
Submitted:
22 August 2024
Posted:
25 August 2024
You are already at the latest version
Abstract
Porphyromonas gingivalis is the major pathogenic bacteria found in the subgingival plaque of patients with periodontitis, leading to neuroinflammation. The bacteria destroy periodontal tissue through its virulence factors which are retained in the bacteria’s outer membrane vesicles (OMV). The purpose of this study was to determine the real time effect of intraperitoneal in-jection of P. gingivalis OMV on the production and expression of inflammatory markers as well as histopathological changes in the central nervous system (CNS) of adult zebrafish. Fol-lowing the LD50 (21 µg of OMV), the zebrafish were injected intraperitoneally with 18 µg of OMV and control group with normal saline at 7 different time points. Brains of experimental zebrafish were dissected at desired time points for colorimetric assays, ELISA and histology. This study discovered that nitric oxide and PGE2 were significantly increased at 45 minutes while IL-1β and IL-6 were expressed at subsequent 12- and 24-hours. respectively. Histopatho-logical changes such as blood coagulation, astrocytosis, oedema, spongiosis, and necrosis were also observed between 6- and 24-hours. The two apoptotic enzymes, caspases 3 and 9 were not expressed at any time point. In summary, the OMV induced neuroinflammatory responses and histopathological changes in adult zebrafish were time point dependent
Keywords:
P. gingivalis
; Outer membrane vesicles
; Neuroinflammation
1. Introduction
Periodontitis patients may experience persistent systemic inflammation, leading to neuroinflammation [1] They have higher C-reactive protein levels, higher proportions of periodontal bacteria especially Porphyromonas gingivalis (P. gingivalis), and elevated levels of proinflammatory cytokines and lymphocytes [2]. Elevated blood levels of interferon gamma (IFN-γ) and tumour Necrosis Factor alpha (TNF-α) are also linked to periodontal disease (Alvarenga et al. 2021). Research outcomes imply that distinct immune cell subsets produce distinct cytokines, the presence of particular periodontal bacteria is associated with diverse subsets of immune cells in the peripheral blood of individuals with periodontitis [3] This condition is a likely cause of persistent neuroinflammation since it causes microglial priming, systemic inflammation, and an increase in proinflammatory cytokine levels in the blood [4]
Paramount Periodontal bacteria, P. gingivalis possesses numerous virulence factors that have been implicated in contributing to its pathogenic impact at both local and systemic levels [5]. Similar to the majority of Gram-negative bacteria, P. gingivalis generates outer membrane vesicles (OMVs) that seem to preserve a variety of virulence factors from the original cell, such as LPS, fimbriae, and gingipains [6]. OMVs have the potential to worsen disease conditions by prompting the upregulation of inflammatory molecules [7] For instance, Helicobacter pylori (H. pylori) OMVs stimulate in vitro production of interleukin 6 (IL-6) in human peripheral blood mononuclear cells, while Salmonella spp. OMVs induce TNF and nitric oxide (NO) production by mouse macrophages [8]. Recently, it has also been found that P. gingivalis OMVs can trigger the production of NO and expression of inducible nitric oxide synthase (iNOS) in mouse macrophages [9]
Lately, there’s a surge in the use of zebrafish models to mimic human neurodegenerative disorders [10,11,12] and some physiological conditions that are strongly associated with neurodegeneration such as neuroinflammation, anxiety [13,14,15,16] neuronal redox imbalance [17,18,19] and dementia [11,20,21,22,23]. Based on the link between periodontitis and neurodegenerative/systemic diseases, P. gingivalis and Gram-negative bacteria derivatives such as LPS [17,24,25], gingipain [26,27] and OMVs [28,29] have been used to establish relevant zebrafish disease models.
The objective of this study was to investigate the effect P. gingivalis OMV on the upregulation of neuroinflammatory markers in real time and the relationship between the markers and histopathological alterations in the brain tissue of OMV-treated zebrafish. The outcome of this study may serve as the basis for future research on a better approach to perpetuating histopathological changes beyond 24 hours and the data reported here could also serve as a background for future studies on the mechanism of P. gingivalis OMV- stimulated neuronal injury in adult zebrafish. It may also provide insight on a better route of administering P. gingivalis and its virulence factors.
2. Results
2.1. Extraction of P. gingivalis Outer Membrane Vesicle and Authentication
The volume of OMV recovered after ultracentrifugation was approximately 1.5ml of OMV from 2 liters of P. gingivalis grown to the late log phase:
1.5: 2000 = 0.075%
Western blotting analysis of OMV sample (30 & 60 µg loaded) in which the arginine-specific gingipains (RgpA) was detected. The arginine-specific gingipain, RgpA is a complex protein with approximately 95 kDa to 110 kDa (Wei et al., 2023). This is a confirmation that the derivation technique was correct and the neurotoxin used in the study was actually OMV.
2.2. LD50 of OMV in Adult Zebrafish
Figure 1 shows that there is a relationship between log dose and the probit i.e. as the log dose increases, the probit increases. The LD50 was determined by plotting log dose against the probit. From the equation of the curve, the LD50 is determined by substituting the value of Y at 50% to get the equivalent value of X. From the calculation, the LD50 was found to be 21 µg.
2.3. OMV Triggers Increased NO Level in Zebrafish
In Figure 2 below, the graph depicts the levels of NO in the zebrafish treated with OMV and the corresponding control groups in the time points shown in the graph. There was a significant increase in the level of NO in the OMV zebrafish group at 45 minutes compared with the untreated control at the same time point (p < 0.05). However, from 6 hours till the end of the study, the levels of NO were statistically not different from their corresponding control groups.
2.4. OMV Has No Effect on Caspase 3 and 9 Activity
Figure 3 shows the activities of apoptotic enzymes, caspase 3 (A) and caspase 9 (B) at 0, 6, 12, 24, 48, 72 hour and 7 days in the OMV treated zebrafish and the untreated control groups. When the means of the treated and untreated were compared statistically, it was observed that there was no difference between the caspases 3 and 9 groups and their corresponding control groups at all time points. This suggest that the two apoptotic enzymes did not display any activity throughout the study.
2.5. P. gingivalis OMV Stimulates Production of IL-1β in Zebrafish
Figure 4 shows the concentration of IL-1β in the Y-axis at 7 time points as depicted in the X- axis of the graph. Statistical analysis shows that there was no difference between the OMV treated zebrafish and the control at 45 minutes and 6-hour time points with the P values of 0.84 and 0.828 respectively. Nevertheless, there was significant rise in the concentration of the OMV treated zebrafish against the untreated control at 12 hours (P = 0.0384). Post 12 hours, there was no statistical difference between the concentration of IL-1β in the brains of the treated zebrafish and the untreated groups.
2.6. P. gingivalis OMV Activates Release of IL-6 in Zebrafish
Figure 5 below shows the concentration of IL-6 in OMV induced zebrafish and the corresponding control group injected only normal saline. It was observed that the OMV did not have any effect on the concentration of IL-6 at 0, 6 and 12 hours with P values of 0.975, 0.308, and 0.425 respectively (t-test comparing the treated zebrafish with the control). However, the effect of OMV on the concentration was significantly higher than the untreated control at 24 hours’ time (0.0145). Nonetheless, the effect of OMV on the concentration of IL-6 was not sustained beyond the 24 hours’ time point as the concentration of IL-6 in OMV treated zebrafish and the untreated control were not statistically different from each other at 48 hour, 72 hours and 7 days’ time point.
The values represent the mean ± standard error of the mean (n = 3 per group). The zebrafish that were subjected to OMV treatment exhibited a statistically significant difference compared to the control group, with a p-value of less than 0.05.
2.7. P. gingivalis OMV Triggers High Concentration of PGE2 in Zebrafish
In Figure 6 below, the effect of OMV on the concentration of PGE2 was noticeably high at the early stage of the study with the zebrafish induced with OMV having higher concentration of PGE2 than the control at 0 hour time point. Nevertheless, the other time points showed that the concentration ofPGE2 in OMV induced zebrafish and the control was not significantly different from each other.
