
Submitted:
20 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
The Bax inhibitor-1 (BI-1) gene family, which is required for plant growth, development, and stress tolerance, remains largely unexplored in cauliflower. In this study, we identified and characterized cauliflower BI-1 family genes. On the basis of aligned homologous sequences and collinearity with Arabidopsis genes, we identified nine cauliflower BI-1 genes, which encoded proteins that varied regarding length, molecular weight, isoelectric point, and subcellular localization (i.e., Golgi apparatus, plasma membrane, cell membrane, and chloroplast). Phylogenetic analyses detected evolutionary conservation and divergence among these genes. Ten structural motifs were identified, with Motif 5 crucial for inhibiting apoptosis. According to the cis-regulatory elements in their promoters, these genes likely influence hormone signaling and stress responses. Expression profiles among tissues highlighted the diversity in the functions of these genes, which were significantly expressed in the silique and root. Focusing on BobBIL4, we investigated its role in brassinosteroid (BR)-mediated root development and salt stress tolerance. BobBIL4 expression levels increased in response to BR and salt treatments. The functional characterization of this gene in Arabidopsis revealed it enhances root growth and salinity tolerance. These findings provide insights into BI-1 gene functions in cauliflower, while also highlighting the potential utility of BobBIL4 for improving crop stress resistance.
Keywords:
Cauliflower
; Bax inhibitor-1
; Expression pattern
; BobBIL4
; Abiotic stress
1. Introduction
Cauliflower (Brassica oleracea var. botrytis), which belongs to the family Brassicaceae, is characterized by an enlarged inflorescence (i.e., curd) that serves as its primary edible organ [1]. Notably, cauliflower is cultivated globally and widely consumed as a vegetable because of its exceptional nutritional value. Its curd comprises short, thick, and fleshy stems along with an undifferentiated inflorescence meristem, making it a distinctive feature of the plant. Cauliflower curd growth and development exhibits unique reproductive characteristics that are extremely sensitive to environmental changes. This sensitivity can lead to premature flowering, formation of loose or “hairy” curds, and other developmental defects that adversely affect curd quality, potentially resulting in significant yield losses or even total crop failure. Consequently, compared with the cultivation of other commercially produced vegetables, the cultivation of cauliflower faces greater risks and challenges. Hence, the genes and associated epigenetic and physiological mechanisms regulating cauliflower responses to abiotic stresses should be identified and characterized. Elucidating these underlying mechanisms is crucial for understanding how cauliflower adapts to environmental changes. This knowledge has significant implications for breeding new varieties with enhanced stress resistance, which is essential for stabilizing yield and quality under variable environmental conditions. Developing stress-resistant varieties will mitigate production risks, while also ensuring the sustainable cultivation of cauliflower, with beneficial effects on consumer health.
Programmed cell death (PCD) is a widespread phenomenon in plants and animals [2]. While PCD is primarily known for its role in responses to various biotic and abiotic stresses, it also plays a general role in certain aspects of plant growth and development [3]. Apoptosis, a specific form of PCD, refers to a gene-regulated systematic process involving cellular self-destruction and is characterized by the loss of euchromatin structural information, leading to chromatin condensation and cell death [4]. More specifically, PCD in plants generally necessitates the activation of certain genes [5]. The Bax inhibitor-1 (BI-1) family of highly conserved transmembrane proteins has been extensively studied regarding its contributions to cell protection, ion homeostasis regulation, and anti-apoptotic activities [6]. BI-1 family genes encode critical inhibitors of apoptosis that are responsive to endogenous and exogenous stimuli [7]. These genes have been thoroughly studied in terms of their effects on stress tolerance in various species. They are crucial for enhancing the tolerance of different organisms to a wide range of environmental stresses. Earlier research indicated the heterologous expression of Arabidopsis BI-1 increases the tolerance of monocotyledonous and polycarpic plants to menadione sodium bisulfite, hydrogen peroxide, and drought-induced stress [8,9,10] Under water stress conditions, the expression of AtBI1 regulates PCD in root tips [11]. In barley, BI-1 is reportedly responsive to an infection by Blumeria graminis, resulting in compromised osmotic homeostasis [12]. Additionally, melatonin increases BI-1 expression levels in Medicago sativa roots under highly saline conditions [13]. Arabidopsis contains five BI-1 homologs, which have been designated as LIFEGUARD 1-5 (LFG1-5). Both LFG1 and LFG3 inhibit apoptosis induced by endoplasmic reticulum (ER) stress, after which they participate in the inositol-requiring enzyme 1 (IRE1) signaling pathway to enable the plant to resume normal growth [14]. These two proteins also interact with membrane-bound progesterone receptor 3 (MAPR3) to modulate ER stress-induced apoptosis mediated by the IRE1 pathway [14,15,16]. In contrast, LFG5 inhibits apoptosis through the IRE1 signaling pathway by regulating the balance of oxidized and reduced glutathione (GSH) [17]. Hence, BI-1 family genes are important for plant resistance to abiotic and biotic stresses. However, BI-1 gene families in horticultural crops have not been comprehensively investigated. More specifically, there has been relatively little research on the cauliflower BI-1 gene family and its role in abiotic stress resistance. Among BI-1 family genes, LFG2, which is also known as BRZ-INSENSITIVE-LONG HYPOCOTYLS 4 (BIL4), encodes a protein with seven transmembrane domains. The considerable interest in this gene is due to its pivotal role in plant processes, especially responses to brassinosteroid (BR) and various abiotic stresses. BIL4 positively regulates BR signaling by preventing the degradation of the BR receptor BRI1, thereby ensuring plants are appropriately responsive to BR, which is necessary for optimal growth and adaptations to environmental changes [18]. A previous study showed BIL4 is essential for maintaining cell turgor and overall stress tolerance, suggesting that it helps preserve cellular structures critical for stress tolerance [18]. BIL4 may interact with other stress-responsive genes to enhance the ability of plants to cope with adverse environmental conditions [19]. In our previous comparative genomic study, we identified structural variations in the BIL4 promoter region between cauliflower and cabbage, implying that BIL4 may differentially affect various B. oleracea morphotypes [20]. This raises several important questions: What are the characteristics and tissue-specific features of the BI-1 gene family in cauliflower? How do these genes (e.g., BIL4) modulate cauliflower growth and development, particularly in response to abiotic stress?
In this study, nine BI-1 homologs were identified in cauliflower via BLAST and collinearity analyses. A bioinformatics analysis characterized these genes in terms of their properties, phylogenetic relationships, chromosomal locations, structures, conserved motifs, and predicted cis-acting elements. In addition, BobBIL4 was expressed specifically in the ER membrane in the roots, cotyledons, petals, and apical regions of siliques. Furthermore, BobBIL4 expression was responsive to BR and resulted in substantial abiotic stress tolerance. These findings offer valuable scientific and practical insights relevant to future research on the biological role of BobBIL4 in cauliflower, with implications for clarifying the evolution of stress tolerance in Brassica species.
2. Results
2.1. Identification of Cauliflower BI-1 Family Genes
Using the Arabidopsis BI-1 gene family for a sequence homology analysis, we identified nine BI-1 family genes in cauliflower (Supplementary Figure S1). These genes were named according to the corresponding Arabidopsis genes. Specifically, the AtLFG1-4 Arabidopsis genes had one homologous gene in cauliflower, whereas AtLFG5 had three homologous genes, which resulted from whole-genome duplication and tandem duplication events. Additionally, two homologous genes of AtBI-1 were identified in cauliflower. The characteristics of the proteins encoded by the nine cauliflower BI-1 family genes were analyzed, revealing lengths ranging from 239 to 256 amino acids, molecular weights between 26.98 and 30.94 kDa, and pI between 5.88 and 9.05. The instability index varied from 24.72 to 46.95, hydrophilicity scores ranged from 0.474 to 0.932, and the lipid index was between 107.77 and 130.50. These proteins were predicted to be predominantly localized in the Golgi apparatus, plasma membrane, cell membrane, and chloroplast. Notably, these BI-1 family proteins were revealed to contain seven transmembrane domains and no signal peptides (Table 1). In terms of their chromosomal distribution, the nine cauliflower BI-1 family genes were detected on chromosomes C01, C02, C04, C05, C07, C08, and C09, of which chromosomes C02 and C07 each contained two genes, whereas the remaining five chromosomes contained one gene each (Supplementary Figure S2).
2.2. Phylogenetic Analysis of BI-1 Family Genes
To investigate the evolutionary relationships of BI-1 family genes, a phylogenetic tree was constructed using the amino acid sequences encoded by the nine BI-1 genes identified in cauliflower as well as the amino acid sequences of six, seven, and four BI-1 proteins in Arabidopsis, rice, and maize, respectively [17,21] (Figure 1). In the phylogenetic tree, the cauliflower BI-1 gene family was divided into five groups. Most cauliflower BI-1 family genes were clustered with their Arabidopsis homologs in the same group, suggestive of the evolutionary conservation within this gene family. Intriguingly, BobLFG1 in cauliflower was not clustered with AtLFG1 in Arabidopsis, indicative of a possible sequence variation and functional divergence. Sequence-based phylogenetic analyses offer valuable insights for further functional analyses of the BI-1 gene family in cauliflower.
2.3. Structural and Conserved Motif Analyses of Cauliflower BI-1 Genes
To assess the diversity in protein structures, we used Hidden Markov Models, CDD, and MEME to analyze the conserved motifs and domains of BI-1 proteins. Ten motifs were predicted within BobBI-1 amino acid sequences. Motif 5 (EKDKKKEKK) was shared by all nine BobBI-1 proteins (Figure 2A-B, D). This motif, which was located at the C-terminal end of most proteins, is crucial for inhibiting apoptosis induced by extensive stimulation [22,23]. BI-1 proteins contain seven transmembrane domains [24]. Additionally, the exon–intron organization of the nine cauliflower BI-1 genes was analyzed to gain insights into gene structures (Figure 2C). Both BobLFG5.1 and BobLFG5.2 comprised three exons, whereas BobBIL4, BobLFG4, BobLFG5.3, and BobLFG3 contained four exons. In contrast, six exons were detected in each of the remaining BI-1 genes. The diversity in the number and distribution of exons and introns among the BI-1 genes reflects a complex and heterogeneous genomic landscape.
