
Submitted:
21 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
Rhizosphere microorganisms can utilize nutrient resources in the rhizosphere efficiently, while rhizosphere growth-promoting bacteria play a crucial role in regulating soil fertility and promoting plant health. In this study, we employed a combination of isolation, purification, identification of dominant microorganisms, and high-throughput sequencing technology to analyze the main microbial groups and diversity in the rhizosphere soil of Helichrysum arenarium(L.)Moench. in Altay, Xinjiang,China. By isolating bacterial strains from the rhizosphere soil using a dilution coating method, we successfully obtained 43 strains. Subsequently, selective medium was utilized to screen for growth-promoting characteristics among these isolated strains from the rhizosphere soil of H. arenarium(L.)Moench.. The results obtained through high-throughput amplification sequencing revealed that the rhizosphere soil harbored diverse bacterial communities belonging to 35 phyla, 93 orders, 215 families, 324 genera and 231 species associated with H. arenarium(L.)Moench., as well as fungal communities consisting of 14 phyla ,47 orders ,96 families ,204 genera and 571 species. Among these bacterial communities identified, eight phyla exhibited relative abundances exceeding 1%, with Actinobacteriota being most predominant at an abundance rate of 41.30%. Additionally,five bacterial genera were found to have relative abundances exceeding 1% ,with Arenimicrobium_luteum being most abundant at 6.11% . Furthermore,four fungal phyla displayed relative abundances surpassing1 % ,with Ascomycota accounting for 67.21% of total fungal community composition.Finally,four fungal genera demonstrated relative abundances higher than 1% ,with Knufia representing 3.80% of total fungal community composition.The analysis on culturable bacteria's promotion activity within the rhizospheric soil indicated that two strains,S16and S29, exhibited the highest solubility index for inorganic phosphorus. However, it lacked the ability to dissolve organophosphorus and potassium. On the other hand, strains S7 and S10, which were screened, demonstrated nitrogen-fixing capabilities. Nine strains with excellent iron-bearing capacity were identified, with strain S16 exhibiting the highest D/d value and strongest iron-bearing capacity among them. Our findings revealed that 43 rhizosphere soil bacteria belonged to three genera. The ten growth-promoting bacteria were identified as Kocuria rosea, Priestia megaterium, Bacillus mobilis, B. bataviensis, B. mycoides (two strains), B. paramobilis, and Alcaligenes faecalis. This study provides a particular research basis for the influence of H. arenarium(L.)Moench. rhizosphere microorganisms on soil nutrient release and depicts a solution for improving the yield and quality cultivation of H. arenarium(L.)Moench. by isolating, screening, and identifying rhizosphere soil growth-promoting bacteria, including screening strains with growth-promoting functions and analyzing the population structure of rhizosphere and habitat soil in combination with high-throughput sequencing technology.
Keywords:
Helichrysum arenarium(L.)Moench.
; Rhizosphere microorganisms
; Community structure
; Diversity
; Growth-promoting bacteria
; Xinjiang
1. Introduction
Helichrysum arenarium (L.) Moench. is a perennial herb of the genus Helichrysum in the family of Asteraceae, with a woody, thick rhizome. Its medicinal value is very high, the whole herb contains tannin and volatile oil, which has the function of promoting bile secretion, anti-inflammatory and antioxidant. It grows at altitudes of 900 m-2400 m on earthy hillocks, semi-sandy dunes, slopes of wet saline soils, gravelly soils, sand dunes, grasslands and under pine forests [1,2]. It is distributed in Russia, Europe and Mongolia and Xinjiang of China. It is mainly distributed in Altay City, Habahe County, Fuyun County and Qinghe County of Xinjiang . Altay region has dry and hot summers, cold winters, low precipitation, high evaporation, large temperature difference between day and night, and sufficient light, which makes it a continental north-temperate cold climate zone[3].
The major chemical constituents of H. arenarium (L.) Moench wax chrysanthemum are flavonoids and polyphenolic compounds, while tannins and volatile oils can be found in the entire herb. Additionally, the flower sap exhibits bile secretion-promoting effects [4,5]. This plant possesses a wide range of medicinal properties including choleretic, hepatoprotective, and antiviral effects. Furthermore, its flowers exhibit lipid-lowering activity with identified components such as lcariside F2, icari-side D1, phenethanol-β-vicianoside, benzyl alcohol-β-vicianoside, benzoic acid-β-D-gentiobioside and phenethanol-β-D-gentiobioside[6,7,8].
The root system of this plant enriches soil microorganisms through root secretions. Rhizosphere microorganisms significantly influence plant growth and development through various metabolic activities. Bacteria and fungi are the main types of rhizosphere microorganisms; beneficial ones regulate soil nutrient cycling, secrete hormone compounds to promote growth, and inhibit pathogenic bacteria. Conversely, harmful microorganisms can infect plants leading to hindered growth or death. Plants rely on the rhizosphere microbial community for their functions by obtaining nutrients from them and being influenced by their activities and composition. The diversity of rhizosphere microbes is crucial for maintaining stable soil ecosystems [9,10,11,12]. Numerous studies have analyzed the rhizosphere microbial communities of various plants worldwide resulting in isolation of functional strains widely used in biopromotion studies or antagonism researches. However, no study has reported on the rhizosphere microorganisms specifically associated with H. arenarium (L.) Moench.. Therefore,in this study we aimed to explore the diversity of rhizosphere microorganisms associated with H. arenarium (L.) Moench using high-throughput sequencing techniques. The study provides a research foundation for elucidating the rhizosphere microbial species of H. arenarium (L.) Moench., with a focus on rhizosphere probiotics and their role in regulating soil fertility and plant health.
2. Materials and Methods
2.1. Sample Collection
Soil samples were collected from Burqin County, Altay Region, Xinjiang Uygur Autonomous Region, at an altitude of 737.4m in late July 2023 (48°18N, 86°78E).

Figure 1.
Soil sample collection site of H. arenarium (L.) Moench.
2.1.1. Sample Pre-Treatment
The collected soil samples were thoroughly mixed, sieved through a 2 mm sieve, divided into three portions, labeled, and stored at -80 ℃ and 4 ℃ respectively in a refrigerator.
2.2. Measurement of Soil Nutrient Indicators
Organic matter content was determined using potassium dichromate oxidation-added thermal oxidation; total nitrogen content was determined using Kjeldahl method; hydrolyzable nitrogen content was determined using the alkaline diffusion method; effective phosphorus content was determined using colorimetric method with an ultraviolet spectrophotometer; fast-acting potassium content was determined using flame photometer method.
2.3. High-Throughput Sequencing Analysis of Rhizosphere Microbial Amplicons
Genomic DNA extraction from the soil samples was performed using TianGen magnetic bead-based soil genomic DNA extraction kit followed by purity and concentration assessment via agarose gel electrophoresis (1%). Bacterial 16S rRNA sequence and fungal ITS sequence were amplified by PCR. Library construction utilized NEB Next® Ultra™ II FS DNA PCR-free Library Prep Kit (New England Biolabs) with library quantification done by Qubit and Q-PCR methods. After quality control checks on libraries’ qualification status, PE250 sequencing was conducted on NovaSeq6000 platform.
Bioinformatics analysis involved initial splicing of PE reads based on overlap relationship along with quality control filtering to obtain high-quality sequences. Subsequent steps included sample distinction analysis as well as clustering analysis for species taxonomy determination based on OTU clustering results. Diversity index analysis was also performed. Based on the taxonomic information, statistical analyses of community structure were conducted at each taxonomic level. Multivariate analysis and significance tests were performed on the community composition and phylogenetic information of multiple species based on the aforementioned analyses.