2.8. OMV Induced Histopathological Changes in the Brain of Zebrafish
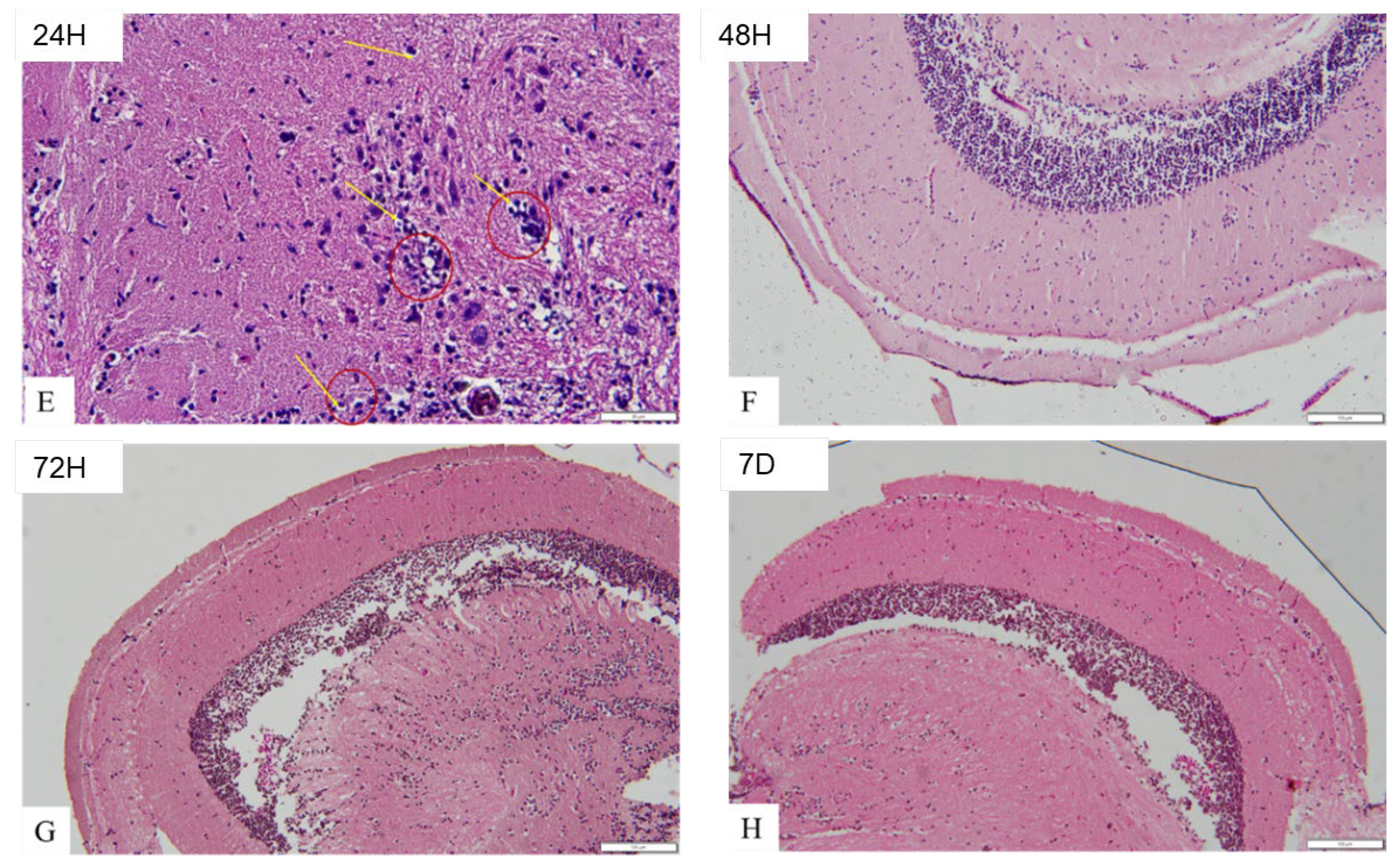
Brain histology with haematoxylin and eosin staining as shown in Figure 7a revealed control (A) and the OMV treated zebrafish at 45 minutes time point (B), the optic tectum with well organised cells and there was no congestion of blood vessels. The astrocytes are not more than usual. In (A) and (B), normal architecture of the brain was retained. However, the OMV treated zebrafish brain at 6 hours’ time points (C), apart from congestion of the blood vessels that are very obvious (black arrow), other cells and the architecture of the brain remain normal and undistorted and are similar to the control group. Nevertheless, in OMV treated zebrafish at 12 hours’ time point (D), in addition to congestion of blood vessels (black arrow), the astrocytes are more than usual (dark red circle) i.e. astrocytosis is beginning to set in. Similarly at 24 hours’ time point (E) in Figure 7b, the architecture of the tectum is beginning to change, the astrocytes and glial cells are becoming active and there are signs of oedema (yellow arrow), spongiosis as well as necrosis. Post 24 hours i.e. from 48 hours till the end of the study at 7 days, the normal architecture of the brain was observed, the cells are not crowded and there was no congestion of blood vessels. The architecture of the brain and the arrangement of the cells at 48-hour, 72 hour and 7 days’ time point were not different from the control groups.
3. Discussion
The brain's innate immune response can result in neuroinflammation, which can manifest in two ways: 1) acutely, when inflammatory mediators are transiently expressed, and 2) chronically, when the resolution of inflammation takes a long time. The latter phase, which gradually contributes to the loss of neuronal cells, is characterized by persistent, low-grade inflammation brought on by glial cells' proinflammatory cytokine release. Unresolved systemic inflammatory stimulation results in a condition where hypersensitive, continuously active microglia react aggressively to new immunological triggers [30,31] Research have shown that the brain experiences a proinflammatory response when LPS activates TLR4 in astrocytes and microglia [23,32].
A single intraperitoneal injection of 18 µg of P. gingivalis OMV triggered an inflammatory response in the brain of adult zebrafish from 45 minutes time point up till 24 hours’ time point. While the levels of nitric oxide (NO) and prostaglandin E2 (PGE2) were significantly high at 45 minutes time points, the histopathological alterations due to OMV treatment was apparent between 6-hour time point and 24-hour time point. However, IL-1β and IL-6 which are the two inflammatory cytokines were expressed at 12- and 24-hour time points respectively. The activity of astrocytes and glial cells between 12- and 24-hours’ time points which culminated in astrocytosis, spongiosis, oedema and necrosis are pointer to the fact that there is a strong relationship between these histopathological changes and the expression of inflammatory cytokines. We have demonstrated here that OMV-induced neuroinflammation was a series of physiological events that are time point dependent in which astrocytes seemed to have played a central role by altering the inflammatory process before 48 hours’ time point. The relationship between histopathological alterations and the upregulation of inflammatory markers shown in this study are supported by the outcomes of some identical studies.
For instance, a rodent model that simulate cerebral venous congestion resulting from arteriovenous anastomosis also demonstrated symptoms of cognitive impairment [33,34]. In addition, there is substantial evidence supporting the association between mice experiencing cerebral venous congestion, disruption of blood brain barrier (BBB), increased extravasation of IgG and potentially other plasma components, and an exacerbated neuroinflammatory response [35,36] This is evident through the increased presence of activated microglia and upregulation of inflammatory mediators within the hippocampus. It has also been observed that pro-inflammatory cytokines, chemokines, proteases, and reactive oxygen species produced by activated microglia contribute to neuronal dysfunction [36].
Several research efforts focused on re-evaluation of the importance of inflammation in neurodegeneration are ongoing and neuroinflammation is being recognized as a key factor in the development of neurodegenerative disease [4]. Despite the neurodegenerative diseases including Alzheimer’s disease (AD), Parkison’s disease, multiple sclerosis, Huntington’s disease having different pathogenic mechanism such as genetic differences and varying protein aggregates, the common denominator among them is chronic neuroinflammation. The endogenous pathologic protein aggregation in each of the disorders is capable of triggering neuroinflammation thereby prompting neurodegeneration [37].
The P. gingivalis OMV was adopted for this study because OMV contains gingipains, LPS and DNA that have all been detected in AD patients [38]. P. gingivalis OMV has a crucial advantage over the P. gingivalis cells because of their ability to enter the brain[39]. This attribute is reliant on three critical factors. Firstly, P. gingivalis OMV have numerical advantage over the P. gingivalis cells [38]. Secondly, they are smaller with a diameter of 80 nm compared with P. gingivalis diameter of 600 nm (Gong et al. 2022). Thirdly, P. gingivalis OMV contains gingipain and LPS which have been reported to cause AD-like pathological changes [40].
Outer membrane vesicles (OMVs) derived from P. gingivalis have been shown to possess the ability to transmit the required signals for initiating and activating the inflammasome, a condition that could guarantee a powerful activation of pyroptosis in macrophages. The increased levels of gingipains identified in OMVs compared to their parent cells could be a possible explanation for this discovery [41]. Furthermore, the enzymatic activity of gingipains has been recognized as a catalyst for the activation of inflammasomes and the triggering of cell death in different types of cells (Fleetwood et al. 2017; Xu et al. 2020). In contrast to their parent cells, OMVs [44] lack nucleoside-diphosphate kinase (NDK), which means they cannot rely on NDK suppression to prevent ATP/ROS-mediated inflammasome activation [9].
Bacterial OMVs transport lipopolysaccharide (LPS) to the cytosol, leading to the activation of inflammasomes and pyroptosis in macrophages, as demonstrated by Vanaja et al. in 2016 [45]Therefore, it is reasonable to suggest that the high concentration of gingipains in outer membrane vesicles (OMVs), together with their ability to access the cytosolic inflammasome complex, is responsible for their notable capacity to trigger pyroptotic cell death in macrophages [43]. The activation of pyroptotic cell death by P. gingivalis OMVs (as well as other pathogens like A. actinomycetemcomitans) could help explain the increased levels of lactate dehydrogenase (LDH) detected in the saliva of individuals with periodontitis [46].