To functionally characterize BobBI-1 genes and gain insights into the precise regulation of their stress-responsive expression, we analyzed the promoter cis-elements within the 2 kb sequence upstream of the ATG start codon of BobBI-1 genes. All BobBI-1 promoters contained elements responsive to various hormones (e.g., methyl jasmonate, gibberellin, salicylic acid, abscisic acid, and auxin) as well as elements responsive to light. Specifically, the BobLFG1 promoter included regulatory elements related to anaerobic induction and low-temperature responsiveness. The BobLFG3 promoter contained elements associated with anaerobic induction and expression in differentiated tissue. Furthermore, elements related to anaerobic induction, stress response, cold response, and hypoxia-specific induction were detected in the BobBI1.2 promoter. The promoter of BobBIL4 contained elements related to anaerobic, stress, and hypoxia-specific induction as well as circadian rhythm control. The BobLFG4 and BobLFG5.1 promoters included drought response-related elements. The BobBI1.1 promoter harbored elements associated with stress responses, including those specific to anaerobic conditions. Similarly, the BobLFG5.2 promoter contained elements related to anaerobic induction, stress induction, and differentiated tissue expression. The promoter of BobBI1.2 contained elements associated with anaerobic induction and low-temperature responsiveness. Only BobLFG5.3 and BobLFG5.2 had promoters with MYB transcription factor-binding sites and were involved in anthocyanin synthesis (Supplementary Figure S3). In addition, the promoters of all family members, except for BobLFG3, contained G-boxes. With the exception of BobBI1.1, all family members had promoters comprising Box-4 elements. The BobBI-1 promoters were particularly rich in stress-responsive elements (42 in total). In contrast, there were relatively few light-responsive (4CK-CMA1b) elements and elements related to tissue-specific expression (RY-elements) (i.e., one each). These findings suggest that cauliflower BobBI-1 genes may encode key regulators of hormone signaling pathways, stress responses, and growth and development.
2.4. BobBI-1 Expression Profiles in Different Cauliflower Tissues
To investigate the potential differences in cauliflower BI-1 gene functions, we analyzed the expression patterns of the nine BI-1 family genes in various tissues (e.g., root, stem, leaf, curd, bud, flower, and silique) using transcriptome data [20]. BobLFG3 was expressed at low levels in all examined tissues, with no detectable expression in the root, stem, leaf, and flower. This suggests that in cauliflower, BobLFG3 may play a very limited role under normal growth conditions. BobLFG1 was highly expressed only in the silique. BobLFG5.3 had the highest average expression level in all tissues, with the highest expression levels in each tissue. Additionally, its two homologous genes (BobLFG5.1 and BobLFG5.2) were also highly expressed in all tissues. Hence, these three homologous LFG5 genes, which were retained after a whole-genome triplication event, likely have important functions in cauliflower. The BobLFG4 expression level, which was relatively low in most tissues, was highest in the flower. The average BI-1 expression level was highest in the silique, followed by the root, and was lowest in the leaf. The variability in the expression of these genes was greatest in the root. BobLFG5.1, BobLFG5.2, and BobLFG1 were expressed at relatively high levels in the silique, whereas BobBIL4, BobLFG5.3, BobBI1.1, and BobBI1.2 were most highly expressed in the root (Figure 3A-C). Correlation analyses of gene expression revealed the following significant correlations among the nine BI-1 genes: BobBI1.2 with BobBIL4, BobLFG5.1, and BobLFG5.2; BobLFG1 with BobLFG3; and BobBI1.1 with BobLFG5.3 (Figure 3D). These findings suggest that the nine BI-1 family genes are differentially expressed and have undergone functional differentiation in cauliflower. Moreover, the gene expression data provide a theoretical basis for future studies on gene functions.
2.5. Analyses of BobBIL4 Expression and Subcellular Localization of the Encoded Protein
In plants, BIL4 is critical for responses to BR and stress resistance. Additionally, structural differences in the BIL4 promoter region revealed by a comparison between cauliflower and cabbage may be related to cauliflower development and stress resistance. To further functionally characterize BobBIL4 in cauliflower, we analyzed its expression pattern in various Arabidopsis tissues using a ProBobBIL4-GUS reporter line. We also investigated the subcellular localization of BobBIL4 in tobacco leaf epidermal cells. According to the detection of GUS signals, ProBobBIL4 was active in transgenic Arabidopsis root tips, cotyledons, leaves, inflorescences, and siliques (Figure 4A-E). In tobacco leaf epidermal cells, the green fluorescence due to the transient expression of an eGFP-BobBIL4 fusion protein overlapped with the red fluorescence from the RFP-HDEL fusion construct (i.e., ER marker) (Figure 4F-G). This observation is consistent with the findings of an earlier study that determined BI-1 is predominantly localized in the ER [25].
2.6. BobBIL4 Plays a Pivotal Role in BR-Mediated Root Development
Earlier research revealed the crucial regulatory effects of BR on plant root development (e.g., promoting root growth, root hair formation, and lateral root initiation), enabling roots to adapt to environmental stimuli and enhancing stress tolerance [26,27,28,29]. Previous studies in Arabidopsis have indicated that brassinosteroids (BRs) exhibit spatiotemporal dynamics in root tissues. In the current study, BobBIL4, BobLFG5.3, BobBI1.1, and BobBI1.2 were highly expressed specifically in the cauliflower root. To determine if these genes are responsive to BR in the root, we analyzed the changes in their expression within 12 h after an 24-epibrassinolide (EBL) treatment. The elongation of Arabidopsis roots is highly sensitive to BR concentration, with EBL being one of the most bioactive brassinosteroid biosynthetic intermediates. There was a continuous increase in the BobBIL4 expression level following the EBL treatment, with a significant increase detected after 8 h and peak expression at 12 h. The expression of BobLFG5.3 was affected soon after the EBL treatment, with a significant increase at 2 h, peak expression at 8 h, and a decrease at 12 h. The BobBI1.1 expression level was highest at 2 h post-treatment, after which it decreased. In contrast, BobBI1.2 expression decreased significantly after the EBL treatment. These results suggest that although these four genes were highly expressed in the root, they are differentially responsive to BR signals. Notably, BobBIL4 expression in the root increased continuously in response to BR (Figure 5A).
To further investigate the effects of BobBIL4 on BR-mediated root development, we utilized wild-type Arabidopsis (Columbia) plants, bil4 mutants (in which the endogenous BIL4 gene is knocked out), and Arabidopsis lines complemented with BobBIL4 under the control of the 35S promoter in the bil4 mutant background. As detailed in the Materials and Methods section, the expression levels of BIL4 in the complemented lines BobBIL4 in bil4 #2 and BobBIL4 in bil4 #7 are approximately 90 times higher than in the wild type. Consequently, we have designated these transgenic lines as BobBIL4-overexpressing lines/plants. The inhibitory effects of the BR antagonist Brassinazole (Brz) on BR biosynthesis were examined. Brz, a known suppressor of BR synthesis, targets plant hormones crucial for proper developmental processes. Seeds were sown on solid MS medium containing 2 µM EBL or 3 µM Brz to examine root development in 3-day-old seedlings. On normal MS medium, bil4 mutant plants had significantly shorter roots than wild-type plants. The expression of cauliflower BobBIL4 in bil4-deficient lines restored normal root development. The EBL and Brz treatments of the wild-type, bil4 mutant, and BobBIL4-overexpressing plants inhibited root growth and development, although the extent of the inhibition varied. Following the EBL treatment, root growth was inhibited the most in the bil4 mutant (according to the average root length). However, the average root length of the BobBIL4-overexpressing lines was similar to that of the wild-type control, indicating that BobBIL4 expression may mitigate the inhibitory effects of exogenous BR on root growth (Figure 5B-C, E). The addition of Brz to the MS medium inhibited root development in the examined genotypes, but the differences were not significant, suggesting that BobBIL4 likely regulates root growth and development through BR (Figure 5B, D-E). These findings highlight the importance of BobBIL4 for BR-mediated root development.
2.7. BobBIL4 Expression Enhances Salt Stress Tolerance
To clarify the effects of salt stress on the expression of BobBIL4, BobLFG5.3, BobBI1.1, and BobBI1.2, which are relatively highly expressed in the root, we analyzed the changes in their expression within 12 h after a 100 mM NaCl treatment. The expression levels of BobBIL4 and BobBI1.2 increased significantly after the salt treatment. More specifically, their expression levels initially increased, peaking at 4 h post-treatment, and then decreased. Interestingly, the extent of the expression level change was greater for BobBIL4 than for the other genes. In contrast, BobBI1.1 expression was relatively unchanged under saline conditions, suggesting it may not play a crucial role in the salt stress response. The expression of BobLFG5.3 increased significantly at 4 h post-treatment, but decreased at 12 h (Figure 6A). These results indicate that these four genes are differentially responsive to salt stress, with BobBIL4 exhibiting the most rapid and significant response.
