
Submitted:
22 August 2024
Posted:
23 August 2024
You are already at the latest version
Abstract
Based on molecular phylogenetic analyses, but also considering morphological characters, four new families are separated from the family Glomeraceae within the Glomerales and the class Glomeromycetes. The revised family Glomeraceae comprises only four genera: the type genus Glomus, Complexispora, Sclerocarpum and Simiglomus. Septoglomeraceae fam. nov. comprises, besides Septoglomus, also Funneliformis, Funneliglomus, Blaszkowskia and Viscospora. Sclerocystaceae fam. nov. is represented by the type genus Sclerocystis, but also by Halonatospora, Oehlia, Parvocarpum, Rhizoglomus and Silvaspora. Kamienskiaceae fam. nov. encompasses Kamienskia, Microkamienskia and Epigeocarpum. Finally, Dominikiaceae fam. nov. includes the genera Dominikia, Macrodominikia gen. nov. Microdominikia, Nanoglomus and Orientoglomus. The genera Oehlia and Halonatospora form two other clades well separated from Silvaspora, Sclerocystis and Rhizoglomus and might represent two further families within the Glomerales. Such deeper-going separation is to our opinion fully supported by molecular phylogeny, but in view of the low numbers of taxa, this separation is not yet needed at this stage of research progress.
Keywords:
classification
; systematics
; taxonomy
; arbuscular mycorrhizal fungi
1. Introduction
The fungal phylum Glomeromycota (arbuscular mycorrhizal fungi, AMF) currently comprises three classes, six orders, sixteen families and > 50 genera [1,2,3,4,5,6]. In the class Glomeromycetes, four orders were described, which are called Diversisporales, Entrophosporales, Gigasporales and Glomerales [4,7,8,9,10]. Within the Glomerales and Entrophosporales there has been just one family for each other, the Glomeraceae and the Entrophosporaceae [9,11], while Diversisporales comprises the type family (Diversisporaceae) and three other families: Acaulosporaceae, Pacisporaceae and Sacculosporaceae [9,12,13,14]. The order Gigasporales comprises the five families Gigasporaceae, Scutellosporaceae, Racocetraceae, Dentiscutataceae and Intraornatosporaceae [2,3,15,16]. Finally, in the most ancient clades of AM fungi there are Archaeosporales, including the families Archaeosporaceae, Ambisporaceae, Geosiphonaceae and Polonosporaceae, and Paraglomerales, comprising Paraglomeraceae and Pervetustaceae [2,3].
Within the last few years, new genera were separated within the Glomerales, due to concomitant morphological and molecular analyses and especially phylogenetic progresses [5,6,17,18,19,20,21]. The Glomeraceae is the family with highest species richness in Glomeromycota (> 150), and the latest phylogenetic trees for Glomeraceae species revealed at least four or five major clades [4,6,17,19,20,21]. Glomus macrocarpum, Septoglomus constrictum, Sclerocystis coremioides, Kamienskia bistrata and Dominikia minuta might be the most representative species of these five major clades. The family Glomeraceae also includes two genera of outstanding importance, which are counted among the 50 most cited fungal genera within the last decades, Funneliformis and Rhizoglomus due to the species F. mosseae and R. irregulare, and often used in basic research, applied sciences and also commercially applied, e.g. to enhance soil fertility and improve plant nutrition and root health [22]. Two complexes of genera were already reported in previous works, the Dominikia and the Septoglomus complexes [5,19). Based on our observations (phylogenetic and morphological analyses), our hypothesis is that at least five major clades/complexes exist in Glomeraceae, and thus the family could be divided into five different families, based on these five major clades/complexes. Thus, the objectives of this study were to perform a thorough phylogenetic analysis on the described Glomerales species and to re-organize all the families and genera justified within the order. At the end, a revision of the family Glomeraceae will be needed, and new families would have to be described, based on significative type species.
2. Materials and Methods
2.1. Phylogenetic Analyses
To reconstruct the phylogeny, two alignments (datasets) were generated with AM fungal sequences from the genera of Glomerales (Supplementary Material, Spreadsheet S1). The first dataset was used to reconstruct the phylogeny based on partial SSU, ITS region, and partial LSU nrDNA. The second was used to reconstruct the phylogeny based on partial SSU, 5.8S, and partial LSU nrDNA. Entrophospora etunicata (W.N. Becker & Gerd.) Błaszk., Niezgoda, B.T. Goto & Magurno was included as outgroup in all analyses. For some isolates just sequences from the partial LSU nrDNA were used, which are indicated in the phylogenetic trees generated. Sequences from Septoglomus titan were not used in the phylogenetic analyses because we decided do not include sequences in the alignment with less than 500bp (sequences from S. titan have 457bp). According to Silva et al. [5] it is not possible to be sure about the phylogenetic position of this species due to the short LSU nrDNA fragment and unavailability of its ITS sequences. The datasets were aligned in Mafft [23], using the default parameters. Prior to phylogenetic analyses, the model of nucleotide substitution was estimated using Topali 2.5 [24]. Bayesian (two runs over 5 × 106 generations, with a sample frequency of 500 and a burnin value of 25%) and maximum likelihood (1,000 bootstrap) analyses were performed, respectively, in MrBayes 3.1.2 [25] and PhyML [26], launched from Topali 2.5, using the GTR + G model.
2.2. Specimen Analyses
We analyzed morphologically all species belonging to the Glomerales, and in detail specimens representing > 120 taxa. Holotype, isotype, paratype and ex-type materials were examined along with representatives from institutional herbaria — OSC, FH, Z+ZT, URM (Recife, Brazil), Embrapa Agrobiologia (Seropédica, Brazil), DCS-UFLA (Lavras, Brazil), International Culture Collection of Vesicular-Arbuscular Mycorrhizal Fungi (INVAM), Swiss collection of Arbuscular Mycorrhizal Fungi (SAF, Agroscope ART Zurich) — and private AM fungal collections curated by Sieverding, Oehl, Trappe, Błaszkowski, and McGee, see also Oehl et al. [9]. The Hall & Abbott [27] photographic slide collection was also reviewed. Older specimens (mounted on microscopic slides prior to 1990) were mostly mounted in lactophenol, while others were fixed with polyvinyl alcohol-lactic acid-glycerol (PVLG) or in a mixture of PVLG + Melzer’s reagent, which post-1990 are the principal fixing media [28]. Newly mounted spores and sporocarps from collections or from cultures were fixed using the latter two fixing media, or occasionally also in a mixture of 1:1 lactic acid to water, in Melzer’s reagent, and in water. When available, spores freshly isolated from soils or bait cultures were also mounted and analyzed. All spore observations and all information on spore characteristics are based on spores extracted from soil, from trap cultures or from single or multiple spore-derived pure cultures. No information is provided from in-vitro-cultured materials. Spore wall terminology follows the nomenclature of Walker [29] and Stürmer & Morton [30]. Analyses of the spore walls, the germination structures and all other mycorrhizal structures were performed using compound microscopes at 100–1000×. For this paper, all original species descriptions and published species emendations were also considered and thoroughly studied.
