
Submitted:
21 August 2024
Posted:
23 August 2024
You are already at the latest version
Abstract
The aim of this study was to identify differentially expressed lncRNAs (DElncRNAs) and mRNAs (DEmRNAs) in the endometrium of individuals with and without EMS during the proliferative (P) and secretory (S) phases of the menstrual cycle. Tissues were obtained from 18 control (CT; P-phase [pCT], n=8; S-phase [sCT], n=13) and 23 EMS patients (P-phase [pEMS], n=13; S-phase [sEMS], n=12). DElncRNAs and DEmRNAs were analyzed using total RNA-sequencing. In P-phase, expression of NONHSAG019742.2 and NONHSAT120701.2 was significantly higher in EMS than control patients, that of while NONHSAG048398.2 and NONHSAG016560.2 was lower in EMS patients. In S-phase, expression of NONHSAT000959.2, NONHSAT203423.1, and NONHSAG053769.2 was significantly increased in EMS patients, while that of NONHSAG012105.2 and NONHSAG020839.2 was lower. In addition, the expression of HSD11B2, THBS1, GPX3, and SHISA6 was similar to that of neighboring lncRNAs in both P- and S-phases. In contrast, ELP3 and NR4A1, respectively, were up- or downregulated in pEMS tissues. In sEMS, expression of LAMB3 and HIF1A was increased, while expression of PAM was reduced. Our findings on lncRNAs and mRNAs encourage not only to explore the potential clinical applications of lncRNAs and mRNAs as prognostic or diagnostic biomarkers for EMS but also to gain valuable insights into its pathogenesis.
Keywords:
Endometriosis
; endometrium
; menstrual cycle
; long noncoding RNA
; neighboring gene
1. Introduction
Endometriosis (EMS) is a chronic gynecological disease in which endometrial tissue, which is normally located inside the uterus, grows outside of the uterus. This abnormal tissue may be found on or within various organs and structures, including the fallopian tubes, ovaries, or peritoneum. Both eutopic and ectopic uterine endometrial tissues respond to hormonal signals to grow, thicken, and break down in a similar manner during the menstrual cycle; however, ectopic endometrial tissue is not expelled from the body during menstruation as is normal endometrial tissue, and its presence may trigger inflammation and scar tissue formation (including adhesions) and may also impact the endometrial tissue in the uterus. EMS occurs in 10 to 15% of women of reproductive age and causes various symptoms such as pelvic pain, lower or abdominal pain, pain with bowel movements or urination, pain with intercourse, longer and heavier menstrual bleeding, and infertility [1,2,3]. Therefore, a better understanding of the etiology and pathophysiology of EMS is needed to identify biomarkers and to develop more effective strategies to prevent, diagnose and treat EMS and prevent relapses; such investigations should include genomic assessment.
The human genome is composed of coding and noncoding DNAs that are transcribed to coding and noncoding RNAs, respectively. Coding messenger RNAs (mRNAs) are translated into proteins. Among noncoding RNAs (ncRNAs), there are three main categories based on size: small RNAs (17−25 nucleotides), which include microRNAs (miRNAs); mid-size RNAs (20−200 nucleotides); and long ncRNAs (lncRNAs), consisting of more than 200 nucleotides [4]. The novel high-throughput RNA-sequencing (RNA-Seq) technology has revolutionized transcriptomics, allowing discovery of known and unknown transcripts and noncoding RNAs [5,6,7,8], including rapidly identifying the existence and function of several novel lncRNAs [9]. As a result, lncRNAs are emerging as promising therapeutic targets in numerous diseases including EMS, neurodegenerative diseases, and cancer due to newly identified roles in various key processes, such as epigenetic and transcriptional modifications and cell signaling pathways [10,11,12].
A microarray study predicted that lncRNAs participate in crucial processes associated with the pathogenesis of EMS, including immune responses, estrogen production, tissue adhesion, and angiogenesis [13]. Additional high-throughput studies have found differential expression of multiple immune-related lncRNAs in ectopic and eutopic endometrial samples, which included lncRNAs that modulate signaling pathways such as PI3-K-AKT and MAPK [14,15]. Another study investigated differential expression of lncRNAs, circRNAs, and mRNAs in eutopic and ectopic endometrial tissue using next generation sequencing (NGS) technology and identified endogenous networks among them [16]. Earlier reports showed lncRNAs to be involved in various pathological mechanisms by regulating mRNA stability and mRNA translation [17,18], Nevertheless, there have been few studies investigating lncRNA expression and the correlations between lncRNAs and mRNAs in EMS tissue.
The endometrial transition from the proliferative (P) to secretory (S) phase during the menstrual cycle entails maturation of secretory glands and adjustment of uterine fluid composition through secretion of factors that govern implantation and embryo development [19]. We hypothesized that the patterns of lncRNA and mRNA expression in endometrial tissue during the menstrual cycle would differ between normal control (CT) and EMS patients. Thus, we aimed to analyze the expression of lncRNAs and mRNAs in eutopic tissue and to identify their networks. To accomplish this, we performed high-throughput profiling of lncRNAs and mRNAs in eutopic endometrial tissues from CT and EMS patients using total RNA-Seq and functional annotation of differentially expressed mRNAs. In addition, we validated genes selected from total RNA-Seq using qRT-PCR and identified some candidate genes as EMS-related biomarkers. Our findings provide new data regarding the expression of lncRNAs and mRNAs and their correlated expression in eutopic endometrium during the menstrual cycle.
2. Results
2.1. Identification of DElncRNAs in the Control and EMS Groups during the Menstrual Cycle
We used total RNA-Seq to perform high-throughput transcriptomic profiling of eutopic endometrial tissues in the P and S phases of the menstrual cycle in the CT and EMS groups. After mining a large amount of data for lncRNAs from the CT and EMS groups using log2 transformation (fold change cutoff of 2), heatmaps were used to assess correlations between the differentially expressed lncRNAs (DElncRNAs) using ExDEGA (Ebiogen Inc., Seoul, Korea). As shown in Figures 1a and b, the expression of endometrial lncRNAs was correlated among groups. When comparing the expression patterns between the CT and EMS groups, 221 lncRNAs of the 240 DElncRNAs identified in P-phase endometrium samples were downregulated in the pEMS group compared with the pCT group, while 154 lncRNAs of the 189 DElncRNAs in S-phase samples were upregulated in the sEMS group compared with sCT group. The Venn diagrams presented in Figure 1c show up-/downregulated lncRNAs among comparison pairs: the largest number of DElncRNAs was identified between the pEMS and sEMS groups, and second largest was for comparison of the pEMS and pCT groups.
2.2. Identification of DEmRNAs between NC and EMS Groups during the Menstrual Cycle
After selecting differentially expressed mRNAs (DEmRNAs) with fold-change values ≥ 2, correlations between the DEmRNAs were analyzed using clustering heatmaps generated by ExDEGA (Excel-based differentially expressed gene analysis; Ebiogen Inc., Seoul, Korea). The expression of endometrial mRNAs was correlated by group (Figure 2a and b). When comparing the expression patterns of the NC and EMS groups, 365 mRNAs of the 407 DEmRNAs in P-phase endometrial samples were downregulated in the pEMS group compared with the pCT group, while 431 mRNAs among the 520 DEmRNAs in S-phase samples were upregulated in the sEMS group compared with the sCT group, showing very similar expression patterns. When the numbers of up/downregulated mRNAs or overlapped mRNAs among comparison pairs were presented as Venn diagrams (Figure 2c), the 4 comparison pairs showed many DEmRNAs but little overlap of mRNAs.
2.3. Functional Annotation and Pathway Networks of mRNAs
After performing GO enrichment and functional annotation analysis using DAVID bioinformatics resources, we identified three GO categories (biological process [BP], cellular component [CP], and molecular function [MF]) and Kyoto Encyclopedia of Genes and Genomics (KEGG) pathways related to DEmRNAs. As a result of enrichment of the top 20 GO terms that satisfied the adjusted p values in the BP category, genes related to cell adhesion, angiogenesis, and negative regulation of apoptotic process were differentially expressed in the pEMS group compared with the pCT group (Figure 3a and Table S1). On the other hand, genes related to cellular response to zinc ion, negative regulation of apoptotic process, and response to hypoxia were differentially expressed in the sEMS group compared with the sCT group (Figure 3b and Table S1). In addition, genes related to apoptotic processes and wound healing were differentially expressed in the pCT and sCT groups, while genes related to angiogenesis and cell adhesion were differentially expressed in the pEMS and sEMS groups (Figure S1a and b). The KEGG pathway enrichment analyses indicated that genes related to proteoglycans in cancer, osteoclast differentiation, and focal adhesion were differentially expressed in the pEMS group compared with the pCT group, while genes related to mineral absorption, focal adhesion, and pathways in cancer were differentially expressed in the sEMS group compared with the sCT group (Figure 3c, d, and Table S1). When comparing the pCT and sCT groups, genes related to mineral absorption were differentially expressed; genes related to focal adhesion were differentially expressed between pEMS and sEMS groups (Figure S1c and d).
2.4. Pathway Network Identification of DEmRNAs
To better understand the biological mechanisms and cellular pathways in endometrial tissue, DEmRNAs were analyzed using IPA software (Ingenuity System). When comparing DEmRNAs between the pCT and pEMS groups, 20 networks were identified (Table S2). In addition, 28 focus molecules were included in the top-ranked network (IPA score: 41) and were associated with cardiovascular disease, cell death and survival, and connective tissue disorders (Figure 4a and Table S2). When comparing the sCT and sEMS groups, 20 networks were identified; 29 focus molecules were included in the top-ranked network (IPA score: 42) and were associated with organismal injury and abnormalities, organismal survival, and cellular movement (Figure 4b and Table S2). When comparing the pCT and sCT groups, 13 networks were identified; 24 focus molecules were included in the top-ranked network (IPA score: 43) and were associated with neurological disease, organismal injury and abnormalities, and cardiovascular system development and function (Figure S2a and Table S2). When comparing the pEMS and sEMS groups (Figure S2b), 31 focus molecules were included in the top-ranked network (IPA score: 40).