To further analyze the contribution of BobBIL4 to the root defense against salt stress, wild-type Arabidopsis plants, bil4 mutants, and BobBIL4-overexpressing lines with a bil4-deficient genetic background were grown on MS medium containing 100 mM NaCl to compare the root development of 9-day-old seedlings. Under normal growth conditions, bil4 mutant plants had significantly shorter roots than wild-type plants and BobBIL4-overexpressing lines, implying that BIL4 plays an essential role in root growth. Moreover, BobBIL4 expression is conducive to restoring normal root development in the Arabidopsis bil4 mutant. The 100 mM NaCl treatment resulted in shortened roots in all four genotypes, indicative of inhibited root development. However, the inhibitory effects of the salt treatment on root growth were weakest for the BobBIL4-overexpressing lines (Figure 6B-D). On the basis of electrical conductivity, we assessed membrane permeability, which indicated cellular damage was most severe in the bil4 mutant, whereas the cellular damage of the BobBIL4-overexpressing lines was similar to that of the wild-type control (Supplementary Figure S4). This finding suggests that BobBIL4 expression can enhance the salt stress tolerance of Arabidopsis, highlighting its critical role in the plant response to salinity.
3. Discussion
During ontogenic and morphogenetic development, plants are exposed to multiple stressors; the detrimental effects of these stressors on intracellular homeostasis are mitigated by complex regulatory networks [30,31]. The BI-1 gene family plays a crucial role in modulating cellular stress responses and maintaining homeostasis in animal and plant species exposed to biotic and abiotic stressors [32]. This gene family has been detected in many important crops, including Triticum aestivum [33], Daucus carota [34], and Brassica napus L. [35]. However, comprehensive investigations on the functions and phylogenetic relationships of this gene family have not been conducted. In this study, we identified and characterized nine BI-1 genes in the cauliflower genome. Using a range of bioinformatics approaches, we analyzed these genes and their transcriptional profiles across various tissues and in response to abiotic stressors. The BobBIL4 response to stress was validated via heterologous expression in Arabidopsis. The results of these analyses indicate that the BobBI-1 sequence is relatively conserved and its expression patterns vary slightly in different tissues and under various stress conditions. The study findings provide the foundation for future investigations on the biological functions of BobBIL4.
Adaptations to environmental changes may involve alterations to exon–intron structures in genes [36]. Analyzing gene structures can reveal phylogenetic relationships and elucidate the evolution of gene families [37]. In the present study, analyses of BobBI-1 genes detected conserved domains (Figure 3C) as well as variations in exon–intron architecture and motifs (Figure 3B), suggestive of diverse biological roles. Moreover, although BI-1 genes were revealed to contain conserved sequences, which implies functional similarities, they differed in terms of tissue-specific expression and stress-induced expression. The characterization of the cauliflower BI-1 gene family has enhanced our understanding of stress tolerance-related mechanisms, with potential implications for optimizing crop tolerance to environmental stresses.
Identifying cis-acting elements is a standard approach to investigating temporospatial gene expression patterns as well as gene expression associated with tissue growth and development [38]. The cis-elements located within promoter regions are critical for controlling transcription and often serve as binding sites for specific transcriptional regulators [39,40]. Earlier research showed BI-1 genes mediate adaptations to abiotic and biotic stressors, especially physiological or environmental conditions that induce ER stress [41]. In Arabidopsis, AtBI-1 is expressed in a tissue-specific manner, AtLFG1 is highly expressed in young leaves, during the floral development period, and in tender siliques, and AtLFG2 (AtBIL4) is highly expressed in tender tissues and developing flowers; this gene also influences hypocotyl cell elongation via the oleuropein lactone signaling pathway [18,21]. AtLFG3 is highly expressed during seed germination and floral development stages, but its expression is also significantly upregulated under ER and salt stress conditions[15,16]. In the present study, BobBI-1 expression differed slightly among cauliflower tissues. In addition, BobBIL4, BobLFG5.3, BobBI1.1, and BobBI1.2 were highly expressed in the root. Moreover, the tissue-specific expression pattern of BobBIL4 is consistent with that of AtBIL4. Our analysis of subcellular localization indicated most of the examined BI-1 genes encode proteins primarily located in the ER membrane and Golgi apparatus. We also confirmed the subcellular localization of BobBIL4 in the ER membrane (Figure 4E-J). Future studies on BobBIL4 should focus on the effects of its expression in root tissue on membrane protein dynamics.
Apoptosis, as a form of programmed cell death, not only involves the systematic self-destruction of cells but is also marked by the progressive loss of euchromatin structural information. This loss contributes to chromatin condensation, a key feature of the apoptotic process, and underscores the complex epigenetic regulation underlying apoptosis.
The expression of BI-1 genes can be induced by various stresses, indicating that these genes may differentially contribute to stress tolerance [23]. In D. carota, the overexpression of HvBI-1 in the root confers resistance to nematodes [34]. The overexpression of AtBI-1 enhances plant tolerance to various cell death-associated stresses. For example, the heterologous expression of AtBI-1 increases the drought tolerance of transgenic sugarcane plants [10], whereas it enhances the adaptation of transgenic rice plants to oxidative stress [8]. Furthermore, PCD is affected by multiple plant hormones. A recent study showed jasmonic acid modulates the expression of PCD-related genes in salt-stressed rice seedlings [42]. Both TaBI1.1 and AtBI-1, which have highly conserved sequences, positively affect heat stress tolerance [43]. In the current study, the expression levels of four BI-1 family genes increased in the root following BR and salt treatments. The distinct expression profiles of these genes under different stress conditions may reflect their diverse roles associated with plant adaptations to stress. In darkness, the bil4 mutant is insensitive to Brz; this mutant was previously compared with BIL4-overexpressing seedlings in terms of hypocotyl elongation [21]. Consistent with the findings of an earlier study [44], we determined that BR inhibits root elongation under light. Additionally, after a Brz treatment, the BobBIL4-overexpressing lines had the shortest roots, which was in accordance with the findings of previous research on hypocotyl elongation [18,21]. The enhanced tolerance of BobBIL4-overexpressing lines to salt stress suggests BobBIL4 may be useful for developing stress-resistant crops. Research on its homologs related to abiotic stress responses and their regulatory mechanisms is currently lacking, highlighting the need for studies on the functions of these genes related to abiotic stress resistance in cauliflower.
4. Materials and Methods
4.1. Identification of BobBI-1 Gene Family Members
The ‘Korso’ cauliflower reference genome (https://figshare.com/collections/Korso_and_OX_heart_genome_assemblies_and_annotations/5392466) was used to identify BobBI-1 family genes in cauliflower [20]. PF01027 (Bax1) (http://pfam.xfam.org/) from the Pfam database was employed to identify BI-1 gene family members. Amino acid sequences were obtained from Arabidopsis and cauliflower databases and analyzed using the NCBI Conserved Domain Database (CDD) (https://www.ncbi.nlm.nih.gov/). Homology models were constructed using SMART (http://smart.embl-heidelberg.de/), whereas incomplete sequences were filtered using InterPro (https://www.ebi.ac.uk/interpro/) [45].
4.2. Determination of Biochemical Properties
BI-1 family genes were examined regarding their length as well as the amino acid composition, molecular weight, isoelectric point (pI), and hydrophilicity of the encoded proteins using ExPaSy ProtParam (https://web.expasy.org/protparam/) [46]. Additionally, protein subcellular localization was predicted using the Cell-PLoc 2.0 suite [47].
4.3. Chromosomal Localization of Cauliflower BI-1 Family Genes
For the chromosomal mapping of BI-1 genes, we used TBtools to analyze the GFF3 data for the cauliflower genome. Subsequently, gene repetition was examined using MCScanX, with the results visualized using TBtools (v2.034) [48].
4.4. Phylogenetic Analyses
Six, seven, and four BI-1 sequences from Arabidopsis, rice, and maize, respectively, were retrieved from the Ensembl Plants database (https://plants.ensembl.org) and aligned with the cauliflower BI-1 sequences using MUSCLE within MEGA-X [49]. On the basis of the aligned sequences, a phylogenetic tree was constructed according to the neighbor-joining method, with 1,000 bootstrap replicates. The tree was visualized using EvolView (https://evolgenius.info//evolview-v2/#mytrees/1/2) [49].
4.5. Analyses of Gene Structures, Conserved Motifs, and Conserved Sequences
The exon–intron architecture of genes was determined using GSDS2.0 (http://gsds.gao-lab.org/), whereas conserved motifs were identified using the MEME platform (https://meme-suite.org/meme/tools/meme) configured to detect 10 motifs [50]. In addition, DNAMAN was used for the comparative analysis of BI-1 protein sequences. Results were visualized using TBtools (v2.034).
4.6. Analysis of Cis-Acting Elements
PlantCare was used to screen the 2 kb sequence upstream of BI-1 genes for cis-acting elements [51], which were visualized using TBtools (v2.034).
4.7. Gene Duplication and Covariance Analyses
According to the chromosomal locations of BI-1 genes, intra- and inter-genic covariances among cauliflower and Arabidopsis BI-1 genes were analyzed using MapChart (v2.0) [52]. MCScanX was used to examine segmental duplication events and homologous relationships.
4.8. Subcellular Localization of BobBIL4
The open reading frames (ORFs) of BobBIL4 was cloned into pFGC-eGFP for the subsequent transformation of tobacco (Nicotiana benthamiana) epidermal cells. Seedlings were grown in an insect-free chamber at 25 °C with an 8 h light:16 h dark cycle. Epidermal cells were transformed with the recombinant plasmid using Agrobacterium tumefaciens GV3101. For the co-expression of eGFP-BobBIL4, RFP-HDEL, and p19, bacterial cultures were mixed at a 1:1:1 ratio and incubated at room temperature for 1-3 h prior to the infiltration of tobacco at the 5- to 6-leaf stage using a 1 mL syringe. Fluorescence was captured at 48 h post-infiltration using a LSM780 (Carl Zeiss, Jena, Germany), with settings adjusted for detecting fluorophores.
4.9. Preparation of Plant Materials and Stress Treatments
Stress treatments were completed using cauliflower ‘Korso’ plants [20]. Seeds were surface-sterilized and germinated on solid Murashige and Skoog (MS) medium for 6 days in a temperature-controlled incubator. Uniformly growing seedlings were selected for the following liquid MS medium-based stress treatments (0, 2, 4, 8, and 12 h): 2 μM 24-epibrassinolide (EBL) or 3 μM brassinazole (Brz) (hormones) and 200 μM NaCl (salinity). Root samples were harvested, flash-frozen in liquid nitrogen, and stored at -80 °C until analyzed.