Regarding the apoptotic enzymes (caspase 9 and caspase 3), it was observed that they did not show activity at any of the time points. All of the wild-type strains of P. gingivalis, namely A7A1-28, ATCC 49417 (the strain used for this study) and W83, exhibited the ability to hinder the activation of caspase-3 induced by chemical means. The strains that impede apoptosis have a unique characteristic. They don’t produce the long (FimA) and short (Mfa) fimbriae [47,48]. Interestingly, these fimbrae play crucial roles in causing apoptotic cell death in the host cells by inactivating caspase 3 [49]. Since our strain belongs to the FimA and Mfa non-producing strain, an inference can be drawn that our OMV lacked these two fimbra. However, a study reported the capacity of P. gingivalis to inhibit the activation of caspase 9 [50,51]. Although both caspases 3 and 9 are apoptotic caspases, other apoptotic caspases depend on caspase 9 for their activity. In other words, they cannot be activated if caspase 9 was not activated. So, caspase 9 is referred to as initiator apoptotic enzyme while caspases 3 and 6 are tagged executioner enzymes [52].
The cascade of events in this study shows that OMV induced neuroinflammation in adult zebrafish is a process involving a number of inflammatory mediators which are time point dependent. While PGE2 and NO production were significant at 45 minutes time points, blood coagulation was the only event observed at 6 hours’ time point. However, there were obvious relationship at 12 hour and 24-hour time point between expression of inflammatory cytokines and histopathological alterations such as blood coagulation, astrocytosis, necrosis, oedema and spongiosis.
4. Materials and Methods
4.1 Porphyromonas Gingivalis Derived Outer Membrane Vesicles (P. gingivalis OMV) Preparation
P. gingivalis OMV derivation protocol was according to Ho et al., (2015) with some modifications. P. gingivalis was grown for between 5 to 7 days to reach the late log phase. Thereafter the culture was subjected to low and high-speed centrifugation which was followed by the harvesting of the OMV pellet. The OMV sample was subjected to western blotting analysis by targeting gingipain R1 (RgpA) [53]
Following the utilization of the Bradford reagent to quantify the protein content in the sample, the total protein (30 and 50 ug) was segregated using sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) utilizing the subsequent methodology. The separated proteins were transferred to PVDF membrane by sandwiching the gel and the membrane between sponge and filter papers in the following order (sponge – filter paper – gel – membrane – filter paper – sponge) starting from the black side of the gel holder cassette to the clear side. The cassette was placed in the electrode assembly and electric current was made to run through the cassette. After successful transfer of proteins to the membrane, they were treated with Rabbit anti-P. gingivalis RgpA Polyclonal antibody (Cusabio, Houston, Texas USA) at a concentration of 1:1,000, rinsed five times, and then incubated with horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G secondary antibodies (Gene Tex, USA) at a concentration of 1:2,000. The samples underwent six cycles of washing in PBST, and the blots were visualized using the Chemidoc™ xrs system.
4.2. Fish Husbandry and Care
All procedures were carried out at the Zebrafish Laboratory, Faculty of Medicine, Universiti Kebangsaan Malaysia in compliance with ethical approval by the Animal Ethics Committee of Universiti Kebangsaan Malaysia (FPG/2021/NURRUL SHAQINAH/28-JULY/1189-JULY-2021-JUNE-2024). The fish were housed in 6L 3-dimentional transparent acrylic aquaria tank with a 3-dimentional shape (L,27cm X W, 12cm X H, 15cm) connected to an automated circulating aquatic system. The fish were stocked at a density of 5 fish per liter and acclimatized for at least 7 days before any experimental procedure. All fish will be fasted 24h prior to the experimental procedure. The fish were maintained at 27°C ± 0.5°C and a 14:10 hour light-dark cycle. A twice- daily feeding arrangement (during and pre-experimental period) was practiced with a commercial flaked food (New Life Spectrum, Thera + A, USA), throughout the experimental duration.
4.3. Intraperitoneal Injection of P. gingivalis Derived Outer Membrane Vesicles (OMV)
Fully acclimatized fish were selected randomly and assigned to control and treatment groups. Fish were anesthetized by transferring them one at a time to a beaker containing 0.0035% of Benzocaine with the aid of a hand net . Immediately the fish slows the movement of operculum with no response to touch, the anesthetized fish was moved to an improvised surgical bed that has been slit in the middle. The fish is positioned belly up followed by a careful insertion of the needle between the pelvic fins. The OMV group received 18 µg of OMV while the control group received normal saline. The injections were administered using a micro syringe (Hamilton Gastight syringe, 1700 series, Luer tip, Hamilton company, Nevada, USA). After injection, the fish was transferred to a beaker containing one-third of system water and placed in the designated group accordingly. However, sometimes water needed to be swirled towards the gills to hasten recovery or assist struggling fish attain full recovery[54,55]
4.4. Median Lethal Dose Determination
After a week of acclimatization including 24h of fasting, 160 of zebrafish was divided equally into eight groups to receive different doses of OMV (40, 30, 20, 15, 10, 5, 2.5, 1.0 µg). The fish have been anesthetized in a beaker containing 0.0035% benzocaine, and once they showed reduced motor function, each fish was injected respective OMV doses intraperitoneally (using 30G syringe with a Hamilton micro syringe) . Post OMV intraperitoneal injection, zebrafish were put in a beaker with one-third of the system water for recovery from anaesthesia. Fresh system water is swirled to the gills of the fish until the fish starts swimming freely up and down the beaker [54]. Fish that failed to recover fully from anaesthesia were not selected for the experiment. The number of deaths in each group was monitored for 7 days. The health of experimental fish was maintained by keeping them in freshly cleaned tanks containing system water every 2 days or immediately after an incidence of mortality in a tank. The LD50 was determined using the Miller and Tainter methods (a simple regression method in which the probit is plotted against the log dose) [56].
4.5. Determination of Levels of Nitric Oxide (NO), Caspase 3 and 9 Using Colorimetric Assay
A total of 252 zebrafish was selected for nitric oxide (NO), Caspase 3 and Caspase 9 measurement. Each OMV group and the control group received 18 µg of OMV and 10 µl of normal saline via intraperitoneal route respectively as described before. Experimental fish were euthanized by immersing them in ice water at designated time point (45 minutes, 6, 12, 24, 48, 72 hours and 7 days)., which was followed by brain dissection and homogenization according to kit manufacturer’s instruction, (Elabscience, Houston, Texas, USA).
4.6. Determination of Concentrations of Interleukin 6 (IL-6), Interleukin 1 Beta (IL-1β) and Prostaglandin E2 (PGE2) Using ELISA
In experimental procedures using 126 adult zebrafish, concentrations of IL-6, IL-1β and PGE2 were estimated from the homogenate of brain tissues of zebrafish induced with OMV and the uninduced control group at all the 7 time points (45 minutes, 6, 12, 24, 48, 72 hours and 7 days) using the ELISA protocol according to the manufacturer’s instructions (ELK Biotechnology Denver, USA).
4.7. Evaluation of Zebrafish Brain Histology
After the expiration of each time point, the whole of zebrafish head was dissected for brain histological examination. (The head was fixed in 10% formalin solution for 48 hours. The tissue samples were processed and stained with routine H&E protocol. The histology images were analysed using a light microscope (Olympus, Center Valley, Pennsylvania, USA).
4.8. Statistical Analysis
The LD50 determination was evaluated using the Miller and Tainter method. The other results were presented as mean ± standard deviation (SD) of each experimental group, determined using unpaired t-test. The results were selected from at least three independent experiments carried out in triplicate. P-values of 0.05 or less were statistically significant.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.A, N.S.N. and S.L.T.; methodology, M.A, H.M and R.A; validation, M.N.A.A, N.A.B, N.S.N.; investigation, M.A, H.M and R.A.; resources, S.L.T..; writing—original draft preparation, M.A; writing—review and editing, M.N.A.A, S.L.T, N.A.B and N.S.N.; supervision, M.N.A.A, S.L.T, N.A.B and N.S.N.; ; project administration, S.L.T and N.S.N.; funding acquisition, N.S.N All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Fundamental Research Grant Scheme, Ministry of Education; FRGS/1/2020/SKK0/UKM/02/10.
Institutional Review Board Statement
The animal study was approved by the Universiti Kebangsaan Malaysia Animal Ethics Committee (Reference: FPG/2021/NURRUL SHAQINAH/28-JULY/1189-JULY-2021-JUNE-2024). The study was conducted in compliance with local legislation and institutional requirements.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable
Acknowledgments
This study was financially supported by the Malaysian Government Fundamental Research Grant Scheme (FRGS) (FRGS/1/2020/SKK0/UKM/02/10).
