
Submitted:
20 August 2024
Posted:
22 August 2024
You are already at the latest version
Abstract
Influenza A Virus (IAV) is a multi-host pathogen maintained in water birds and capable of spillover into humans, wildlife, and livestock. Prior research has focused on dabbling ducks as a known IAV reservoir species, yet our understanding of influenza dynamics in other water birds, including gulls, is lacking. Here, we quantify morphological and environmental drivers of serological (antibody detection by ELISA) and virological (viral RNA detection by PCR) prevalence in two gull species: Ring-billed (\emph{Larus delawarensis}) and Franklin's (\emph{Leucophaeus pipixcan}) gulls. Across 12 months and 10 locations, we tested over 1500 gulls for influenza viral RNA, and additionally tested antibody levels in nearly 1000 of these. We find substantial virus prevalence and a large, nonoverlapping seroprevalence, with significant differences across age and species classifications. Body condition index had minimal explanatory power to predict (sero)positivity, and the effect of the surrounding environment was idiosyncratic. Our results hint at a nontrivial relationship between virus and seropositivity, highlighting serological surveillance as a valuable counterpoint to sequencing. By providing indication of both past infections and susceptibility to future infections, serosurviellance can help inform the distribution of limited resources to maximize surveillance effectiveness for a disease of high human, wildlife, and livestock concern.
Keywords:
Influenza A virus (IAV)
; gull
; prevalence
; transmission
; Larus delawarensis
; Leucophaeus pipixcan
; Charadriiformes
; positivity
; seroprevalence
; seropositivity
1. Introduction
Spillover of multihost pathogens between humans, domestic animals, and wildlife is increasingly common [1,2,3,4]. Pathogen spillover can have negative conservation implications for wildlife [5], cripple livestock farming economies and communities [6,7], and occasionally result in devastating global pandemics in humans (including SARS-CoV-2 from bats; [4,8]). In order to understand spillover risk and better control multihost pathogens, it is important to understand pathogen dynamics within the reservoir of infection [9,10]. Infection reservoirs can be defined as “one or more epidemiologically connected populations or environments in which a pathogen can be permanently maintained and from which infection is transmitted to the target population” [11], while the target population of interest can be humans or a specific animal species. Pathogen dynamics in infection reservoirs are driven by, among other things, host density, prevalence and intensity of infection, existing immunity, movement, and migration (which impact contact rates between infected and susceptible hosts; [9,12,13]).
Influenza A Virus (IAV) is a classic multihost pathogen which is maintained in wild water birds and can spillover into humans (e.g. 1918 influenza pandemic; [14]), pigs [15], wild birds of conservation concern (e.g. the Caspian Tern Hydroprogne caspia and Common Tern Sterna hirundo; [16]), and domestic poultry (e.g. chickens and turkeys; [17,18]). In the wild bird reservoir, especially in migratory water birds like ducks, geese, and gulls, IAV circulates year round with seasonal peaks of infection occurring when immature birds migrate en masse to and from the breeding grounds [19]. When migratory birds come in contact, either directly or indirectly, with domestic poultry, the likelihood of interspecies transmission increases with the number of wild birds and the frequency and duration of visits with domestic poultry [20]. In domestic poultry, IAV are classified as highly pathogenic avian influenza (HPAI) or having low pathogenicity (LP or LPAI) based on laboratory criteria that include the presence of basic amino acids at the IAV hemagglutinin protein cleavage site and whether they cause mortality in at least 75% of intravenously-inoculated susceptible chickens within 10 days post-inoculation [21]. Regardless of the IAV being classified as LPAI or HPAI, all IAV have the potential to mutate and change and are an ongoing threat for susceptible humans, pigs, wild birds, and domestic poultry [22,23].
Although known infection reservoir species such as dabbling ducks can survive most IAV infections (depending on the strain) and potentially distribute IAV, there are gaps in our understanding of avian influenza dynamics in birds such as gulls. Gulls are a neglected infection reservoir for IAV [24]. Gulls are not only commonly infected with IAV, but are a species that, when infected, facilitate IAV reassortment and genetic change [25]. Gulls could be important for IAV spillover by being a source population for IAV from wild birds to humans [26] or from wild birds to domestic birds [27]. Furthermore, gulls also migrate long distances (i.e. internationally) and could bring novel influenza strains to new areas [28,29]. Since the literature review conducted by [24], IAV surveillance in gull species has increased and has been conducted more globally. However, the majority of the surveillance has occurred in Europe [25,30,31] and the studies on gulls in North America have primarily focused on the Atlantic flyway and Arctic regions [26,32,33,34,35,36,37], with studies of inland gulls less commonly conducted [29]. Regardless of their location and relative paucity, studies like these have demonstrated that gull IAV infection dynamics are quite variable due to differences in species, geography, diet, local ecology, and focal ecosystem.
To further add to the body of knowledge regarding gulls as an IAV infection reservoir, we studied IAV dynamics in two species of gulls in the state of Minnesota, USA: Ring-billed gulls (Larus delawarensis) and Franklin’s gulls (Leucophaeus pipixcan). Poultry and grain farmers frequently report large flocks of gulls on farms and in fields and have questioned the role these birds may play in distributing avian influenza on the landscape. In the 2021/2022 HPAI H5 outbreak in North America, spatiotemporal analyses of HPAI-positive premises in the United States showed that HPAI detection was most likely to occur within seven days of heavy wild bird observations [38]. Seasonally, gulls, like all wild birds, have variable locations and activities in Minnesota, some of which bring them close to domestic poultry and others that preclude interactions with any domestic birds. For example, in the spring (February, March, and April), migratory gulls come back to their breeding grounds in Minnesota and are often sighted at landfills. In the summer, gulls are often found at a breeding colony where adult gulls nest during the first part of the summer (May to early June). During the second part of the summer (late June through July), the breeding colonies have susceptible flightless hatch-year birds (with waning egg-yolk derived antibodies to IAV) that are mixing with adult gulls. These immunologically naïve hatch-year birds are important for understanding IAV dynamics as they have higher IAV prevalence than the adults and those with a higher body condition index are less likely to be IAV positive [39]. In the fall (August, September, and October) the migratory gulls slowly leave Minnesota during which time they are foraging at places like landfills to stock up for the migration and are mixing with other species.
Age of bird, season of the year, and location are known drivers of IAV dynamics in wild birds, yet gulls have previously been studied at just the species level, rather than considering characteristics such as host age or other spatiotemporal characteristics. For example, three published studies of gulls in 2014-2016 all demonstrated a high seroprevalence in gulls and a low viral prevalence, findings which would discourage gull IAV surveillance since it appeared unlikely that IAV would be detectable in the seemingly immune gull population [30,32,33]. However, surveillance and research efforts conducted on both inland and coastal gulls of Massachusetts revealed higher viral prevalence as detected by IAV PCR in juveniles [26], and further explored the relationship between seroprevalence, viral prevalence, and age in their multi-year longitudinal study. [26] recommend, “...targeted sampling of the sites where juvenile gulls first congregate on the mainland after leaving the colony, as this is where most virus will be detected.”
Encouraged by efforts of IAV researchers who studied gulls of the coastal regions [26] and having experience with effective capturing methods and IAV assays [29,39], we conducted this research in an effort to explore drivers of serological (antibody detection by ELISA) prevalence and virological (viral RNA detection by PCR) prevalence in two species, Ring-billed gulls (Larus delawarensis) and Franklin’s gulls (Leucophaeus pipixcan), in the non-coastal, i.e. inland/continental, state of Minnesota in the midwestern United States. We analyzed location effects to ascertain if gulls in Minnesota wetlands, i.e. wetlands historically postulated to harbor many wild IAV-positive Anatidae, were more likely to be IAV PCR positive than gulls in dense poultry production areas, i.e. areas with a highly dense population of IAV-susceptible domestic turkeys with previous HPAI and LPAI infected flocks. In addition to location, we analyzed any correlations between gull age, weight, and species with antibody or viral RNA prevalence. Our goal was to further elucidate the drivers of IAV infections in gulls, thus generating information to help guide future surveillance efforts and control IAV interspecies transmission between wild and domestic animals, with implications for public health.
2. Materials and Methods
2.1. Gull Sampling
Using the methods described in Froberg et al. [39] and Rasmussen et al. [29], gulls were trapped, banded, measured, sampled, and released at landfill and breeding colony locations across Minnesota (Supplementary Material supp:sec:sampling,supp:fig:samplemap). Six of these locations were landfills that have generally maintained sizable numbers of gulls (100) available to be captured and sampled. Three landfills sites were located within counties with a high density of poultry facilities (Blue Earth, Cottonwood, and Kandiyohi Counties) and three landfills were located in counties with a low density of poultry facilities (Dakota, Kanabec and Rice Counties). Gulls were sampled between November 2016 and October of 2017 (Supplementary Material supp:fig:sampleweeks,supp:fig:sampleseasons).
When the birds were handled, the age was recorded as described in Froberg et al. [39]. Briefly, we recorded the age of birds as a categorical variable: hatch-year, juvenile, or adult. We based age on plumage, bill coloring, iris coloring and leg coloring [40,41,42]. During the breeding season, birds incapable of sustained flight, i.e. in some stage of pre-juvenile molt, and judged to be approximately 3 to 5 weeks old were categorized as hatch-year birds. This cohort is effectively considered immunologically naïve [43]. Based on phenotypic characteristics, we classified birds years of age as juveniles and birds years old as adults. In addition to age classification, the weight of each bird in grams (g) was measured using a spring scale (©PESOLA AG, Schindellegi, Switzerland). Finally, each bird’s keel (mm) and head-to-beak (mm) measurements were taken to calculate a body condition index (BCI) as outlined by Boersma and Ryder [44], namely the ratio of bird weight to the summed keel and head-to-beak lengths.