3. Results
3.1. Molecular Phylogeny
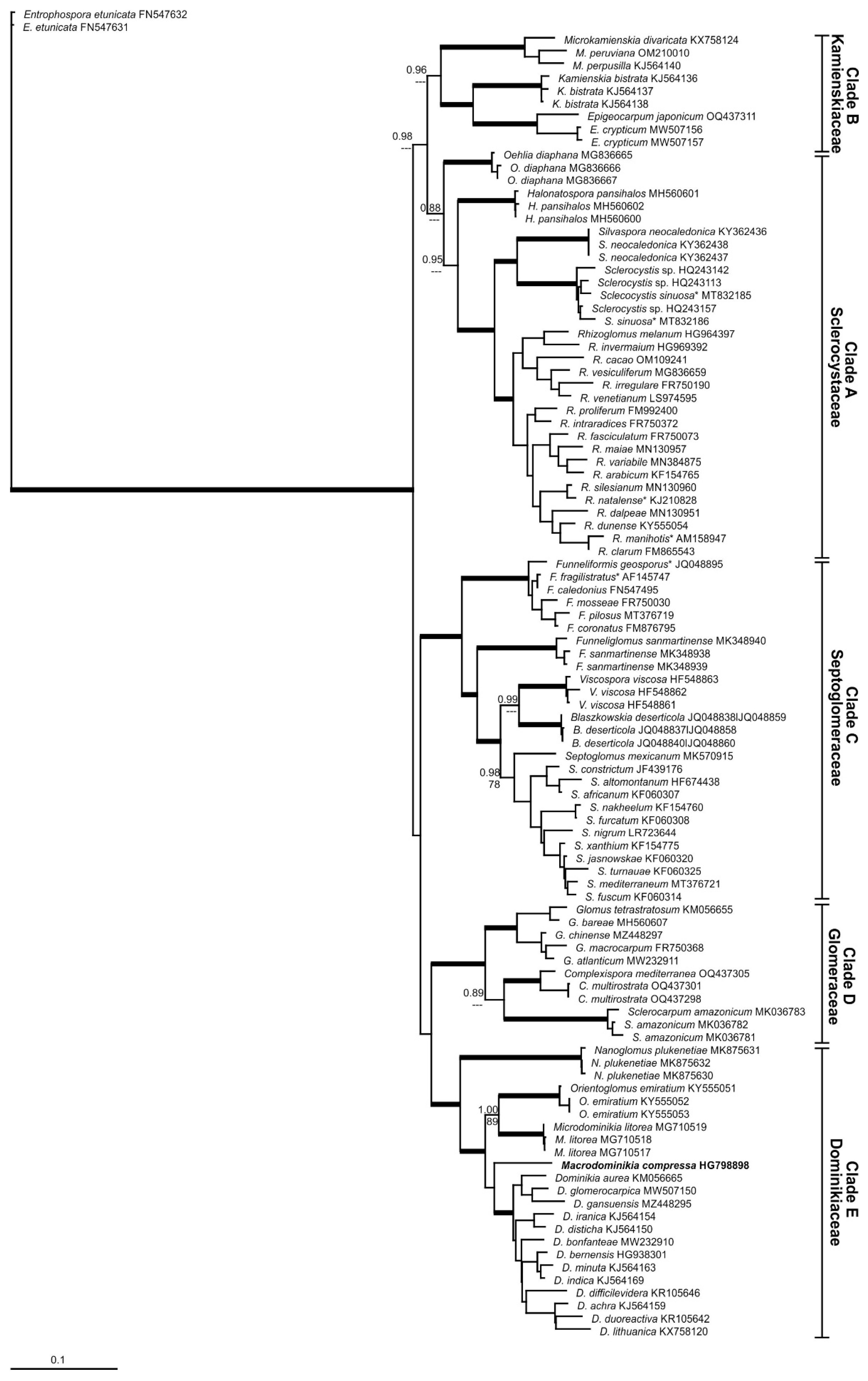
By observing the generated phylogenetic trees, five different clades were separated in the Glomerales (Figure 1 and Figure 2). The clade “A”, composed by the genera Halonatospora, Oehlia, Rhizoglomus, Sclerocystis and Silvaspora, had strong support for Bayesian inference (BI) and ML analyses (1.00/88%) in the first tree obtained (Figure 1), however was not supported by ML analyses and presented a support of 0.88 for BI in the second tree (Figure 2). The clade “B” presented strong support for BI in the two trees obtained, however shown support for ML (94%) just in the first tree. This clade comprises the genera Epigeocarpum, Kamienskia and Microkamienskia. The clades “C”, “D” and “E” showed strong support in all analyses of the two trees. Clade “C” is represented by Blaszkowskia, Funneliformis, Funneliglomus, Septoglomus and Viscospora, clade “D” by Complexispora, Glomus and Sclerocarpum, and clade “E” by Dominikia, Macrodominikia, Microdominikia, Nanoglomus and Orientoglomus, respectively. Our findings indicate that each clade represent a different family in Glomerales with in total 22 AMF genera, which can be separated morphologically by a combination of spore morphology, characteristics of the subtending hyphae and pore closure, and spore formation features in sporocarps, spore clusters or exclusively singly in soils (Supplementary Material, Table S1).
3.2. Taxonomy
Glomerales J.B. Morton and Benny, emend. Błaszk., B.T. Goto, and Magurno, Mycotaxon 37: 473. 1990.
MycoBank MB 90425
Description: Spores formed in soils or in roots, terminally on or intercalary in hyphae, either singly, in loose spore clusters, or in multiple-spored loose to compact sporocarps, when compact spores are randomly distributed or organized around a central plexus of hyphae. Compact sporocarps with or without peridium, and with or without intrasporocarpic hyphal gleba. Spores with one mono-to-multiple layered wall. Wall of the subtending hyphae (SH) conspicuously continuous and concolorous with the spore wall, or slightly lighter in color than the spore wall; SH funnel-shaped, cylindrical or constricted; straight, curved or flared; pore at spore base open, or closed by a bridging, straight or curved septum or by introverted wall thickening, or SH closed at some distance from the spore base by a septum or plug-like structure forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue in trypan blue.
Type family: Glomeraceae Piroz. & Dalpé
Other families: Septoglomeraceae Oehl et al., Sclerocystaceae Oehl et al., Dominikiaceae G.A. Silva et al., Kamienskiaceae G.A. Silva et al.
Type genus: Glomus Tul. & C. Tul.
Other genera: Sclerocarpum B.T. Goto et al., Complexispora Błaszk. et al., Simiglomus Sieverd. et al., Blaszkowskia G.A. Silva & Oehl, Septoglomus Sieverd. et al., Funneliformis C. Walker & A. Schüssler, Funneliglomus Corazon-Guivin et al., Viscospora Sieverd. et al., Rhizoglomus Sieverd. et al., Oehlia Błaszk. et al., Sclerocystis Berk. & Broome, Halonatospora Błaszk. et al., Kamienskia et al. Błaszk. et al., Microkamienskia G.A. Silva et al., Dominikia Błaszk. et al., Macrodominikia Oehl et al., Microdominikia Oehl et al., Nanoglomus Corazon-
Guivin et al., Orientoglomus G.A. Silva et al., Epigeocarpum Błaszk., et al., Silvaspora Błaszk., et al., Parvocarpum Magurno.
Glomeraceae Piroz. & Dalpé emend. Oehl, G.A. Silva & Sieverd., Symbiosis 7: 19. 1989.
MycoBank 82026
Emended description: Spores formed in soil and sometimes in roots, terminally on or intercalary in hyphae, either singly, in loose spore clusters, or in multiple-spored loose to compact sporocarps, when compact, then spores are randomly distributed, but not organized around a central plexus of hyphae. Compact sporocarps with or without peridium, and with or without intrasporocarpic hyphal gleba. Spores with one (mono-)bi-to-multiple layered wall. SH wall conspicuously continuous and concolorous with the spore wall, or slightly lighter in color than the spore wall; SH cylindrical or slightly funnel-shaped or slightly constricted; straight, curved or flared; pore at spore base open, or closed at spore base by a straight or curved septum or by introverted wall thickening, or at some distance from the spore base by a bridging septum arising from the SH wall; forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue in trypan blue.
Type genus: Glomus Tul. & C. Tul.
Other genera: Sclerocarpum B.T. Goto et al., Simiglomus Sieverd. et al., Complexispora Błaszk. et al.
Glomus Tul. & C. Tul., Giorn. Bot. Ital., Anno 1, 2(7–8): 63. 1845.
MycoBank 20244
Emended description: Spores formed singly within soil or sometimes roots, in disorganized, multiple-spored loose spore clusters or in compact sporocarps; compact, but not radially organized around a central hyphal plexus, without or with peridium, spores are either in sporocarp, or organized. Spores with a mono-to-multiple layered wall. SH wall conspicuously continuous and concolorous with the spore wall, or slightly lighter in color than the spore wall. Spore pore closure often by introverted wall thickening, sometimes supported by a short bridging septum, rarely open. Forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue in trypan blue.
Type species: Glomus macrocarpum Tul & C. Tul., Giorn. Bot. Ital., Anno 1, 2(7–8): 63. 1845.
MycoBank MB 240247
≡ Endogone macrocarpa (Tul. & C. Tul.) Tul. & C. Tul., Fungi Hypog.: 182. 1851.
MycoBank MB 218537
≡ Endogone guttulata E. Fisch., Ber. Schweiz. Bot. Ges. 32: 13. 1923.
≡ Endogone nuda Petch., Ann. R. Bot. Gdns Peradeniya 9: 322. 1925.
≡ Endogone pampaloniana Bacc., Nuovo Giorn. Bot. Ital., n.s. 10: 90. 1903.