Arabidopsis thaliana Columbia-0 ecotype seeds were preserved in the laboratory. The bil4 mutant (Salk_052507C) was obtained from the Salk Institute T-DNA collection, with the T-DNA insertion verified by PCR using plant genomic DNA and primers flanking the T-DNA and gene-specific sequences. Transgenic Arabidopsis were generated via floral dip transformation. For the wild-type Columbia background, we introduced the BobBIL4 promoter (ProBobBIL4) fused with the GUS reporter gene to analyze tissue-specific expression patterns. In the bil4 mutant background, we introduced the coding sequence (CDS) of BobBIL4 under the control of the 35S promoter to complement the bil4 mutation and further investigate the functional role of BobBIL4. We utilized quantitative fluorescence PCR to measure the expression of the BIL4 gene in wild-type plants and the BobBIL4 gene in the complemented lines BobBIL4 in bil4 #2 and BobBIL4 in bil4 #7. The results showed that the expression levels of BobBIL4 in the two transgenic lines were 84 and 90 times higher than the BIL4 expression in the wild type, respectively. The bil4 mutant line shows significantly reduced expression compared to the wild type, consistent with the findings of Wang et al (2019) [16] (Figure S4).Seeds underwent stratification at4°C for a period of 2 days to induce germination. Subsequently, the plants were cultivated under a regime of continuous on a 1/2 strength Murashige and Skoog (MS) medium (22°C, 16 h light/8 h dark cycle and 60% humidity). To examine promoter activity patterns among tissues and developmental stages. Seedlings and the inflorescence organs of proBobBIL4 were incubated for 12 h, at 37℃ in the staining buffer, respectively.
For the salt stress experiments, 3-day-old seedlings with the same growth trend were transferred to a medium containing 100 mM NaCl, 2 μM EBL and 3 μM Brz. After 9-day treatment, root length was measured and photographed, and lon Leakage Measurement.
4.10. Ion Leakage Measurement
The permeability of the cell membrane was evaluated through the quantification of ion efflux from seedlings post-salinity treatment. The conductivity was assessed as previously reported [15]. Following each assay, 30 seedlings were submerged in 20 mL of distilled water with mild agitation for 2 h at room temperature. Each sample was subjected to triplicate biological replicates.
4.11. Total RNA Extraction and Expression Analysis
Total RNA was extracted using an RNAprep Pure Kit (Vazyme, Nanjing, China). RNA quality was assessed via 1% agarose gel electrophoresis, whereas the RNA concentration and purity were determined using a NanoDrop 2000 spectrophotometer (ThermoFisher Science, China). First-strand cDNA was synthesized from 1 μg RNA using HiScript® III All-in-one RT SuperMix (Vazyme) for the qRT-PCR analysis of gene expression using Taq Pro Universal SYBR qPCR Master Mix (Takara, Japan) and a CFX96 Real-time PCR instrument (BioRad, USA). Gene-specific primers were designed using Integrated DNA Technologies online software (idtdna.com). BobACTIN (BolK_3g69850) was selected as an internal reference gene. The qRT-PCR analysis (three technical replicates per reaction) was completed using a reaction mixture comprising 10 μL Taq Pro Universal SYBR qPCR Master Mix, 0.5 μL forward/reverse primers, 1 μL cDNA template, and 8 μL ddH2O. The qRT-PCR program was as follows: 95 °C for 2 min; 39 cycles of 95 °C for 10 s and 60 °C for 30 s. Relative expression levels were calculated according to the 2-∆∆Ct method [53].
Seven different tissues of Korso (root, stem, leaf, curd, bud, flower, and silique) were collected for transcriptome sequencing. The tissues were assessed as previously reported [20]. Tissue-specific expression patterns were visualized using heatmaps generated by R (https://www-project.org).
4.12. Statistical Analysis
Data analyzed were processed and visualized by GraphPad Prism 9.0 (GraphPad Software, San Diego, CA, USA) and shown in mean ± SD. Significant differences were identified using one-way ANOVA. Student’s t-test was used to calculate P values (* P < 0.05, *** P < 0.01, **** P < 0.001).
5. Conclusions
In this study, we identified and characterized the cauliflower BI-1 gene family. The evolutionary conservation and divergence among these genes may reflect a complex interplay between functional redundancy and specialization that enables cauliflower to adapt to diverse external stresses. The critical roles of specific BI-1 genes, such as BobBIL4, in hormone signaling and stress responses, are relevant to future research aimed at improving crop resilience through genetic manipulation. Additional studies focusing on the mechanistic basis of BI-1-mediated stress responses are necessary to ensure these findings are exploited to improve the production of cauliflower and other agriculturally important crops.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Multiple sequence alignment of nine Bax inhibitor-1 proteins in Brassica oleracea var. botrytis. Figure. S2: Schematic representation of the chromosome distribution of the cauliflower BI-1 genes. Figure. S3: The distributions of predicted cis-acting elements in cauliflower BI-1 genes, with different colors representing cis-acting elements with distinct functions. Figure S4: Relative expression levels of the BIL4 gene in wild-type Arabidopsis (Col) and BobBIL4 in bil4 mutant lines. Figure. S5: BobBIL4 reduced cell death induced by salt stress. Table S1: Primer sequences of RT-qPCR.
Author Contributions
F.L., N.G., and J.Z. conceived and designed the research. X.L. completed the experiment and wrote the manuscript. N.G., J.Z. and F.L. revised and edited the manuscript. X.L., G.W. and N.G. designed the entire experiment. X.L., S.L. and M.D. performed data analysis. X.L., G.W., M.Z., S.H. and Z.W. performed experiments. N.G., F.L. and X.L. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by grants from the National Natural Science Foundation of China (32072576), the Innovation Program of the Beijing Academy of Agricultural and Forestry Sciences (KJCX20230121), Collaborative innovation program of the Beijing Vegetable Research Center (XTCX202302), Establishment of the International Cooperation Base for Young Scientific and Technological Talent in Vegetable Germplasm Improvement (030210110).
Data Availability Statement
All data are included in the article. RNA-seq raw data are available at the NCBI BioProject database under accession numbers PRJNA546441.
Acknowledgments
We thank the State Key Laboratory of Vegetable Biobreeding and Beijing Key Laboratory of Vegetable Germplasms Improvement. We thank Liwen Bianji (Edanz) (www.liwenbianji.cn) for editing the English text of a draft of this manuscript.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- DR., S., Flower development. Origin of the cauliflower. Current Biology 1995, 5(4), 361-3.
- Metzstein, M., Stanfield, G., Horvitz, H,. , Genetics of programmed cell death in C. elegans: past, present and future Trends Genet, 1998, (410-416), 14.
- Watanabe, N.; Lam, E., Arabidopsis bax inhibitor-1. Plant Signaling & Behavior 2008, 3, (8), 564-566.
- Kabbage, M.; Kessens, R.; Bartholomay, L. C.; Williams, B., The Life and Death of a Plant Cell. Annu Rev Plant Biol 2017, 68, 375-404.
- Richael, C.; Lincoln, J. E.; Bostock, R. M.; Gilchrist, D. G., Caspase inhibitors reduce symptom development and limit bacterial proliferation in susceptible plant tissues. Physiological and Molecular Plant Pathology 2001, 59, (4), 213-221.
- Kerstin Reimers, C. Y. U. C., Vesna Bucan and Peter M. Vogt, The Bax Inhibitor-1 (BI-1) Family in Apoptosis and Tumorigenesis. Current Molecular Medicine 2008, 8, 148-156.
- B Li, R. K. Y., G S Jeong, H-R Kim, H-J Chae, The Characteristics of Bax Inhibitor-1 and its Relate. Current Molecular Medicine, 2014, (14(5)), 603-615.
- Ishikawa, T.; Takahara, K.; Hirabayashi, T.; Matsumura, H.; Fujisawa, S.; Terauchi, R.; Uchimiya, H.; Kawai-Yamada, M., Metabolome Analysis of Response to Oxidative Stress in Rice Suspension Cells Overexpressing Cell Death Suppressor Bax Inhibitor-1. Plant and Cell Physiology 2010, 51, (1), 9-20.
- Ishikawa, T.; Uchimiya, H.; Kawai-Yamada, M., The Role of Plant Bax Inhibitor-1 in Suppressing H2O2-Induced Cell Death. In Hydrogen Peroxide and cell signaling, Part B, 2013; pp 239-256.
- Ramiro, D. A.; Melotto-Passarin, D. M.; Barbosa, M. d. A.; Santos, F. d.; Gomez, S. G. P.; Massola Júnior, N. S.; Lam, E.; Carrer, H., Expression of Arabidopsis Bax Inhibitor-1 in transgenic sugarcane confers drought tolerance. Plant Biotechnology Journal 2016, 14, (9), 1826-1837.
- Duan, Y.; Zhang, W.; Li, B.; Wang, Y.; Li, K.; Sodmergen; Han, C.; Zhang, Y.; Li, X., An endoplasmic reticulum response pathway mediates programmed cell death of root tip induced by water stress in Arabidopsis. New Phytologist 2010, 186, (3), 681-695.
- Eichmann R, B. M., Weis C, Shaw J, Lacomme C, Schweizer P, Duchkov D, Hensel G, Kumlehn J, Hückelhoven R. , BAX INHIBITOR-1 is required for full susceptibility of barley to powdery mildew. Mol Plant Microbe Interact. 2010, 23(9):, 1217-27.
- Jalili, S.; Ehsanpour, A. A.; Javadirad, S. M., The role of melatonin on caspase-3-like activity and expression of the genes involved in programmed cell death (PCD) induced by in vitro salt stress in alfalfa (Medicago sativa L.) roots. Botanical Studies 2022, 63, (1).