Correlations between all covariates are reported in Supplementary Material supp:sec:correlations,supp:fig:corsub,supp:fig:corfull,supp:fig:cormorph.
2.2. Influenza A Virus RNA and Antibody Detection
Briefly, blood was collected from the birds for influenza antibody tests and swabs (cloacal and oral) were collected for virus RNA testing. Specifically, the swabs were tested for influenza A virus with rRT-PCR following standard methods [21] in the Midwest Central Research and Outreach Center (MCROC) laboratory in Willmar, Minnesota, USA. To determine virus positivity, a bird’s swab with an rRT-PCR cycle threshold (CT) value was considered positive for IAV. Any bird swab with a CT value , or having no CT value, i.e. no amplification of viral RNA, was considered negative for IAV. Sera was separated from the blood and tested for IAV antibodies at MCROC with a commercial blocking enzyme-linked immunosorbent assay (bELISA, MultiS-Screen ELISA [FlockCheck], Idexx™, Westbrook, ME). To determine seropositivity to IAV, a bird’s serum sample was considered negative for IAV antibodies if the sample result-to-negative control absorbance ratio (s/n ratio) was greater than or equal to 0.50. A sample was considered positive for IAV if the s/n ratio was less than 0.50 [45]. Sera with detectable antibodies by bELISA were forwarded to the Diagnostic Virology Laboratory at the USDA National Veterinary Services Laboratory in Ames, Iowa, USA for Hemagglutination Inhibition (HI) assays to detect IAV subtype-specific antibodies. The HI assays were performed via standard procedures [21] and used the viruses listed in tab:antigens as HI antigens. Continuous variables (i.e. CT value or s/n ratio) were compared between species/ages using two-sample Student’s t-tests, while discrete variables (i.e. “positive” / “negative”) were compared with 2-sample tests of equal proportions.
2.3. Generalized Linear Models
We sought to identify covariates that are most predictive of virus positivity and seropositivity using generalized linear regression models (GLMs) in which the response variable was binomially distributed (i.e. positive or negative test result). We identified the best fitting model in a two-step process, first identifying the best way to incorporate location-specific effects (i.e. sampling site), and then considering the incorporation of other covariates.
For the first part, three potential methods for incorporating location-effects in the model were considered. First, we included location as a fixed effect in the GLM, such that each location received a unique coefficient in the model. Second, we considered including location as a random effect, that is, where the location-specific coefficients were considered as independent, identically random draws from a distribution. Finally, we considered omitting explicit location from the models, instead including the percentage of habitat within a 10 mile radius of the sampling site that was classified as wetland (“wetland percentage”), and the number of commercially raised poultry within a 10 mile radius of the sampling site (“poultry production”), each of which were unique for each location. Wetland habitat classifications were provided by the National Wetland Inventory for Minnesota [46]. The number of poultry within a 10 mile radius was based on geospatial information obtained from Oklahoma State University’s Spatial Agrometrics Tool for Livestock Estimation (available at https://osu-geog.maps.arcgis.com/apps/MapSeries/index.html?appid=b1bf901f4e9b4ab7ae83798642b793b0) and in consultation with the Minnesota Turkey Growers Association and the Minnesota Board of Animal Health regarding proximity to poultry facilities. Thus, Kanabec, Dakota, and Rice County landfills are in low-density poultry areas, while Kandiyohi, Cottonwood, and Blue Earth landfills are in high-density poultry areas [39]. We then proceeded to use stepwise model selection on binomial GLMs that initially included location (included as identified above), species, age, body condition index, wetland percentage, and poultry production. Variables were algorithmically removed or re-added according to changes in Akaike information criterion (AIC) until the model could no longer be improved. From all models within two units of the lowest AIC value, the one with the highest area under the receiver operating characteristic curve was identified as the “best” model. Final model effects of categorical explanatory variables were explored in more detail using Tukey’s honestly significant difference test. Binomial model fit was further quantified using McKelvey-Zavoina Pseudo-R2 [47,48].
All analyses were conducted in R v. 4.4.1 [49], with functions from the broom [50], fuzzyjoin [51], GGally [52], ggbeeswarm [53], ggmap [54], ggpubr [55], GLMMadaptive [56], janitor [57], kableExtra [58], lme4 [59], lubridate [60], magrittr [61], multcomp [62], patchwork [63], pROC [64], scatterpie [65], and tidyverse [66] packages.
3. Results
3.1. Sampling Distribution
A total of 1592 gulls were sampled between October, 2016 and September, 2017 at 10 locations across Minnesota, USA: of which 244 were Franklin’s Gulls (Leucophaeus pipixcan) and 1348 were Ring-billed Gulls (Larus delawarensis). Of these, 1588 were swabbed for influenza A virus RNA, and 990 of these had their blood drawn for antibody titer. The majority, 164 (67%), of the Franklin’s gulls were adults, 79 (32%) were juvenile, and 1 was undetermined. For the Ring-billed gulls, 726 (46%) were adults, 258 (16%) were juvenile, 297 (19%) were hatch-year, and 67 were undetermined. No hatch-year Franklin’s Gulls were captured, as no sampling was conducted at Franklin’s Gull breeding sites. IAV was detected in 424 of the swabbed birds (27% virus positivity), and 553 of the blood samples tested were found to positively detect IAV antibodies (54% seropositivity).
Analyses presented in the main text are restricted to site-date combinations in which at least five birds of each species were captured. This corresponds to birds captured at the Burnsville, Kandiyohi County, or Rice County landfills in August of 2017Ḟurthermore, we only considered samples for which both rRT-PCR and antibody titer results were available. Thus, 243 birds were considered in these analyses: 142 Franklin’s Gulls [103 (73%) adults and 39 (27%) juveniles] and 101 Ring-billed Gulls [57 (56%) adults and 44 (44%) juveniles]. IAV was detected in 145 of the swabbed birds (60% virus positivity), and 107 of the blood samples were found to positively detect IAV antibodies (44% seropositivity).
3.2. Observational Results
Body condition index (BCI) was found to have minimal explanatory power for virus positivity and seropositivity (Pseudo consistently less than 0.2), though, when significant, such relationships tended to be for virus positivity and positive in sign. That is, when such relationships existed, gulls with higher BCI were slightly more likely to be found with virus RNA. BCI did, however, differ significantly between species (, ), and between ages within species (Franklin’s Gulls: , ; Ring-billed gulls: , ; fig:bci).
Regardless of species, adult gulls were found to have higher seropositivity than juveniles (, ), but not virus positivity (, ; fig:age). Between species, Ring-billed gulls were more likely to be found virus positive (, ), while Franklin’s gulls were more likely to be found seropositive (, ; fig:species).
The effect of the surrounding habitat on virus/seropositivity was circumstantial, we found a slight positive relationship between Franklin’s Gull virus positivity and the percentage of the surrounding habitat classified as wetlands (, , Pseudo ), and a slight negative relationship between Ring-billed Gull virus positivity and the estimated number of commercially raised poultry in the surrounding habitat (, , Pseudo ; Supplementary Material supp:fig:regvirus). Seropositivity was unaffected by either habitat variable (Supplementary Material supp:fig:regsero).
When considering all available samples (i.e. rather than just the subset of sites/dates where both species were sampled), we find a significant negative relationship between virus positivity and seropositivity (, ), however this is not the case for the subsetted data (, ). In particular, 57 of the sampled birds in the subsetted data were both virus positive and seropositive. Birds that were “dual” positive did not have significantly different PCR CT values or ELISA s/n ratios than did birds who were just virus- or seropositive. Birds that were just virus positive did, however, have higher s/n ratios (mean: 0.779, standard deviation: 0.148) than those that were negative for both virus and antibodies (mean: 0.701, standard deviation: 0.143; , ; fig:dualpos).
All analyses were likewise performed on the full dataset (Supplementary Material supp:fig:regvirusfull,supp:fig:regserofull,supp:fig:corfull,supp:sec:fulldatafigs,supp:sec:fulldatacontinuous).
3.3. Model Results
The best fitting model for virus positivity incorporated location, species, age, and body condition index, yet only the coefficients associated with location were found to be significantly different from 0 (using bold typeface to denote variables with significant model coefficients; ). In contrast, the best fitting model for seropositivity included only species and age as explanatory variables (). Considering the quantitative response variables of cycle threshold (for PCR amplification) and s/n ratio (for ELISA), the best fitting models were and , respectively. Best fitting models for the full dataset are reported in Supplementary Material supp:sec:fulldatamodels.
4. Discussion
Influenza is a devastating, widespread disease in humans and livestock, but little is known of influenza dynamics in reservoir and “mixing vessel” species such as gulls. In this work, we identified differences in gull virus- and seroprevalence across space and time, but also by species and age. We found that younger gulls were unlikely to be seropositive, but more likely to be virus-positive than not, while adult birds were often virus- or seropositive (or both). Between species, Ring-billed Gulls were more likely to be virus positive and seronegative, while Franklin’s Gulls were equally likely to be positive and negative for both tests. Understanding these nuances of influenza disease dynamics can help inform surveillance and public- and livestock-health interventions.
4.1. Serological Sampling Can Complement Current Surveillance
Wildlife disease surveillance is often conducted through monitoring active infection status, for instance through the detection of live pathogen propagules via genetic sequencing [67,68]. While such efforts provide valuable insight into the current disease dynamics, they are limited to providing a snapshot in time. In contrast, serology can reveal the signature of past infection [69], reducing the sensitivity of sample timing and providing insight into disease dynamics in other locations along migration routes despite temporally and geographically limited sampling.
