
Submitted:
20 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
Abstract: Conventional antibiotic and multidrug treatments become less and less effective and a discovery of new effective and safe antibacterial agents got a global priority. The returning to the natural antibacterial product is relatively new current trend. Terrestrial biota is a rich source of biologically active substances whose antibacterial potential was not fully utilized.
Aim of this review is to present the current state-of-the art of terrestrial biota-derived antibacterial agents and inspired by nature treatments. It is a summary of the most important sources and new identified or modified antibacterial agents and treatments during the last 5 years. It focuses on the significance of plants- animals- and bacteria-derived biologically active agents as powerful alternatives to antibiotics as well as on the advantages of utilizing natural antibacterial molecules alone or in combinations with antibiotics.
The main conclusion is that the terrestrial biota derived antibacterial products and substances open variety of new ways to modern improved therapeutic strategies. New terrestrial sources of known antibacterial agents and new antibacterial agents from terrestrial biota were discovered during the last 5 years, which are under investigation together with some long ago known but now surviving their renaissance for the development of new medical treatments. Use of natural antibacterial peptides as well as combinational therapy by commercial antibiotics and natural products outlines as the most promising for treating of bacterial infections. In vivo testing and clinical trials are necessary to reach clinical application.
Keywords:
terrestrial biota
; anti-bacterial products
; antibacterial compounds
; bacteriocines
; biosynthesized nanoparticles
; combination treatments
1. Introduction
Increasing resistance to the traditional antibiotics is expanding across the world at an alarming rate. Expecting to be the next global pandemic, antimicrobial resistance (AMR) is already recognized as one of the top-most serious threats to human health, [1,2].
The conventional antibiotic and multidrug treatments become less and less effective. Every year, various microbial pathogens cause infectious diseases of a large number of humans with lethal issue for some of them. It was estimated that the bacterial AMR was directly responsible for 1.27 million and contributing to 4.95 million global deaths in 2019 [2]. The first World Health Organization (WHO) regional assessment on health predicts that 5.2 million people in the Western Pacific Region will die between 2023 and the end of 2030 because of drug-resistant bacterial infections, [3]. Without new and better treatments, the global deaths number is expected to rise up to 10 million by 2050, that is higher than cancer (8.2 millions) and diabetes (1.5 millions) death, [4,5,6,7]. In addition to death and disability, the AMR has significant economic costs. The World Bank estimates that AMR could result in losses of gross domestic product at 1 trillion to 3.4 trillion USD per year by 2030 and at 1 trillion USD additional healthcare costs by 2050, [8]. According to 2021, WHO report the pool of new antimicrobial compounds in clinical testing is limited. Only six of thirty-two antibiotic agents in the clinical establishment (that meet the WHO listing of critical pathogens) were categorized as novel in the year 2019, [9]. During the period 2019-2023, FDA approves, according to our checking, the following antibacterial drugs: Likmez, Voquezna, Twyneo, Xaciato, Zilxi, Recarbrio, and Fetroja.
In early 2017, the WHO published Priority Pathogens List of bacteria that are most frequently involved in multidrug-resistant infections. The updated 2024 WHO Bacterial Priority Pathogens list (WHO BPPL) covers 24 pathogens, spanning 15 families, which require the greatest action with regard to supplementary antibacterial therapies. Among them are Gram-negative bacteria resistant to last-resort antibiotics: drug-resistant mycobacterium tuberculosis, and other high-burden resistant pathogens such as Salmonella, Shigella, Neisseria gonorrhoeae, Pseudomonas aeruginosa, and Staphylococcus aureus, [10].
Global concern for human morbidity and mortality caused by multi drug-resistant organisms requires urgent development of creative and innovative approaches, from chemical identification and analysis to assessment of bioactivity of new antimicrobial agents, [11,12,13].
Many investigations are devoted to understanding the mechanism of different bacterial species drug resistance and thus to help the development of new approaches mitigating the problems of the AMR, [1,14,15,16,17,18,19]. Mir et al. [20] and lately, Helmy et al. [21] discuss alternative of antibiotic approaches that might be used to control AMR, including probiotics, prebiotics, antimicrobial peptides, small molecules, organic acids, essential oils, bacteriophages, fecal transplants, and nanoparticles. With the ability to kill Gram-negative and Gram-positive bacteria or to inhibit their growth, numerous antibacterial peptides (ABPs) have already demonstrated a potential as novel therapeutic agents. However, as drugs, they have some undesirable properties including instability, toxicity, etc. that should be overcome for their clinical translation, [22].
The lack of good quality antibacterial drugs remains a major problem and the discovery of effective and safe antibacterial agents got already a global priority. Natural biologically active products play a vital role in human survival for millennia. The returning to them is relatively new current trend. The interest to natural biologically active product increases because of their high efficiency, structural diversity, offering potential lead compounds, low toxicity and miss or delay development of resistance. Antibacterial activity toward Gram-negative and Gram-positive bacteria of many natural compounds isolated from plants, animals and microorganisms was experimentally proved [23] but the clinical developments are not enough, [24].
Natural antimicrobial and anti-biofilm agents from marine biota, including antioxidant active substances [25,26,27,28,29] as well as terrestrial biota derived anti-biofilm agents were a subject of our former studies.
The aim of this review is to present the current state-of-the art of terrestrial biota-derived antibacterial agents and inspired by nature treatments. It is a summary of the most important sources and new identified or modified antibacterial agents and treatments during the last 5 years up to now. This review is focused on the significance of herbs, plants- animals- and bacteria-derived biologically active agents as powerful alternatives to antibiotics as well as the advantages of utilizing natural antibacterial molecules alone or in combination with antibiotics. It is expected this review to aid the future development of more effective antibacterial agents and approaches to the combating the rising number of bacterial infections. It includes natural antibacterial agents derived from plants, animals and bacteria, their experimentally found or machine (in silico) predicted antibacterial activity as well as potential advantages, limitations and disadvantages as well as modifications to reduce the last ones. It is expected this review to aid the future development of more effective antibacterial drugs and approaches to combating the rising number of bacterial infections.
2. Approaches to Avoid/Mitigate the Bacterial Resistance
The medical applications of terrestrial biota derived antibacterial agents are associated with treatment of antibiotic resistant bacterial infections. Therefore, currently used or under study approaches and potential places in them of natural products, to overcome/mitigate antibacterial resistance are shortly presented here, [11].
As it is evident in Figure 1, the antibacterial approaches include: reducing a overuse/misuse of antibiotics [11]; drug repurposing [30]; treatments by nanomaterials and use of nanodelivery systems [31]; application of natural antibacterial agents, bacteriophage therapy, immunomodulation, etc., [32]; molecular docking (ethno-pharmacological approaches; synthetic strategies inspired by nature; chemical modifications of existing natural antimicrobials, semi-synthesis; computer-aided design, etc.) [11,33]; combinational therapy (combinations of phytochemicals and antibiotics; antibiotics and adjuvants) [34,35,36,37,38,39].
2.1. Use of Natural Antibacterial Agents: Plant Products, Antibacterial Peptides, Bacteriocines and Bacteriophages
The study of natural products as alternative or adjuvants to the current antibiotics becomes more and more popular approach in medical treatments due to their generally low toxicity and chemical diversity, which provides important therapeutic effects and make the microbes unable to copy themselves for creating resistance, [23]. Bacteriophage therapies and peptide therapies are studied as novel options to control the development of multidrug resistance (MDR). CRISPR, an innovative genome editing technology, offers multiple safeguard applications to overcome different challenges of the MDR. Immunotherapy is a way to improving the host defence and combating issues of bacterial drug resistance using mainly plant products, [11].
2.2. Combination Drug Therapy
This approach includes combination of antibiotics and phytochemicals or antibiotics and adjuvants, etc. Herbal extracts, essential oils and isolated pure compounds are reported to act synergistically with existing antibiotics and to increase their activity, [34]. Theoretical and practical framework are outlined for the development of effective combinations of antibiotics and antibiotics with non-antibiotic compounds, [35,36]. Combination therapies of existing antibiotics and adjuvants are accepted as promising for mitigation the AMR problems, [37,38].
The combination of different drugs offers many advantages over their use as individual chemical moieties, namely reduction of dosage; fewer side effects; reduced risk for the development of drug resistance; better combined response; wide-spectrum antibacterial action; and ability to attack simultaneously multiple target sites. An appropriate combination approach provides a pathway to development of antimicrobial therapeutics with broad-spectrum antibacterial action, bactericidal instead of bacteriostatic mechanisms of action, and better efficacy against multidrug-resistant bacteria, [39].
2.3. Nanotechnologies
Nanotechnologies are emerging, widely popular approaches for treatment of multi drug-resistant infectious diseases. The use of nanomaterials, mainly biologically synthetized metal, metal oxide and composites nanoparticles as well as based on nanocarriers drug delivery systems is a relatively new trend to improvement of pharmacokinetics, stability and solubility, to reducing toxicity and providing controlled release of therapeutic agents or other compounds at the target site. Some nanoparticles, extracted from natural products, such as Glycyrrhiza glabra L. green tea, Allium sativum L. and Ginkgo biloba L. demonstrate god antimicrobial potential, [40]. Liposome-based nanoparticles can restore the potency of antibiotics such as ceftazidime, imipenem, and cefepime against multidrug-resistant P. aeruginosa, amikacin for K. pneumoniae, and chloramphenicol for MRSA-65 by effective drug administration, [31,40,41].
2.4. Molecular Docking Inspired by Nature
Inspired by nature chemical synthesis, chemical modifications of existing natural antimicrobials, semi-synthesis, and computer-aided design are relatively new approaches to development of novel antibacterial agents. The antimicrobial characteristics of some naturally occurring chemicals produced by bacteria, plants and animals are now studied for using as lead molecules, [42]. Structure-guided drug discovery [43] and ethno-pharmacological approaches also contribute to the molecular docking. Their goals are to update knowledge on natural antimicrobial products and their ethno-medicinal uses in preventing and treating infections as well as future research directions for the discovery of new antibiotics from natural products, [44]. For example, the efficacy of developed in 1930s, maggot debridement therapy (involving the use of maggots of the green-bottle fly) of wounds to remove necrotic, sloughy and/or infected tissue, now is studied on S. aureus and P. aeruginosa in diabetic foot ulcers [45]. The ethnobotany and plant natural products are discussed currently as a promising source of antibacterial lead compounds that could help the new drug discovery, [46].
The demand of new antibiotics: antimicrobial peptides, nanoparticles, combinatorial therapies and structure-guided drugs is emerging as the most important future strategy in the design of antibacterial agents, including such that can selectively interact with a target site (a gene or a cellular process) or a specific pathogen, [39]. Natural antimicrobial agents acting on different mechanisms are under development now to attack the developed by the bacteria immune defense elements and mechanisms, such as virulence factors, [47]; immune system elements [48,49,50] or; to directly inhibit bacterial immune system, [51] or; targeting oxidative stress in bacteria [52], etc.
3. Terrestrial Sources of Antibacterial Agents
Terrestrial sources of antibacterial agents are plants, animals and even bacteria (Figure 2). The most promising natural compounds used to combat bacteria include plant extracts, essential oils, and small antimicrobial peptides of animal origin, bacteriocins and various groups of plant compounds (triterpenoids; alkaloids; phenols; flavonoids) with antimicrobial and antiviral activity, [23].
Antioxidants such as polyphenols, vitamins, and carotenoids, derived from natural sources and dominantly involved in boosting of the defence system of the organisms are accepted as natural antibiotics, [53]. Inspired by the nature solutions are promising for discovery of new antibiotics and antibacterial treatments, [54]. Currently, natural products continue to be one of the most essential resources of modern therapeutics since they have a wide range of chemical and functional variability as well as fewer side effects, synergy, and capacity to combat drug tolerance. Secondary metabolites, synthesized by plants, animals and microbes can be used to isolate biologically active molecules, [55].
Natural products provide a significant number new chemical compounds that exhibits biological or pharmacological properties with therapeutic characteristics (lead compounds) and integral components of currently accessible drug formulations. The two crucial elements that propel natural materials from precursors to medicines are pharmacological activity and drug ability. Therefore, structural traits are under investigation for their adjustment to the corresponding application, [1]. Both short-term and long-term solutions are proposed to overcome the limitations in various research sectors with aim to bridge the gap between academic, industrial and political stakeholders, and to unite interdisciplinary expertise in benefit of development of future generations antibiotics, [56].
Antibacterial potential of natural products isolated from plants, animals and bacteria, either alone or in combination with conventional antibiotics against multidrug resistant pathogens is of especial interest. Prebiotics, probiotics, synbiotics, bacteriophages, nanoparticles, and bacteriocins are presented now as new tools supporting the progress of effective antibiotics to combating antibiotic-resistant bacteria, [57]. The development of bacterial vaccines is also of current interest, [58].
As it is evident from Figure 2, plants originating antibacterial agents include plant extracts, essential oils and fatty acids as well as antibacterial compounds: phenols, alkaloids, terpenoids, flavonoids and peptides. The animal originating antimicrobial agents are mainly peptides. Bacteriocines and bacteriophages originate from bacteria. Biogenic synthesis is a relatively new source of antibacterial nanoparticles. All these antibacterial agents and their sources are presented more detailed in the next sections.
3.1. Plant-Derived Antibacterial Products
Since ancient time, the world uses plants as medicine. Herbs, herbal components, and materials containing several parts of plants or other plant-based compounds are traditionally used to treat multiple health ailments. Medicinal plants, including garlic (Allium sativum), ginger (Zingiber officinale), green tea (Camellia sinensis), St. John’s wart (Hypericum perforatum), black cumin (Nigella sativa), licorice (Glycyrrhiza glabra), Mongolian milkvetch (Astragalus membranaceus), and purple coneflower (Echinacea spp.), possess a notable history of efficacy in managing of microbial diseases. Exhibiting noticeable immune-boosting properties and potential to combat bacterial pathogens these plants are thoroughly searched and effectively utilized, [59].
The use of herbal antibacterial (clove, portulaca, tribulus, eryngium, cinnamon, turmeric, ginger, thyme, pennyroyal, mint, fennel, chamomile, burdock, eucalyptus, primrose, lemon balm, mallow, and garlic), extracts or derived active components, instead of synthetic chemical drugs is increasing because of their less side effects, strong antimicrobial properties and primary healthcare benefits, [60]. Five herbal antimicrobials: echinacea, manuka, thyme, olive leaf and astragalus are reported as fighting infections naturally, [61]. Echinacea, also known as coneflower, is called “toothache plant” in many Native American cultures, because of its antibiotic and antimicrobial action. Experts for treating of toothaches, [60], recommend infusing a few drops of fresh leaves of this herb to create a mouthwash concoction. Ginger, clove, garlic and turmeric can also be used in different ways to reduce tooth infections, [62].
Strain-specific activity was found for curcumin (from Curcuma longa L.), that is for a long time known for its antibacterial properties. Experimental results confirm a much greater sensitivity of Gram-positive than Gram-negative bacteria and suggest that numerous clinical strains of widespread pathogens have poor sensitivity to curcumin (the MICs of the multi drug resistant types of Staphylococcus aureus, S. haemolyticus, Escherichia coli, and Proteus mirabilis are high, ≥ 2000 µg/mL). However, curcumin is effective against some species and strains: Streptococcus pyogenes (median MIC = 31.25 µg/mL), methicillin-sensitive S. aureus (250 µg/mL), Acinetobacter lwoffii (250 µg/mL), and individual strains of Enterococcus faecalis and Pseudomonas aeruginosa (62.5 µg/mL). Based on these results, the curcumin is classified as a promising antibacterial agent with a very selective activity, [63]. The curcumin blocks bacterial growth owing to its structural characteristics and the generation of antioxidant products. It can inhibit bacterial virulence factors and bacterial biofilm formation and prevents bacterial adhesion to host receptors through the bacterial quorum sensing regulation system. As a photosensitizer, the curcumin acts under blue light irradiation to induce photo toxicity and inhibit bacterial growth. Moreover, it can exert a synergistic antibacterial effect with other antibacterial substances, [64]. In-silico design and mechanistic study of niosomes (non-ionic surfactant-based vesicles) encapsulated curcumin suggest that it is active against multidrug-resistant Staphylococcus aureus biofilms and can be extended to be active against biofilms of other pathogens, [65].
Simultaneously with direct antibacterial activities, some phytoproducts demonstrate in vitro synergistic effects in combination with conventional antibiotics, [66]. It is generally agreed that medicinal plants could contribute in avoiding the crisis with microbial resistance to conventional antibiotics and multidrug treatments because of their attractive features. The plants are readily available and cheap, extracts or compounds from plants often demonstrate high activity against bacterial pathogens and they rarely have severe side effects. The large variety of plant-derived compounds provides diverse chemical structures that may supply both novel mechanisms of antimicrobial action and new targets within the bacterial cell. The rapid development of modern biotechnologies opens a ways for obtaining of bioactive compounds in environmentally friendly and low-toxic conditions, [67]. The rich and unique chemo diversity, the worldwide distribution and ease access, the various antibacterial modes of action, and the proved clinical effectiveness of plant extracts are the major strength of plant natural products as a promising source of antibacterial lead compounds, [45].
Plant-derived antibacterial products: extracts, oils, fatty acids, biosurfactants and large variety of biologically active compounds are under intense study for their potential to fight the AMR as adjuvants to antibiotics or as led molecules for development of new, more efficient antibacterial drugs.
3.1.1. Antibacterial Plant Extracts
Antibacterial plant extracts are prepared on technologies whose simple sketch is presented in Figure 3. The extraction technologies usually include several steps, staring with grinding of plant parts followed by crude extracts preparation, separation by sequential extraction, purification, evaporation, analysis and collection of the obtained antibacterial material. In some cases, grinding is to a very fine powder that can be directly applied as antibacterial material. In most cases, the grinded powder passes crude extraction using different solvents/solvent systems. Together with the type of the extraction technology and its parameters, the solvents are very important for the total yield and the composition of the extract. Some crude extracts demonstrate high antibacterial activity and after purification, drying and analysis they are used as antibacterial agents. The drying could be avoided if the crude extract will be used in liquid form.
The crude extracts contain a mixture of natural biologically active compounds and therefore they usually pass sequential extraction to isolate different components of the mixture. Sequential extracts are further processed similarly of the crude extracts: thy pass purification, evaporation (if it is necessary) and analysis to obtain the final product, [68].
Plant extracts often demonstrate high activity against bacterial pathogens. The screening of the antimicrobial potential of four different plant extracts against twelve pathogenic microorganisms and two reference bacterial strains demonstrates that the most extracts exhibit different antimicrobial activity in vitro. The extracts of Oxalis corniculata are most efficient against Gram-negative bacteria (Escherichia coli, Salmonella Typhi, Klebsiella pneumoniae, and Citrobacter koseri. The methanol extracts of Artemisia vulgaris, Cinnamomum tamala, and Ageratina adenophora show efficacy against Staphylococcus aureus, [69].
In vitro and in vivo evaluation of anti-salmonella effect of pectin extracts and hydrolysates from "Cas mango" (Spondias dulcis) show inhibition zones of 12.0 to 15.0 mm (for a pectin solution of 100 μg/ ml, by disk diffusion test) and MIC values for the different strains of Salmonella spp. of 5.68 to 44.45 μg/ml. The treatment with these extracts of mice infected by Salmonella spp. prolongs their lifespan, [70].
Inhibition of Staphylococcal pathogenesis by Witch-Hazel and green tea extracts [71], antimicrobial activity of Hibiscus acid and chromatographic fractions from Hibiscus Sabdariffa Calyces against multidrug-resistant pathogenic bacteria [72], as well as antibacterial potential of ethanol extracts of neem leafs (Azadirachta indica) against uropathogens producing beta-lactamase enzymes [73] were found by in vitro tests.
Influence of extraction solvent on the phenolic profile and bioactivity of two Achillea species was shown. The ethanol extract of A. abrotanoides Vis. (yarrow) demonstrates more significant antimicrobial activity against Gram-positive bacteria Enterococcus faecalis than the antibiotic ampicillin. The very high activity against E. faecalis is ascribed to the estimated high concentration of the flavanone, naringenin, in the ethanolic extract, [74].
The antimicrobial properties of Kalanhoe pinnata extract, bioactive compounds content and its mode of action against pathogenic microorganisms were studied aimed at further exploration as an alternative medicine to the current synthetic antibacterial drugs, [75].
The phytochemical constituents of ethanol extract of Sida acuta leafs and their effects on pathogenic Escherichia coli, Staphylococcus aureus, Pseudomonas aeruginosa and Bacillus subtilis bacterial species were determined using commercial ciprofloxacin as control. The screening reveals that ethanol extract of S. acuta leafs possesses secondary metabolites such as alkaloids, flavonoids, phenols, tannins, terpenoids, glycosides and cardiac glycosides. The extract exhibits significant inhibitory effects against the test bacteria. The highest antibacterial activity is exhibited at the highest concentration of the extract (300mg/ml). The minimum inhibitory concentrations (MIC) and minimum bactericidal concentrations (MBC) were found at 37.5 and 75mg/ml, respectively against all reference isolates of the test bacteria, [76].
Nasution et al. [77] aim to discover potential of herbal plants as natural antibiotic candidates by a machine learning approach based on traditional Indonesian herbal medicine system called Jamu (Jamu is a mixute of several herbs). The input data consists of a list of herbal formulas with their plants constituents. The target class corresponds to bacterial diseases that can be cured by herbal formulations. The best model was those implementing Random Forest (RF) algorithm. This study shows that 14 plants can be potentially used as natural antibiotic candidates. Furthermore, according to scientific journals, 10 of the 14 selected plants have direct or indirect antibacterial activity.
The screening of the antimicrobial activity of Camellia japonica flowers extracts demonstrates their potential as antibacterial agents with promising applications in the pharmaceutical and food industries. Extracts were obtained using 60 % methanol as a solvent and both, conventional and cost effective maceration method (50°C, 1 h). The results of agar diffusion assay reveal significant antimicrobial activity against S. aureus (10.29 mm), P. aeruginosa (9.24 mm), and Salmonella enteritidis (6.95 mm). However, the extracts do not exhibit activity against E. coli, S. epidermidis, and B. cereus, unlike other varieties of C. japonica that displayed activity against these microorganisms, [78].
The composition, antioxidant, antimicrobial and cytotoxic characteristics of methanol and ethanol extracts from leaves of the plants, both Juniperus sabina and Ferula communis (Cyprus) were reported in the year 2023. Total phenolic and flavonoids content of methanol and ethanol extracts were quantified. Momeinositol is the predominant component in the J. Sabina’s extracts. The most dominant component of F. communis ethanol extract is phytol, while in methanol extract it is 1,3,4,5 tetrahydroxycyclohexanecarboxylic acid. Antioxidant activity (by DPPH free radical-scavenging ability) testing reveals concentration dependent activity for methanol and ethanol extracts from the plant leaves. Sufficient antibacterial activity (by disk diffusion and minimal inhibitory concentration methods) of the plant extracts was found against Gram-negative and Gram-positive bacteria, [79].
The shrubby plant Opuntia stricta is known to have medicinal properties due to its phytochemical composition. Impact of different solvents (80% ethanol, 80% methanol and 80% acetone (v/v) in water) on extraction yield, phenolic composition, in vitro antioxidant and antibacterial activities of deseeded O. stricta fruit was investigated. It appears that the extraction solvents influence significantly the total phenolic content (TPC), total flavonoids content (TFC) and antioxidant capacity. The 80% ethanol shows highest percentage of extraction yield while 80% acetone extract shows the lowest extraction yield but the highest TPC, TFC and antioxidant activity. The antibacterial tests demonstrates that both, 80% methanol and 80% acetone extracts exhibit the highest inhibition zones (of 11.66 and 11.33 mm respectively, p > 0.05) against Salmonella thyphimurium. The 80% acetone extract demonstrates the best inhibitory effect against Escherichia coli (inhibition zone of 11 cm). Less sensitivity to all O. stricta extracts was observed of Gram-positive bacteria than that of Gram-negative bacteria, [80].
The bioactive components and phytochemicals of the methanol extract of Rhanterium epapposum oliv. (locally known as Al-Arfaj), belonging to the family Asteraceae, were studied using Agilent gas chromatography-mass spectrometry (GC-MS). The methanol extract of R. epapposum aerial parts shows presence of sixteen compounds. The determined phytochemicals in this extract were saponins, flavonoids, and phenolic compounds. A quantitative analysis reveals the presence of high content of flavonoids, total phenolic, and tannins. These data made possible use of R. epapposum aerial parts as herbal remedy for various diseases, [68].
It is reported that extracts from some endemic plants demonstrate different antibacterial activity against Gram-negative and Gram-positive bacteria. For example, the antibacterial potential of methanol extracts from three Iran endemic species of Acantholimon family (Acantholimon austroiranicum Rech.f. & Schiman-Czeika, Acantholimon serotinum Rech.f. & Schiman-Czeika and Acantholimon chlorostegium Rech.f. & Schiman-Czeika) is reported to be significantly higher toward Gram-negative bacteria (E. coli and P. aeruginosa) than against Enterococcus faecalis and S. aureus, [81].
Methanol and macerated methanol extracts of Anabasis aretioides, Morocco and Algeria endemic plant, widely used in traditional medicine, demonstrate antimicrobial activity against Proteus mirabilis, Bacillus subtilis, S. aureus and P. aeruginosa, [82].
Good antibacterial activity of extracts and oils of Doronicum macrolepis, an endemic plant from Turkey, was found to be due to the high phenolic content of ethyl acetate extract. The essential oil shows inhibitory effect on E. coli, S. epidermidis, Enterococcus faecium, Yersinia pseudotuberculosis, C. albican and C. tropicalis, [83].
Özcan et al. [83] investigate the administration of inhalable liposomes loaded with licorice extract (Glycyrrhiza glabra) for the treatment of tuberculosis. In vivo lung deposition studies of liposomal dry powder for inhalation (LDPI) in mice show that nearly 46% of the administered drug reach the lungs and 16% of the administered drug remain in the lungs after 24 h of administration. In vivo pharmacodynamic evaluation of the LDPI against Mycobacterium tuberculosis shows a significant reduction of bacterial counts in the lungs and spleen. Ginkgo biloba L. extract was also encapsulated in nanoparticles and shows good bioavailability, [84].
3.1.2. Essential Oils and Fatty Acids
A number of publications highlights the antimicrobial activity of essential oils (EOs) and their chemical constituents, including against drug-resistant microbial pathogens. Concerning the mode of action, it was shown that EOs primarily destabilize the cellular architecture of bacterial pathogens that leads to breakdown of membrane integrity and disruption of many cellular activities, including energy production and membrane transport. Membrane rupture induced by essential oils can lead to leakage of cellular components and loss of ions, [85]. Among reported 250 commercially available essential oils, about a dozen possess high antimicrobial potential and the antimicrobial activities of EOs are not pre-eminent for all strains. Therefore, further investigations should be focused on the targeting of EOs and microorganisms, [86].
Terpenes and their derivatives comprising of hydrocarbons, that are usually found in EOs are reported as having potential antimicrobial activity, exhibiting bacteriostatic or bactericidal effects against test pathogens and possible mechanisms exerted by each terpene class are discussed, [87]. The predominant antibacterial mechanism of action exhibited by EOs derived from polyphenol- and terpene-rich plants (such as Cuminum cyminum, Mentha piperita, Thymus daenensis, Pimenta dioica, Myrtus communis), is the disruption of the membrane function and the structure of bacterial cells and others. EOs, particularly those derived from the Lamiaceae and Verbenaceae families commonly found in the Mediterranean region, exhibit anti-quorum sensing and anti-biofilm properties against bacterial pathogens. Moreover, secondary metabolites could interfere with intermediary metabolism, as well as to disrupt DNA/RNA synthesis and functionality, and modulate critical events within the pathogenic progression, [88].
The antibacterial activity of fatty acids (FAs) is accepted as a promising option for developing of next-generation broad-spectrum antibacterial agents. Originating from the defense system of living organisms, FAs combined with other antibacterial agents usually demonstrate a remarkable ability to enhance their bactericidal properties. In vitro testing the antibacterial activity of eight plant oils (non hydrolyzed and hydrolyzed) containing medium-chain fatty acids (palm, red palm, palm kernel, coconut, babassu, murumuru, tucuma, and cuphea oil) against Gram-positive pathogenic bacteria clearly demonstrates the selective effect of the hydrolyzed forms of tested oils. While the hydrolyzed oils were active against all tested bacteria (Clostridium perfringens, Enterococcus cecorum, Listeria monocytogenes, and S. aureus) the same oils do not show any effect on commensal bacteria (Bifidobacterium spp. and Lactobacillus spp.). Tucuma and Cuphea seed oils show the strongest antibacterial activity. Non-hydrolyzed forms of all tested oils exerted no antibacterial effect against any test bacteria. This study creates a basis for the development of selective bacterial inhibitors, [89].
