
Submitted:
20 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
Somatic cell nuclear transfer (SCNT) has great potential in the replication of high commercial value animals, threatened wild species for conservation purposes, and transgenic animals for biomedical purposes. However, SCNT has a low success rate, due to intrinsic factors of the technique itself, which will lead to low rates of embryonic development and epigenetic alterations in the cloned embryos. The objective of the present study was to evaluate the effect of OF-UF on the intracellular concentration of ROS, GSH, and the development of Ovis aries cloned and parthenogenetic embryos. The results did not show a beneficial effect on the development of parthenogenetic and cloned embryos at concentrations of 0.5% OF-UF. Furthermore, at 1% OF-UF, an adverse effect was observed in cloned embryos at the blastocyst stage and 2% OF and UF in parthenogenetic embryos in the first divisions. A decrease in ROS and GSH levels was observed in the parthenogenetic blastocysts treated with 1% OF-UF, but not in the clones, in which a higher concentration of GSH and a similar concentration of ROS was observed. No effect was observed of OF-UF on embryonic development and redox balance on sheep clone embryos by Handmade Cloning.
Keywords:
Nuclear transfer
; Embryonic development
; Oviductal fluid
; Uterine fluid
; Reactive oxygen species
; Glutathione
1. Introduction
According to data from the International Embryo Technology Association (IETS), the in vitro production of sheep embryos increased more than 340% from 2021 to 2022 [1], being four productive species such as cattle, goats and horses, those that indicate growth and efficiency in the use of assisted reproduction techniques (ART). Within the ART, there is the SCNT the one that allows the multiplication of individuals with genetic characteristics, for example: animals of zootechnical interest of high commercial value [2], threatened wild species for conservation purposes [3,4] companion animals [5] and transgenic animals for biomedical purposes [6].
However, this biotechnology has not had a practical application like other ART, due to the small number of cloned embryos that reach term with the birth of healthy offspring. In sheep, efficiencies of 5.3 to 42% of cloned embryos reaching the blastocyst stage have been reported, while the efficiency in newborn cloned lambs per blastocyst transferred is 5.7 to 15% [7]. This is because of causes inherent to the technique, such as: trauma due to manipulation of the cells used, inadequate capacity of the recipient oocyte to reprogram the nucleus of the somatic cell, resistance to reprogramming by the somatic cells or abnormalities induced by the in vitro culture [8].
This leads to epigenetic alterations in embryos produced by SCNT such as elevated levels of DNA methylation in cloned embryos, compared to those produced by IVF or in vivo [9]. This has been observed in sheep, particularly in the trophectoderm cells in the blastocyst [10] that give rise to placentas with a reduced number or abnormalities in the placentomes and with histopathological alterations [11,12,13]. Newborn cloned lambs have also been described as having a high incidence of Abnormal Offspring Syndrome (AOS), due to the variety of reported phenotypes [14]. This is related to a deregulation of different imprinted genes [15,16,17].
In this context, it has been observed that by supplementing the embryo development media with OF and UF, or with the extracellular vesicles present in them, in bovines and pigs the quality of embryos produced in vitro [18,19] have a higher number of cells [20,21], lower levels of ROS and better survival after vitrification [21]. Furthermore, the epigenetic marks in them are more like those in embryos obtained in vivo [22,23].
Embryos produced by SCNT typically have higher levels of ROS than embryos produced by IVF [24,25,26], due to the excessive manipulation that the cells receive during enucleation, fusion and activation [27]. The imbalance between the generation of ROS and the antioxidant system of the embryo results in oxidative stress, which has a negative impact during embryonic development that can extend to fetal development [28].
Given that OF and UF contain elements that promote development and improve the quality of embryos produced in vitro, then supplementation of these fluids during the in vitro culture of Ovis aries cloned embryos may improve embryonic quality. Therefore, the aim of the present study was to evaluate the effect of OF and UF on the balance of ROS, GSH and the development of O. aries cloned and parthenogenetic embryos.
2. Materials and Methods
All reagents used were purchased from Sigma-Aldrich. The incubation conditions for cell cultures, in vitro oocyte maturation and in vitro embryonic development were: 38.5 °C, 5% CO2 and saturation humidity.
2.1. Obtaining Reproductive Fluids (OF and UF)
Reproductive tracts (RT) of adult Creole type sheep (O. aries) were collected from a local slaughterhouse and transported to the laboratory at 8 °C in disinfected plastic bags. Upon arrival, within no more than two hours, the RT were washed in sterile Dulbecco’s Phosphate Buffered Saline (DPBS) supplemented with 1% antibiotic-antimycotic (Streptomycin 10,000 µg mL-1, Amphotericin B 25 µg mL-1 and 10,000 IU of penicillin, In vitro S.A. de C.V. CDMX, México) at 4 °C. Subsequently, the RT were classified based on the ovarian structures present (to determine the phase of the estrous cycle), with regular size and shape, without cysts, according to the methodology described and adapted for sheep by Carrasco et al. [29]. The luteal phase was divided into early and middle, the early phase was determined by the presence of corpora hemorrhagicum (CH), while the middle phase was determined by the presence of developing corpora lutea (CL). Fluids were recovered from the oviduct and uterine horn ipsilateral to the ovary with the presence of CH and developing CL. Once the RT were classified, the suspensory ligaments were dissected, and the uterine-tubal junction was sectioned to separate the oviduct from the uterine horn. The oviducts and uterine horns were maintained in DPBS at 4 °C. For the extraction of the OF, the oviduct was placed on a flat surface and pressure was exerted with a slide from the utero-tubal junction towards the ampulla. A fire-polished Pasteur pipette was then introduced to recover the OF, which was deposited in a 300 µL tube. The sample was centrifuged at 2000 x g for 5 min at 4 °C, the supernatant was recovered and centrifuged at 7000 x g for 10 min at 4 °C. Finally, the supernatant was recovered and stored at -80 °C until use. To obtain the UF, the RT was placed vertically so that the fluid descended by gravity to the uterine-tubal junction. Immediately afterward, a fire-polished Pasteur pipette was introduced, and the fluid was collected. This was placed in a 300 µL tube, processed and stored in the same way as the OF.
2.2. Derive and Culture Skin Fibroblasts for Use as Karyoplasts
Ear skin fibroblasts were obtained according to Navarro-Maldonado et al. [30]. First, a sample of ear skin was obtained from an adult sheep (O. aries) and fibroblasts were derived by explant sowing and enzymatic disintegration, then they were cultured in DMEM medium (Dulbecco's Modified Eagle's Medium, In Vitro S.A. de C.V. CDMX, Mexico) supplemented with 2.3% antibiotic-antimycotic, 10% Newborn Calf Serum (NCS) and 13.6 mg mL-1 NaHCO3. Fibroblasts were taken to the 4th cell passage and cryopreserved at −196 °C. One week before SCNT, they were reseeded and left at confluence for at least three days so that they could reach the 5th passage and synchronize their cell cycle (G0/G1).