2.4. Screening of Rhizosphere Growth-Promoting Bacteria in H. arenarium(L.)Moench.
2.4.1. Isolation and Purification of Bilberry Rhizosphere Cultureable Bacteria
The collected rhizosphere soil was air-dried under natural conditions, refined using a sterile mortar, and weighed 1 g per rhizosphere soil sample. Subsequently, 9 ml of sterile water was added to the sample which was then mixed using a vortex mixer for 30 minutes. The mixture was diluted with gradient dilutions (10-2, 10-3, 10-4, 10-5, and 10-6) using the dilution coating method. A suspension of 100 μL was coated onto LB, PDA, and Gau I medium[13] followed by incubation at 28℃ for seven days. Single colonies were selected from these cultures for purification and numbering purposes before being stored in a refrigerator at an inclined surface with a temperature of 4℃.
2.4.2. Analysis of Phosphorus-Solubility Ability of Strains
The obtained strains were inoculated into inorganic phosphorus medium as well as organophosphorus medium; each pure culture strain was inoculated into three different mediums to observe phosphorus solubilization circles at a temperature of 28°C over seven days. Strains exhibiting significant phosphate-solubilizing circles were screened based on measurements taken for both the diameter of the phosphorus solubilizing circle (D) and colony diameter (d). The ratio between these two measurements determined the strength of their phosphorus solubilizing ability[14].
2.4.3. Nitrogen-Fixing Bacteria Screening
The isolated strains were streaked onto Ashby medium using sterile toothpicks followed by incubation at a temperature of 28°C for seven days. Bacterial colonies displaying viscous translucent growth with white or brown coloration or dark brown coloration indicated nitrogen-fixing ability which was determined based on their growth diameters.
2.4.4. Screening of Iron-Producing Carrier Strains
The obtained strains were streaked onto CAS detection medium; each pure culture strain was inoculated at 28 ℃ for 7 days with three duplicates.Subsequently, the nitrogen fixation circle was observed, and strains exhibiting a significant orange chelating ring were selected. The iron-producing capacity of the strain was determined based on the size of the orange chelate ring [15].
2.4.5. Screening of Potassium Solubilizing Bacteria
The strain was then inoculated onto silicate bacteria medium using the dot method and incubated at 28 ℃ for 7 days. The yellow halo surrounding each colony was monitored every 24 hours. After 7 days, the potassium solubility of the strain was preliminarily assessed according to halo size.
2.5. Molecular Biology Identification of Growth-Promoting Bacteria
Selected growth-promoting bacteria were subjected to DNA extraction and amplification of their 16SrDNA region. Amplification products were analyzed by electrophoresis and sent for sequencing to Shanghai Shenggong Sequencing Company. Following completion of sequencing, alignment analysis was performed on NCBI website (www.ncbi.nlm.nih.gov) using MEGA11 software to construct a phylogenetic tree.

Table 1.
The main instruments and equipment used in this experiment.
| Instrument and equipment | Model number | Purchasing company |
|---|---|---|
| Ultra-clean table | SW-CJ-1F | Suzhou Antai Air Technology Co., LTD |
| Biochemical incubator | SPX-250BF | Shanghai Fuma test Equipment Co., LTD |
| autoclave | HVE-50 | HIRAYAMA Corporation of Japan |
| PCR apparatus Gel imager |
DL9700 GellDoc GO |
Donglin Changsheng Limited company American Bole limited |
| Optical microscope | CH20BIMF200 | Olympus Corporation of Japan |
| centrifuge | TGL-16B | Shanghai Anting scientific instrument factory |
Table 2.
Strain identification and related primers sequence.
| Amplified gene | primer | Primer sequence | Fragment size (bp) |
|---|---|---|---|
| 16S rDNA | 27F | 5’-AGAGTTTGATCCTGGCTCAG-3’ | 1500 |
| 1492R | 5’-GGTTACCTTGTTACGACTT-3’ |
Table 3.
PCR reaction system.
| Bulk volume | 30µL |
|---|---|
| ddH2O | 11µL |
| 27F | 1µL |
| 1492R | 1µL |
| Taq enzyme | 15µL |
| Template DNA | 2µL |
Table 4.
PCR reaction conditions.
| Reaction step | time |
|---|---|
| Predenaturation at 94 ℃ | 5min |
| 94 ℃ denaturation | 30s |
| Annealing at 54 ℃ | 30s |
| 72 ℃ extension | 30s |
| It extends after 72 ℃ | 7min |
| Store at 4 ℃ | Hold |
3. Results
3.1. rhizosphere Soil Nutrients
The soil at H. arenarium (L.) Moench sample site consisted mainly of gravel with a content of approximately 13.14%. Soil water content ranged from 5.12% to 6.21%, while pH levels varied between neutral and slightly acidic (6.68-7.06). Nutrient elements and organic matter content in soil were relatively low; alkaline dissolved nitrogen measured at around 68.6 mg/kg, organic matter at approximately 24.l g/kg, effective phosphorus at about8 .3mg/kg , quick-acting potassium recorded as129 mg/kg ,and total nitrogen estimated as1 .26g /kg.The sandy gravel soil provided suitable conditions for growth of Helichrysum arenarium (L.) Moench., which exhibited lower demand for nitrogen and phosphorus along with low organic matter content due to its well-drained nature.
Table 1.
rhizosphere soil test results of Helichrysum arenarium (L.) Moench. sampling sites.
| Soil Sample | HN (mg/kg) | AP (mg/kg) | AK (mg/kg) | OM (g/kg) | TN (g/kg) |
| H. arenarium (L.) Moench. | 68.6 | 8.3 | 129 | 24.1 | 1.26 |
3.2. Sequencing Data Evaluation
After high-throughput sequencing data obtained from rhizosphere soil samples underwent splicing, quality control measures including chimera removal [16,17], it was found that more than90 % of bases had values greater than or equal to 20(Q20) and 30 (Q30), while over90 %had values greater than the total bases, and the GC content was also in the range of 50-60%,which indicated that the processed data were qualified for analysis in the next step.
Table 2.
Results of data analysis of H. arenarium (L.) Moench. sequencing samples.
| Sample | GC | Q20 | Q30 |
| SS | 57.88 | 97.92 | 93.58 |
3.3. Sequencing Data Accuracy and Sequencing Volume Assessment
The dilution curve was constructed by randomly extracting the sequences obtained from the samples, the number of sequences obtained and the number of OUTs that can be represented based on that number of sequencing entries [18,19,20]. As can be seen in Figure 2, the curve flattens out with increasing amount of sequencing. It indicates that the number of OUTs, Shannon index and Simpson index are close to saturation, and the current sequencing amount is reasonable, which can reflect the diversity information of most of the bacteria in the samples to be tested, and can be used for subsequent data analysis.
3.4. Alpha Diversity of Rhizosphere Bacterial Microbial Community
The diversity index (Shannon index) and richness index (Simpson index and Chao l index) of bacteria and fungi in plant rhizosphere soil were significantly different from those in corresponding soil samples (P>0.05), and the differences were significant (P <0.05). There were significant differences in the diversity and richness of microbial communities in H. arenarium (L.) rhizosphere soil.
Table 3.
α diversity of microbial community in rhizosphere soil of H. arenarium (L.) Moench.
| Sample | chao1 | shannon | simpson | dominance | goods_coverage | pielou_e |
| SS(bacteria) | 3037.445±531.36 | 10.525±0.22 | 0.999±0.0004 | 0.001±0.0004 | 0.986±0.0053 | 0.918±0.003 |
| SS(fungi) | 561.318±58.923 | 6.486±0.264 | 0.962±0.018 | 0.038±0.018 | 1 | 0.711±0.028 |
3.5. Similarities and Differences in Soil Microbial Community Composition
There were differences in the structure of the soil microbial community between the roots of H. arenarium (L.) Moench. at the OTU level, as shown by the results of NMDS analysis (Figure a, b),.