The profile analysis (GC-MS) of FAs of tengkawangplant (Shorea Sumatrana) oil indicates the presence of palmitic acid, stearic acid, oleic acid, oleic acid chloride, stearic acid chloride, glycidyl stearate, diethyl phthalate, and 2-monopalmitin with predominance of stearic acid (60.68%). The diameter of the inhibition zone against S. enteritidis, E. coli, S. aureus, and B. cereus at a concentration of 12.5%, 25%, 50% (by disc diffusion test and tetracycline as a positive control), shows that the tengkawang oil is an antibacterial agent with concentration optimum at 25% with more susceptibility to Gram-positive than Gram-negative bacteria, [90]. The study of antibacterial properties of FAs and how their chemical structures influence the antibacterial activity aims to better understanding of both traditional and non-traditional mechanisms involved in the antibacterial activities of FAs, [91]. Omega-3 fatty acids, offering enormous nutritional benefits, are shown as therapeutically significant in treating of several infectious diseases. To avoid the undesirable odor and flavor, heavy metal contamination, extinction of fish species, etc. of fish produced FAs oleaginous microorganisms were studied as a promising alternative for the production of a more sustainable, consistent and quality production of Omega-3 fatty acids, [92]. Unsaturated FAs acting as vancomycin adjuvants are reported to rapidly killing a range of Gram-positive bacteria, including vancomycin-tolerant and resistant populations (S. aureus and other Gram-positive bacteria). Synergistic bactericidal activity relies on the accumulation of membrane-bound cell wall intermediates that generate large fluid patches in the membrane leading to protein delocalization, aberrant septal formation, and loss of membrane integrity, [93].
Although their promising antibacterial activity, some plant products have drawbacks that limit their therapeutic use. For instance, allicin, berberine, curcumin, emodin, linalool, oleanolic acid, quercetin, and thymol are poorly bioavailable when are administered as single compound therapies. Other disadvantages of plant-derived compounds are a high volatility (e.g., linalool), low chemical stability (e.g., quercetin), pungent odor (e.g., allicin) and toxicity (e.g., sanguinarine). Structural modifications and/or loading them in drug delivery systems are two of the possible ways to avoid such drawbacks, [46].
3.1.3. Propolis and Honey
Resins, waxes, polyphenols, polysaccharides, volatile materials and secondary metabolites presenting in the composition of propolis are responsible for its bioactivities including antibacterial. These compounds positively modulate the antimicrobial resistance of multidrug resistant bacteria. Published research indicates that the propolis and its derivatives contain many natural antimicrobial compounds that enhance the efficacy of conventional antibiotics. The activity of propolis strongly depends on seasonal and regional factors. In combinations with honey, propolis demonstrates a synergistic effect against E. coli and S. aureus. Clinical trials are necessary to better elucidate the potential application of propolis and its main flavonoid ingredients in various medical fields, [94].
For centuries, honey has been used as a natural remedy to several types of illnesses and to treat wounds. Honey is known as very effective in almost all cases of infections and in the improvement of healing especially in burn injury and wounds. Periodically, reviews cover the antibacterial activity of honey, its use in the treatment of infections and diseases as well as the features that are relevant to its activity, [95,96]. The antibacterial activity of honey is ascribed to the presence of various components (such as sugars, polyphenol compounds, hydrogen peroxide, 1,2-dicarbonyl compounds, and bee defensin-1) at different concentrations (depending on the source of nectar, bee type, and storage) that work synergistically. The effectiveness of honey depends on its botanical origin, the health of the bee, and processing method. The application of antibiotics with honey yields better antimicrobial potential. In medicine, honey were used in the treatment of surface wounds, burns, and inflammation, and demonstrates a synergistic effect when applied with antibiotics. Tissue repair is enhanced by the low pH of honey (3.5–4), that causes a reduction in protease activity on the wound site, elevating of oxygen release from hemoglobin and stimulating of fibroblast and macrophage activity. Furthermore, H2O2 has antiseptic effects, and it disinfects the wound site and stimulates production of vascular endothelial growth. Honey can promote fast healing, reduces scarring and is very convenient for plastic surgery. In non-infected areas, the honey reduces pain and inflammation. In general, the use of honey in medical settings could reduce economic loss and provide economic benefits by lowering direct costs in comparison to conventional treatments and by using less antibiotics, faster healing and less hospitalization stay, [97]. Honey contains reactive oxygen species that are naturally produced by the body of flight insects and destroy bacteria. However, the honey is sticky and difficult to apply in the correct dose. Therefore research team of Sophie Cox [98] is working on ways for delivering a sterile honey product to wounds, including spray, cream, and powder making it effective in surgery, war zones and potentially in all of our homes, [99].
3.1.4. Plants-Derived Antibacterial Compounds
Plants have two major groups of metabolites: primary and secondary. Carbohydrates and lipids are products of the primary metabolism while phenolic compounds, carotenoids, alkaloids, saponins, and terpenoids are considered as secondary metabolites, [66]. Numerous plants-derived chemical compounds with significant antibacterial activity were already identified, belonging to different classes: triterpenoids; alkaloids; phenols; flavonoids and large group antimicrobial peptides (defensins, cathelicidins, cecropins, melitin, thionins, nisin and mitacin), [88]. In the most cases, the bioactive plant extracts contain complex mixtures of these groups, and their combined action can yield an enhanced effect, [66]. For example, comparative metabolite analysis of organs of Piper sarmentosum (Piperaceae), a traditional medicinal and food plant widely distributed in the tropical and subtropical regions of Asia, indicates 154 tentatively identified metabolites with a predominant occurrence of flavonoids, lignans and phenyl propanoids in the leaves, aporphines in the stems, piperamides in the fruits and lignan-amides in the roots. These extensive data on the metabolite composition of P. sarmentosum supply useful information for a bioactive compounds discovery and patterns of their preferential biosynthesis or storage in specific organs. They can be used to optimize the production and to maximize the plant’s economic value as herbal medicine, [100].
Plant-derived compounds possess many interesting biological properties combined with multiple antimicrobial activity, [101,102]. The antibacterial properties of medical plants derived substances, such as alkaloids, tannins, flavonoids, peptides, etc. are periodically discussed as a base for development of new medicines, [59]. Systematical literature review of plant-derived compounds with experimentally proved antibacterial activity for the period of 2012 to 2019, shows that of totally 459 compounds, 50.8 % are phenolic derivatives, 26.6 % are terpenoids, 5.7 % are alkaloids, and 17 % are classified as other metabolites. Only 10 of them have been in clinical trials, [46]. The major chemical classes and sub-classes antibacterial plant-derived natural products are presented in Figure 4.
According to current literature review, major phytochemical classes with potent antibacterial activity include phenolic compounds, alkaloids, saponins, terpenoids and others, [88].
3.1.4.1. Phenolic Compounds
More than 8000 bioactive phenolic compounds are known now, including phenolic acids and aldehydes, flavonoids, chalcones, benzophenones, xanthones, stilbenes, benzoquinones, and polyphenols, among others, which can be extracted from different parts of the plant, such as the leaves, roots, and fruits (bark and seeds). Experimentally was found that these compounds are more effective against Gram-positive bacteria. This could be explained by the presence of a thick peptidoglycan layer and the absence of an external membrane in them. Such outer membrane presents in Gram-negative bacteria (Figure 5) and exerts a hydrophobic action, preventing the penetration of hydrophilic molecules, such as phenolic compounds into the bacterial cells.
Main mechanism of action of the phenolic compounds is associated with their ability to reduce the expression of efflux pumps. However, there are molecules, like tannins and anthraquinones that are reported to inhibit DNA gyrase (and thus, capable to inhibit microbial growth), [88].
Resveratrol is a plant compound that tends to be concentrated mostly in the skins and seeds of grapes and berries. It acts as polyphenolic antioxidant receiving attention for its potential antimibacterial, anti-carcinogenic and anti-aging properties. Resveratrol displays in vitro antimicrobial activity against a surprisingly wide range of bacterial, viral and fungal species. In combination with conventional antibiotics, resveratrol enhances the activity of aminoglycosides against Staphylococcus aureus, [103]. Phenolic compounds from extracts of Hibiscus acetosella were reported to inhibit the growth of S. aureus, in addition to its efficiency against P. aeruginosa growth, [104]. Prophylactic efficacy against bacterial infections by driving phagocyte influx was found for a non-bactericidal cathelicidin, [105].The available information on different types of plant phenolic compounds was systematized with emphasis on their extraction, analysis and potential biological activity, [106].
Novel cathelicidin from Hydrophis cyanocinctus with antimicrobial and anti-inflammatory activity was identified and characterised in year 2023, [107]. The instability of phenolic compounds to light and oxygen raises some difficulties for their extraction that could be avoided by use of adequate modern extraction, concentration and separation technologies, [108].
Flavonoids are a family of widely distributed natural phenolic compounds, produced as secondary metabolites of plants in response to diverse biotic and abiotic factors. Some of the highest amounts of flavonoids are in berries, apples, citrus fruit (oranges, lemons, grapes), spinach, legumes, kale, broccoli, soybeans, onions, tea, and cocoa, [109,110]. The family of the flavonoid includes more than 6000 low molecular weight phenolic compounds that are derivatives of flavan. Flavonoids exhibiting strong antibacterial, anti-inflammatory, antioxidant, antiplatelet and other effects are reported, [111]. In some cases, flavonoids (especially chalcones) show up to six fold stronger antibacterial activities than standard drugs on the market. Some synthetic derivatives of flavonoids also exhibit remarkable antibacterial activities against multidrug-resistant Gram-negative and Gram-positive bacteria (including E. coli, P. aeruginosa and S. aureus): 20- to 80-fold higher than the activity of the standard drugs, [49,112]. The naturally occurring flavonoid trans-cinnamaldehyde and its derivatives demonstrate in some cases, improved antibacterial activity compared to that of the commonly used antibiotics, [113].
Chalcones are a class of flavonoids known as having antibacterial, anti-inflammatory, antifungal, antioxidant, cytotoxic, antitumor and chemo preventive activities. The largest number of natural chalcones was isolated from species of the Leguminosae, Asteraceae and Moraceae families, [114]. Chalcones, containing α,β-unsaturated ketone fragment, are accepted as an important pharmacologically active agents because of their diverse mechanisms of antibacterial action. The relationships structure-biological activity and action mechanisms provide some important guidance for development of more candidate - antibacterial agents, [115].
Progress in the development of environmentally friendly methods for extraction of flavonoids from plant materials was presented. It includes ultrasound, pressurized liquid extraction, mechano-chemical, high hydrostatic pressure, supercritical fluid, negative pressure cavitation, intensification of vaporization by decompression to the vacuum, microwave, infrared, pulsed electric field, high-voltage electrical discharges, deep eutectic solvents and enzyme-assisted extraction as well as the impact of the operation conditions, [116,117]. The current advanced technologies for flavonoids extraction from natural sources are faster, more environmentally friendly, and with higher automation levels, compared to conventional extraction techniques, [118].
Experimental design combined with modeling could reduce the number of experiments that should be performed to achieve maximum extraction yield, [119]. Based on the single factor experiment and Plackett-Burman design results, the microwave-assisted extraction of flavonoids from Phyllostachys heterocycla leaves was further optimized, using the response surface methodology, [120]. The investigation of the effects on total flavonoid content, flavonoid composition, and stability in jujube extracts, prepared by five extraction methods: (i) water-water bath, (ii) ethanol-water bath, (iii) deep eutectic solvent, combined with ultrasound-assisted extraction (DES-UAE), (iv) microwave-assisted extraction, and (v) enzyme-assisted extraction, demonstrates that DES-UAE would be an most efficient method for flavonoid extraction from jujube, [121]. Effect of extraction solvents (hexane, ethyl acetate and methanol) on phenolic compounds and flavonoids from different parts of pongame oil tree (Derris indica) (that could be a potential source to develop pharmaceutical products) against bacterial pathogens demonstrates that the ethyl acetate extract is with highest content of phenolic compounds and flavonoids, [122]. Analysis of flavonoid compounds isolated from the leaves of Akalifa (Acalypha wilkesiana Muell. Arc.) by maceration and methanol solvent extraction, addition of ethyl acetate and partitionation with n-hexane, proves that the flavonoid is an isoflavone, [123].
3.1.4.2. Alkaloids
More than 12,000 alkaloid compounds isolated from plant extracts are already known with medicinal actions, such as antibacterial, antitumor and analgesic (morphine and codeine) properties, [124]. The alkaloids have chemical structure with heterocyclic rings containing N-heterocyclic nitrogen and could be classified according to their carbon precursors and structure. Examples of alkaloid compounds, commonly found in plants include pyridine, piperidine, quinoline, alkaloidal amines, and terpenoids, [88]. Tryptanthrin is a natural alkaloid with indoloquinazoline content. This alkaloid could be synthesized chemically and could be derived from natural sources like plant extracts or cell cultures, including yeast. Its good anti-cryptococcal activity and a synergistic effect in combination with calcineurin inhibitors in vitro, are less expressed in vivo due to a poor blood brain barrier penetration, [125]. Good antimicrobial action of benzyltetrahydroisoquinolin alkaloids, derived from the leaves of Doryphora aromatica was demonstrated against methicillin resistant isolates of Mycobacteria spp. and S. aureus, [126].
3.1.4.3. Terpenoids
Terpenoids, or terpenes, are a class of metabolites that encompass a variety of natural substances, which have in common the presence of C5 isoprene units in their chemical structure. Depending on the amount of C5 isoprene involved in their synthesis, terpenes are monoterpenoids, sesquiterpenoids, diterpenoids, sesterterpenoids, and triterpenoids. More than 40,000 terpenoid substances are known with different applications: pharmaceutical, aromatic, agricultural, and industrial, [88]. Antibacterial action of terpene compounds from Eremophila lucida was demonstrated against S. aureus isolates, [127]. Terpenoids from Commiphora resin were isolated and identified with good antibacterial action against sensitive and resistant isolates of Mycobacterium tuberculosis, [128]. Andrographolide, a terpene derivative from Andrographis paniculata, was reported as able to inhibit invasive microbe virulence factors and regulate the host immunity. Controlled clinical trials reveal that A. paniculata treatment is safe and efficient against acute respiratory tract infections like common cold and sinusitis. Therefore, it was considered that A. paniculata and andrographolide, could be accepted as excellent candidates for antimicrobial drug development, [129]. Pech-Puch et al. [130] verify the good (MIC in the range of 1–8 µg/mL) and moderate (MIC value of 16 µg/mL) antimicrobial action of diterpene alkaloids from Agelas citrina against the Gram-positive pathogens S. aureus, S. pneumoniae, and E. faecalis and the Gram-negative pathogens A. baumannii, P. aeruginosa, and K. pneumoniae.
Saponins were found in a variety of plants. They are chemically characterized with presence of glycosylated groups, formed by a hydrophilic and a lipophilic part. This structure confers detergent and surfactant properties of the saponins, [66]. It is known that the chemical structure of the saponins directly interferes with the effectiveness of their antimicrobial action. Saponins with tri-saccharide chains exhibit good antifungal action, whereas saponins with mono- or di-saccharide chains did not show good antimicrobial action, [131].
3.1.4.4. Other Compounds
Lipids (essential oils, fixed oils, sterols, waxes, phospholipids, and fat-soluble vitamins) are other class of naturally occurring compounds. They are categorized as primary metabolites, nevertheless studies show that they have secondary metabolite functions, [132].
The pelargonic acid micelles, extracted from tomatoes, were found to demonstrate antimicrobial efficacy against Salmonella that varies by the surfactant, strain serotype and stress response, [133]. A recently published report confirms the antibacterial activity of garlic and onions, exhibiting inhibitory effects on diverse bacteria. The inhibitory effect is due to their abundant sulfoxide contents imparting them with antimicrobial properties. On the other hand, the horseradish, mustard seeds, and wasabi demonstrate inhibition activity that is attributed to their elevated levels of allyl glucosinolates, [88].
Seven bioactive compounds (berberine, catechin, chelerythrine, cinnamaldehyde, ellagic acid, proanthocyanidin, and sanguinarine) originating from plants were presented lately with antibacterial activity against Staphylococcus aureus, Enterococcus spp., Klebsiella pneumoniae, Acinetobacter baumannii, Escherichia coli, Serratia marcescens and Pseudomonas aeruginosa, as natural antimicrobial agents for the treatment of wound infections, [134].
It is thought that the utilization of plant derived phenolic compounds, alkaloids, saponins, terpenoids etc. as effective antibacterial drugs, could be optimized by omics technologies and network pharmacology in order to identify optimal combinations among these compounds or in conjunction with antibiotics, [88].
3.1.4.5. Possible Antibacterial Modes of Action of Plant-Derived Antibacterial Agents
Most natural antibacterial agents appear to affect the bacterial membrane permeability, leading to membrane rupture and cell lysis. However, not all mechanisms of action were elucidated and sometimes the mechanism may be indirect, stimulating the host immune system or inhibiting adhesion to the host cell. Due to the structural differences between Gram-negative and Gram-positive bacteria (Figure 5) the efficacy of antimicrobial agents varies, [135,136].
Generally, the antibacterial properties of medicinal plants are hypothesized to be connected to two mechanisms: chemical interference with the synthesis or functioning of most important bacterial components and/or bypassing the conventional mechanisms of antibacterial resistance. The possible antibacterial modes of action of plant-derived antibacterial agents are illustrated in Figure 6.
They include: the immune system activation; increase permeability; disruption of membrane function and structure; interference with intermediary metabolism; potential alternation; anti-quorum sensing and anti-biofilm activity and restore physiological balance. The last one is a holistic mechanism that is totally absent in the modern antibiotic treatment. Plants may alter the physiological balance of the body and to make it more resistant to pathogens, whereas in the modern medicine, the drugs are in a form of single bioactive compounds that are designed to target a specific disorder or infection. A general feature of the traditional medicine is the synergism, which provides multiple targets against specific diseases. More detailed information about the mechanisms of action of plant-derived antibacterial agents could be fined in [88].
3.2. Antibacterial Peptides Derived from Terrestrial Biota
Natural antibacterial peptides (ABPs) are low-molecular-weight peptides obtained by animals, plants and microbial, where they act as a significant part of the innate immune system and demonstrate a broad range of antimicrobial and immunomodulatory effects against bacteria. The natural ABPs are positively charged and usually contain fewer amino acids, [137]. The large diversity of natural ABPs makes challenging their classification. Nevertheless, they are categorized according to the origin as plants-, animals- and microbial-derived; according to the activity as antibacterial, antifungal, antiviral, antiparasitic, and antitumor and according to the secondary structure as β-sheet, α-helix, loops and extended peptides. The secondary structure of AMPs: α-helix, β-sheet, loop and extended is depicted in Figure 7. The majority of AMPs belongs to the β-sheet and α-helix groups, [41,137,138,139,140].
The natural ABPs have numerous advantages compared to conventional antibiotics that explains the high interest to their utilization in the therapy of bacterial, including multi-drug resistant infections, [141,142]. They are effective at low concentrations and against certain types of bacteria that are resistant to common antibiotics such as vancomycin-resistant Enterococcus and methicillin resistante Staphylococcus aureus and others. Furthermore, AMPs in combination with conventional antibiotics demonstrate a synergistic effect. They are considered as safe for use, with less or lack of side effects. As an added benefit, they possess a broad spectrum antimicrobial properties as compared to traditional antibiotics [143,144], high sensitivity [145] and multifunctional activity, i.e. they are able to target several points of interest, [146]. Many ABPs can directly kill pathogenic microbes, whereas some operate indirectly by regulating various host defensive systems. In addition to direct bactericidal action, several AMPs exhibit complex immunomodulatory activity thereby indirectly promoting pathogen purification of the host, [147]. The immunomodulatory functions of ABPs include: enhanced chemotaxis of immune cells activation and differentiation, including dendritic cell maturation and initiation of adaptive immunity; repression of cytokine-mediated and Toll-like receptor (TLR)-mediated release of pro-inflammatory cytokines and reactive oxygen species; induction of anti-inflammatory cytokines; scavenging of bacterial endotoxins, stimulating angiogenesis, enhancing wound healing, and reducing scar formation, [148]. The most ABPs exert:
i) direct biocidal effect by disrupting the membrane integrity of the target organism and/or by translocating across the microbial membrane to reach the intra cellular targets;
ii) membrane interactions, mediated by the electrostatic forces between the positively charged ABPs and negatively charged bacterial surface;
iii) destabilization of the bacterial membrane by decreasing or increasing its thickness or by causing closterization of phospholipid head groups in the membrane; iv) affecting of membrane permeabilization by formation of a complex with small organic anions carrying them across the membrane, [149,150].
The low ability to develop drug resistance and the high antibacterial activity of the ABPs are explained to some extend by possible mechanisms of their action that could be simplified summarized as including: (1) Inhibition of NRA/protein synthesis; (2) Degradation of DNA; (3) Ion-permeable pore formation in the cell membrane and; (4) Peptidoglycan synthesis inhibition, [42,150].
In contrast to the antibiotics, ABPs realize contact with the cell membranes by neutralizing the charge, and then pass through the membranes to destroy the bacteria and lessen the likelihood, those they will develop drug resistance, [143]. Since ABPs have antimicrobial mechanisms separate from conventional antibiotics, they are accepted as one of the best choice for treatment of pathogenic and drug-resistant bacterial infections, [137].
The distribution of ABPs among virtually all living organisms, complemented by exceptional structural and functional variety, stipulate a range of different antimicrobial activities including development of new antibiotic lead molecules, [151,152].
The first antimicrobial peptide (AMP) was discovered in 1957 by Robert Skarnes in blood cells, [146]. The key advantages of the ABPs: slow emergence of resistance, broad-spectrum anti-bacterial activity, high sensitivity, ability to modulate the host immune response, etc. rapidly gained attention and now ABPs are accepted as promising next-generation antibiotics that can be used to combat drug-resistant pathogens, [141,142].
From the large amount of ABPs discovered until 2020, FDA approved seven: gramicidin, daptomycin, colistin, vancomycin, oritavancin, dalbavancin, and telavancin. Analyses of the FDA approved drugs database, demonstrates that all FDA-approved ABPs were found in Gram-positive soil bacteria, and 98% of the known ABPs come from natural sources (skin secretions of frogs and toxins from different species), [147].
Encouraging examples of several ABPs already introduced into medical practice exist, such as the well-characterized cyclic anti-infective peptides gramicidins and polymyxins. Numerous other ABPs were under evaluating in non-clinical settings and in clinical trials up to 2020. Examples of ABPs that have progressed into late-stage clinical studies include omiganan, pexiganan, and DPK060 developed for the treatment of bacterial infections; LL-37 developed for improved healing of venous leg ulcers; and PXL01 evaluated for preventing postsurgical adhesions. These ABPs are linear, cationic and with a molecular length varying from 12 (omiganan) to 37 (LL-37) amino acids, [150].
Up to year 2022, more than 1,500 natural AMPs were identified and some of them like novexatin, omiganan, pexiganan, thionins, and thioninetic are passing or passed preclinical or clinical trials, [42].
In 2023, the size of the market of natural antimicrobial peptides was evalued at USD 223.00 Mn. It is expected this market to reach USD 532.02 Mn by the year 2031 and to increases by 11.65 % during the forecast period of 2024 – 2031, [3].
Up to 2024, only small number of the thousands discovered ABPs, were approved by FDA for clinical use. Because of their extremely short half-lives, they were recommended only for intravenous administration, topical treatments and, in certain cases, oral administration in the form of a very small tablet or capsule formulation. FDA approved ABPs formulations are: Atazanavir; Boceprevir; Bulevirtide, Colistin, Dalbavancin, Daptomycin, Enfuvirtide, Glecaprevir, Indinavir, Lopinavir, Nelfinavir, Oritavancin, Oseltamivir, Peramivir, Remdesivir, Ritonavir, Saquinavir, Teicoplanin, Telaprevir, Telavancin, Tesamorelin, and Vancomycin. Some ABPs based formulations as drugs that are in clinical trials include: Murepavadin; EA 230; Ghrelin; SGX942; p2TA (AB103); PMX-30063 (Brilacidin); hLF1–11 (Fungal infections); Friulimicin B; PLG0206; IDR1; Omiganan (MBI-226); Pexiganan (MSI-78); LTX-109; OP-145; DPK-060; NP101 and NP108; Novexatin (NP213); P113 (PAC-113); SGX942; Iseganan (IB-367); Brilacidin (PMX-30063); Ctx(Ile21)-Ha, [137].
The discovery of novel antibiotics using natural ABPs remains a significant current challenge. Some drawbacks like metabolic instability and/or toxicity, difficulties in the design for some specific targets and other make their clinical application problematic. Structural and functional limitations, combined with strict environmental regulations hamper the clinical translation of antimicrobial peptides as potential therapeutic agents, [145].
3.2.1. Plant Antibacterial Peptides
Plant antimicrobial peptides (ABPs) that play an important role in their innate immunity have broad-spectrum antibacterial activity, rapid killing, and cell selectivity, [153]. The plant ABPs primary target cell membranes or intracellular components in a variety of ways, which enables them to effectively kill a wide range of microorganisms and reduce the chance of pathogens to develop resistance, [154,155].
Plant ABPs have advantages compared with current antibiotic drugs because they possess a naturally occurring defence mechanisms used by plants since antiquity in fighting pathogenic bacteria. The plant ABPs are underutilized and their products offer great promise as a novel source of drug discovery for treating human infections and other diseases to solve myriad problems confounding pathogen resistance and lack of antibiotics sensitivity, [156]. Their potential applicability in bacterial diseases treatments gathers much and much interest. The molecular-based delivery, classification, production, mode of action, and chemical synthesis get growing popular over the years, [157]. Extraction methods are used for isolation of ABPs from plant organs, directed to obtain some specific structural types of ABPs, [158]. Optimized methods for chemical extraction of ABPs from roots and leaves of extremophile plants Anthyllis sericea and Astragalus armatus (collected from the Tunisian desert) are described based on using of sulfuric acid, dichloromethane, phosphate buffer, acetic acid and sodium acetate as solvents. The most appropriate solvents for extraction of ABPs from both An. sericea and As. armatus appear to be acetic acid and sodium acetate. Respectively, the corresponding leaves and roots extracts demonstrate activity against Gram-positive and Gram-negative bacteria, [155,156,159].
The most accepted classification of plant ABPs is based on their sequence similarity and tridimensional structures. In these respects, they are classified as thionins, defensins, hevein-like peptides, knottins, stable-like peptides, lipid transfer proteins, snakins and cyclotides, [160].
Thionins were found only in some plant families of Angiosperms. The One Thousand Plant Transcriptomes Initiative (1KP project) sequences the transcriptomes of more than 1000 plant species and use the data to search for new thionin sequences. Many hits were from Angiosperm plant families, which were previously not known to contain thionins. A large gene family for thionins was found in Papaver. A thionin encoded by a genomic clone was found that has antimicrobial activity in vitro. Previously, thionins were grouped into four classes. New data give reason to revise this classification. Now are recognized only class 1 thionins with eight cysteine residues and class 2 thionins with six cysteine residues, [161].
Cruciferins, napins, oil-body proteins and oleosins, are seed storage proteins (SSPs) in mustard and rapeseed (Brassica napus L., B. juncea L., B. nigra L., B. rapa L. and Sinapis alba L.), that have been used in traditional medicinal systems against different infectious diseases. Both in silico and in vitro antibacterial activity of napin and cruciferin rapseed proteins give reason to be accepted as potential candidates for development of new antibacterial agents and to be used in complementary medicine to alleviate bacterial diseases, [162]. Shotgun proteomics of Brassica rapa seed proteins identifies vicilin as a major seed storage protein in the mature seed, [163].
Several bioactive ABPs including defensins (protease inhibitor), lectins, thionin-like peptides, vicilin-like peptides, and snaking were isolated from the plants of Solanaceae family, [164,165]. Several important amino acid–derived classes were presented as plant defensive compounds, including antimicrobial peptides defensins, thionins, and knottins as potential drug leads, examining their mechanisms of action, therapeutic targets, and structure - function relationships, [166].
Solanaceae is an important family of flowering plants (Angiosperms) that comprise over 100 genera and more than 3000 species. The Solanaceae family plants are rich of different bioactive constituents including natural ABPs that have been used in different traditional medicinal systems. Several bioactive ABPs including defensins (protease inhibitor), lectins, thionin-like peptides, vicilin-like peptides, and snaking were isolated from the plants of Solanaceae family, [164,165].
The successful use of plant ABPs requires detailed knowledge of the structure-function relationship. The last one was studied in thionins, α-hairpinins, hevein-like peptides, and the unique Ib-AMP peptides isolated from Impatiens balsamina. It was shown that even subtle changes in amino acid sequences could affect the biological activity of the ABPs, which opens possibilities to creation of molecules with better therapeutic efficacy and cheaper large-scale production, [167].
There are studies reporting some toxicity of plant ABPs to non-target cells or limitations of oral administration. However, it is accepted that ABPs with reduced toxicity or allergenicity or greater resistance to peptidases could be designed by chemical modification strategies and different bioinformatics tools, [160].
It is thought that the discovery and design of ABPs with desired properties (reduced toxicity or allergenicity or greater resistance to peptidases, etc.) could be done by data base analyses, chemical modification strategies and different predictive bioinformatics tools, [155,156].
Multiple classes of antimicrobial peptides in Amaranthus tricolor (red amaranth or Chinese spinach) were revealed by in silico prediction, proteomics, and mass spectrometric characterization. Bottom-up proteomics identify seven novel peptides from three ABP classes including lipid transfer proteins, snakins, and a defensin. Bioactivity screening of isolated Atr-LTP1 shows activity against the high-risk ESKAPE bacterial pathogens: Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, and Enterobacter cloacae. The results highlight the potential for integrating ABP prediction algorithms with complementary OMICS approaches to accelerate characterization of biologically relevant ABP forms, [168].