≡ Paurocotylis fulva var. zelandica Cooke, Grevillea 8: 59. 1879.
Simiglomus Sieverd., G.A. Silva & Oehl, Mycotaxon 116: 104. 2011.
MycoBank MB 518435
Description: Spores formed in soil or in roots, terminally on or intercalary in hyphae, in small to multiple-spored loose clusters, randomly distributed, but not organized around a central plexus of hyphae. Loose sporocarps generally without peridium, and without intrasporocarpic hyphal gleba. Spores with one mono-to-multiple layered wall. SH wall conspicuously continuous and concolorous with the spore wall, or slightly lighter in color than the spore wall; SH cylindrical or slightly to rarely slightly funnel-shaped or constricted; straight, curved or flared; pore open at spore base, but often in some distance by one to multiple bridging, straight to curved, septum, not supported by introverted wall thickening; forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue in trypan blue.
Type species: Simiglomus hoi (S.M. Berch & Trappe) G.A. Silva, Oehl & Sieverd.
MycoBank MB 518461
Basionym: Glomus hoi S.M. Berch & Trappe, Mycologia 77: 654. 1985.
Sclerocarpum B.T. Goto, Błaszk., Niezgoda, Kozłowska & Jobim, Mycol. Progr. 18(3): 375. 2019.
MycoBank MB 828316
Description: Producing spores in scleroid, epigeous and sub-hypogeous, light- to dark-colored unorganized sporocarps, very hard to break, with a peridium and a gleba comprising hyphae and glomoid glomerospores (= spores) with a single SH; spores hyaline; globose to subglobose; small, < 100 μm diam; frequently ovoid; Spore wall composed a few smooth layers, of which one layer laminate and much thicker than the outermost layer, forming the spore surface. SH funnel-shaped with a wall continuous with the spore wall; pore narrow, open or occluded by thickening of the SH wall, or (rarely) occluded by a straight or slightly invaginated septum continuous with some innermost laminae of spore wall layer 2; septum, positioned at the spore base.
Type species: Sclerocarpum amazonicum B.T. Goto, Błaszk., Niezgoda, Kozłowska & Jobim, Mycol. Progr. 18: 377. 2019.
MycoBank MB 828317
Specific morphological observation on Sclerocarpum
Especially Sclerocarpum shows well-defined, outstanding morphological characteristics of their hard sporocarps, their spores, which are exclusively formed within these sporocarps and of their funnel-shaped subtending hyphae including a pronounced wall-thickening at the spore base at the point of pore closure. Three Glomus species described in the past, clearly combine with all these characteristics. These are G. convolutum, G. pellucidum and G. segmentatum [31,32,33]. Consequently, these species are hereafter transferred to the genus Sclerocarpum.
Sclerocarpum convolutum (Gerd. & Trappe) Oehl, Sieverd. & G.A. Silva, comb. nov.
MycoBank MB 855472
Basionym: Glomus convolutum Gerd. & Trappe, Mycol. Mem. 5: 42. 1974. Mycological Memoirs 5: 42. 1974.
MycoBank MB 314590
Sclerocarpum segmentatum (Trappe, Spooner & Ivory) Oehl, Sieverd. & G.A. Silva, comb. nov.
MycoBank MB 855473
Basionym: Glomus segmentatum Trappe, Spooner & Ivory, Trans. Br. Mycol. Soc. 73: 362. 1979.
MycoBank MB 314610
Sclerocarpum pellucidum (McGee & Pattinson) Oehl, Sieverd. & G.A. Silva, comb. nov.
MycoBank MB 855474
Basionym: Glomus pellucidum McGee & Pattinson, Austral. Syst. Bot. 15: 120. 2002.
MycoBank MB 374910
Complexispora Błaszk., B.T. Goto, Niezgoda & Magurno, Mycol. Progr. 22(5, no. 34): 7. 2023.
MycoBank MB 847607
Description: Producing hypogeous glomoid spores singly and in clusters. Spores four to six layers, of which two consist of tightly adherent sublayers (laminae). SH cylindrical to funnel-shaped, concolorous with the spore wall layer, with a wall composed of layers continuous with all spore wall layers, except for the innermost layer. Pore closed by a septum continuous with the innermost spore wall layer, occasionally also by a septum connecting the inner surfaces of the main structural laminate spore wall layer. Forming mycorrhiza with vesicles and arbuscules staining dark in Trypan blue.
Type species: Complexispora multistratosa Błaszk., B.T. Goto, Niezgoda & Magurno, Mycol. Progr. 22(5, no. 34): 6. 2023.
MycoBank MB 847608
Septoglomeraceae Oehl, G.A. Silva, Sieverd. fam. nov.
Mycobank MB 855466
Description: Spores formed in soil and sometimes in roots, terminally on or intercalary in hyphae, either singly, in loose spore clusters or sporocarps. Compact sporocarps with or without peridium, without intrasporocarpic hyphal gleba. Spores with one mono-to-multiple layered wall. SH wall conspicuously continuous and concolorous with the spore wall, or slightly lighter in color than the spore wall; SH pronounced funnel-shaped, slightly funnel-shaped, cylindrical or constricted, rarely inflated; straight, curved or flared; forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue in trypan blue.
Type genus: Septoglomus Sieverd. et al.
Other genera: Funneliformis C. Walker & A. Schüssler, Funneliglomus Corazon-Guivin et al., Viscospora Sieverd. et al., Blaszkowskia G.A. Silva & Oehl
Type species: Septoglomus constrictum (Trappe) Sieverd.
Septoglomus Sieverd., G.A. Silva & Oehl, Mycotaxon 116: 105. 2011.
MycoBank MB 518436
Emended description: Spores formed in soil and sometimes in roots, terminally on or intercalary in hyphae, either singly, in loose spore clusters. Spores with one mono-to-multiple layered wall. SH wall conspicuously continuous and concolorous with the spore wall, or slightly lighter in color than the spore wall; SH constricted to cylindrical, rarely slightly funnel-shaped, or inflated in some distance to the spore base; straight, curved or flared; pore closed by a broad, pronounced septum or a plug at spore base or in some distance from spore; forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue in trypan blue.
Type species: Septoglomus constrictum (Trappe) Sieverd., G.A. Silva & Oehl. Mycotaxon 116: 105. 2011.
MycoBank MB 518462
Basionym: Glomus constrictum Trappe, Mycotaxon 6: 361. 1977.
MycoBank MB 314589
≡ Funneliformis constrictus (Trappe) C. Walker & A. Schüssler, The Glomeromycota – a species list: 14. 2010.
MycoBank MB 542904
Blaszkowskia G.A. Silva & Oehl, Mycol. Progr. 22(11, no. 74): 5. 2023.
MycoBank MB 847414
Description: Spores light yellow to yellow brown to dark brown or dark red brown, generally 20–200 μm, with > 1 spore wall layer; SH funnel-shaped, rarely constricted, thick-walled; spore pores might be open at the spore base, but often closed by one to several thin septa within SH at some distance from the spore base. SH thicker-walled than mycelium hyphae and pigmented over long distances and within the whole spore clusters, including intraradical spore formations (> 100 μm).
Type species: Blaszkowskia deserticola (Trappe, Bloss & J.A. Menge) Oehl & G.A. Silva. Mycol. Progress 22 (11, no. 74): 5. 2023.
MycoBank MB 847415
Basionym: Glomus deserticola Trappe, Bloss & J.A. Menge, Mycotaxon 20: 123. 1984.
MycoBank MB 106847
≡ Septoglomus deserticola (Trappe, Bloss & J.A. Menge) G.A. Silva, Oehl & Sieverd. Mycotaxon 116: 106. 2011.
MycoBank MB 518463
Funneliglomus Corazon-Guivin, G.A. Silva & Oehl, Sydowia 71: 19. 2019.
MycoBank MB 829266
Description: Spores formed in soil or rarely in roots, terminally on or intercalary in hyphae, singly or in small clusters; the conspicuous SH is concolorous with spore wall color (or slightly lighter in color), SH is species-specific and funnel-shaped to slightly funnel-shaped, to rarely cylindrical or slightly constricted, or inflated in some distance to the spore base; straight, curved or flared. Pore regularly closed by a conspicuous septum that species-specifically may arise from the structural wall layer, from an additional adherent innermost, (semi-)flexible lamina, or from both but not by introverted wall thickening, which is lacking. Forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue in trypan blue.