2.3. In Vitro Oocyte Maturation (IVM)
The cumulus-oocyte complexes (COCs) were obtained from the ovaries of slaughterhouse sheep, by aspiration of follicles of 2 to 8 mm in diameter, using 10 mL syringes with 20 G gauge needles. The aspiration medium was Tissue Culture Medium (TCM-199) with Hepes (In Vitro S. A. de C. V. CDMX, México), supplemented with 100 IU mL-1 of heparin. The collected follicular fluid was passed through a cell filter (100 μm, Biologix, Shandong, China) to recover the COCs and were selected based on their cytoplasmic morphology and the number of granulosa cell layers [31]. The selected COCs were cultured in 4-cell dishes (Nunc™) with 0.5 mL of in vitro maturation medium (IVM), based on TCM-199 (In Vitro S. A. de C. V. CDMX, México) supplemented with 1.675x10-04 g mL-1 of cysteine, 0.001 g mL-1 of polyvinyl alcohol, 5.496x10-04 g mL-1 of D-glucose, 1x10-04 g mL-1 of Sodium Pyruvate, 10% Newborn Calf Serum (NCS), 5 IU mL-1 of equine Chorionic Gonadotropin (eCG, Gonaforte Parfarm S.A., CDMX, Mexico), 0.1 IU mL-1 of recombinant Follicle stimulating hormone (FSH, Gonal-F Merck, Darmstadt, Germany), 10 ng mL-1 of Epidermal Grow Factor (EGF) and 0.65% antibiotic-antimycotic. The COCs were incubated for 21 h at 38.5 °C, 5% CO2 in air and saturation humidity.
2.4. Somatic Cell Nuclear Transfer (SNCT) by handmade cloning (HMC)
Once IVM was completed, SNCT by handmade cloning (HMC) was performed. To do this, the expanded cumulus cells of the COCs were disrupted with 0.5 mg mL-1 of hyaluronidase for 8 min incubation and by gentle pipetting. Oocytes were selected in MII (presence of the first polar body) and incubated in 0.5 µg mL-1 of demecolcine in MIV medium for 1 h to induce the cytoplasmic extrusion of the genetic material of the oocyte (the first polar body and the metaphase plate) [32,33].
2.4.1. Cytoplast Preparation
Subsequently, the zona pellucida (ZP) of the oocytes was removed with 2 mg mL-1 of pronase in 30 µL drops of TCM-199 medium with 4-(2-hydroxyethyl) piperazine-1-ethane-sulfonic acid (HEPES). Zona pellucida-free oocytes were manually enucleated with the use of a microblade (Professional Embryo Transfer Supply, Inc. Texas, USA), in 30 µL drops of TCM-199 with HEPES at 10% NCS (T10). For enucleation, the oocytes were placed orienting the extruded portion of their cytoplasm (the one that contained the genetic material) at the north pole, then the microblade was passed manually sectioning that portion of the cytoplasm. The enucleated oocytes (cytoplasts) were placed in 30 µL drops of TCM-199 with HEPES at 20% NCS (T20) for reconstitution, that is, until they return to their spherical shape, as a sign of viability [32,34].
2.4.2. Triplets’ Formation and Fusion
Groups of 5 cytoplasts were taken and immersed in 5 mg mL-1 of phytohemagglutinin in drops of TCM-199 medium with HEPES at 2% NCS (T2) for 3 sec, then pairs of cells were formed with the karyoplasts (karyoplast-cytoplast), dropping each cytoplast on a fibroblast (karyoplast). Then, the triplets (cytoplast-karyoplast-cytoplast) were formed and equilibrated in fusion medium (0.3 M of D-mannitol and 1 mg mL-1 of polyvinyl alcohol), aligned on the positive electrode of the fusion chamber (BTX microslide, 0.5 mm aperture, model 450, Massachusetts, USA). The triplets were fused by means of a single pulse of 1.0 kV cm-1 of direct current (DC) for 9 µs, then these were placed in drops of T20 for reconstitution to form a single cell (a cloned embryo). Meanwhile, the fusion process was repeated with the remaining cytoplasts. Once the triplets were reconstituted in cloned embryos, they were incubated in in vitro development medium (IVD) for 1.5 h to promote their nuclear reprogramming [34].
2.4.3. Activation of Cloned Embryos
Cloned embryos were activated with 8 µg mL-1 of Ca2+ ionophore A23187 in TCM-199 at 2% HEPES NCS (T2) for 5 min and then in 2 mM 6-DMAP in IVD medium (Cleavage, Cook Medical, Bloomington, USA) for 4 h, under the same incubation conditions.
2.4.4. In Vitro Culture of Cloned Embryos in the WOW System
According to Vajta et al. [35], cloned embryos were cultured in the Well of the Well (WOW) system, which consisted of making microwells with a punch inside a cell of a 4-cell box (Nunc™) placing a single-cloned embryo in each microwell. They were cultured for 4 days in IVD Cleavage medium (Cook Medical, Bloomington, USA) and subsequently in Blastocyst medium (Cook Medical, Bloomington, USA) for three more days until they reached the blastocyst stage on day seven. The experimental groups were worked separately and consisted of supplementing the IVD media with OF (for Cleavage medium) and with UF (for Blastocyst medium). Treatments consisted of 0.5, 1.0, and 2.0% OF and UF.
2.5. In Vitro Production of Parthenogenetic Embryos
After IVM, oocytes in MII were selected and activated following the same method described for cloned embryos. They were cultured in Cleavage medium for 4 days and later in Blastocyst medium for three more days, for a total of seven days. The experimental groups were worked separately and were supplemented the same as for the cloned embryos, 0.5, 1.0 and 2.0% of OF and UF.
2.6. Evaluation of ROS and GSH Levels in Parthenogenetic and Cloned Embryos
According to Nadri et al. [26], ROS and GSH levels were evaluated in O. aries parthenogenetic cloned embryos at the blastocyst stage (7 days IVD), which were incubated with H2DCFDA (2',7'-dichlorodihydrofluorescein diacetate, Invitrogen, Eugene Oregon, USA) to determine the presence of ROS and with Cell Tracker Blue™ (Gibco™) to determine the presence of GSH. To do this, blastocysts of each group were washed in DPBS with 1 mg mL-1 of polyvinyl alcohol and then incubated with 10 mM H2DCFDA and 10 mM Cell Tracker Blue™ at 38.5 °C for 30 min, protected from light. Embryos were fixed with 4% paraformaldehyde and mounted on slides, covered with coverslips and sealed with nail polish. They were subsequently evaluated in epifluorescence microscopy using 460 nm UV filters to evaluate ROS and 405 nm UV filters to evaluate GSH. The taken images were analyzed with the Image J program (version 1.53; Wayne Rasband, National Institute of Health, USA).
2.7. Statistic Analysis
Data were expressed as mean ± standard error. The comparison between the means of the different stages of embryo development, as well as the mean of the relative fluorescence intensity of ROS and GSH were analyzed by a one-way ANOVA test. To evaluate the possible relationship between ROS and GSH, the Spearman correlation test was used. The differences were considered significant at P<0.05. For statistical analysis, the GraphPad Prism program (version 9.5., Boston, USA) was used.