A user-friendly web server, available at the URL: http://www.nipgr.ac.in/PTPAMP/ was developed, termed PTPAMP that could be used as a prediction tool for plant-derived antimicrobial peptides, by integration of selected compositional models. So far developed models were based on multiple peptide features, like amino acid - and dipeptide composition as well as physicochemical attributes for predicting plant-derived ABPs. The selected compositional models were integrated now into the web server PTPAMP that is capable to classify a query peptide sequence into four functional activities: antimicrobial, antibacterial, antifungal, and antiviral. The analysis indicates the abundance of cysteine residues in plant-derived ABPs and the distribution of other residues like G, S, K, and R, which differ as per the peptide structural family, [169].
Recently, recombinant ABPs were produced in plants at large scale and low cost. Because the ABPs are less likely to elicit resistance of pathogenic bacteria than conventional antibiotics, it seems that they open new avenues for agricultural and medical applications, [170].
3.2.2. Animal Origin Antibacterial Peptides
Natural antibacterial peptides (ABPs) present in every animal as a part of its innate immune system and ABPs are derived from variety of animals including mammalians, amphibians, snake venom, crabs, insects and others.
3.2.2.1. Antibacterial Peptides Derived from Mammalians
ABP were isolated from different mammalian sources such as granules of neutrophils, Peneth cells, mucosal secretions from epithelial cells and protein degradation products. Three classes of antibacterial peptides, found in abundance in neutrophils defensins, cathelicidins and histatins were studied extensively, [171].
Cathelicidins are a class of natural, short cationic, ABPs, identified in different animal species: birds, fish, reptiles, amphibians, snake venoms and mammalians (cow, pig, rabbit, sheep, mouse, monkey, horse and human), primarily produced in epithelial cells, neutrophils and macrophages. About 30 cathelicidin family members were identified in mammalian species. The cathelicidin family of ABPs with varying antibacterial activities and safety is considered now as a promising alternative to conventional antibiotics, [172].
3.2.2.2. Antibacterial Peptides Derived from Amphibians
Amphibians are important sources of ABPs. Duttaphrynus melanostictus is the main source of traditional Chinese medicine “Chansu”, which has anti-infection effect without a clear mechanism. A study aimed to find a cathelicidin peptide in D. melanostictus, investigates its activity in vivo and in vitro, and an AMP-encoding gene (cathelicidin-DM, GenBank: KJ820824.1) was obtained from the constructed cDNA library of D. melanostictus. A skin wound infection model and in vivo imaging were used for evaluation of possible applications. The results show that cathelicidin-DM is a 37 amino acid AMP with good bactericidal ability, similar to that of melittin. Both can kill bacteria within 15 min. Thus, cathelicidin-DM could be a new template for antimicrobial drug development based on its good antibacterial activity in vivo and in vitro, [173].
Another antimicrobial peptide isolated from frog skin hemocytes is poly(glycolide-co-lactide) (PGLA), and it was shown to prevent bacterial adhesion by causing a conformational change and elimination of bacterial pili. One more antimicrobial peptide, magainin was isolated from the African clawed frog (Xenopus laevis). However, studies show that resistant Escherichia coli strains can easily develop resistance, [23].
3.2.2.3. Antibacterial Peptides Derived from Snake Venom
Cathelicidins are a class of natural, short cationic, AMPs, identified in different animal species: mammalians, birds, fish, reptiles, amphibians and snake venoms. About 30 cathelicidin family members were identified in mammalian species. The data on identified in snake venoms cathelicidins, including their chemistry, characterization, pharmacological action, antimicrobial and antibioflim effects, mechanism of action and a potential for the development of novel antibiotics to combat antibiotic-resistant bacteria were summarized by Barros et al., [172]. Cathelicidins have been at first studied mostly for their direct antimicrobial killing capacity, but nowadays they are more and more appreciated for their immunomodulatory functions, [174]. According to Yang et al. [105], a non-bactericidal cathelicidin provides prophylactic efficacy against bacterial infection by driving phagocyte influx eLife.
Wang et al. [107] reports a new identified cathelicidin peptide, named Hydrostatin-AMP2 from the snake Hydrophis cyanocinctus with excellent antimicrobial activity against both Gram-positive and Gram-negative bacteria, including standard and clinical Ampicillin-resistant strains. The new identified Hydrostatin-AMP2 has faster antimicrobial action than Ampicillin as demonstrated by a kinetic assay. Hydrostatin-AMP2 apparently decreases the production of pro-inflammatory cytokines in the LPS-induced RAW264.7 cell model.
Klubthawee et al. [175] report rationally designed, hybrid antimicrobial peptide, that is inspired by cathelicidin and aurein and exhibits membrane-active mechanisms against Pseudomonas aeruginosa.
3.2.2.4. Antibacterial Peptides Derived from Crabs
Jiang et al. [176] report a novel antibacterial peptide Spampcin56–86 from Scylla paramamosain crab exerting rapid bactericidal and anti-biofilm activity in vitro and anti-infection in vivo. Many diseases in the modern world originate from inflammatory and oxidative stress. Beneficial metabolites with promising antioxidant and anti-inflammatory potentials were reported lately, that were identified in the methanol extracts of the shell of crabs of the genus Charybdis, [177].
3.2.2.5. Antibacterial Peptides Derived from Insects and Others
Lee and Shin [178] identify novel ABPs from venom gland of spider Pardosa astrigera by deep multi-task learning.
Different types of antimicrobial agents, obtained from animals/animal products are known to demonstrate antimicrobial potency against various pathogenic microbes. Such are lactoferrin (Lf), (an ironbinding milk protein), that is active against a spectrum of microbial organisms like E.coli, Carnobacterium, Klebsiella, L. monocytogenes, etc; chitosan, a polycationic biopolymer found naturally in the crustacean and arthropod exoskeletons that is active against various Gram-positive and Gram –negative bacteria; the bacteriolitic enzyme, lysozyme that is found naturally in mammalian milk and hens eggs displaying an excellent antimicrobial action in the case of Listeria innocua and Saccharomyces cerevisiae; certain milk-derived bioactive substances, like casein, that are reported to have antibacterial activities and others, [42].
Arenicins are a group of 3 type of peptides: arenicin-1, arenicin-2 and arenicin-3 that were shown to have good antimicrobial activity against Gram-negative bacteria. Arenicin-1 isolated from Arenicola marina (sandworm) shows potent antimicrobial activity against Escherichia coli and Pseudomonas aeruginosa, with MIC values between 1 and 2 µM. A slightly modified arenicin-3 peptide has considerable antimicrobial activity even against XDR (extensive drug resistance) and MDR (multi-drug resistance) strains such as Pseudomonas aeruginosa, Acinetobacter baumannii, Escherichia coli and Klebsiella pneumonia, [179].
3.2.3. Antibacterial Agents Produced by Microbes
Microbial secondary metabolites are a great source of bioactive molecules. The discovery of penicillin (efficient against Gram-positive bacteria), derived from Penicillium notatum by Alexander Fleming, in the year 1928 makes a fundamental change in the sources for derivation of natural biologically active products from plants to microbes, [42,180,181]. Natural products with diverse structure and biological activity and recombinant proteins, produced by microbes, are already valuable molecules for the medicine. The development of robust and promising microorganisms as cell factories, engineering approaches directed to improve yields of microbial production and generating novel molecules are largely studied as inspiring the development of new therapeutic agents, [182].
Converting of carbon and nitrogen sources into a large diversity of intracellular and extracellular biopolymers, such as polysaccharides, polyamides, polyesters, polyphosphates, extracellular DNA and proteinaceous components by bacteria is also of research interest as a new target for antibacterial drugs. Bacterial polymers have important roles in pathogenicity, and their varied chemical and material properties make them suitable for medical and industrial applications. When produced by pathogenic bacteria they function as major virulence factors, whereas when are produced by non-pathogenic bacteria, they become food ingredients or biomaterials, [183].
Produced by microbes both, bacteriophages and bactericines are two types’ antibacterial agents of especial interest in the combating the antimicrobial resistance (AMR).
3.2.3.1. Bacteriophages
Bacteriophages (phages) are viruses, able to infect bacterial cells and force them to produce viral components, using a lytic or lysogenic cycle. Bacteriophages are potential alternative of chemical antimicrobial agents used against pathogens that are of public health significance. Understanding the phage diversity and host specificity is important for the development of effective phage biocontrol approaches, [184].
Thousands bacteriophages of different types could be find everywhere. Bacteriophages were discovered over a century ago, but their use for treatment of antibiotic-resistant infections regained popularity only recently. In contrast to many antibiotics, which damage harmful bacteria, and simultaneously disturb the all microbiota thus triggering a new set of problems, each phage more narrowly targets bacterial strains or species. This specificity makes phage therapy an attractive approach for treating of antibiotic-resistant bacterial infections, including such caused by multi-drug resistant bacteria. The bacteriophages provide other advantages over antibiotics, like less significant side effects, less time-consuming and less costly development process, [185,186]. Because it’s many advantages, phage therapy survives currently its renaissance after long years of different doubts. Recently, the U.S. National Institutes of Health (NIH) awarded $2.5 million to 12 institutes around the world to study phage therapy, [187,188].
Phages are the only drug that reproduces itself at the site of infection and disintegrates again after lysis of all suitable bacteria. They contain DNA or RNA in their genome that is encapsulated in a protein coat. Many phage proteins, including endolysins (lysins), virion-associated peptidoglycan hydrolases (VAPGHs), depolymerases, and holins display antibacterial activity. Since several phage types may be suitable for one bacterial pathogen from a set of available phage types, the therapeutic use of mixtures of different phage types is optimal. The specificity of phages is their major advantage over antibiotics. It is also the main reason for the individual character of phage application as a tailor-made therapy in individual cases. Main families and characteristics of bacteriophages are in debt presented by Bin Hafeez et al., [139].
Phages against Staphylococcal Infections
In terms of potential medical applications, phages belonging to the Kayvirus genus of the Herelleviridae family are regarded as the most interesting ones. Kayviruses have already demonstrated their efficacy in the treatment of various Staphylococcus aureus infections, both in animal models and human clinical cases, [189,190]. A phage belonging to the kayvirus lytic module was isolated that encodes an additional endolysin, [191]. The polyvalent, Kayvirus genus phages, infecting mostly S. aureus and some CoNS and displaying a broad spectrum biological activity are one of the best agents controlling staphylococcal infections, [192,193,194]. Some of the kayvirus phages have already been used in commercial phage-based preparations to treat DFI, [195]. Studies on phage therapy for staphylococcal infections are focused mainly on S. aureus and only a few on the isolation and characterization of phages infecting the clinical isolates of CoNS, especially Staphylococcus epidermidis, [192,195].
Fanaei Pirlar et al. [196] describe one of the novel bacteriophages specific against S. epidermidis and with antibiofilm activity. Important for the success of phage therapy, in vitro techniques and measurements of phage characteristics are presented, [197]. Comparative assessment of the bacteriophage and antibiotic activity against multidrug-resistant S. aureus biofilms shows that while the antibiotics cannot diffuse through the polymeric matrix of a biofilm, the Kayviruses can effectively penetrate and disrupt S. aureus and S. epidermidis biofilm structures, [198,199].
Alsaadi et al. [200] report the isolation and genome sequencing of 40 bacteriophages from human skin SWABS that infect coagulase-negative Staphylococcus (CoNS) species, which extends the knowledge of phage diversity. Six genetic clusters of phages were identified with two clusters representing novel phage species, one of which was characterized and named Alsa phages. The identified Alsa phages have a greater ability to infect the species S. hominis that were otherwise less infected than other CoNS species by the isolated phages. This indicates an undescribed barrier to phage infection that could be due to numerous restriction-modification systems. The extended diversity of Staphylococcus phages here enables further research to define their contribution to skin microbiome research and the mechanisms that limit phage infection.
Staphylococcus sp. is the most common bacterial genus in infections related to diabetic foot ulcers (DFUs). Plumet et al. [201] isolate six phages (SAVM01 to SAVM06) from effluents of diabetic foot ulcers (DFUs), belonging to the Herelleviridae family, with sequences similar to those of the Kayvirus genus. No lysogeny-associated genes, known virulence or drug resistance genes were identified in the phage genomes. The phages display a strong lytic and antibiofilm activity against DFU clinical isolates, as well as against opportunistic pathogenic coagulase-negative staphylococci. The experimental results suggest that these phages could be effective biocontroling agents against staphylococcal clinical isolates from DFUs.
Phages against Pseudomonas Infections
Bacteria surviving in extreme conditions and the bacteriophages that infect them are sources of heat-stable proteins that are utilized in biotechnological applications but not as antimicrobial agents. Plotka et al. [202] demonstrate that the Ts2631 endolysin from the extremophilic bacteriophage vB_Tsc2631, which infects Thermus scotoductus, is very active against the multidrug-resistant clinical strains of Acinetobacter baumannii, Pseudomonas aeruginosa and pathogens from the Enterobacteriaceae family, i.e. Ts2631 endolysin could be an effective antimicrobial agent against Gram-negative multidrug-resistant bacteria. Transmission electron microscopy (TEM) and fluorescence microscopy observations of A. baumannii cells, treated with Ts2631 endolysin variants, demonstrate that the intrinsic antibacterial activity of Ts2631 endolysin depends on the presence of its N-terminal tail, [202].
A novel phage, pPa_SNUABM_DT01, infecting Pseudomonas aeruginosa canine otitis externa isolates was characterized by its morphology, growth, lysis kinetics, and genomic characteristics. Comparative genome analysis demonstrates that the phage is a novel species in Myoviridae. The nucleotide similarity was moderately high compared to the Pseudomonas virus, Noxifer. However, a phylogenetic analysis and a dot plot indicate that pPa_SNUABM_DT01 is not closely related to the Phikzvirus or Noxifervirus genus, instead, it belongs to a novel one, [203].
Proteus mirabilis and P. aeruginosa are two bacterial species commonly associated with urinary tract infections in humans. Novel bacteriophages and their derived proteins were developed for the biocontrol of Proteus and Pseudomonas biofilms. The identification and utilisation of both, the whole phage and its tail-spike protein with the pectate lyase activity are described to treat P. mirabilis biofilms. The phage and the tail-spike protein were assessed by different in vivo and in vitro assays that demonstrate their antibacterial and antivirulence properties against P. mirabilis biofilms. The combinational treatment of P. aeruginosa biofilms by phage and cold atmospheric plasma establishes that the use of cold atmospheric plasma followed by exposure to P. aeruginosa phages is the most effective for eradication of P. aeruginosa biofilms, [204].
Two bacteriophage genera (targeting B. mycoides and Pseudomonas species) discovered in a groundwater reservoir highlight subsurface environments as underexplored biotopes in bacteriophage ecology, [205].
Phages against Escherichia coli Infections
Sattar et al. [206] report isolation, preliminary characterization, and genome analysis of two novel lytic phage species (Escherichia phage SKA49 and Escherichia phage SKA64) having lysis potential against MDR strain of avian pathogenic E. coli, QZJM25. Both phages, SKA49 and SKA64, are able to keep QZJM25 growth significantly less than the untreated bacterial control for approximately 18 h, being stable at 37 °C only. In contrast to SKA64, SKA49 demonstrates a broader host range against Escherichia coli strains. Genome analysis indicates their safety because no recombination, integration and host virulence genes were identified.
Nicolas et al. [207] isolate and characterize a novel phage collection against avian-pathogenic E. coli (APEC). The collection includes nineteen genetically diverse, lytic E. coli phages, eighth of which were tested in combinations for their efficacy in controlling of avian patogenic E. coli infections. Genome homology analysis reveals that the phages belong to nine different genera, one of them being a novel genus (Nouzillyvirus). The broad host range of some phages is partially explained by the presence of receptor-binding protein carrying a polysaccharidase domain. To demonstrate their therapeutic potential, a phage cocktail consisting of eight phages belonging to eight different genera, was tested against BEN4358, an APEC O2 strain. This phage cocktail fully inhibits in vitro, the growth of BEN4358.
Phages for Global Health (PGH) is training scientists in the region of East Asia to isolate relevant therapeutic phages for pathogenic bacteria within their locality, and thus contributing to making phage technology universally available. During the inaugural PGH workshop in East Africa, samples from Ugandan municipal sewage facilities were collected and two novel E. coli lytic phages were isolated and characterized.
The phages, UP19 and UP30 lysed ∼82% and ∼36% of the 11 clinical isolates examined, respectively. The genomes of UP19 (171.402 kb, 282 CDS) and UP30 (49.834 kb, 75 CDS) closely match the genera Dhakavirus and Tunavirus, respectively. The isolated phages have therapeutic potential for further development of treatments against E. coli infections, [208].
Phages against Salmonella Infections
Plasmid-dependent phages infect bacteria carrying conjugative plasmids by recognizing the plasmid-encoded pilus. Two lytic phages from wastewater were isolated using a virulent strain of Salmonella enterica carrying the conjugative IncN plasmid pKM101. Both phages, named Lu221 and Hi226, are novel dsDNA viruses within the class Caudoviricetes with genomes of approximately 76 kb. They show broad host range infecting E. coli, S. enterica, Kluyvera sp., and Enterobacter sp. carrying conjugative plasmids. They recognize plasmid-encoded receptors from 12 out of 15 tested plasmids, all of them carrying resistance determinants, [209].
Genomic and phenotypic analysis of S. enterica phages identifies two novel phage species. The host range, morphology, and genetic diversity of eight S. enterica phages isolated from a wastewater treatment plant were assessed. The host range analysis revealed that six out of eight phages lysed more than 81% of the 43 S. enterica isolates tested. Whole-genome sequencing (WGS) data revealed that phage genome sizes ranged from 41 to 114 kb, with GC contents between 39.9 and 50.0%. Two of the phages SB13 and SB28 represent new species, Epseptimavirus SB13 and Macdonaldcampvirus, respectively, as designated by the International Committee for the Taxonomy of Viruses (ICTV) using genome-based taxonomic classification. One phage (SB18) belonges to the Myoviridae morphotype while the remaining phages belonge to the Siphoviridae morphotype. None of the phages possesses virulence, toxin, antibiotic resistance, type I–VI toxin–antitoxin modules, or lysogeny genes (by gene content analyses), [210].
Others
Nakonieczna et al. [211] discover three novel bacteriophages, J5a, F16Ba, and z1a, specific for Bacillus anthracis (able to be highly lethal) by screening environmental samples from various regions in Poland and present their basic characteristic. The new phages and their closest relative phages Tavor_SA, Negev_SA, and Carmel_SA, form a separate clade of the Wbetavirus genus, were designated as J5a clade. The comparative genomic analysis indicates that the new bacteriophages encode two receptor-binding proteins, of which one may bind a sugar moiety of B. anthracis cell surface.
The global warming favored becoming of a range of bacteria, such as Aeromonas hydrophila, pathogenic to humans. They are not easy for treatment by traditional methods due to their capacity to form biofilms. Bacteriophage offer a possible alternative approach for controlling the growth of the biofilms.
Kabwe et al. [212] first report the isolation and characterization of bacteriophages which carry intrinsic antibiotic resistance genes and are capable to disrupt biofilms caused by clinical isolates of A. hydrophila.
Kallies et al. [213] identify huge phages (with genomes larger than 200 kilobases) from wastewater metagenomes by screening of 165 wastewater metagenomes for the presence of viral sequences. The dataset of over 600 identified potential huge phage genomes, was reduced using manual curation by excluding these did not containing viral protein-coding genes or consisting of concatemers of several small phage genomes. A phylogenomic analysis of the huge phages and phages with smaller genomes (less than 200 kb) support the hypothesis that huge phages have undergone convergent evolution. The genomes containes typical phage protein-coding genes, sequential gene cassettes for metabolic pathways, and complete inventories of tRNA genes covering all standard and rare amino acids.
3.2.3.2. Bacteriocines
Bacteria produce a range of antimicrobial peptides, the most diverse of which are bacteriocins. Bacteriocines are small antimicrobial peptides (peptide toxins), synthesized by ribosomes of both Gram-positive and Gram-negative bacteria and usually display activity against bacteria (pathogenic and multi-drug resistant), phylogenetically related to the producing strain. The antimicrobial activity spectrum depends on the peptide that can target several bacteria, [214,215].
The selectivity and safety profile of the bacteriocins are their superior advantages over traditional antibiotics; however, the bacteriocins are susceptible to degradation by proteolytic enzymes and therefore have low in vivo stability. In addition, their large-scale production is problematic. It is expected, that such limitations will be avoided by extensive research, including development novel drug delivery systems, [216].
Yount et al. describe discovery of Type II bacteriocins, using a new high dimensional bioinformatic algorithm. In this way, all bacteriocin families of Type II were detected whereupon identified putative bacteriocins with broad-spectrum antimicrobial activity against a range of human pathogens. The putative bacteriocin sequences are from different microorganisms: Bacillus thuringiensis, Eubacterium rectale, B. cereus and Enterococcus pallens, [217].
Bacteriocines from Lactic Acid Bacteria
Lactic acid bacteria (LAB) are one of the most used bacteria to produce bacteriocins that could serve as alternatives of conventional antibiotics. Enterococcus faecalis, Lactobacillus fermentum, L. plantarum, L. helveticus, L. pentosus, L. paracasei subsp. paracasei, L. rhamnosus I, and L. delbrueckii subsp. lactis are strong strains in bacteriocins production. To date, Nisin, Pediocin PA-1, and Micocin are the only FDA-approved bacteriocins to use as food preservatives, [218].
LAB-bacteriocins could be used alone, or as potentiating agents to treat bacterial infections with aim to reduce the use of traditional antibiotics and to develop novel therapeutic options. Most LAB-bacteriocins act by disturbing the cytoplasmic membrane through forming pores, or by cell wall degradation. Some of the bacteriocines that are active against Gram-negative bacteria, still have unknown mode of action. The most bacteriocins-producing strains have immunity mechanism, involving an immunity protein and a dedicated transport system. The immunity mechanisms usually vary from one bacteriocin to another, [184].
Lei et al. [219] present partial purification and characterization of a broad-spectrum bacteriocin, produced by isolated from infant’s feces Lactobacillus plantarum ZRX03. The fermentation supernatant, produced by this strain inhibits E. coli, Staphylococcus aureus, and Listeria monocytogenes (inhibition zone of 12.83 ± 0.62 mm, 15.08 ± 0.31 mm, 6.75 ± 0.20 mm, respectively,) stronger than the lactic acid bacteria N1, N2, M13, M21, M31, and M37. Ethyl acetate was selected as the optimal crude extract solution. A broad-spectrum antimicrobial activity was shown for the obtained bacteriocin, inhibiting Gram-positive bacteria, Gram-negative bacteria and yeast, including S. aureus, Bacillus subtilis, Bacillus anthracis, E. coli, and Salmonella, [219]. Circular bacteriocin, plantacyclin B21AG from Lactiplantibacillus Plantarum B21 (isolated from nem chua, Vietnamese sausage) was discovered with broad spectrum antimicrobial activity against Gram-positive bacteria, high thermostability and proteolytic resistance, [220]. Bacteriocin-like peptide (SLG10), made by Lactobacilus plantarum strain, with antimicrobial activity against both Gram-positive and Gram-negative bacteria, was isolated from kombucha (a fermented bubble tea). Stability studies show that the peptide retains its antimicrobial properties for 14 days at 37° C and for 2 months at 4°C being stable at pH values between 2.0 and 7.0, [221].
A novel bacteriocin LSX01 of Lactobacillus paracasei LS-6 was isolated from a traditional fermented yogurt (produced in Yunnan, China), purified and characterized. The LSX01 exhibits an extensive antimicrobial spectrum against both Gram-positive and Gram-negative bacteria as well as a tolerance to heat, acid-base treatments, and a sensitivity to proteolytic enzymes. The treatment of S. aureus planktonic cells with LSX01 significantly reduces their metabolic activity and disrupts the cell membrane integrity. A biofilm formation of S. aureus is also significantly inhibited, [222].
Thuy et al. [223] characterize the broad-spectrum antibacterial activity of bacteriocin-like inhibitory substance-producing probiotics, isolated from fermented foods. Selected lactic acid bacteria (LAB) with probiotic potential were evaluated by various tests, including exopolysaccharide production, antibiotic susceptibility, acid and bile tolerance, antibacterial activity, and cell adhesion and cytotoxicity to gastric cell lines. Six selected LAB strains demonstrate high viability under gastrointestinal conditions, produce high exopolysaccharides, show no or less cytotoxicity, and adhere successfully to gastric cells. Three strains, Weissella confusa , Lactiplantibacillus plantarum , and Limosilactobacillus fermentum, demonstrate a strong antibacterial effect against drug-resistant Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, Salmonella enterica serovar Choleraesuis, Enterococcus faecium, and Staphylococcus aureus. The whole genome sequencing of these three strains (using the Nanopore platform) shows that they do not harbor genes related to toxins, super antigens, and acquired antimicrobial resistance. The extract of CYLB30 and CYLB47 bacteriocin-like substances (BLIS) inhibit the growth and biofilm formation of drug-resistant P. aeruginosa and methicillin-resistant S. aureus, causing membrane disruption and inhibiting adhesion ability to human skin HaCaT cells.
Lacticaseibacillus paracasei-derived antibacterial peptide NGJ1D was found in the fermentation broth of L. paracasei. The antibacterial peptide NGJ1D has minimum inhibitory concentration (MIC) of 62.5 μg/mL against Staphylococcus aureus and could kill the bacteria within 3 h, [224].
Enterococcus faecalis (CAUM157), a Gram-positive bacteria isolated from raw cow’s milk, was studied for its bacteriocin production. In vitro and in silico characterization of N-formulated, di-peptide bacteriocin from E. faecalis was lately presented with anti-Listeria activity. The antimicrobial activity of CAUM157 was attributed to a two-peptide class IIb bacteriocins with potent activity against food-borne pathogen Listeria monocytogenes and periodontal disease-causing pathogens (Prevotella intermedia KCTC 15693 T and Fusobacterium nucleatum KCTC 2488 T). Although E. faecalis CAUM157 innately has genes for virulence factors and antibiotical resistance (e.g., tetracycline and erythromycin), its bacteriocin production is valuable for the needs of in live microorganisms and pathogens control, [225]. Cui et al. [226] perform purification and characterization of novel bacteriocins, produced by E. faecalis CG9 from human saliva that inhibit the growth of Gram-negative bacteria. Study of the one of the isolated from E. faecium bacteriocins, Plantacyclin B21AG, shows its excellent stability and bactericidal activity against sporulating bacteria such as Clostridium perfringens and non-sporulating Listeria monocytogenes. Sharma et al. [227] found that the produced by vaginal E. faecium enterocin 12 A inhibits multidrug resistant Gram-negative bacteria such as Salmonella enterica, Shigella flexneri, E. coli, and Vibrio cholerae, as well as the proliferation of cancer cells.
Bacteriocines from Staphylococci
Newstead et al. [228] isolate several bacteriocins from commensal coagulase-negative Staphylococci, many of which display in vitro and in vivo inhibiting activity against S. aureus. The ability of these bacteriocins to destroy a biofilm formation, their novel mechanisms of action and efficiency against antibiotic-resistant bacteria make them novel antibacterial candidate therapeutic. Ovchinnikov et al. [229] report successful development of bacteriocins into therapeutic formulation for treatment of methicillin-resistant Staphylococcus aureus (MRSA) skin infection in a murine model. The potential of two broad-spectrum bacteriocins, garvicin KS and micrococcin P1, were explored in this study for skin infection treatments. The two bacteriocins act synergistically with each other and with penicillin G in killing MRSA in vitro. To assess its therapeutic potential, the three-component formulation was involved in a murine skin infection model with a multidrug-resistant luciferase-tagged MRSA Xen31, a strain derived from the clinical isolate S. aureus ATCC 33591. The efficiency of the three-component formulation, in eradicating the pathogen from treated wounds was demonstrated by using of the tagged-luciferase activity as a reporter for the presence of Xen31 in wounds. As compared to Fucidin cream, which is an antibiotic commonly used in skin infection treatments, this formulation appears to be superior in terms of preventing the development of resistance, [229].
Bacteriocines from Streptomyces
Exploring antibacterial properties of bioactive compounds, isolated from Streptomyces sp. in bamboo rhizosphere soil (collected within the Megamalai forest of the Western Ghats in the Theni zone of Tamil Nadu, India) was presented lately. Bioactive compounds were extracted from the culture medium using ethyl acetate. Their antibacterial and antioxidant activities were evaluated through disc diffusion and DPPH radical scavenging methods, respectively. Ethyl acetate extracts were analysed by FT-IR and GC–MS techniques. Among total nine strains of Actinobacteria, the strain BS-16 identified as Streptomyces sp., displays remarkable antibacterial activity against three strains: S. aureus, B. subtilis, and Streptococcus pyogenes with inhibition zone of 19 mm, 12 mm and 10 mm respectively, [230].
3.2.4. Approaches to Address Shortcomings of Natural Antibacterial Peptides
Shortcomings, such as low stability, high toxicity in some cases, lability to proteolytic degradation, poor pharmacokinetics, high production costs, as well as difficulties in the specific targets design of successful in vitro natural antibacterial peptides (ABPs), continue to hamper their clinical applicability, [145]. Existing experimental and computational tools attempt to address some shortcomings of the ABPs and to make ease the preclinical and clinical development of novel therapeutics based on antimicrobial peptides, [151]. Use of nanodelivery systems, as well as modification or synthesis of new AMPs inspired by nature are some of them.