Conflicts of Interest
The authors declared no conflict of interest.
References
- Alvarenga MOP, Frazão DR, Matos IG de, et al. Is There Any Association Between Neurodegenerative Diseases and Periodontitis? A Systematic Review. Frontiers in Aging Neuroscience; 13. Epub ahead of print 24 May 2021. [CrossRef]
- Sirisereephap K, Maekawa T, Tamura H, et al. Osteoimmunology in Periodontitis: Local Proteins and Compounds to Alleviate Periodontitis. International Journal of Molecular Sciences; 23. Epub ahead of print 1 May 2022. [CrossRef]
- Teixeira FB, Saito MT, Matheus FC, et al. Periodontitis and alzheimer’s disease: A possible comorbidity between oral chronic inflammatory condition and neuroinflammation. Front Aging Neurosci; 9. Epub ahead of print 10 October 2017. [CrossRef]
- Webers A, Heneka MT, Gleeson PA. The role of innate immune responses and neuroinflammation in amyloid accumulation and progression of Alzheimer’s disease. Immunology and Cell Biology 2020; 98: 28–41.
- Gare J, Kanoute A, Meda N, et al. Zperiodontal conditions and pathogens associated with pre-eclampsia: A scoping review. International Journal of Environmental Research and Public Health; 18. Epub ahead of print 1 July 2021. [CrossRef]
- Jia Y, Guo B, Yang W, et al. Rho kinase mediates Porphyromonas gingivalis outer membrane vesicle-induced suppression of endothelial nitric oxide synthase through ERK1/2 and p38 MAPK. Arch Oral Biol 2015; 60: 488–495.
- Nakao R, Hasegawa H, Ochiai K, et al. Outer membrane vesicles of porphyromonas gingivalis elicit a mucosal immune response. PLoS One; 6. Epub ahead of print 2011. [CrossRef]
- Avila-Calderón ED, Ruiz-Palma M del S, Aguilera-Arreola MG, et al. Outer Membrane Vesicles of Gram-Negative Bacteria: An Outlook on Biogenesis. Frontiers in Microbiology; 12. Epub ahead of print 4 March 2021. [CrossRef]
- Gong T, Chen Q, Mao H, et al. Outer membrane vesicles of Porphyromonas gingivalis trigger NLRP3 inflammasome and induce neuroinflammation, tau phosphorylation, and memory dysfunction in mice. Front Cell Infect Microbiol; 12. Epub ahead of print 9 August 2022. [CrossRef]
- Omar NA, Kumar J, Teoh SL. Neuroprotective effects of Neurotrophin-3 in MPTP-induced zebrafish Parkinson’s disease model. Front Pharmacol; 14. Epub ahead of print 28 November 2023. [CrossRef]
- Gao X, Zhang P, Chen J, et al. Necrostatin-1 Relieves Learning and Memory Deficits in a Zebrafish Model of Alzheimer’s Disease Induced by Aluminum. Neurotoxicity Research 2022 40:1 2022; 40: 198–214.
- Koehler D, Shah ZA, Hensley K, et al. Lanthionine ketimine-5-ethyl ester provides neuroprotection in a zebrafish model of okadaic acid-induced Alzheimer’s disease. Neurochem Int 2018; 115: 61–68.
- Farías-Cea A, Leal C, Hödar-Salazar M, et al. Behavioral Study of 3- and 5-Halocytisine Derivatives in Zebrafish Using the Novel Tank Diving Test (NTT). Int J Mol Sci; 24. Epub ahead of print 1 July 2023. [CrossRef]
- Fontana BD, Alnassar N, Parker MO. The zebrafish (Danio rerio) anxiety test battery: comparison of behavioral responses in the novel tank diving and light–dark tasks following exposure to anxiogenic and anxiolytic compounds. Psychopharmacology (Berl) 2022; 239: 287–296.
- Aparna S, Patri M. Benzo[a]pyrene exposure and overcrowding stress impacts anxiety-like behavior and impairs learning and memory in adult zebrafish, Danio rerio. Environ Toxicol 2021; 36: 352–361.
- Abu Bakar N, Mohd Sata NSA, Ramlan NF, et al. Evaluation of the neurotoxic effects of chronic embryonic exposure with inorganic mercury on motor and anxiety-like responses in zebrafish (Danio rerio) larvae. Neurotoxicol Teratol 2017; 59: 53–61.
- Singh S, Sahu K, Kapil L, et al. Quercetin ameliorates lipopolysaccharide-induced neuroinflammation and oxidative stress in adult zebrafish. Molecular Biology Reports 2022 49:4 2022; 49: 3247–3258.
- Singh S, Sahu K, Kapil L, et al. Administration of Quercetin Ameliorates Lipopolysaccharide Induced Neuroinammation and Oxidative Stress in Adult Zebrash. Epub ahead of print 2021. [CrossRef]
- Valcarce DG, Martínez-Vázquez JM, Riesco MF, et al. Probiotics reduce anxiety-related behavior in zebrafish. Heliyon; 6. Epub ahead of print 1 May 2020. [CrossRef]
- Kaur K, Narang RK, Singh S. AlCl3 induced learning and memory deficit in zebrafish. Neurotoxicology 2022; 92: 67–76.
- Zanandrea R, Abreu MS, Piato A, et al. Lithium prevents scopolamine-induced memory impairment in zebrafish. Neurosci Lett 2018; 664: 34–37.
- Singsai K, Ladpala N, Dangja N, et al. Effect of Streblus asper Leaf Extract on Scopolamine-Induced Memory Deficits in Zebrafish: The Model of Alzheimer’s Disease. Adv Pharmacol Pharm Sci; 2021. Epub ahead of print 2021. [CrossRef]
- Suganya K, Koo BS. Gut–brain axis: Role of gut microbiota on neurological disorders and how probiotics/prebiotics beneficially modulate microbial and immune pathways to improve brain functions. International Journal of Molecular Sciences 2020; 21: 1–29.
- Mottaz H, Schönenberger R, Fischer S, et al. Dose-dependent effects of morphine on lipopolysaccharide (LPS)-induced inflammation, and involvement of multixenobiotic resistance (MXR) transporters in LPS efflux in teleost fish. Environmental Pollution 2017; 221: 105–115.
- Zhang Q, Kopp M, Babiak I, et al. Low incubation temperature during early development negatively affects survival and related innate immune processes in zebrafish larvae exposed to lipopolysaccharide. Sci Rep; 8. Epub ahead of print 1 December 2018. [CrossRef]
- Widziolek M, Prajsnar TK, Tazzyman S, et al. Zebrafish as a new model to study effects of periodontal pathogens on cardiovascular diseases. Sci Rep; 6. Epub ahead of print 25 October 2016. [CrossRef]
- Farrugia C, Stafford GP, Potempa J, et al. Mechanisms of vascular damage by systemic dissemination of the oral pathogen Porphyromonas gingivalis. FEBS Journal 2021; 288: 1479–1495.
- Farrugia C, Stafford GP, Murdoch C. Porphyromonas gingivalis Outer Membrane Vesicles Increase Vascular Permeability. J Dent Res 2020; 99: 1494–1501.
- Guo J, Lin K, Wang S, et al. Effects and mechanisms of Porphyromonas gingivalis outer membrane vesicles induced cardiovascular injury. BMC Oral Health; 24. Epub ahead of print 1 December 2024. [CrossRef]
- Perry VH, Holmes C. Microglial priming in neurodegenerative disease. Nature Reviews Neurology 2014; 10: 217–224.
- Hampel H, Hardy J, Blennow K, et al. The Amyloid-β Pathway in Alzheimer’s Disease. Molecular Psychiatry 2021; 26: 5481–5503.
- Chen Z, Jalabi W, Shpargel KB, et al. Lipopolysaccharide-induced microglial activation and neuroprotection against experimental brain injury is independent of hematogenous TLR4. Journal of Neuroscience 2012; 32: 11706–11715.
- Hai J, Wan JF, Lin Q, et al. Cognitive dysfunction induced by chronic cerebral hypoperfusion in a rat model associated with arteriovenous malformations. Brain Res 2009; 1301: 80–88.
- Zhang Y, Fang Z, Li R, et al. Design of outer membrane vesicles as cancer vaccines: A new toolkit for cancer therapy. Cancers; 11. Epub ahead of print 1 September 2019. [CrossRef]
- Adamski MG, Sternak M, Mohaissen T, et al. Vascular cognitive impairment linked to brain endothelium inflammation in early stages of heart failure in mice. J Am Heart Assoc; 7. Epub ahead of print 1 April 2018. [CrossRef]
- Fulop GA, Ahire C, Csipo T, et al. Cerebral venous congestion promotes blood-brain barrier disruption and neuroinflammation, impairing cognitive function in mice. Geroscience 2019; 41: 575–589.