For instance, in this work, sampling was restricted to four migration stopover sites in August of 2017. Yet, the lack of seropositive results by bELISA for juveniles of both Ring-billed and Franklin’s gulls at that time allows inference that juvenile birds did not encounter IAV prior to their arrival at these sites [70] or that antibodies had waned to non detectable levels. Furthermore, the relatively high positivity of bELISA and rRT-PCR results in adult gulls suggest that at least some of the adult gulls were infected with IAV prior to our sampling period. Our wider data include additional sampling at breeding sites and throughout the spring-early fall in which gulls are present in Minnesota (Supplementary Material supp:fig:samplemap,supp:fig:sampleweeks,supp:fig:sampleseasons). These data show, for Ring-billed gulls, higher seroprevalence in the spring (with nearly no virus positivity), followed by lower seroprevalence and much higher virus positivity in the fall. While in situ sampling of gulls across their migration would provide the most insight into real-time disease dynamics, sampling across the full spatial and temporal range of long-distance migratory species like gulls would require a substantial investment of time and resources.
Serosurveillance can also facilitate forecasting of susceptibility to future outbreaks, because seropositivity as detected by bELISA and virus positivity as detected by rRT-PCR are negatively correlated. This negative correlation between seropositivity and virus positivity is supported by other studies of trans-hemispheric migratory Charadriiformes [71] and is in agreement with the widely accepted conclusion that age-related patterns of seropositivity and viral positivity observed in the wild result from birds gaining immunity to IAV with repeated infections over the course of their lifespan [72,73].
Moreover, serological surveillance need not be restricted to a binary categorization. Our results revealed that seropositive gulls were more likely to be infected when their antibody titers were lower. Yet seropositive birds had similar result-to-negative control absorbance ratio (s/n) ratios regardless of whether or not the virus was detected by PCR. Similarly, we find greater variance in antibody titers in general than rRT-PCR cycle thresholds. While virus positive gulls have statistically indistinguishable cycle thresholds across age or species (excepting lower cycle thresholds in hatch-year Ring-billed gulls), serologically negative gulls’ s/n ratios varied across both age and species. Future work could explore these patterns more comprehensively, perhaps identifying key transitions in antibody titers that inform infection risk.
Our results suggest that if one wants to target surveillance to maximize likelihood of positive test results, juveniles have more detectable virus than other age groups. Yet, if resources allow for it, the most information is gained by combining the serology and rRT-PCR results. Doing so for our data reveals that adults are actually slightly more likely to be positive for at least one test: while about 56% of adult gulls in our study were found to be virus positive, an additional 27% were seropositive and virus negative, with only 17% of the population actually having no known past infections.
Importantly, we did not find that seropositivity is a reliable indicator that a bird will be virus negative. Half of seropositive birds in our study were also found to be virus positive (25% when considering all available samples), suggesting that either these birds have non-neutralizing antibodies and cannot fight off new infections or that these birds are late in their infection timeline and are seroconverting (convalescing) and their antibodies are causing virus to go down. If gulls are being reinfected while there is still the possibility of persisting virus from a prior infection, this raises the possibility of gulls serving as sources of recombination and new strain development [74], which would be a critical consideration for early detection of strains of interest for public health [75,76]. Future studies with higher resolution and repeated sampling of the same birds is needed to disambiguate these possibilities.
4.2. The Role of the Surrounding Environment
Interestingly, we found only idiosyncratic evidence of the surrounding environment (specifically the density of wetlands and poultry farms in the surrounding landscape) on prevalence. Franklin’s Gulls were more likely to be positive as the proportion of wetlands increased, while Ring-billed Gulls were less likely to be positive as the number of poultry increased. The former could be driven by cross-species infections with other waterfowl in the surrounding wetlands, and can also be considered as an infection risk for those same populations [77]. The latter is harder to explain. Our expectation was that gulls are a potential infection risk for surrounding poultry agriculture, as gulls are often encountered in close proximity to poultry farms [78]. The negative correlation suggests that this risk might not be as large as suspected, as gull populations close to large farming operations are less likely to be carrying the virus.
Importantly, these relationships should be evaluated with caution, as there are only four sites considered in our main analysis. More extensive sampling in both space and time are needed to fully understand the relationship between the surrounding environment (natural and anthropogenic) on disease processes, however such research is costly in terms of both money and labor.
Critically, some of these potential environmental covariates might likewise vary temporally; for instance, the actual coverage of wetlands might depend on recent precipitation. Our sampling revealed a significant variation in (sero)prevalence over time, with Ring-billed gulls being largely virus-negative (yet seropositive) in the spring/summer compared to the fall samples considered elsewhere in the text. This could be due to altering environmental conditions over time, or differential levels of pathogen exposure at other locations in the summer migration in-between our sampling dates.
4.3. Insights into Influenza Reservoirs
Despite several decades of appreciation for the relevance of waterbirds for influenza A transmission dynamics, much of the current literature focuses on ducks as the focal reservoir for potential transmission to livestock and humans [24]. Our results point to the importance of broadening this perspective. Gulls in particular deserve increased attention, as they both migrate further than ducks [79] and are more commonly human-associated [80]. Moreover, we find that influenza infections are common, with more than half of our samples being actively positive, and many more (especially adults) showing serological evidence of recent infection (fig:age).
Gulls occur in relatively large numbers in the Midwestern U.S., yet are under-represented, compared to hunter-harvested ducks, in surveillance efforts in Minnesota, and arguably in the world [39]. Per data obtained from the Influenza Research Database [81], the surveillance for IAV in Laridae (gulls) and Anatidae (ducks) is quite disproportionately in favor of ducks (tab:gullsvducks). Thus, despite the call by Arnal et al. [24] to expand surveillance to gulls and other Laridae, the gap in surveillance still remains.
Gulls migrate long distances [24,82], allowing for both long-distance disease transmission [83] and perhaps increasing recombination risk due to exposure to (and consequent infection with) a more diverse suite of strains. Upon returning to nesting grounds in the proximity of both humans and livestock, these gulls provide ample opportunities for introduction of new strains such as the recent outbreak of HPAI in North America (including Minnesota, USA) in 2021 [84].
Between the two gull species we considered in this study, our results suggest that while both species have relatively high prevalence, Ring-billed Gulls especially showed widespread infection suggesting they might be a more critical reservoir for IAV.
4.4. Limitations of BCI in Cross-Species Analyses
Body condition indices (BCI) are often developed as standardized measures of bird health [85], however our results call the applicability of this measure into question when comparing across ages or species. We found significant differences across species (with Franklin’s Gulls having substantially lower values than did Ring-billed Gulls) and age (with juvenile birds having slightly lower BCI than adult birds in each species), though with substantial overlap in the distributions for the latter (fig:bci). These results suggest that BCI cannot be used reliably across birds of varying species or age. More critically, while our previous results suggest that birds would have lower BCI during and immediately after infection [39], BCI was not found to relate strongly with virus or seropositivity (best fitting binomial glm ) and, when significant, was found to be positively associated with virus positivity. That is, birds with higher BCI were more likely to be found to be virus positive. Altogether, these results cast doubt on the usefulness of BCI in assessing disease status.
5. Conclusions
Despite being spatially and temporally constrained, this study provides insights into the distribution of influenza A virus in wild gull populations. Understanding the dynamics of this critical disease before being confronted by epidemics in livestock and human populations is critical to both preventing such outbreaks and adequately responding when they do occur. Our results highlight the heterogeneity in (sero)prevalence across age and species, as well as hint at the possibility of identifying environmental covariates of disease risk. These findings serve as a foundation for building hypotheses to direct further research, but can also be used to inform surveillance, in particular emphasizing the benefits of adding serological sampling to future surveillance efforts and increasing attention to historically understudied gull species, which are nevertheless critical components in the spread of influenza through humans, livestock, and wildlife.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Section S1. Sampling distribution through space and time; Figure S1. Map of the distribution of sample counts and virus-/seropositivity rates at each site; Figure S2. Timeseries of sample counts and virus-/seropositivity rates in each sampling week; Figure S3. Timeseries of percent sample positivity aggregated by sampling season; Section S2. (Sero)positivity by landscape variables; Figure S4. Binomial regressions of PCR virus detection by landscape variables, subsetting data to include only dates/sites with at least 5 birds captured from each species; Figure S5. Binomial regressions of ELISA antibody detection by landscape variables, subsetting data to include only dates/sites with at least 5 birds captured from each species; Figure S6. Binomial regressions of PCR virus detection by landscape variables, using all available samples; Figure S7. Binomial regressions of ELISA antibody detection by landscape variables, using all available samples; Section S3. Continuous response variable figures analogous to main text; Figure S8. As Figure 2 in the main text, but using continuous response variables for each test (ELISA result-to-negative control absorbance (s/n) ratio for the serology and the minimum of the two PCR cycle thresholds (cloacal and oropharyngeal)); Figure S9. As Figure 3 in the main text, but using continuous response variables for each test (ELISA result-to-negative control absorbance (s/n) ratio for the serology and the minimum of the two PCR cycle thresholds (cloacal and oropharyngeal)); Section S4. Full data figures analogous to main text; Figure S10. As Figure 1, but using all available samples; Figure S11. As Figure 2, but using all available samples; Figure S12. As Figure 3, but using all available samples; Figure S13. As Figure 4, but using all available samples; Section S5. Best fitting full data models; Section S6. Full data figures with continuous response variables; Figure S14. As Figure S8, but using all available samples; Figure S15. As Figure S9, but using all available samples; Section S7. Covariate correlations; Figure S16. Correlations between all covariates used in the main text analyses, subsetting data to include only dates/sites with at least 5 birds captured from each species; Figure S17. As Figure S16, but for all available samples; Figure S18. Correlations between morphometric measurements and body condition index
Author Contributions
Conceptualization, M.M., M.R.C. and M.E.C.; methodology, M.M. and E.R.; validation, M.M., M.R.C. and M.E.C.; formal analysis, M.M.; investigation, E.R. and M.R.C.; resources, M.R.C.; data curation, M.M., E.R. and E.C.; writing—original draft preparation, M.M., E.R., M.R.C. and M.E.C.; writing—review and editing, M.M., M.R.C. and M.E.C..; visualization, M.M.; supervision, M.R.C. and M.E.C.; project administration, M.R.C. and M.E.C.; funding acquisition, M.R.C. and M.E.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the following organizations and research grants: the Minnesota Agricultural Experimental Station 2020 Avian Influenza Program grant (CON 81431, Project 85606), “Capturing movements of avian influenza virus from wild birds to domestic poultry in Minnesota,” Minnesota Agricultural Experimental Station 2015 Avian Influenza Program grant (CON 58786, Project 72634) “Defining the role gull species play in the disease ecology of avian influenza in Minnesota,” and the Environment and Natural Resources Trust Fund from the Legislative-Citizen Commission on Minnesota Resources program grant M.L. 2016, “Avian Influenza distribution, evolution, and impacts on Ring-billed and Herring gulls in Minnesota.” M.E.C. was funded by the National Science Foundation (DEB-2321358).