Mwangi et al. [231] discuskey strategies used to optimize the performance of ABPs, including rational design and de novo synthesis as well as predictive computational tools and utilizing artificial intelligence in the design and synthesis of highly efficient lead drug candidates. Research on discovery and optimization of lead molecules, originating from natural sources [232] and nanotechnology-based delivery are some of them, [145]. Luong et al. [233] discuss approaches to improve biological activity of ABPs either by modifying chemical structure or incorporating into delivery systems.
3.2.4.1. Nanodelivery systems
Nano delivery is known to enhance the antibacterial efficacy of classical antibiotics and ABPs, while reducing their toxicity that is due to its’ capability to improve the bioavailability and permeation across barriers of classical antibiotics and ABPs, and/or to protect against degradation. The biological activity of nanodelivery systems could be timesaving and cost-effective predicted by machine learning, [234,235].
The cyclic peptide antibiotic, polymyxin B (produced by Bacillus polymyxa bacterium) is an example of improved biological activity by nanodelivery. Polymixin B cross-linked with sodium alginate in polyion complex nanoparticles or loaded in niosomes show enhanced bioavailability and increased potential antimicrobial activity against Pseudomonas aeruginosa, [236].
Stan et al. [23] review nano-carriers used for transportation of natural antibacterial compounds to improve their stability, bioavailability, cellular uptake/internalization, pharmacokinetic profile and to reduce the toxicity of these natural compounds. There are a number of nanocarriers such as liposomes, micro-emulsion systems, nanocapsules, solid lipid nanoparticles, polymeric micelles, dendrimers, etc. However, some of the recent studies are focused on the incorporation of natural antimicrobial substances into polymeric nanoparticles, niosomes and silver nanoparticles, having intrinsic antimicrobial activity, are one of them. Many recent in silico or in vitro works show that the internalization of optimal nanocarriers represents the future of the “green therapeutics”.
The ABPs are susceptible to proteolytic degradation in body fluids and therefore the tablet, capsule, and solution are inconvenient when given orally due to peptide degradation and lower absorption in the gastrointestinal tract. Special drug delivery systems and formulations like nanocarriers, hydrogels, cubosomes, wafers, dry powder inhalers, creams, and mouth rinses were established to prevent some major barriers to clinical applications, such as cytotoxicity, bioavailability, and stability. Several specialized drug delivery systems of AMPs were already approved by FDA and some others are under clinical trials, [137].
3.2.4.2. Modified and Synthetic Analogues of Antibacterial Peptides Inspired by Nature
It was experimentally found that natural peptides with special chemical structure demonstrate high antibacterial activity and some modifications of the peptide backbone and structure can additionally improve their biological activity and stability as well as to reduce the toxicity of these ABPs. Inspired by nature development of modified and synthetic analogues of natural ABPs is an extensive current approach that is based on different chemical strategies assisted by in silico research. Gan et al. [237] present various chemical synthetic strategies to develop ABPs with improved properties, including chemical modifications of existing peptides, semi-synthesis, and computer-aided design. Novel ABP structures, including hybrid, dendrimer and polypeptide, peptidomimetic, and ABP–drug conjugates are also highlighted. Zhou et al. [238] demonstrate that poly(2-oxazoline) (POX) can work as a functional mimic of peptides. POX-based glycine pseudo-peptides, a host defense peptide mimic, have potent activities against methicillin-resistant S. aureus, which causes formidable infections. S. aureus does not develop resistance to POX owning to the reactive oxygen species related antimicrobial mechanism.
It is thought that the understanding the AMPs structure–function relationship have potential to help the conceptualization of the development of new ABPs by evaluation the role of each residue and essential amino acids for their biological activity. This feature could help the development of a second-generation AMPs with high potential antimicrobial and other activity, [239]. Li et al. [6] summarize recent synthetic efforts, their impact on analogue design and various applications in the development of ABPs for next generation antimicrobial agents. Focusing on antimicrobial peptidomimetics, they present modifications to enhance antimicrobial activity, such as lipidation, glycosylation and multimerization through a broad application of novel bio-orthogonal chemistry.
Ajish et al. [240] design novel short antimicrobial peptide BP100-W with antimicrobial, anti-biofilm and anti-inflammatory activities by replacement with tryptophan. Wiman et al. [241] develop novel broad-spectrum antimicrobial lipopeptides derived from plantaricin NC8 β. Chaudhary et al. [242] report efficient production of bioactive amidated ABPs by transient expression of glycine-extended ABPs in Nicotiana benthamiana line expressing the mammalian enzyme peptidylglycine α-amidating mono-oxygenase (PAM). Cationic ABPs accumulate to substantial levels in PAM transgenic plants compare to non-transgenic N. benthamiana. Moreover, ABPs purified from plants exhibit robust killing activity against six highly virulent and antibiotic resistant ESKAPE pathogens, prevent their biofilm formation, analogous to their synthetic counterparts and synergize with antibiotics. The experimental data and techno-economic analysis demonstrate the potential use of plant chassis for large-scale production of clinical-grade ABPs [243]. The study of antimicrobial cyclic peptides become a hot topic. Among them, macrocyclic AMPs receive extensive attention. The structures and functions of the dominant cyclic natural and synthetic AMPs are presented with outlook on the future direction of cyclic AMPs, [144].
Many databases were created to collect both fundamental and pharmacological information summarizing sources, structures, modes of action, and classifications of ABPs, valuable computational tools for prediction the antimicrobial activity and new machine learning approaches that can be employed to improve ABP activity. Multi-label classifiers with binary relevance and algorithm adaptation techniques are used to predict different functions of ABPs across a wide range of pathogen bacteria. Forward feature selection identifies sequences order and charge as critical, with specific amino acids as discriminative. These findings provide valuable insights for the design of ABPs with multiple functionalities, thus contributing to the broader effort to combat drug-resistant pathogens [139]. Peptide databases and different computational tools are available as resources to collect ABPs and beneficial tools for prediction and design of computational models for new biologically active AMPs, [243].
The computer-aided design puts together crucial information on chemical parameters and bioactivities in AMP sequences, thus providing modes of prediction to evaluate the antibacterial potential of a candidate sequence before synthesis. Quantitative structure-activity relationship (QSAR) computational models, for instance, greatly contribute to AMP sequence optimization aimed at improved biological activities. In addition to machine-learning methods, the de novo design, linguistic model, pattern insertion methods, and genetic algorithms, show a potential to boost the automated design of AMPs. As expected from automated design strategies, diverse candidate sequences with different structural arrangements were generated and deposited in the databases, [244].
Boone et al. [245] present combining genetic algorithm with machine learning strategies for designing potent antimicrobial peptides with an example of supervised machine learning and a genetic algorithm to find a peptide active against S. epidermidis (a common bacterial strain for implant infections) with an improved aggregation propensity average for an improved ease of synthesis. This is the first time when codon-based genetic algorithms combined with rough set theory methods is used for computational search on peptide sequences.
C. Rumancev et al. [246] demonstrate that emerging BioSAXS method could help to elucidate the mode of action of ABPs that can contribute in the development of antibiotics against resistant bacteria. The effect of two highly active short, broad-spectrum ABPs (14D and 69D) was investigated against E. coli and methicillin-resistant S. aureus (MRSA) as well as further studied the ultrastructural changes in E. coli and MRSA in response to these AMPs using the BioSAXS technique.
Jianxun et al. prepare α-Helical antimicrobial peptide YHX-1 by de novo design and estimate its antibacterial activity. To obtain a novel antimicrobial peptide, the sequence length, charge numbers and amino acid composition of this antimicrobial peptide were determined and optimized based on the existing database of antimicrobial peptides. The newly designed antibacterial peptide YHX-1 is broad-spectrum bactericidal efficient and safe, [247].
The results of peptide-protein docking study of Mustafa et al. [248], based on 10 selected ABPs, among which napin and snaking-1, confirmed by molecular dynamics simulations, demonstrates that the peptide-protein docking computational approach supports the evidence of efficiency of these ABPs as potential inhibitors of bacterial strain specific proteins. Lin et al. [249] perform intelligent de novo design of novel antimicrobial peptides against antibiotic-resistant bacterial strains. They train a Wasserstein generative adversarial network with gradient penalty (WGAN-GP) based on known ABPs to generate novel ABP candidates. The quality of the GAN-designed peptides was evaluated in silico, and eight of them were selected by an ABP Artificial Intelligence (AI) classifier and synthesized for further experiments. This approach shows an efficient way to discover ABPs effective against general and antibiotic-resistant bacteria strains. Such strategy allows other novel functional peptides to be quickly designed, identified, and synthesized for validation on the wet bench, [249].
Szymczak et al. [250] propose a novel approach to a potent antibacterial peptide generation, using a deep generative model HydrAMP (that is a cVAE-based model), which is specifically trained to perform analogue generation of both positives and negatives, as well as unconstrained generation. Nedyalkova et al. [251] present the progress and future of the computational design of antimicrobial peptides (ABPs) as bio-inspired functional molecules. The latest scientific breakthroughs and technologies that could offer new opportunities and alternative strategies for developing novel ABPs are in the focus of this discussion.
Aguilera-Puga et al. [252] discuss the evolution and applications of predictive and generative modelling to discover and design safe and effective antimicrobial peptides as well as the accelerating of discovery and design of antimicrobial peptides with artificial intelligence. Peptides, which modulate many processes of human physiology (targeting ion channels, protein receptors, or enzymes) represent valuable starting points for the development of new biologics against communicable and non-communicable disorders. However, turning native peptide ligands into drug gable materials requires high selectivity and efficacy, predictable metabolism, and good safety profiles.
4. Biogenic Synthesized Metal, Metal Oxide and Composite Nanoparticles
The use of nanomaterials is accepted as promising approach in the fighting of bacterial infections and antibiotic resistance. Unlike the antibiotics, which are administered to treat diseases and infections in the patients, the nanomaterials provide an opportunity to limit microbial growth prior to human infection, [253].
Many novel antibacterial nanomaterials were developed during the last years and some of them are already applied in some hospitals, [254]. The experimentally proved remarkable antimicrobial activity of metal and metal oxide nanoparticles as well as of metal nanocomposites seems to be a very interesting for alternative treatment of multidrug resistant microbial infections, [255,256]. The nanoparticles (NPs) are small enough to pass through the cell membranes of pathogenic bacteria and interfere with essential molecular pathways, [256]. The antibacterial activity of the metallic nanoparticles (NPs) is accepted to be due to interactions with important cellular organelles and biomolecules like DNA, enzymes, ribosomes, and lysosomes that can affect cell membrane permeability, oxidative stress, gene expression, protein, and enzyme activation, [257]. Since the NPs target multiple biomolecules concurrently, it becomes difficult for the bacteria to develop resistance against them, [258].
NPs could be utilized by their including in delivery systems, as intrinsic antimicrobials, or in mixtures. The techniques for obtaining are very important for the properties of the produced metal NPs and hybrid nanostructured materials, the particle size and distribution on size being among the most important ones, [259].
The developed up to now synthesis methods are of three categories: physical, chemical, and biological. The physical methods use a top-down strategy, i.e. a large piece of metal is fragmented into small parts by physical action into progressively smaller fragments. The produced NPs are with a somewhat scattering size distribution. The NPs size mainly determines their antibacterial activity. Therefore, the physical methods are not preferable for preparation of metallic NPs. The chemical methods are based on bottom-up strategies leading to formation of more narrow size distributed metallic NPs. They utilize chemical procedures that use chemical solvents and certain drawbacks such as toxicity and others limit their utilization in the synthesis of metallic NPs. The biosynthesis does not use toxic chemicals, it is eco-friendly and therefore it is favoured among the other methods of NPs creation, [256].
The biogenic synthesis of metal NPs receives global attention as eco-friendly and cost-effective approach. Some microbes were shown to naturally produce metal-based NPs as a method for the detoxifying of heavy metals [260]. However, the biogenic synthesis of metal NPs was in details investigated in recent decades, and since then, metal-based NPs find application in the production of cosmetics and textiles. The adaptability of these substances attracts the world’s scientific attention, and the development of novel formulations, applications, and synthesis techniques continue, [256].
It is known that living organisms like bacteria, yeast, fungi, and plants can reduce inorganic metal ions into metal NPs by their cellular metabolites and thus to produce various metal and metal oxide NPs. This is the base of the current biogenic synthesis of NPs. Both the yield and stability of the biogenic NPs are quite satisfactory. Photosynthesis of NPs from plants seems to be a very effective way for the development of a rapid, clean, nontoxic, and eco-friendly technologies for obtaining of a variety of biologically active nanomaterials for medical applications, [258].
Extracts from different plant species are used for biosynthesis of NPs, such as silver (Ag), cerium dioxide (Ce2O2), copper oxide (CuO), gold (Au), titanium dioxide (TiO2), and zinc oxide (ZnO). Plant extracts could reduce the use of hazardous compounds and a harsh reactions in the production of metallic NPs, [261]. Plant, fungal, algal, and cyanobacterial extract solutions were employed as nucleation/capping agents in the green bioprocess to produce effective nanomaterials for medical applications. Biogenic synthesized NPs show wound-healing antibacterial, anticancer, and antifungal properties, [262]. The biogenic syntheses of metallic NPs from seed extracts is presented as a cost-effective alternate to other biological methods due to low maintenance costs, culture/growth independence for biomass, and diversity of phytochemicals as reducing and capping agents. Recently, seed extracts become popular because of the single-step green synthesis of metal NPs with satisfactory antimicrobial activity, [263].
The utilization of phytochemicals from plant extracts become unique technology for the synthesis of NPs, as they possess dual nature of reducing and capping agents to the NPs. Green synthetized NPs attract extensive interest worldwide because of their biocompatibility and huge potential for utilization as antimicrobial agents, as well as in the cancer/gene therapy, in energy harvesting etc. The biological reduction of metals provides NPs of desirable size and morphology with better physicochemical characteristics, lower toxicity, and high stability. Fungi are the most preferred for the biogenic synthesis of NPs due to high biomass production, secretion of a high quantity of extracellular proteins that stabilize the NPs, ease in handling, and high metal tolerance, [264,265].
Currently, NPs synthesized using plant or microbial extracts and conjugated with biologically active components are discussed as a safe alternative approach for therapeutic applications as they are environmentally friendly and cost-effective. Furthermore, NPs conjugated with natural biomolecules have improved bioavailability and minimal side effects, as they are smaller in size and have higher permeability in addition to being reducing and stabilizing agents possessing excellent antioxidant activity. NPs serve as potential antimicrobial agents due to their affinity towards sulphur-rich amino acids, adhere to microbial cell walls by means of electrostatic attraction, and disrupt the cytoplasmic membrane along with the nucleic acid of microbes, [266].
Bhati [267] describes the green methods for the synthesis of biogenic NPs, involving various plant extracts which are nontoxic, environmentally friendly and value effective. The natural plant extracts contain metabolites like flavonoids, terpenoids, polyphenols, alkaloids, etc. which acts as both reduction and stabilization agents for synthesis of biogenic NPs with desired shape and size. Furthermore, the employment of assorted plant extracts and bacteria, fungi, proteins, etc. within the biogenic synthesis of NPs are described shortly. It is anticipated that transition metals will be the ideal choice for a production of metal-based nanoparticles because they have partially filled d-orbitals, which makes them more redox-active, [268].
Numerous approaches to green synthesis of different metal NPs, metal oxide NPs, and nanocomposites by using a variety of natural reducing agents originating from terrestrial biota are described in the special literature. Strategies for improving the antibacterial capability of the NPs through surface modification and their potential for applications in antimicrobial performance targeted at the eradicating of multi-drug resistant bacteria are also investigated. Although research was recently extended to less-common metals, the materials most extensively studied and used in metal-based NPs, remain silver, gold, copper, iron, and zinc.
4.1. Biosynthesized Silver Nanoparticles
Due to their intrinsic therapeutic properties and the broad-spectrum antibacterial activity, the silver NPs were recognized as potential candidates for development of novel approaches to the treatment of various bacterial diseases, [269,270,271]. Their bactericidal activity is associated with a triggering the oxidative stress, protein dysfunction, membrane and DNA damage that leads to damage of the microbial cells, [272]. The size, shape, and concentration play an important role in the antimicrobial activity of the silver NPs. As antibacterial agents, the silver ions are usually more effective than the silver NPs. However, modified or functionalized silver NPs could be extremely active to kill bacteria than pure silver NPs, [273]. For example, the modification of silver nanoparticles with titanate nanotubes changes their physicochemical properties (such as stability, size, oxidation state, and shape), resulting in enhanced antibacterial, catalytic, and photocatalytic activity, [254]. Exhibiting multiple and simultaneous mechanisms of action, the silver NPs are active against Gram-negative and Gram-positive bacteria, including multidrug resistant strains. In combination with antibiotics or organic compounds as antibacterial agents, they demonstrate synergistic effect against pathogen bacteria such as Escherichia coli and Staphylococcus aureus, [274].
Different approaches to green synthesis of silver NPs by use of natural reducing agents originating from the terrestrial biota were presented in the literature. Gupta et al. [275] accomplish green synthesis of silver NPs for wound dressing applications using curcumin-cyclodextrin complexes, loaded into bacterial cellulose-based hydrogels. Curcumin, that is a natural polyphenolic compound (well known as a wound-healing agent) is used as a natural reducing agent. The hydrophobicity of the curcumin overcomes by its microencapsulation in cyclodextrin. These novel dressings exhibit antimicrobial activity against three common wound-infecting pathogenic microbes: S. aureus, Pseudomonas aeruginosa, and Candida auras. Das et al. [276] highlight silver NPs synthesis mediated by plant and algae derivatives and discuss their application as antibacterial agents with emphasize of their role for providing further better health, environment and prevention from infectious diseases. Feroze et al. [277] perform fungal mediated biosynthesis of silver NPs in presence of fungal metabolites of Penicillium oxalicum (characterized by XRD and SEM). The evaluation of the antimicrobial activity (by disc diffusion test and UV-Visible Spectrophotometry) of the biosynthesized silver NPs, demonstrates their excellent activity against Staphylococcus aureus, S. dysenteriae, and Salmonella typhi and indicates that such silver NPs might be useful as bactericidal agents against drug resistant bacteria. Garibo et al. [278] find that green synthesized silver NPs by using Lysiloma acapulcensis (a perennial tree used in the traditional Mexico medicine) exhibit higher antimicrobial activity than that of chemically produced silver NPs, maintaining their low-cytotoxicity. The obtained antimicrobial potency was as follows: E. coli ≥ S. aureus ≥ P. aeruginosa > C. albicans. FTIR and LC–MS results show the presence of chemical groups that could act as either (i) reducing agents stabilizing the silver NPs or (ii) antimicrobial capping agents enhancing antimicrobial properties of silver NPs. Rahimi et al. [279] present recent progress in the antimicrobial wound dressings based on carbohydrate polymer-based silver nanocomposites. The methods of synthesis, physicochemical properties, healing efficiencies, toxicity against human tissues, antibacterial and antifungal effects of each material are discussed. Barabadi et al. [280] use simple, non-toxic and fast method to fabricate Zataria multiflora-derived silver NPs. The antibacterial activity evaluation against Staphylococcus aureus demonstrates the higher activity of the plant-mediated fabricated silver NPs as compared to that of commercial silver NPs. Ramires-Rosas et al. [281] present a green route to produce silver NPs using the bioactive flavonoid quercetin as a reducing agent and food anti-caking agents as stabilizers.
Chavan et al. [282] describe a green synthetic approach to produce silver NPs using an Artocarpus heterophyllus leaf extract and Design Expert Ver. 13 to optimize their parameters. The optimized silver NPs were characterized by UV–Vis and FTIR spectroscopy. Antioxidant and antimicrobial potential were determined in vitro using standard protocols. The optimized nanoparticles appear to be spherical, with average particle diameter of 100–110 nm (SEM, TEM) and show effective antibacterial, antioxidant, and antifungal activity.
Mahalingam et al. [283] synthetize biogenic silver NPs with a spherical shape, average particle size of 15 to 25 nm, that are stable and monodispersed, using ethanol extract of Catharanthus roseus flower. UV–Vis and FTIR spectroscopy; XRD; Particle Size Analysis; TEM; and EDX characterized the NPs. These silver NPs reveal superior antibacterial activity against human pathogenic bacteria with a remarkable inhibition zone for Salmonella typhimurium (10–14 mm), Bacillus subtilis (6–11 mm), for Enterococcus faecalis (11–14 mm) and Shigella boydii (9–10 mm).
Said et al. [284] synthetize silver NPs which are active against the common pathogens for urinary tract infections, using Egyptian henna leaves (Lawsonia inermis) extracts. Plant extract components were identified by GC-MS and the analysis of the prepared silver NPs was carried out through UV–Vis, XRD, TEM, SEM, and FTIR spectroscopy. Antibacterial activities of the obtained silver NPs were examined against common pathogens from urinary tract. Very high sensitivity of all test pathogens to the biologically synthesized silver NPs was observed.
Hani et al. [285] use orange peel extract in a biogenic synthesis of silver NPs with aim to utilize an agro-industrial byproduct, specifically Citrus sinensis peels, as a reservoir of polyphenols. The natural chemicals presented in C. sinensis peels serve as reducing agents in an environmentally benign method for synthesis of silver NPs. The last one were characterized by UV–Vis spectroscopy, Dynamic Light Scattering (DLS), SEM, EDX, and TEM. Their effectiveness in inhibiting growth and biofilm formation of E. coli, Staphylococcus aureus, and Candida albicans was demonstrated simultaneously with significant toxic effects against human prostate cancer cell line DU145 (as investigated by anti-apoptotic, 4′,6-diamidino-2-phenylindole (DAPI), reactive oxygen species (ROS), and acridine orange/ethidium bromide (AO/EtBr) assays). All findings confirm that this approach can serve as a cost-effective, non-toxic, and environmentally friendly technology for green synthesis of medical silver NPs, that offers an alternative recycling strategy and contributes to the sustainable use of biological by-products.
Silver/silica nanocomposite (Ag/SiO2) was biosynthesized at room temperature using the crude extract of Escherichia coli metabolites in presence of sunlight. It was characterized by UV-Vis spectrophotometry, XRD, FTIR and TEM. The Ag/SiO2 NPs show average size of ~32 – 48 nm whereas silver NPs show a mean size of 18 – 24 nm. The negative charged Ag/SiO2 (-31.0 mV) indicate potential antimicrobial activity against Bacillus cereus, Klebsiella pneumoniae, Staphylococcus aureus, E. coli, Candida albicans, and Botrytis cinerea. The minimum inhibitory concentration (MIC) test shows a dose-dependent antimicrobial action of the Ag/SiO2 NPs. In addition, a formation of a mucilage matrix connecting the hyphal cells were observed as well as a big vacuoles and lipid droplets appearance with severe leakage of cytoplasmic contents of the treated B. cinerea, [286].
4.2. Biosynthesized Zinc Oxide Nanoparticles
Naseer et al. [287] develop green route to synthesis of zinc oxide nanoparticles (NPs) using leaf extracts of Cassia fistula and Melia azadarach and prove their antibacterial potential. Gur et al. [288] also present green synthesis of biogenic zinc oxide NPs, their characterization and bioactivity evaluation. Since ZnO and TiO2 have no impact on human cells, NPs-based wound care solutions by utilizing these two oxides are presented as relatively new approach compared to conventional materials. Veselova et al. demonstrate the long-term antibacterial efficacy of textiles coated with ZnO and TiO2 nanoparticles in a tropical environment, [289]. Biogenic synthesis and characterization of ZnO nanoparticles for degradation of synthetic dyes were performed as a sustainable environmentally clean approach, [290].
Green synthesized ZnO and vanadium-doped ZnO nanoparticles, using Vinca rosea plant leaf, were characterized by FTIR, XRD, and SEM-EDX. Their testing for biomedical applications (antibacterial, anticancer, antioxidant and antidiabetic) show the higher antioxidant, antidiabetic, and anticancer activity of Vinca rosea capped ZnO NPs than vanadium-doped ZnO NPs, both prepared by one the same green approach, [291].
ZnO NPs and Sesbania grandiflora are known for their biocompatibility and medicinal properties, such as anti-cancer, anti-microbial, anti-diabetic, and anti-oxidant. Ramasubbu et al. [292] present green synthesis of ZnO NPs using S. grandiflora and the evaluation of antimicrobial, antidiabetic and cytotoxic effects.
4.3. Biosynthesized Coper Nanoparticles
A study on the biosynthesis of copper nanoparticles (NPs) mediated by a rare medicinal plant Cissus arnotiana was reported together with the antibacterial activity against Gram-negative and Gram-positive bacteria. The biosynthesized coper NPs show high antibacterial activity against the Gram-negative bacterium, E. coli with an inhibition zone of 22.20 ± 0.16 mm at 75 μg/ml, [293].
A strategy for the synthesis of coper NPs was developed that is based on the use of enzymes as stabilizing agents generating metal-enzyme nanobiohybrids. The enzymes facilitate the in situ formation of NPs under mild synthesis conditions (water medium and room temperature) in absence of reducing agents. Furthermore, the use of a protein allows a control over the size and shape as well as over the production of monodisperse metal NPs, [294].
Various physical, chemical, and biological methods used for CuO NPs preparation as well as affected by them physicochemical and biological properties of the synthetized CuO NPs are under discussion regarding potential biomedical applications, [295]. A recent study of Ortega-Nieto et al. [296] demonstrates the effect of a reduction step inclusion in the coper nanobiohybrids synthesis. Changes in the metal structure and antibacterial efficiencies were observed that are depending on the added amount of reducing agent. Copper sulphate as a metal salt, lipase from Candida antarctica (CAL-B) as a scaffold enzyme, and sodium borohydride (NaBH4) as a reducing agent were used. Hybrids were synthetized by using of different percentages NaBH4 from 0% to 100%. XRD and TEM analyses demonstrate different oxidation states of the coper and sizes for the coper NPs depending on the degree of reduction. Predominant Cu(0), larger NPs (with a maximum size of 13 nm) and agglomerated nanohybrids were found at higher NaBH4 amounts whereas at lower NaBH4 amounts, Cu(I) species and smaller NPs (particle size less than 6 nm) are predominant.
4.4. Biosynthesized Iron Oxide Nanoparticels
Biosynthesis of iron oxide NPs via a composite of Psidium guavaja-Moringa oleifera is reported as a benign, facile, biocompatible, cost-effective and eco-friendly approach. The antibacterial and photocatalytic studies (FTIR, UV-Vis, XRD, TEM and VSM, Vibrating Sample Magnetometer) demonstrate that the in this way synthetized FeO NPs inhibit the growth of six human pathogens with higher activity at lower concentrations compared to the orthodontic antibacterial drugs, [297].
4.5. Biosynthesized Selenium Nanoparticles
Biogenic selenium NPs were synthesized using herbal extract of Triphala. The assessment of their antimicrobial and antioxidant activity and toxicity indicates that the Triphala-augmented biogenic selenium NPs have a great potential as oral antimicrobial agents with superior biocompatibility and antioxidant properties. The antimicrobial activity was assessed against Streptococcus mutans, S. aureus, E. faecalis and Candida albicans by agar diffusion test, and DPPA assay was used to evaluate the antioxidant potential of the selenium NPs. Their toxicity was tested on brine shrimp, [298].
4.6. Others
Combinations of metal NPs and other antibacterial agents, like farnesol for example, are also studied as a potential tool to mitigate the problem of AMR. The assessment of the effectiveness of antibacterial agents, composed of farnesol and nanoparticles (silver, gold, copper, and zinc oxide), in the degradation of biofilms, produced by pathogenic microorganisms (Escherichia coli, Enterococcus faecalis, Staphylococcus aureus, Pseudomonas aeruginosa, and Candida albicans) demonstrates the great changes in the biofilm structure caused by Ag-farnesol composite altering the process of biofilm formation, [299].
Biogenic synthetized metal and metal oxide NPs with their narrow particles size distribution, large surface area-to-volume ratio and high biological activity offer numerous options for development of antibacterial agents to treat bacterialial infections [300].
5. Concluding Remarks
Terrestrial biota derived antibacterial products and substances open a variety of new ways to improved modern therapeutic strategies. Natural products serve as key sources of novel chemical diversity as well as integral components of currently accessible drug formulations. Originating from plants, animals and microbes they have a wide range of chemical and functional variability and show great efficiency for treating of infectious diseases with fewer side effects, synergy, and capacity to combating drug tolerance.
New terrestrial sources of known antibacterial agents and new antibacterial agents from terrestrial biota were discovered during the last 5 years, which are under investigation together with long ago known but now surviving their renaissance for the development of new medical treatments. Some natural terrestrial biota derived antibacterial materials and based on them antibacterial agents are more efficient, and with lower side effects than the chemically synthesized counter parts.
Development of novel derivatives that avoid some disadvantages of the natural antibacterial compounds and modification of natural products to adjust their properties to different medical applications using natural led molecules and mashing design were performed for the creation of new pharmaceutical products active against different bacterial pathogens including against multi-drug resistant such.