Type species: Funneliglomus sanmartinense Corazon-Guivin, G.A. Silva & Oehl. Sydowia 71: 21. 2019
MycoBank MB 829267 (Funneliglomus sanmartinensis), MycoBank MB 830216 (corrected to Funneliglomus sanmartinense by Mycobank after publication)
Funneliformis C. Walker & A. Schüssler, emend. Oehl, G.A. Silva & Sieverd., The
Glomeromycota – a species list with new families and genera: 13. 2010.
MycoBank MB 542894
Description: Spores formed within soil or rarely in roots, singly or sometimes in sporocarps with a few to several spores per sporocarp only; the conspicuous SH is concolorous with spore wall color (or slightly lighter in color), SH is species-specific and generally pronounced funnel-shaped to slightly funnel-shaped to rarely cylindrical. Wall differentiation and pigmentation may continue over long distances from the spore base (often > 50–250 μm), then mycelium may become hyaline. Pore regularly closed by a conspicuous septum that species-specifically arises from the structural wall layer, from an additional adherent innermost, (semi-)flexible lamina, or from both but not by introverted wall thickening, which is lacking. Forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue in trypan blue.
Type species: Funneliformis mosseae (T.H. Nicolson & Gerd.) C. Walker & A. Schüssler, The Glomeromycota – a species list with new families and genera: 13. 2010.
MycoBank MB 542895
Basionym: Endogone mosseae T.H. Nicolson & Gerd., Mycologia 60: 314. 1968.
MycoBank MB 330367
≡ Glomus mosseae (T.H. Nicolson & Gerd.) Gerd. & Trappe, Mycol. Mem. 5: 40. 1974.
MycoBank MB 314604
Viscospora Sieverd., Oehl & G.A. Silva, Mycotaxon 116: 108: 2011.
MycoBank MB 518439
Emended description: Spores hyaline to white or subhyaline, terminally or intercalary on pronounced funnel-shaped to slightly funnel-shaped to cylindrical or inflated subtending hyphae, hyaline, singly, or in loose clusters with up to 100 spores per cluster. Outer layer is evanescent, viscose, gathering large amounts of debris during degradation. SWL2 is persistent, unite to laminate, and might stain whitish yellow to yellowish, when exposed to Melzer's reagent. SH straight or recurved, sometimes folded to rarely flared. Regularly, one to several straight to curved septa are formed in the SH close to the spore base or in a distance of up to 10–100 μm to the spore base. The spore wall layers continue in the SH, with similar to slightly smaller thickness towards the mycelium hyphae.
Type species: Viscospora viscosa (T.H Nicolson) Sieverd., Oehl & G.A. Silva. Mycotaxon 116: 108. 2011.
MycoBank MB 518471
Basionym Glomus viscosum T.H. Nicolson, Mycol. Res. 99: 1502. 1995.
MycoBank MB 413125
≡ Septoglomus viscosum (T.H. Nicolson) C. Walker, D. Redecker, D. Stiller & A. Schüßler: Mycorrhiza 23: 524. 2013
MycoBank MB 550089
Sclerocystaceae Oehl, G.A. Silva, & Sieverd. fam. nov.
MycoBank MB 855467
Description: Spores formed in loose sporocarps, in clusters, or singly in soil, and frequently in roots. When formed in compact sporocarps, spores can be randomly distributed on SH of different length, or well-organized around a central plexus of hyphae. SH wall continuous with the spore wall, and for a certain distance concolorous with the spore wall, or slightly lighter in color. SH cylindrical or seldom slightly funnel shaped at spore base. Pore at spore base open, or closed by a septum. Spore walls show rarely one, to generally two to three (and up to five) distinct layers, of which one or several of the outermost may separate when pressure is applied to spores. Forming vesicular-arbuscular mycorrhizae, whose fungal structures stain blue to dark blue in trypan blue.
Type genus: Sclerocystis Berk. & Broome
Other genera: Oehlia Błaszk. et al., Rhizoglomus Sieverd. et al., Halonatospora Błaszk. et al., Silvaspora Błaszk. et al., Parvocarpum Magurno
Sclerocystis Berk. & Broome, J. Linn. Soc., Bot. 14 (73 & 74): 137. 1873.
MycoBank MB20512
Emended description: Spores formed in compact sporocarps, in soils or roots, in cultures sometimes also singly or in loose clusters. When formed in compact sporocarps, then spores are regularly organized, arising radially around a central plexus of hyphae. SH wall continuous with the spore wall, and for a certain distance concolorous with the spore wall, or slightly lighter in color. SH funnel-shaped to cylindrical at spore base. Pore at spore base regularly closed by a fine septum, which is formed within the small pore channel within the spore base. Spore walls show more than one, and generally two or rarely > two distinct layers. Forming vesicular-arbuscular mycorrhizae, whose fungal structures stain blue to dark blue in trypan blue.
Type species: Sclerocystis coremioides Berk. & Broome, J. Linn. Soc., Bot. 14(no. 74): 137 (1873) [1875]
MycoBank MB 213141
≡ Glomus coremioides (Berk. & Broome) D. Redecker & J.B. Morton
MycoBank MB 464612
Rhizoglomus Sieverd., G.A. Silva & Oehl, Mycotaxon 129: 377. 2015.
MycoBank MB 803191
Description: Spores formed in loose sporocarps, in clusters, or singly, in soil and frequently also in roots. When formed in compact sporocarps, they are not organized around a central plexus of hyphae. SH wall continuous with the spore wall, and for a certain distance concolorous with the spore wall, or slightly lighter in color. SH cylindrical or seldom slightly funnel shaped at spore base. Pore at spore base regularly open, rarely closed by a septum. Spore walls show more than one, and generally two to three (and up to five) distinct layers, of which one or several of the outermost may separate when pressure is applied to spores. Forming vesicular-arbuscular mycorrhizae, whose fungal structures stain blue to dark blue in trypan blue.
Type species: Rhizoglomus intraradices (N.C. Schenck & G.S. Sm.) Sieverd., G.A. Silva & Oehl, Mycotaxon 129(2): 378 (2015) [2014]
MycoBank MB 803192
Basionym: Glomus intraradices N.C. Schenck & G.S. Sm., Mycologia 74: 78. 1982.
MycoBank MB 110704
≡ Rhizophagus intraradices (N.C. Schenck & G.S. Sm.) C. Walker & A. Schussler, The Glomeromycota: 19. 2010.
MycoBank MB 542910
Oehlia Błaszk., Kozłowska, Niezgoda, B.T. Goto & Dalpe, Nova Hedwigia 107: 507. 2018.
MycoBank MB 824689
Emended description: Spores formed in loose sporocarps, in clusters, or singly, in soil and frequently also in roots. When formed in compact sporocarps, they are not organized around a central plexus of hyphae. SH wall continuous with the spore wall, and for a certain distance concolorous with the spore wall, or slightly lighter in color. SH cylindrical or seldom slightly funnel shaped at spore base. Pore at spore base regularly closed by a septum. Spore wall shows more than one, and generally two to three (and up to five) distinct layers. Forming vesicular-arbuscular mycorrhizae, whose fungal structures stain blue to dark blue in trypan blue.
Type species: Oehlia diaphana (J.B.Morton & C.Walker) Błaszk., Kozłowska & Dalpe, Nova Hedwigia 107(3-4): 507 (2018)
MycoBank MB 824693
Basionym: Glomus diaphanum J.B.Morton & C.Walker. Mycotaxon 21: 433, 1984.
MycoBank MB 106161
Parvocarpum Magurno, MycoKeys 107: 283. 2024.
MycoBank MB 853558
Description: Producing glomoid spores mainly in small sporocarps, irregularly around a small central plexus of interwoven, intrasporocarpic hyphae, or singly in soils. SH generally short and cylindrical, often breaking at the spore base during sporocarp degradation. Pore closed by introverted wall thickening at the spore base, and additionally by a septum arising from the laminated wall layer and/or an additional flexible inner layer.
Type species: Parvocarpum badium (Oehl, Redecker & Sieverd.) Magurno, MycoKeys 107: 284. 2024
MycoBank MB 853560
Basionym: Glomus badium Oehl, D. Redecker & Sieverd., J. Appl. Bot. Food Qual. 79: 39. 2005.
MycoBank MB 341387
≡ Funneliformis badius (Oehl, D. Redecker & Sieverd.) C. Walker & A. Schüssler, The Glomeromycota – a species list: 13. 2010.