Institutional Review Board Statement
The animal study protocol was reviewed and approved by the Institutional Animal Care and Use Committee at the University of Minnesota (Protocol # 1910-37530A).
Data Availability Statement
All data used in these analyses are available at https://doi.org/10.13020/9b7h-0973.
Acknowledgments
We would like to thank Francesca Cuthbert, Carol Cardona, and Colette Pollard for input at early stages in this work. We thank Mia Torchetti and Mary Lea Killian of the Diagnostic Virology Laboratory, National Veterinary Services Laboratories, USDA, for the HI testing of the sera. We are grateful to field team members, Todd Froberg and Madeline Grunklee, for capturing, sampling, and releasing gulls with E.R. and M.R.C. To all the owners and operators of the landfills, we express our gratitude for allowing us access to your sites.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| IAV | Influenza A Virus |
| RNA | Ribonucleic acid |
| PCR CT | Polymerase Chain Reaction cycle threshold |
| ELISA S/N | Enzyme-linked Immunosorbent Assay sample result-to-negative control absorbance ratio |
| HPAI | High Pathogenic Avian Influenza |
| LPAI | Low Pathogenic Avian Influenza |
| BIC | Body Condition Index |
References
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging Infectious Diseases of Wildlife—Threats to Biodiversity and Human Health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.J.; Albery, G.F.; Merow, C.; Trisos, C.H.; Zipfel, C.M.; Eskew, E.A.; Olival, K.J.; Ross, N.; Bansal, S. Climate change increases cross-species viral transmission risk. Nature 2022, 607, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Meadows, A.J.; Stephenson, N.; Madhav, N.K.; Oppenheim, B. Historical trends demonstrate a pattern of increasingly frequent and severe spillover events of high-consequence zoonotic viruses. BMJ Global Health 2023, 8, e012026. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.F.; Acevedo-Whitehouse, K.; Pedersen, A.B. The role of infectious diseases in biological conservation. Animal Conservation 2009, 12, 1–12. [Google Scholar] [CrossRef]
- Musoke, J.; Hlokwe, T.; Marcotty, T.; du Plessis, B.J.; Michel, A.L. Spillover of Mycobacterium bovis from Wildlife to Livestock, South Africa. Emerging Infectious Diseases 2015, 21, 448–451. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Miller, R.S.; Farnsworth, M.L.; Malmberg, J.L. Diseases at the livestock-wildlife interface: Status, challenges, and opportunities in the United States. Preventive Veterinary Medicine 2013, 110, 119–132. [Google Scholar] [CrossRef]
- Ruiz-Aravena, M.; McKee, C.; Gamble, A.; Lunn, T.; Morris, A.; Snedden, C.E.; Yinda, C.K.; Port, J.R.; Buchholz, D.W.; Yeo, Y.Y.; Faust, C.; Jax, E.; Dee, L.; Jones, D.N.; Kessler, M.K.; Falvo, C.; Crowley, D.; Bharti, N.; Brook, C.E.; Aguilar, H.C.; Peel, A.J.; Restif, O.; Schountz, T.; Parrish, C.R.; Gurley, E.S.; Lloyd-Smith, J.O.; Hudson, P.J.; Munster, V.J.; Plowright, R.K. Ecology, evolution and spillover of coronaviruses from bats. Nature Reviews Microbiology 2022, 20, 299–314. [Google Scholar] [CrossRef]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to zoonotic spillover. Nature Reviews Microbiology 2017, 15, 502–510. [Google Scholar] [CrossRef]
- Viana, M.; Mancy, R.; Biek, R.; Cleaveland, S.; Cross, P.C.; Lloyd-Smith, J.O.; Haydon, D.T. Assembling evidence for identifying reservoirs of infection. Trends in Ecology & Evolution 2014, 29, 270–279. [Google Scholar] [CrossRef]
- Haydon, D.T.; Cleaveland, S.; Taylor, L.; Laurenson, M.K. Identifying Reservoirs of Infection: A Conceptual and Practical Challenge. Emerging Infectious Diseases 2002, 8, 1468–1473. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wilber, M.Q.; Yang, A.; Boughton, R.; Manlove, K.R.; Miller, R.S.; Pepin, K.M.; Wittemyer, G. A model for leveraging animal movement to understand spatio-temporal disease dynamics. Ecology Letters 2022, 25, 1290–1304. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.K.; Craft, M.E.; Zuk, M.; Binning, S.A. Host migration strategy is shaped by forms of parasite transmission and infection cost. Journal of Animal Ecology 2019, 88, 1601–1612. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Morens, D.M. The 1918 Influenza Pandemic and Its Legacy. Cold Spring Harbor Perspectives in Medicine 2020, 10, a038695. [Google Scholar] [CrossRef] [PubMed]
- Bourret, V. Avian influenza viruses in pigs: An overview. The Veterinary Journal 2018, 239, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Loeb, J. Scottish seabirds hit by avian influenza. Veterinary Record 2022, 190, 488–488. [Google Scholar] [CrossRef]
- Garber, L.; Bjork, K.; Patyk, K.; Rawdon, T.; Antognoli, M.; Delgado, A.; Ahola, S.; McCluskey, B. Factors Associated with Highly Pathogenic Avian Influenza H5N2 Infection on Table-Egg Layer Farms in the Midwestern United States, 2015. Avian Diseases 2016, 60, 460–466. [Google Scholar] [CrossRef]
- Ssematimba, A.; Malladi, S.; Hagenaars, T.J.; Bonney, P.J.; Weaver, J.T.; Patyk, K.A.; Spackman, E.; Halvorson, D.A.; Cardona, C.J. Estimating within-flock transmission rate parameter for H5N2 highly pathogenic avian influenza virus in Minnesota turkey flocks during the 2015 epizootic. Epidemiology & Infection 2019, 147, e179. [Google Scholar] [CrossRef]
- van Dijk, J.G.B.; Hoye, B.J.; Verhagen, J.H.; Nolet, B.A.; Fouchier, R.A.M.; Klaassen, M. Juveniles and migrants as drivers for seasonal epizootics of avian influenza virus. Journal of Animal Ecology 2014, 83, 266–275. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Gonzales, J.L. Quantification of visits of wild fauna to a commercial free-range layer farm in the Netherlands located in an avian influenza hot-spot area assessed by video-camera monitoring. Transboundary and Emerging Diseases 2020, 67, 661–677. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Spackman, E. A Brief Introduction to Avian Influenza Virus. In Animal Influenza Virus: Methods and Protocols; Spackman, E., Ed.; Methods in Molecular Biology, Springer US: New York, NY, 2020; pp. 83–92. [Google Scholar] [CrossRef]
- Chen, H.; Yuan, H.; Gao, R.; Zhang, J.; Wang, D.; Xiong, Y.; Fan, G.; Yang, F.; Li, X.; Zhou, J.; Zou, S.; Yang, L.; Chen, T.; Dong, L.; Bo, H.; Zhao, X.; Zhang, Y.; Lan, Y.; Bai, T.; Dong, J.; Li, Q.; Wang, S.; Zhang, Y.; Li, H.; Gong, T.; Shi, Y.; Ni, X.; Li, J.; Zhou, J.; Fan, J.; Wu, J.; Zhou, X.; Hu, M.; Wan, J.; Yang, W.; Li, D.; Wu, G.; Feng, Z.; Gao, G.F.; Wang, Y.; Jin, Q.; Liu, M.; Shu, Y. Clinical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: a descriptive study. The Lancet 2014, 383, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.H.; Taubenberger, J.K. . The origin of the 1918 pandemic influenza virus: a continuing enigma. Journal of General Virology 2003, 84, 2285–2292. [Google Scholar] [CrossRef]
- Arnal, A.; Vittecoq, M.; Pearce-Duvet, J.; Gauthier-Clerc, M.; Boulinier, T.; Jourdain, E. Laridae: A neglected reservoir that could play a major role in avian influenza virus epidemiological dynamics. Critical Reviews in Microbiology 2015, 41, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Postnikova, Y.; Treshchalina, A.; Boravleva, E.; Gambaryan, A.; Ishmukhametov, A.; Matrosovich, M.; Fouchier, R.A.M.; Sadykova, G.; Prilipov, A.; Lomakina, N. Diversity and Reassortment Rate of Influenza A Viruses in Wild Ducks and Gulls. Viruses 2021, 13, 1010. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ineson, K.M.; Hill, N.J.; Clark, D.E.; MacKenzie, K.G.; Whitney, J.J.; Laskaris, Y.; Ronconi, R.A.; Ellis, J.C.; Giroux, J.F.; Lair, S.; Stevens, S.; Puryear, W.B.; Runstadler, J.A. Age and season predict influenza A virus dynamics in urban gulls: consequences for natural hosts in unnatural landscapes. Ecological Applications 2022, 32, e2497. [Google Scholar] [CrossRef]