Well-known herb and medicinal plant extracts as well as bacteriophages survive now their renaissance as a base for development of antibacterial drugs, antibiotic adjuvants and individual therapies. In the area of plant drug discovery, much progress is made in the generation and analysis of large chemical datasets.
Special efforts were devoted to the locking for novel antibacterial peptides and bacteriophages as well as for biogenic synthetized metal, metal oxide and composite nanoparticles. A number of new antimicrobial peptides and their modified variations, with in vitro proved high antibacterial activity, are adjusted/in adjustment for development of drugs for specific bacterial diseases, some of them being in clinical trials. The peptide antibiotics are already accepted as one of the most important tools in the combat with multi-drug resistant bacteria. Bacteriophages, especially designed for concerned individuals, are another hope for successful treatment of heavy multidrug resistant infections.
Antibacterial activity of biogenic synthetized metal, metal oxide and composite nanoparticles is very promising and some of them are already in clinical application, for example in healing of wounds, orthopedic coatings, etc.
The use of engineered nanomaterials, such as drug delivery systems containing natural antibacterial substances lids to increased efficiency of the antibacterial treatments improving the availability, pharmacokinetic and others.
Author Contributions
Conceptualization, T.V.; software, D.G.; data curation, T.V., D.G.; writing - original draft preparation, T.V., D.G.; writing - review and editing, D.G., Y.S., B.M.; supervision, T.V.; project administration, T.V; funding acquisition, T.V. All authors have read and agreed to the published version of the manuscript.
Funding
Bulgarian National Scientific Fund, grant number КП-06-КОСТ/11/07.08.2023, funded this research.
Institutional Review Board Statement
The study did not require ethical approval.
Data Availability Statement
This is review paper and no new data were created.
Acknowledgments
Authors gratefully acknowledge the financial support from the Bulgarian National Scientific Fund, (Grand КП-06-КОСТ/11/07.08.2023) and COST Action CA21145 EURESTOP for providing a stimulating environment that led to this research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ye, L.; Zhang, J.; Xiao, W.; Liu, S. Efficacy and Mechanism of Actions of Natural Antimicrobial Drugs. Pharmacology & Therapeutics 2020, 216, 107671. [Google Scholar] [CrossRef]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. The Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Maarten van Dongen. AMR Insights in 2023 in a Bird’s Eye View; AMR Insights: Amsterdam, The Netherlands, 2024; p. 18. [Google Scholar]
- World Health Organization 10 Threats to Global Health in 2018. Available online: https://medium.com/who/10-threats-to-global-health-in-2018-232daf0bbef3 (accessed on 12 August 2024).
- Health Promotion Board Antimicrobial Resistance. Available online: https://hpb.gov.sg/healthy-living/preventive-health/antimicrobial-resistance (accessed on 12 August 2024).
- Li, W.; Separovic, F.; O’Brien-Simpson, N.M.; Wade, J.D. Chemically Modified and Conjugated Antimicrobial Peptides against Superbugs. Chem. Soc. Rev. 2021, 50, 4932–4973. [Google Scholar] [CrossRef] [PubMed]
- Pulingam, T.; Parumasivam, T.; Gazzali, A.M.; Sulaiman, A.M.; Chee, J.Y.; Lakshmanan, M.; Chin, C.F.; Sudesh, K. Antimicrobial Resistance: Prevalence, Economic Burden, Mechanisms of Resistance and Strategies to Overcome. European Journal of Pharmaceutical Sciences 2022, 170, 106103. [Google Scholar] [CrossRef]
- Jonas, O.B.; Irwin, A.; Berthe, F.C.J.; Le Gall, F.G.; Marquez, P. Drug-Resistant Infections : A Threat to Our Economic Future (Vol. 2) : Final Report. Available online: https://documents.worldbank.org/en/publication/documents-reports/documentdetail/323311493396993758/final-report (accessed on 10 August 2024).
- World Health Organization. World Health Statistics 2021: Monitoring Health for the SDGs, Sustainable Development Goals; World Health Organization: location, 2021; p. 136. [Google Scholar]
- World Health Organization. WHO Bacterial Priority Pathogens List, 2024 Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance; World Health Organization: location, 2024; p. 72. [Google Scholar]
- Qadri, H.; Shah, A.H.; Mir, M. Novel Strategies to Combat the Emerging Drug Resistance in Human Pathogenic Microbes. Current Drug Targets 2021, 22, 1424–1436. [Google Scholar] [CrossRef] [PubMed]
- Symochko, L.; Demyanyuk, O.; Symochko, V.; Grulova, D.; Fejer, J.; Mariychuk, R. The Spreading of Antibiotic-Resistant Bacteria in Terrestrial Ecosystems and the Formation of Soil Resistome. Land 2023, 12, 769. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Bush, K.; Harbarth, S.; Paul, M.; Rex, J.H.; Tacconelli, E.; Thwaites, G.E. Critical Analysis of Antibacterial Agents in Clinical Development. Nat Rev Microbiol 2020, 18, 286–298. [Google Scholar] [CrossRef]
- Abushaheen, M.A.; Muzaheed; Fatani, A.J.; Alosaimi, M.; Mansy, W.; George, M.; Acharya, S.; Rathod, S.; Divakar, D.D.; Jhugroo, C.; et al. Antimicrobial Resistance, Mechanisms and Its Clinical Significance. Disease-a-Month 2020, 66, 100971. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.R.M.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K.; et al. Antibiotic Resistance in Microbes: History, Mechanisms, Therapeutic Strategies and Future Prospects. Journal of Infection and Public Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef]
- Bösch, A.; Macha, M.E.; Ren, Q.; Kohler, P.; Qi, W.; Babouee Flury, B. Resistance Development in Escherichia Coli to Delafloxacin at pHs 6.0 and 7.3 Compared to Ciprofloxacin. Antimicrobial Agents and Chemotherapy 2023, 67, e01625-22. [Google Scholar] [CrossRef] [PubMed]
- Jubeh, B.; Breijyeh, Z.; Karaman, R. Resistance of Gram-Positive Bacteria to Current Antibacterial Agents and Overcoming Approaches. Molecules 2020, 25, 2888. [Google Scholar] [CrossRef]
- Breijyeh, Z.; Jubeh, B.; Karaman, R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules 2020, 25, 1340. [Google Scholar] [CrossRef] [PubMed]
- Mir, M.A.; Aisha, S.; Qadri, H.; Jan, U.; Yousuf, A.; Jan, N. Chapter 2 - Evolution of Antimicrobial Drug Resistance in Human Pathogenic Bacteria. In Human Pathogenic Microbes; Developments in Microbiology; Mir, M.A., Ed.; Academic Press: lication, 2022; pp. 31–52. ISBN 978-0-323-96127-1. [Google Scholar]
- Helmy, Y.A.; Taha-Abdelaziz, K.; Hawwas, H.A.E.-H.; Ghosh, S.; AlKafaas, S.S.; Moawad, M.M.M.; Saied, E.M.; Kassem, I.I.; Mawad, A.M.M. Antimicrobial Resistance and Recent Alternatives to Antibiotics for the Control of Bacterial Pathogens with an Emphasis on Foodborne Pathogens. Antibiotics 2023, 12, 274. [Google Scholar] [CrossRef]
- Browne, K.; Chakraborty, S.; Chen, R.; Willcox, M.D.; Black, D.S.; Walsh, W.R.; Kumar, N. A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides. International Journal of Molecular Sciences 2020, 21, 7047. [Google Scholar] [CrossRef]
- Stan, D.; Enciu, A.-M.; Mateescu, A.L.; Ion, A.C.; Brezeanu, A.C.; Stan, D.; Tanase, C. Natural Compounds With Antimicrobial and Antiviral Effect and Nanocarriers Used for Their Transportation. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- Butler, M.S.; Paterson, D.L. Antibiotics in the Clinical Pipeline in October 2019. J Antibiot 2020, 73, 329–364. [Google Scholar] [CrossRef] [PubMed]
- Vladkova, T.; Georgieva, N.; Staneva, A.; Gospodinova, D. Recent Progress in Antioxidant Active Substances from Marine Biota. Antioxidants 2022, 11, 439. [Google Scholar] [CrossRef]
- Vladkova, T.G.; Martinov, B.L.; Gospodinova, D.N. Anti-Biofilm Agents from Marine Biota. Journal of Chemical Technology and Metallurgy 2023, 58, 825–839. [Google Scholar] [CrossRef]
- Vladkova, T.G.; Staneva, A.D.; Avramova, I.A.; Ivanova, I.A.; Gospodinova, D.N. Fucoidan-Containing, Low-Adhesive Siloxane Coatings for Medical Applications: Inhibition of Bacterial Growth and Biofilm Development. Materials 2023, 16, 3651. [Google Scholar] [CrossRef]
- Vladkova, T.G.; Martinov, B.L.; Staneva, A.D.; Ivanova, I.A.; Gospodinova, D.N.; Albu-Kaya, M.G. Preparation and Antimicrobial Activity of Fucoidan Containing Collagen/(ZnTiO3/SiO2) Composites. Journal of Chemical Technology and Metallurgy 2023, 58, 654–663. [Google Scholar] [CrossRef]
- Vladkova, T.G.; Monov, D.M.; Akuzov, D.T.; Ivanova, I.A.; Gospodinova, D. Comparative Study of the Marinobacter Hydrocarbonoclasticus Biofilm Formation on Antioxidants Containing Siloxane Composite Coatings. Materials 2022, 15, 4530. [Google Scholar] [CrossRef]
- Farha, M.A.; Brown, E.D. Drug Repurposing for Antimicrobial Discovery. Nat. Microbiol. 2019, 4, 565–577. [Google Scholar] [CrossRef]
- Qi, M.; Chi, M.; Sun, X.; Xie, X.; Weir, M.D.; Oates, T.W.; Zhou, Y.; Wang, L.; Bai, Y.; Xu, H.H. Novel Nanomaterial-Based Antibacterial Photodynamic Therapies to Combat Oral Bacterial Biofilms and Infectious Diseases. IJN 2019, 14, 6937–6956. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Ahmad, M.N.; Dasgupta, A.; Rana, P.; Srinivas, N.; Chopra, S. Novel Approaches for the Treatment of Infections Due to Multidrug-Resistant Bacterial Pathogens. Future Medicinal Chemistry 2022, 14, 1133–1148. [Google Scholar] [CrossRef] [PubMed]
- Moo, C.-L.; Yang, S.-K.; Yusoff, K.; Ajat, M.; Thomas, W.; Abushelaibi, A.; Lim, S.-H.-E.; Lai, K.-S. Mechanisms of Antimicrobial Resistance (AMR) and Alternative Approaches to Overcome AMR. Current Drug Discovery Technologies 2020, 17, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Ullah, F.; Ovais, M.; Ahmed, J.; Devkota, H.P. Synergistic Interactions of Phytochemicals with Antimicrobial Agents: Potential Strategy to Counteract Drug Resistance. Chemico-Biological Interactions 2019, 308, 294–303. [Google Scholar] [CrossRef]
- Tyers, M.; Wright, G.D. Drug Combinations: A Strategy to Extend the Life of Antibiotics in the 21st Century. Nat Rev Microbiol 2019, 17, 141–155. [Google Scholar] [CrossRef]
- Zharkova, M.S.; Orlov, D.S.; Golubeva, O.Y.; Chakchir, O.B.; Eliseev, I.E.; Grinchuk, T.M.; Shamova, O.V. Application of Antimicrobial Peptides of the Innate Immune System in Combination With Conventional Antibiotics—A Novel Way to Combat Antibiotic Resistance? Front. Cell. Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef]
- Douafer, H.; Andrieu, V.; Phanstiel, O.I.; Brunel, J.M. Antibiotic Adjuvants: Make Antibiotics Great Again! J. Med. Chem. 2019, 62, 8665–8681. [Google Scholar] [CrossRef]
- Mir, M.A.; Kumawat, M.; Nabi, B.; Kumar, M. Chapter 8 - Combinatorial Approach to Combat Drug Resistance in Human Pathogenic Bacteria. In Human Pathogenic Microbes; Developments in Microbiology; Mir, M.A., Ed.; Academic Press: location, 2022; pp. 187–206. ISBN 978-0-323-96127-1. [Google Scholar]
- León-Buitimea, A.; Garza-Cárdenas, C.R.; Garza-Cervantes, J.A.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. The Demand for New Antibiotics: Antimicrobial Peptides, Nanoparticles, and Combinatorial Therapies as Future Strategies in Antibacterial Agent Design. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Moammeri, A.; Chegeni, M.M.; Sahrayi, H.; Ghafelehbashi, R.; Memarzadeh, F.; Mansouri, A.; Akbarzadeh, I.; Abtahi, M.S.; Hejabi, F.; Ren, Q. Current Advances in Niosomes Applications for Drug Delivery and Cancer Treatment. Materials Today Bio 2023, 23, 100837. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Qadri, H.; Haseeb Shah, A.; Mudasir Ahmad, S.; Alshehri, B.; Almilaibary, A.; Ahmad Mir, M. Natural Products and Their Semi-Synthetic Derivatives against Antimicrobial-Resistant Human Pathogenic Bacteria and Fungi. Saudi Journal of Biological Sciences 2022, 29, 103376. [Google Scholar] [CrossRef]
- Arrowsmith, C.H. Structure-Guided Drug Discovery: Back to the Future. Nat Struct Mol Biol 2024, 31, 395–396. [Google Scholar] [CrossRef]
- Süntar, I. Importance of Ethnopharmacological Studies in Drug Discovery: Role of Medicinal Plants. Phytochem Rev 2020, 19, 1199–1209. [Google Scholar] [CrossRef]
- Malekian, A.; Esmaeeli Djavid, G.; Akbarzadeh, K.; Soltandallal, M.; Rassi, Y.; Rafinejad, J.; Rahimi Foroushani, A.; Farhoud, A.R.; Bakhtiary, R.; Totonchi, M. Efficacy of Maggot Therapy on Staphylococcus Aureus and Pseudomonas Aeruginosa in Diabetic Foot Ulcers: A Randomized Controlled Trial. Journal of Wound Ostomy & Continence Nursing 2019, 46, 25. [Google Scholar] [CrossRef]
- Porras, G.; Chassagne, F.; Lyles, J.T.; Marquez, L.; Dettweiler, M.; Salam, A.M.; Samarakoon, T.; Shabih, S.; Farrokhi, D.R.; Quave, C.L. Ethnobotany and the Role of Plant Natural Products in Antibiotic Drug Discovery. Chem. Rev. 2021, 121, 3495–3560. [Google Scholar] [CrossRef]
- Wu, S.-C.; Liu, F.; Zhu, K.; Shen, J.-Z. Natural Products That Target Virulence Factors in Antibiotic-Resistant Staphylococcus Aureus. J. Agric. Food Chem. 2019, 67, 13195–13211. [Google Scholar] [CrossRef]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial Plant Compounds, Extracts and Essential Oils: An Updated Review on Their Effects and Putative Mechanisms of Action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial Activity of Flavonoids and Their Structure–Activity Relationship: An Update Review. Phytotherapy Research 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, B.A.; Bhat, B.A.; Ahmad, Z.; Mir, M.A. Strategies Employed to Evade the Host Immune Response and the Mechanism of Drug Resistance in Mycobacterium Tuberculosis: In Search of Finding New Targets. Current Pharmaceutical Biotechnology 2022, 23, 1704–1720. [Google Scholar] [CrossRef]
- Mayo-Muñoz, D.; Pinilla-Redondo, R.; Camara-Wilpert, S.; Birkholz, N.; Fineran, P.C. Inhibitors of Bacterial Immune Systems: Discovery, Mechanisms and Applications. Nat Rev Genet 2024, 25, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Vaishampayan, A.; Grohmann, E. Antimicrobials Functioning through ROS-Mediated Mechanisms: Current Insights. Microorganisms 2022, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.A.R.; Nadeem, S.; Komal, S.; Naqvi, S.A.A.; Mubarik, M.S.; Qureshi, S.Y.; Ahmad, S.; Abbas, A.; Zahid, M.; Khan, N.-U.-H.; et al. Antioxidants: Natural Antibiotics. In Antioxidants; IntechOpen: location, 2019; ISBN 978-1-78923-920-1. [Google Scholar]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic Discovery: History, Methods and Perspectives. International Journal of Antimicrobial Agents 2019, 53, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Franco, C.M.; Vázquez, B.I. Natural Compounds as Antimicrobial Agents. Antibiotics 2020, 9, 217. [Google Scholar] [CrossRef]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the Sustainable Discovery and Development of New Antibiotics. Nat Rev Chem 2021, 5, 726–749. [Google Scholar] [CrossRef]
- Elmaidomy, A.H.; Shady, N.H.; Abdeljawad, K.M.; Elzamkan, M.B.; Helmy, H.H.; Tarshan, E.A.; Adly, A.N.; Hussien, Y.H.; Sayed, N.G.; Zayed, A.; et al. Antimicrobial Potentials of Natural Products against Multidrug Resistance Pathogens: A Comprehensive Review. RSC Adv. 2022, 12, 29078–29102. [Google Scholar] [CrossRef]
- Mir, M.A.; Usman, M.; Qadri, H.; Aisha, S. Chapter 10 - Recent Trends in the Development of Bacterial and Fungal Vaccines. In Human Pathogenic Microbes; Developments in Microbiology; Mir, M.A., Ed.; Academic Press: location, 2022; pp. 233–259. ISBN 978-0-323-96127-1. [Google Scholar]
- Jabborova, D.; Davranov, K.; Egamberdieva, D. Antibacterial, Antifungal, and Antiviral Properties of Medical Plants. In Medically Important Plant Biomes: Source of Secondary Metabolites; Egamberdieva, D., Tiezzi, A., Eds.; Springer: Singapore, 2019; pp. 51–65. ISBN 9789811395666. [Google Scholar]
- Parham, S.; Kharazi, A.Z.; Bakhsheshi-Rad, H.R.; Nur, H.; Ismail, A.F.; Sharif, S.; RamaKrishna, S.; Berto, F. Antioxidant, Antimicrobial and Antiviral Properties of Herbal Materials. Antioxidants 2020, 9, 1309. [Google Scholar] [CrossRef]
- Goodhealth Herbal Antimicrobials: Fighting Infections Naturally. Available online: https://goodhealth.co.nz/herbal-antibiotics-fighting-infections-naturally-2/ (accessed on 10 August 2024).
- Gong, C. What Is the Most Effective Natural Antibiotic for Tooth Infections. Available online: https://europe.oclean.com/blogs/tips/what-is-the-strongest-natural-antibiotic-for-tooth-infection (accessed on 10 August 2024).
- Adamczak, A.; Ożarowski, M.; Karpiński, T.M. Curcumin, a Natural Antimicrobial Agent with Strain-Specific Activity. Pharmaceuticals 2020, 13, 153. [Google Scholar] [CrossRef]
- Zheng, D.; Huang, C.; Huang, H.; Zhao, Y.; Khan, M.R.U.; Zhao, H.; Huang, L. Antibacterial Mechanism of Curcumin: A Review. Chemistry & Biodiversity 2020, 17, e2000171. [Google Scholar] [CrossRef]
- Khaleghian, M.; Sahrayi, H.; Hafezi, Y.; Mirshafeeyan, M.; Moghaddam, Z.S.; Farasati Far, B.; Noorbazargan, H.; Mirzaie, A.; Ren, Q. In Silico Design and Mechanistic Study of Niosome-Encapsulated Curcumin against Multidrug-Resistant Staphylococcus Aureus Biofilms. Front. Microbiol. 2023, 14. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Fazly Bazzaz, B.S. Review on Plant Antimicrobials: A Mechanistic Viewpoint. Antimicrobial Resistance & Infection Control 2019, 8, 118. [Google Scholar] [CrossRef]
- Gorlenko, C.L.; Kiselev, H.Y.; Budanova, E.V.; Zamyatnin, A.A.; Ikryannikova, L.N. Plant Secondary Metabolites in the Battle of Drugs and Drug-Resistant Bacteria: New Heroes or Worse Clones of Antibiotics? Antibiotics 2020, 9, 170. [Google Scholar] [CrossRef] [PubMed]
- El-Ashmawy, I.M.; Aljohani, A.S.M.; Soliman, A.S. Studying the Bioactive Components and Phytochemicals of the Methanol Extract of Rhanterium Epapposum Oliv. Appl Biochem Biotechnol 2024, 196, 2414–2424. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, S.; Luitel, S.; Dahal, R.K. In Vitro Antimicrobial Activity of Some Medicinal Plants against Human Pathogenic Bacteria. Journal of Tropical Medicine 2019, 2019, 1895340. [Google Scholar] [CrossRef]
- Zofou, D.; Shu, G.L.; Foba-Tendo, J.; Tabouguia, M.O.; Assob, J.-C.N. In Vitro and In Vivo Anti-Salmonella Evaluation of Pectin Extracts and Hydrolysates from “Cas Mango” (Spondias Dulcis). Evidence-Based Complementary and Alternative Medicine 2019, 2019, 3578402. [Google Scholar] [CrossRef]
- Rasooly, R.; Molnar, A.; Choi, H.-Y.; Do, P.; Racicot, K.; Apostolidis, E. In-Vitro Inhibition of Staphylococcal Pathogenesis by Witch-Hazel and Green Tea Extracts. Antibiotics 2019, 8, 244. [Google Scholar] [CrossRef]
- Portillo-Torres, L.A.; Bernardino-Nicanor, A.; Gómez-Aldapa, C.A.; González-Montiel, S.; Rangel-Vargas, E.; Villagómez-Ibarra, J.R.; González-Cruz, L.; Cortés-López, H.; Castro-Rosas, J. Hibiscus Acid and Chromatographic Fractions from Hibiscus Sabdariffa Calyces: Antimicrobial Activity against Multidrug-Resistant Pathogenic Bacteria. Antibiotics 2019, 8, 218. [Google Scholar] [CrossRef]
- Faujdar, S.S.; Bisht, D.; Sharma, A. Antibacterial Potential of Neem (Azadirachta Indica) against Uropathogens Producing Beta-Lactamase Enzymes: A Clue to Future Antibacterial Agent? Biomedical and Biotechnology Research Journal (BBRJ) 2020, 4, 232. [Google Scholar] [CrossRef]
- Kaczorová, D.; Karalija, E.; Dahija, S.; Bešta-Gajević, R.; Parić, A.; Ćavar Zeljković, S. Influence of Extraction Solvent on the Phenolic Profile and Bioactivity of Two Achillea Species. Molecules 2021, 26, 1601. [Google Scholar] [CrossRef] [PubMed]
- Tajudin, N.J.; Ismail, I.N.A. Antimicrobial Activity of Kalanchoe Pinnata: A Review. Malaysian Journal of Science Health & Technology 2022, 8, 31–37. [Google Scholar] [CrossRef]
- Hassan, M.; Musa, F.M.; Aliyu, F.; Adamu, A. Antibacterial Activity of Ethanol Leaf Extract of Sida Acuta Against Some Clinical Bacterial Isolates. Int. J. Life Sci. Biotechnol. 2022, 5, 572–580. [Google Scholar] [CrossRef]
- Nasution, A.K.; Wijaya, S.H.; Gao, P.; Islam, R.M.; Huang, M.; Ono, N.; Kanaya, S.; Altaf-Ul-Amin, M. Prediction of Potential Natural Antibiotics Plants Based on Jamu Formula Using Random Forest Classifier. Antibiotics 2022, 11, 1199. [Google Scholar] [CrossRef]
- Pereira, A.G.; Silva, A.; Grosso, C.; Echave, J.; Chamorro, F.; Seyyedi-Mansour, S.; Donn, P.; Fraga-Corral, M.; Barroso, M.F.; Prieto, M.A. Antimicrobial Activity Screening of Camellia Japonica Flowers (Var. Carolyn Tuttle) for Potential Drug Development. Engineering Proceedings 2023, 56, 314. [Google Scholar] [CrossRef]
- Kavaz, D.; Faraj, R.E. Investigation of Composition, Antioxidant, Antimicrobial and Cytotoxic Characteristics from Juniperus Sabina and Ferula Communis Extracts. Sci Rep 2023, 13, 7193. [Google Scholar] [CrossRef] [PubMed]
- El Mannoubi, I. Impact of Different Solvents on Extraction Yield, Phenolic Composition, in Vitro Antioxidant and Antibacterial Activities of Deseeded Opuntia Stricta Fruit. J.Umm Al-Qura Univ. Appll. Sci. 2023, 9, 176–184. [Google Scholar] [CrossRef]
- Soltanian, S.; Sheikhbahaei, M.; Mirtadzadini, M.; Kalantari Khandani, B. Evaluation of Anticancer, Antioxidant and Antibacterial Properties of Methanol Extract of Three Acantholimon Boiss. Species. Avicenna J Phytomed 2020, 10, 641–652. [Google Scholar] [PubMed]
- Senhaji, S.; Lamchouri, F.; Toufik, H. Phytochemical Content, Antibacterial and Antioxidant Potential of Endemic Plant Anabasis Aretioïdes Coss. & Moq. (Chenopodiaceae). BioMed Research International 2020, 2020, 6152932. [Google Scholar] [CrossRef]
- Özcan, K. Antibacterial, Antioxidant and Enzyme Inhibition Activity Capacities of Doronicum Macrolepis (FREYN&SINT): An Endemic Plant from Turkey. Saudi Pharmaceutical Journal 2020, 28, 95–100. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, X.; Yang, F.; Wu, W.; Liu, Y.; Wang, L.; Wang, L.; Wang, Z. Enhanced Bioaccessibility in Vitro and Bioavailability of Ginkgo Biloba Extract Nanoparticles Prepared by Liquid Anti-Solvent Precipitation. International Journal of Food Science & Technology 2019, 54, 2266–2276. [Google Scholar] [CrossRef]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A Comprehensive Review of the Antibacterial, Antifungal and Antiviral Potential of Essential Oils and Their Chemical Constituents against Drug-Resistant Microbial Pathogens. Microbial Pathogenesis 2019, 134, 103580. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef]
- Mahizan, N.A.; Yang, S.-K.; Moo, C.-L.; Song, A.A.-L.; Chong, C.-M.; Chong, C.-W.; Abushelaibi, A.; Lim, S.-H.E.; Lai, K.-S. Terpene Derivatives as a Potential Agent against Antimicrobial Resistance (AMR) Pathogens. Molecules 2019, 24, 2631. [Google Scholar] [CrossRef]
- Abdallah, E.M.; Alhatlani, B.Y.; de Paula Menezes, R.; Martins, C.H.G. Back to Nature: Medicinal Plants as Promising Sources for Antibacterial Drugs in the Post-Antibiotic Era. Plants 2023, 12, 3077. [Google Scholar] [CrossRef]
- Hovorková, P.; Laloučková, K.; Skřivanová, E. Determination of in Vitro Antibacterial Activity of Plant Oils Containing Medium-Chain Fatty Acids against Gram-Positive Pathogenic and Gut Commensal Bacteria. Czech Journal of Animal Science 2018, 63, 119–125. [Google Scholar] [CrossRef]
- Riski, D.G.; Maulana, R.G.R.; Permana, E.; Lestari, I.; Tarigan, I.L. Profile Analysis of Fatty Acids of Tengkawang (Shorea Sumatrana) Oil Using GC-MS and Antibacterial Activity. Indonesian Journal of Chemical Research 2020, 8, 114–119. [Google Scholar] [CrossRef]
- Casillas-Vargas, G.; Ocasio-Malavé, C.; Medina, S.; Morales-Guzmán, C.; Del Valle, R.G.; Carballeira, N.M.; Sanabria-Ríos, D.J. Antibacterial Fatty Acids: An Update of Possible Mechanisms of Action and Implications in the Development of the next-Generation of Antibacterial Agents. Progress in Lipid Research 2021, 82, 101093. [Google Scholar] [CrossRef] [PubMed]
- Kannan, N.; Rao, A.S.; Nair, A. Microbial Production of Omega-3 Fatty Acids: An Overview. Journal of Applied Microbiology 2021, 131, 2114–2130. [Google Scholar] [CrossRef]
- Sidders, A.E.; Kedziora, K.M.; Arts, M.; Daniel, J.-M.; de Benedetti, S.; Beam, J.E.; Bui, D.T.; Parsons, J.B.; Schneider, T.; Rowe, S.E.; et al. Antibiotic-Induced Accumulation of Lipid II Synergizes with Antimicrobial Fatty Acids to Eradicate Bacterial Populations. eLife 2023, 12, e80246. [Google Scholar] [CrossRef]
- Almuhayawi, M.S. Propolis as a Novel Antibacterial Agent. Saudi Journal of Biological Sciences 2020, 27, 3079–3086. [Google Scholar] [CrossRef]
- Mandal, M.D.; Mandal, S. Honey: Its Medicinal Property and Antibacterial Activity. Asian Pacific Journal of Tropical Biomedicine 2011, 1, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Albaridi, N.A. Antibacterial Potency of Honey. International Journal of Microbiology 2019, 2019, 2464507. [Google Scholar] [CrossRef] [PubMed]
- Almasaudi, S. The Antibacterial Activities of Honey. Saudi Journal of Biological Sciences 2021, 28, 2188–2196. [Google Scholar] [CrossRef]
- University of Birmingham Honey and the Antibiotic Crisis. Available online: https://www.birmingham.ac.uk/alumni/giving/honey-and-the-antibiotic-crisis.aspx (accessed on 10 August 2024).