3. Results
3.1. Effect of FO and FU on the In Vitro Development of Parthenogenetic Embryos
No significant differences were observed in development of parthenogenetic embryos treated with 0.5 and 1.0% of OF and UF (P>0.05), when compared to the control group (Table 1). Although it was observed that there was a higher rate of 4 to16- cells embryos at 2% of OF and UF when compared to the control group (100 vs. 31.8%, respectively, P<0.05), these embryos did not continue their development (Table 1). For this reason, this concentration was not considered for O. aries cloned embryos.
3.2. Effect of OF and UF on the In Vitro Development of Cloned Embryos
No differences were observed in the development of cloned embryos treated with 0.5% OF and UF when compared to the control group (P>0.05, Table 2). An increase in the 4 to 16- cells rate was observed in embryos treated with 1.0% OF and UF, when compared to the control group (62.1 vs 27.9%, respectively, P<0.05, Table 2). However, a decrease in the morula and blastocyst rates was observed vs. the control group (27.9 vs 47.3 and 7.7 vs. 28.1%, respectively; P<0.05, Table 2).
3.3. Effect of OF and UF on the Presence and Levels of ROS and GSH in Parthenogenetic Embryos
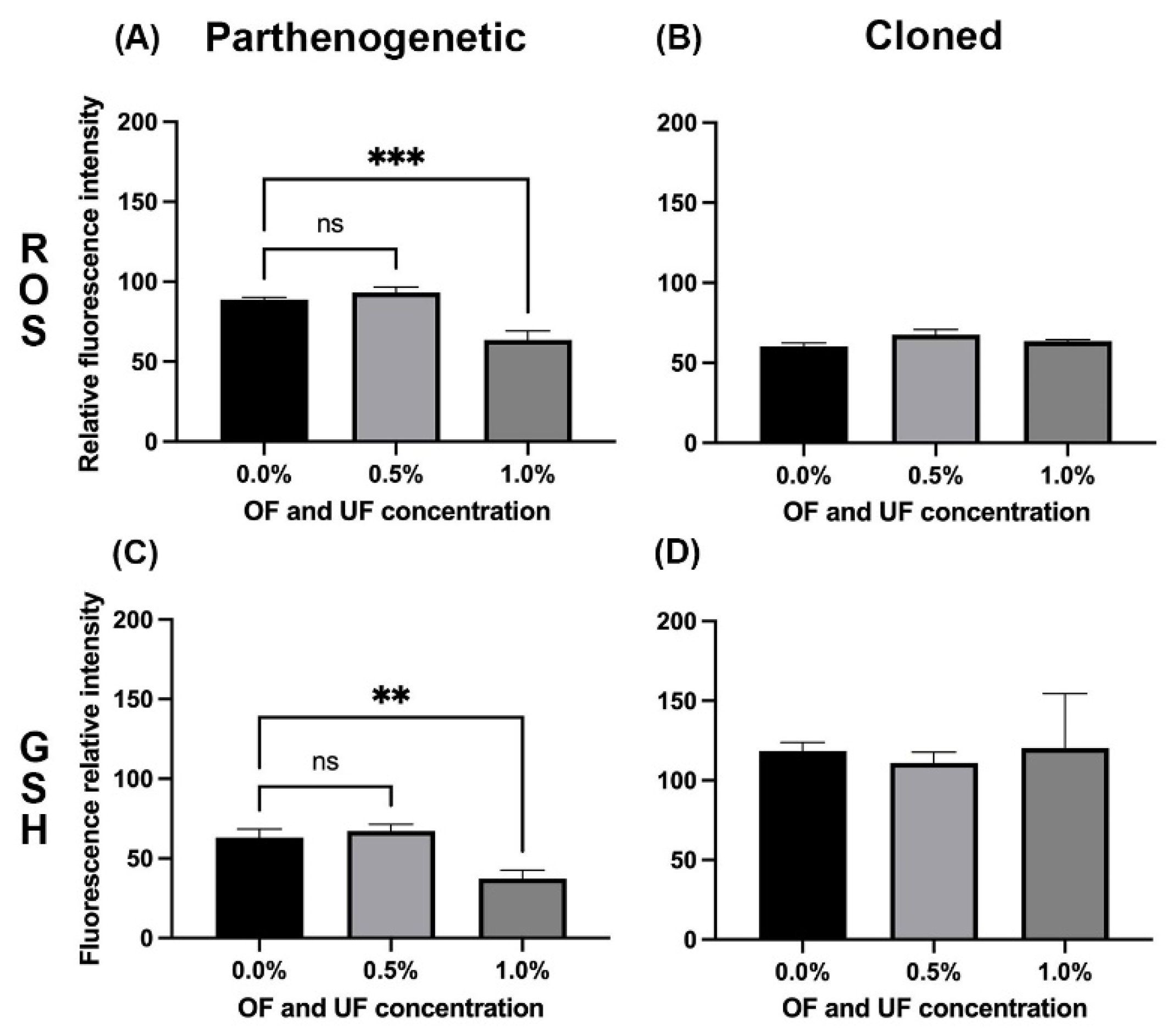
Figure 1 shows parthenogenetic blastocyst stained to determine ROS and intracellular GSH. In the parthenogenetic blastocysts treated with 0.5% OF and UF, no significant differences were observed in ROS and GSH levels with respect to the control group (Figure 3A and Figure 3C).
At 1.0% OF and UF, ROS and GSH levels decreased significantly compared to the control group (P<0.05) (Figure 3A and Figure 3C). At this same concentration, both variables presented a directly proportional positive correlation (r = 0.8727; P<0.001), that is, as ROS levels increased, GSH levels also increased and vice versa.
At 2.0% OF and UF, there was no development into blastocysts.
3.4. Effect of OF and UF on the Presence and Levels of ROS and GSH in Cloned Embryos
Figure 2 shows cloned blastocyst stained to determine ROS and intracellular GSH. No effect was observed on the levels of ROS and GSH for blastocysts treated with 0.5 and 1% OF and UF, with respect to the control group (Figure 3B and Figure 3D).
The levels of ROS and GSH presented blastocysts in the control group showed a negative, inversely proportional correlation (r = -0.8095; P<0.022), that is, as GSH levels increased, ROS levels decreased.
Figure 2.
Effect of OF and UF on the presence of ROS and GSH in cloned embryos. Panel photographs (light and dark fields) of embryos in the blastocyst stage are observed. Presence of ROS in green. Presence of GSH in blue. 20X Magnification. Scale bar 200 μm.
Figure 2.
Effect of OF and UF on the presence of ROS and GSH in cloned embryos. Panel photographs (light and dark fields) of embryos in the blastocyst stage are observed. Presence of ROS in green. Presence of GSH in blue. 20X Magnification. Scale bar 200 μm.