- Hill, N.J.; Takekawa, J.Y.; Cardona, C.J.; Meixell, B.W.; Ackerman, J.T.; Runstadler, J.A.; Boyce, W.M. Cross-Seasonal Patterns of Avian Influenza Virus in Breeding and Wintering Migratory Birds: A Flyway Perspective. Vector-Borne and Zoonotic Diseases 2012, 12, 243–253. [Google Scholar] [CrossRef]
- Anderson, C.M.; Gilchrist, H.G.; Ronconi, R.A.; Shlepr, K.R.; Clark, D.E.; Fifield, D.A.; Robertson, G.J.; Mallory, M.L. Both short and long distance migrants use energy-minimizing migration strategies in North American herring gulls. Movement Ecology 2020, 8, 26. [Google Scholar] [CrossRef]
- Rasmussen, E.A.; Czaja, A.; Cuthbert, F.J.; Tan, G.S.; Lemey, P.; Nelson, M.I.; Culhane, M.R. Influenza A viruses in gulls in landfills and freshwater habitats in Minnesota, United States. Frontiers in Genetics 2023, 14, 2024-02–14]. [Google Scholar] [CrossRef]
- Arriero, E.; Müller, I.; Juvaste, R.; Martínez, F.J.; Bertolero, A. Variation in Immune Parameters and Disease Prevalence among Lesser Black-Backed Gulls (Larus fuscus sp.) with Different Migratory Strategies. PLOS ONE 2015, 10, e0118279. [Google Scholar] [CrossRef]
- Dusek, R.J.; Hallgrimsson, G.T.; Ip, H.S.; Jónsson, J.E.; Sreevatsan, S.; Nashold, S.W.; TeSlaa, J.L.; Enomoto, S.; Halpin, R.A.; Lin, X.; Fedorova, N.; Stockwell, T.B.; Dugan, V.G.; Wentworth, D.E.; Hall, J.S. North Atlantic Migratory Bird Flyways Provide Routes for Intercontinental Movement of Avian Influenza Viruses. PLOS ONE 2014, 9, e92075. [Google Scholar] [CrossRef]
- Huang, Y.; Wille, M.; Benkaroun, J.; Munro, H.; Bond, A.L.; Fifield, D.A.; Robertson, G.J.; Ojkic, D.; Whitney, H.; Lang, A.S. Perpetuation and reassortment of gull influenza A viruses in Atlantic North America. Virology 2014, 456-457, 353–363. [Google Scholar] [CrossRef]
- Guinn, K.; Fojtik, A.; Davis-Fields, N.; Poulson, R.L.; Krauss, S.; Webster, R.G.; Stallknecht, D.E. Antibodies to Influenza A Viruses in Gulls at Delaware Bay, USA. Avian Diseases 2016, 60, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Reeves, A.B.; Hall, J.S.; Poulson, R.L.; Donnelly, T.; Stallknecht, D.E.; Ramey, A.M. Influenza A virus recovery, diversity, and intercontinental exchange: A multi-year assessment of wild bird sampling at Izembek National Wildlife Refuge, Alaska. PLOS ONE 2018, 13, e0195327. [Google Scholar] [CrossRef]
- Bevins, S.N.; Pedersen, K.; Lutman, M.W.; Baroch, J.A.; Schmit, B.S.; Kohler, D.; Gidlewski, T.; Nolte, D.L.; Swafford, S.R.; DeLiberto, T.J. Large-Scale Avian Influenza Surveillance in Wild Birds throughout the United States. PLOS ONE 2014, 9, e104360. [Google Scholar] [CrossRef]
- Krauss, S.; Stucker, K.M.; Schobel, S.A.; Danner, A.; Friedman, K.; Knowles, J.P.; Kayali, G.; Niles, L.J.; Dey, A.D.; Raven, G.; Pryor, P.; Lin, X.; Das, S.R.; Stockwell, T.B.; Wentworth, D.E.; Webster, R.G. Long-term surveillance of H7 influenza viruses in American wild aquatic birds: are the H7N3 influenza viruses in wild birds the precursors of highly pathogenic strains in domestic poultry? Emerging Microbes & Infections 2015, 4, 1–9. [Google Scholar] [CrossRef]
- Criado, M.F.; Moresco, K.A.; Stallknecht, D.E.; Swayne, D.E. Low-pathogenicity influenza viruses replicate differently in laughing gulls and mallards. Influenza and Other Respiratory Viruses 2021, 15, 701–706. [Google Scholar] [CrossRef]
- USDA-APHIS. Epidemiologic and other analyses of HPAI affected poultry flocks: , 2022 Interim Report. Technical report, USDA:APHIS:VS: Center for Epidemiology and Animal Health, Fort Collins, CO, 2022. [Online; accessed 2024-02-14]. 1 July.
- Froberg, T.; Cuthbert, F.; Jennelle, C.S.; Cardona, C.; Culhane, M. Avian Influenza Prevalence and Viral Shedding Routes in Minnesota Ring-Billed Gulls (Larus delawarensis). Avian Diseases 2018, 63, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.N.G.; Dunn, J. A Reference Guide to Gulls of the Americas; Houghton Mifflin Harcourt, 2007. Google-Books-ID: J7lQsufWfdkC.
- Pyle, P. Identification Guide to North American Birds. Part II: Anatidae to Alcidae; Slate Creek Press: Point Reyes Station Cal, 2008. [Google Scholar]
- Pollet, I.L.; Shutler, D.; Chardine, J.W.; Ryder, J.P. Ring-billed Gull (Larus delawarensis), version 2.0. Birds of North America. [CrossRef]
- Velarde, R.; Calvin, S.E.; Ojkic, D.; Barker, I.K.; Nagy, E. Avian Influenza Virus H13 Circulating in Ring-Billed Gulls (Larus delawarensis) in Southern Ontario, Canada. Avian Diseases 2010, 54, 411–419. [Google Scholar] [CrossRef]
- Boersma, D.; Ryder, J.P. Reproductive Performance and Body Condition of Earlier and Later Nesting Ring-Billed Gulls. Journal of Field Ornithology 1983, 54, 374–380. [Google Scholar]
- Brown, J.D.; Stallknecht, D.E.; Berghaus, R.D.; Luttrell, M.P.; Velek, K.; Kistler, W.; Costa, T.; Yabsley, M.J.; Swayne, D. Evaluation of a Commercial Blocking Enzyme-Linked Immunosorbent Assay To Detect Avian Influenza Virus Antibodies in Multiple Experimentally Infected Avian Species. Clinical and Vaccine Immunology 2009, 16, 824–829. [Google Scholar] [CrossRef]
- of Natural Resources, M.D. National Wetland Inventory for Minnesota, 2019. [Online; accessed 2024-07-11].
- McKelvey, R.D.; Zavoina, W. A statistical model for the analysis of ordinal level dependent variables. The Journal of Mathematical Sociology 1975, 4, 103–120. [Google Scholar] [CrossRef]
- Veall, M.R.; Zimmermann, K.F. Evaluating Pseudo-R2’s for binary probit models. Quality and Quantity 1994, 28, 151–164. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, 2024.
- Robinson, D.; Hayes, A.; Couch, S. broom: Convert Statistical Objects into Tidy Tibbles, 2024. R package version 1.0.6.
- Robinson, D. fuzzyjoin: Join Tables Together on Inexact Matching, 2020. R package version 0.1.6.
- Schloerke, B.; Cook, D.; Larmarange, J.; Briatte, F.; Marbach, M.; Thoen, E.; Elberg, A.; Crowley, J. GGally: Extension to ’ggplot2’, 2024. R package version 2.2.1.
- Clarke, E.; Sherrill-Mix, S.; Dawson, C. ggbeeswarm: Categorical Scatter (Violin Point) Plots, 2023. R package version 0.7.2.
- Kahle, D.; Wickham, H. ggmap: Spatial Visualization with ggplot2. The R Journal 2013, 5, 144–161. [Google Scholar] [CrossRef]
- Kassambara, A. ggpubr: ’ggplot2’ Based Publication Ready Plots, 2023. R package version 0.6.0.
- Rizopoulos, D. GLMMadaptive: Generalized Linear Mixed Models using Adaptive Gaussian Quadrature, 2023. R package version 0.9-1.
- Firke, S. janitor: Simple Tools for Examining and Cleaning Dirty Data, 2023. R package version 2.2.0.
- Zhu, H. kableExtra: Construct Complex Table with ’kable’ and Pipe Syntax, 2024. R package version 1.4.0.
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Grolemund, G.; Wickham, H. Dates and Times Made Easy with lubridate. Journal of Statistical Software 2011, 40, 1–25. [Google Scholar] [CrossRef]
- Bache, S.M.; Wickham, H. magrittr: A Forward-Pipe Operator for R, 2022. R package version 2.0.3.
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biometrical Journal 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Pedersen, T.L. patchwork: The Composer of Plots, 2024. R package version 1.2.0.
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Müller, M. pROC: an open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinformatics 2011, 12, 77. [Google Scholar] [CrossRef]
- Yu, G. scatterpie: Scatter Pie Plot, 2024. R package version 0.2.3.