- Zhu, M.; Tang, X.; Wang, Z.; Xu, W.; Zhou, Y.; Wang, W.; Li, X.; Li, R.; Guo, K.; Sun, Y.; Zhang, W.; Xu, L.; Li, X., Arabidopsis GAAPs interacting with MAPR3 modulate the IRE1-dependent pathway upon endoplasmic reticulum stress. J Exp Bot 2019, 70, (21), 6113-6125.
- Guo, K.; Wang, W.; Fan, W.; Wang, Z.; Zhu, M.; Tang, X.; Wu, W.; Yang, X.; Shao, X.; Sun, Y.; Zhang, W.; Li, X., Arabidopsis GAAP1 and GAAP3 Modulate the Unfolded Protein Response and the Onset of Cell Death in Response to ER Stress. Front Plant Sci 2018, 9, 348.
- Wang, W.; Li, X.; Zhu, M.; Tang, X.; Wang, Z.; Guo, K.; Zhou, Y.; Sun, Y.; Zhang, W.; Li, X., Arabidopsis GAAP1 to GAAP3 Play Redundant Role in Cell Death Inhibition by Suppressing the Upregulation of Salicylic Acid Pathway Under Endoplasmic Reticulum Stress. Front Plant Sci 2019, 10, 1032.
- Tang, X. H.; Li, X.; Zhou, Y.; He, Y. T.; Wang, Z. Y.; Yang, X.; Wang, W.; Guo, K.; Zhang, W.; Sun, Y.; Li, H. Q.; Li, X. F., Golgi anti-apoptotic proteins redundantly counteract cell death by inhibiting production of reactive oxygen species under endoplasmic reticulum stress. J Exp Bot 2022, 73, (8), 2601-2617.
- Yamagami, A.; Saito, C.; Nakazawa, M.; Fujioka, S.; Uemura, T.; Matsui, M.; Sakuta, M.; Shinozaki, K.; Osada, H.; Nakano, A.; Asami, T.; Nakano, T., Evolutionarily conserved BIL4 suppresses the degradation of brassinosteroid receptor BRI1 and regulates cell elongation. Scientific Reports 2017, 7, (1).
- Yamagami, A.; Chieko, S.; Sakuta, M.; Shinozaki, K.; Osada, H.; Nakano, A.; Asami, T.; Nakano, T., Brassinosteroids regulate vacuolar morphology in root meristem cells of Arabidopsis thaliana. Plant Signaling & Behavior 2018, 13, (4).
- Guo, N.; Wang, S.; Gao, L.; Liu, Y.; Wang, X.; Lai, E.; Duan, M.; Wang, G.; Li, J.; Yang, M.; Zong, M.; Han, S.; Pei, Y.; Borm, T.; Sun, H.; Miao, L.; Liu, D.; Yu, F.; Zhang, W.; Ji, H.; Zhu, C.; Xu, Y.; Bonnema, G.; Li, J.; Fei, Z.; Liu, F., Genome sequencing sheds light on the contribution of structural variants to Brassica oleracea diversification. BMC Biol 2021, 19, (1), 93.
- Yamagami, A.; Nakazawa, M.; Matsui, M.; Tujimoto, M.; Sakuta, M.; Asami, T.; Nakano, T., Chemical Genetics Reveal the Novel Transmembrane Protein BIL4, Which Mediates Plant Cell Elongation in Brassinosteroid Signaling. Bioscience, Biotechnology, and Biochemistry 2009, 73, (2), 415-421.
- Carrara, G.; Saraiva, N.; Gubser, C.; Johnson, B. F.; Smith, G. L., Six-transmembrane Topology for Golgi Anti-apoptotic Protein (GAAP) and Bax Inhibitor 1 (BI-1) Provides Model for the Transmembrane Bax Inhibitor-containing Motif (TMBIM) Family. Journal of Biological Chemistry 2012, 287, (19), 15896-15905.
- Watanabe, N.; Lam, E., Bax Inhibitor-1, a Conserved Cell Death Suppressor, Is a Key Molecular Switch Downstream from a Variety of Biotic and Abiotic Stress Signals in Plants. International Journal of Molecular Sciences 2009, 10, (7), 3149-3167.
- Reimers K, C. C., Mau-Thek E, Vogt PM, Sequence analysis shows that Lifeguard belongs to a new evolutionarily conserved cytoprotective family. INTERNATIONAL JOURNAL OF MOLECULAR MEDICINE 2006, 18, 729-734.
- Reed, Q. X. a. J. C., Bax Inhibitor-1, a Mammalian Apoptosis Suppressor Identified by Functional Screening in Yeast. Molecular Cel 1998, 1, (227-346).
- Nolan, T. M.; Vukasinovic, N.; Liu, D.; Russinova, E.; Yin, Y., Brassinosteroids: Multidimensional Regulators of Plant Growth, Development, and Stress Responses. Plant Cell 2020, 32, (2), 295-318.
- Jiroutová; Mikulík; Novák; Strnad; Oklestkova, Brassinosteroids Induce Strong, Dose-Dependent Inhibition of Etiolated Pea Seedling Growth Correlated with Ethylene Production. Biomolecules 2019, 9, (12).
- Della Rovere, F.; Piacentini, D.; Fattorini, L.; Girardi, N.; Bellanima, D.; Falasca, G.; Altamura, M. M.; Betti, C., Brassinosteroids Mitigate Cadmium Effects in Arabidopsis Root System without Any Cooperation with Nitric Oxide. International Journal of Molecular Sciences 2022, 23, (2).
- Nolan, T. M.; Vukašinović, N.; Hsu, C.-W.; Zhang, J.; Vanhoutte, I.; Shahan, R.; Taylor, I. W.; Greenstreet, L.; Heitz, M.; Afanassiev, A.; Wang, P.; Szekely, P.; Brosnan, A.; Yin, Y.; Schiebinger, G.; Ohler, U.; Russinova, E.; Benfey, P. N., Brassinosteroid gene regulatory networks at cellular resolution in theArabidopsisroot. Science 2023, 379, (6639).
- Waadt, R.; Seller, C. A.; Hsu, P. K.; Takahashi, Y.; Munemasa, S.; Schroeder, J. I., Plant hormone regulation of abiotic stress responses. Nat Rev Mol Cell Biol 2022, 23, (10), 680-694.
- Huang, Y.; Yang, R.; Luo, H.; Yuan, Y.; Diao, Z.; Li, J.; Gong, S.; Yu, G.; Yao, H.; Zhang, H.; Cai, Y., Arabidopsis Protein Phosphatase PIA1 Impairs Plant Drought Tolerance by Serving as a Common Negative Regulator in ABA Signaling Pathway. Plants 2023, 12, (14).
- Robinson, K. S.; Clements, A.; Williams, A. C.; Berger, C. N.; Frankel, G., Bax Inhibitor 1 in apoptosis and disease. Oncogene 2011, 30, (21), 2391-2400.
- Weis, C.; Huckelhoven, R.; Eichmann, R., LIFEGUARD proteins support plant colonization by biotrophic powdery mildew fungi. J Exp Bot 2013, 64, (12), 3855-67.
- Imani, J.; Baltruschat, H.; Stein, E.; Jia, G.; Vogelsberg, J.; Kogel, K. H.; Huckelhoven, R., Expression of barley BAX Inhibitor-1 in carrots confers resistance to Botrytis cinerea. Mol Plant Pathol 2006, 7, (4), 279-84.
- Chae, H. J.; Ke, N.; Kim, H. R.; Chen, S.; Godzik, A.; Dickman, M.; Reed, J. C., Evolutionarily conserved cytoprotection provided by Bax Inhibitor-1 homologs from animals, plants, and yeast. Gene 2003, 323, 101-13.
- Feng, X.; Xu, Y.; Peng, L.; Yu, X.; Zhao, Q.; Feng, S.; Zhao, Z.; Li, F.; Hu, B., TaEXPB7-B, a beta-expansin gene involved in low-temperature stress and abscisic acid responses, promotes growth and cold resistance in Arabidopsis thaliana. J Plant Physiol 2019, 240, 153004.
- Xue, J. Y.; Wang, Y.; Wu, P.; Wang, Q.; Yang, L. T.; Pan, X. H.; Wang, B.; Chen, J. Q., A primary survey on bryophyte species reveals two novel classes of nucleotide-binding site (NBS) genes. PLoS One 2012, 7, (5), e36700.
- Zhao, Q.; Luo, Z.; Chen, J.; Jia, H.; Ai, P.; Chen, A.; Li, Y.; Xu, G., Characterization of two cis-acting elements, P1BS and W-box, in the regulation of OsPT6 responsive to phosphors deficiency. Plant Growth Regulation 2021, 93, (3), 303-310.
- Zou, C.; Sun, K.; Mackaluso, J. D.; Seddon, A. E.; Jin, R.; Thomashow, M. F.; Shiu, S. H., Cis-regulatory code of stress-responsive transcription in Arabidopsis thaliana. Proc Natl Acad Sci U S A 2011, 108, (36), 14992-7.
- Molina, C.; Grotewold, E., Genome wide analysis of Arabidopsis core promoters. BMC Genomics 2005, 6, 25.
- Cybulsky, A. V., Endoplasmic reticulum stress, the unfolded protein response and autophagy in kidney diseases. Nature Reviews Nephrology 2017, 13, (11), 681-696.
- Khan, M. S.; Hemalatha, S., Autophagy and Programmed Cell Death Are Critical Pathways in Jasmonic Acid Mediated Saline Stress Tolerance in Oryza sativa. Appl Biochem Biotechnol 2022, 194, (11), 5353-5366.