- Zhang W, Xiao D, Mao Q, et al. Role of neuroinflammation in neurodegeneration development. Signal Transduction and Targeted Therapy; 8. Epub ahead of print 1 December 2023. [CrossRef]
- Toyofuku M, Nomura N, Eberl L. Types and origins of bacterial membrane vesicles. Nature Reviews Microbiology 2019; 17: 13–24.
- Yang WW, Guo B, Jia WY, et al. Porphyromonas gingivalis-derived outer membrane vesicles promote calcification of vascular smooth muscle cells through ERK1/2-RUNX2. FEBS Open Bio 2016; 6: 1310–1319.
- Wei S, Peng W, Mai Y, et al. Outer membrane vesicles enhance tau phosphorylation and contribute to cognitive impairment. J Cell Physiol 2020; 235: 4843–4855.
- Mantri CK, Chen CH, Dong X, et al. Fimbriae-mediated outer membrane vesicle production and invasion of Porphyromonas gingivalis. Microbiologyopen 2015; 4: 53–65.
- Xu W, Zhou W, Wang H, et al. Roles of Porphyromonas gingivalis and its virulence factors in periodontitis. Adv Protein Chem Struct Biol 2020; 120: 45–84.
- Fleetwood AJ, Lee MKS, Singleton W, et al. Metabolic remodeling, inflammasome activation, and pyroptosis in macrophages stimulated by Porphyromonas gingivalis and its outer membrane vesicles. Front Cell Infect Microbiol; 7. Epub ahead of print 4 August 2017. [CrossRef]
- Johnson L, Atanasova KR, Bui PQ, et al. Porphyromonas gingivalis attenuates ATP-mediated inflammasome activation and HMGB1 release through expression of a nucleoside-diphosphate kinase. Microbes Infect 2015; 17: 369–377.
- Vanaja SK, Russo AJ, Behl B, et al. Bacterial Outer Membrane Vesicles Mediate Cytosolic Localization of LPS and Caspase-11 Activation. Cell 2016; 165: 1106–1119.
- Lu F, Lan Z, Xin Z, et al. Emerging insights into molecular mechanisms underlying pyroptosis and functions of inflammasomes in diseases. Journal of Cellular Physiology 2020; 235: 3207–3221.
- Ma L, Liu H, Wang X, et al. CXXC5 orchestrates Stat3/Erk/Akt signaling networks to modulate P. gingivalis-elicited autophagy in cementoblasts. Biochim Biophys Acta Mol Cell Res; 1868. Epub ahead of print 1 March 2021. [CrossRef]
- Noiri Y, Li L, Yoshimura F, et al. Localization of Porphyromonas gingivalis-carrying fimbriae in situ in human periodontal pockets. J Dent Res 2004; 83: 941–945.
- Chopra A, Radhakrishnan R, Sharma M. Porphyromonas gingivalis and adverse pregnancy outcomes: a review on its intricate pathogenic mechanisms. Critical Reviews in Microbiology 2020; 46: 213–236.
- Lamont RJ, Fitzsimonds ZR, Wang H, et al. Role of Porphyromonas gingivalis in oral and orodigestive squamous cell carcinoma. Periodontology 2000 2022; 89: 154–165.
- Yao L, Jermanus C, Barbetta B, et al. Porphyromonas gingivalis infection sequesters pro-apoptotic Bad through Akt n primary gingival epithelial cells. Mol Oral Microbiol 2010; 25: 89–101.
- Hegazy RR, Mansour DF, Salama AA, et al. Regulation of PKB/Akt-pathway in the chemopreventive effect of lactoferrin against diethylnitrosamine-induced hepatocarcinogenesis in rats. Pharmacological Reports 2019; 71: 879–891.
- Ho MH, Chen CH, Goodwin JS, et al. Functional advantages of Porphyromonas gingivalis vesicles. PLoS One; 10. Epub ahead of print 21 April 2015. [CrossRef]
- Omar NA, Kumar J, Teoh SL. Parkinson’s disease model in zebrafish using intraperitoneal MPTP injection. Front Neurosci; 17. Epub ahead of print 2023. [CrossRef]
- Kinkel MD, Eames SC, Philipson LH, et al. Intraperitoneal injection into adult zebrafish. Journal of Visualized Experiments. Epub ahead of print 2010. [CrossRef]
- Chandra M, Raj J, Das Dogra T, et al. Determination of median lethal dose of triazophos with DMSO in wistar rats. Article in Asian Journal of Pharmaceutical and Clinical Research; 6. https://www.researchgate.net/publication/265378795 (2014).
Figure 1.
Log10 dose versus the probits. The probits value is the standardized conversion value derived from the corrected mortality percentage as adapted from Finney’s table.
Figure 1.
Log10 dose versus the probits. The probits value is the standardized conversion value derived from the corrected mortality percentage as adapted from Finney’s table.
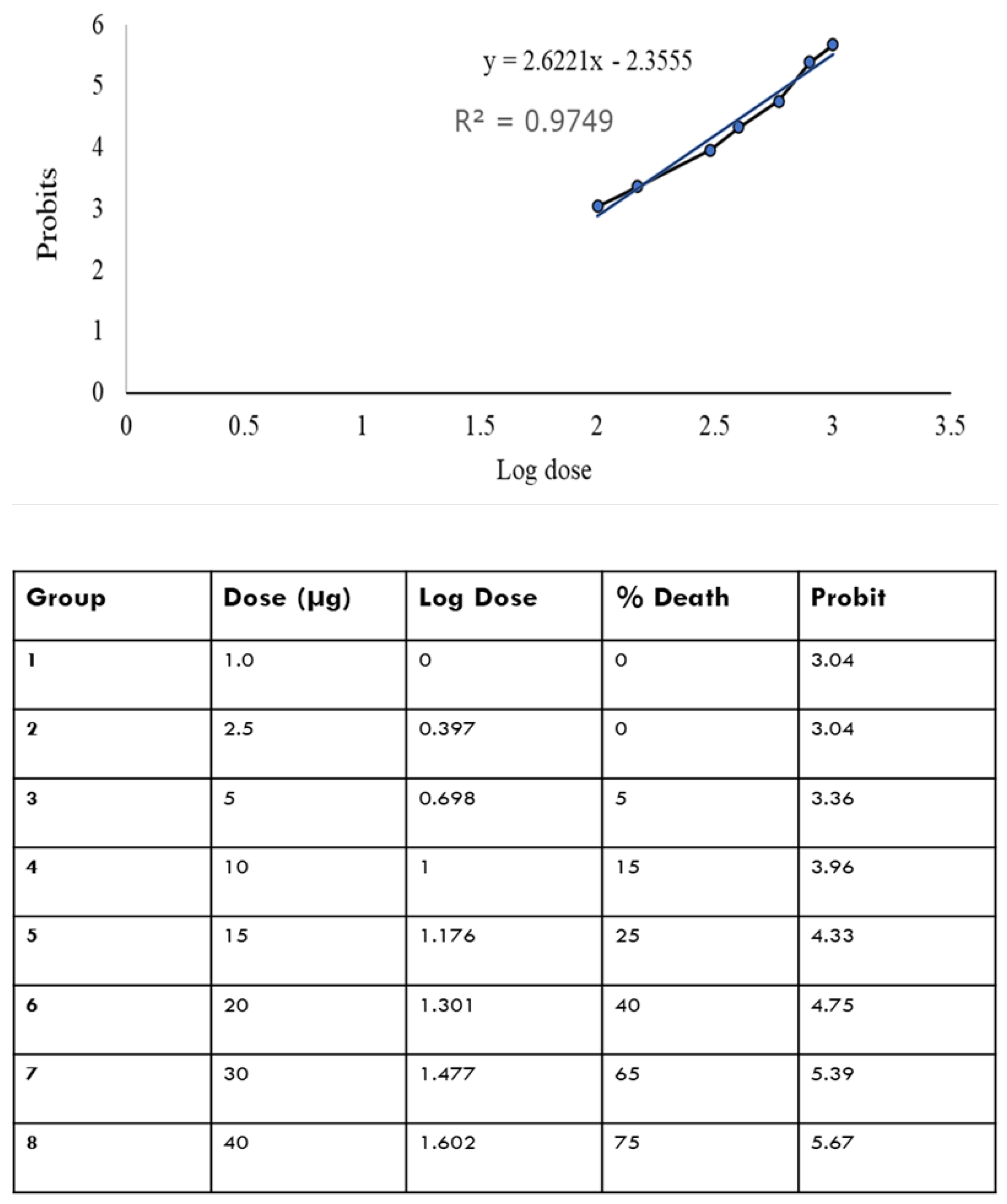
Figure 2.
The level of NO in the brains of OMV- treated and the control group of zebrafish. *** this symbol represents OMV treated zebrafish group was significantly different from the control group at p < 0.001.