- Zakir-Hussain, M. Scientists Turn to Honey in Their Urgent Search for Alternative Antibiotics. Available online: https://www.aol.com/scientists-turn-honey-urgent-search-103812541.html (accessed on 10 August 2024).
- Ware, I.; Franke, K.; Frolov, A.; Bureiko, K.; Kysil, E.; Yahayu, M.; El Enshasy, H.A.; Wessjohann, L.A. Comparative Metabolite Analysis of Piper Sarmentosum Organs Approached by LC–MS-Based Metabolic Profiling. Nat. Prod. Bioprospect. 2024, 14, 30. [Google Scholar] [CrossRef]
- Mickymaray, S. Efficacy and Mechanism of Traditional Medicinal Plants and Bioactive Compounds against Clinically Important Pathogens. Antibiotics 2019, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Gioia, S.D.; Hossain, M.N.; Conese, M. Biological Properties and Therapeutic Effects of Plant-Derived Nanovesicles. Open Medicine 2020, 15, 1096–1122. [Google Scholar] [CrossRef]
- Vestergaard, M.; Ingmer, H. Antibacterial and Antifungal Properties of Resveratrol. International Journal of Antimicrobial Agents 2019, 53, 716–723. [Google Scholar] [CrossRef]
- Lyu, J.I.; Ryu, J.; Jin, C.H.; Kim, D.-G.; Kim, J.M.; Seo, K.-S.; Kim, J.-B.; Kim, S.H.; Ahn, J.-W.; Kang, S.-Y.; et al. Phenolic Compounds in Extracts of Hibiscus Acetosella (Cranberry Hibiscus) and Their Antioxidant and Antibacterial Properties. Molecules 2020, 25, 4190. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, J.; Li, Q.; Wang, J.; Mu, L.; Hui, L.; Li, M.; Xu, W.; Yang, H.; Wei, L. A Non-Bactericidal Cathelicidin Provides Prophylactic Efficacy against Bacterial Infection by Driving Phagocyte Influx. eLife 2022, 11, e72849. [Google Scholar] [CrossRef]
- Zhang, Y.; Cai, P.; Cheng, G.; Zhang, Y. A Brief Review of Phenolic Compounds Identified from Plants: Their Extraction, Analysis, and Biological Activity. Natural Product Communications 2022, 17, 1934578X211069721. [Google Scholar] [CrossRef]
- Wang, S.; Fan, L.; Pan, H.; Li, Y.; Zhao, X.; Qiu, Y.; Lu, Y. Identification and Characterization of a Novel Cathelicidin from Hydrophis Cyanocinctus with Antimicrobial and Anti-Inflammatory Activity. Molecules 2023, 28, 2082. [Google Scholar] [CrossRef]
- Shi, L.; Zhao, W.; Yang, Z.; Subbiah, V.; Suleria, H.A.R. Extraction and Characterization of Phenolic Compounds and Their Potential Antioxidant Activities. Environ Sci Pollut Res 2022, 29, 81112–81129. [Google Scholar] [CrossRef]
- Flavonoid. Wikipedia 2024.
- Harvard Health Publishing The Thinking on Flavonoids. Available online: https://www.health.harvard.edu/mind-and-mood/the-thinking-on-flavonoids (accessed on 10 August 2024).
- Kozłowska, A.; Szostak-Węgierek, D. Flavonoids – Food Sources, Health Benefits, and Mechanisms Involved. In Bioactive Molecules in Food; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, 2019; pp. 53–78. ISBN 978-3-319-78030-6. [Google Scholar]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as Natural Phenolic Compounds and Their Role in Therapeutics: An Overview. Future Journal of Pharmaceutical Sciences 2021, 7, 25. [Google Scholar] [CrossRef]
- Doyle, A.A.; Stephens, J.C. A Review of Cinnamaldehyde and Its Derivatives as Antibacterial Agents. Fitoterapia 2019, 139, 104405. [Google Scholar] [CrossRef]
- Xu, M.; Wu, P.; Shen, F.; Ji, J.; Rakesh, K.P. Chalcone Derivatives and Their Antibacterial Activities: Current Development. Bioorganic Chemistry 2019, 91, 103133. [Google Scholar] [CrossRef]
- Dan, W.; Dai, J. Recent Developments of Chalcones as Potential Antibacterial Agents in Medicinal Chemistry. European Journal of Medicinal Chemistry 2020, 187, 111980. [Google Scholar] [CrossRef]
- Skarpalezos, D.; Detsi, A. Deep Eutectic Solvents as Extraction Media for Valuable Flavonoids from Natural Sources. Applied Sciences 2019, 9, 4169. [Google Scholar] [CrossRef]
- Rodríguez De Luna, S.L.; Ramírez-Garza, R.E.; Serna Saldívar, S.O. Environmentally Friendly Methods for Flavonoid Extraction from Plant Material: Impact of Their Operating Conditions on Yield and Antioxidant Properties. The Scientific World Journal 2020, 2020, 6792069. [Google Scholar] [CrossRef]
- Chaves, J.O.; de Souza, M.C.; da Silva, L.C.; Lachos-Perez, D.; Torres-Mayanga, P.C.; Machado, A.P.d.F.; Forster-Carneiro, T.; Vázquez-Espinosa, M.; González-de-Peredo, A.V.; Barbero, G.F.; et al. Extraction of Flavonoids From Natural Sources Using Modern Techniques. Front. Chem. 2020, 8. [Google Scholar] [CrossRef]
- Jurinjak Tušek, A.; Šamec, D.; Šalić, A. Modern Techniques for Flavonoid Extraction—To Optimize or Not to Optimize? Applied Sciences 2022, 12, 11865. [Google Scholar] [CrossRef]
- Xu, J.; Wu, J.; Qi, J.; Li, J.; Liu, Y.; Miao, Z.; Qiu, G.; Jia, W. Microwave-Assisted Extraction of Flavonoids from Phyllostachys Heterocycla Leaves: Optimization, Mechanism, and Antioxidant Activity in Vitro. BioResources 2021, 16, 8060–8081. [Google Scholar] [CrossRef]
- Liu, X.; Liu, Y.; Shan, C.; Yang, X.; Zhang, Q.; Xu, N.; Xu, L.; Song, W. Effects of Five Extraction Methods on Total Content, Composition, and Stability of Flavonoids in Jujube. Food Chemistry: X 2022, 14, 100287. [Google Scholar] [CrossRef]
- Vittaya, L.; Charoendat, U.; Ui-eng, J.; Leesakul, N. Effect of Extraction Solvents on Phenolic Compounds and Flavonoids from Pongame Oiltree (Derris Indica [Lamk.] Bennet) Aerial Parts and Their Growth Inhibition of Aquatic Pathogenic Bacteria. Agriculture and Natural Resources 2022, 56, 569–582. [Google Scholar]
- Simangunsong, S.N.; Lenny, S.; Marpaung, L. Isolation of Flavonoids Compounds from Akalifa (Acalypha Wilkesiana Muell. Arc.) Plant Leaves. Journal of Chemical Natural Resources 2023, 5, 40–45. [Google Scholar] [CrossRef]
- Bribi, N. Pharmacological Activity of Alkaloids: A Review. 2018, 1.
- Lin, C.-J.; Chang, Y.-L.; Yang, Y.-L.; Chen, Y.-L. Natural Alkaloid Tryptanthrin Exhibits Novel Anticryptococcal Activity. Medical Mycology 2021, 59, 545–556. [Google Scholar] [CrossRef]
- Liu, M.; Han, J.; Feng, Y.; Guymer, G.; Forster, P.I.; Quinn, R.J. Antimicrobial Benzyltetrahydroisoquinoline-Derived Alkaloids from the Leaves of Doryphora Aromatica. J. Nat. Prod. 2021, 84, 676–682. [Google Scholar] [CrossRef]
- Biva, I.J.; Ndi, C.P.; Semple, S.J.; Griesser, H.J. Antibacterial Performance of Terpenoids from the Australian Plant Eremophila Lucida. Antibiotics 2019, 8, 63. [Google Scholar] [CrossRef]
- Zhu, C.-Z.; Hu, B.-Y.; Liu, J.-W.; Cai, Y.; Chen, X.-C.; Qin, D.-P.; Cheng, Y.-X.; Zhang, Z.-D. Anti-Mycobacterium Tuberculosis Terpenoids from Resina Commiphora. Molecules 2019, 24, 1475. [Google Scholar] [CrossRef]
- Hossain, S.; Urbi, Z.; Karuniawati, H.; Mohiuddin, R.B.; Moh Qrimida, A.; Allzrag, A.M.M.; Ming, L.C.; Pagano, E.; Capasso, R. Andrographis Paniculata (Burm. f.) Wall. Ex Nees: An Updated Review of Phytochemistry, Antimicrobial Pharmacology, and Clinical Safety and Efficacy. Life 2021, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Pech-Puch, D.; Forero, A.M.; Fuentes-Monteverde, J.C.; Lasarte-Monterrubio, C.; Martinez-Guitian, M.; González-Salas, C.; Guillén-Hernández, S.; Villegas-Hernández, H.; Beceiro, A.; Griesinger, C.; et al. Antimicrobial Diterpene Alkaloids from an Agelas Citrina Sponge Collected in the Yucatán Peninsula. Marine Drugs 2022, 20, 298. [Google Scholar] [CrossRef]
- Miyakoshi, M.; Tamura, Y.; Masuda, H.; Mizutani, K.; Tanaka, O.; Ikeda, T.; Ohtani, K.; Kasai, R.; Yamasaki, K. Antiyeast Steroidal Saponins from Yucca Schidigera (Mohave Yucca), a New Anti-Food-Deteriorating Agent. J. Nat. Prod. 2000, 63, 332–338. [Google Scholar] [CrossRef]
- Hussein, R.A.; El-Anssary, A.A.; Hussein, R.A.; El-Anssary, A.A. Plants. In Herbal Medicine; IntechOpen: location, 2018; ISBN 978-1-78984-783-3. [Google Scholar]
- Dev Kumar, G.; Mis Solval, K.; Mishra, A.; Macarisin, D. Antimicrobial Efficacy of Pelargonic Acid Micelles against Salmonella Varies by Surfactant, Serotype and Stress Response. Sci Rep 2020, 10, 10287. [Google Scholar] [CrossRef]
- Pacyga, K.; Pacyga, P.; Topola, E.; Viscardi, S.; Duda-Madej, A. Bioactive Compounds from Plant Origin as Natural Antimicrobial Agents for the Treatment of Wound Infections. International Journal of Molecular Sciences 2024, 25, 2100. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.S.; Grove, A. Do Global Regulators Hold the Key to Production of Bacterial Secondary Metabolites? Antibiotics 2019, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, N.; Nazir, R.; Khan, M.; Khaliq, A.; Adnan, M.; Ullah, M.; Yang, H. Antibacterial Activities, Phytochemical Screening and Metal Analysis of Medicinal Plants: Traditional Recipes Used against Diarrhea. Antibiotics 2019, 8, 194. [Google Scholar] [CrossRef]
- Asif, F.; Zaman, S.U.; Arnab, M.K.H.; Hasan, M.; Islam, M.M. Antimicrobial Peptides as Therapeutics: Confronting Delivery Challenges to Optimize Efficacy. The Microbe 2024, 2, 100051. [Google Scholar] [CrossRef]
- Koehbach, J.; Craik, D.J. The Vast Structural Diversity of Antimicrobial Peptides. Trends in Pharmacological Sciences 2019, 40, 517–528. [Google Scholar] [CrossRef]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. International Journal of Molecular Sciences 2021, 22, 11691. [Google Scholar] [CrossRef]
- Chung, C.-R.; Liou, J.-T.; Wu, L.-C.; Horng, J.-T.; Lee, T.-Y. Multi-Label Classification and Features Investigation of Antimicrobial Peptides with Various Functional Classes. iScience 2023, 26. [Google Scholar] [CrossRef]
- Sharma, H.; Dave, V.; Tyagi, E.; Prakash, A. Antimicrobial Peptides: A Novel and Natural Approach as Antibiofouling Mediator. Biologia 2024, 79, 2515–2533. [Google Scholar] [CrossRef]
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.-Y. Antimicrobial Peptides: New Hope in the War against Multidrug Resistance. zr 2019, 40, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Xuan, J.; Feng, W.; Wang, J.; Wang, R.; Zhang, B.; Bo, L.; Chen, Z.-S.; Yang, H.; Sun, L. Antimicrobial Peptides for Combating Drug-Resistant Bacterial Infections. Drug Resistance Updates 2023, 68, 100954. [Google Scholar] [CrossRef]
- Lai, S.; Zhang, Q.; Jin, L. Natural and Man-Made Cyclic Peptide-Based Antibiotics. Antibiotics 2023, 12, 42. [Google Scholar] [CrossRef]
- Bakare, O.O.; Gokul, A.; Niekerk, L.-A.; Aina, O.; Abiona, A.; Barker, A.M.; Basson, G.; Nkomo, M.; Otomo, L.; Keyster, M.; et al. Recent Progress in the Characterization, Synthesis, Delivery Procedures, Treatment Strategies, and Precision of Antimicrobial Peptides. International Journal of Molecular Sciences 2023, 24, 11864. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The Antimicrobial Peptides and Their Potential Clinical Applications. Am J Transl Res 2019, 11, 3919–3931. [Google Scholar]
- Chen, C.H.; Lu, T.K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef]
- van der Does, A.M.; Hiemstra, P.S.; Mookherjee, N. Antimicrobial Host Defence Peptides: Immunomodulatory Functions and Translational Prospects. In Antimicrobial Peptides: Basics for Clinical Application; Matsuzaki, K., Ed.; Springer: Singapore, 2019; pp. 149–171. ISBN 9789811335884. [Google Scholar]
- Raheem, N.; Straus, S.K. Mechanisms of Action for Antimicrobial Peptides With Antibacterial and Antibiofilm Functions. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial Peptides as Therapeutic Agents: Opportunities and Challenges. Critical Reviews in Biotechnology 2020, 40, 978–992. [Google Scholar] [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The Value of Antimicrobial Peptides in the Age of Resistance. The Lancet Infectious Diseases 2020, 20, e216–e230. [Google Scholar] [CrossRef] [PubMed]
- Uddin, S.J.; Shilpi, J.A.; Nahar, L.; Sarker, S.D.; Göransson, U. Editorial: Natural Antimicrobial Peptides: Hope for New Antibiotic Lead Molecules. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Phazang, P.; Negi, N.P.; Raina, M.; Kumar, D. Plant Antimicrobial Peptides: Next-Generation Bioactive Molecules for Plant Protection. In Phyto-Microbiome in Stress Regulation; Kumar, M., Kumar, V., Prasad, R., Eds.; Springer: Singapore, 2020; pp. 281–293. ISBN 9789811525766. [Google Scholar]
- Zhou, X.; Zhang, J.; Shen, J.; Cheng, B.; Bi, C.; Ma, Q. Branched-Chain Amino Acid Modulation of Lipid Metabolism, Gluconeogenesis, and Inflammation in a Finishing Pig Model: Targeting Leucine and Valine. Food Funct. 2023, 14, 10119–10134. [Google Scholar] [CrossRef]
- Zou, F.; Tan, C.; Shinali, T.S.; Zhang, B.; Zhang, L.; Han, Z.; Shang, N. Plant Antimicrobial Peptides: A Comprehensive Review of Their Classification, Production, Mode of Action, Functions, Applications, and Challenges. Food Funct. 2023, 14, 5492–5515. [Google Scholar] [CrossRef]
- Bakare, O.O.; Gokul, A.; Fadaka, A.O.; Wu, R.; Niekerk, L.-A.; Barker, A.M.; Keyster, M.; Klein, A. Plant Antimicrobial Peptides (PAMPs): Features, Applications, Production, Expression, and Challenges. Molecules 2022, 27, 3703. [Google Scholar] [CrossRef]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant Antimicrobial Peptides: Structures, Functions, and Applications. Botanical Studies 2021, 62, 5. [Google Scholar] [CrossRef] [PubMed]
- Barashkova, A.S.; Rogozhin, E.A. Isolation of Antimicrobial Peptides from Different Plant Sources: Does a General Extraction Method Exist? Plant Methods 2020, 16, 143. [Google Scholar] [CrossRef]
- Ben Brahim, R.; Ellouzi, H.; Fouzai, K.; Asses, N.; Neffati, M.; Sabatier, J.M.; Bulet, P.; Regaya, I. Optimized Chemical Extraction Methods of Antimicrobial Peptides from Roots and Leaves of Extremophilic Plants: Anthyllis Sericea and Astragalus Armatus Collected from the Tunisian Desert. Antibiotics 2022, 11, 1302. [Google Scholar] [CrossRef]
- Lima, A.M.; Azevedo, M.I.G.; Sousa, L.M.; Oliveira, N.S.; Andrade, C.R.; Freitas, C.D.T.; Souza, P.F.N. Plant Antimicrobial Peptides: An Overview about Classification, Toxicity and Clinical Applications. International Journal of Biological Macromolecules 2022, 214, 10–21. [Google Scholar] [CrossRef]
- Höng, K.; Austerlitz, T.; Bohlmann, T.; Bohlmann, H. The Thionin Family of Antimicrobial Peptides. PLOS ONE 2021, 16, e0254549. [Google Scholar] [CrossRef]
- Chen, K.; Yin, Y.; Liu, S.; Guo, Z.; Zhang, K.; Liang, Y.; Zhang, L.; Zhao, W.; Chao, H.; Li, M. Genome-Wide Identification and Functional Analysis of Oleosin Genes in Brassica Napus L. BMC Plant Biology 2019, 19, 294. [Google Scholar] [CrossRef]
- Rahman, M.; Guo, Q.; Baten, A.; Mauleon, R.; Khatun, A.; Liu, L.; Barkla, B.J. Shotgun Proteomics of Brassica Rapa Seed Proteins Identifies Vicilin as a Major Seed Storage Protein in the Mature Seed. PLOS ONE 2021, 16, e0253384. [Google Scholar] [CrossRef]
- Afroz, M.; Akter, S.; Ahmed, A.; Rouf, R.; Shilpi, J.A.; Tiralongo, E.; Sarker, S.D.; Göransson, U.; Uddin, S.J. Ethnobotany and Antimicrobial Peptides From Plants of the Solanaceae Family: An Update and Future Prospects. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Kar, A.; Biswas, S.; Chaudhary, S.K.; Banerjee, S. Chapter 37 - Hyphenated Analytical Techniques for Validation of Herbal Medicine. In Evidence-Based Validation of Herbal Medicine (Second Edition); Mukherjee, P.K., Ed.; Elsevier: location, 2022; pp. 811–827. ISBN 978-0-323-85542-6. [Google Scholar]
- Parthasarathy, A.; Borrego, E.J.; Savka, M.A.; Dobson, R.C.J.; Hudson, A.O. Amino Acid–Derived Defense Metabolites from Plants: A Potential Source to Facilitate Novel Antimicrobial Development. Journal of Biological Chemistry 2021, 296. [Google Scholar] [CrossRef] [PubMed]
- Slezina, M.P.; Odintsova, T.I. Plant Antimicrobial Peptides: Insights into Structure-Function Relationships for Practical Applications. Current Issues in Molecular Biology 2023, 45, 3674–3704. [Google Scholar] [CrossRef] [PubMed]
- Moyer, T.B.; Allen, J.L.; Shaw, L.N.; Hicks, L.M. Multiple Classes of Antimicrobial Peptides in Amaranthus Tricolor Revealed by Prediction, Proteomics, and Mass Spectrometric Characterization. J. Nat. Prod. 2021, 84, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, M.; Singh, A.; Kumar, S. PTPAMP: Prediction Tool for Plant-Derived Antimicrobial Peptides. Amino Acids 2023, 55, 1–17. [Google Scholar] [CrossRef]
- Nazarian-Firouzabadi, F.; Torres, M.D.T.; de la Fuente-Nunez, C. Recombinant Production of Antimicrobial Peptides in Plants. Biotechnology Advances 2024, 71, 108296. [Google Scholar] [CrossRef]
- Mojsoska, B.; Jenssen, H. Peptides and Peptidomimetics for Antimicrobial Drug Design. Pharmaceuticals 2015, 8, 366–415. [Google Scholar] [CrossRef]
- de Barros, E.; Gonçalves, R.M.; Cardoso, M.H.; Santos, N.C.; Franco, O.L.; Cândido, E.S. Snake Venom Cathelicidins as Natural Antimicrobial Peptides. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef]
- Shi, Y.; Li, C.; Wang, M.; Chen, Z.; Luo, Y.; Xia, X.; Song, Y.; Sun, Y.; Zhang, A.-M. Cathelicidin-DM Is an Antimicrobial Peptide from Duttaphrynus Melanostictus and Has Wound-Healing Therapeutic Potential. ACS Omega 2020, 5, 9301–9310. [Google Scholar] [CrossRef]
- Scheenstra, M.R.; van Harten, R.M.; Veldhuizen, E.J.A.; Haagsman, H.P.; Coorens, M. Cathelicidins Modulate TLR-Activation and Inflammation. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W.; Somsri, S.; Aunpad, R. A Novel, Rationally Designed, Hybrid Antimicrobial Peptide, Inspired by Cathelicidin and Aurein, Exhibits Membrane-Active Mechanisms against Pseudomonas Aeruginosa. Sci Rep 2020, 10, 9117. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Chen, R.; Zhang, J.; Chen, F.; Wang, K.-J. A Novel Antimicrobial Peptide Spampcin56–86 from Scylla Paramamosain Exerting Rapid Bactericidal and Anti-Biofilm Activity In Vitro and Anti-Infection In Vivo. International Journal of Molecular Sciences 2022, 23, 13316. [Google Scholar] [CrossRef]
- Galal-Khallaf, A.; Samir Aboali, E.; El-Sayed Hassab El-Nabi, S.; El-Tantawy, A.I.; Schott, E.J.; Mohammed-Geba, K. As Healthy as Invasive: Charybdis Natator Shell Extract Reveals Beneficial Metabolites with Promising Antioxidant and Anti-Inflammatory Potentials. Front. Mar. Sci. 2024, 11. [Google Scholar] [CrossRef]
- Lee, B.; Shin, M.K.; Yoo, J.S.; Jang, W.; Sung, J.-S. Identifying Novel Antimicrobial Peptides from Venom Gland of Spider Pardosa Astrigera by Deep Multi-Task Learning. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Elliott, A.G.; Huang, J.X.; Neve, S.; Zuegg, J.; Edwards, I.A.; Cain, A.K.; Boinett, C.J.; Barquist, L.; Lundberg, C.V.; Steen, J.; et al. An Amphipathic Peptide with Antibiotic Activity against Multidrug-Resistant Gram-Negative Bacteria. Nat Commun 2020, 11, 3184. [Google Scholar] [CrossRef]
- Fleming, A. On the Antibacterial Action of Cultures of a Penicillium, with Special Reference to Their Use in the Isolation of B. Influenzæ. Br J Exp Pathol 1929, 10, 226–236. [Google Scholar] [CrossRef]
- Tan, S.; Tatsumura, Y. Alexander Fleming (1881–1955): Discoverer of Penicillin. smedj 2015, 56, 366–367. [Google Scholar] [CrossRef]
- Pham, J.V.; Yilma, M.A.; Feliz, A.; Majid, M.T.; Maffetone, N.; Walker, J.R.; Kim, E.; Cho, H.J.; Reynolds, J.M.; Song, M.C.; et al. A Review of the Microbial Production of Bioactive Natural Products and Biologics. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Moradali, M.F.; Rehm, B.H.A. Bacterial Biopolymers: From Pathogenesis to Advanced Materials. Nat Rev Microbiol 2020, 18, 195–210. [Google Scholar] [CrossRef]
- Pérez-Ramos, A.; Madi-Moussa, D.; Coucheney, F.; Drider, D. Current Knowledge of the Mode of Action and Immunity Mechanisms of LAB-Bacteriocins. Microorganisms 2021, 9, 2107. [Google Scholar] [CrossRef] [PubMed]
- Romero-Calle, D.; Guimarães Benevides, R.; Góes-Neto, A.; Billington, C. Bacteriophages as Alternatives to Antibiotics in Clinical Care. Antibiotics 2019, 8, 138. [Google Scholar] [CrossRef]
- Plumet, L.; Ahmad-Mansour, N.; Dunyach-Remy, C.; Kissa, K.; Sotto, A.; Lavigne, J.-P.; Costechareyre, D.; Molle, V. Bacteriophage Therapy for Staphylococcus Aureus Infections: A Review of Animal Models, Treatments, and Clinical Trials. Front. Cell. Infect. Microbiol. 2022, 12. [Google Scholar] [CrossRef]
- Barron, M. Phage Therapy: Past, Present and Future. Available online: https://asm.org:443/Articles/2022/August/Phage-Therapy-Past,-Present-and-Future (accessed on 10 August 2024).
- Parkinson, J. What’s Old Is New: Bacteriophage Is a Therapy That May Combat Antimicrobial Resistance. Available online: https://www.contagionlive.com/view/what-s-old-is-new-bacteriophage-is-a-therapy-that-may-combat-antimicrobial-resistance (accessed on 10 August 2024).
- Ji, Y.; Cheng, M.; Zhai, S.; Xi, H.; Cai, R.; Wang, Z.; Zhang, H.; Wang, X.; Xue, Y.; Li, X.; et al. Preventive Effect of the Phage VB-SavM-JYL01 on Rabbit Necrotizing Pneumonia Caused by Staphylococcus Aureus. Veterinary Microbiology 2019, 229, 72–80. [Google Scholar] [CrossRef]
- Barylski, J.; Enault, F.; Dutilh, B.E.; Schuller, M.B.; Edwards, R.A.; Gillis, A.; Klumpp, J.; Knezevic, P.; Krupovic, M.; Kuhn, J.H.; et al. Analysis of Spounaviruses as a Case Study for the Overdue Reclassification of Tailed Phages. Systematic Biology 2020, 69, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Głowacka-Rutkowska, A.; Ulatowska, M.; Empel, J.; Kowalczyk, M.; Boreczek, J.; Łobocka, M. A Kayvirus Distant Homolog of Staphylococcal Virulence Determinants and VISA Biomarker Is a Phage Lytic Enzyme. Viruses 2020, 12, 292. [Google Scholar] [CrossRef]
- Göller, P.C.; Elsener, T.; Lorgé, D.; Radulovic, N.; Bernardi, V.; Naumann, A.; Amri, N.; Khatchatourova, E.; Coutinho, F.H.; Loessner, M.J.; et al. Multi-Species Host Range of Staphylococcal Phages Isolated from Wastewater. Nat Commun 2021, 12, 6965. [Google Scholar] [CrossRef] [PubMed]
- Łubowska, N.; Grygorcewicz, B.; Kosznik-Kwaśnicka, K.; Zauszkiewicz-Pawlak, A.; Węgrzyn, A.; Dołęgowska, B.; Piechowicz, L. Characterization of the Three New Kayviruses and Their Lytic Activity Against Multidrug-Resistant Staphylococcus Aureus. Microorganisms 2019, 7, 471. [Google Scholar] [CrossRef]
- Oduor, J.M.O.; Kadija, E.; Nyachieo, A.; Mureithi, M.W.; Skurnik, M. Bioprospecting Staphylococcus Phages with Therapeutic and Bio-Control Potential. Viruses 2020, 12, 133. [Google Scholar] [CrossRef]
- Kifelew, L.G.; Warner, M.S.; Morales, S.; Vaughan, L.; Woodman, R.; Fitridge, R.; Mitchell, J.G.; Speck, P. Efficacy of Phage Cocktail AB-SA01 Therapy in Diabetic Mouse Wound Infections Caused by Multidrug-Resistant Staphylococcus Aureus. BMC Microbiology 2020, 20, 204. [Google Scholar] [CrossRef]
- Fanaei Pirlar, R.; Wagemans, J.; Ponce Benavente, L.; Lavigne, R.; Trampuz, A.; Gonzalez Moreno, M. Novel Bacteriophage Specific against Staphylococcus Epidermidis and with Antibiofilm Activity. Viruses 2022, 14, 1340. [Google Scholar] [CrossRef] [PubMed]
- Glonti, T.; Pirnay, J.-P. In Vitro Techniques and Measurements of Phage Characteristics That Are Important for Phage Therapy Success. Viruses 2022, 14, 1490. [Google Scholar] [CrossRef]
- Kaźmierczak, N.; Grygorcewicz, B.; Roszak, M.; Bochentyn, B.; Piechowicz, L. Comparative Assessment of Bacteriophage and Antibiotic Activity against Multidrug-Resistant Staphylococcus Aureus Biofilms. International Journal of Molecular Sciences 2022, 23, 1274. [Google Scholar] [CrossRef] [PubMed]
- Podlesek, Z.; Žgur Bertok, D. The DNA Damage Inducible SOS Response Is a Key Player in the Generation of Bacterial Persister Cells and Population Wide Tolerance. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Alsaadi, S.E.; Lu, H.; Zhang, M.; Dykes, G.F.; Allison, H.E.; Horsburgh, M.J. Bacteriophages from Human Skin Infecting Coagulase-Negative Staphylococcus: Diversity, Novel Species and Host Resistance 2023, 2023.11.07.565964.