- Lu, P. P.; Zheng, W. J.; Wang, C. T.; Shi, W. Y.; Fu, J. D.; Chen, M.; Chen, J.; Zhou, Y. B.; Xi, Y. J.; Xu, Z. S., Wheat Bax Inhibitor-1 interacts with TaFKBP62 and mediates response to heat stress. BMC Plant Biol 2018, 18, (1), 259.
- Bingsheng Lv, H. T., Feng Zhang, Jiajia Liu1, Songchong Lu, Mingyi Bai, Chuanyou Li, Zhaojun Ding, Brassinosteroids regulate root growth by controlling reactive oxygen species homeostasis and dual effect on ethylene synthesis in Arabidopsis. PLOS Genetics 2018, 14, (1).
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G. A.; Sonnhammer, E. L. L.; Tosatto, S. C. E.; Paladin, L.; Raj, S.; Richardson, L. J.; Finn, R. D.; Bateman, A., Pfam: The protein families database in 2021. Nucleic Acids Res 2021, 49, (D1), D412-D419.
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C., Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res 2021, 49, (W1), W216-W227.
- Horton, P.; Park, K. J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C. J.; Nakai, K., WoLF PSORT: protein localization predictor. Nucleic Acids Res 2007, 35, (Web Server issue), W585-7.
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H. R.; Frank, M. H.; He, Y.; Xia, R., TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol Plant 2020, 13, (8), 1194-1202.
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K., MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol 2018, 35, (6), 1547-1549.
- Bailey, T. L.; Boden, M.; Buske, F. A.; Frith, M.; Grant, C. E.; Clementi, L.; Ren, J.; Li, W. W.; Noble, W. S., MEME SUITE: tools for motif discovery and searching. Nucleic Acids Research 2009, 37, (Web Server), W202-W208.
- Magali Lescot, P. D., Gert Thijs, Kathleen Marchal, Yves Moreau, Yves Van de Peer, Pierre Rouzé, Stephane Rombauts, PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Research 2002, 30(1), 325-7.
- Wang, Y.; Tang, H.; DeBarry, J. D.; Tan, X.; Li, J.; Wang, X.; Lee, T. h.; Jin, H.; Marler, B.; Guo, H.; Kissinger, J. C.; Paterson, A. H., MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Research 2012, 40, (7), e49-e49.
- Jo Vandesompele , K. D. P., Filip Pattyn, Bruce Poppe, Nadine Van Roy, Anne De Paepe, Frank Speleman, Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biology 2002, 3(7), RESEARCH0034.
Figure 1.
Phylogenetic tree of BI-1 genes in cauliflower, Arabidopsis, rice, and maize. Neighbor-joining trees are generated using the MEGA software, with 1000 bootstrap replicates. All BI-1 genes were classified into five subgroups, which are differentiated by color.
Figure 1.
Phylogenetic tree of BI-1 genes in cauliflower, Arabidopsis, rice, and maize. Neighbor-joining trees are generated using the MEGA software, with 1000 bootstrap replicates. All BI-1 genes were classified into five subgroups, which are differentiated by color.
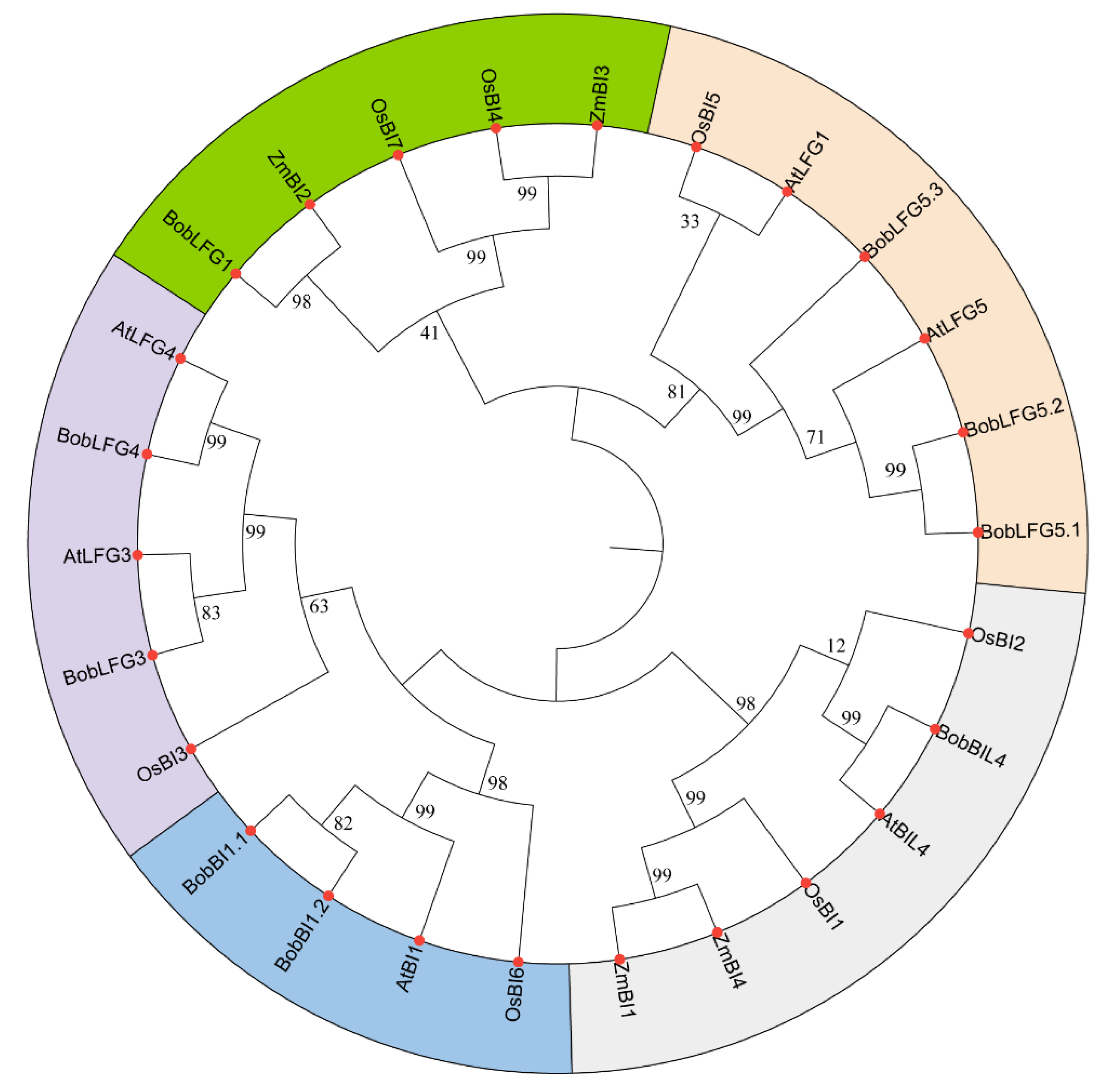
Figure 2.
Structural characteristics of domains and promoter cis-regulatory elements of BI-1 family genes in cauliflower. A. Phylogenetic tree. B. Conserved motifs. C. Structure of BobBI-1 genes. D. Classification and number of promoter cis-acting elements, with the color intensity reflecting the number of cis-acting elements.
Figure 2.
Structural characteristics of domains and promoter cis-regulatory elements of BI-1 family genes in cauliflower. A. Phylogenetic tree. B. Conserved motifs. C. Structure of BobBI-1 genes. D. Classification and number of promoter cis-acting elements, with the color intensity reflecting the number of cis-acting elements.

Figure 3.
Expression profiles and correlations analysis of BI-1 family genes in cauliflower across different tissues. A. Boxplot showing the expression levels (FPKM) of nine BI-1 family genes in cauliflower across various tissues including root, stem, leaf, curd, bud, flower, and silique. B. Boxplot depicting the overall expression distribution of the nine BI-1 family genes in each tissue type, indicating differential gene expression patterns across the tissues. C. Heatmap illustrating the expression profiles of the nine BI-1 family genes in different cauliflower tissues. The color scale represents the expression level, with red indicating higher expression and blue indicating lower expression. D. Correlation heatmap showing the Pearson correlation coefficients between the expression levels of the nine BI-1 family genes in different cauliflower tissues.
Figure 3.
Expression profiles and correlations analysis of BI-1 family genes in cauliflower across different tissues. A. Boxplot showing the expression levels (FPKM) of nine BI-1 family genes in cauliflower across various tissues including root, stem, leaf, curd, bud, flower, and silique. B. Boxplot depicting the overall expression distribution of the nine BI-1 family genes in each tissue type, indicating differential gene expression patterns across the tissues. C. Heatmap illustrating the expression profiles of the nine BI-1 family genes in different cauliflower tissues. The color scale represents the expression level, with red indicating higher expression and blue indicating lower expression. D. Correlation heatmap showing the Pearson correlation coefficients between the expression levels of the nine BI-1 family genes in different cauliflower tissues.