Figure 2.
The level of NO in the brains of OMV- treated and the control group of zebrafish. *** this symbol represents OMV treated zebrafish group was significantly different from the control group at p < 0.001.
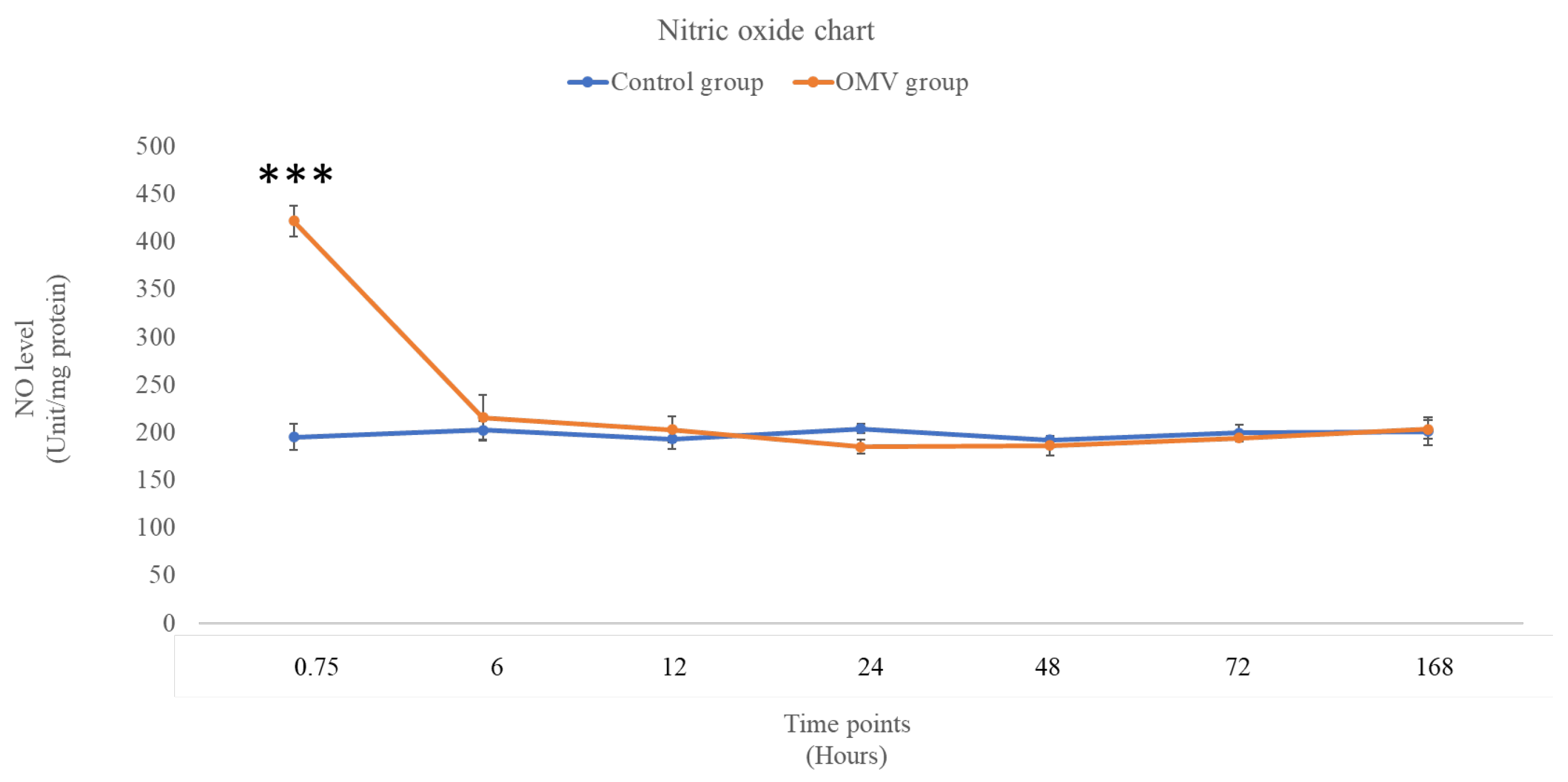
Figure 3.
(A). The activity of caspase 3 in the brains of OMV- treated group and the control group of zebrafish. (B). The activity of caspase 9 in the brains of OMV- treated and the control group of zebrafish.Values are mean ± SEM (n = 6/group). OMV treated zebrafish was not significantly different from the control group at p < 0.05.
Figure 3.
(A). The activity of caspase 3 in the brains of OMV- treated group and the control group of zebrafish. (B). The activity of caspase 9 in the brains of OMV- treated and the control group of zebrafish.Values are mean ± SEM (n = 6/group). OMV treated zebrafish was not significantly different from the control group at p < 0.05.
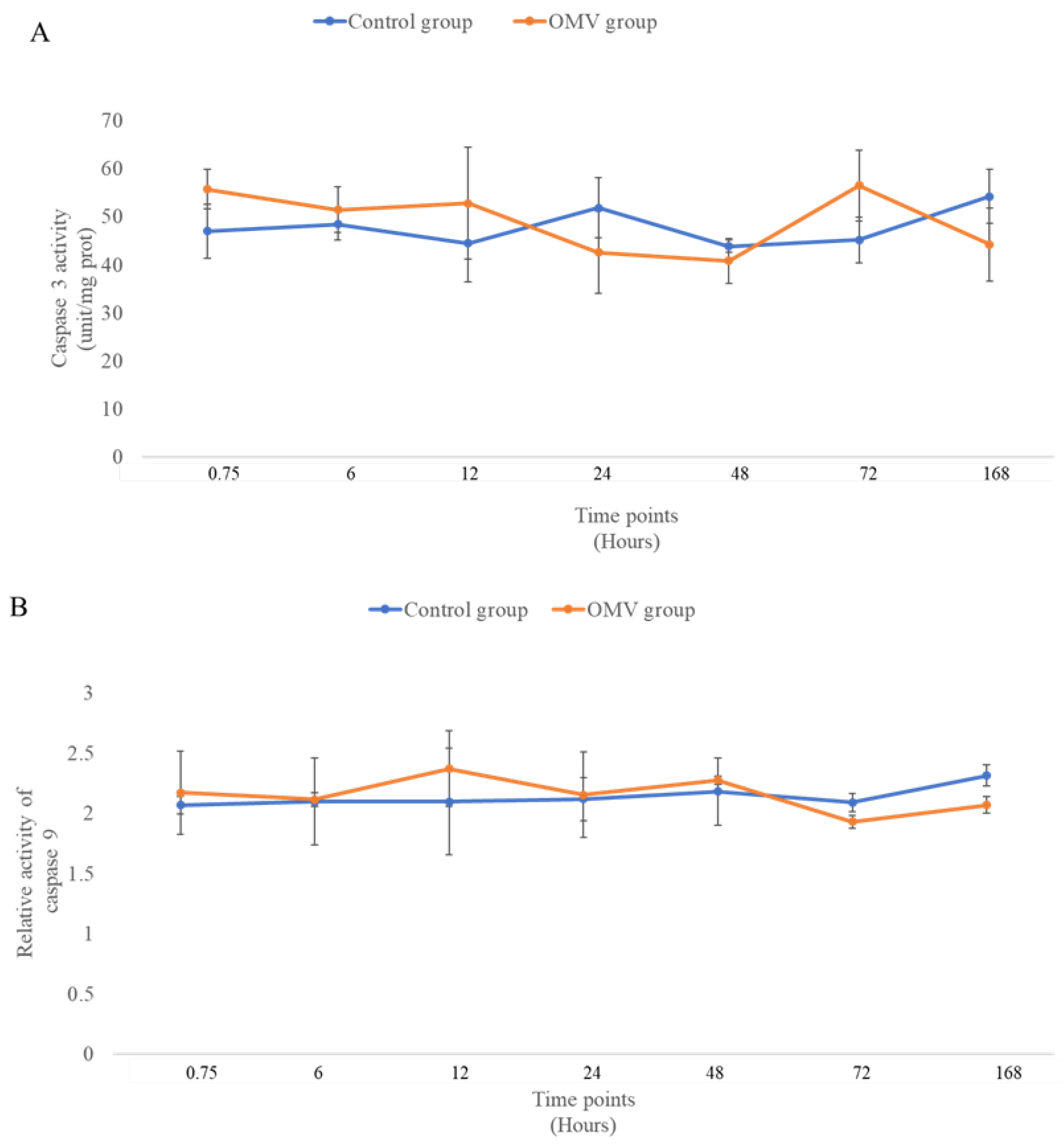
Figure 4.
The concentration of IL-1β in the brains of OMV treated zebrafish and the control group of zebrafish.Values are mean ± SEM (n = 3/group). * OMV treated zebrafish was significantly different from the control group at p < 0.05.
Figure 4.