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; Kuhn, M.; Pedersen, T.L.; Miller, E.; Bache, S.M.; Müller, K.; Ooms, J.; Robinson, D.; Seidel, D.P.; Spinu, V.; Takahashi, K.; Vaughan, D.; Wilke, C.; Woo, K.; Yutani, H. Welcome to the tidyverse. Journal of Open Source Software 2019, 4, 1686. [Google Scholar] [CrossRef]
- Michel, A.L.; Van Heerden, H.; Crossley, B.M.; Al Dahouk, S.; Prasse, D.; Rutten, V. Pathogen detection and disease diagnosis in wildlife: challenges and opportunities. Revue Scientifique Et Technique (International Office of Epizootics) 2021, 40, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; Luttrell, M.P.; Berghaus, R.D.; Kistler, W.; Keeler, S.P.; Howey, A.; Wilcox, B.; Hall, J.; Niles, L.; Dey, A.; Knutsen, G.; Fritz, K.; Stallknecht, D.E. Prevalence of Antibodies to Type A Influenza Virus in Wild Avian Species Using Two Serologic Assays. Journal of Wildlife Diseases 2010, 46, 896–911. [Google Scholar] [CrossRef]
- Gilbert, A.T.; Fooks, A.R.; Hayman, D.T.S.; Horton, D.L.; Müller, T.; Plowright, R.; Peel, A.J.; Bowen, R.; Wood, J.L.N.; Mills, J.; Cunningham, A.A.; Rupprecht, C.E. Deciphering Serology to Understand the Ecology of Infectious Diseases in Wildlife. EcoHealth 2013, 10, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Salewski, V.; Bruderer, B. The evolution of bird migration—a synthesis. Naturwissenschaften 2007, 94, 268–279. [Google Scholar] [CrossRef]
- Johnson, J.A.; DeCicco, L.H.; Ruthrauff, D.R.; Krauss, S.; Hall, J.S. Avian Influenza Virus Antibodies in Pacific Coast Red Knots (Calidris canutus roselaari). Journal of Wildlife Diseases 2014, 50, 671–675. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Munster, V.J.; Majoor, F.; Lexmond, P.; Vuong, O.; Stumpel, J.B.G.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Schutten, M.; Slaterus, R.; Fouchier, R.A.M. Avian Influenza A Virus in Wild Birds in Highly Urbanized Areas. PLOS ONE 2012, 7, e38256. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.M.; Hall, J.S.; Flint, P.L.; Franson, J.C.; Ely, C.R.; Schmutz, J.A.; Samuel, M.D. High Seroprevalence of Antibodies to Avian Influenza Viruses among Wild Waterfowl in Alaska: Implications for Surveillance. PLOS ONE 2013, 8, e58308. [Google Scholar] [CrossRef]
- Lu, L.; Lycett, S.J.; Leigh Brown, A.J. Reassortment patterns of avian influenza virus internal segments among different subtypes. BMC Evolutionary Biology 2014, 14, 16. [Google Scholar] [CrossRef]
- Morse, S.S. Public Health Surveillance and Infectious Disease Detection. Biosecurity and Bioterrorism: Biodefense Strategy, Practice, and Science 2012, 10, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Fuller, T.L.; Gilbert, M.; Martin, V.; Cappelle, J.; Hosseini, P.; Njabo, K.Y.; Abdel Aziz, S.; Xiao, X.; Daszak, P.; Smith, T.B. Predicting Hotspots for Influenza Virus Reassortment. Emerging Infectious Diseases 2013, 19, 581–588. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fereidouni, S. Ecology of Avian Influenza Viruses in Wild Birds. In Ecology of Wild Bird Diseases; CRC Press, 2024. number-of-pages: 18.
- Ljubojević, D.; Radosavljević, V.; Milanov, D. The role of gulls (Laridae) in the emergence and spreading of antibiotic resistance in the environment. World’s Poultry Science Journal 2016, 72, 853–864. [Google Scholar] [CrossRef]
- Endo, A.; Nishiura, H. The Role of Migration in Maintaining the Transmission of Avian Influenza in Waterfowl: A Multisite Multispecies Transmission Model along East Asian-Australian Flyway. Canadian Journal of Infectious Diseases and Medical Microbiology 2018, 2018, 3420535. [Google Scholar] [CrossRef]
- Raven, S.; Coulson, J. The distribution and abundance of Larus gulls nesting on buildings in Britain and Ireland. Bird Study 1997, 44, 13–34. [Google Scholar] [CrossRef]
- Zhang, Y.; Aevermann, B.D.; Anderson, T.K.; Burke, D.F.; Dauphin, G.; Gu, Z.; He, S.; Kumar, S.; Larsen, C.N.; Lee, A.J.; Li, X.; Macken, C.; Mahaffey, C.; Pickett, B.E.; Reardon, B.; Smith, T.; Stewart, L.; Suloway, C.; Sun, G.; Tong, L.; Vincent, A.L.; Walters, B.; Zaremba, S.; Zhao, H.; Zhou, L.; Zmasek, C.; Klem, E.B.; Scheuermann, R.H. Influenza Research Database: An integrated bioinformatics resource for influenza virus research. Nucleic Acids Research 2017, 45, D466–D474. [Google Scholar] [CrossRef]
- Hatch, J.J. Threats to public health from gulls (Laridae). International Journal of Environmental Health Research 1996, 6, 5–16. [Google Scholar] [CrossRef]
- Ahlstrom, C.A.; van Toor, M.L.; Woksepp, H.; Chandler, J.C.; Reed, J.A.; Reeves, A.B.; Waldenström, J.; Franklin, A.B.; Douglas, D.C.; Bonnedahl, J.; Ramey, A.M. Evidence for continental-scale dispersal of antimicrobial resistant bacteria by landfill-foraging gulls. Science of The Total Environment 2021, 764, 144551. [Google Scholar] [CrossRef] [PubMed]
- Bevins, S.N.; Shriner, S.A.; Cumbee, J.C.; Dilione, K.E.; Douglass, K.E.; Ellis, J.W.; Killian, M.L.; Torchetti, M.K.; Lenoch, J.B. Intercontinental Movement of Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4 Virus to the United States, 2021. Emerging Infectious Diseases 2022, 28, 1006–1011. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Labocha, M.K.; Hayes, J.P. Morphometric indices of body condition in birds: a review. Journal of Ornithology 2012, 153, 1–22. [Google Scholar] [CrossRef]
Figure 1.
Body condition index (BCI) significantly differs between species (blue vs. green) and (to a lesser extent) age within species (light vs. dark within colors). This complicates the attribution of cross-species differences in IAV positivity to differences in BCI per se. Only includes samples collected from dates, sites where at least five birds of each species were sampled. See Supplementary Material supp:fig:bcifull for an analogous figure with the full dataset.
Figure 1.
Body condition index (BCI) significantly differs between species (blue vs. green) and (to a lesser extent) age within species (light vs. dark within colors). This complicates the attribution of cross-species differences in IAV positivity to differences in BCI per se. Only includes samples collected from dates, sites where at least five birds of each species were sampled. See Supplementary Material supp:fig:bcifull for an analogous figure with the full dataset.
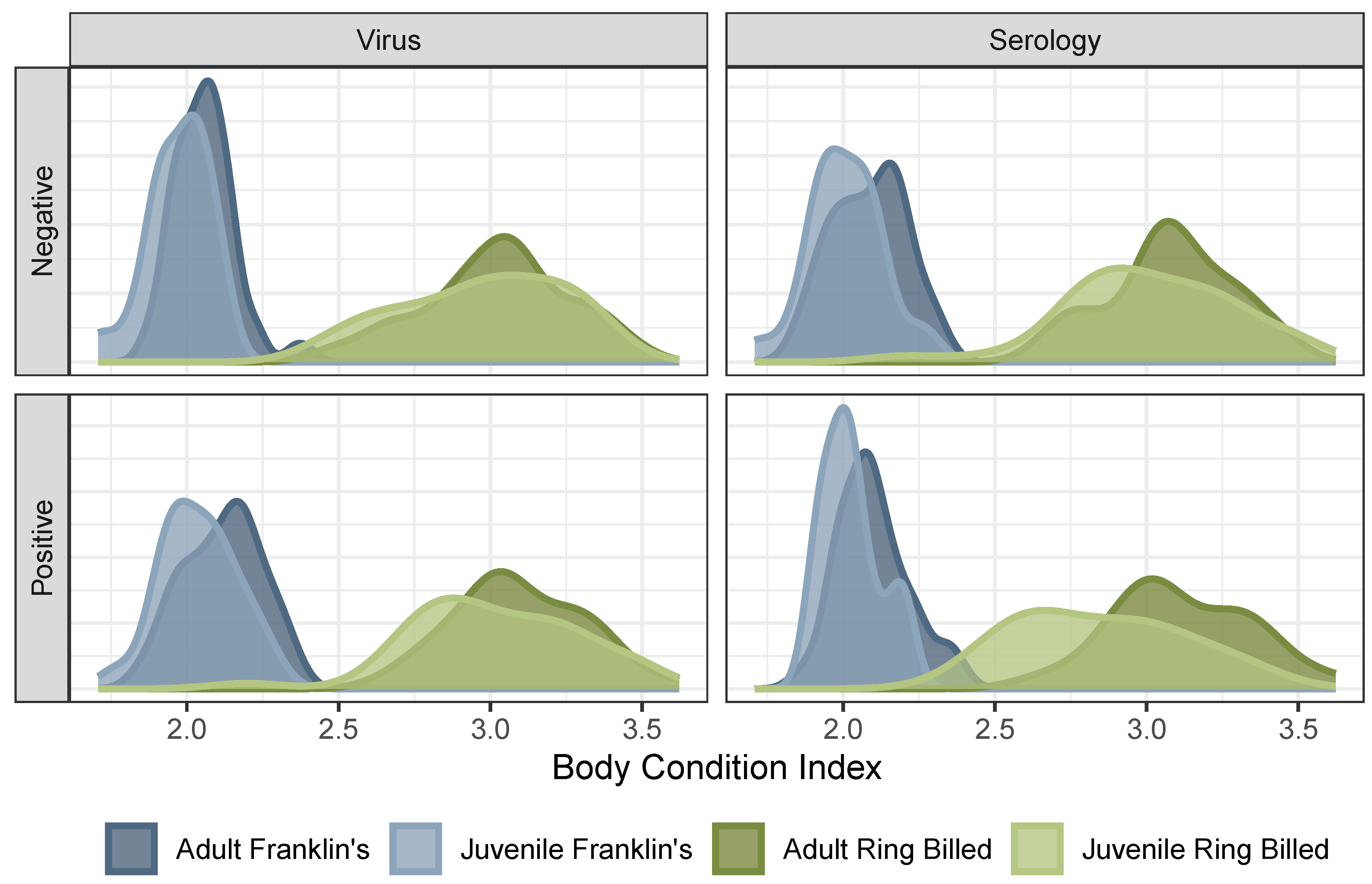
Figure 2.