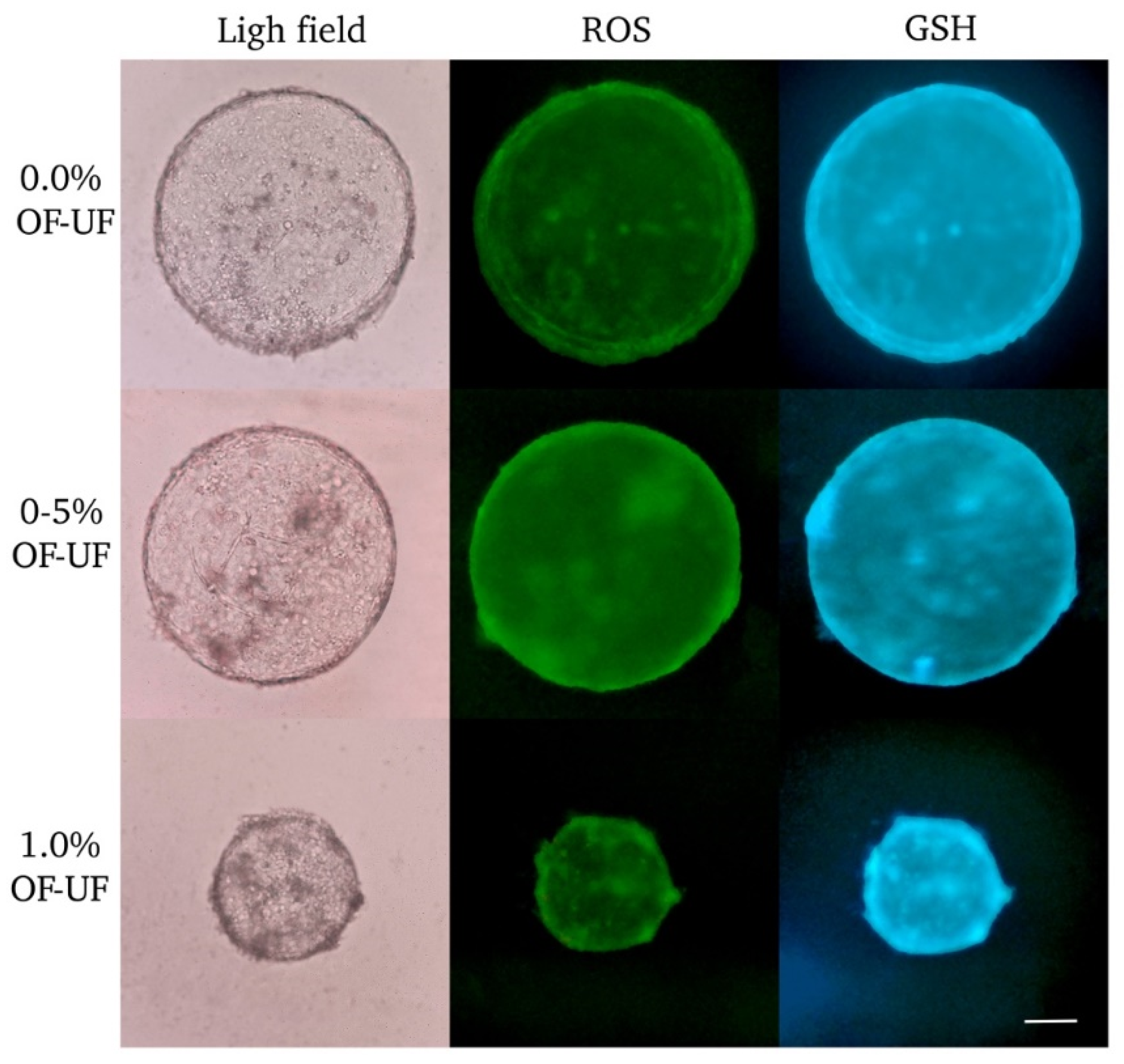
Figure 3.
Effect of OF and UF on ROS and GSH levels in O. aries parthenogenetic and cloned embryos at the blastocyst stage. A, C) Relative fluorescence intensity of ROS and GSH in parthenogenetic blastocysts. B, D) Relative fluorescence intensity of ROS and GSH in cloned blastocysts. (Image J). Bars represent mean ± S.E. ***P = 0.0009, **P = 0.0032.
Figure 3.
Effect of OF and UF on ROS and GSH levels in O. aries parthenogenetic and cloned embryos at the blastocyst stage. A, C) Relative fluorescence intensity of ROS and GSH in parthenogenetic blastocysts. B, D) Relative fluorescence intensity of ROS and GSH in cloned blastocysts. (Image J). Bars represent mean ± S.E. ***P = 0.0009, **P = 0.0032.
4. Discussion
Somatic cell nuclear transfer (SCNT) has great potential in the propagation of animals with genetic and phenotypic characteristics of productive, ecological or biomedical interest. However, its practical application has been limited due to intrinsic factors of the technique itself [8] that promote alterations at the genetic and epigenetic level, at the beginning of the preimplantation embryo development and to the fetal development. In some cases, and, if carried to term, clonal offspring may present AOS [14].
This is a novel study because, given the positive effect of OF and UF on the development and quality of embryos produced by IVF [21,22,23]. For the first time it was proposed to analyze its effect on the in vitro development of sheep-cloned embryos produced by handmade cloning (HMC), to counteract the incidence of these syndromes in cloned offspring. Interestingly, it was observed that in cloned and parthenogenetic sheep embryos, there were no differences in the segmentation rate at the evaluated concentrations of OF and UF vs. the control group. This agrees with what has been reported for bovine embryos generated by IVF [20,21,22,36]. At the lowest concentration (0.5%), OF and UF showed no effect on the development of cloned nor parthenogenetic embryos. However, at 1.0%, OF and UF reduced the morula and blastocyst rates in sheep-cloned embryos. An exacerbated effect was observed at 2.0% of OF and UF in parthenogenetic embryos, which were blocked at the 4 to 16- cell stage. In pig-cloned embryo, it was reported that supplementation with 14 and 28 μg mL-1 of OF in PZM-5 medium significantly increased the blastocyst rate (27 and 26%, respectively) with respect to the control group (14%). But at higher levels (56 and 100 μg mL-1) it had no effect (18.1 and 19.2%) [37]. Unlike these authors who supplemented OF in μg mL-1, in the present study levels were evaluated as percentage. The difference in results between both studies could be due to the compounds generated by the metabolism of embryos and the degradation of carbohydrates, amino acids and proteins in the culture medium, which could accumulate until reaching toxic levels. Therefore, it is essential to maintain an ideal concentration of reproductive fluids that benefit embryonic development without generating high levels of toxic compounds for embryos.
Other studies found that high levels of OF (5, 10 and 25%) or UF (2.5 and 5%) have a negative effect on the blastocyst rate in embryos produced by IVF (11, 10 and 1% for FO; 13.2 and 3% for FU, respectively) [20,21]. Data similar to those observed in the present study for cloned embryos at 1% OF and UF, and for parthenogenetic embryos that blocked their development at cleavage at 2.0% OF and UF.
Lopera-Vasquez et al. [20] supplemented SOF medium with 0.625, 1.25 and 2.5% OF for bovine embryos produced by IVF, observing that on day 7 the blastocyst rate was significantly lower at all OF concentrations (16, 17 and 13.9%, respectively) with respect to the control group (22.9%). However, on day 9 the blastocyst rate was not different from the control group at 0.625 and 1.25% of OF, like the present study for blastocyst rate of cloned embryos treated with 1.0% OF and UF on day 7 of culture.