The PCA principal component analysis fully reflects the difference and similarity in the composition of the rhizosphere communities of different samples. The points of different colors or shapes represent different samples, and the closer the two points are the more similar the species composition of the two samples is[21]. When PC1 was 9.11%(Figure c), from the sample species, two of the H. arenarium (L.) Moench. rhizosphere bacterial community samples showed a long distance from other samples, indicating that there were certain differences in the species composition of the samples, but the other samples were relatively close to each other, with a very high correlation. The distance of repeated samples in each group was relatively close, and the complexity of spatial distribution of flora was relatively uniform.When PC1 was 14.15% (Figure d), from the perspective of sample species, most samples of rhizosphere fungal communities were very close to each other, indicating that the species composition of samples was similar and the correlation was extremely high. Two samples were far apart and had great differences. The distance of repeated samples in each group was relatively close, and the complexity of spatial distribution of flora was relatively uniform.
The PCoA analysis presents the differences in community species composition between samples from the similarity between distances, different color or shape of points represent different samples, the closer the distance between two points in space indicates the more similar species composition between samples. When PC1 was 62.45% and PC2 was 30.72%(Figure e,f), the bacterial community of each sample of H. arenarium (L.) Moench. has obvious aggregation, and the distance is extremely close to each other, which indicates that there is no obvious difference in the bacterial flora of the microbes; each data point in the figure represents a sample, and the distribution of the data points in Figure c showed that the bacterial community composition of each rhizosphere sample is centralized, but a few of them have large differences. When PC1 was 13.41%(Figure g,h), the rhizosphere soil fungal community presents a certain distance between each sample, indicating that there was a certain degree of variability among the sample species compositions; when PC2 was 21.77%, the rhizosphere fungal community aggregation of three samples have a large difference with the other samples of the fungal community.
Each data point in the figure represents was a sample, and the distribution of data points in the figure showed that the microbial community composition of each rhizosphere sample of H. arenarium (L.) Moench. was relatively decentralized, but a few have large differences.
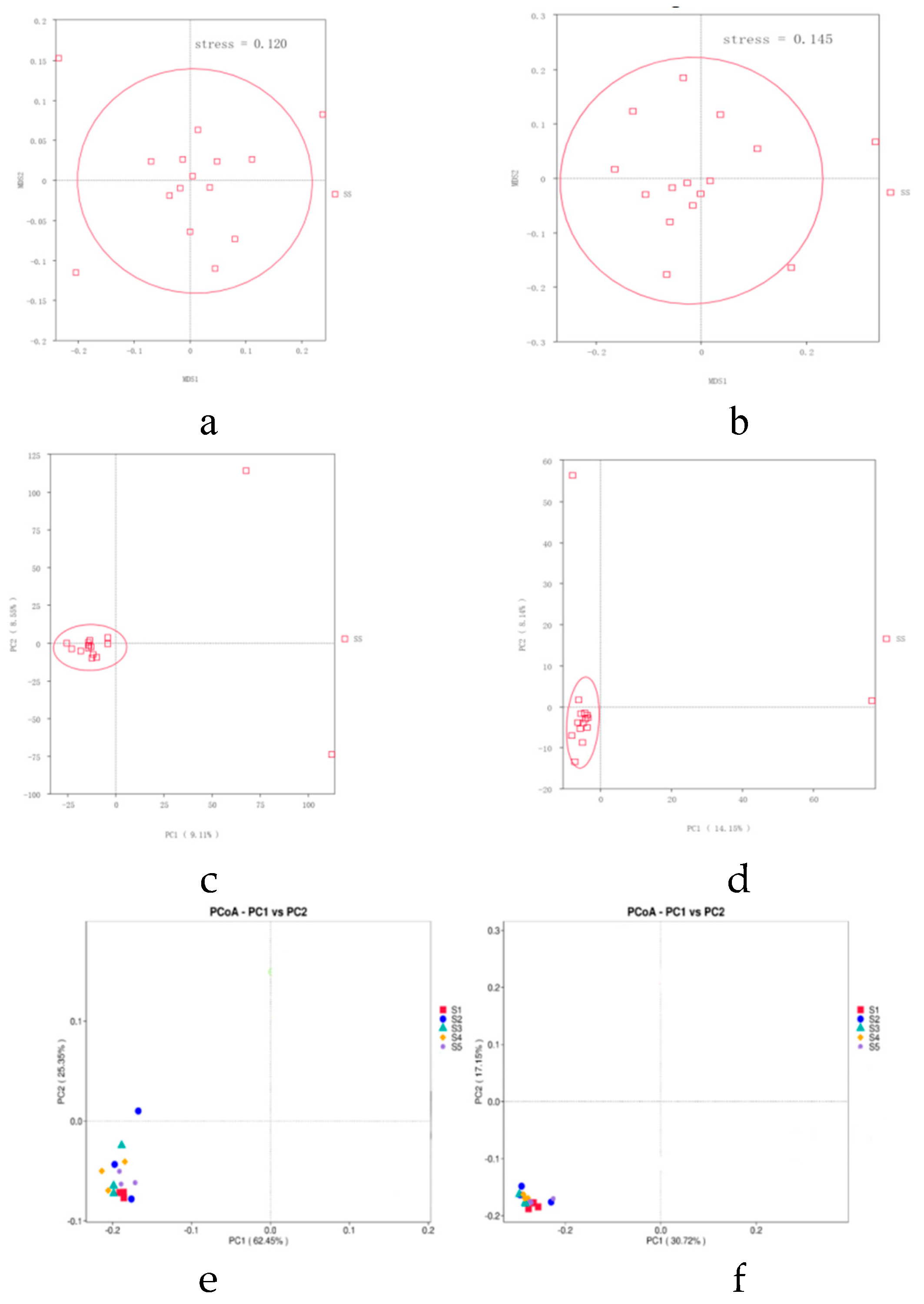
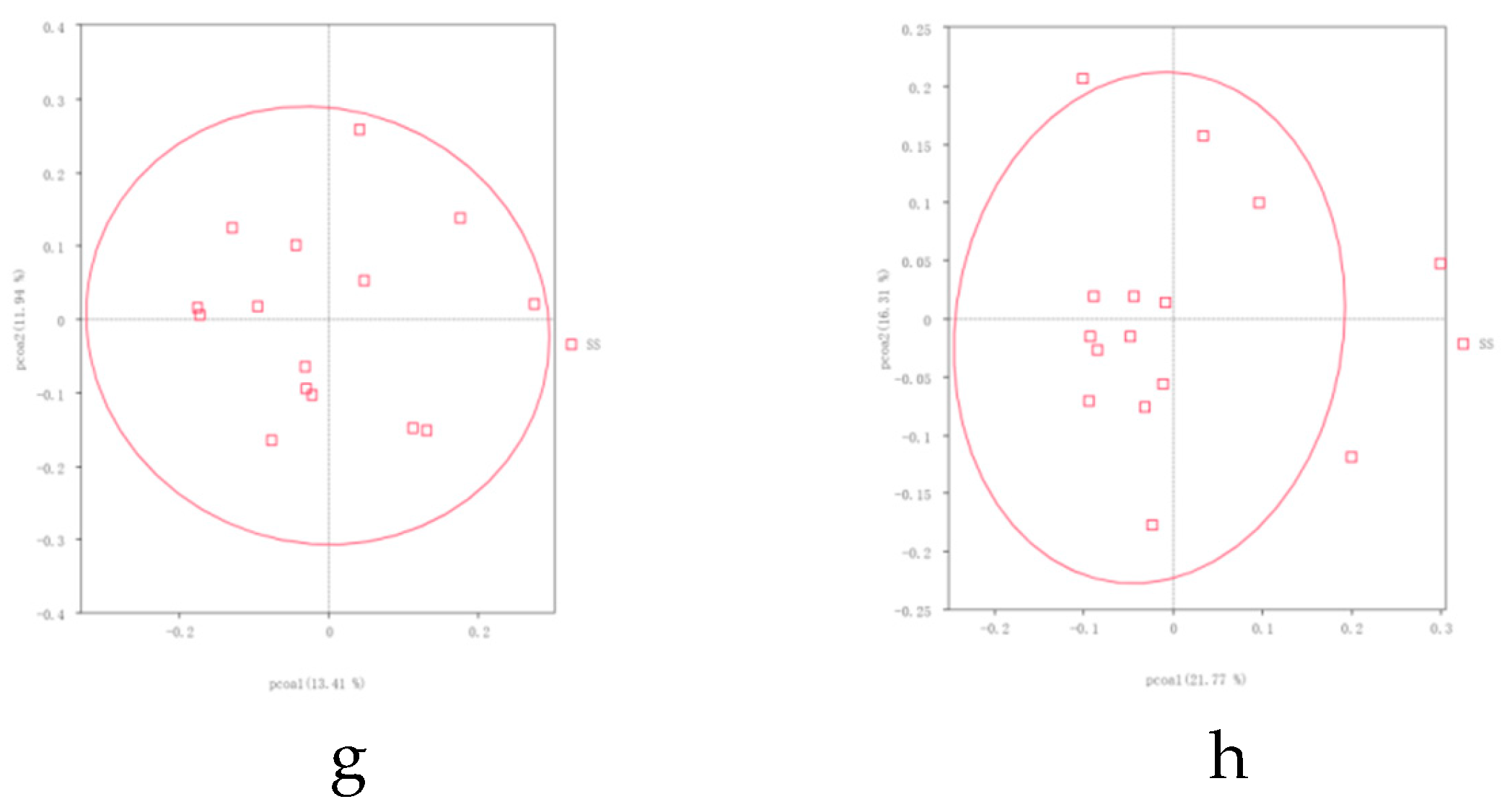
Figure 3.