2.5. Validation of lncRNAs and Neighboring Genes Based on qRT-PCR
To validate the expression of lncRNAs identified in the total RNA-Seq analyses, we selected 16 lncRNAs and analyzed them using qRT-PCR. In P-phase endometrial samples, 3 lncRNAs (NONHSAT008415.2, NONHSAG019742.2, and NONHSAT120701.2) were significantly upregulated, and 3 (NONHSAG048398.2, NONHSAG016560.2, and NONHSAT14106.1) were significantly downregulated in the pEMS group compared with the pCT group (Figure 5a). In addition, NONHSAG019547.2 and NONHSAG026040.2 showed a tendency to be up- or downregulated, respectively, in the pEMS group. On the other hand, in S-phase samples, 4 lncRNAs (NONHSAT000959.2, NONHSAT203423.1, NONHSAG001763.2, and NONHSAG053769.2) were significantly upregulated and 3 (NONHSAG012105.2, NONHSAG020839.2, and NONHSAG039821.2) were significantly downregulated in the sEMS group compared with the sCT group (Figure 5b). All validated lncRNAs except NONHSAG012543.2 showed similar expression patterns to those observed in the total RNA-Seq analyses (Figure 1d and e).
In light of reports that lncRNAs cis-regulate the expression of neighboring genes [20,21,22], we evaluated neighboring genes of validated lncRNAs. In P-phase, HSD11B2, a neighboring gene of NONHSAG019742.2, was significantly upregulated like the expression of NONHSAG019742.2 (Figure 6). In contrast, expression of THBS1 and SGIP1, which are neighboring genes of NONHSAG016560.2 and NONHSAG026040.2, respectively, was significantly downregulated similar to their related lncRNAs. In S-phase endometrial samples, GPX3, a neighboring gene of NONHSAT203423.1, was significantly upregulated, similar to the expression of NONHSAT203423.1. On the other hand, IGF1 and SHISA6 that are neighboring genes of NONHSAG012105.2 and NONHSAG020839.2, respectively, were significantly downregulated, similar to the related lncRNAs (Figure 6).
2.6. Validation of mRNAs Based on qRT-PCR
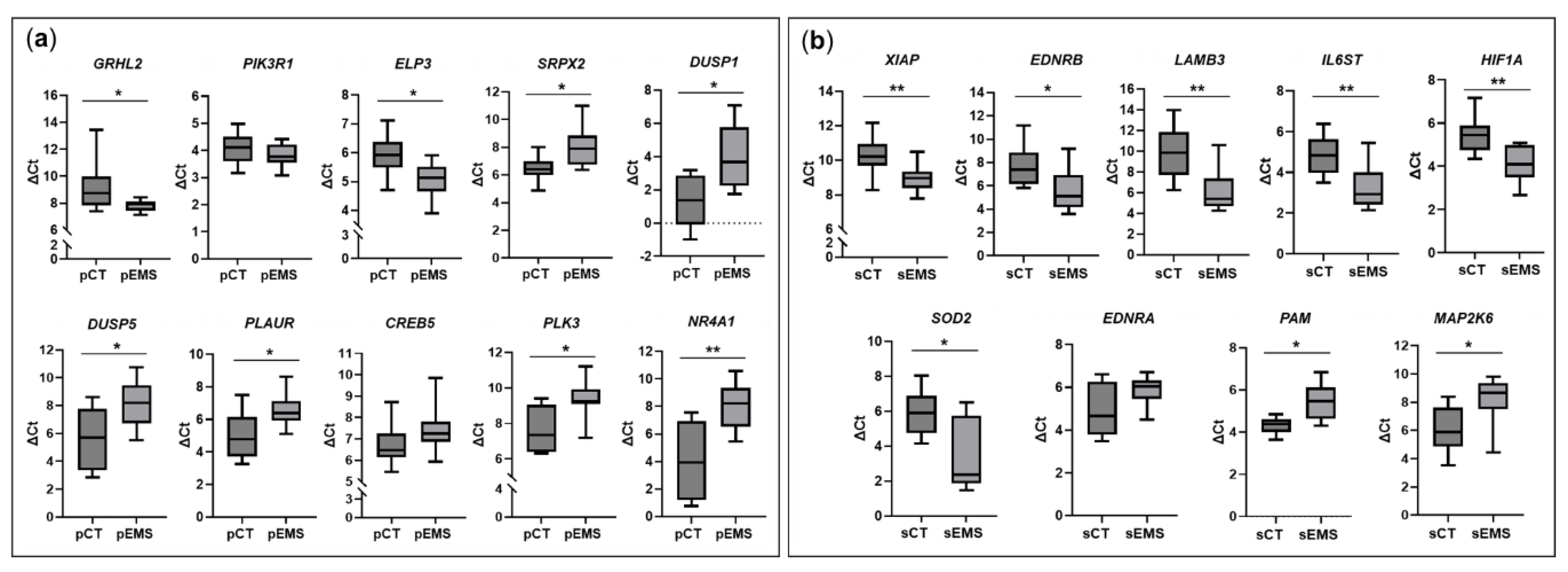
Based on the total RNA-Seq results (Table S3), we selected 10 and 9 DEmRNAs for the P and S phases, respectively, and validated them using qRT-PCR. The sample number used to validate mRNA expression changes was the same as that used to validate lncRNA expression changes. In the P-phase endometrial samples, genes related to cell adhesion (GRHL2) and positive regulation of cell migration (ELP3) were significantly upregulated in the pEMS group compared with the pCT group, while genes related to angiogenesis (SRPX2), the MAPK signaling pathway (DUSP1 and DUSP5), proteoglycans in cancer (PLAUR), negative regulation of apoptotic process (PLK3), and positive regulation of endothelial cell proliferation (NR4A1) were significantly downregulated in the pEMS group compared with the pCT group (Figure 7a). On the other hand, in the S-phase samples, genes related to negative regulation of apoptotic processes (XIAP, EDNRB, and SOD2) and pathways in cancer (LAMB3, IL6ST, and HIF1A) were significantly upregulated in the sEMS group compared with the sCT group, while genes related to responses to hypoxia (PAM) and the Rap1 signaling pathway (MAP2K6) were significantly downregulated in the sEMS group compared with the sCT group (Figure 7b). Based on these results, all validated mRNAs showed similar expression patterns to those observed in the total RNA-Seq analyses.
3. Discussion
Given that the human endometrium is a highly dynamic tissue that experiences a variety of molecular and cellular changes throughout the menstrual cycle, transcriptomic studies can provide insights into the complex mechanisms driving endometriosis. Recently, lncRNAs and mRNAs have been postulated to modulate a variety of processes involved in the pathogenesis of EMS, and lncRNAs, in particular, have emerged as possible biomarkers of EMS. Therefore, we profiled differentially expressed lncRNAs and mRNAs in both P- and S-phase endometrial samples obtained from CT and EMS patients.
In the current study, P-phase endometrial tissues from EMS patients showed changes in the expression of genes related to cell adhesion, positive regulation of angiogenesis, cellular response to tumor necrosis factor, and positive regulation of endothelial cell proliferation. Previous studies have reported that endometrium in EMS patients exhibits increased adhesion and proliferative capacity, and that the development of EMS is associated with proangiogenic processes [11,23]. However, these studies did not consider the phase of the menstrual cycle. When assessing the S phase, we found altered expression of genes involved in cellular response to zinc ion, negative regulation of growth, negative regulation of apoptotic process, angiogenesis, and response to hypoxia. Metal ions such as zinc and copper have been shown to perform various physiological functions, including enzymatic activation, DNA synthesis and repair, anti-oxidative function, hormonal and cellular signaling, and immunomodulation in the S-phase of the menstrual cycle [24,25]. Given that the fertilized egg implants in the endometrium during the S phase, assessing the expression of genes related to the regulation of these metal ions in the endometrial tissues of EMS patients is expected to be helpful in understanding infertility as well as the pathophysiology of EMS. As the endometrium transitions from the P phase to the S phase, with maturation of secretory glands and thickening of the endometrium, there are multiple changes in gene expression, and it is reasonable that some gene functions between P- and S-phases differ. These differences in gene function suggest that the menstrual cycle be considered when searching for biomarkers targeting EMS.
Even though lncRNAs do not code for proteins, they control mRNA expression both transcriptionally and post-transcriptionally. Various mechanisms are responsible for lncRNA function. A well-known example is lncRNAs functioning as competing endogenous RNA (ceRNA) to suppress miRNA and manage target mRNA [26]. Additionally, lncRNA can either cis-regulate the expression of neighboring genes or trans-regulate the expression of distant genes [27]. A growing body of evidence suggests that lncRNA plays a crucial role in endometriosis, with dysregulated lncRNAs showing potential for use as diagnostic biomarkers or targets for therapy. In this study, many lncRNAs were up- or downregulated in tissue samples from patients with EMS compared to CT patients during both the P and S phases. Among DElncRNAs in P-phase tissues from EMS patients, NONHSAG019742.2 was significantly upregulated. Recently, it has been reported that lncRNA was downregulated in the plasma of patients with dilated cardiomyopathy (DCM) [28], as were other circulating lncRNAs. These findings suggest their association with cardiac function and their potential as prognostic biomarkers for DCM. On the other hand, Cai et al. identified that NONHSAG026040.2, also known as lnc-FOSB-1-1, was downregulated in the blood of patients with systemic lupus erythematous (SLE), and that the decrease was associated with greater risk of renal damage [29]. The expression of NONHSAG026040.2 was also downregulated in the P-phase tissues of EMS patients in this study. The results in both SLE and EMS patients indicate that changes in the expression of NONHSAG026040.2 affect the development of some diseases, and that the gene may have value as a diagnostic biomarker for these diseases.