Gulls differed in their likelihood of being found virus- or seropositive between species (reported p-values are the results of 2-Sample Tests of Equal Proportions). While Ring-billed gulls were more likely to be virus positive than Franklin’s Gulls, they were less likely to be seropositive than Franklin’s Gulls. For both virus- and seropositivity, Franklin’s Gulls were found to be approximately equally likely to be positive or negative. Analogous figure considering continuous measures of test response (cycle threshold and result-to-negative control absorbance ratio Supplementary Material supp:fig:agecont). Only includes samples collected from dates, sites where at least five birds of each species were sampled. See Supplementary Material supp:fig:agefull,supp:fig:agecontfull for analogous figures with the full dataset.
Figure 2.
Gulls differed in their likelihood of being found virus- or seropositive between species (reported p-values are the results of 2-Sample Tests of Equal Proportions). While Ring-billed gulls were more likely to be virus positive than Franklin’s Gulls, they were less likely to be seropositive than Franklin’s Gulls. For both virus- and seropositivity, Franklin’s Gulls were found to be approximately equally likely to be positive or negative. Analogous figure considering continuous measures of test response (cycle threshold and result-to-negative control absorbance ratio Supplementary Material supp:fig:agecont). Only includes samples collected from dates, sites where at least five birds of each species were sampled. See Supplementary Material supp:fig:agefull,supp:fig:agecontfull for analogous figures with the full dataset.
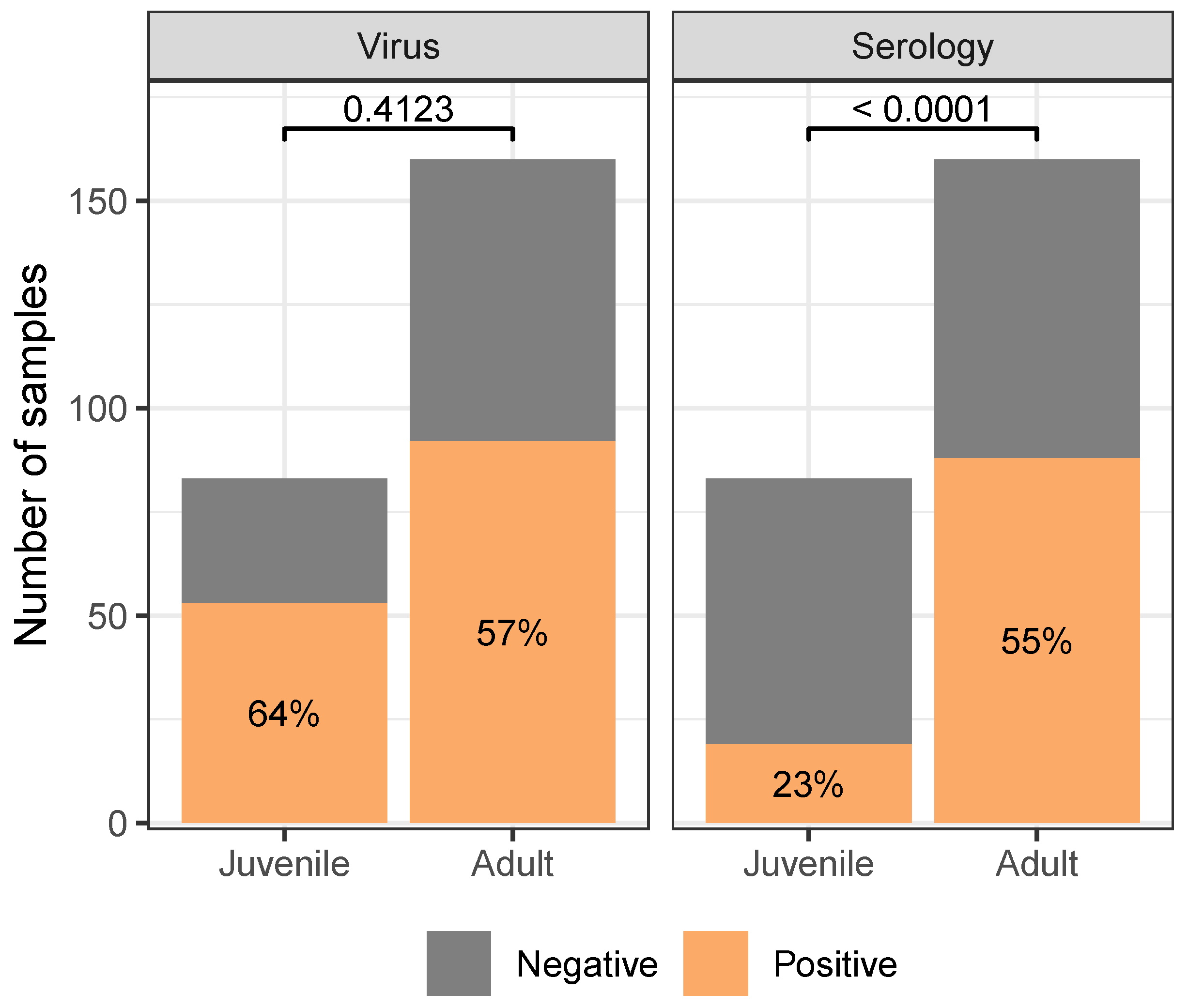
Figure 3.
Gulls differed in their likelihood of being found seropositive, but not virus positive, between age classes (reported p-values are the results of 2-Sample Tests of Equal Proportions). Juvenile gulls were unlikely to be seropositive, while adult gulls were more likely to be seropositive than not. For virus positivity, both age classes were more likely to be found positive than not. Analogous figure considering continuous measures of test response (cycle threshold and result-to-negative control absorbance ratio in Supplementary Material supp:fig:speciescont). Only includes samples collected from dates, sites where at least five birds of each species were sampled. See Supplementary Material supp:fig:speciesfull,supp:fig:speciescontfull for analogous figures with the full dataset.
Figure 3.
Gulls differed in their likelihood of being found seropositive, but not virus positive, between age classes (reported p-values are the results of 2-Sample Tests of Equal Proportions). Juvenile gulls were unlikely to be seropositive, while adult gulls were more likely to be seropositive than not. For virus positivity, both age classes were more likely to be found positive than not. Analogous figure considering continuous measures of test response (cycle threshold and result-to-negative control absorbance ratio in Supplementary Material supp:fig:speciescont). Only includes samples collected from dates, sites where at least five birds of each species were sampled. See Supplementary Material supp:fig:speciesfull,supp:fig:speciescontfull for analogous figures with the full dataset.