(a) H. arenarium (L.) Moench. bacterial NMDS map; (b) H. arenarium (L.) Moench. fungal NMDS map; (c) H. arenarium (L.) Moench. bacterial PCA distribution; (d) H. arenarium (L.) Moench. fungal PCA distribution; (e) and (f) H. arenarium (L.) Moench. bacterial PCoA distribution; (g) and (h) H. arenarium (L.) Moench. fungal PCoA distribution;.
Figure 3.
(a) H. arenarium (L.) Moench. bacterial NMDS map; (b) H. arenarium (L.) Moench. fungal NMDS map; (c) H. arenarium (L.) Moench. bacterial PCA distribution; (d) H. arenarium (L.) Moench. fungal PCA distribution; (e) and (f) H. arenarium (L.) Moench. bacterial PCoA distribution; (g) and (h) H. arenarium (L.) Moench. fungal PCoA distribution;.
3.6. Numerical Analysis of OTUs
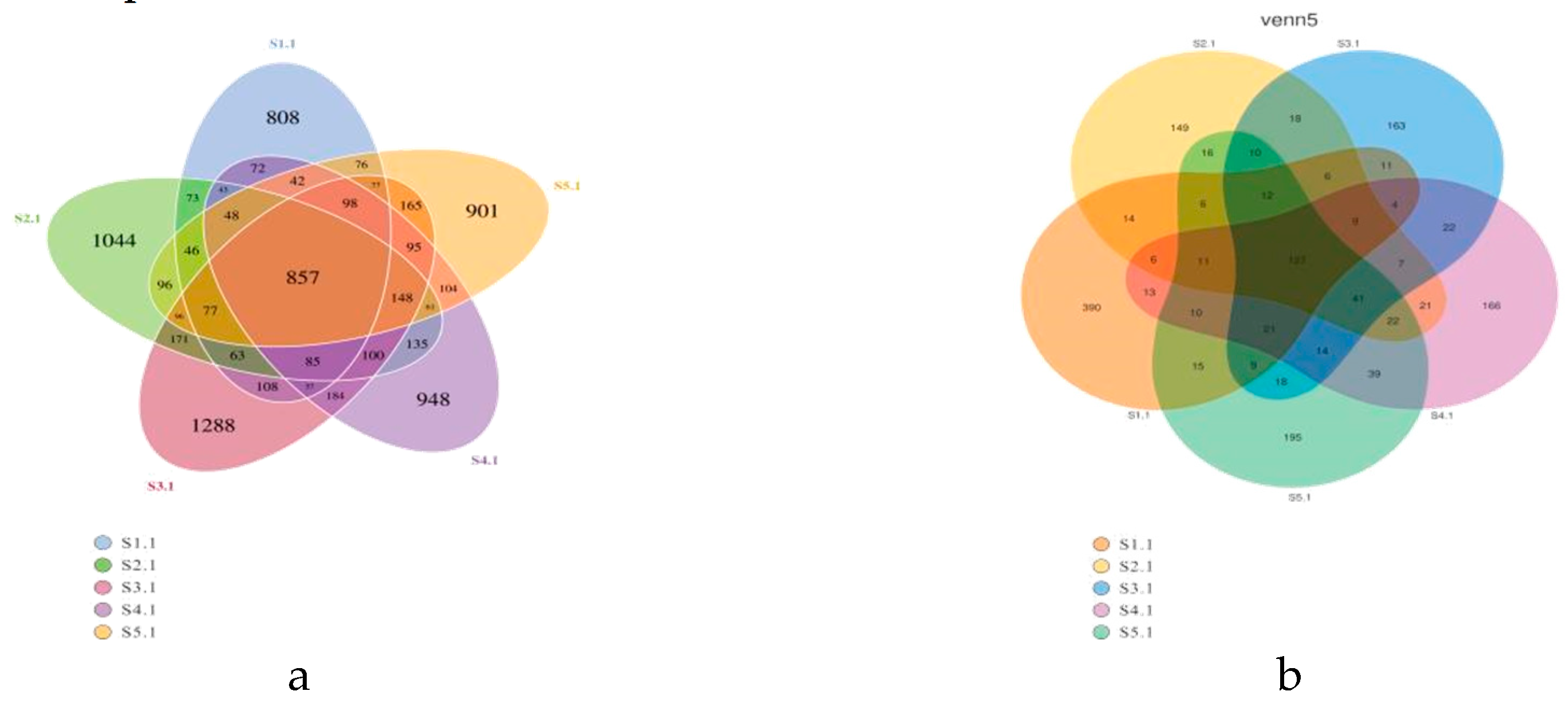
The results of the analysis of the differences in the rhizosphere microbial communities of the samples are shown in Figure 4. The rhizosphere microbial communities of the samples were rich and differentiated, with specific OUT values for each sample.
There were 8,261 OTUs in the rhizosphere soil bacteria, and all soil samples have shared and unique OUTs with each other, in which sample S1.1 has 808 unique OTUs, accounting for 9.78%; sample S2.1 has 1044 unique OTUs, accounting for 12.64%; sample S3.1 has 1288 unique OTUs, accounting for 15.59% ; sample S4.1 is 948, accounting for 11.48%; the value of OTUs specific to sample S5.1 is 901, accounting for 10.91%.
There were 1,465 in the rhizosphere soil fungi in H. arenarium (L.) Moench., and there were common and unique OUT numbers between each soil sample, in which sample S1.1 were 390, accounting for 26.62%; sample S2.1 were 149, accounting for 10.17% of the total number of OUTs; sample S3.1 were 163, which was 11.13% of the total number of OUTs; sample S4.1 were 166, which was11.33%; sample S5.1 were 195, which was 13.31% .