In the present study, when assessing S-phase tissues, NONHSAG053769.2 was upregulated in the EMS group compared with the CT group. In luteinizing hormone (LH) + 7 (receptive) human endometrium, lncRNA was highly expressed during the luteal phase of the ovarian cycle, coinciding with the S-phase of the menstrual cycle [30]. However, there was no difference in lncRNA expression between patients with or without recurrent implantation failure (RIF), suggesting that the lncRNA does not affect implantation failure [30]. Taken together, these findings indicate that NONHSAG053769.2, which is highly expressed during the S phase, influences the development of endometriosis but not RIF; however, such research is still in the early stages.
Based on reports that lncRNA positively cis-regulates the expression of neighboring genes [20,21], we analyzed the neighboring genes HSD11B2, THBS1, SGIP1, GPX3, IGF1, and SHISA6 of NONHSAG019742.2, NONHSAG016560.2, NONHSAG026040.2, NONHSAT203423.1, NONHSAG012105.2, and NONHSAG020839.2, respectively. These neighboring genes showed the same expression pattern as lncRNAs. In P-phase tissues, HSD11B2, located near NONHSAG019742.2, was significantly upregulated in EMS group compared to the CT group; in contrast, in S-phase tissues, the gene was significantly downregulated in EMS group in total RNA-Seq (data not shown). HSD11B2 deactivates cortisol by catalyzing its transformation into cortisone [31]. Patients with EMS exhibit abnormalities in the synthesis, degradation, and binding of steroid hormones and in the expression of genes or proteins involved in steroid hormone metabolism and regulation [32]. In the study by the Monsivais team [32], who analyzed the transcriptome in P-phase eutopic and ectopic (ovarian) endometrial tissue obtained from the same EMS patient (n = 8), HSD11B2 expression was significantly lower in the eutopic endometrium than in the ectopic endometrium, contrary to our findings. The opposing pattern of HSD11B2 expression in this study and our study may have been due to Monsivia et al. having obtained both normal and EMS tissues from the same patients, whereas we obtained normal and EMS tissues from NC and EMS patients, respectively. In the present study, SHISA6 was shown to be located near NONHSAG020839.2 and both transcripts were downregulated in S-phase tissues from EMS patients. It has been reported that SHISA6 regulates self-renewal and differentiation of spermatogonia stem cells through Wnt/beta-catenin signaling, implying that SHISA6 may be related to fertility [33,34]. These findings indicate that NONHSAG020839.2 in S-phase endometrium may also play a role in Wnt/beta-catenin signaling either directly or indirectly. Thus, we postulate the effects of as-yet unidentified lncRNAs on the development of EMS through the actions of neighboring genes.
In the present study, ELP3 involved in positive regulation of cell migration in GO enrichment was significantly up-regulated in P-phase endometrial samples from EMS patients. The ELP3 protein enhances tumorigenesis by stabilizing c-myc, and ELP3 and c-Myc are overexpressed in hepatocellular carcinoma and colorectal cancer [35]. In contrast, the ELP3 protein exhibited high expression in endometrioid adenocarcinoma, and its expression was inversely correlated with progression of the disease [36]. In a microarray transcriptomic study of endometrial tissues obtained from normal controls and IVF patients, ELP3 was up-regulated 2.7-fold in IVF patients, implying that its expression is negatively correlated with fertility [37]. Taken together, these findings indicate ELP3 to be associated with endometrial tissue function and the occurrence of endometrium-related diseases, but the mechanism remains to be elucidated.
An orphan nuclear receptor, NR4A1, is rapidly activated in response to diverse stressors, serving as an immediate-early gene. It plays a pivotal role in cancer cell functions including cell cycle progression, survival, migration, and invasion in diverse solid tumors such as melanoma, pancreatic, lung, and ovarian cancers [38,39]. In the present study, NR4A1 was significantly down-regulated in the endometrium of EMS patients during both the P phase (in both the total RNA-Seq and qRT-PCR analyses) and S phase (in the total RNA-Seq analyses). In a study assessing the phosphorylation of NR4A1 (pNR4A1) in ectopic (ovarian) endometrial tissues of EMS patients and P-phase eutopic endometrial tissues of normal controls, ovarian endometriotic tissues had higher pNR4A1 expression than normal endometrial tissues [39]. However, since the specific phase of the menstrual cycle was not identified in the tissues of EMS patients, unlike the P-phase tissues of normal controls, further evaluation is necessary to determine whether the same results are obtained when analyzing autopsy tissues from EMS patients and normal controls during the same phase of menstrual cycle. On the other hand, a study found NR4A1 expression in S-phase endometrial tissue to be lower in patients with EMS or primary infertility than in normal controls [40], consistent with the findings of our study. Thus, the expression patterns of NR4A1 appear to differ between endometriosis and endometrial or ovarian cancers, suggesting different regulatory mechanisms. In addition, it is thought that altered expression of NR4A1 in reproductive-age women with EMS may be associated with infertility.
In this study, when validating select genes for which the expression was altered in S-phase endometria tissues from EMS patients (LAMB3, IL6ST, and HIF1A belonging to pathways in cancer), the significant increase expression of all of these genes was consistent with the results of the total RNA-Seq analyses. The LAMB3 protein is a major component of the extracellular matrix and basal membrane and promotes cell migration and tumorigenicity in SCID mice and lung adenocarcinoma [41,42]. The expression of both LAMB3 mRNA and protein has been reported to be greater in ovarian cancer than in normal ovarian tissue, based on studies that examined correlations between laminin expression and prognosis in ovarian cancer using several open sources including cBioPortal and ONCOMINE [43]. Another study found LAMB3 mRNA expression to be greater in endometrial cancer than in ovarian cancer [44]. A hypoxic environment induces EMS, endometrioid endometrial cancer, and ovarian endometrial carcinoma, and these diseases induce upregulation of HIF1A in response to hypoxia [45,46,47]. Increased expression of HIF1A triggers various physiological responses, including glycolysis, to mitigate oxygen deficiency and promote angiogenesis. The serum concentration of HIF1A was reported to be greater in patients with EMS than in controls, and the expression of HIF1A increased in both serum and tissues as the severity of EMS increased [48]. Similarly, we found that HIF1A mRNA expression in endometrial tissues was greater in EMS patients than in controls, confirming that endometrial hypoxia occurs in EMS, resulting in upregulation of HIF1A. Considering the above-noted findings, our observation that the expression of genes such as LAMB3 and HIF1A increased in EMS patients suggests that early treatment of EMS may be very important to prevent progression to endometrial cancer.
In summary, we profiled DElncRNAs and DEmRNAs in EMS patients during the P and S phases of the menstrual cycle. In P-phase endometrial tissues, the expression of both NONHSAG019742.2 lncRNA and its neighboring gene, HSD11B2, increased in EMS patients, while the expression of both NONHSAG016560.2 lncRNA and its neighboring gene, THBS1, decreased in EMS patients. In S-phase endometrial tissues, the expression of both NONHSAT203423.1 lncRNA and its neighboring gene, GPX3, increased in EMS patients, while the expression of both NONHSAG020839.2 lncRNA and its neighboring gene, SHISA6, decreased in EMS patients. In addition, ELP3 and NR4A1 that positively regulate cell migration and endothelial cell proliferation, respectively, were up- or downregulated in P-phase obtained from EMS patients. In S-phase tissues, the expression of LAMB3 and HIF1A, associated with pathways in cancer, increased in EMS patients, while the expression of PAM, related to responses to hypoxia, was reduced. To the best of our knowledge, this is the first study to identify changes in the expression of both lncRNAs and mRNAs depending on the phase of the menstrual cycle in women with EMS. Given that EMS, an estrogen-dependent disease, encounters different endocrine profiles during the P- and S-phases, our results suggest the need for further exploration of the clinical applications of DElncRNAs and DEmRNAs as prognostic or diagnostic biomarkers for EMS and may to lead to valuable insights into the pathogenesis of EMS.
4. Materials and Methods
4.1. Sample Collection
The study recruited premenopausal women (aged 27-45) undergoing surgery for conditions such as EMS, benign gynecological disorders (e.g., uterine fibroids, endometrial polyps, benign ovarian tumors), at Ajou University Hospital. After explaining the study protocol and obtaining informed consent, all participants provided written consent for the study. Women taking hormonal medications that could affect endometrial conditions or having a malignant disease were excluded. Participants were divided into two groups: a control group (CT group) confirmed to have no pelvic or ovarian endometriosis at the time of surgery, and an endometriosis group (EMS group) confirmed to have pelvic EMS or endometriotic lesions on histopathological examination. Eutopic endometrial tissues were collected via Pipelle biopsy or hysteroscopy during the surgical procedure. Samples were analyzed based on the menstrual cycle phase at the time of collection, categorized into P- and S-phases.
Three samples (CT group; n=1, EMS group; n=2) were provided by the Biobank of AJOU University Hospital, a member of Korea Biobank Network. After excluding unsuitable subjects as described above, the study utilized endometrial tissues from a total of 43 participants (the CT group, n=18; the EMS group, n=25).