MycoBank MB 541897
Silvaspora Błaszk., Niezgoda, B.T. Goto, Crossay & Magurno, Frontiers in Microbiology 12 (no. 655910): 14. 2021.
MycoBank 838881
Description: Forming pigmented spores with 2-3 wall layers, of which only layer 1, forming the spore surface, is impermanent and hyaline to brightly colored. SH colored similarly to the spore wall, cylindrical, slightly funnel-shaped, or constricted at the spore base, with a pore occluded due to thickening of that SH wall layer continuous with the laminate spore wall layer, rarely slightly open. Forming mycorrhiza with arbuscules, vesicles, and hyphae staining dark in trypan blue.
Type species: Silvaspora neocaledonica (D. Redecker, Crossay & Cilia) Błaszk., Niezgoda, B.T. Goto, Crossay & Magurno, Frontiers in Microbiology 12(no. 655910): 14 (2021)
MycoBank MB 838882
≡ Rhizoglomus neocaledonicum (D. Redecker, Crossay & Cilia) Oehl, Turrini & Giovann.
MycoBank MB 827095
Basionym: Rhizophagus neocaledonicus D. Redecker, Crossay & Cilia, Mycological Progress 17: 739. 2018.
MycoBank MB 820537
Halonatospora Błaszk., Niezgoda, B.T. Goto & Kozłowska, Botany 96(11): 743. 2018.
MycoBank MB 826963
Description: Spores formed in loose clusters, or singly, in soil and frequently in roots. When formed in sporocarps, they are not organized around a central plexus of hyphae. SH wall continuous with the spore wall, and for a certain distance concolorous with the spore wall, or slightly lighter in color. Outer spore wall layer strongly swells in PVLG, forming a halo with radiate columns. SH cylindrical or slightly flared, sometimes slightly constricted at spore base, straight or slightly curved. Pore open or closed by a septum at spore base. Spore wall shows more than one, and generally two to three (and up to five) distinct layers. Forming vesicular-arbuscular mycorrhizae, whose fungal structures stain blue to dark blue in trypan blue.
Type species: Halonatospora pansihalos (S.M. Berch & Koske) Błaszk., Niezgoda, B.T. Goto & Kozłowska, Can. J. Bot. 96(11): 743 (2018)
MycoBank MB 826964
Basionym: Glomus pansihalos S.M. Berch & Koske. Mycologia 78: 832, 1986.
MycoBank MB 358213
Kamienskiaceae G.A. Silva, Sieverd. & Oehl
MycoBank MB 855468
Description: Spores hypogeous and/or intraradical, produced in loose clusters to compact sporocarps, and never organized around a central plexus of hyphae. Spores hyaline, small, up to 50 μm diam. when globose. Spore wall with two permanent, smooth layers, of which one may stain in Melzer’s reagent. The structural laminate layer is layer 1 or 2. SH cylindrical to funnel-shaped with an open pore. Forming vesicular-arbuscular mycorrhizae staining dark in trypan blue.
Type genus: Kamienskia Błaszk. et al.,
Other genus: Microkamienskia Corazon-Guivin, G.A. Silva & Oehl, Epigeocarpum Błaszk., B.T. Goto, Jobim, Niezgoda & Marguno
Kamienskia Błaszk., Chwat & Kovács, Nova Hedwigia 100(1-2): 230. (2014) [2015].
MycoBank MB 808260
Emended description: Spores hypogeous and/or intraradical, produced in loose clusters to compact, but fast degrading sporocarps, but never organized around a central plexus of hyphae. Spores hyaline, small, up to 50 μm diam. when globose. Spore wall with two permanent, smooth layers. The structural laminate layer might be layer 1 or 2. SH regularly funnel-shaped with an open pore. Forming vesicular-arbuscular mycorrhizae staining dark in trypan blue.
Type species: Kamienskia bistrata (Błaszk., D. Redecker, Koegel, Symanczik, Oehl & Kovács) Błaszk., Chwat & Kovács, Nova Hedwigia 100(1-2): 230. (2014) [2015].
MycoBank MB 808261
Basionym: Glomus bistratum Błaszk., D. Redecker, Koegel, Symanczik, Oehl & Kovács, Botany 87: 267. 2009.
MycoBank MB 512540
Microkamienskia Corazon-Guivin, G.A. Silva & Oehl, Nova Hedwigia 109: 359. 2019.
MycoBank MB 830814
Description: Spores hypogeous and/or intraradical, produced in loose clusters to compact, but fast degrading sporocarps, but never organized around a central plexus of hyphae. Spores hyaline, small, up to 50 μm diam. when globose. Spore wall with two permanent, smooth layers, of which one may stain in Melzer’s reagent. The structural laminate layer might be layer 1 or 2. SH cylindrical to rarely slightly funnel-shaped with an open pore. Forming vesicular-arbuscular mycorrhizae staining dark in trypan blue.
Type species: Microkamienskia perpusilla (Błaszk. & Kovács) Corazon-Guivin, G.A. Silva & Oehl. Nova Hedwigia 109: 361. 2019.
MycoBank MB 830815
Basionym: Glomus perpusillum Błaszk. & Kovács. Mycologia 101: 249. 2009.
MycoBank MB 512346
≡ Kamienskia perpusilla (Błaszk. & Kovács) Błaszk., Chwat & Kovács. Nova Hedwigia 100: 231. 2015-
MycoBank MB 808264
Epigeocarpum Błaszk., B.T. Goto, Jobim, Niezgoda & Marguno, Frontiers in Microbiology 12 (no. 655910): 10. 2021.
MycoBank MB 838879
Description: Spores hyaline to light yellow, formed in compact, unorganized sporocarps. Spores have 2–3 wall layers, laminate layer usually transferring into a crown-like structure due to contracting in spores crushed in PVLG and PVLG + Melzer’s reagent. The channel connecting the lumen of the SH with the interior of spores is closed by a septum usually positioned at half the thickness of the laminate layer; SH funnel-shaped, SH lumen gradually narrowing in maturing spores due to internal thickening of the laminate wall layer.
Type species: Epigeocarpum crypticum Jobim, Błaszk., Niezgoda, Magurno & B.T. Goto, Frontiers Microbiol. 12(no. 655910): 14. 2021.
MycoBank MB 838880
Dominikiaceae G.A. Silva, Sieverd. & Oehl
MycoBank MB 855469
Description: Spores hypogeous and/or intraradical, produced in loose clusters to compact sporocarps, but never organized around a central plexus of hyphae. Spores hyaline to creamy or yellow to yellow brown; spore wall with one to regularly two, or > 2 layers. SH wall continuous with the spore wall, and for a certain distance concolorous with the spore wall, or slightly lighter in color. Spore pore often closed at spore base by a thin bridging septum, sometimes supported by a certain degree of introverted wall thickening, rarely continuing in the SH for a short distance (2-10 μm). Forming vesicular-arbuscular mycorrhizae staining dark in trypan blue.
Type genus: Dominikia Błaszk. et al.,
Other genera: Macrodominikia Oehl et al., Microdominikia Oehl et al., Nanoglomus Corazon-Guivin et al., Orientoglomus G.A. Silva et al.
Dominikia Błaszk., Chwat & Kovács, Nova Hedwigia 100(1-2): 228. (2014) [2015].
MycoBank MB 808255
Emended description: Spores hypogeous and/or intraradical, produced in loose clusters to compact sporocarps but never organized around a central plexus of hyphae. Spores hyaline to creamy or yellow to yellow brown, small, up to 70 μm diam; spore wall with one to regularly two, or > 2 layers. SW wall continuous with the spore wall, and for a certain distance concolorous with the spore wall, or slightly lighter in color. Spore pore often closed at spore base by a thin bridging septum, sometimes supported by a certain degree of introverted wall thickening, rarely continuing in the SH for a short distance. Forming vesicular-arbuscular mycorrhizae staining dark in trypan blue.
Type species: Dominikia minuta (Błaszk., Tadych & Madej) Błaszk., Chwat & Kovács, Nova Hedwigia 100: 230. (2014) [2015].
MycoBank MB 808256
Basionym: Glomus minutum Błaszk., Tadych & Madej. Mycotaxon 76: 189. 2000.
Macrodominikia Oehl, Sieverd. & G.A. Silva, gen. nov.