The concentration of IL-1β in the brains of OMV treated zebrafish and the control group of zebrafish.Values are mean ± SEM (n = 3/group). * OMV treated zebrafish was significantly different from the control group at p < 0.05.
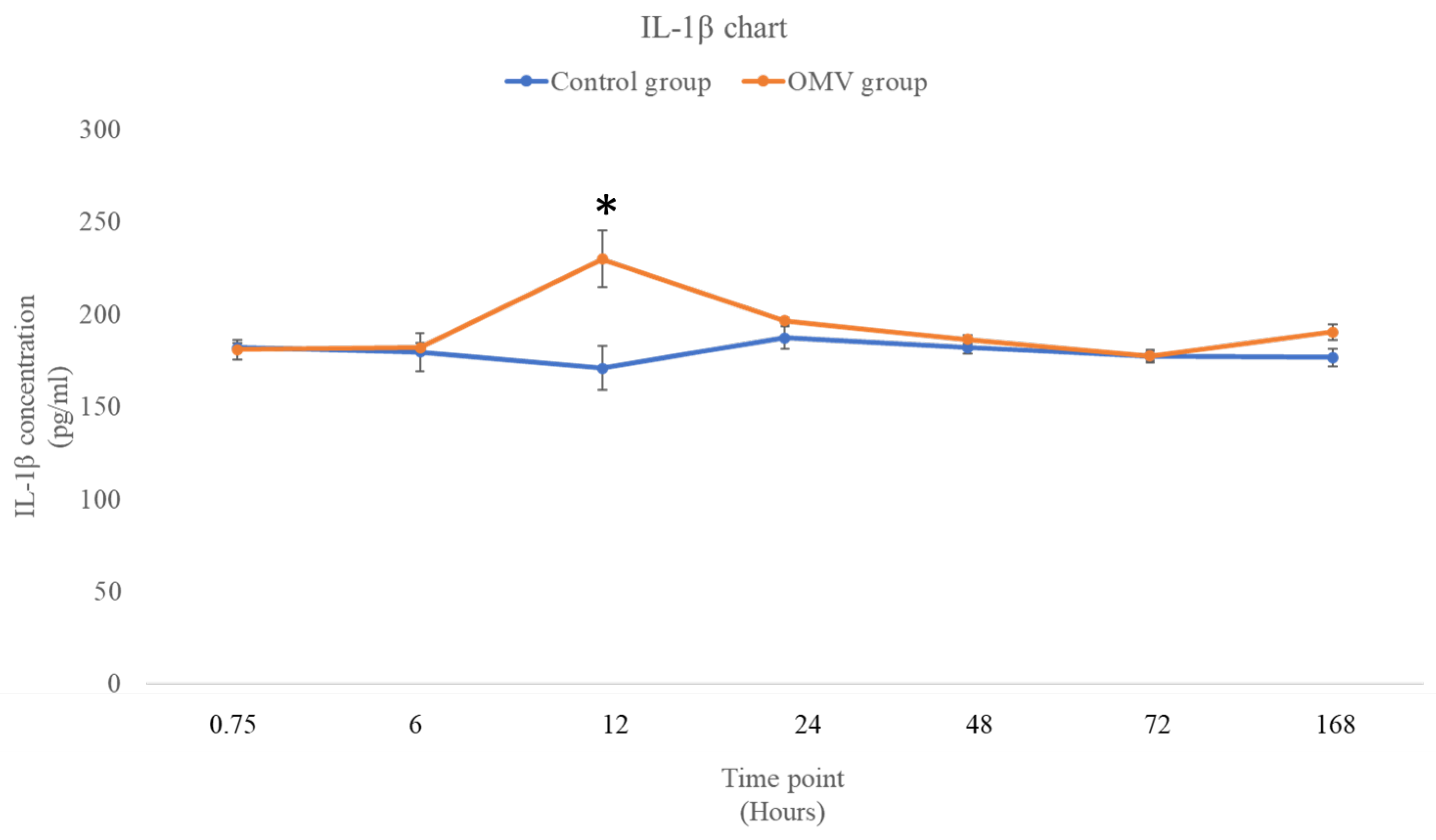
Figure 5.
The concentration of IL-6 in the brains of OMV-treated and the control group of zebrafish.
Figure 5.
The concentration of IL-6 in the brains of OMV-treated and the control group of zebrafish.
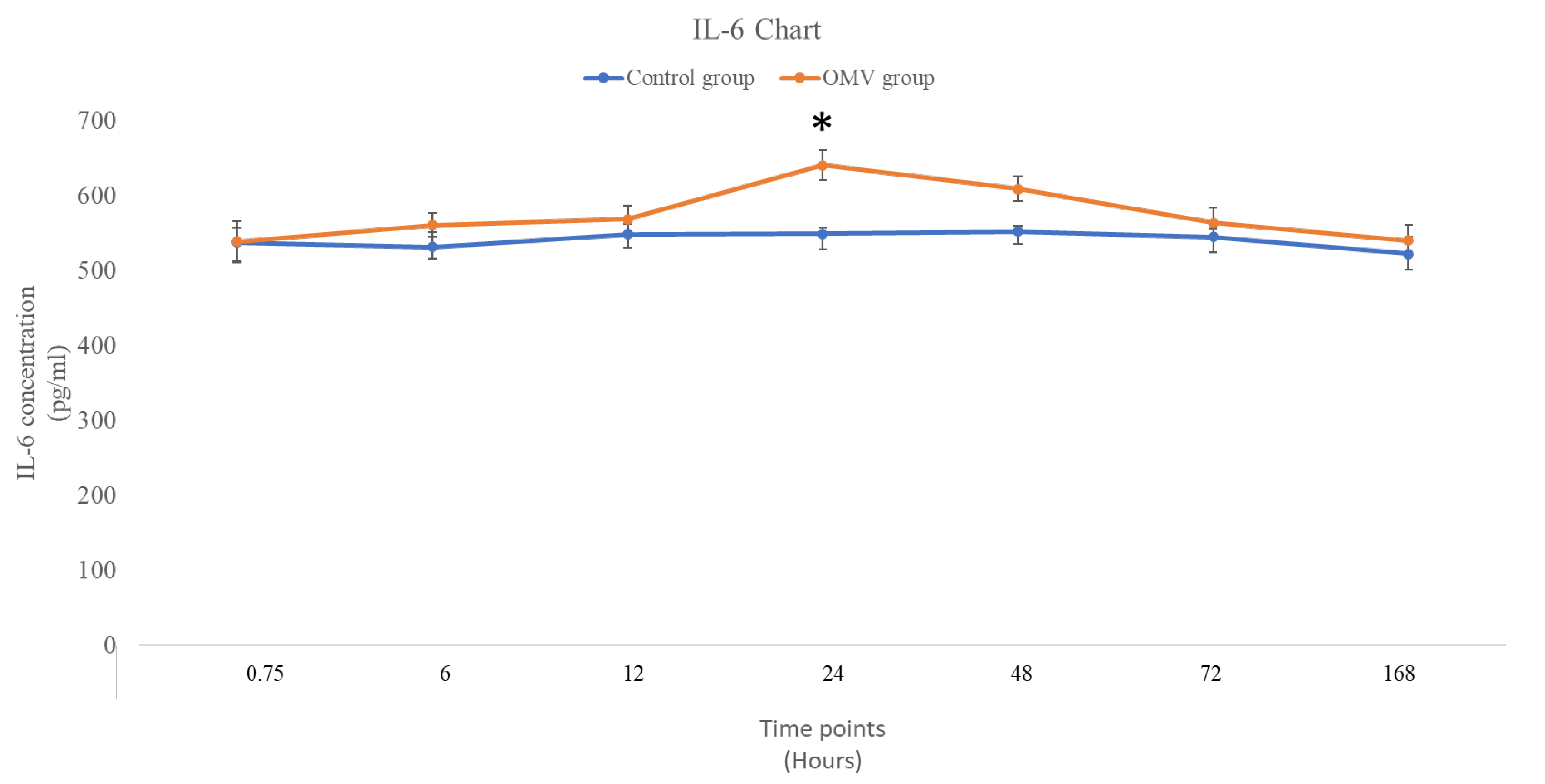
Figure 6.
The concentration of PGE2 in the brains of OMV- treated and the control group of zebrafish. The values represent the mean ± standard error of the mean (n = 3 per group). *** The zebrafish group treated with OMV exhibited a statistically significant difference compared to the control group, with a p-value of less than 0.001.
Figure 6.
The concentration of PGE2 in the brains of OMV- treated and the control group of zebrafish. The values represent the mean ± standard error of the mean (n = 3 per group). *** The zebrafish group treated with OMV exhibited a statistically significant difference compared to the control group, with a p-value of less than 0.001.