4.2. Library Preparation and Sequencing
Total RNA was extracted from eutopic endometrial tissues obtained from patients in the P phase (CT group [proliferative control, pCT], n=4; EMS group [proliferative EMS, pEMS], n=5) and S phase (CT group [secretory control, sCT], n=5; EMS group [secretory EMS, sEMS], n=5) of their menstrual cycle using TRIzol™ reagent (Thermo Fisher Scientific, Waltham, MA, USA). The quality of the total RNA recovered was assessed using an Agilent 4200 TapeStation system (Agilent Technologies, Santa Clara, CA, USA), and the RNA integrity number (RIN) was higher than 7.8 in all samples. After removing undesired ribosomal RNA (rRNA) using a RiboCop rRNA Depletion kit (LEXOGEN Inc., Austria), libraries were constructed using an NEBNext Ultra II Directional RNA Library Prep kit (New England BioLabs Inc., MA, USA) according to the manufacturer’s instructions. The enrichment of the libraries was carried out using PCR, and the libraries were checked using an Agilent 2100 bioanalyzer (DNA High Sensitivity Kit) to evaluate the mean fragment size. The constructed libraries were 101-bp paired-end sequenced using a NovaSeq 6000 system (Illumina, San Diego, CA, USA).
4.3. Differerential Gene Expression Analysis and Functional Annotation
Quality-control assessment of raw sequencing data was performed using FastQC [49]. Adapter contamination and low-quality reads (<Q20) were removed using FASTX_Trimmer [50] and BBMap [51]. The trimmed reads were mapped to the reference genome using TopHat [52]. The expression of genes, isoforms, and lncRNAs was estimated using Fragments Per kb per Million reads (FPKM) values by Cufflinks [53]. The FPKM values were normalized based on the quantile normalization method using edgeR. All mRNAs and ncRNAs for which the fold change in expression was ≥ 2.0 in at least one of the comparison pairs were selected. For DEmRNA and DElncRNA sets, hierarchical clustering analyses were performed using ExDEGA (Ebiogen Inc., Seoul, Korea). Gene ontology (GO) enrichment and functional annotation analysis of DEmRNAs were performed using DAVID Bioinformatics Resources (https://david.ncifcrf.gov). Additional analyses of the biological responses and various canonical pathways associated with DEmRNAs were performed using ingenuity pathway analysis (IPA) software (Ingenuity System, Redwood City, CA, USA). The IPA identified cellular networks associated with DEmRNAs based on previously known associations between genes or proteins.
4.4. qRT-PCR Analysis
We performed qRT-PCR to validate mRNAs and lncRNAs differentially expressed in total RNA-Seq assays. The number of samples per group was increased for the qRT-PCR assay as follows: pCT (n=8), pEMS (n=13), sCT (n=10), and sEMS (n=12). Total RNA was extracted from tissues using TRIzol™ reagent (Thermo Fisher Scientific, Waltham, MA, USA) and reverse transcribed into cDNA using PrimeScriptTM Reverse Transcriptase (TaKaRa, Shiga, Japan) according to the manufacturer’s instructions. Details of the qRT-PCR method have been described previously [54]. The expression of the lncRNAs and mRNAs in each sample was normalized to that of U6 and GAPDH, respectively. The relative expression differences between the NC and EMS groups were calculated using the 2-ΔΔCT method. The housekeeping gene used to analyze the expression of lncRNA was U6, and that used to analyze the expression level of mRNA was GAPDH. The expression of each gene was quantified as a threshold cycle (Ct) value, and the larger was the Ct value, the lower was the gene expression. The ΔCt value was obtained by subtracting the Ct value of the housekeeping gene from the Ct value of the target gene. Therefore, the larger was the ΔCt value in the sample, the lower was the expression of the target gene. The primers used for amplification of candidate genes are presented in Table S4.
4.5. Statistical Analyses
All statistical analyses were conducted using GraphPad Prism 10 software (San Diego, CA, USA). Differences between the CT and EMS groups with respect to the qRT-PCR results were analyzed using unpaired t-tests and Mann-Whitney U tests for normally distributed variables and non-parametric variables, respectively. All data were expressed as the mean ± standard error of the mean (SEM). A P value < 0.05 was considered statistically significant.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1,.
Author Contributions
Conceptualization, S.-R.L., M.K., M.R.C. and H.J.C.; methodology, M.R.C., J.-H.H. S.H.Y. and E.J.; validation, M.R.C. and J.-H.H.; formal analysis, M.K. and H.J.C.; data curation, M.R.C.; writing—original draft preparation, M.R.C. and H.J.C.; writing—review & editing, S.-R.L. and M.K.; supervision, S.-R.L. and M.K.; funding acquisition, S.-R.L. and M.R.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the intramural research fund of Ajou University medical center (M2021C046000083), Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2022R1I1A1A01064306), a grant of the Korea Bio-health Technology R&D Project (RS-2023-00267453), and a grant of the Korea Health Technology R&D Project (grant No. HR21C1003) through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board of Ajou University Hospital (approval number, AJOUIRB-SMP-2021-583; date of approval, 28 December 2021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The datasets generated and analyzed in this study are available from the corresponding author upon reasonable request.
Acknowledgments
Some biospecimens and data used for this study were provided by the Biobank of Ajuo University Hospital, a member of Korea Biobank Network.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Giudice, L.C.; Kao, L.C. Endometriosis. Lancet 2004, 364, 1789–1799. [Google Scholar] [CrossRef]
- Ozkan, S.; Murk, W.; Arici, A. Endometriosis and infertility: epidemiology and evidence-based treatments. Ann N Y Acad Sci 2008, 1127, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, M.; Macpherson, A.; Healy, D.L.; Rogers, P.A. Cell proliferation is increased in the endometrium of women with endometriosis. Fertil. Steril. 1995, 64, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.R.; Jin, Y.B.; Kim, H.N.; Lee, H.; Chai, Y.G.; Lee, S.R.; Kim, D.J. Differential Gene Expression in the Hippocampi of Nonhuman Primates Chronically Exposed to Methamphetamine, Cocaine, or Heroin. Psychiatry Investig 2022, 19, 538–550. [Google Scholar] [CrossRef]
- Choi, M.R.; Cho, S.; Kim, D.J.; Choi, J.S.; Jin, Y.B.; Kim, M.; Chang, H.J.; Jeon, S.H.; Yang, Y.D.; Lee, S.R. Effects of Ethanol on Expression of Coding and Noncoding RNAs in Murine Neuroblastoma Neuro2a Cells. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.R.; Jin, Y.B.; Kim, H.N.; Chai, Y.G.; Im, C.N.; Lee, S.R.; Kim, D.J. Gene expression in the striatum of cynomolgus monkeys after chronic administration of cocaine and heroin. Basic Clin Pharmacol Toxicol 2021, 128, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.R.; Jin, Y.B.; Bang, S.H.; Im, C.N.; Lee, Y.; Kim, H.N.; Chang, K.T.; Lee, S.R.; Kim, D.J. Age-related Effects of Heroin on Gene Expression in the Hippocampus and Striatum of Cynomolgus Monkeys. Clin Psychopharmacol Neurosci 2020, 18, 93–108. [Google Scholar] [CrossRef]
- Khatoon, Z.; Figler, B.; Zhang, H.; Cheng, F. Introduction to RNA-Seq and its applications to drug discovery and development. Drug Dev. Res. 2014, 75, 324–330. [Google Scholar] [CrossRef]
- Fang, Y.; Fullwood, M.J. Roles, Functions, and Mechanisms of Long Non-coding RNAs in Cancer. Genomics Proteomics Bioinformatics 2016, 14, 42–54. [Google Scholar] [CrossRef]
- Hudson, Q.J.; Proestling, K.; Perricos, A.; Kuessel, L.; Husslein, H.; Wenzl, R.; Yotova, I. The role of long non-coding RNAs in endometriosis. Int. J. Mol. Sci. 2021, 22, 11425. [Google Scholar] [CrossRef]
- Zhang, M.; He, P.; Bian, Z. Long Noncoding RNAs in Neurodegenerative Diseases: Pathogenesis and Potential Implications as Clinical Biomarkers. Front Mol Neurosci 2021, 14, 685143. [Google Scholar] [CrossRef]
- Sun, P.R.; Jia, S.Z.; Lin, H.; Leng, J.H.; Lang, J.H. Genome-wide profiling of long noncoding ribonucleic acid expression patterns in ovarian endometriosis by microarray. Fertil. Steril. 2014, 101, 1038–1046. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Wang, D.; Cui, L.; Yang, Q. RNA sequencing-based long non-coding RNA analysis and immunoassay in ovarian endometriosis. Am J Reprod Immunol 2021, 85, e13359. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Luo, Y.; Wang, G.; Yang, Q. Circular RNA expression profiles and bioinformatics analysis in ovarian endometriosis. Mol Genet Genomic Med 2019, 7, e00756. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Zhai, L.; Wang, J.; Yu, Q.; Li, T.; Xu, X.; Guo, X.; Mao, X.; Zhou, J.; Zhang, X. Comprehensive Analysis of RNA-Seq in Endometriosis Reveals Competing Endogenous RNA Network Composed of circRNA, lncRNA and mRNA. Front Genet 2022, 13, 828238. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Song, X. The Working Modules of Long Noncoding RNAs in Cancer Cells. Adv Exp Med Biol 2016, 927, 49–67. [Google Scholar] [CrossRef]