- Plumet, L.; Morsli, M.; Ahmad-Mansour, N.; Clavijo-Coppens, F.; Berry, L.; Sotto, A.; Lavigne, J.-P.; Costechareyre, D.; Molle, V. Isolation and Characterization of New Bacteriophages against Staphylococcal Clinical Isolates from Diabetic Foot Ulcers. Viruses 2023, 15, 2287. [Google Scholar] [CrossRef]
- Plotka, M.; Kapusta, M.; Dorawa, S.; Kaczorowska, A.-K.; Kaczorowski, T. Ts2631 Endolysin from the Extremophilic Thermus Scotoductus Bacteriophage vB_Tsc2631 as an Antimicrobial Agent against Gram-Negative Multidrug-Resistant Bacteria. Viruses 2019, 11, 657. [Google Scholar] [CrossRef]
- Kwon, J.; Kim, S.W.; Kim, S.G.; Kang, J.W.; Jung, W.J.; Lee, S.B.; Lee, Y.M.; Giri, S.S.; Chi, C.; Park, S.C. The Characterization of a Novel Phage, pPa_SNUABM_DT01, Infecting Pseudomonas Aeruginosa. Microorganisms 2021, 9, 2040. [Google Scholar] [CrossRef]
- Rice, C. Novel Bacteriophages and Their Derived Proteins for the Biocontrol of Proteus and Pseudomonas Biofilms. Doctoral Thesis, Queen’s University Belfast, Belfast, 2022. [Google Scholar]
- Hylling, O.; Carstens, A.B.; Kot, W.; Hansen, M.; Neve, H.; Franz, C.M.A.P.; Johansen, A.; Ellegaard-Jensen, L.; Hansen, L.H. Two Novel Bacteriophage Genera from a Groundwater Reservoir Highlight Subsurface Environments as Underexplored Biotopes in Bacteriophage Ecology. Sci Rep 2020, 10, 11879. [Google Scholar] [CrossRef]
- Sattar, S.; Bailie, M.; Yaqoob, A.; Khanum, S.; Fatima, K.; Altaf, A.U.R.B.; Ahmed, I.; Shah, S.T.A.; Munawar, J.; Zehra, Q.A.; et al. Characterization of Two Novel Lytic Bacteriophages Having Lysis Potential against MDR Avian Pathogenic Escherichia Coli Strains of Zoonotic Potential. Sci Rep 2023, 13, 10043. [Google Scholar] [CrossRef]
- Nicolas, M.; Trotereau, A.; Culot, A.; Moodley, A.; Atterbury, R.; Wagemans, J.; Lavigne, R.; Velge, P.; Schouler, C. Isolation and Characterization of a Novel Phage Collection against Avian-Pathogenic Escherichia Coli. Microbiology Spectrum 2023, 11, e04296-22. [Google Scholar] [CrossRef]
- Nale, J.Y.; Chan, B.; Nnadi, N.E.; Cheng, J.K.J.; Matts, S.; Nezam-Abadi, N.; Turkington, C.J.R.; Charreton, L.M.; Bola, H.; Nazir, R.; et al. Novel Escherichia Coli-Infecting Bacteriophages Isolated from Uganda That Target Human Clinical Isolates. PHAGE 2023, 4, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Parra, B.; Cockx, B.; Lutz, V.T.; Brøndsted, L.; Smets, B.F.; Dechesne, A. Isolation and Characterization of Novel Plasmid-Dependent Phages Infecting Bacteria Carrying Diverse Conjugative Plasmids. Microbiology Spectrum 2023, 12, e02537-23. [Google Scholar] [CrossRef] [PubMed]
- Bhandare, S.; Lawal, O.U.; Colavecchio, A.; Cadieux, B.; Zahirovich-Jovich, Y.; Zhong, Z.; Tompkins, E.; Amitrano, M.; Kukavica-Ibrulj, I.; Boyle, B.; et al. Genomic and Phenotypic Analysis of Salmonella Enterica Bacteriophages Identifies Two Novel Phage Species. Microorganisms 2024, 12, 695. [Google Scholar] [CrossRef] [PubMed]
- Nakonieczna, A.; Rutyna, P.; Fedorowicz, M.; Kwiatek, M.; Mizak, L.; Łobocka, M. Three Novel Bacteriophages, J5a, F16Ba, and Z1a, Specific for Bacillus Anthracis, Define a New Clade of Historical Wbeta Phage Relatives. Viruses 2022, 14, 213. [Google Scholar] [CrossRef] [PubMed]
- Kabwe, M.; Brown, T.; Speirs, L.; Ku, H.; Leach, M.; Chan, H.T.; Petrovski, S.; Lock, P.; Tucci, J. Novel Bacteriophages Capable of Disrupting Biofilms From Clinical Strains of Aeromonas Hydrophila. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Kallies, R.; Hu, D.; Abdulkadir, N.; Schloter, M.; Rocha, U. Identification of Huge Phages from Wastewater Metagenomes. Viruses 2023, 15, 2330. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, Synthesis, Mechanism of Action and Resistance Development in Food Spoilage Causing Bacteria. Microbial Pathogenesis 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, Antimicrobial Peptides from Bacterial Origin: Overview of Their Biology and Their Impact against Multidrug-Resistant Bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, Potent Antimicrobial Peptides and the Fight against Multi Drug Resistant Species: Resistance Is Futile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Yount, N.Y.; Weaver, D.C.; de Anda, J.; Lee, E.Y.; Lee, M.W.; Wong, G.C.L.; Yeaman, M.R. Discovery of Novel Type II Bacteriocins Using a New High-Dimensional Bioinformatic Algorithm. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Aljohani, A.B.; Al-Hejin, A.M.; Shori, A.B. Bacteriocins as Promising Antimicrobial Peptides, Definition, Classification, and Their Potential Applications in Cheeses. Food Sci. Technol 2023, 43, e118021. [Google Scholar] [CrossRef]
- Lei, S.; Zhao, R.; Sun, J.; Ran, J.; Ruan, X.; Zhu, Y. Partial Purification and Characterization of a Broad-Spectrum Bacteriocin Produced by a Lactobacillus Plantarum Zrx03 Isolated from Infant’s Feces. Food Science & Nutrition 2020, 8, 2214–2222. [Google Scholar] [CrossRef]
- Golneshin, A.; Gor, M.-C.; Williamson, N.; Vezina, B.; Van, T.T.H.; May, B.K.; Smith, A.T. Discovery and Characterisation of Circular Bacteriocin Plantacyclin B21AG from Lactiplantibacillus Plantarum B21. Heliyon 2020, 6, e04715. [Google Scholar] [CrossRef]
- Pei, J.; Jin, W.; Abd El-Aty, A.M.; Baranenko, D.A.; Gou, X.; Zhang, H.; Geng, J.; Jiang, L.; Chen, D.; Yue, T. Isolation, Purification, and Structural Identification of a New Bacteriocin Made by Lactobacillus Plantarum Found in Conventional Kombucha. Food Control 2020, 110, 106923. [Google Scholar] [CrossRef]
- Jiang, Y.-H.; Xin, W.-G.; Yang, L.-Y.; Ying, J.-P.; Zhao, Z.-S.; Lin, L.-B.; Li, X.-Z.; Zhang, Q.-L. A Novel Bacteriocin against Staphylococcus Aureus from Lactobacillus Paracasei Isolated from Yunnan Traditional Fermented Yogurt: Purification, Antibacterial Characterization, and Antibiofilm Activity. Journal of Dairy Science 2022, 105, 2094–2107. [Google Scholar] [CrossRef]
- Thuy, T.T.D.; Lu, H.-F.; Bregente, C.J.B.; Huang, F.-C.A.; Tu, P.-C.; Kao, C.-Y. Characterization of the Broad-Spectrum Antibacterial Activity of Bacteriocin-like Inhibitory Substance-Producing Probiotics Isolated from Fermented Foods. BMC Microbiology 2024, 24, 85. [Google Scholar] [CrossRef]
- Xing, Y.; Li, W.; Aweya, J.J.; Jin, R.; Lin, R.; Liang, D.; Weng, W.; Yang, S. Lacticaseibacillus Paracasei-Derived Antibacterial Peptide NGJ1D and Its Mechanism of Action Against Staphylococcus Aureus. Food Bioprocess Technol 2024. [Google Scholar] [CrossRef]
- Elnar, A.G.; Kim, G.-B. In Vitro and In Silico Characterization of N-Formylated Two-Peptide Bacteriocin from Enterococcus Faecalis CAUM157 with Anti-Listeria Activity. Probiotics & Antimicro. Prot. 2024. [Google Scholar] [CrossRef]
- Cui, G.; Pan, C.; Xu, P.; Li, Y.; Wang, L.; Gong, B.; Li, X.; Huang, S. Purification and Characterization of a Novel Bacteriocin Produced by Enterococcus Faecalis CG-9 from Human Saliva. Biotechnology & Biotechnological Equipment 2020, 34, 1224–1233. [Google Scholar] [CrossRef]
- Sharma, P.; Kaur, S.; Chadha, B.S.; Kaur, R.; Kaur, M.; Kaur, S. Anticancer and Antimicrobial Potential of Enterocin 12a from Enterococcus Faecium. BMC Microbiology 2021, 21, 39. [Google Scholar] [CrossRef]
- Newstead, L.L.; Varjonen, K.; Nuttall, T.; Paterson, G.K. Staphylococcal-Produced Bacteriocins and Antimicrobial Peptides: Their Potential as Alternative Treatments for Staphylococcus Aureus Infections. Antibiotics 2020, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, K.V.; Kranjec, C.; Thorstensen, T.; Carlsen, H.; Diep, D.B. Successful Development of Bacteriocins into Therapeutic Formulation for Treatment of MRSA Skin Infection in a Murine Model. Antimicrobial Agents and Chemotherapy 2020, 64, 10–1128. [Google Scholar] [CrossRef]
- Sriragavi, G.; Sangeetha, M.; Santhakumar, M.; Lokesh, E.; Nithyalakshmi, M.; Saleel, C.A.; Balagurunathan, R. Exploring Antibacterial Properties of Bioactive Compounds Isolated from Streptomyces Sp. in Bamboo Rhizosphere Soil. ACS Omega 2023, 8, 36333–36343. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, J.; Kamau, P.M.; Thuku, R.C.; Lai, R. Design Methods for Antimicrobial Peptides with Improved Performance. zr 2023, 44, 1095–1114. [Google Scholar] [CrossRef]
- Verma, S.; Pathak, R.K. Chapter 16 - Discovery and Optimization of Lead Molecules in Drug Designing. In Bioinformatics; Singh, D.B., Pathak, R.K., Eds.; Academic Press: location, 2022; ISBN 978-0-323-89775-4. [Google Scholar]
- Luong, H.X.; Thanh, T.T.; Tran, T.H. Antimicrobial Peptides – Advances in Development of Therapeutic Applications. Life Sciences 2020, 260, 118407. [Google Scholar] [CrossRef]
- Teixeira, M.C.; Carbone, C.; Sousa, M.C.; Espina, M.; Garcia, M.L.; Sanchez-Lopez, E.; Souto, E.B. Nanomedicines for the Delivery of Antimicrobial Peptides (AMPs). Nanomaterials 2020, 10, 560. [Google Scholar] [CrossRef]
- Sowers, A.; Wang, G.; Xing, M.; Li, B. Advances in Antimicrobial Peptide Discovery via Machine Learning and Delivery via Nanotechnology. Microorganisms 2023, 11, 1129. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, M.K.; Bhatt, N. Bioavailability Enhancement of Polymyxin B With Novel Drug Delivery: Development and Optimization Using Quality-by-Design Approach. JPharmSci 2019, 108, 1521–1528. [Google Scholar] [CrossRef]
- Gan, B.H.; Gaynord, J.; Rowe, S.M.; Deingruber, T.; Spring, D.R. The Multifaceted Nature of Antimicrobial Peptides: Current Synthetic Chemistry Approaches and Future Directions. Chem. Soc. Rev. 2021, 50, 7820–7880. [Google Scholar] [CrossRef]
- Zhou, M.; Qian, Y.; Xie, J.; Zhang, W.; Jiang, W.; Xiao, X.; Chen, S.; Dai, C.; Cong, Z.; Ji, Z.; et al. Poly(2-Oxazoline)-Based Functional Peptide Mimics: Eradicating MRSA Infections and Persisters While Alleviating Antimicrobial Resistance. Angewandte Chemie International Edition 2020, 59, 6412–6419. [Google Scholar] [CrossRef]
- Ouertani, A.; Mosbah, A.; Cherif, A.; Ouertani, A.; Mosbah, A.; Cherif, A. Anti-Microbial Peptides: The Importance of Structure-Function Analysis in the Design of New AMPs. In Insights on Antimicrobial Peptides; IntechOpen: location, 2022; ISBN 978-1-83969-714-2. [Google Scholar]
- Ajish, C.; Kumar, S.D.; Kim, E.Y.; Yang, S.; Shin, S.Y. A Short Novel Antimicrobial Peptide BP100-W with Antimicrobial, Antibiofilm and Anti-Inflammatory Activities Designed by Replacement with Tryptophan. Journal of Analytical Science and Technology 2022, 13, 46. [Google Scholar] [CrossRef]
- Wiman, E.; Zattarin, E.; Aili, D.; Bengtsson, T.; Selegård, R.; Khalaf, H. Development of Novel Broad-Spectrum Antimicrobial Lipopeptides Derived from Plantaricin NC8 β. Sci Rep 2023, 13, 4104. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.; Ali, Z.; Tehseen, M.; Haney, E.F.; Pantoja-Angles, A.; Alshehri, S.; Wang, T.; Clancy, G.J.; Ayach, M.; Hauser, C.; et al. Efficient in Planta Production of Amidated Antimicrobial Peptides That Are Active against Drug-Resistant ESKAPE Pathogens. Nat Commun 2023, 14, 1464. [Google Scholar] [CrossRef]
- Ramazi, S.; Mohammadi, N.; Allahverdi, A.; Khalili, E.; Abdolmaleki, P. A Review on Antimicrobial Peptides Databases and the Computational Tools. Database 2022, 2022, baac011. [Google Scholar] [CrossRef]
- Cardoso, M.H.; Orozco, R.Q.; Rezende, S.B.; Rodrigues, G.; Oshiro, K.G.N.; Cândido, E.S.; Franco, O.L. Computer-Aided Design of Antimicrobial Peptides: Are We Generating Effective Drug Candidates? Front. Microbiol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Boone, K.; Wisdom, C.; Camarda, K.; Spencer, P.; Tamerler, C. Combining Genetic Algorithm with Machine Learning Strategies for Designing Potent Antimicrobial Peptides. BMC Bioinformatics 2021, 22, 239. [Google Scholar] [CrossRef]
- Rumancev, C.; Rosenhahn, A.; Hilpert, K. BioSAXS–an Emerging Method to Accelerate, Enrich and de-Risk Antimicrobial Drug Development. Front. Pharmacol. 2022, 13. [Google Scholar] [CrossRef]
- Jianxun; Yushan, B.Y.; Yinxue; Yisuo; Boxing; Wei; Huaxi. De Novo Design and Antibacterial Activity of α-Helical Antimicrobial Peptide YHX-1. 食品科学技术学报 2022, 40, 98–104. [Google Scholar] [CrossRef]
- Mustafa, G.; Mehmood, R.; Mahrosh, H.S.; Mehmood, K.; Ahmed, S. Investigation of Plant Antimicrobial Peptides against Selected Pathogenic Bacterial Species Using a Peptide-Protein Docking Approach. BioMed Research International 2022, 2022, 1077814. [Google Scholar] [CrossRef]
- Lin, T.-T.; Yang, L.-Y.; Lin, C.-Y.; Wang, C.-T.; Lai, C.-W.; Ko, C.-F.; Shih, Y.-H.; Chen, S.-H. Intelligent De Novo Design of Novel Antimicrobial Peptides against Antibiotic-Resistant Bacteria Strains. International Journal of Molecular Sciences 2023, 24, 6788. [Google Scholar] [CrossRef]
- Szymczak, P.; Możejko, M.; Grzegorzek, T.; Jurczak, R.; Bauer, M.; Neubauer, D.; Sikora, K.; Michalski, M.; Sroka, J.; Setny, P.; et al. Discovering Highly Potent Antimicrobial Peptides with Deep Generative Model HydrAMP. Nat Commun 2023, 14, 1453. [Google Scholar] [CrossRef] [PubMed]
- Nedyalkova, M.; Paluch, A.S.; Vecini, D.P.; Lattuada, M. Progress and Future of the Computational Design of Antimicrobial Peptides (AMPs): Bio-Inspired Functional Molecules. Digital Discovery 2024, 3, 9–22. [Google Scholar] [CrossRef]
- Aguilera-Puga, M. d. C.; Cancelarich, N.L.; Marani, M.M.; de la Fuente-Nunez, C.; Plisson, F. Accelerating the Discovery and Design of Antimicrobial Peptides with Artificial Intelligence. In Computational Drug Discovery and Design; Gore, M., Jagtap, U.B., Eds.; Springer US: New York, NY, 2024; pp. 329–352. ISBN 978-1-07-163441-7. [Google Scholar]
- Ogunsona, E.O.; Muthuraj, R.; Ojogbo, E.; Valerio, O.; Mekonnen, T.H. Engineered Nanomaterials for Antimicrobial Applications: A Review. Applied Materials Today 2020, 18, 100473. [Google Scholar] [CrossRef]
- Díez-Pascual, A.M. Recent Progress in Antimicrobial Nanomaterials. Nanomaterials 2020, 10, 2315. [Google Scholar] [CrossRef] [PubMed]
- Alavi, M.; Rai, M. Recent Advances in Antibacterial Applications of Metal Nanoparticles (MNPs) and Metal Nanocomposites (MNCs) against Multidrug-Resistant (MDR) Bacteria. Expert Review of Anti-infective Therapy 2019, 17, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Karnwal, A.; Kumar, G.; Pant, G.; Hossain, K.; Ahmad, A.; Alshammari, M.B. Perspectives on Usage of Functional Nanomaterials in Antimicrobial Therapy for Antibiotic-Resistant Bacterial Infections. ACS Omega 2023, 8, 13492–13508. [Google Scholar] [CrossRef]
- Shaikh, S.; Nazam, N.; Rizvi, S.M.D.; Ahmad, K.; Baig, M.H.; Lee, E.J.; Choi, I. Mechanistic Insights into the Antimicrobial Actions of Metallic Nanoparticles and Their Implications for Multidrug Resistance. International Journal of Molecular Sciences 2019, 20, 2468. [Google Scholar] [CrossRef]
- Singh, A.; Gautam, P.K.; Verma, A.; Singh, V.; Shivapriya, P.M.; Shivalkar, S.; Sahoo, A.K.; Samanta, S.K. Green Synthesis of Metallic Nanoparticles as Effective Alternatives to Treat Antibiotics Resistant Bacterial Infections: A Review. Biotechnology Reports 2020, 25, e00427. [Google Scholar] [CrossRef]
- Ortega-Nieto, C.; Losada-Garcia, N.; Prodan, D.; Furtos, G.; Palomo, J.M. Recent Advances on the Design and Applications of Antimicrobial Nanomaterials. Nanomaterials 2023, 13, 2406. [Google Scholar] [CrossRef]
- Vaghela, H.M.; Pathan, A.; SHAH, R. Biogenic Synthesis of Au, Pd and Pt Metal: Nanoparticles Using Various Plants and Its Medicinal Applications: A Review; ISBN 978-3-330-01803-7.
- Kushwah, K.S.; Verma, D.K.; Kushwah, K.S.; Verma, D.K. Biological Synthesis of Metallic Nanoparticles from Different Plant Species. In 21st Century Nanostructured Materials - Physics, Chemistry, Classification, and Emerging Applications in Industry, Biomedicine, and Agriculture; IntechOpen: location, 2021; ISBN 978-1-80355-085-5. [Google Scholar]
- Rizki, I.N.; Klaypradit, W. Patmawati Utilization of Marine Organisms for the Green Synthesis of Silver and Gold Nanoparticles and Their Applications: A Review. Sustainable Chemistry and Pharmacy 2023, 31, 100888. [Google Scholar] [CrossRef]
- Bhardwaj, K.; Chopra, C.; Bhardwaj, P.; Dhanjal, D.S.; Singh, R.; Najda, A.; Cruz-Martins, N.; Singh, S.; Sharma, R.; Kuča, K.; et al. Biogenic Metallic Nanoparticles from Seed Extracts: Characteristics, Properties, and Applications. Journal of Nanomaterials 2022, 2022, 2271278. [Google Scholar] [CrossRef]
- Chauhan, A.; Anand, J.; Parkash, V.; Rai, N. Biogenic Synthesis: A Sustainable Approach for Nanoparticles Synthesis Mediated by Fungi. Inorganic and Nano-Metal Chemistry 2023, 53, 460–473. [Google Scholar] [CrossRef]
- Nandhini, S.N.; Sisubalan, N.; Vijayan, A.; Karthikeyan, C.; Gnanaraj, M.; Gideon, D.A.M.; Jebastin, T.; Varaprasad, K.; Sadiku, R. Recent Advances in Green Synthesized Nanoparticles for Bactericidal and Wound Healing Applications. Heliyon 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Mayegowda, S.B.; Sarma, G.; Gadilingappa, M.N.; Alghamdi, S.; Aslam, A.; Refaat, B.; Almehmadi, M.; Allahyani, M.; Alsaiari, A.A.; Aljuaid, A.; et al. Green-Synthesized Nanoparticles and Their Therapeutic Applications: A Review. Green Processing and Synthesis 2023, 12. [Google Scholar] [CrossRef]
- Bhati, M. Biogenic Synthesis of Metallic Nanoparticles: Principles and Applications. Materials Today: Proceedings 2023, 81, 882–887. [Google Scholar] [CrossRef]
- Jayasinghe, M.K.; Lee, C.Y.; Tran, T.T.T.; Tan, R.; Chew, S.M.; Yeo, B.Z.J.; Loh, W.X.; Pirisinu, M.; Le, M.T.N. The Role of in Silico Research in Developing Nanoparticle-Based Therapeutics. Front. Digit. Health 2022, 4. [Google Scholar] [CrossRef] [PubMed]
- Kailasa, S.K.; Park, T.-J.; Rohit, J.V.; Koduru, J.R. Chapter 14 - Antimicrobial Activity of Silver Nanoparticles. In Nanoparticles in Pharmacotherapy; Grumezescu, A.M., Ed.; William Andrew Publishing: location, 2019; ISBN 978-0-12-816504-1. [Google Scholar]
- Das, G.; Patra, J.K.; Debnath, T.; Ansari, A.; Shin, H.-S. Investigation of Antioxidant, Antibacterial, Antidiabetic, and Cytotoxicity Potential of Silver Nanoparticles Synthesized Using the Outer Peel Extract of Ananas Comosus (L.). PLOS ONE 2019, 14, e0220950. [Google Scholar] [CrossRef]
- Paladini, F.; Pollini, M. Antimicrobial Silver Nanoparticles for Wound Healing Application: Progress and Future Trends. Materials 2019, 12, 2540. [Google Scholar] [CrossRef]
- Anees Ahmad, S.; Sachi Das, S.; Khatoon, A.; Tahir Ansari, M.; Afzal, M.; Saquib Hasnain, M.; Kumar Nayak, A. Bactericidal Activity of Silver Nanoparticles: A Mechanistic Review. Materials Science for Energy Technologies 2020, 3, 756–769. [Google Scholar] [CrossRef]
- Hamad, A.; Khashan, K.S.; Hadi, A. Silver Nanoparticles and Silver Ions as Potential Antibacterial Agents. J Inorg Organomet Polym 2020, 30, 4811–4828. [Google Scholar] [CrossRef]
- Bruna, T.; Maldonado-Bravo, F.; Jara, P.; Caro, N. Silver Nanoparticles and Their Antibacterial Applications. International Journal of Molecular Sciences 2021, 22, 7202. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Briffa, S.M.; Swingler, S.; Gibson, H.; Kannappan, V.; Adamus, G.; Kowalczuk, M.; Martin, C.; Radecka, I. Synthesis of Silver Nanoparticles Using Curcumin-Cyclodextrins Loaded into Bacterial Cellulose-Based Hydrogels for Wound Dressing Applications. Biomacromolecules 2020, 21, 1802–1811. [Google Scholar] [CrossRef] [PubMed]
- Das, C.G.A.; Kumar, V.G.; Dhas, T.S.; Karthick, V.; Govindaraju, K.; Joselin, J.M.; Baalamurugan, J. Antibacterial Activity of Silver Nanoparticles (Biosynthesis): A Short Review on Recent Advances. Biocatalysis and Agricultural Biotechnology 2020, 27, 101593. [Google Scholar] [CrossRef]
- Feroze, N.; Arshad, B.; Younas, M.; Afridi, M.I.; Saqib, S.; Ayaz, A. Fungal Mediated Synthesis of Silver Nanoparticles and Evaluation of Antibacterial Activity. Microscopy Research and Technique 2020, 83, 72–80. [Google Scholar] [CrossRef]
- Garibo, D.; Borbón-Nuñez, H.A.; de León, J.N.D.; García Mendoza, E.; Estrada, I.; Toledano-Magaña, Y.; Tiznado, H.; Ovalle-Marroquin, M.; Soto-Ramos, A.G.; Blanco, A.; et al. Green Synthesis of Silver Nanoparticles Using Lysiloma Acapulcensis Exhibit High-Antimicrobial Activity. Sci Rep 2020, 10, 12805. [Google Scholar] [CrossRef]
- Rahimi, M.; Noruzi, E.B.; Sheykhsaran, E.; Ebadi, B.; Kariminezhad, Z.; Molaparast, M.; Mehrabani, M.G.; Mehramouz, B.; Yousefi, M.; Ahmadi, R.; et al. Carbohydrate Polymer-Based Silver Nanocomposites: Recent Progress in the Antimicrobial Wound Dressings. Carbohydrate Polymers 2020, 231, 115696. [Google Scholar] [CrossRef]
- Barabadi, H.; Mojab, F.; Vahidi, H.; Marashi, B.; Talank, N.; Hosseini, O.; Saravanan, M. Green Synthesis, Characterization, Antibacterial and Biofilm Inhibitory Activity of Silver Nanoparticles Compared to Commercial Silver Nanoparticles. Inorganic Chemistry Communications 2021, 129, 108647. [Google Scholar] [CrossRef]
- Ramírez-Rosas, S.L.; Delgado-Alvarado, E.; Sanchez-Vargas, L.O.; Herrera-May, A.L.; Peña-Juarez, M.G.; Gonzalez-Calderon, J.A. Green Route to Produce Silver Nanoparticles Using the Bioactive Flavonoid Quercetin as a Reducing Agent and Food Anti-Caking Agents as Stabilizers. Nanomaterials 2022, 12, 3545. [Google Scholar] [CrossRef]
- Chavan, R.R.; Bhutkar, M.A.; Bhinge, S.D. Design, Synthesis, and Optimization of Silver Nanoparticles Using an Artocarpus Heterophyllus Lam. Leaf Extract and Its Antibacterial Application. Nano Biomedicine and Engineering 2023, 15, 239–252. [Google Scholar] [CrossRef]
- Mahalingam, S.; Govindaraji, P.K.; Solomon, V.G.; Kesavan, H.; Neelan, Y.D.; Bakthavatchalam, S.; Kim, J.; Bakthavatchalam, P. Biogenic Synthesis and Characterization of Silver Nanoparticles: Evaluation of Their Larvicidal, Antibacterial, and Cytotoxic Activities. ACS Omega 2023, 8, 11923–11930. [Google Scholar] [CrossRef] [PubMed]
- Said, A.; Abu-Elghait, M.; Atta, H.M.; Salem, S.S. Antibacterial Activity of Green Synthesized Silver Nanoparticles Using Lawsonia Inermis Against Common Pathogens from Urinary Tract Infection. Appl Biochem Biotechnol 2024, 196, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Hani, U.; Kidwan, F.N.; Albarqi, L.A.; Al-qahtani, S.A.; AlHadi, R.M.; AlZaid, H.A.; Haider, N.; Ansari, M.A. Biogenic Silver Nanoparticle Synthesis Using Orange Peel Extract and Its Multifaceted Biomedical Application. Bioprocess Biosyst Eng 2024, 47, 1363–1375. [Google Scholar] [CrossRef]
- El-Zahed, M.M.; Abou-Dobara, M.I.; El-Sayed, A.K.A.; Baka, Z.A.M. Ag/SiO2 Nanocomposite Mediated by Escherichia Coli D8 and Their Antimicrobial Potential. Nova Biotechnologica et Chimica 2022, 21, e1023–e1023. [Google Scholar] [CrossRef]
- Naseer, M.; Aslam, U.; Khalid, B.; Chen, B. Green Route to Synthesize Zinc Oxide Nanoparticles Using Leaf Extracts of Cassia Fistula and Melia Azadarach and Their Antibacterial Potential. Sci Rep 2020, 10, 9055. [Google Scholar] [CrossRef]
- Gur, T.; Meydan, I.; Seckin, H.; Bekmezci, M.; Sen, F. Green Synthesis, Characterization and Bioactivity of Biogenic Zinc Oxide Nanoparticles. Environmental Research 2022, 204, 111897. [Google Scholar] [CrossRef]
- Veselova, V.O.; Plyuta, V.A.; Kostrov, A.N.; Vtyurina, D.N.; Abramov, V.O.; Abramova, A.V.; Voitov, Y.I.; Padiy, D.A.; Thu, V.T.H.; Hue, L.T.; et al. Long-Term Antimicrobial Performance of Textiles Coated with ZnO and TiO2 Nanoparticles in a Tropical Climate. Journal of Functional Biomaterials 2022, 13, 233. [Google Scholar] [CrossRef] [PubMed]
- Rasool, A.; Kiran, S.; Gulzar, T.; Abrar, S.; Ghaffar, A.; Shahid, M.; Nosheen, S.; Naz, S. Biogenic Synthesis and Characterization of ZnO Nanoparticles for Degradation of Synthetic Dyes: A Sustainable Environmental Cleaner Approach. Journal of Cleaner Production 2023, 398, 136616. [Google Scholar] [CrossRef]
- Chandrasekaran, S.; Anbazhagan, V. Green Synthesis of ZnO and V-Doped ZnO Nanoparticles Using Vinca Rosea Plant Leaf for Biomedical Applications. Appl Biochem Biotechnol 2024, 196, 50–67. [Google Scholar] [CrossRef]
- Ramasubbu, K.; Rajeswari, V.D. Green Synthesising ZnO Nanoparticle Using Sesbania Grandiflora and Their Evaluation of Anti-Diabetic Anti-Advanced Glycation End Products and Cytotoxic Effects. Appl Biochem Biotechnol 2024, 196, 2652–2672. [Google Scholar] [CrossRef]
- Rajeshkumar, S.; Menon, S.; Venkat Kumar, S.; Tambuwala, M.M.; Bakshi, H.A.; Mehta, M.; Satija, S.; Gupta, G.; Chellappan, D.K.; Thangavelu, L.; et al. Antibacterial and Antioxidant Potential of Biosynthesized Copper Nanoparticles Mediated through Cissus Arnotiana Plant Extract. Journal of Photochemistry and Photobiology B: Biology 2019, 197, 111531. [Google Scholar] [CrossRef] [PubMed]
- Losada-García, N.; Rodríguez-Otero, A.; Palomo, J.M. Tailorable Synthesis of Heterogeneous Enzyme–Copper Nanobiohybrids and Their Application in the Selective Oxidation of Benzene to Phenol. Catal. Sci. Technol. 2020, 10, 196–206. [Google Scholar] [CrossRef]
- Naz, S.; Gul, A.; Zia, M.; Javed, R. Synthesis, Biomedical Applications, and Toxicity of CuO Nanoparticles. Appl Microbiol Biotechnol 2023, 107, 1039–1061. [Google Scholar] [CrossRef]
- Ortega-Nieto, C.; Losada-Garcia, N.; Pessela, B.C.; Domingo-Calap, P.; Palomo, J.M. Design and Synthesis of Copper Nanobiomaterials with Antimicrobial Properties. ACS Bio Med Chem Au 2023, 3, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Madubuonu, N.; Aisida, S.O.; Ali, A.; Ahmad, I.; Zhao, T.; Botha, S.; Maaza, M.; Ezema, F.I. Biosynthesis of Iron Oxide Nanoparticles via a Composite of Psidium Guavaja-Moringa Oleifera and Their Antibacterial and Photocatalytic Study. Journal of Photochemistry and Photobiology B: Biology 2019, 199, 111601. [Google Scholar] [CrossRef] [PubMed]
- Chellapa, L.R.; Shanmugam, R.; Indiran, M.A.; Samuel, S.R. Biogenic Nanoselenium Synthesis, Its Antimicrobial, Antioxidant Activity and Toxicity. Bioinspired, Biomimetic and Nanobiomaterials 2020, 9, 184–189. [Google Scholar] [CrossRef]
- Lange, A.; Matuszewski, A.; Kutwin, M.; Ostrowska, A.; Jaworski, S. Farnesol and Selected Nanoparticles (Silver, Gold, Copper, and Zinc Oxide) as Effective Agents Against Biofilms Formed by Pathogenic Microorganisms. NSA 2024, 17, 107–125. [Google Scholar] [CrossRef]
- Yılmaz, G.E.; Göktürk, I.; Ovezova, M.; Yılmaz, F.; Kılıç, S.; Denizli, A. Antimicrobial Nanomaterials: A Review. Hygiene 2023, 3, 269–290. [Google Scholar] [CrossRef]
Figure 1.