The effect that reproductive fluids have on the in vitro development of embryos depends on their characteristics conditioned by variables intrinsic to the origin of the fluids, whether if they are collected in vivo or Postmortem, the recovery method: by mechanical pressure with forceps, by compression with a slide, by aspiration with an automatic pipette, by scraping with a curette or by washing with physiological solution [38,39]. Depending on the recovery method, the fluids may contain a greater or lesser number of impurities such as: erythrocytes, oviductal cells and cellular debris that are not always efficiently eliminated by centrifugation [38]. Another variable to consider is whether the fluids are pure and concentrated, or diluted, which is important since most studies carry out a supplementation (v/v), without considering the concentration of its components which can vary from one to another sample and may not be reproducible. The best to do would be the supplementation of these fluids based on their protein concentration (μg mL-1), which are the main molecules that constitute them that culture media lack.
Regarding the levels of ROS and GSH present in sheep parthenogenetic embryos, in the present study it was observed that at 1.0% of OF and UF, the levels of both decreased significantly in blastocysts, results like that reported in other studies, where ROS concentrations, as well as the expression of genes related to oxidative stress, decreased in bovine embryos produced by IVF, treated with OF and UF [21,40]. In the proteome of sheep-reproductive system, 940 proteins have been identified, 4% related to oxidation-reduction processes [41] and 5% to oxidative stress [42]. Some of the most abundant proteins in OF in the luteal phase are ceruloplasmin and lactotransferrin, in UF they are peroxiredoxin-1 and glutathione S-transferase [41,42]. These proteins are part of the non-enzymatic antioxidant system of reproductive fluids and were probably responsible for lowering intracellular ROS levels in parthenogenetic embryos, that is why these embryos did not need to increase intracellular GSH to counteract ROS. Hence, by adding 1.0% of OF and UF, these levels will decrease.
For cloned embryos, no effect on levels of ROS and GSH was observed for the different OF and UF treatments. Likewise, ROS levels were similar between cloned and parthenogenetic embryos, which is noteworthy, given the intense manipulation to which the former are subjected that causes exposure to high concentrations of oxygen, changes in temperature and prolonged exposure to light [28,43]. The opposite was observed for GSH, which levels were different between both types of embryos, being about 30% more concentrated in cloned embryos than in parthenogenetic ones. Possibly this was because to form each cloned embryo, two cytoplasts were used, which retained around 70% of the initial volume of cytoplasm after enucleation; therefore, each cloned embryo had about 40% more cytoplasm than the parthenogenetic embryos, which increased the intracellular concentration of GSH and possibly this kept the ROS levels in balance in the cloned embryos [44].
It has been described that increasing the cytoplasmic volume of cloned embryos improves their quality and implantation potential, due to the addition of mitochondria and other cytoplasmic factors that help nuclear reprogramming [45,46,47], but its effect on intracellular GSH levels has not been described.
Glutathione (GSH) is a non-enzymatic antioxidant synthesized mainly in the cytosol (80-85%) and mitochondria (10-15%), and in lesser quantities in the nucleus and endoplasmic reticulum. Its antioxidant action can be direct or as a cofactor of antioxidant and detoxifying enzymes, it acts on various free radicals and pro-oxidants such as hydrogen peroxide, which is a precursor of ROS [48].
The present study is a pioneer in evaluating the effect of OF and UF on the development rate and quality of sheep cloned embryos, produced by HMC. Still, due to the characteristics of this technique, where cloned embryos lack ZP and must be cultured in a Well of the Well (WOW) system, it made complicated the sequential supplementation with OF and UF.
The study allowed to know about some effects that OF and UF have on the development rate in sheep cloned embryos and provides the possibility for further research about the impact they may have on the quality of cloned embryos, in terms of total cell number, gene expression and epigenetic regulation.
5. Conclusions
In the present study, no effect on the development rate of Ovis aries cloned vs. parthenogenetic embryos was observed when supplementing culture media with 0.5% of OF and UF. Moreover, higher levels had negative effects since the first divisions of the in vitro development in parthenogenetic embryos, and in the blastocyst stage for cloned embryos. Still, a positive effect on redox regulation was observed at 1% OF in parthenogenetic embryos, but not in cloned embryos. We concluded that supplementation with OF and UF did not improved the in vitro development nor the quality of Ovis aries cloned embryos produced by HMC.
Author Contributions
Conceptualization, J.R.V.-A; methodology, J.R.V.-A.; J.L.R.-S., J.E.H.-P.; software, J.R.V.-A.; validation, M.d.C.N.-M., D.A.A.-G., C.C.-R.; formal analysis, J.R.V.-A.; investigation, J.R.V.-A. M.d.C.N.-M., D.A.A.-G., C.C.-R; resources, D.A.A.-G., J.E.H.-P. and M.d.C.N.-M.; data curation, J.R.V.-A.; writing-original draft preparation, J.R.V.-A.; writing-review and editing, M.d.C.N.-M., D.A.A.-G., C.C.-R. J.L.R.-S., J.E.H.-P.; supervision, M.d.C.N.-M.; project administration, M.d.C.N.-M. and D.A.A.-G., funding acquisition, M.d.C.N.-M. and D.A.A.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Consejo Nacional de Humanidades, Ciencia y Tecnología CONAHCyT (Grant No.: C-/2024 459).
Institutional Review Board Statement
Ethical review and approval were waived for this study due the fact that institutional codes of ethics were not affected, as sheep ovaries as well as skin ear biopsy were collected from slaughterhouses.
Data Availability Statement
Data are available from first author José Roberto Vazquez-Avendaño (robertmizer@gmail.com) upon request.
Acknowledgments
The authors would like to thanks Dr. Miriam Fahiel Casillas Ávalos from the Department of Biology of Reproduction and Dr. Miguel Betancourt Rule from the Department of Health Sciences, Division of Biological and Health Sciences at the Universidad Autónoma Metropolitana Unidad Iztapalapa, for their support in the evaluation of the presence and levels of ROS and GSH in O. aries cloned embryos. Also, to CONAHCyT for the scholarship for postgraduate studies awarded to the main author with Registration Number 763618.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Viana, J.H.M. Statistics of embryo production and transfer in domestic farm animals. In: Embryo Technology Newsletter, 2022; 2021, 40 https://www.iets.org/Portals/0/Documents/Public/Committees/DRC/IETS_Data_Retrieval_Report_2021.pdf [accessed on 10 April 2024].