- Yang, G.; Lu, X.; Yuan, L. LncRNA: a link between RNA and cancer. Biochim. Biophys. Acta 2014, 1839, 1097–1109. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Q.; Wang, H.; Duan, E. Uterine Fluid in Pregnancy: A Biological and Clinical Outlook. Trends Mol Med 2017, 23, 604–614. [Google Scholar] [CrossRef]
- Anderson, K.M.; Anderson, D.M.; McAnally, J.R.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Transcription of the non-coding RNA upperhand controls Hand2 expression and heart development. Nature 2016, 539, 433–436. [Google Scholar] [CrossRef]
- Joung, J.; Engreitz, J.M.; Konermann, S.; Abudayyeh, O.O.; Verdine, V.K.; Aguet, F.; Gootenberg, J.S.; Sanjana, N.E.; Wright, J.B.; Fulco, C.P.; et al. Genome-scale activation screen identifies a lncRNA locus regulating a gene neighbourhood. Nature 2017, 548, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Lu, J.Y.; Liu, L.; Yin, Y.; Chen, C.; Han, X.; Wu, B.; Xu, R.; Liu, W.; Yan, P.; et al. Divergent lncRNAs regulate gene expression and lineage differentiation in pluripotent cells. Cell Stem Cell 2016, 18, 637–652. [Google Scholar] [CrossRef]
- Wang, Y.; Nicholes, K.; Shih, I.M. The Origin and Pathogenesis of Endometriosis. Annu Rev Pathol 2020, 15, 71–95. [Google Scholar] [CrossRef] [PubMed]
- Nasiadek, M.; Stragierowicz, J.; Klimczak, M.; Kilanowicz, A. The role of zinc in selected female reproductive system disorders. Nutrients 2020, 12, 2464. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Yue, X.; Zhang, Y.; Shen, L.; Zhang, H.; Wang, X.; Yin, T.; Zhang, H.; Peng, J.; Wang, X.; et al. Elevated levels of Zn, Cu and Co are associated with an increased risk of endometriosis: Results from a casecontrol study. Ecotoxicol Environ Saf 2024, 271, 115932. [Google Scholar] [CrossRef] [PubMed]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Villegas, V.E.; Zaphiropoulos, P.G. Neighboring gene regulation by antisense long non-coding RNAs. Int. J. Mol. Sci. 2015, 16, 3251–3266. [Google Scholar] [CrossRef]
- Zhang, X.; Nie, X.; Yuan, S.; Li, H.; Fan, J.; Li, C.; Sun, Y.; Zhao, Y.; Hou, H.; Wang, D.W.; et al. Circulating Long Non-coding RNA ENST00000507296 Is a Prognostic Indicator in Patients with Dilated Cardiomyopathy. Mol Ther Nucleic Acids 2019, 16, 82–90. [Google Scholar] [CrossRef]
- Cai, B.; Cai, J.; Yin, Z.; Jiang, X.; Yao, C.; Ma, J.; Xue, Z.; Miao, P.; Xiao, Q.; Cheng, Y.; et al. Long non-coding RNA expression profiles in neutrophils revealed potential biomarker for prediction of renal involvement in SLE patients. Rheumatology (Oxford) 2021, 60, 1734–1746. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, S.; Yao, G.; Zhu, Q.; He, Y.; Lu, Y.; Qi, J.; Xu, R.; Ding, Y.; Li, J.; et al. A Novel Molecule in Human Cyclic Endometrium: LncRNA TUNAR Is Involved in Embryo Implantation. Front Physiol 2020, 11, 587448. [Google Scholar] [CrossRef]
- Krozowski, Z.; MaGuire, J.A.; Stein-Oakley, A.N.; Dowling, J.; Smith, R.E.; Andrews, R.K. Immunohistochemical localization of the 11 beta-hydroxysteroid dehydrogenase type II enzyme in human kidney and placenta. J. Clin. Endocrinol. Metab. 1995, 80, 2203–2209. [Google Scholar] [CrossRef] [PubMed]
- Monsivais, D.; Bray, J.D.; Su, E.; Pavone, M.E.; Dyson, M.T.; Navarro, A.; Kakinuma, T.; Bulun, S.E. Activated glucocorticoid and eicosanoid pathways in endometriosis. Fertil. Steril. 2012, 98, 117–125. [Google Scholar] [CrossRef]
- Tokue, M.; Ikami, K.; Mizuno, S.; Takagi, C.; Miyagi, A.; Takada, R.; Noda, C.; Kitadate, Y.; Hara, K.; Mizuguchi, H.; et al. SHISA6 Confers Resistance to Differentiation-Promoting Wnt/β-Catenin Signaling in Mouse Spermatogenic Stem Cells. Stem Cell Reports 2017, 8, 561–575. [Google Scholar] [CrossRef]
- Woodward, E.; Schlingmann, K.; Tobias, J.; Turner, R. Characterisation of the testicular transcriptome in stallions with age-related testicular degeneration. Equine Vet J 2023, 55, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Ouyang, C.; Huang, C.; Zhang, J.; Xiao, Q.; Zhang, F.; Wang, H.; Lin, F.; Wang, J.; Wang, Z.; et al. ELP3 stabilizes c-Myc to promote tumorigenesis. J. Mol. Cell. Biol. 2024, 15, mjad059. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ikeda, J.I.; Rahadiani, N.; Mamat, S.; Ueda, Y.; Tian, T.; Enomoto, T.; Kimura, T.; Aozasa, K.; Morii, E. Prognostic significance of elongator protein 3 expression in endometrioid adenocarcinoma. Oncol Lett 2012, 3, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kalma, Y.; Granot, I.; Gnainsky, Y.; Or, Y.; Czernobilsky, B.; Dekel, N.; Barash, A. Endometrial biopsy-induced gene modulation: first evidence for the expression of bladder-transmembranal uroplakin Ib in human endometrium. Fertil. Steril. 2009, 91, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Karki, K. The Paradoxical Roles of Orphan Nuclear Receptor 4A (NR4A) in Cancer. Mol Cancer Res 2021, 19, 180–191. [Google Scholar] [CrossRef]
- Zeng, X.; Yue, Z.; Gao, Y.; Jiang, G.; Zeng, F.; Shao, Y.; Huang, J.; Yin, M.; Li, Y. NR4A1 is Involved in Fibrogenesis in Ovarian Endometriosis. Cell Physiol Biochem 2018, 46, 1078–1090. [Google Scholar] [CrossRef]
- Tang, X.; Zheng, H.; Xu, H.; Wang, M.; Kang, X.; Wu, Z. NR4A1 Affects endometrial receptivity by participating in mesenchymal–epithelial transition of endometrial stromal cells. Reprod. Sci. 2022, 29, 133–142. [Google Scholar] [CrossRef]
- Ii, M.; Yamamoto, H.; Taniguchi, H.; Adachi, Y.; Nakazawa, M.; Ohashi, H.; Tanuma, T.; Sukawa, Y.; Suzuki, H.; Sasaki, S.; et al. Co-expression of laminin β3 and γ2 chains and epigenetic inactivation of laminin α3 chain in gastric cancer. Int. J. Oncol. 2011, 39, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Li, J.; Yan, M.X.; Liu, L.; Jia, D.S.; Geng, Q.; Lin, H.C.; He, X.H.; Li, J.J.; Yao, M. Integrative analyses identify osteopontin, LAMB3 and ITGB1 as critical pro-metastatic genes for lung cancer. PLoS One 2013, 8, e55714. [Google Scholar] [CrossRef]
- Diao, B.; Yang, P. Comprehensive Analysis of the Expression and Prognosis for Laminin Genes in Ovarian Cancer. Pathol Oncol Res 2021, 27, 1609855. [Google Scholar] [CrossRef] [PubMed]
- Guirguis, A.; Elishaev, E.; Oh, S.H.; Tseng, G.C.; Zorn, K.; DeLoia, J.A. Use of gene expression profiles to stage concurrent endometrioid tumors of the endometrium and ovary. Gynecol. Oncol. 2008, 108, 370–376. [Google Scholar] [CrossRef]
- Kumari, P.; Sharma, I.; Saha, S.C.; Srinivasan, R.; Sharma, A. Promoter methylation status of key genes and its implications in the pathogenesis of endometriosis, endometrioid carcinoma of ovary and endometrioid endometrial cancer. J Cancer Res Ther 2022, 18, S328–s334. [Google Scholar] [CrossRef]
- Sivridis, E.; Giatromanolaki, A.; Gatter, K.C.; Harris, A.L.; Koukourakis, M.I.; Tumor and Angiogenesis Research Group. Association of hypoxia-inducible factors 1α and 2α with activated angiogenic pathways and prognosis in patients with endometrial carcinoma. Cancer 2002, 95, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.M.; Yang, W.X. Epithelial-to-mesenchymal transition in the development of endometriosis. Oncotarget 2017, 8, 41679–41689. [Google Scholar] [CrossRef]
- Zhang, F.; Liu, X.L.; Wang, W.; Dong, H.L.; Xia, Y.F.; Ruan, L.P.; Liu, L.P. Expression of MMIF, HIF-1α and VEGF in Serum and Endometrial Tissues of Patients with Endometriosis. Curr Med Sci 2018, 38, 499–504. [Google Scholar] [CrossRef]
- Simon, A. Babraham Bioinformatics. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 24 May 2023).
- Hannon Lab. FASTX-Toolkit. Available online: http://hannonlab.cshl.edu/fastx_toolkit/index.html (accessed on 24 April 2023).
- Bushnell, B. BBMap. Available online: https://sourceforge.net/projects/bbmap (accessed on 3 May 2023).
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol 2011, 12, R22. [Google Scholar] [CrossRef]
- Choi, M.R.; Jung, K.H.; Park, J.H.; Das, N.D.; Chung, M.K.; Choi, I.G.; Lee, B.C.; Park, K.S.; Chai, Y.G. Ethanol-induced small heat shock protein genes in the differentiation of mouse embryonic neural stem cells. Arch Toxicol 2011, 85, 293–304. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Differential expression of long noncoding RNA (lncRNA) in the control (CT) and endometriosis (EMS) groups. (a) Heatmap generated from hierarchical clustering analysis of differentially expressed lncRNAs (DElncRNAs) in the proliferative-phase CT (pCT) and EMS (pEMS) groups. (b) Heatmap generated from hierarchical clustering analysis of DElncRNAs in the secretory-phase CT (sCT) and EMS (sEMS) groups. (c) Venn diagram showing overlap of lncRNA expression among the pCT, pEMS, sCT, and sEMS groups. (d, e) Expression of select lncRNAs in the CT and EMS groups as assessed using qRT-PCR.