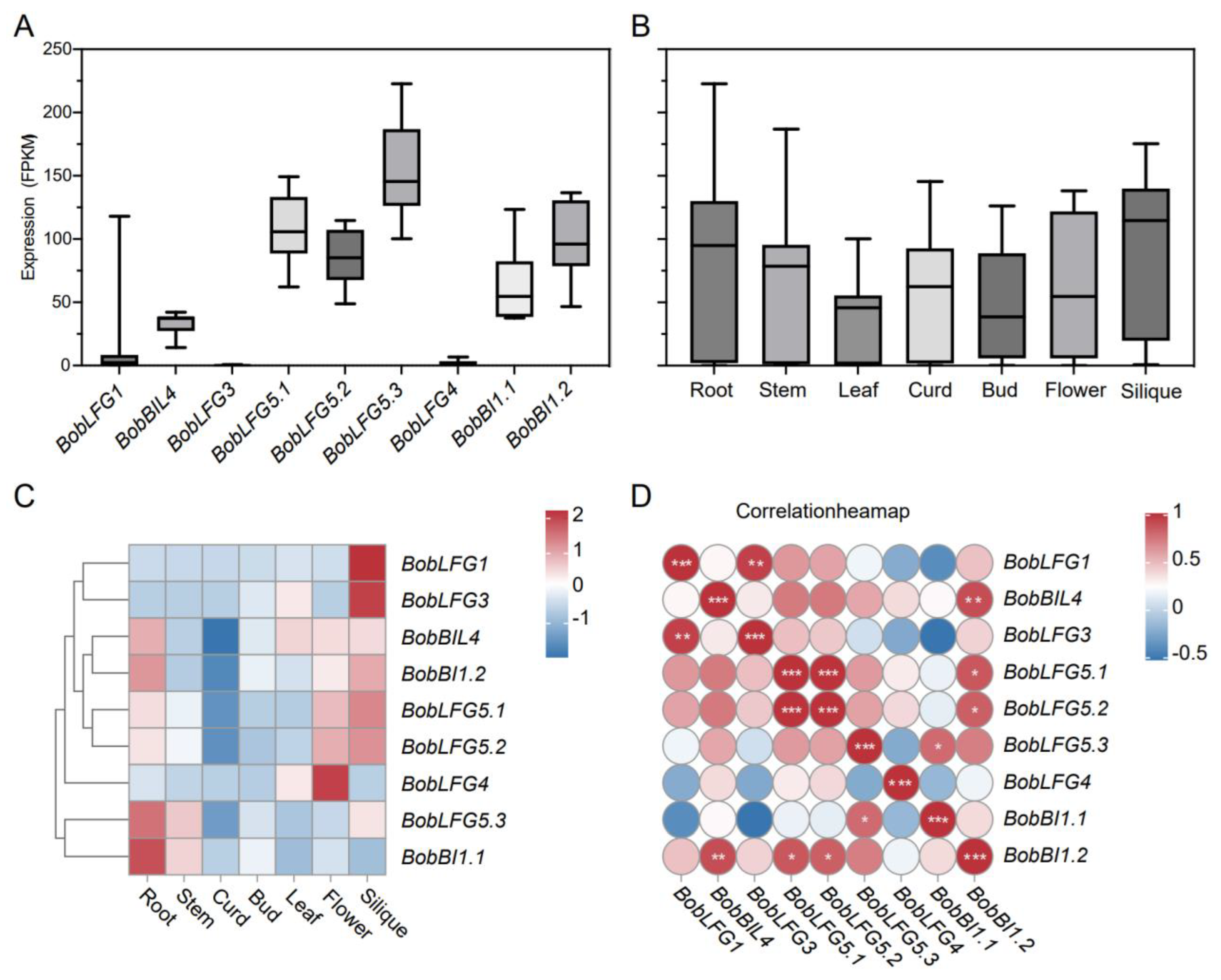
Figure 4.
BobBIL4 expression patterns in Arabidopsis and subcellular localization. A-E. Tissue-specific expression of BobBIL4 in transgenic Arabidopsis. F-J. Subcellular localization of BobBIL4 in Nicotiana benthamiana leaf epidermal cells. Cells containing the empty vector pFGC-eGFP served as the control. F. eGFP image. G. Merged eGFP and bright field images. H. Subcellular localization of eGFP-BobBIL4. I. Endoplasmic reticulum marker RFP-HDEL. J. Merged eGFP-BobBIL4, RFP-HDEL, and bright field images. Scale bars = 50 μm (F-J).
Figure 4.
BobBIL4 expression patterns in Arabidopsis and subcellular localization. A-E. Tissue-specific expression of BobBIL4 in transgenic Arabidopsis. F-J. Subcellular localization of BobBIL4 in Nicotiana benthamiana leaf epidermal cells. Cells containing the empty vector pFGC-eGFP served as the control. F. eGFP image. G. Merged eGFP and bright field images. H. Subcellular localization of eGFP-BobBIL4. I. Endoplasmic reticulum marker RFP-HDEL. J. Merged eGFP-BobBIL4, RFP-HDEL, and bright field images. Scale bars = 50 μm (F-J).
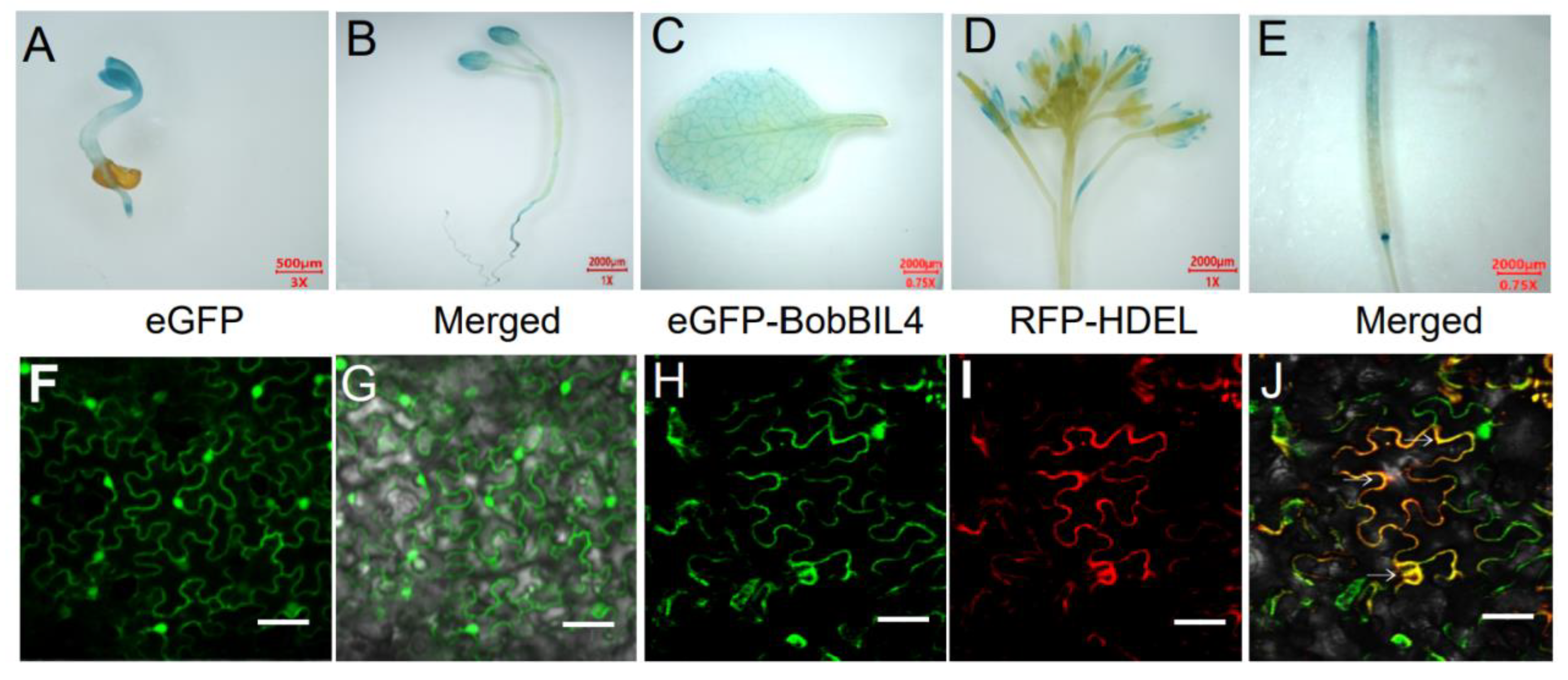
Figure 5.
BobBI-1 expression patterns following an exposure to an inducer or inhibitor of brassinolide stress as well as the involvement of BobBIL4 in the induction and inhibition of brassinolide stress. A. Roots were collected at 0, 2, 4, 8, and 12 h following the initiation of the 2 μM epibrassinolide (EBL) treatment. B-D. Images of wild-type, atbil4 mutant, and BobBIL4-overexpressing Arabidopsis seedlings grown on normal MS medium, MS medium containing 2 μM EBL, and MS medium containing 3 μM brassinazole (Brz) for 9 days. E. Root length. Error bars indicate mean ± SD (n = 15). Asterisks signify significant differences among samples (α = 0.05, Duncan's multiple range test). Scale bars = 1 cm (B-D).
Figure 5.
BobBI-1 expression patterns following an exposure to an inducer or inhibitor of brassinolide stress as well as the involvement of BobBIL4 in the induction and inhibition of brassinolide stress. A. Roots were collected at 0, 2, 4, 8, and 12 h following the initiation of the 2 μM epibrassinolide (EBL) treatment. B-D. Images of wild-type, atbil4 mutant, and BobBIL4-overexpressing Arabidopsis seedlings grown on normal MS medium, MS medium containing 2 μM EBL, and MS medium containing 3 μM brassinazole (Brz) for 9 days. E. Root length. Error bars indicate mean ± SD (n = 15). Asterisks signify significant differences among samples (α = 0.05, Duncan's multiple range test). Scale bars = 1 cm (B-D).