- Galli, C.; Lazzari, G. 25th Anniversary of cloning by somatic-cell nuclear transfer: Current applications of SCNT in advanced breeding and genome editing in livestock. Reproduction. 2021, 162, F23-F32. [CrossRef]
- Borges, A.A.; Pereira, A.F. Potential role of intraspecific and interspecific cloning in the conservation of wild mammals. Zygote. 2019, 27, 111-117. [CrossRef]
- Bolton, R.L.; Mooney, A.; Pettit, M.T.; Bolton, A.E.; Morgan, L.; Drake, G.J.; Appeltant, R.; Walker, S.L.; Gillis, J.D.; Hvilsom, C. Resurrecting biodiversity: advanced assisted reproductive technologies and biobanking. Reprod Fertil. 2022, 3, R121-R146. [CrossRef]
- Loi, P.; Palazzese, L.; Scapolo, P.A.; Fulka, J.; Fulka, H.; Czernik, M. 25th Anniversary of cloning by somatic-cell nuclear transfer: Scientific and technological approaches to improve SCNT efficiency in farm animals and pets. Reproduction. 2021, 162, F33-F43. [CrossRef]
- Kalds, P.; Gao, Y.; Zhou, S.; Cai, B.; Huang, X.; Wang, X.; Chen, Y. Redesigning small ruminant genomes with CRISPR toolkit: Overview and perspectives. Theriogenology. 2020, 147, 25-33. [CrossRef]
- Vazquez-Avendaño, J.R.; Ambríz-Garcia, D.A.; Cortez-Romero, C.; Trejo-Cordova, A.; Navarro-Maldonado, M.C.; Current state of the efficiency of sheep embryo production through somatic cell nuclear transfer. Small Rumin Res. 2022, 212, 106702. [CrossRef]
- Simmet, K.; Wolf, E.; Zakhartchenko, V.; Manipulating the Epigenome in Nuclear Transfer Cloning: Where, When and How. Int J Mol Sci. 2020, 22, 236. [CrossRef]
- Sawai, K.; Takahashi, M.; Fujii, T.; Moriyasu, S.; Hirayama, H.; Minamihashi, A.; Hashizume, T.; Onoe, S. DNA methylation status of bovine blastocyst embryos obtained from various procedures. J Reprod Dev. 2011, 57, 236-41. [CrossRef]
- Beaujean, N.; Taylor, J.; Gardner, J.; Wilmut, I.; Meehan, R.; Young, L. Effect of limited DNA methylation reprogramming in the normal sheep embryo on somatic cell nuclear transfer. Biol Reprod, 2004, 71, 185–193. [CrossRef]
- Loi, P.; Clinton, M.; Vackova, I.; Fulka, J.; Jr, Feil, R.; Palmieri, C.; Della Salda, L.; Ptak, G. Placental abnormalities associated with post-natal mortality in sheep somatic cell clones. Theriogenology. 2006, 65, 1110-21. [CrossRef]
- Palmieri, C.; Loi, P.; Ptak, G.; Della Salda, L. Review paper: a review of the pathology of abnormal placentae of somatic cell nuclear transfer clone pregnancies in cattle, sheep, and mice. Vet Pathol. 2008, 45, 865-80. [CrossRef]
- Ni, W.; You, S.; Cao, Y.; Li, C.; Wei, J.; Wang, D.; Qiao, J.; Zhao, X.; Hu, S.; Quan, R. Aberrant expression of miR-127, miR-21 and miR-16 in placentas of deceased cloned sheep. Res Vet Sci. 2016, 105, 200-4. [CrossRef]
- Nava-Trujillo, H.; Rivera, R.M. Review: Large offspring syndrome in ruminants: current status and prediction during pregnancy. Animal. 2023, 17, Suppl 1, 100740. [CrossRef]
- Young, L.E.; Schnieke, A.E.; McCreath, K.J.; Wieckowski, S.; Konfortova, G.; Fernandes, K.; Ptak, G.; Kind, A. J.; Wilmut, I.; Loi, P.; Feil, R. Conservation of IGF2-H19 and IGF2R imprinting in sheep: effects of somatic cell nuclear transfer. Mech Dev. 2003, 120, 1433-42. [CrossRef]
- Wang, F.; Pan, J.; Zhao, L.X.; Liu, Y.Y.; Zhang, L.; Wang, S.Y.; Li, L.; Zhou, H. M.; Zhang, D. Discovery of DNA Methylation Status of Peg3, Cdkn1c and Gtl2 in Cloned and Natural Lambs. Prog Biochem Biophys. 2016, 43, 706-715. [CrossRef]
- Luo, J.; Zhang, Y.; Guo, Y.; Tang, H.; Wei, H.; Liu, S.; Wang, X.; Wang, L.; Zhou, P. TRIM28 regulates Igf2-H19 and Dlk1-Gtl2 imprinting by distinct mechanisms during sheep fibroblast proliferation. Gene. 2017, 637, 152-160. [CrossRef]
- Cajas, Y.N.; Cañón-Beltrán, K.; de la Blanca, M.G.M.; Sánchez, J.M.; Fernandez-Fuertes, B.; González, E.M.; Rizos, D. Role of reproductive fluids and extracellular vesicles in embryo–maternal interaction during early pregnancy in cattle. Reprod Fertil Dev. 2021, 34, 117-138. [CrossRef]
- Coy, P.; Romar, R.; Romero-Aguirregomezcorta, J. The embryo culture media in the era of epigenetics: is it time to go back to nature? Anim Reprod. 2022, 19, e20210132. [CrossRef]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Gutierrez-Adan, A.; Bermejo-Alvarez, P.; Ramírez, M.Á.; Yáñez-Mó, M.; Rizos, D. Effect of bovine oviductal extracellular vesicles on embryo development and quality in vitro. Reproduction. 2017, 153, 461-470. [CrossRef]
- Hamdi, M.; Lopera-Vasquez, R.; Maillo, V.; Sanchez-Calabuig, M.J.; Núnez, C.; Gutierrez-Adan, A.; Rizos, D. Bovine oviductal and uterine fluid support in vitro embryo development. Reprod Fertil Dev. 2018, 30, 935-945. [CrossRef]
- Barrera, A.D.; García, E.V.; Hamdi, M.; Sánchez-Calabuig, M.J.; López-Cardona, Á.P.; Balvís, N.F.; Rizos, D.; Gutiérrez-Adán, A. Embryo culture in presence of oviductal fluid induces DNA methylation changes in bovine blastocysts. Reproduction. 2017, 154, 1-12. [CrossRef]
- Canovas, S.; Ivanova, E.; Romar, R.; García-Martínez, S.; Soriano-Úbeda, C.; García-Vázquez, F.A.; Saadeh, H.; Andrews, S.; Kelsey, G.; Coy, P. DNA methylation and gene expression changes derived from assisted reproductive technologies can be decreased by reproductive fluids. Elife. 2017, 6, e23670. [CrossRef]
- You, J.; Kim, J.; Lim, J.; Lee, E. Anthocyanin stimulates in vitro development of cloned pig embryos by increasing the intracellular glutathione level and inhibiting reactive oxygen species. Theriogenology. 2010, 74, 777-85. [CrossRef]
- Su, J.; Wang, Y.; Xing, X.; Zhang, L.; Sun, H.; Zhang, Y. Melatonin significantly improves the developmental competence of bovine somatic cell nuclear transfer embryos. J Pineal Res. 2015, 59, 455-68. [CrossRef]