Figure 1.
Differential expression of long noncoding RNA (lncRNA) in the control (CT) and endometriosis (EMS) groups. (a) Heatmap generated from hierarchical clustering analysis of differentially expressed lncRNAs (DElncRNAs) in the proliferative-phase CT (pCT) and EMS (pEMS) groups. (b) Heatmap generated from hierarchical clustering analysis of DElncRNAs in the secretory-phase CT (sCT) and EMS (sEMS) groups. (c) Venn diagram showing overlap of lncRNA expression among the pCT, pEMS, sCT, and sEMS groups. (d, e) Expression of select lncRNAs in the CT and EMS groups as assessed using qRT-PCR.
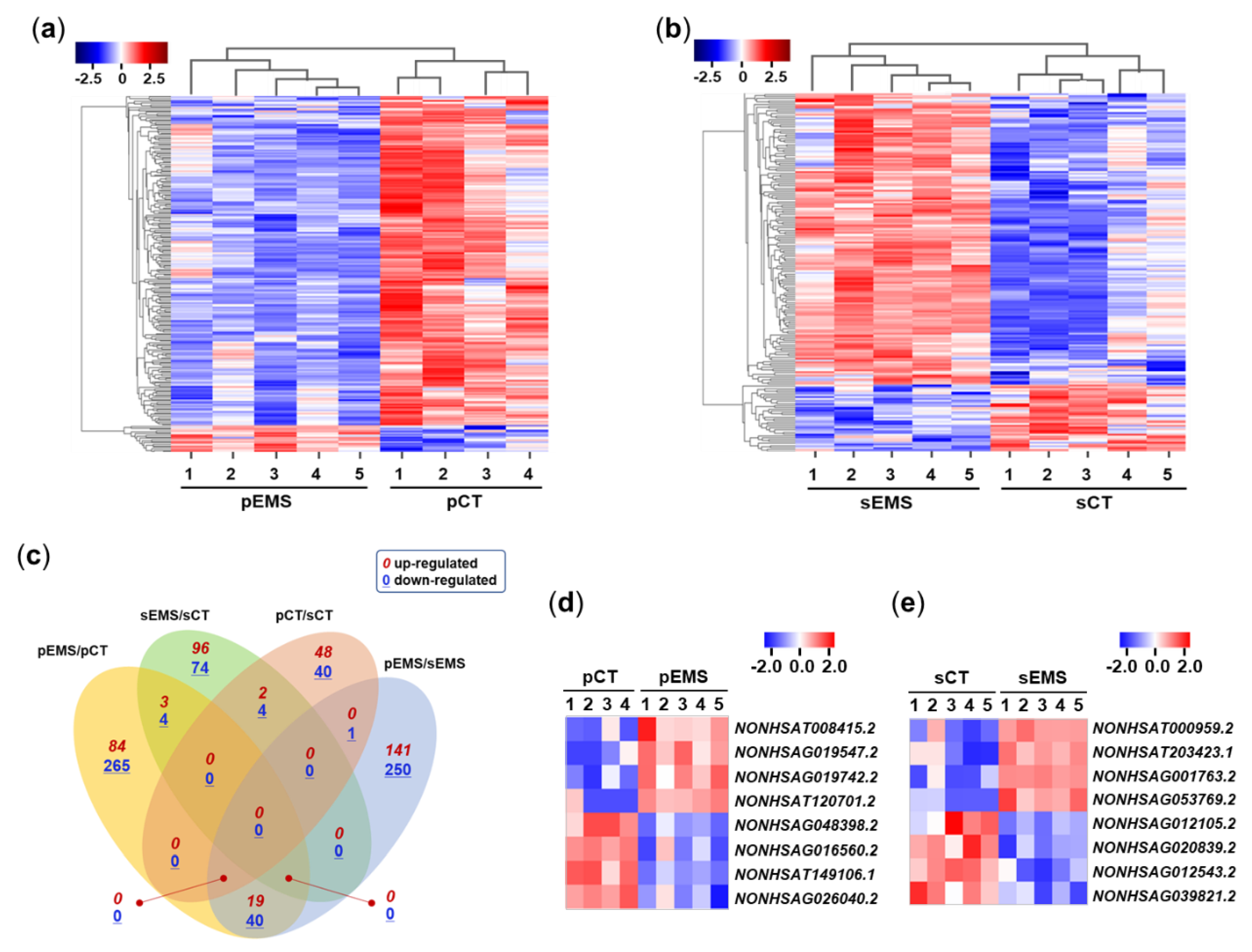
Figure 2.
Differential expression (DE) of mRNAs in the control (CT) and endometriosis (EMS) groups. (a) Heatmap generated from hierarchical clustering analysis of differentially expressed mRNAs (DEmRNAs) in the proliferative-phase CT (pCT) and EMS (pEMS) groups. (b) Heatmap generated from hierarchical clustering analysis of DEmRNAs in the secretory-phase CT (sCT) and EMS (sEMS) groups. (c) Venn diagram showing overlap of mRNAs among the pCT, pEMS, sCT, and sEMS groups. (d, e) Expression of select DEmRNAs in the CT and EMS groups as assessed using qRT-PCR.
Figure 2.
Differential expression (DE) of mRNAs in the control (CT) and endometriosis (EMS) groups. (a) Heatmap generated from hierarchical clustering analysis of differentially expressed mRNAs (DEmRNAs) in the proliferative-phase CT (pCT) and EMS (pEMS) groups. (b) Heatmap generated from hierarchical clustering analysis of DEmRNAs in the secretory-phase CT (sCT) and EMS (sEMS) groups. (c) Venn diagram showing overlap of mRNAs among the pCT, pEMS, sCT, and sEMS groups. (d, e) Expression of select DEmRNAs in the CT and EMS groups as assessed using qRT-PCR.
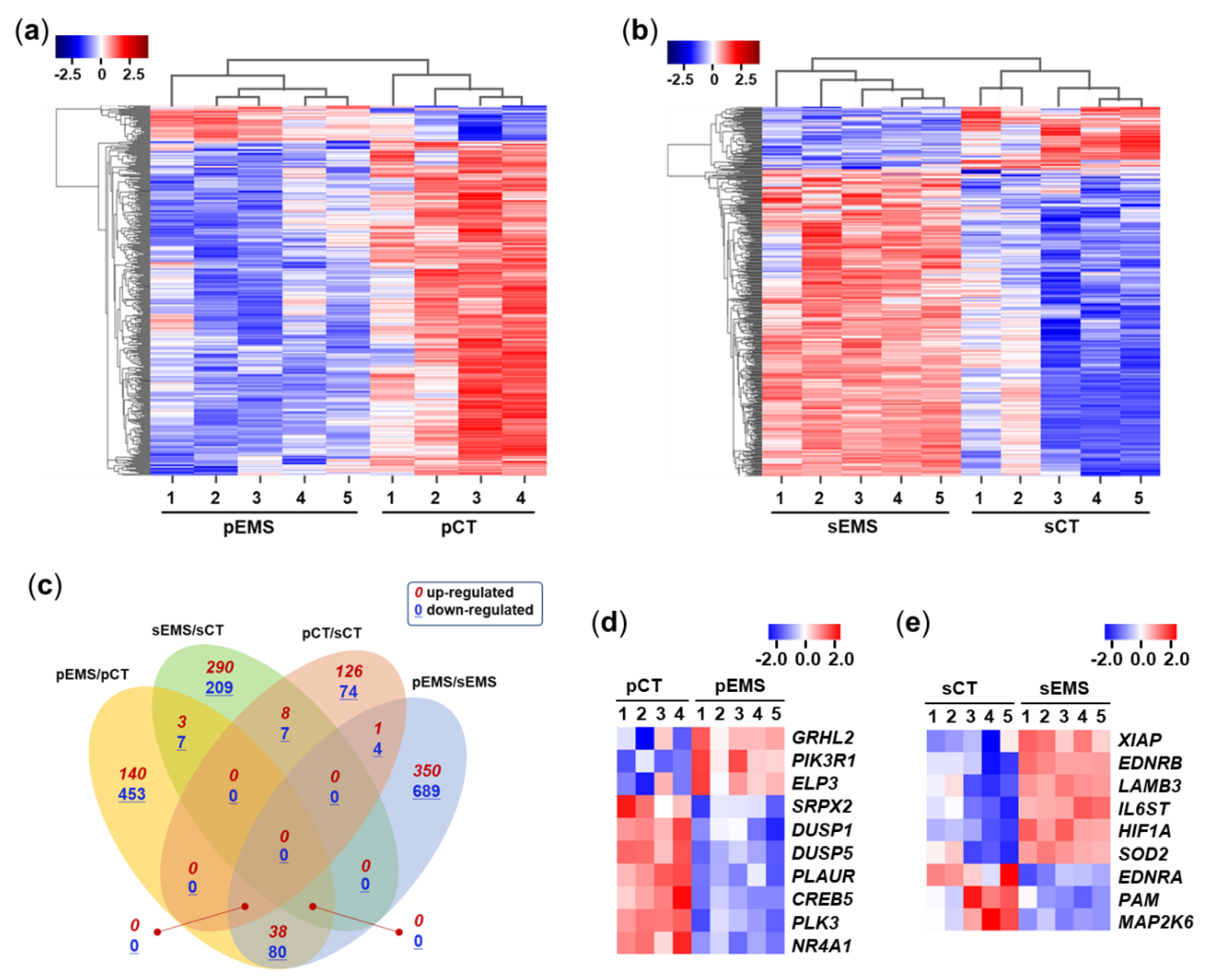
Figure 3.