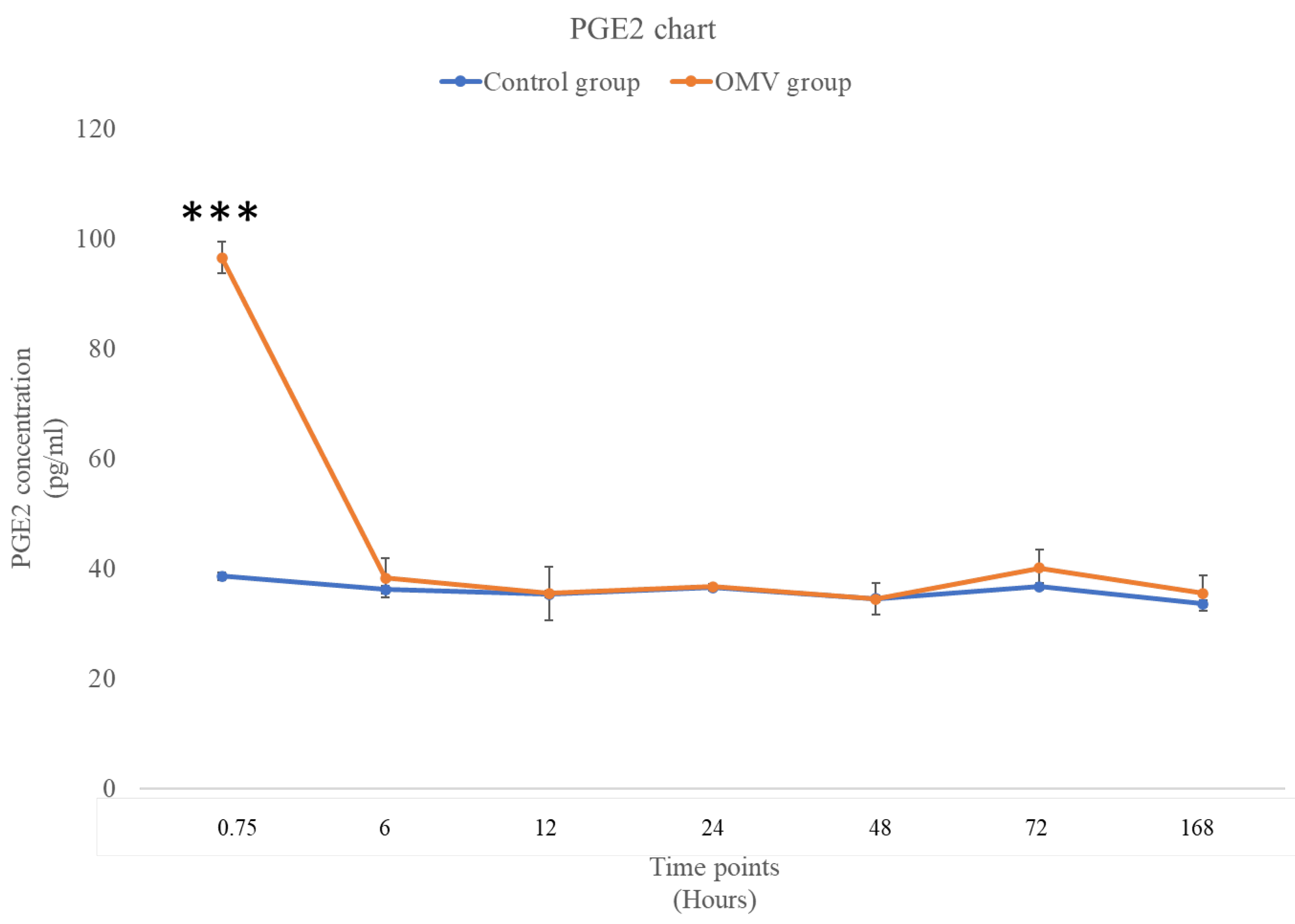
Figure 7.
a. Histological changes in the brain of zebrafish. (A) control group that is injected with normal saline only; (B) the brain of zebrafish injected with OMV at 45 minutes time point;(C) the brain of zebrafish injected with OMV at 6 hours’ time point; (D) the brain of zebrafish injected with OMV at 12 hours’ time points.
Figure 7.
a. Histological changes in the brain of zebrafish. (A) control group that is injected with normal saline only; (B) the brain of zebrafish injected with OMV at 45 minutes time point;(C) the brain of zebrafish injected with OMV at 6 hours’ time point; (D) the brain of zebrafish injected with OMV at 12 hours’ time points.
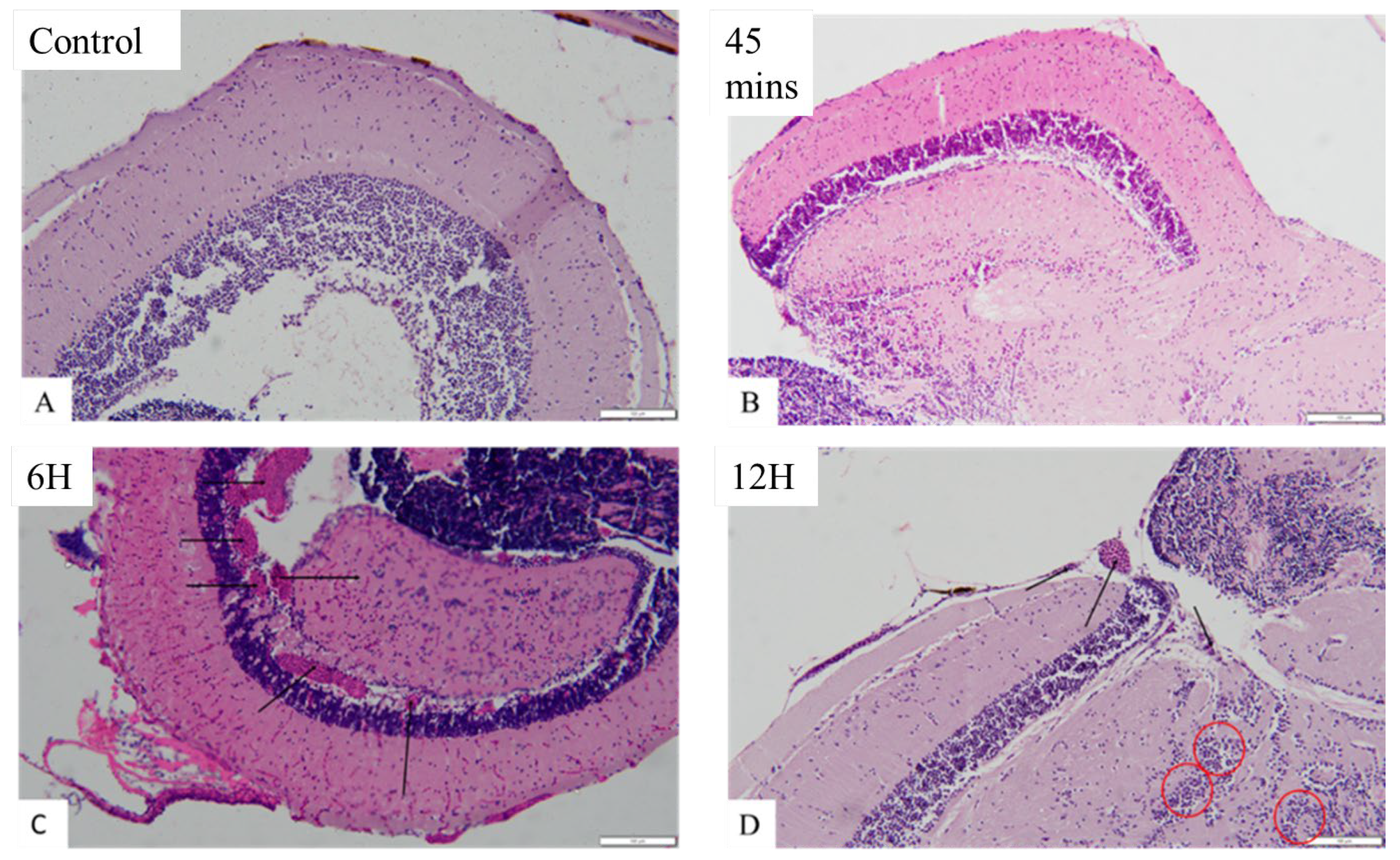
Figure 7.
b. Histological changes in the brain of zebrafish. (E) the brain of zebrafish injected with OMV at 24 hours’ time point; (F) brain of zebrafish injected with OMV at 48 hours’ time point; (G) brain of zebrafish injected with OMV at 72 hours’ time point; (H) brain of zebrafish injected with OMV at 7 days’ time point.
Figure 7.
b. Histological changes in the brain of zebrafish. (E) the brain of zebrafish injected with OMV at 24 hours’ time point; (F) brain of zebrafish injected with OMV at 48 hours’ time point; (G) brain of zebrafish injected with OMV at 72 hours’ time point; (H) brain of zebrafish injected with OMV at 7 days’ time point.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