3.7. Analysis of Bacterial Species Abundance
The results of high-throughput amplification sequencing showed that the soil bacteria in the rhizosphere area of H. arenarium (L.) Moench. belonged to 35 phylums, 93 classes, 215 orders, 324 families, 521 genera and 231 species(Table 4). There were seveal bacterial communities of H. arenarium (L.) Moench. with relative abundance more than 1% at the phylum level.They were Actinobacteriota, accounting for 41.30%; Proteobacteria, accounting for 14.08%; Acidobacteriota, accounting for 8.32%; Verrucomicrobiota, accounting for, 7.86%; Gemmatimonadota, accounting for 9.21%; Bacteroidota, accounting for 3.02%; Chloroflexi, accounting for 7.47% and Planctomycetota, accounting for 1.45%. There were seveal bacterial communities with relative abundances more than 1% at the genus level. They were Arenimicrobium_luteum, accounting for 6.11%; 67-14, accounting for 4.48%; Gemmatimonas, accounting for 3.26%; MB-A2-108, accounting for 3.26% and RB41, accounting for 2.67%. The Actinobacteria was dominated at the phylum level.
The results of high-throughput amplification sequencing results showed that the soil fungi in the rhizosphere area of H. arenarium (L.) Moench. belonged to 14 phylums, 47 classes, 96 orders, 204 families, 378 genera and 571 species.(Table 4) There were seveal fungl communities of H. arenarium (L.) Moench. with relative abundance more than 1% at the phylum level. They were Basidiomycota accounting for 7.80%, Ascomycota accounting for 67.21%, Mortierellomycota accounting for 12.05%, and Chytridiomycota accounting for4.61%. There were seveal fungal communities of H. arenarium (L.) Moench.were more than 1% at the genus level.They were Talaromyces accounting for 2.94%, Knufia accounting for 3.80%, Tetracladium accounting for 1.28% and Thyrostroma accounting for 1.73%. At the phylum level Ascomycota was dominated.
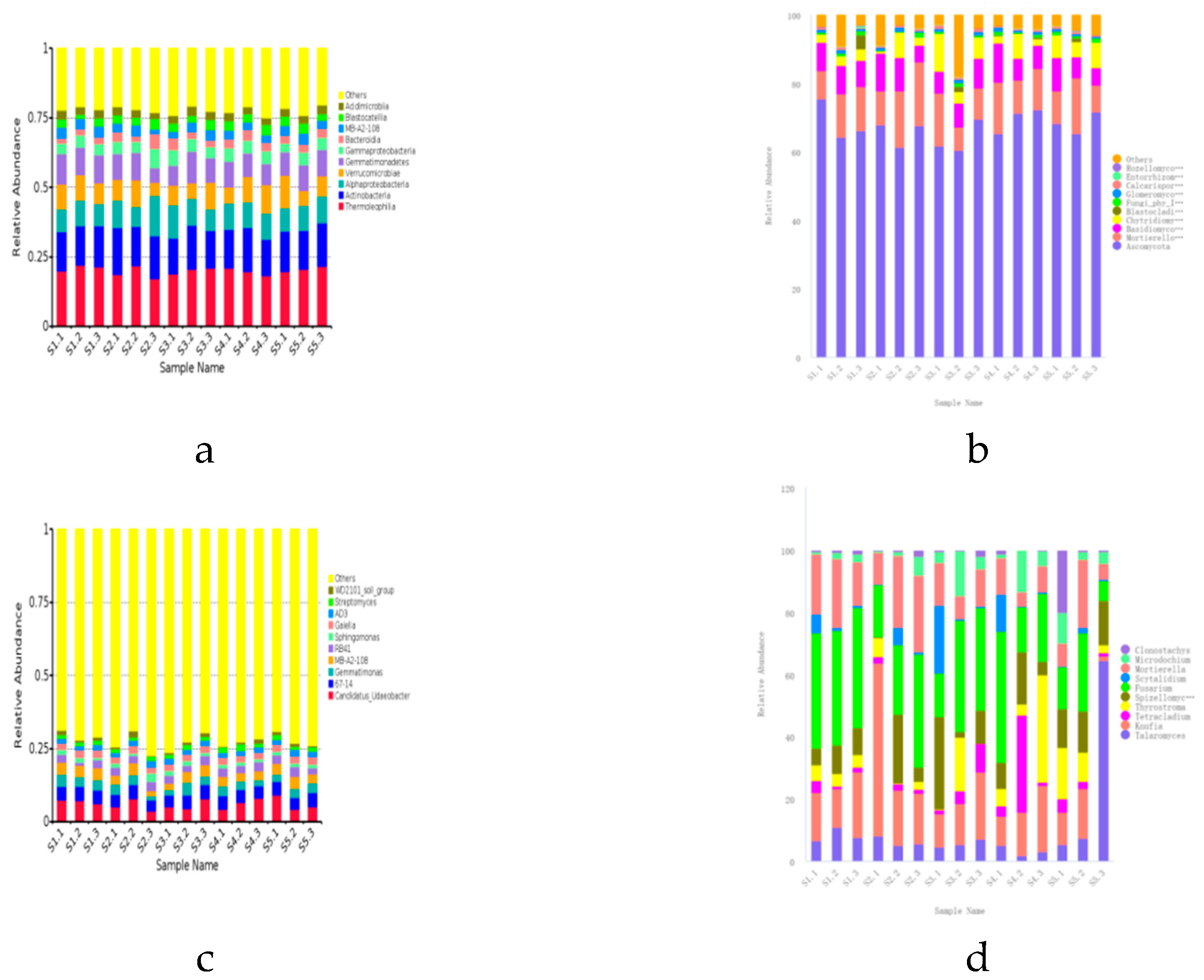
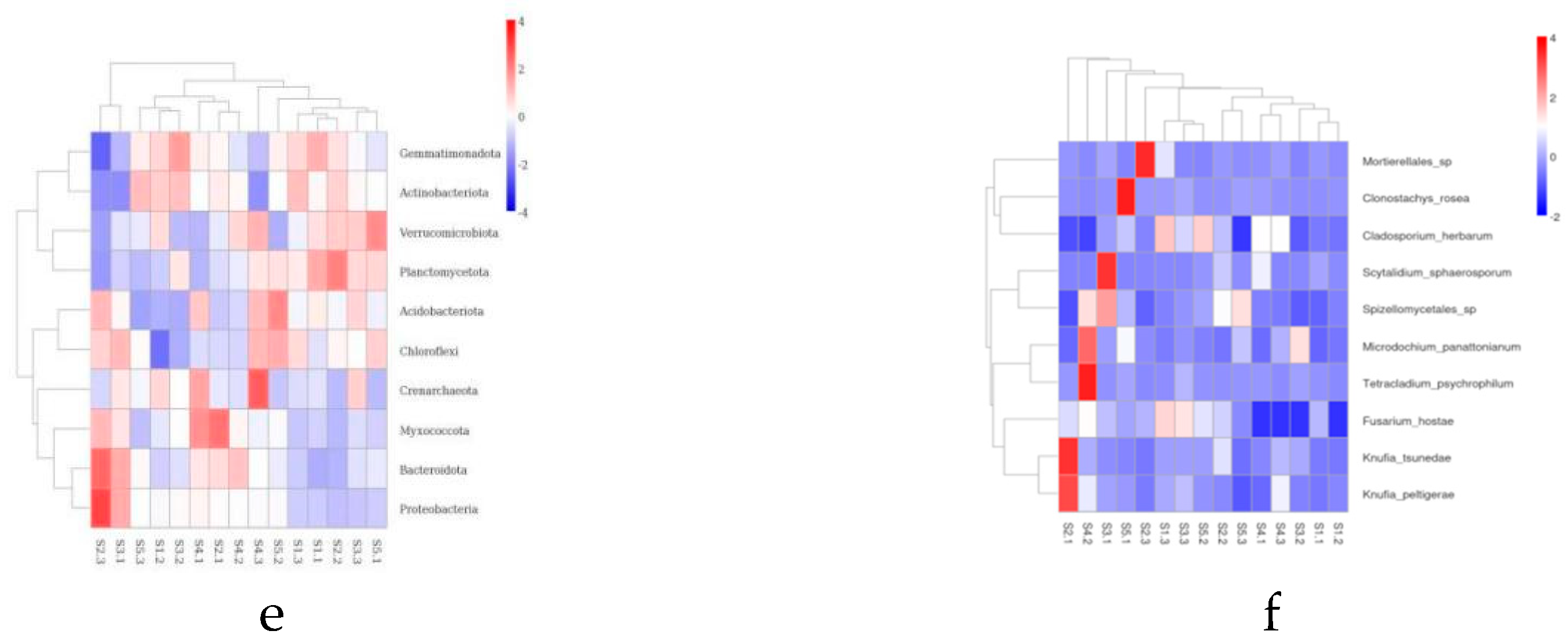
Table 4.
