≤ 0,05, ** p ≤ 0,01, *** p ≤ 0,001, **** p ≤ 0,0001, ns - non-significant. It was analyzed n=10 saliva for MAYV and CHIKV single-infected groups and n=30 for MAYV or CHIKV superinfected groups.
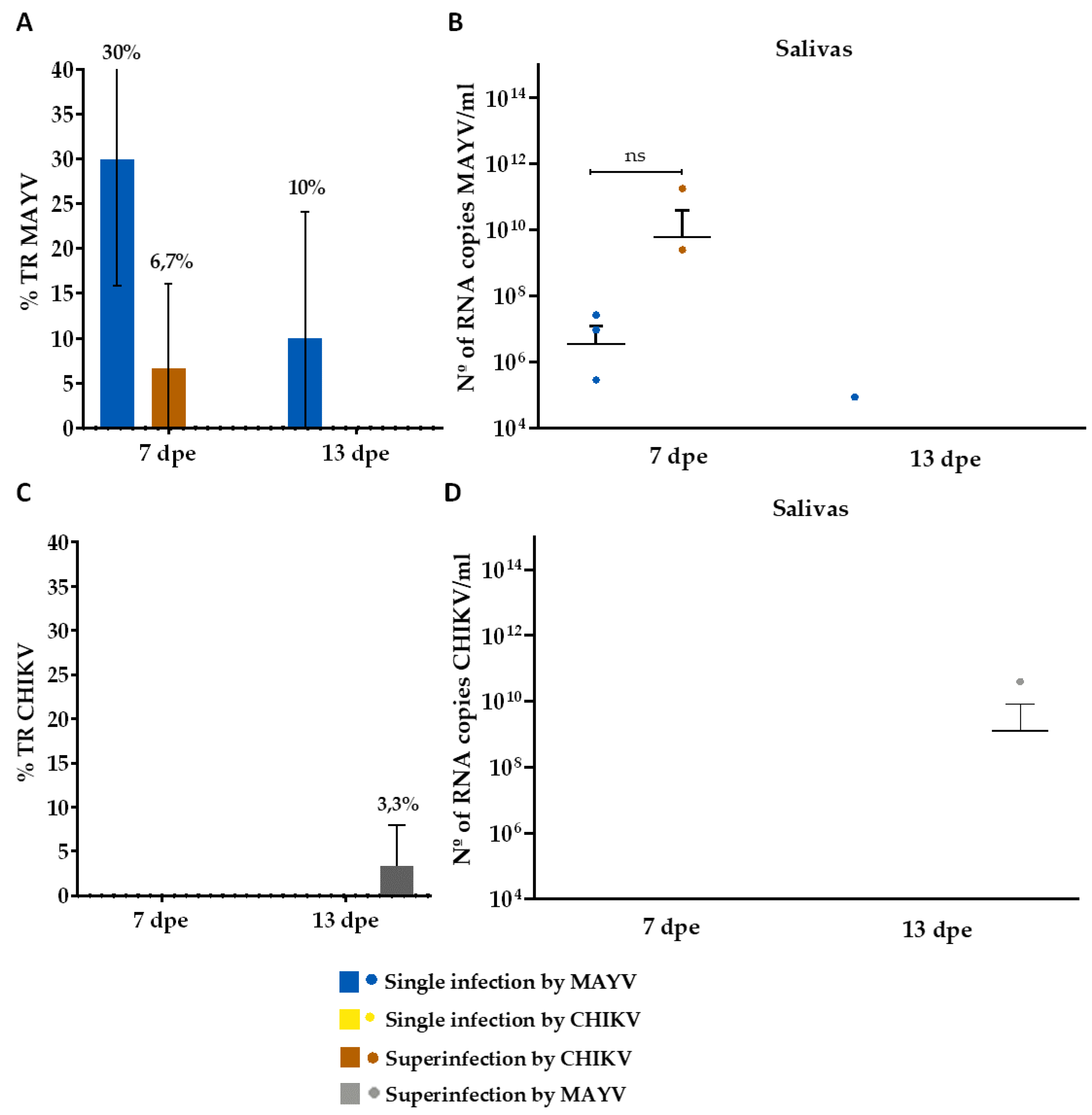

Table 1.
Average initial viral titer, measured in Plaque Forming Units (PFU), for MAYV and CHIKV used on the single infection, coinfection and superinfection assays.
Table 1.
Average initial viral titer, measured in Plaque Forming Units (PFU), for MAYV and CHIKV used on the single infection, coinfection and superinfection assays.
| Group | Virus | 1st blood-meal | 2nd blood-meal |
|---|---|---|---|
| Single infection- MAYV | MAYV | 5,5 x 107 | -* |
| Single infection- CHIKV | CHIKV | 3,1 x 107 | -* |
| Coinfection MAYV + CHIKV | MAYV | 5,5 x 107 | -* |
| CHIKV | 3,1 x 107 | -* | |
| Superinfection by MAYV | CHIKV | 1,4 x 107 | - |
| MAYV | - | 3,1 x 107 | |
| Superinfection by CHIKV | MAYV | 3,5 x 107 | - |
| CHIKV | - | 8,5 x 106 |
* non-infectious blood meal.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