Classification of differentially expressed mRNAs (DEmRNAs) between the control (CT) and endometriosis (EMS) groups as assessed using gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomics (KEGG) PATHWAY enrichment. (a) Top 20 enriched GO terms in the biological process (BP) category among DEmRNAs in the pEMS group compared with the pCT group. (b) Top 20 enriched GO terms in the BP category among DEmRNAs in the sEMS group compared to the sCT group. (c) Top 10 enriched KEGG pathways in the pEMS group compared with the pCT group. (d) Top 10 enriched KEGG pathways in the sEMS group compared with the sCT group. The X-axis represents the number of genes. **p < 0.001, ***p < 0.0001.
Figure 3.
Classification of differentially expressed mRNAs (DEmRNAs) between the control (CT) and endometriosis (EMS) groups as assessed using gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomics (KEGG) PATHWAY enrichment. (a) Top 20 enriched GO terms in the biological process (BP) category among DEmRNAs in the pEMS group compared with the pCT group. (b) Top 20 enriched GO terms in the BP category among DEmRNAs in the sEMS group compared to the sCT group. (c) Top 10 enriched KEGG pathways in the pEMS group compared with the pCT group. (d) Top 10 enriched KEGG pathways in the sEMS group compared with the sCT group. The X-axis represents the number of genes. **p < 0.001, ***p < 0.0001.
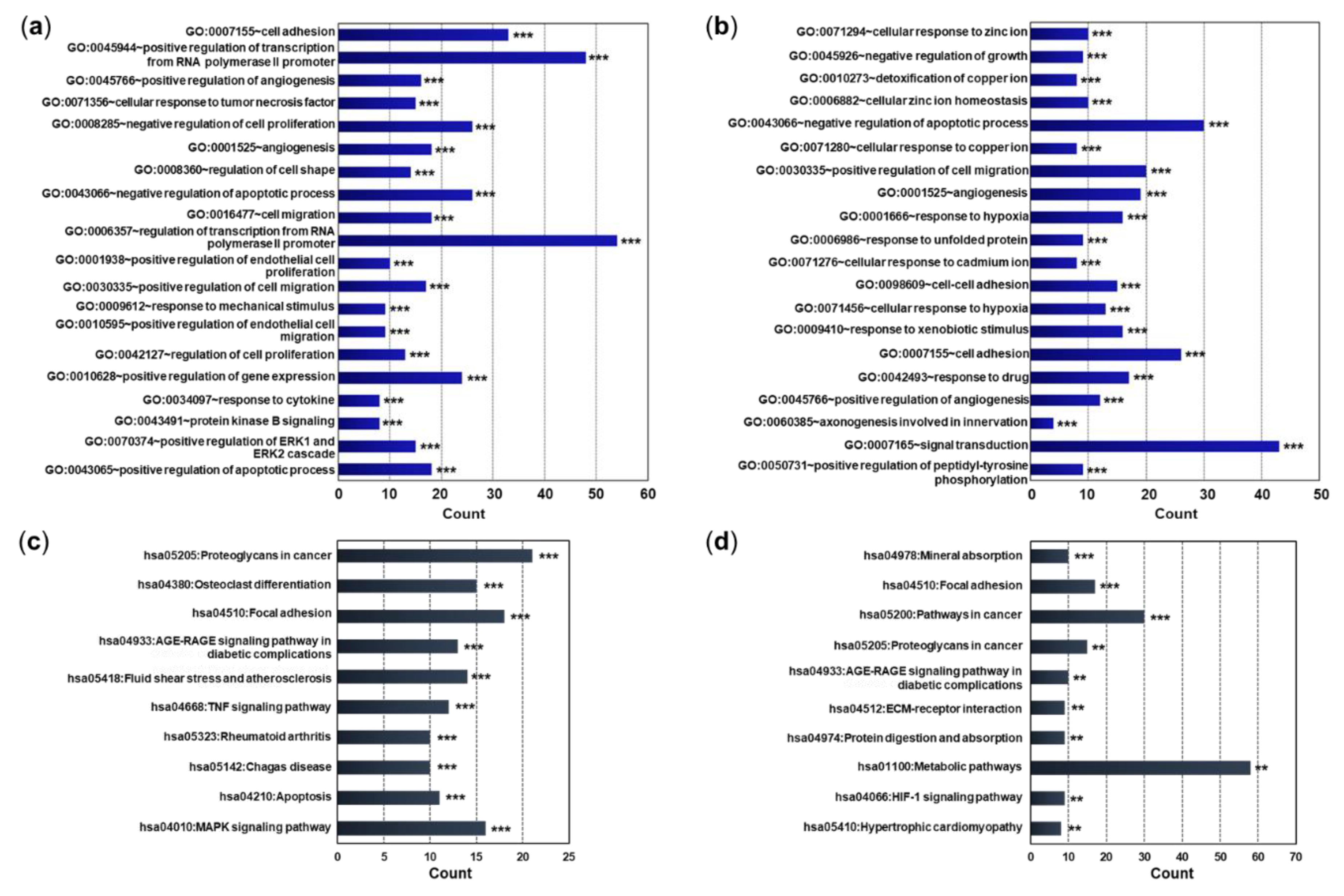
Figure 4.
Top networks of differentially expressed mRNAs (DEmRNAs) in the control (CT) and endometriosis (EMS) groups, as identified using the Ingenuity Pathway Analysis (IPA) software (QIAGEN, Germany). (a) The top network of DEmRNAs in the pEMS group compared with the pCT group. (b) The top network of DEmRNAs in the sEMS group compared with the sCT group.
Figure 4.
Top networks of differentially expressed mRNAs (DEmRNAs) in the control (CT) and endometriosis (EMS) groups, as identified using the Ingenuity Pathway Analysis (IPA) software (QIAGEN, Germany). (a) The top network of DEmRNAs in the pEMS group compared with the pCT group. (b) The top network of DEmRNAs in the sEMS group compared with the sCT group.
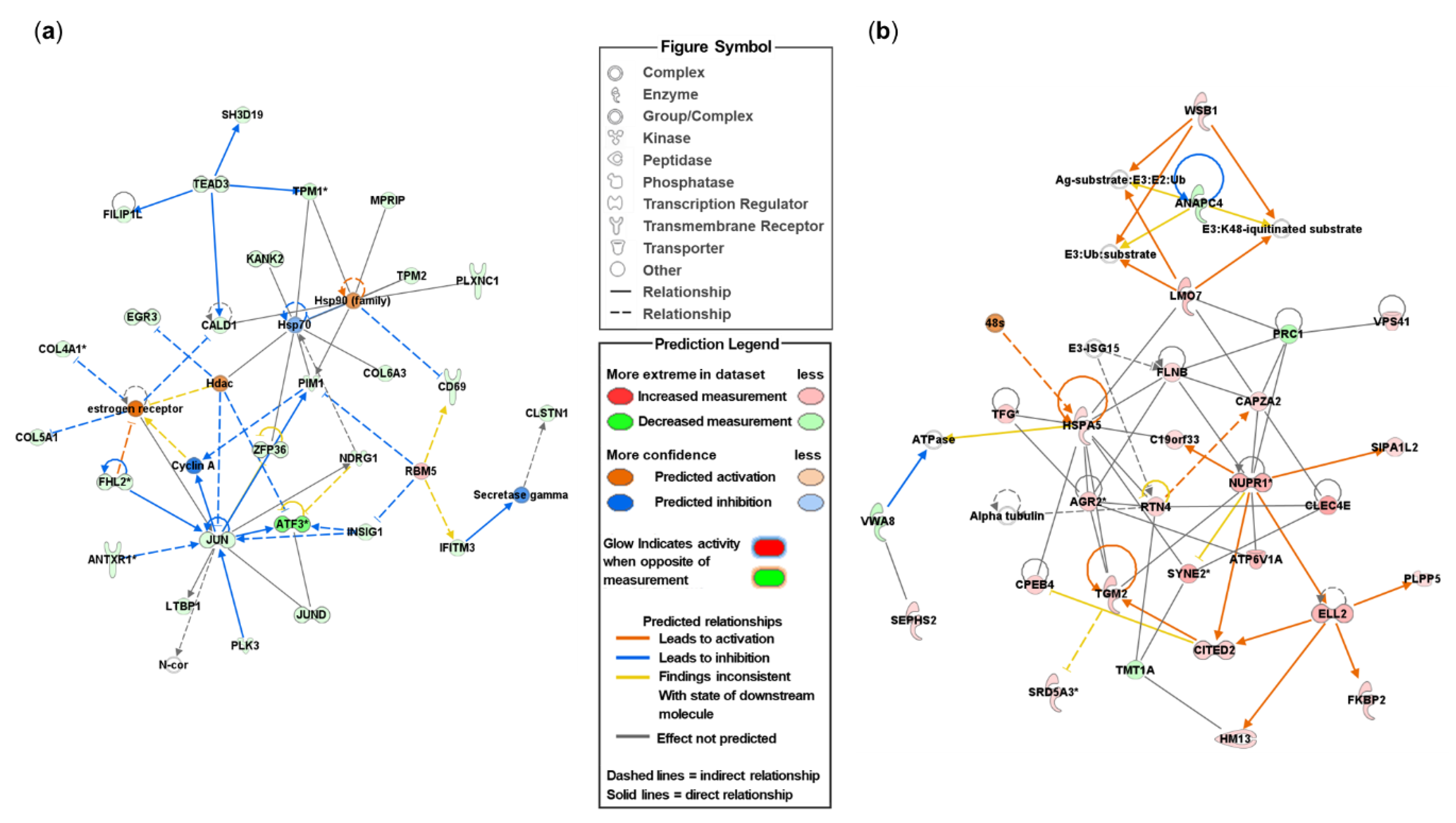
Figure 5.