Species distribution of habitat and rhizosphere H. arenarium (L.) Moench. soil microbe at the taxonomic level.
Table 4.
Species distribution of habitat and rhizosphere H. arenarium (L.) Moench. soil microbe at the taxonomic level.
| Component | Kingdom | Phylum | Class | Order | Family | Genus | Species |
|---|---|---|---|---|---|---|---|
| SS(bacteria) | 1 | 35 | 93 | 215 | 324 | 521 | 231 |
| SS(fungai) | 1 | 14 | 47 | 96 | 204 | 378 | 571 |
Figure 5.
Histogram of relative abundance of species at phylum and genus level. Note: (a) is a histogram of relative abundance of species at the H. arenarium (L.) Moench. bacterial phylum level, (b) is a histogram of relative abundance of species at the H. arenarium (L.) Moench. bacterial genus level; (c) is a heat map analysis of clustering of bacterial species abundance, (d) Species evolutionary tree at bacterial genus level.; (e) is a histogram of relative abundance of species at the H. arenarium (L.) Moench. fungal phylum level, (f) is a histogram of relative abundance of species at the H. arenarium (L.) Moench. fungal genus level; (g) is a heat map analysis of clustering of fungal species abundance,(h) Species evolutionary tree at fungal genus level.
Figure 5.
Histogram of relative abundance of species at phylum and genus level. Note: (a) is a histogram of relative abundance of species at the H. arenarium (L.) Moench. bacterial phylum level, (b) is a histogram of relative abundance of species at the H. arenarium (L.) Moench. bacterial genus level; (c) is a heat map analysis of clustering of bacterial species abundance, (d) Species evolutionary tree at bacterial genus level.; (e) is a histogram of relative abundance of species at the H. arenarium (L.) Moench. fungal phylum level, (f) is a histogram of relative abundance of species at the H. arenarium (L.) Moench. fungal genus level; (g) is a heat map analysis of clustering of fungal species abundance,(h) Species evolutionary tree at fungal genus level.
3.8. Rhizosphere Growth-Promoting Bacteria Screening
3.8.1. Phosphorus-Solubilizing Ability Screening
Phosphorus-solubilizing cycle method was used to screen the phosphorus-solubilizing ability of soil rhizosphere bacteria isolated from H. arenarium (L.) Moench..The experimental results showed that there were two strains of bacteria that produced phosphorus solubilizing circles, namely S29 and S16, and the inorganic phosphorus soluble index were 1.18 and 1.23(Table 5). At the same time,there were no strains with organophosphorus dissolving ability.

Figure 6.
Colony morphology of phosphorus zing medium.
Table 5.
The ability of the strain to dissolve inorganic phosphorus.
| Strain number | Colony diameter (d) /㎝ | Hydrolytic ring diameter (D) /㎝ | Solubility index (D/d) |
| S29 | 1.3 | 1.53 | 1.18 |
| S16 | 0.43 | 0.53 | 1.23 |
3.8.2. Screening of Nitrogen Fixation Capacity of Strains
The experimental results showed that, two strains could fix nitrogen to S7 and S10 and there were obvious transparent circles on the medium of Ashu shellfish. The soluble index were 1.10 and 1.23(Table 6)

Figure 7.
Colony morphology of nitrogen fixation medium.
Table 6.
The ability of the strain to dissolve inorganic phosphorus.
| Strain number | Colony diameter (d) /㎝ | Hydrolytic ring diameter (D) /㎝ | Solubility index (D/d) |
| S7 | 0.3 | 0.33 | 1.10 |
| S10 | 0.43 | 0.53 | 1.23 |
3.8.3. Screening of Siderophore Production Capacity of Strains
A total of tseven strains with good iron-bearing capacity were screened with orange chelation circles on CAS medium. They were S2-1, S3, S13, S14, S16, S18 and S29. The D/d value of S16 was up to 2.96, indicating that S16 had the strongest iron-bearing capacity(Table 7).
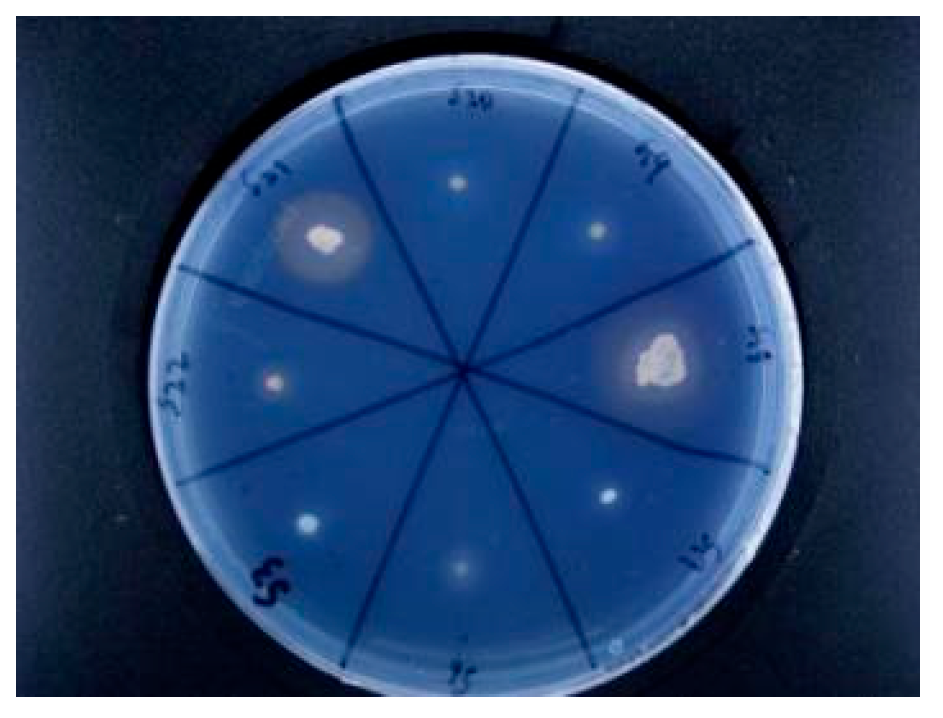
Figure 8.
Colony morphology of CAS detection medium.
Table 7.
Ability of strain to produce iron carrier.
| Strain number | Colony diameter (d) /㎝ | Hydrolytic ring diameter (D) /㎝ | Solubility index (D/d) |
| S2-1 | 0.15 | 0.17 | 1.13 |
| S3 | 0.21 | 0.34 | 1.62 |
| S7 | 0.14 | 0.18 | 1.29 |
| S10 | 0.17 | 0.33 | 1.94 |
| S13 | 0.7 | 1.53 | 2.19 |
| S14 | 0.25 | 0.31 | 1.24 |
| S16 | 0.53 | 1.57 | 2.96 |
| S18 | 0.53 | 1.5 | 2.83 |
| S29 | 0.27 | 0.4 | 1.48 |
3.8.4. Screening of Potassium-Solubilizing Ability of Strains
Strains with potassium-sowing ability were not screened.