MycoBank 855470
Description: Spores hypogeous and/or intraradical, produced singly or in loose clusters to loose sporocarps but never organized around a central plexus of hyphae. Spores creamy to yellow to yellow brown, generally 70-110 μm diam; spore wall with one to regularly two, or > 2 layers. SH wall continuous with the spore wall, and for a certain distance concolorous with the spore wall, or slightly lighter in color. Spore pore often closed at spore base by a thin bridging septum, supported by a large degree of introverted wall thickening, continuing in the SH for 10-50 μm distance, forming an irregular, tortuous pore channel. Forming vesicular-arbuscular mycorrhizae staining dark in trypan blue.
Etymology: Macro-, -dominikia, referencing to the spores, which are larger than known hitherto for other genera in the Dominikiaceae
Type species: Macrodominikia compressa (Sieverd., Oehl, Palenz., Sánchez-Castro & G.A. Silva) Oehl, Siev. & G.A. Silva. comb. nov.
MycoBank MB 855471
Basionym: Glomus compressum Błaszk., Tadych & Mad Sieverd., Oehl, Palenz., Sánchez-Castro & G.A. Silva. Nova Hedwigia 99: 433. 2014.
MycoBank MB 807530
≡ Dominikia compressa (Sieverd., Oehl, Palenz., Sánchez-Castro & G.A. Silva) Oehl, Palenz., Sánchez-Castro & G.A. Silva. Nova Hedwigia. 101: 71. (2014) [2015]
MycoBank MB 809861
Microdominikia Oehl, Corazon-Guivin & G.A. Silva, Mycological Progress 18(12): 1400. 2019.
MycoBank MB 831098
Description: Spores formed singly or in clusters in soils or rarely in roots, terminally or intercalary on SH, globose to subglobose, generally hyaline to subhyaline, 10–45 μm in diameter, rarely egg-shaped or irregular, 25–50 × 20–35 μm, with a permanent outer spore wall layer. SH straight or recurved, usually funnel-shaped, more rarely cylindrical, rarely slightly constricted at the spore base. Spore pores open at the spore base, but closed within SH at some distance.
Type species: Microdominikia litorea (Błaszk. & Kozłowska) Oehl, Corazon-Guivin & G.A. Silva, Mycological Progress 18 (12): 1400 (2019)
MycoBank MB 831099
Basionym: Dominikia litorea Błaszk. & Kozłowska, Phytotaxa 338(3): 246 (2018)
MycoBank MB 823832
Nanoglomus Corazon-Guivin, G.A. Silva & Oehl, Mycological Progress 18(12): 1398. 2019.
MycoBank MB 831096
Description: Nanoglomus species differentiate small spores, generally <40 μm in diameter, when globose, singly or in loose clusters, terminally or intercalary on cylindrical to slightly funnel-shaped, rarely inflating SH, which are concolorous with the spores and have a fine, straight, or rarely curved septum closing the pore at the spore base. So far, they can be differentiated from Dominikia spp. by the generally smaller and thinner walled spores, and by molecular phylogeny on the partial SSU, ITS region, and partial LSU rDNA.
Type species: Nanoglomus plukenetiae Corazon-Guivin, G.A. Silva & Oehl, Mycological Progress 18 (12): 1398 (2019)
MycoBank MB 831097.
Orientoglomus G.A. Silva, Oehl & Corazon-Guivin, Mycological Progress 18(12): 1400. 2019.
MycoBank MB 831100
Description: Glomeraceae spores formed singly or in clusters in soil or rarely in roots, terminally on SH, globose to subglobose, 40–90 μm in diameter, rarely egg-shaped, 50–100 × 40–85 μm; SH wall with significant and regular thickening, resulting in a cylindrical pore channel. Outermost spore wall layer permanent, smooth, at least one spore wall layer laminate. Pore open or closed by a straight to curved to septum.
Type species: Orientoglomus emiratium (Błaszk., Kozłowska, Mullath, AlDhaheri & Al-Yahya'ei) G.A. Silva, Oehl & Corazon-Guivin, Mycological Progress 18 (12): 1403 (2019)
MycoBank MB 831101.
Basionym: Dominikia emiratia Błaszk., Kozłowska, Mullath, AlDhaheri & Al-Yahya'ei, Botany 95(7): 632 (2017)
MycoBank MB 819815
4. Discussion
In the present study, the family Glomeraceae of the order Glomerales was divided into five different families, based on five different phylogenetic major clades, well known and increasingly elaborated within the last fifteen years of research on the ribosomal gene. Also in this study, the clades “B”, “C”, “D” and “E” were very well supported by BI and ML analyses, using different datasets (except, clade “B”, supported by ML just in the first tree) to generate the trees and indicate very stable taxa. The clade “A” was supported with good values just in the first tree (Figure 1). The clade “A” formed a more heterogeneous group, and we believe that in the future this group will be divided in different families. For now, we understand that the better solution is maintain this group as a single family in Glomerales, mainly because the support in the first tree was very strong. These same clades were found in the tree generated by Corazon-Guivin et al. [21] and Tedersoo et al. [6]. The families Glomeraceae, Dominikiaceae, Septoglomeraceae and Sclerocystaceae are also supported by Corazon-Guivin et al. (19,20).
The genera Oehlia and Halonatospora form two other clades well separated from Silvaspora, Sclerocystis and Rhizoglomus and might represent two further families within the Glomerales. Such deeper-going separation is to our opinion fully supported by molecular phylogeny, but in view of the low numbers of taxa, this separation is also not yet needed at this stage of research progress. In our tree, Halotospora was placed near Oehlia, Silvaspora, Sclerocystis and Rhizoglomus. However, according to the tree from Tedersoo et al. [6], Halonatospora is near Kamienskia, Microkamienskia and Epigeocarpum. The tree generated by Tedersoo et al. [6] has different orders from Glomeromycota. The position of some genera can be divergent in relation to our trees from Glomerales, which confirms also the lower degree of relationship between Halonatospora, Oehlia and the other genera clustering together within the Sclerocystaceae.
Morphologically, all genera listed within Glomerales, can clearly be identified based on the combination of their major sporocarp, spore, pore closure, subtending hyphae and spore formation characteristics (see also Silva et al. [5], including an identification key for all genera within the Glomerales). However, attribution to a distinct family is not unequivocally possible, since none of the major morphological characteristics are unique for one of the five families separated here. Some simple indications however, might at least give hints to the family attribution, which is the genus identification itself. Otherwise, our simplified supplementary material (Table S1), shall help to differentiate and attribute fast a specific morphotype to a specific genus and thus to the correct family, to which the morphotype and the genus belongs to. In this study, we accordingly transferred three Glomus species to the genus Sclerocarpum, based on their unique morphological characteristics, fitting exactly to the latter genus. For other Glomeraceae species, new combination would also be justified, either based on molecular phylogeny or morphology of the ‘fruit bodies’, including sporocarps, spores, and their subtending hyphae, but at this stage there is no urgent need to do so.
The genus Parvocarpum [6], was not placed in our analyses because the only sequence available for the P. badium (the type genus) represent a SSU fragment (564bp) and 53bp of ITS1. We conclude that this genus was described mainly based on the high quantity of environmental sequences, which could be attributed to this short SSU fragment [6]. Solely for morphological reasons, there were already sufficient arguments to describe this genus, when the species P. badium was originally described, but at that time the phylogenetic resolution was not given [34], and the most related genus Sclerocystis had been synonymized with Glomus just a few years before, instead of clearly separated.
In our first tree (Figure 1), Macroominikia compressa is not grouped with other Dominikia spp. In the second tree (Figure 2), M. compressa forms a clade with other Dominikia spp., but with no support. According to Tedersoo et al. [6], this species represents a new genus, which is confirmed here and also supported by its distinct spore morphology [5,34]. By including environmental sequences to reliable spore and sequence identifications [36], most probably more genera and even families can be separated, since the phylogenetic resolution will further increase. Rhizoglomus spp. for instance might then be separated from Sclerocystaceae. This separation is already suggested by our phylogenetic trees.
Blaszkowskia was invalidated by Tedersoo et al. [6]. The tree generated by these authors present two different major clades for Viscospora sensu Terderso et al. [6], one clade is divided in two subclades, the first with one sequence from B. deserticola and one environmental sequence and the second with one sequence from V. viscosa and six environmental sequences. The second clade has just environmental sequences (28 sequences) and for us can be a new different genus represented by these sequences. Thus, to our opinion, Blaszkowskia continues to be a valid genus. For morphological reasons, the genera Viscospora and Blaszkowskia, can be easily separated and continued [5].