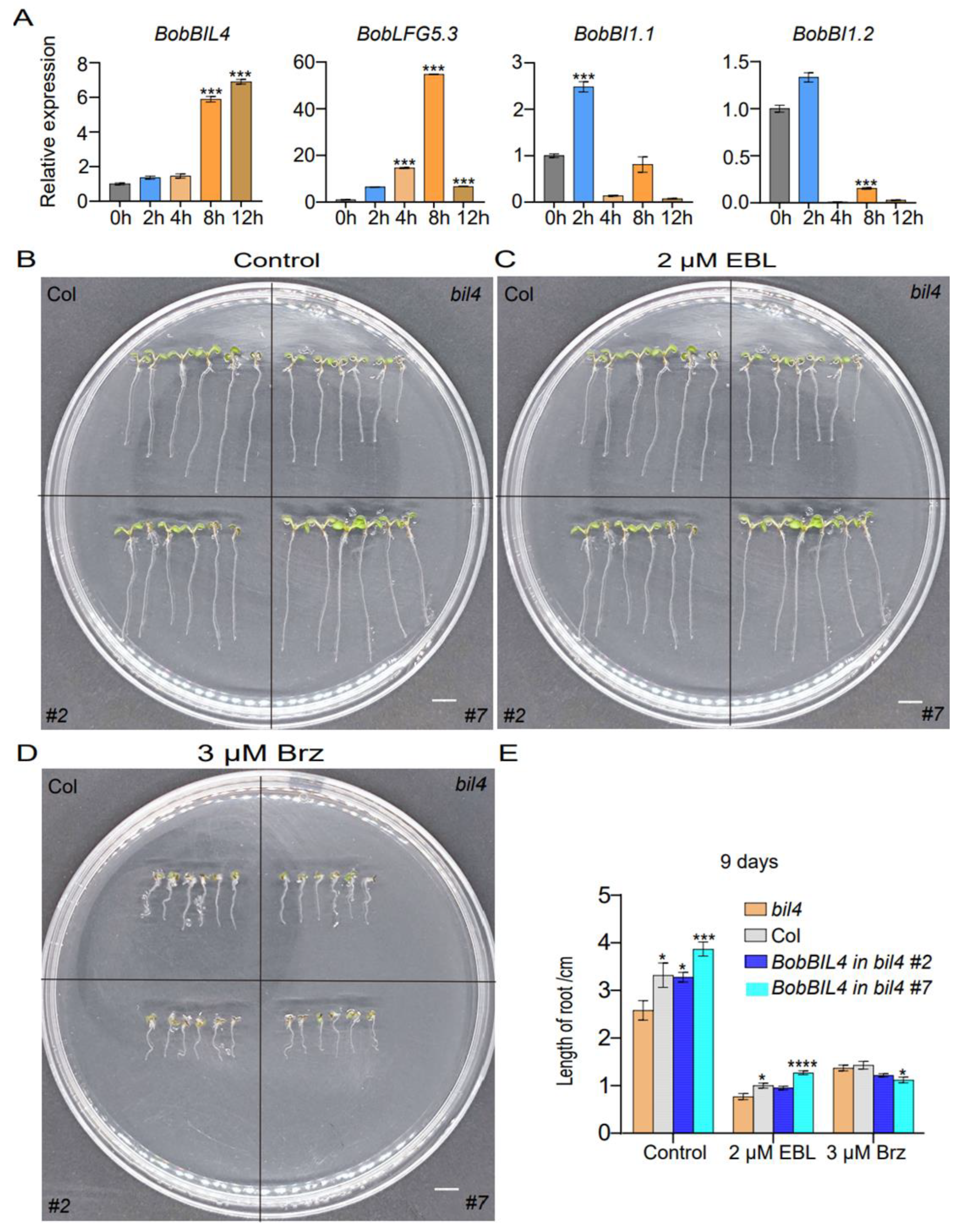
Figure 6.
BobBI-1 expression patterns following an exposure to NaCl stress as well as the involvement of BobBIL4 in the response to NaCl stress. A. Roots were collected at 0, 2, 4, 8 and 12 h following the initiation of the NaCl treatment. B. Images of wild-type, atbil4 mutant, and BobBIL4-overexpressing Arabidopsis seedlings grown on normal MS medium and MS medium containing 100 mM NaCl for 9 days. C. Root length. D. Root growth inhibition rate. Error bars indicate mean ± SD (n = 15). Asterisks signify significant differences among samples (α = 0.05, Duncan's multiple range test).
Figure 6.
BobBI-1 expression patterns following an exposure to NaCl stress as well as the involvement of BobBIL4 in the response to NaCl stress. A. Roots were collected at 0, 2, 4, 8 and 12 h following the initiation of the NaCl treatment. B. Images of wild-type, atbil4 mutant, and BobBIL4-overexpressing Arabidopsis seedlings grown on normal MS medium and MS medium containing 100 mM NaCl for 9 days. C. Root length. D. Root growth inhibition rate. Error bars indicate mean ± SD (n = 15). Asterisks signify significant differences among samples (α = 0.05, Duncan's multiple range test).
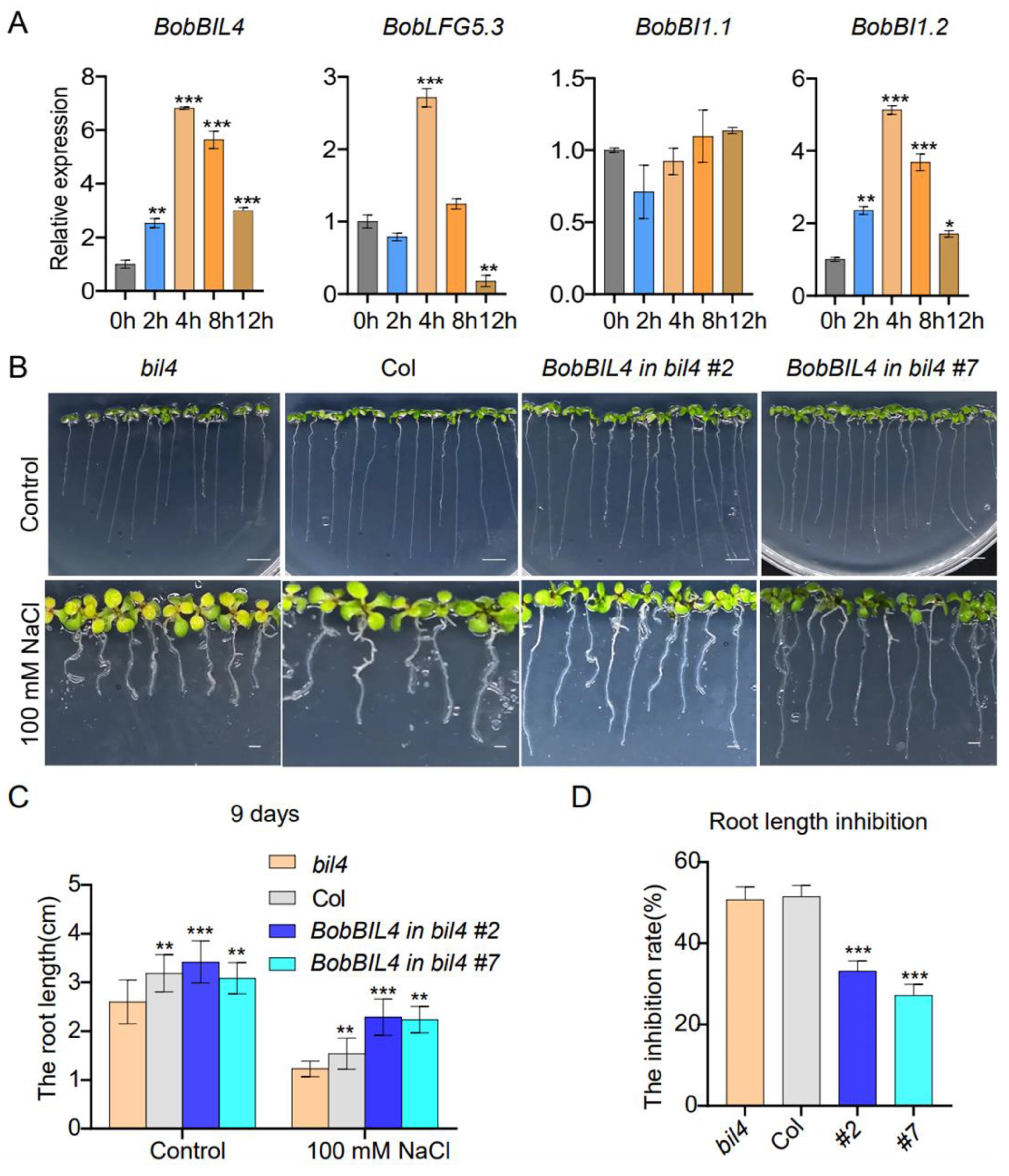

Table 1.
Biochemical properties of proteins encoded by BI-1 genes.
| Gene ID | Accession number | Amino Acid (bp) | Molecular weight (kDa) | pI | Instability index (II) | Aliphatic index | GRAVY | Subcellular Localization | Signal peptide |
|---|---|---|---|---|---|---|---|---|---|
| BobLFG1 | BolK_1g28270 | 277 | 30941.96 | 6.36 | 28.18 | 128.66 | 0.932 | GA | NO |
| BobLFG2 | BolK_4g29470 | 248 | 28079.38 | 9.05 | 26.49 | 125.77 | 0.831 | C | NO |
| BobLFG3 | BolK_2g39070 | 247 | 27526.34 | 8.94 | 46.95 | 107.77 | 0.474 | C | NO |
| BobLFG5.1 | BolK_7g47640 | 239 | 26980.26 | 7.67 | 24.72 | 130.50 | 0.971 | CM | NO |
| BobLFG5.2 | BolK_7g47660 | 248 | 27949.24 | 8.27 | 30.42 | 127.38 | 0.877 | GA | NO |
| BobLFG5.3 | BolK_8g15090 | 256 | 28111.26 | 5.88 | 26.37 | 128.28 | 0.753 | GA | NO |
| BobLFG4 | BolK_5g00210 | 256 | 28111.26 | 5.88 | 26.37 | 128.28 | 0.753 | GA | NO |
| BobBI1.1 | BolK_2g48750 | 256 | 28215.43 | 5.88 | 28.75 | 128.28 | 0.757 | GA | NO |
| BobBI1.2 | BolK_9g27570 | 247 | 27503.40 | 6.89 | 44.91 | 108.58 | 0.500 | CM, C, GA | NO |
1 GA, Golgi apparatus; C, chloroplast; CM, cell membrane.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