3.9. Construct a Phylogenetic Tree
Genomic DNA was extracted from nine grown-promoting strains, 16SrDNA sequence was determined and compared on NCBI website. MEGA11 was used to construct phylogenetic tree. The sequencing results showed that the nine growth-promoting strains were all higher than 99%. They belong to the genus Bacillus under Firmicutes, the genus Alcaligenes under Proteobacteria, and the genus Kocuria under Actinomyces, of which five belong to the genus Bacillus, three belong to the genus Alcaligenes, and one belong to the genus Kocuria. In the genus Bacillus, strain S2-1 was preliminarily identified as Priestia megaterium, strain S3 as Bacillus mobilis and strain S10 as Bacillus bataviensis. Strain S14 was identified as Bacillus mycoides and strain S29 as Bacillus paramobilis. Strains S18, S13 and S16 of the genus Alcaligenes were identified as Alcaligenes faecalis. In the genus Kocuria, S7 was tentatively identified as Kocuria rosea.

Figure 9.
Phylogenetic tree based on 16SrDNA.
4. Discussion
The microbial community structure in the rhizosphere soil is crucial for H. arenarium (L.) Moench. to obtain nutrients. So, we studied and analyzed the microbial community structure in the rhizosphere soil as well as the screening of growth promotion and antagonistic bacteria. Then we found that in the rhizosphere soil environment, bacteria had a higher total number of species, community diversity and abundance than fungi, indicating that the root environment was still bacteria-dominated, which was consistent with the view that soil microorganisms were mainly bacteria[19,20], higher bacterial diversity indicates higher soil resistance[21]. Based on the analysis results of OTUs, there are eight main bacterial phyla in the rhizosphere microbial community (relative abundance greater than 1%), among which Actinobacteriota and Proteobacteria are the dominant phyla, especially the relative abundance of Proteobacteria accounts for more than 40% of all phyla. Proteobacteria are enriched in the rhizosphere soil of alfalfa[22], strawberry[23], potato[24] and asparagus[25], indicating that Proteobacteria can adapt to the rhizosphere environment of various plants and are the main components of rhizosphere bacteria. However, the rhizosphere soils of different plants showed species characteristics for microbial diversity. The rhizosphere soils of the plant grew in mounds, semi-dunes, slopes of wet saline soil, gravel soil, sand dunes, grassland and under pine forests at an altitude of 900 m-2,400 m, with pH ranging from 6.68 to 7.06, mostly in neutral and slightly acidic soil. Based on this special growth environment, Actinobacteriota, Proteobacteria, Acidobacteriota, Verrucomicrobiota, and Gemmatimonadota are the main dominant phyla of H. arenarium (L.) Moench.. Though, soil fungi is fewer than bacteria, it is critical to maintaining ecosystems[26]. There are four major fungal phyla (relative abundance >1%) in the rhizosphere microbial community of H. arenarium (L.) Moench., among which, Ascomycota and Mortierellomycota, Basidiomycota are dominant phyla, and especially Ascomycota is the absolute dominant phyla, with the highest relative abundance accounting for more than 70% of all the phyla, which is is consistent with the study of Li Qingshan et al.[27].Ascomycota is the main decomposers of difficult organic matter such as plant litters and lignin in soil. Ren Neifan et al. isolated a strain of Penicillium cinerea BIBA-G563 from the root of healthy Ruthenica Ruthenica, which belongs to Ascomycota and has a control effect of 75.4% on the root rot of Ruthenica Ruthenica[28]. H. arenarium (L.) Moench. is a root-breeding plant, and its root system provides a large number of attachment points for rhizosphere microorganisms, thus increasing the abundance and diversity of rhizosphere microorganisms and facilitating the enrichment of dominant flora.
Rhizosphere growth-promoting bacteria promote plant nutrient absorption through biological nitrogen fixation, phosphorus solubilization, potassium solubilization, secretion of plant hormones and production of iron carriers. In this study, nine growth-promoting bacteria were screened, which belonged to the genus Bacillus under Firmicutes, the genus Alcaligenes under Proteobacteria, and the genus Kocuria under Actinomyces, of which five belong to the genus Bacillus, three belong to the genus Alcaligenes, and one belong to the genus Kocuria. Most of the growth-promoting bacteria isolated by previous authors were Bacillus and Pseudomonas, S16 has the strongest ability to detoxify phosphorus and produce iron carriers and belongs to Alcaligenes; S10 has the strongest ability to fix nitrogen and belongs to Bacillus. Nitrogen-fixing bacteria reduce nitrogen to ammonia to increase the N content in the soil; phosphorus-solubilizing bacteria convert insoluble phosphorus in the soil into phosphorus that can be easily absorbed by plants to increase the effective phosphorus content in the soil; iron-carrier-producing bacterial strains promote iron nutrition in plants through the production and utilization of iron carriers[29,30,31]. In the later stage of this study, the optimal growth-promoting bacteria will be screened by combining the strain with the growth-promoting experiment, which will provide an experimental basis for the development and promotion of microbial fertilizer for the mulberry, provide a research basis for the protection of wild resources of the mulberry and the development and utilization of rhizosphere microorganisms, and provide a strong support for the future application of the mulberry in food and medicine.
In the later stage of this study, the optimal growth-promoting bacteria will be screened by combining the strain with the growth-promoting experiment, which will provide an experimental basis for the development and promotion of microbial fertilizer for the H. arenarium (L.) Moench., provide a research basis for the protection of wild resources of the H. arenarium (L.) Moench. and the development and utilization of rhizosphere microorganisms, and provide a strong support for the future application of the H. arenarium (L.) Moench..
5. Conclusions
The rhizosphere microbial communities of H. arenarium (L.) Moench. with relative abundance more than 1% at the phylum level were Actinobacteriota, Proteobacteria, Acidobacteriota, Verrucomicrobiota, Gemmatimonadota, Bacteroidota, Chloroflexi and Planctomycetota; relative abundances more than 1% at the genus level were Arenimicrobium_luteum, 67-14, Gemmatimonas, MB-A2-108 and RB41. The Actinobacteria dominate at the phylum level and the Arenimicrobium_luteum dominate at the genus level
High-throughput sequencing results showed that there were 11 fungal phyla and 11 fungal genera in the rhizosphere soil of Saxifrage. The relative abundance of rhizosphere microbial communities of H. arenarium (L.) Moench. at the phylum level was more than 1% for the phyla, Basidiomycota, Ascomycota, Mortierellomycota, and Chytridiomycota. The relative abundance of rhizosphere microbial communities of H. arenarium (L.) Moench.was more than 1% at the genus level, Talaromyces, Knufia, Tetracladium, Thyrostroma.
The nine growth-promoting strains belong to the genus Bacillus under Firmicutes, the genus Alcaligenes under Proteobacteria, and the genus Kocuria under Actinomyces, of which five belong to the genus Bacillus, three belong to the genus Alcaligenes, and one belong to the genus Kocuria. In the genus Bacillus, strain S2-1 was preliminarily identified as Priestia megaterium, strain S3 as Bacillus mobilis and strain S10 as Bacillus bataviensis. Strain S14 was identified as Bacillus mycoides and strain S29 as Bacillus paramobilis. Strains S18, S13 and S16 of the genus Alcaligenes were identified as Alcaligenes faecalis. In the genus Kocuria, S7 was tentatively identified as Kocuria rosea.The above results provide experimental basis for further understanding and utilization of rhizosphere microbial resources and development of rhizosphere microbial fertilizer of H. arenarium (L.) Moench..
Funding
This research was found by the Research and application of high quality and efficient production technology and equipment of Vaccinium myrtillus L. and Helichrysum arenarium (2023B02023-2).
Acknowledgments
This study was supported by the Research and application of high quality and efficient production technology and equipment of Vaccinium myrtillus L. and Helichrysum arenarium (2023B02023-2).