Approaches to overcome bacterial resistance.
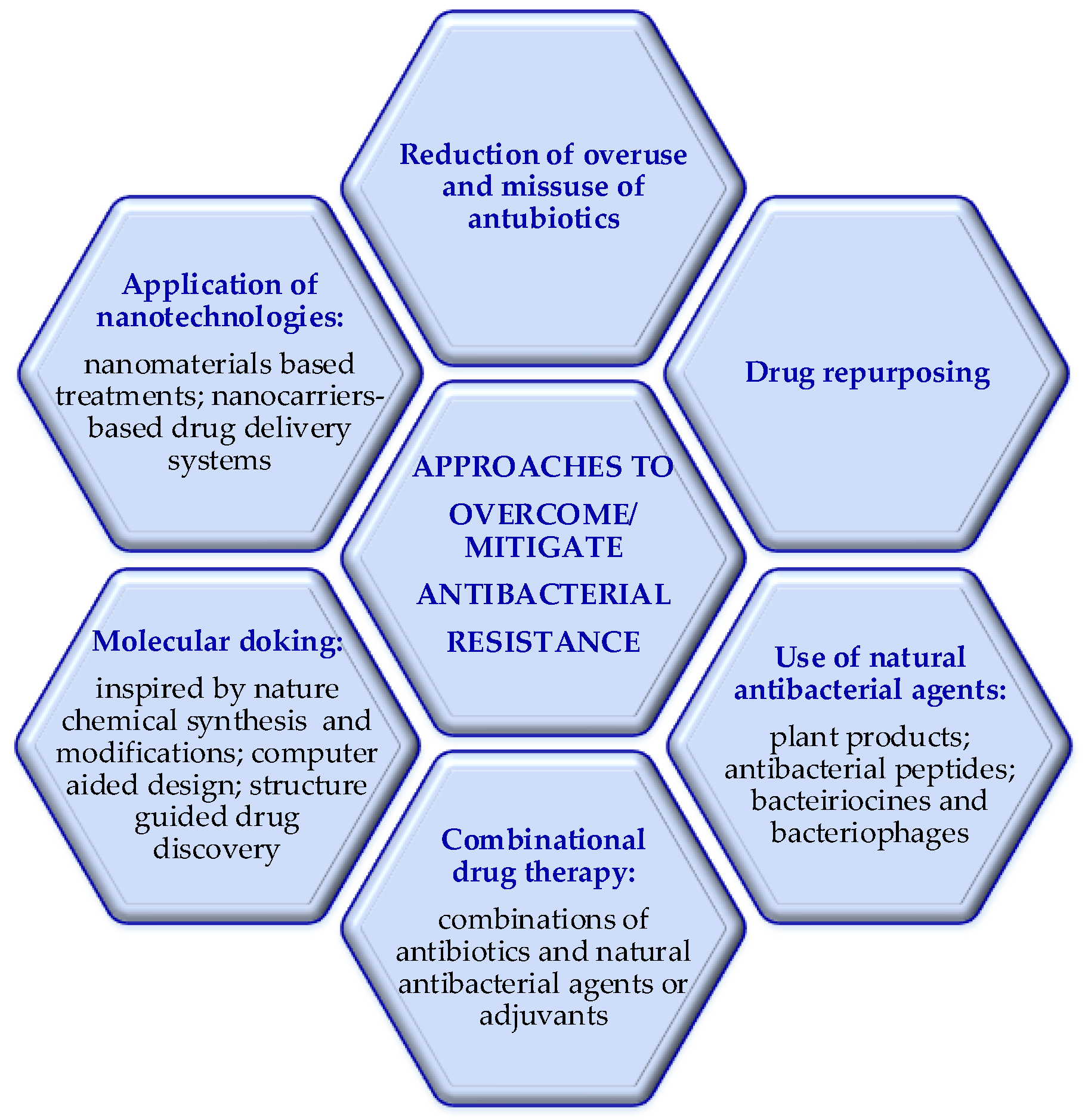
Figure 2.
Terrestrial sources of antibacterial agents.
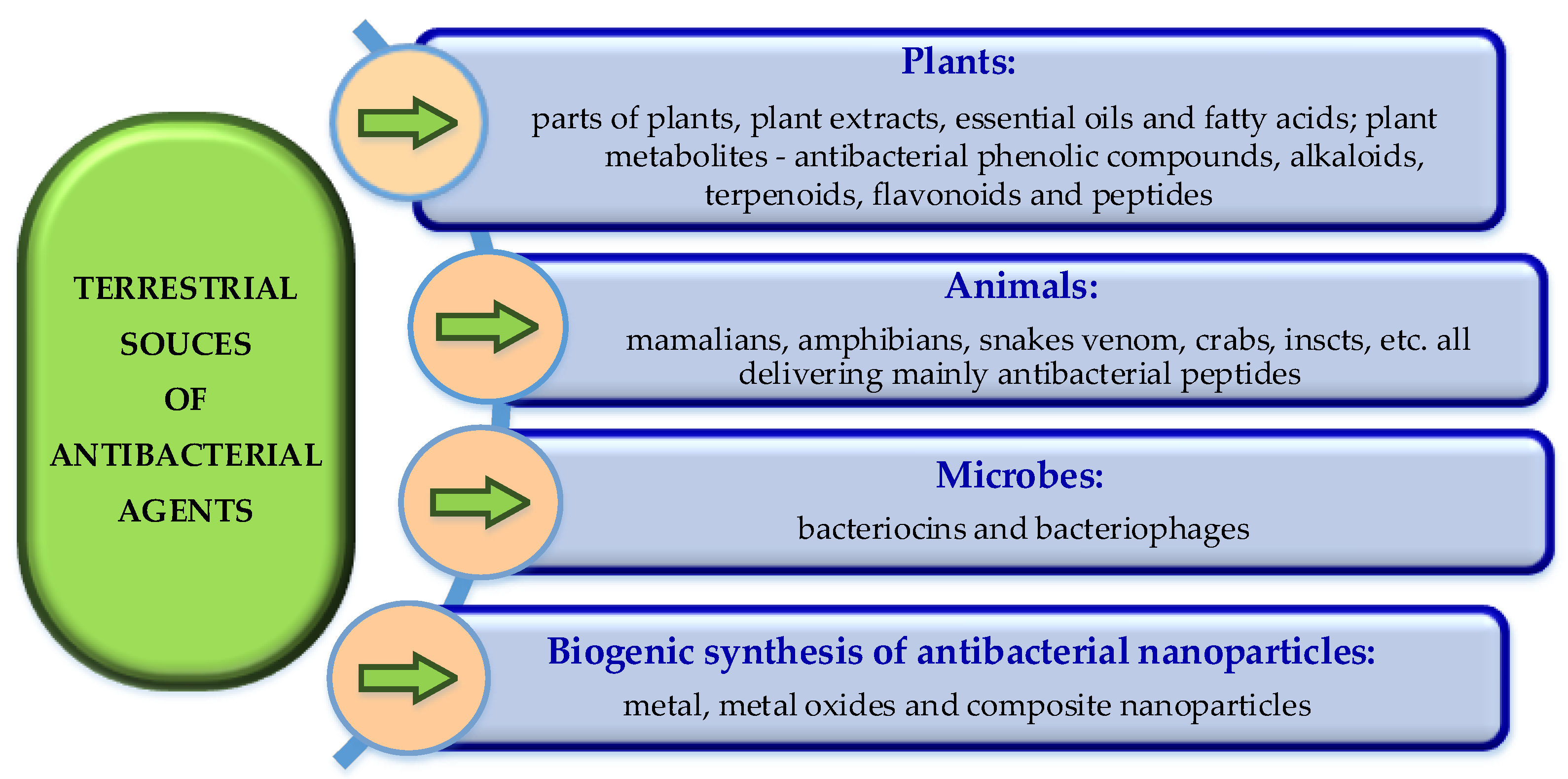
Figure 3.
Principle sketch of plant extracts preparation.
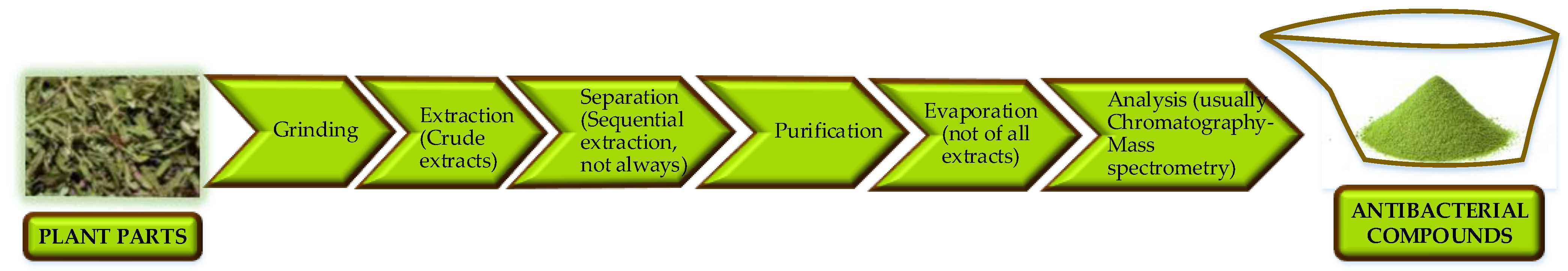
Figure 4.
Plant derived antibacterial compounds.
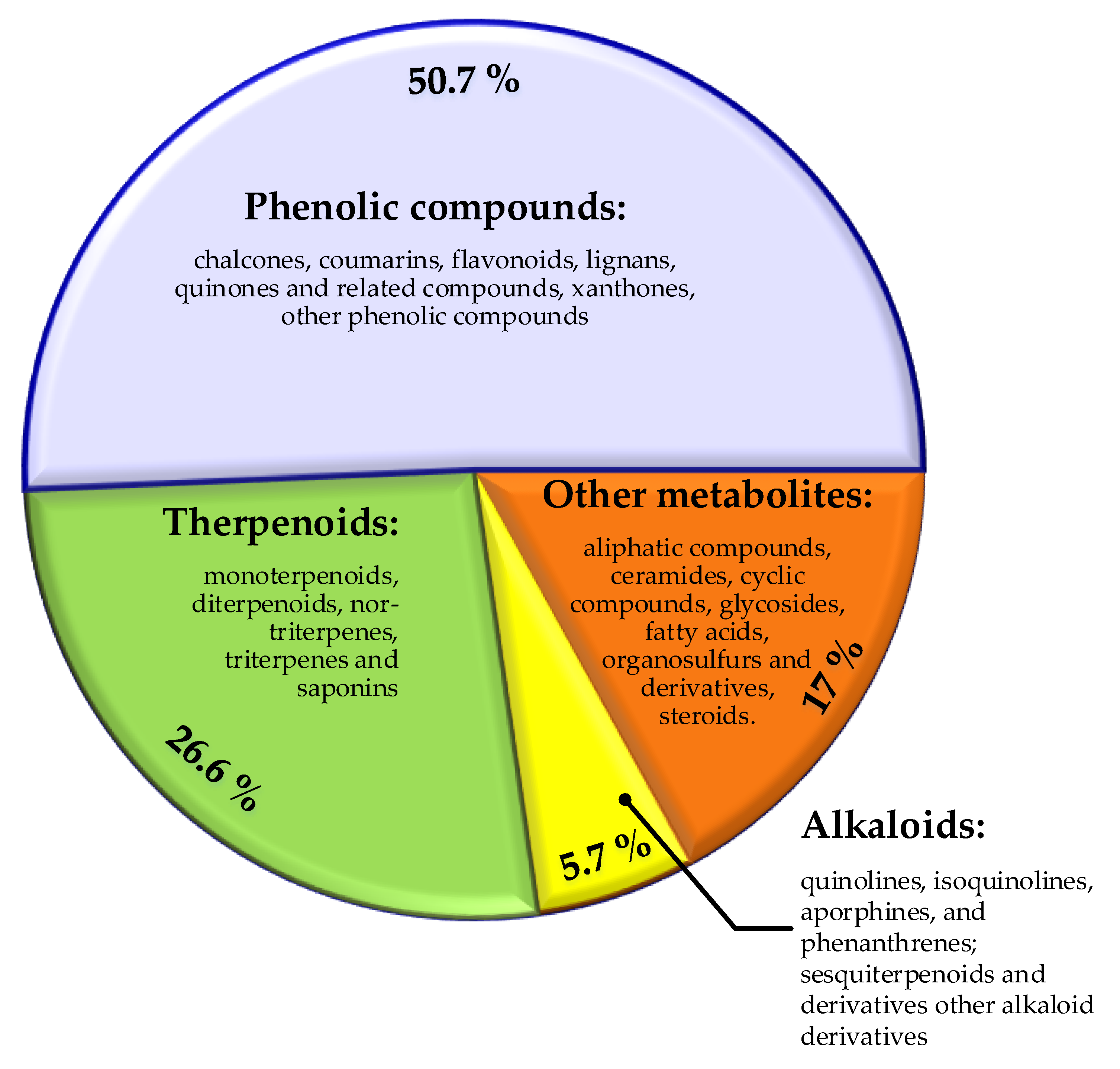
Figure 5.
Structural differences between the cell wall of Gram-negative and Gram-positive bacteria.
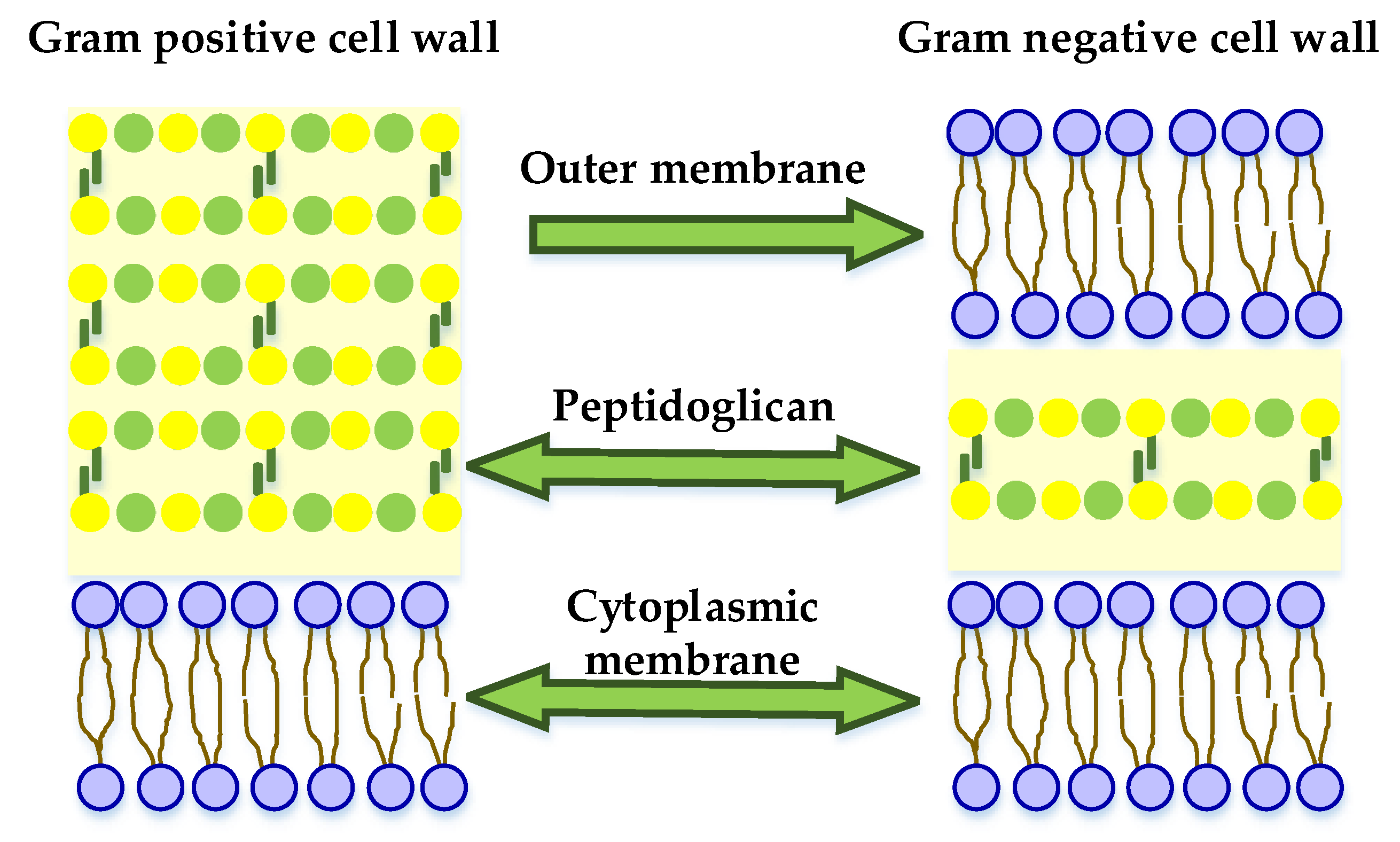
Figure 6.
Possible mode of antibacterial action of medicinal plants.
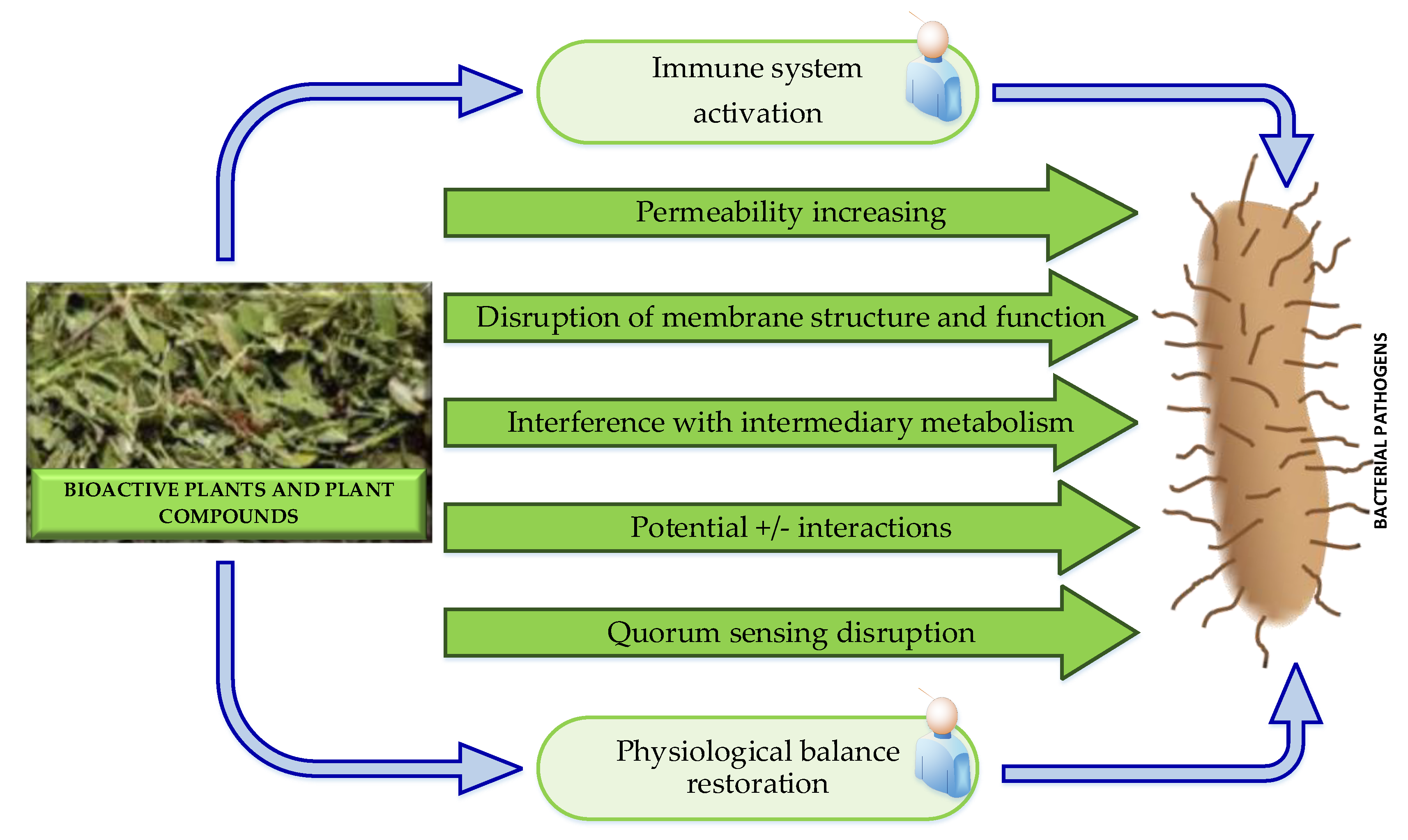
Figure 7.
Secondary structure of antibacterial peptides.
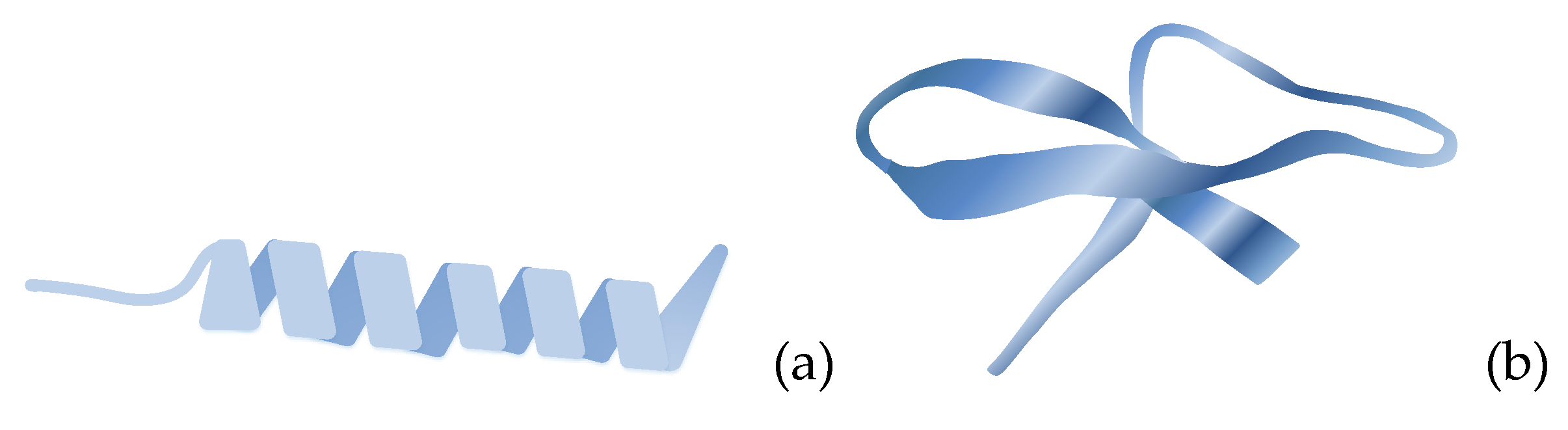
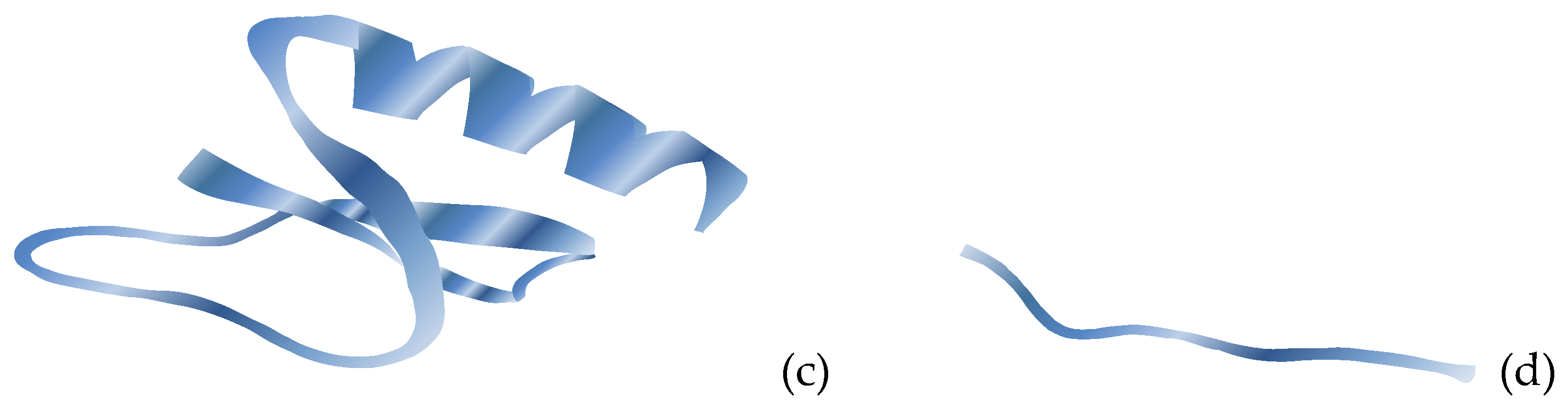
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