- [26] Nadri, P.; Ansari-Mahyari, S.; Jafarpour, F.; Mahdavi, A.H.; Tanhaei Vash, N.; Lachinani, L.; Dormiani, K.; Nasr-Esfahani, M. H. Melatonin accelerates the developmental competence and telomere elongation in ovine SCNT embryos. PLoS One. 2022, 17, e0267598. [CrossRef]
- Koo, O.J.; Jang, G.; Kwon, D.K.; Kang, J.T.; Kwon, O.S.; Park, H.J.; H. J.; Kang, S. K.; Lee, B. C. Electrical activation induces reactive oxygen species in porcine embryos. Theriogenology. 2008, 70, 1111-8. [CrossRef]
- Deluao, J.C.; Winstanley, Y.; Robker, R.L.; Pacella-Ince, L.; Gonzalez, M.B.; McPherson, N.O. Oxidative stress and reproductive function: Reactive oxygen species in the mammalian pre-implantation embryo. Reproduction. 2022, 164, F95-F108. [CrossRef]
- Carrasco, L.C.; Coy, P.; Avilés, M.; Gadea, J.; Romar, R. Glycosidase determination in bovine oviducal fluid at the follicular and luteal phases of the oestrous cycle. Reprod Fertil Dev. 2008, 20, 808-17. [CrossRef]
- Navarro-Maldonado, M.C.; Hernández-Martínez, S.; Vázquez-Avendaño, J.R.; Martínez-Ibarra, J.L.; Zavala-Vega, N.L.; Vargas-Miranda, B.; Rivera-Rebolledo J.A.; Ambríz-García D.A. Deriva de células epiteliales de tejido de piel descongelado de Ovis canadensis mexicana para la formación de un banco de germoplasma. Acta Zool. Mex. (n.s.). 2015, 31, 275-282. [CrossRef]
- Ward, F. A., Lonergan, P., Enright, B. P., Boland, M. P. (2000) Factor affecting recovery and quality of oocytes for bovine embryo production in vitro using ovum pick-up technology, Theriogenology, 54(3): 433-446. [CrossRef]
- Vazquez-Avendaño, J.R.; Hernández-Martínez, S.; Hernández-Pichardo, J.E.; Rivera-Rebolledo, J.A.; Ambríz-García, D.A.; Navarro-Maldonado, M.C. Efecto del uso de medio secuencial humano en la producción de blastocistos de hembra Ovis canadensis mexicana por clonación manual. Acta Zool. Mex. (n.s.). 2017, 33, 328-338. [CrossRef]
- Hernández Martínez, S.; Hernández Pichardo, J.E.; Vazquez Avendaño, J.R.; Ambríz García, D.A.; Navarro Maldonado, M.C. Developmental dynamics of cloned Mexican bighorn sheep embryos using morphological quality standards. Vet Med Sci, 2020, 6, 382–392. [CrossRef]
- Vajta, G.; Lewis, I.M.; Hyttel, P.; Thouas, G.A.; Trounson, A.O. Somatic cell cloning without micromanipulators. Cloning. 2001, 3, 89-95. [CrossRef]
- Vajta, G.; Korösi, T.; Du, Y.; Nakata, K.; Ieda, S.; Kuwayama, M.; Nagy.; Z.P. The Well-of-the-Well system: an efficient approach to improve embryo development. Reprod Biomed Online. 2008, 17, 73-81. [CrossRef]
- Nina, M.; Ayala, C.; Susaño, R. Fluido uterino de llama (Lama glama), como medio para potenciar el desarrollo embrionario de vacas (Bos taurus) en cultivos in vitro. RIIARn. 2021, 8, 138-145. [CrossRef]
- Zhang, Y.H.; Song, E.S.; Kim, E.S.; Cong, P.Q.; Lee, S.; Lee, J.I.; Yi, Y.J.; Park, C.S. Effects of Oviductal Fluid, Culture Media and Zona Pellucida Removal on the Development of Porcine Embryos by Nuclear Transfer. Asian-australas J Anim Sci. 2009, 22, 962-968. [CrossRef]
- Velazquez, M.A.; Parrilla, I.; Van Soom, A.; Verberckmoes, S.; Kues, W.; Niemann, H. Sampling techniques for oviductal and uterine luminal fluid in cattle. Theriogenology. 2010, 73, 758-67. [CrossRef]
- Itze-Mayrhofer, C.; Brem, G. Quantitative proteomic strategies to study reproduction in farm animals: Female reproductive fluids. J Proteomics. 2020, 225, 103884. [CrossRef]
- Cebrian-Serrano, A.; Salvador, I.; García-Roselló, E.; Pericuesta, E.; Pérez-Cerezales, S.; Gutierrez-Adán, A.; Coy, P.; Silvestre, M. A. Effect of the bovine oviductal fluid on in vitro fertilization, development and gene expression of in vitro-produced bovine blastocysts. Reprod Domest Anim. 2013, 48, 331-8. [CrossRef]
- Soleilhavoup, C.; Riou, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Kohnke, P.; Reynaud, K.; de Graaf, S.P.; Gerard, N.; Druart, X. Proteomes of the Female Genital Tract During the Oestrous Cycle. Mol Cell Proteomics. 2016, 15, 93-108. [CrossRef]
- Koch, J.M.; Ramadoss, J.; Magness, R.R. Proteomic profile of uterine luminal fluid from early pregnant ewes. J Proteome Res. 2010, 9, 3878-85. [CrossRef]
- Soto-Heras, S.; Paramio, M.T. Impact of oxidative stress on oocyte competence for in vitro embryo production programs. Res Vet Sci. 2020, 132, 342-350. [CrossRef]
- Liu, R.H.; Li, Y.H.; Jiao, L.H.; Wang, X.N.; Wang, H.; Wang, W.H. Extracellular and intracellular factors affecting nuclear and cytoplasmic maturation of porcine oocytes collected from different sizes of follicles. Zygote. 2002, 10, 253-60. [CrossRef]
- Panda, S.K.; George, A.; Saha, A.P.; Sharma, R.; Manik, R.S.; Chauhan, M.S.; Palta, P.; Singla, S.K. Effect of cytoplasmic volume on developmental competence of buffalo (Bubalus bubalis) embryos produced through hand-made cloning. Cell Reprogram. 2011, 13, 257-62. [CrossRef]
- Liu, X.; Luo, C.; Deng, K.; Wu, Z.; Wei, Y.; Jiang, J.; Lu, F.; Shi, D. Cytoplasmic volume of recipient oocytes affects the nucleus reprogramming and the developmental competence of HMC buffalo (Bubalus bubalis) embryos. J Vet Med Sci. 2018, 80, 1291-1300. [CrossRef]
- Raja, A.K.; Sahare, A.A.; Jyotsana, B.; Priya, D.; Palta, P.; Chauhan, M.S.; Manik, R.S.; Singla, S.K. Reducing the cytoplasmic volume during hand-made cloning adversely affects the developmental competence and quality, and alters relative abundance of mRNA transcripts and epigenetic status of buffalo (Bubalus bubalis) embryos. Anim Reprod Sci. 2019, 208, 106136. [CrossRef]
- Averill-Bates, D.A. The antioxidant glutathione. Vitam Horm. 2023, 121, 109-141. [CrossRef]
Figure 1.