References
- Xinjiang Flora Editorial Committee. Flora of Xinjiang (Volume IV) [M]. Urumqi: Xinjiang Science, Technology and Health Press, 2004:10.
- Ma Zhongjie, Yang Changyou. Survey of Chinese herbal medicines in Altay region of Xinjiang[J]. China J. Chin. Mater. Medica 2002, 27, 70–71.
- Liu Rui. Research on synergistic development of ethnic economy in Burzin County under the domination of tourism [D]. Shihezi University 2011.
- Czinner E, Hagymási K, Blázovics A, et al. In vitro antioxidant properties of H. arenarium (L.) Moench[J]. J. Ethnopharmacol. 2000, 73, 437–443. [Google Scholar] [CrossRef]
- Wang Libo, Wang Jianwei, Wang Ce, et al. Chemical composition of the lipid-lowering active site of the sandy wax chrysanthemum flower (Ⅲ)[J]. Chin. J. Med. Chem. 2012, 22, 220–222. [Google Scholar]
- LI Yiwei, WANG Ying, LAN Wei. Study on the optimal extraction process of total flavonoids from Bupleurum officinale[J]. Journal of Xinjiang Medical University. 2019, 42, 805–807+812. [Google Scholar]
- Zhang Yuwei. Studies on the chemical composition and bioactivity of the aboveground parts of Corydalis alba and Corydalis alba[D]. Shenyang: Shenyang Pharmaceutical University 2009.
- Kihyuck C, Raees K, Woo S L. Dissection of plant microbiota and plant-microbiome interactions[J]. J. Microbiol. 2021, 59, 281–291. [Google Scholar] [CrossRef]
- WU Chang-Hao, LIU Jing-Ze. Progress in the study of rhizosphere microbial influences and their interactions with plants[J]. J. Hebei Norm. Univ. (Nat. Sci. Ed.) 2022, 46, 603–613. [Google Scholar]
- Garima Y, Mukesh M. Bioprospecting of endophytes in medicinal plants of Thar Desert: An attractive resource for biopharmaceuticals[J]. Biotechnol. Rep. 2021, 345, 89–92. [Google Scholar]
- Santoyo G, Moreno-Hagelsieb G, Orozco-Mosqueda CDM, et al. Plant growth-promoting bacterial endophytes[J]. Microbiol. Res. 2016, 128, 92–99. [Google Scholar]
- Huang Xuezhen, Zhao Longfei. Progress on the mechanism of endophytic bacteria of medicinal plants on host plants[J]. Microbiol. Bull. 2023, 50, 1653–1665. [Google Scholar]
- Wang, Y.M.; Fu, J.H.; Qin, X.Z.; Wang, J.; Ruan, W.W.; Cui, F.Z.; Nie, H.L. Isolation, screening and growth promoting characteristics of growth-promoting bacteria from mossy crusty soil in Gurbantunggut Desert [J]. J. Microbiol. 2019, 43, 47–57. [Google Scholar]
- Wang X X, Chen J Y, Fan C, et al. Screening, identification, salt tolerance and growth promotion effect of two strains of phosphorus solubilizing fungi[J]. Jiangsu Agric. Sci. 2024, 52, 219–226. [Google Scholar]
- Du, L.; Wang, S.P.; Chen, G.; Hong, J.; Huang, X.; Zhang, L.H.; Ye, L.X; Lian, Z.C.; Zhang, G.Y. Screening, identification and phosphorus solubilization capacity of a highly efficient phosphorus solubilizing bacterium [J]. China Soil Fertil. 2017, 23, 136–141. [Google Scholar]
- LI Jie, Wang Fei, Xu Lingchuan. High-throughput analysis of soil bacterial community structure in the rhizosphere zone of Taishan white foxglove[J]. Guangdong Agric. Sci. 2022, 49, 75–84. [Google Scholar]
- Zhang Chen. Structural analysis of endophytic and rhizosphere microflora and saponin biotransformation in American ginseng [D]. Zhengzhou University 2021.
- Garima Y, Mukesh M. Bioprospecting of endophytes in medicinal plants of Thar Desert: An attractive resource for biopharmaceuticals[J]. Biotechnol. Rep. 2021, 345, 89–92. [Google Scholar]
- Santoyo G, Moreno-Hagelsieb G, Orozco-Mosqueda C D M, et al. Plant growth-promoting bacterial endophytes[J]. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Huang Xuezhen, Zhao Longfei. Progress on the mechanism of endophytic bacteria of medicinal plants on host plants[J]. Microbiol. Bull. 2023, 50, 1653–1665. [Google Scholar]
- Chen Ke, Chen Min, Lin Li et al. Characterization of microbial communities contaminated by nine traditional Chinese medicine tablets based on 16S rRNA high-throughput sequencing[J]. J. Tradit. Chin. Med. 2022, 28, 53–56. [Google Scholar]
- Wang Fang, Li Wei, Liu Xin, et al. Response of rhizosphere soil bacterial community of alfalfa to decomposed cow manure [J]. Acta Acreologica Sin. 2022, 30, 603–611. [Google Scholar]
- Wang Qing-Feng, Zhou De-Ping, Chu Chang-Bin, et al. Difference of rhizosphere bacterial community structure and function between healthy and anthracnose infected strawberries [J]. Chin. J. Soil Sci. 2022, 53, 1404–1412. [Google Scholar]
- Ge Yinglan, Sun Ting. Microbial community structure and diversity in potato rhizosphere and non-rhizosphere soil [J]. Chin. J. Ecol. Environ. 2020, 29, 141–148. [Google Scholar]
- Sheng Yuzhen, Huang Ling, Ye Pengsheng, et al. Microbial community structure and diversity in rhizosphere and non-rhizosphere soil of asparagus [J]. J. Sichuan Agric. Univ. 2024, 42, 330–338. [Google Scholar]
- Zhao Ji, Liao Yangnan. Characteristics and effects of decomposer subsystem in Leymus chinensis grassland [J]. Acta Ecol. Sin. 1995, 15, 359–364. [Google Scholar]
- Li Qingshan, Wang Mengjiao, Liu Jieran. Diversity and differences of rhizosphere soil microbial community structure in different periods of rapeseed [J]. Northern Horticulture 2023, 72–80.
- Ren Naipeng, Liu Xiangping, Tang Jialin, et al. Effect of Penicillium griseus BIBA-G563 on Fusarium root rot of alfalfa [J]. Chin. J. Grassl. Sci. 2023, 45, 95–103. [Google Scholar]
- Persello-Cartieaux F, Nussaume L, Robaglia C. Tales from the underground: molecular plant–rhizobacteria interactions[J]. Plant Cell Environ. 2003, 26, 189–199. [Google Scholar] [CrossRef]
- Jeffries P, Gianinazzi S, Perotto S, et al. The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility[J]. Biol. Fertil. Soils 2002, 37, 1–16. [Google Scholar]
- Ren Xin-Ping, Li Sheng, Qi Yi-Lin, et al. Screening, identification and application of iron-producing carrier fungus Ti-11 [J]. J. Hunan Univ. Technol. 2023, 37, 15–21. [Google Scholar]
Figure 2.
Dilution curves. Note: a H. arenarium (L.) Moench. Chao1 curve; b H. arenarium (L.) Moench. Shannon curve; c H. arenarium (L.) Moench. Simpson dilution curve.
Figure 2.
Dilution curves. Note: a H. arenarium (L.) Moench. Chao1 curve; b H. arenarium (L.) Moench. Shannon curve; c H. arenarium (L.) Moench. Simpson dilution curve.
Figure 4.
Venn diagram of OUT distribution of H. arenarium (L.) Moench. Note: a is bacteria, b is fungi;.
Figure 4.
Venn diagram of OUT distribution of H. arenarium (L.) Moench. Note: a is bacteria, b is fungi;.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