During the first stages of molecular phylogenetic inclusion into the taxonomy and classification, there were several major synonymizations of AMF genera, also in the Glomeraceae, even for genera with a distinct spore morphology, followed by a time of doubts and resurrections of genera such as Sclerocystis. In the meantime, molecular phylogeny has become the driving force to separate taxa, especially on the genus and species levels. So far, morphology still finds arguments to separate such taxa. With the expected inclusion of environmental sequences directly into taxonomy, morphological identification might lose its importance and significance, and taxa numbers might strongly increase in Glomeromycota, and especially in Glomerales. However, then researchers experienced in morphological identification will start to find the morphs behind such environmental-sequences-born, exclusively molecular-based taxa.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Spreadsheet S1: GenBank accession numbers for the sequences used in this study; Table S1: Main morphological characters of the families and genera within the order Glomerales.
Author Contributions
All authors contributed to the study’s conception and design. Phylogenetic analyses were performed by Gladstone Alves da Silva and morphological analyses by Ewald Sieverding and Fritz Oehl. The manuscript was written and revised by all authors.
Funding
This research received no external funding.
Data Availability Statement
All data (sequences) presented in this article are in GenBank (https://www.ncbi.nlm.nih.gov)
Acknowledgments
Daniele Magna Azevedo de Assis thanks to the Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE) for providing a fellowship. Gladstone A. Silva has a fellowship from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (Proc. 312606/2022-2).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Baltruschat, H.; Santos, V.M.; Silva, D.K.A.; Schellenberg, I.; Deubel, A.; Sieverding, E.; Oehl, F. Unexpectedly high diversity of arbuscular mycorrhizal fungi in fertile Chernozem croplands in Central Europe. Catena 2019, 182, 104135. [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Becerra, A.G.; Thines, M. et al. Outline of fungi and fungus-like taxa. Mycosphere 2020, 11: 1060–1456. [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M, et al. Outline of fungi and fungus-like taxa - 2021. Mycosphere 2022, 13, 53–453. [CrossRef]
- Błaszkowski, J.; Sánchez-García, M.; Niezgoda, P.; Zubek, S.; Fernández, F.; Vila, A.; Al-Yahya’ei, M.N.; Symanczik, S.; Milczarski, P.; Malinowski, R.; Cabello, M.; Goto, B.T.; Casieri, L.; Malicka, M.; Bierza, W.; Magurno, F. A new order, Entrophosporales, and three new Entrophospora species in Glomeromycota. Front. Microbiol. 2022, 13, 962856. [CrossRef]
- Silva, G.A.; Corazon-Guivin, M.A.; Assis, D.M.A.; Oehl, F. Blaszkowskia, a new genus in Glomeraceae. Mycol. Prog. 2023, 22, 74. [CrossRef]
- Tedersoo, L.; Magurno, F.; Alkahtani, S.; Mikryukov, V. Phylogenetic classification of arbuscular mycorrhizal fungi: new species and higher-ranking taxa in Glomeromycota and Mucoromycota (class Endogonomycetes). MycoKeys 2024, 107, 249–271. [CrossRef]
- Morton, J.B.; Benny, G.L. Revised classification of arbuscular mycorrhizal fungi (Zygomycetes): a new order, Glomales, two new suborders, Glomineae and Gigasporineae, and two new families, Acaulosporaceae and Gigasporaceae, with an emendation of Glomaceae. Mycotaxon 1990, 37, 471–491.
- Schüßler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [CrossRef]
- Oehl, F.; Silva, G.A.; Goto, B.T.; Sieverding, E. Glomeromycota: three new genera, and glomoid species reorganized. Mycotaxon 2011, 116, 75–120. [CrossRef]
- Silva, G.A.; Maia, L.C.; Oehl, F. Phylogenetic systematics of the Gigasporales. Mycotaxon 2013, 122, 207–220. [CrossRef]
- Błaszkowski, J.; Yamato, M.; Niezgoda, P.; Zubek, S.; Milczarski, P.; Malinowski, R.; Goto, B.T.; Uszok, S.; Casieri, L.; Magurno, F. A new genus, Complexispora, with two new species, C. multistratosa and C. mediterranea, and Epigeocarpum japonicum sp. nov. Mycol. Prog. 2023, 22, 1–15. [CrossRef]
- Willis, A.; Błaszkowski, J.; Prabhu, T.; Chwat, G.; Góralska, A.; Sashidhar, B.; Harris, P.; D’Souza, J.; Vaingankar, J.; Adholeya, A. Sacculospora felinovii, a novel arbuscular mycorrhizal fungal species (Glomeromycota) from dunes on the west coast of India. Mycol. Prog. 2016, 15, 791-798. [CrossRef]
- Błaszkowski, J.; Kozłowska, A.; Crossay, T.; Symanczik, S.; Al-Yahya’ei, M.N. A new family, Pervetustaceae with a new genus, Pervetustus, and P. simplex sp. nov. (Paraglomerales), and a new genus, Innospora with I. majewskii comb. nov. (Paraglomeraceae) in the Glomeromycotina. Nova Hedwigia 2017, 105, 397-410. [CrossRef]
- Lin, T.C.; Silva, G.A.; Oehl, F. Acaulospora tsugae, a new species in the Glomeromycetes from Taiwan, and a key to species in Acaulosporaceae. Nova Hedwigia 2019, 108, 475–488. [CrossRef]
- Oehl, F.; Souza, F.A.; Sieverding, E. Revision of Scutellospora and description of five new genera and three new families in the arbuscular mycorrhiza-forming Glomeromycetes. Mycotaxon 2008, 106, 311–360.
- Goto, B.T.; Silva, G.A.; Assis, D.M.A.; Silva, D.K.A.; Souza, R.G.; Ferreira, A.C.A.; Jobim, K.; Mello, C.M.A.; Vieira, H.E.E.; Maia, L.C.; Oehl, F. Intraornatosporaceae, (Gigasporales), a new family with two new genera and two new species. Mycotaxon 2012, 119, 117–132. [CrossRef]
- Błaszkowski, J.; Kozłowska, A.; Niezgoda, P.; Goto, B.T.; Dalpé, Y. A new genus, Oehlia with Oehlia diaphana comb. nov. and an emended description of Rhizoglomus vesiculiferum comb. nov. In the Glomeromycotina. Nova Hedwigia 2018, 107, 501–518. [CrossRef]
- Jobim, K.; Błaszkowski, J.; Niezgoda, P.; Kozłowska, A.; Zubek, S.; Mleczko, P.; Chachuła, P.; Ishikawa, N.K.; Goto, B.T. New sporocarpic taxa in the phylum Glomeromycota: Sclerocarpum amazonicum gen. et sp. nov. in the family Glomeraceae (Glomerales) and Diversispora sporocarpia sp. nov. in the Diversisporaceae (Diversisporales). Mycol. Prog. 2019, 18, 369-384. [CrossRef]
- Corazon-Guivin, M.A.; Cerna-Mendoza, A.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Carballar-Hernández, S.; Silva, G.A.; Oehl, F. Nanoglomus plukenetiae, a new fungus from Peru, and a key to small-spored Glomeraceae species, including three new genera in the “Dominikia complex/clades”. Mycol. Prog. 2019, 18, 1395–1409. [CrossRef]
- Corazon-Guivin, M.A.; Mendoza, A.C.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Carballar-Hernández, S.; Silva, G.A.; Oehl, F. Funneliglomus, gen. nov., and Funneliglomus sanmartinensis, a new arbuscular mycorrhizal fungus from the Amazonia region in Peru. Sydowia 2019, 71, 17–24. [CrossRef]
- Corazon-Guivin, M.A.; Mendoza, A.C.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Carballar-Hernández, S.; Silva, G.A.; Oehl, F. Microkamienskia gen. nov. and Microkamienskia peruviana, a new arbuscular mycorrhizal fungus from Western Amazonia. Nova Hedwigia 2019, 109, 355–368. [CrossRef]
- Bhunjun, C.S.; Chen, Y.J.; Phukhamsakda, C.; Boekhout, T.; Groenewald, J.Z.; Mckenzie, E.H.C.; Crous, P. W. et al. What are the 100 most cited fungal genera? Studies in mycology 2024, 108, 1-412. [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinformatics 2019, 20, 1160–1166. [CrossRef]
- Milne, I.; Wright, F.; Rowe, G.; Marshal, D.F.; Husmeier, D.; McGuire, G. TOPALi: Software for automatic identification of recombinant sequences within DNA multiple alignments. Bioinformatics 2004, 20, 1806–1807. [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19: 1572–1574. [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. System Biol. 2003, 52, 696–704. [CrossRef]
- Hall, I.R.; Abbott, L.K. Photographic slide collection illustrating features of the Endogonaceae. 3rd edition. Invery Agricultural Research Centre and Soil Science Department, University of Western Australia, Australia, plus 400 color transparencies, 1979; pp. 1–27.