Effect of OF and UF on the presence of ROS and GSH in parthenogenetic embryos. Panel photographs (light and dark fields) of embryos in the blastocyst stage are observed. Presence of ROS in green. Presence of GSH in blue. 100X Magnification. Scale bar 200 μm.
Figure 1.
Effect of OF and UF on the presence of ROS and GSH in parthenogenetic embryos. Panel photographs (light and dark fields) of embryos in the blastocyst stage are observed. Presence of ROS in green. Presence of GSH in blue. 100X Magnification. Scale bar 200 μm.
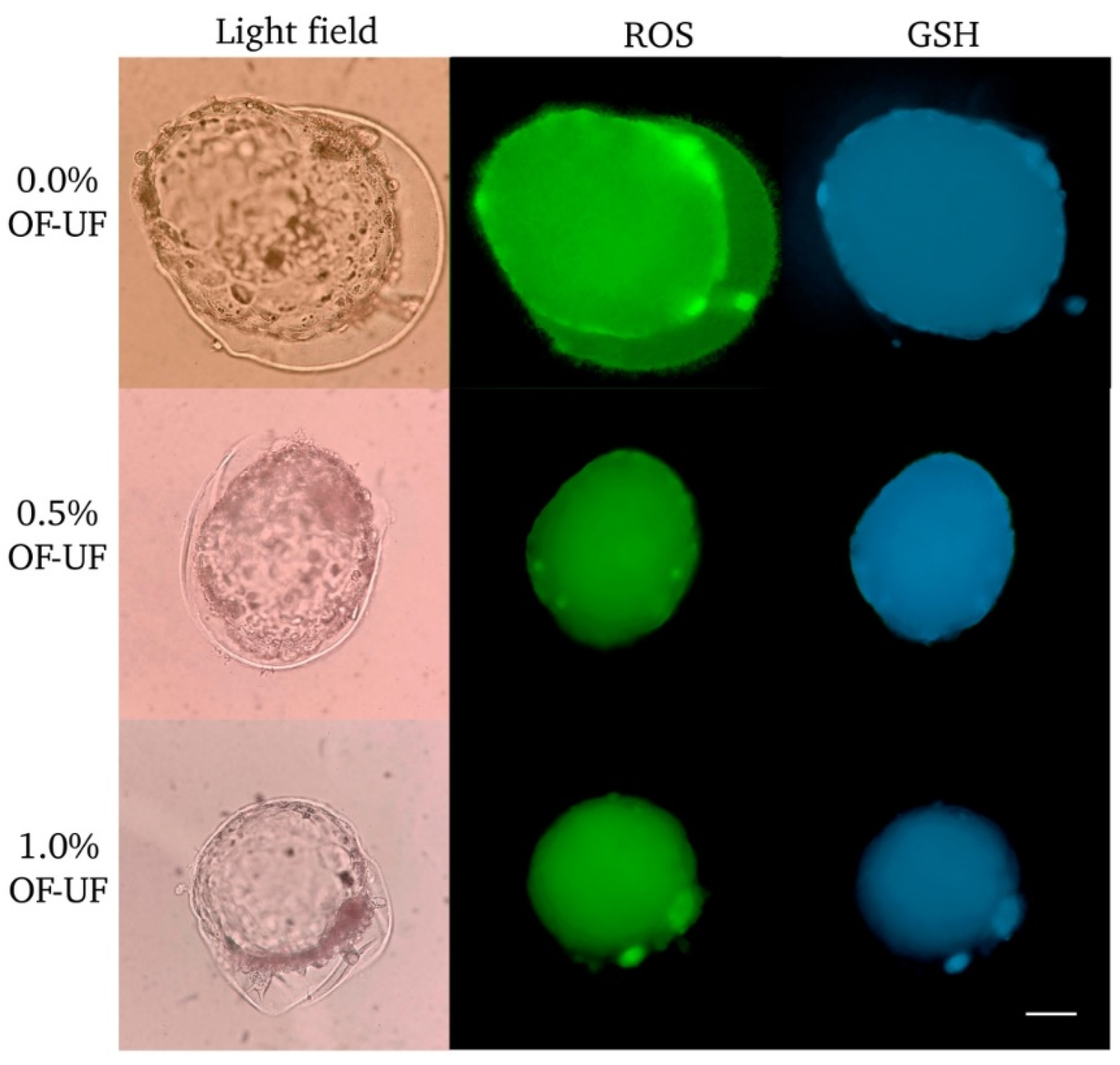

Table 1.
Effect of OF and UF on the in vitro development of O. aries parthenogenetic embryos.
| OF-UF (%) | No. | Cleavage | 4 to 16- cells | Morula | Blastocysts | Fragmented |
|---|---|---|---|---|---|---|
| 0.0 | 140 | 81.6 ± 3.6a (116) | 31.8 ± 4.9a (40) | 52.7 ± 8.1a (62) | 44.9 ± 7.5a (50) | 18.6 ± 8.6a (14) |
| 0.5 | 132 | 81.1 ± 4.0a (107) | 45.4 ± 6.5a (49) | 46 ± 6.7a (52) | 35.7 ± 5.7a (34) | 10.3 ± 4.8a (6) |
| 1.0 | 129 | 71.5 ± 6.1a (95) | 37.6 ± 10.9a (31) | 54.9 ± 12.6a (55) | 28 ± 4.0a (28) | 9.3 ± 3.9a (9) |
| 2.0 | 55 | 67.3 ± 12.5a (41) | 100b (41) | 0b | 0b | 0a |
Means ± SE (%) of six repetitions were carried out for each treatment. The cleavage rate was determined after 36 h of in vitro culture. The cleavage rate of parthenogenetic embryos at the 4 to 16- cells, morula, and fragmented stages was determined on day 4 of culture. The blastocyst rate was determined on day 7. Different literals (a, b) between columns mean statistical differences (P<0.05).
Table 2.
Effect of OF and UF on the in vitro development of O. aries cloned embryos.
| OF-UF (%) |
No. | Cleavage | 4 to 16- cells |
Morula | Blastocysts |
Fragmented |
|---|---|---|---|---|---|---|
| 0.0 | 78 | 97.1 ± 2.9a (76) | 27.9 ± 8.0a (20) | 47.3 ± 9.2a (37) | 28.1 ± 5.5a (21) | 24.8 ± 3.7a (19) |
| 0.5 | 76 | 99.1 ± 0.9a (75) | 36.8 ± 9.4a (28) | 45.1 ± 6.5a (34) | 21.2 ± 4.1a (15) | 18.1 ± 4.7a (13) |
| 1.0 | 66 | 100a (66) | 62.1 ± 7.0a (41) | 27.1 ± 7.0b (18) | 7.7 ± 3.0b (5) | 10.9 ± 4.0a (7) |
Means ± SE (%) of seven repetitions were carried out for each treatment. The cleavage rate was determined after 36 h of in vitro culture. The cleavage rate of cloned embryos at the 4 to16- cells, morula, and fragmented stages was determined at day 4 of culture. The blastocyst rate was determined on day 7. Different literals (a, b) in columns mean statistical differences (P<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