Validation of differentially expressed long noncoding RNAs (DElncRNAs) during the menstrual cycle of the control (CT) and endometriosis (EMS) groups. Select DElncRNAs were validated using qRT-PCR. (a) DElncRNAs in the EMS group (n = 13) compared with the CT group (n = 8) during the proliferative phase of the menstrual cycle. (b) DElncRNAs in the EMS group (n = 12) compared with the CT group (n = 13) during the secretory phase of the menstrual cycle. *p < 0.05.
Figure 5.
Validation of differentially expressed long noncoding RNAs (DElncRNAs) during the menstrual cycle of the control (CT) and endometriosis (EMS) groups. Select DElncRNAs were validated using qRT-PCR. (a) DElncRNAs in the EMS group (n = 13) compared with the CT group (n = 8) during the proliferative phase of the menstrual cycle. (b) DElncRNAs in the EMS group (n = 12) compared with the CT group (n = 13) during the secretory phase of the menstrual cycle. *p < 0.05.
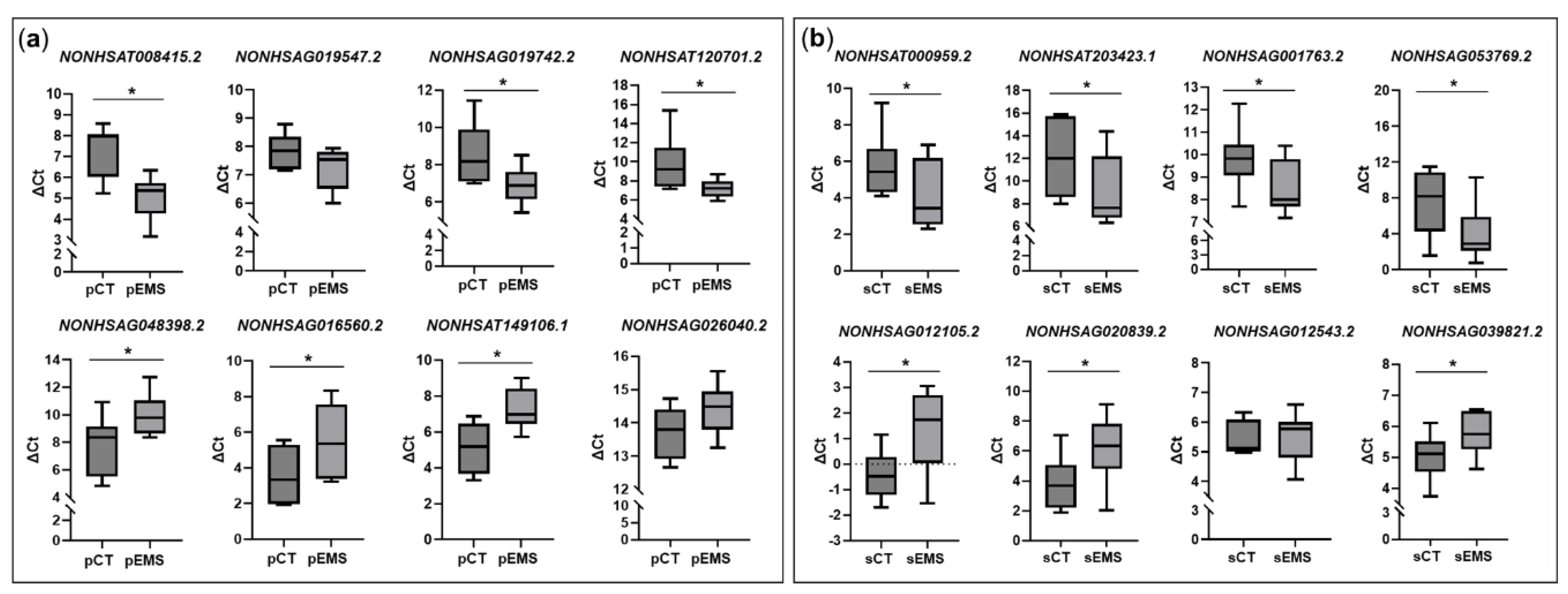
Figure 6.
Validation of the differential expression of genes near select validated lncRNAs in the control (CT) and endometriosis (EMS) groups during the menstrual cycle. (a) Changes in the expression of neighboring genes in the EMS group (n = 13) compared with the CT group (n = 8) during the proliferative phase of the menstrual cycle. (b) Changes in the expression of neighboring genes in the EMS group (n = 12) compared with the CT group (n = 13) during the secretory phase of the menstrual cycle. *p < 0.05.
Figure 6.
Validation of the differential expression of genes near select validated lncRNAs in the control (CT) and endometriosis (EMS) groups during the menstrual cycle. (a) Changes in the expression of neighboring genes in the EMS group (n = 13) compared with the CT group (n = 8) during the proliferative phase of the menstrual cycle. (b) Changes in the expression of neighboring genes in the EMS group (n = 12) compared with the CT group (n = 13) during the secretory phase of the menstrual cycle. *p < 0.05.
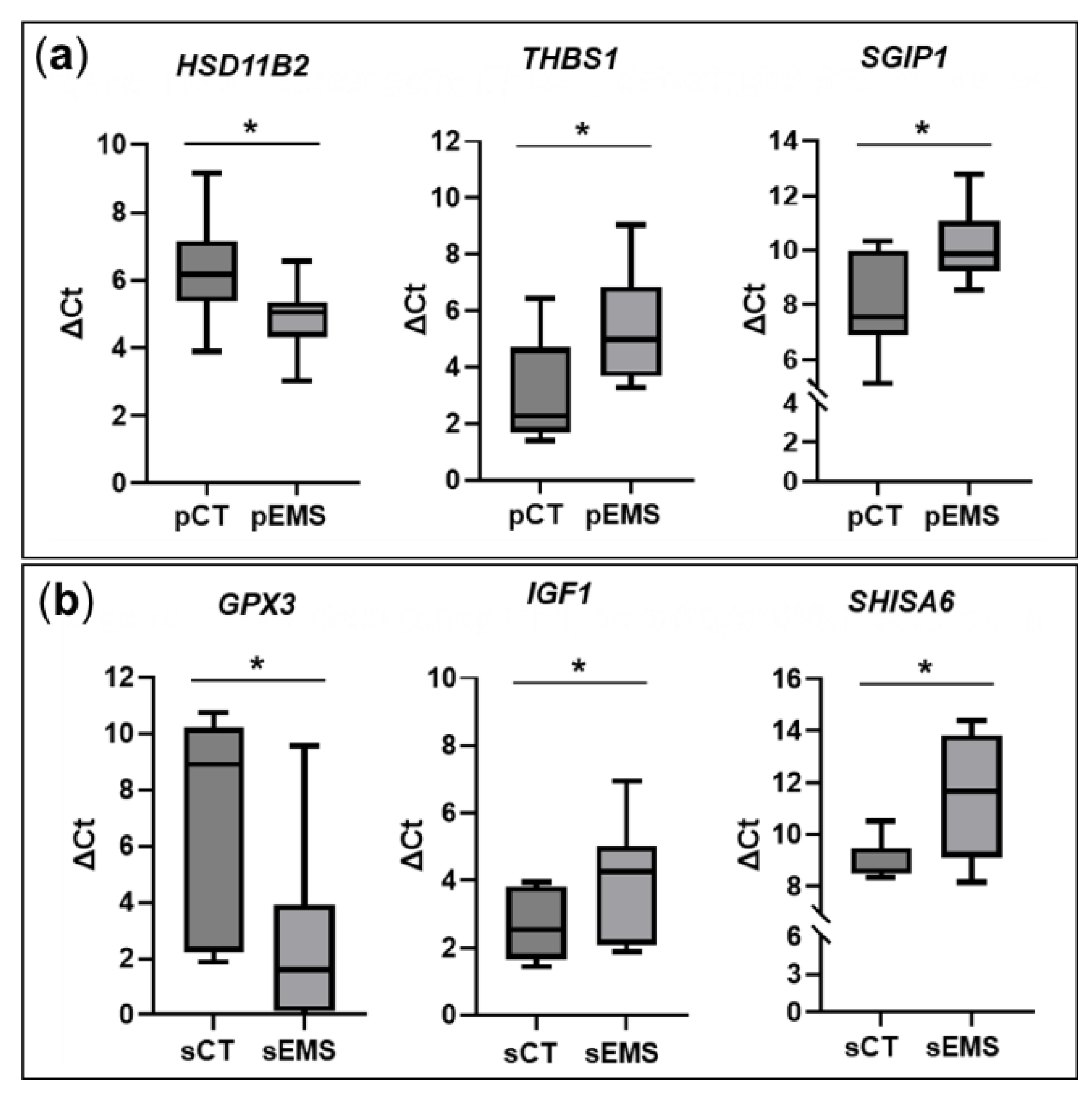
Figure 7.
Validation of differentially expressed mRNAs (DEmRNAs) in the control (CT) and endometriosis (EMS) groups during the menstrual cycle. (a) Changes in the expression of select mRNAs in the EMS group (n = 13) compared with the CT group (n = 8) during the proliferative phase of the menstrual cycle. (b) Changes in the expression of some mRNAs in the EMS group (n = 12) compared with the CT group (n = 13) during the secretory phase of the menstrual cycle. *p < 0.05, **p < 0.01.
Figure 7.
Validation of differentially expressed mRNAs (DEmRNAs) in the control (CT) and endometriosis (EMS) groups during the menstrual cycle. (a) Changes in the expression of select mRNAs in the EMS group (n = 13) compared with the CT group (n = 8) during the proliferative phase of the menstrual cycle. (b) Changes in the expression of some mRNAs in the EMS group (n = 12) compared with the CT group (n = 13) during the secretory phase of the menstrual cycle. *p < 0.05, **p < 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