- Brundrett, M.; Melville, L.; Peterson, L. Practical methods in mycorrhizal research; Mycologue Publications, University of Guelph: Guelph, Canada, 1994.
- Walker, C. Taxonomic concepts in the Endogonaceae: spore wall characteristics in species descriptions. Mycotaxon 1983, 18, 443–455.
- Stürmer, S.L.; Morton, J.B. Developmental patterns defining morphological characters in spores of four species in Glomus. Mycologia 1997, 89, 72-81. [CrossRef]
- Gerdemann, J.W.; Trappe, J.M. The Endogonaceae in the Pacific North West. Mycologia Memoirs 1974, 5, 1-76.
- Trappe, J.M. Glomus segmentatus sp. nov. Trans. Br. Mycol. Soc. 1979, 73, 361-362.
- McGee, P.A.; Trappe, J.M. The Australian zygomycetous mycorrhizal fungi. II. Further Australian sporocarpic Glomaceae. Aust. Syst. Bot. 2002, 15, 115-124. [CrossRef]
- Oehl, F.; Redecker, D.; Sieverding, E. Glomus badium, a new sporocarpic mycorrhizal fungal species from European grasslands with higher soil pH. J. Appl. Bot. Food. Qual. 2005, 79, 38-43.
- Oehl, F.; Sanchez-Castro, I.; Palenzuela, J.; Silva, G.A.; Sieverding, E. Glomus compressum, a new arbuscular mycorrhizal fungus from different agro-ecosystems in central europe. Nova Hedwigia 2014, 99, 429-439. [CrossRef]
- Delavaux, C.S.; Ramos, R.J.; Stürmer, S.L.; Bever, J.D. An updated LSU database and pipeline for environmental DNA identification of arbuscular mycorrhizal fungi. Mycorrhiza 2024, 34, 369–373. [CrossRef]
Figure 1.
Phylogenetic tree obtained by analysis from partial SSU, ITS region, and partial LSU nrDNA sequences of Glomerales. Sequences are labeled with their database accession numbers. Support values from Bayesian inference (BI) and maximum likelihood (ML) are shown just for genera level or above. Only support values of at least 70% are shown. Thick branches represent clades with more than 90% of support in all analyses. The new genus is in bold. The tree was rooted by Entrophospora etunicata. Sequences with only the partial LSU nrDNA are indicated by *.
Figure 1.
Phylogenetic tree obtained by analysis from partial SSU, ITS region, and partial LSU nrDNA sequences of Glomerales. Sequences are labeled with their database accession numbers. Support values from Bayesian inference (BI) and maximum likelihood (ML) are shown just for genera level or above. Only support values of at least 70% are shown. Thick branches represent clades with more than 90% of support in all analyses. The new genus is in bold. The tree was rooted by Entrophospora etunicata. Sequences with only the partial LSU nrDNA are indicated by *.
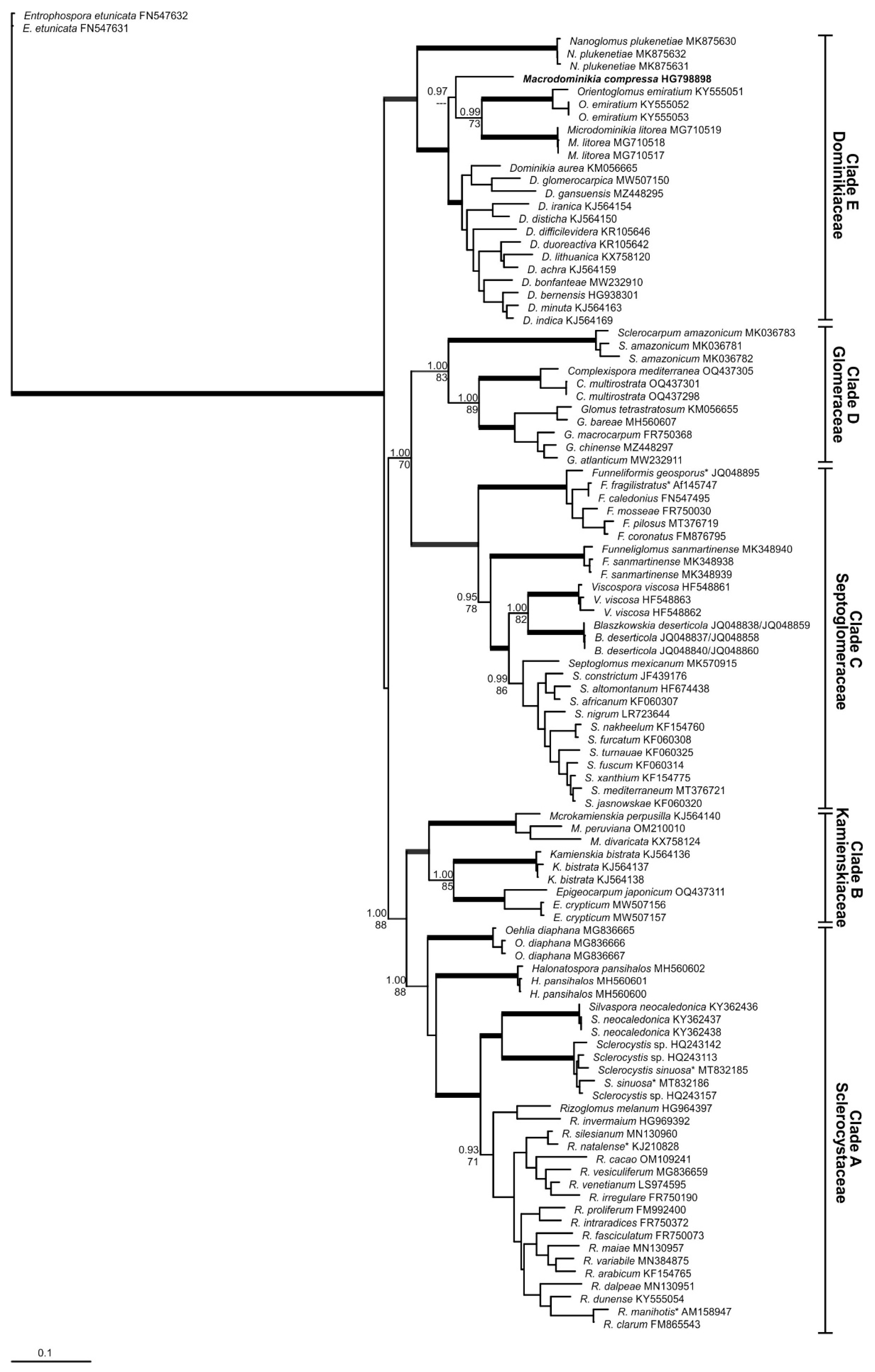
Figure 2.
Phylogenetic tree obtained by analysis from partial SSU, 5.8S, and partial LSU nrDNA sequences of Glomerales. Sequences are labeled with their database accession numbers. Support values from Bayesian inference (BI) and maximum likelihood (ML) are shown just for genera level or above. Only support values of at least 70% are shown. Thick branches represent clades with more than 90% of support in all analyses. The new genus is in bold. The tree was rooted by Entrophospora etunicata. Sequences with only the partial LSU nrDNA are indicated by *.
Figure 2.
Phylogenetic tree obtained by analysis from partial SSU, 5.8S, and partial LSU nrDNA sequences of Glomerales. Sequences are labeled with their database accession numbers. Support values from Bayesian inference (BI) and maximum likelihood (ML) are shown just for genera level or above. Only support values of at least 70% are shown. Thick branches represent clades with more than 90% of support in all analyses. The new genus is in bold. The tree was rooted by Entrophospora etunicata. Sequences with only the partial LSU nrDNA are indicated by *.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