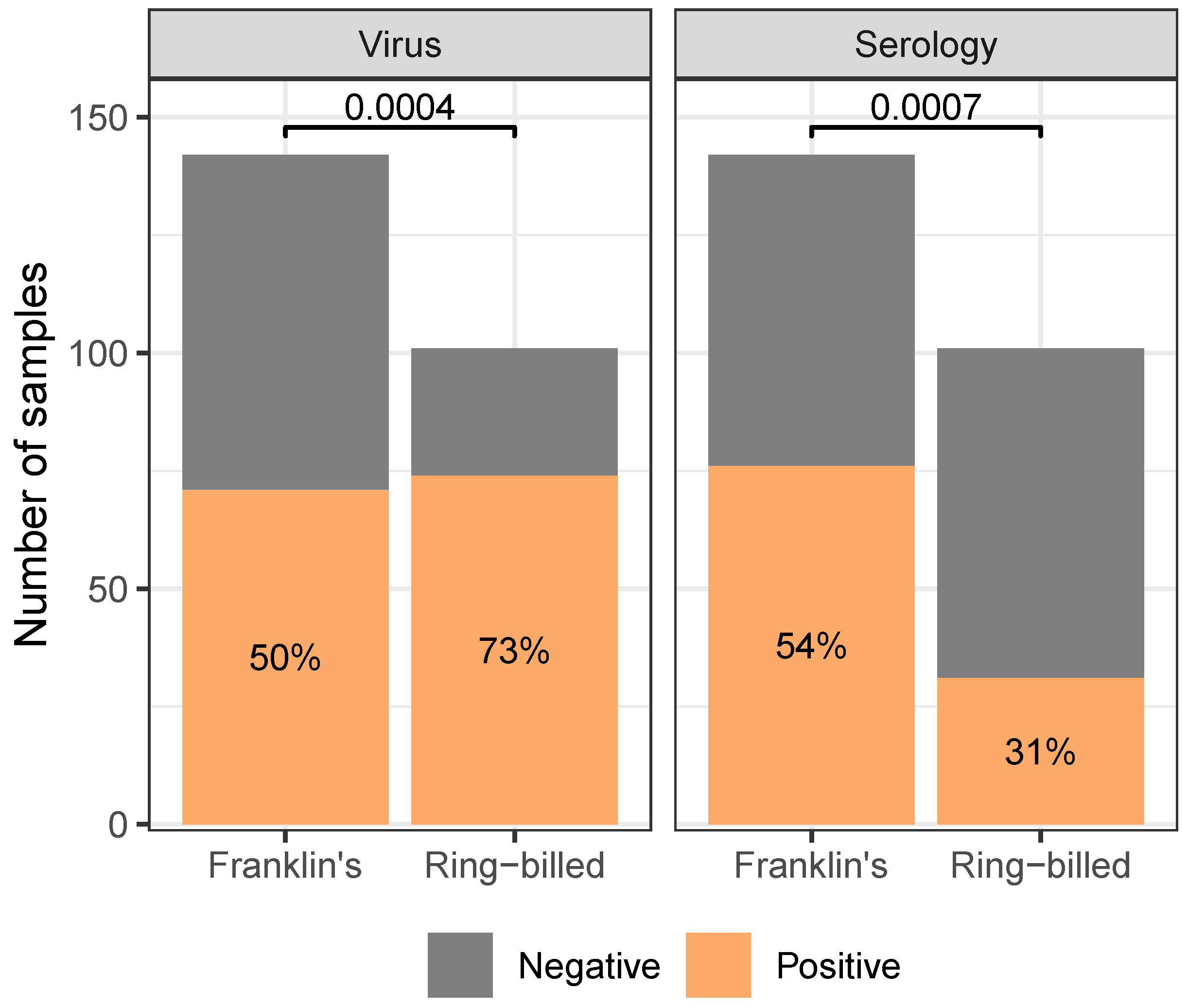
Figure 4.
Relationship between ELISA result-to-negative control absorbance (s/n) ratio across different results for virus detection by PCR amplification (reported p-values are the results of 2-sample Wilcoxon Rank Sum Tests). While gulls did not differ by virus detection in their s/n ratio when found to be seropositive, they did differ when seronegative, with virus-positive birds having slightly higher s/n ratios than virus-negative birds. That is, given seronegativity, virus-negative birds were slightly closer to our threshold for seropositivity (indicated by a dashed horizontal line) than were virus-positive birds. Interestingly, there is no difference in S/N ratio (level of antibodies) of seropositive birds that are virus positive and birds that are virus negative. Thus, antibody titer does not predict virus detection in seropositive birds; however, if birds are seronegative, then virus positivity is correlated with higher SN ratios, i.e. weaker antibody response. Only includes samples collected from dates, sites where at least five birds of each species were sampled. See Supplementary Material supp:fig:dualposfull for an analogous figure with the full dataset.
Figure 4.
Relationship between ELISA result-to-negative control absorbance (s/n) ratio across different results for virus detection by PCR amplification (reported p-values are the results of 2-sample Wilcoxon Rank Sum Tests). While gulls did not differ by virus detection in their s/n ratio when found to be seropositive, they did differ when seronegative, with virus-positive birds having slightly higher s/n ratios than virus-negative birds. That is, given seronegativity, virus-negative birds were slightly closer to our threshold for seropositivity (indicated by a dashed horizontal line) than were virus-positive birds. Interestingly, there is no difference in S/N ratio (level of antibodies) of seropositive birds that are virus positive and birds that are virus negative. Thus, antibody titer does not predict virus detection in seropositive birds; however, if birds are seronegative, then virus positivity is correlated with higher SN ratios, i.e. weaker antibody response. Only includes samples collected from dates, sites where at least five birds of each species were sampled. See Supplementary Material supp:fig:dualposfull for an analogous figure with the full dataset.
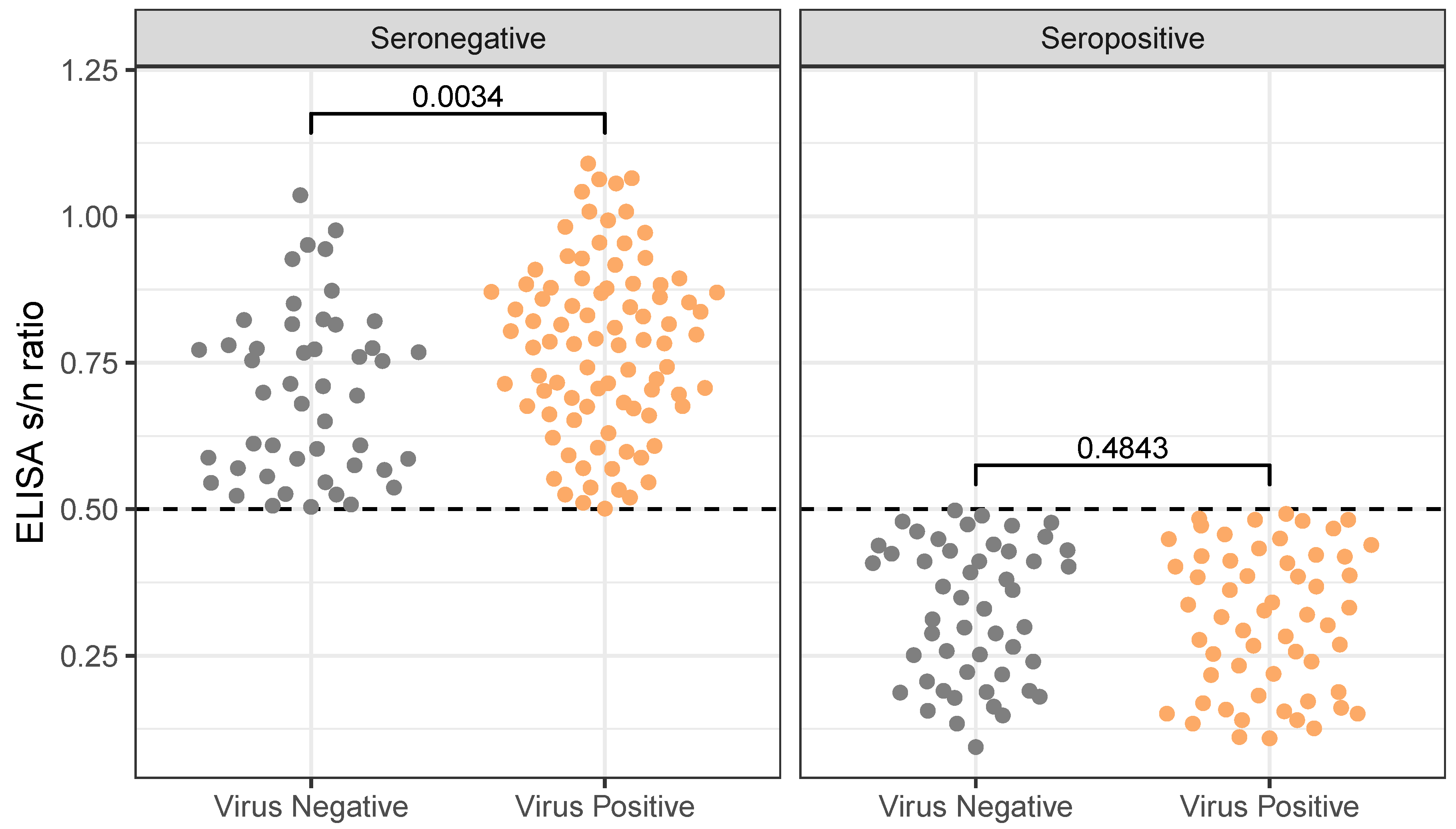

Table 1.
Names and abbreviations of influenza A viruses used as antigens in the hemagglutination inhibition antibody assays. The abbreviation format is Location/year/subtype/pathogenicity.
Table 1.
Names and abbreviations of influenza A viruses used as antigens in the hemagglutination inhibition antibody assays. The abbreviation format is Location/year/subtype/pathogenicity.
| Virus Name | Subtype | Pathogenicity | Abbreviation | Lab designator |
|---|---|---|---|---|
| A/Chicken/Mexico/1433-2/2008 | H5N2 | Low | MX/08/H5N2/LP | none |
| A/Turkey/Wisconsin/1968 | H5N9 | Low | WI/68/H5N9/LP | none |
| A/Turkey/Oregon/1971 | H7N3 | Low | OR/71/H7N3/LP | none |
| A/Turkey/Minnesota/9845-4/2015 | H5N2 | High | MN/15/H5N2/HP | 133 ADV 1501 |
Table 2.
Summary of IAV Surveillance Results for Laridae and Anatidae species for eight years pre- and post-publication by Arnal et al. [24] using the cutoff date of 7/3/2013 from Arnal’s data inquiry.
Table 2.
Summary of IAV Surveillance Results for Laridae and Anatidae species for eight years pre- and post-publication by Arnal et al. [24] using the cutoff date of 7/3/2013 from Arnal’s data inquiry.
| 1/4/2005 to 7/3/2013 | 7/4/2013 to 12/31/2021 | |||||
|---|---|---|---|---|---|---|
| Positive | Tested | Positivity | Positive | Tested | Positivity | |
| Anatidae | 14,624 | 164,215 | 8.9% | 29,584 | 258,631 | 11.4% |
| Laridae | 1,396 | 28,225 | 4.9% | 3,206 | 31,134 | 10.3% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



