
Submitted:
23 August 2024
Posted:
24 August 2024
You are already at the latest version
Abstract
FtsZ is a bacterial protein that plays a crucial role in cytokinesis by forming the Z-ring. This ring acts as a scaffold to recruit other division proteins and guide the synthesis of septal peptidoglycan, which leads to cell constriction. In its native state, the FtsZ protein from Escherichia coli (EcFtsZ) exists as a multi-oligomer comprising dimers, trimers, tetramers, and hexamers in dynamic equilibrium of self-association and dissociation is dependent on protein concentration. This study employed classical biochemical techniques, including native PAGE, size-exclusion chromatography, sucrose gradient, and chemical cross-linking with formaldehyde, to characterize the trimeric state of EcFtsZ. The results of size-exclusion chromatography demonstrate that the trimer of EcFtsZ has a mass of 131 kDa and an experimental friction quotient (Rs/Rmin) of 1.9, which is very similar to the theoretical friction quotient (fn/f1) of 1.8 for a linear trimer. Cross-linking EcFtsZ with formaldehyde resulted in the occurrence of a polypeptide band of 128 kDa, which was recognized by anti-FtsZ antibodies. The result of the sedimentation of EcFtsZ cross-linking with formaldehyde returned a value (Smax/S20,w) equal to 1.9. This experimental value was found to be equal to the theoretical calculation of the sedimentation quotient (Sn/S1) for a lateral trimer. These findings indicate that at micromolar concentrations, the EcFtsZ trimer may assume a linear or lateral structure in the protein. Finally, the protein-protein interaction prediction program (PEPPI) indicated the potential for EcFtsZ to form two protein-protein contacts in its C-terminal domain, which may be involved in trimer formation.
Keywords:
FtsZ
; Oligomers
; friction quotient
; sedimentation quotient
Introduction
The temperature-sensitive mutant Z-filament protein (FtsZ) is a bacterial cytokinesis protein with a molecular weight of 40.3 kDa. It is homologous to tubulin and plays a role in the initial stages of single-cell fission division [1,2,3]. FtsZ assembles in a GTP-dependent manner to form protofilaments, which ultimately form a contractile ring called a Z-ring along the inner circumference of the bacterial cytoplasmic membrane [4]. The protofilament strands are cooperatively assembled from FtsZ units [5]. Z-ring formation and contraction are essential for bacterial cell division. Analytical ultracentrifugation has demonstrated that FtsZ exists in a multimeric state prior to the formation of protofilaments [6]. The dimeric state is responsible for the protein’s GTPase activity, as the catalytic site is formed by the association of two monomeric units [7].
The dimeric state of the protein is of crucial importance for the polymerization mechanism, as it serves as the core for filament formation. Therefore, the oligomerization of FtsZ is an important process for bacterial cleavage [8]. In solution, FtsZ exists as a homomultioligomer in self-association-dissociation equilibrium [9]. Studying a single type of FtsZ oligomer can be challenging. Nevertheless, by combining chemical crosslinking of the protein with separation techniques, it is possible to isolate one or several crosslinked oligomers with high purity. Dimethylpimelimidate (DMP) has been tested for chemical cross-linking of FtsZ. The reaction produces cross-linked dimers, trimers, and tetramers that can be separated by SDS-PAGE [6].
The types of FtsZ oligomers vary with the protein concentration in the solution [6]. At sub-micromolar concentrations, the monomer is the predominant structure. However, between 4 to 10 μM, the amount of dimers and trimers increases up to 70% and 20%, respectively. According to Sossong et al. [6], the trimer is the most abundant oligomer when the concentration of FtsZ is equal to or greater than 100 μM. The function of trimers, tetramers, and other oligomers of greater size of FtsZ is not well understood. One hypothesis is that these oligomers participate in the longitudinal assembly of protofilaments, similar to the trimers and hexamers of Caulobacter crescentus Polar Organizing Protein Z (PopZ) [10]. Another hypothesis is that the larger FtsZ oligomers, such as trimers and tetramers, function as storage structures for the protein in an inactive state when the intracellular concentration of dimers decreases during filament formation. This study characterized two trimers of the EcFtsZ protein, one with a linear structure and the other with a lateral structure, using classical biochemical techniques, including native PAGE, size-exclusion chromatography, sucrose gradient, and chemical cross-linking with formaldehyde.
Materials and methods
Recombinant EcFtsZ
Bacteria were seeded onto a plate with LB agar/ampicillin (0.1 mg/mL) using a nickel loop. The glycerol stock used was from Escherichia coli (E. coli) strain BL21(DE3) transformed with the PMFV57 vector. This vector provides an isopropyl-β-D-1-thiogalactopyranoside (IPTG)-inducible EcFtsZ gene transcription system that confers ampicillin resistance. The plate was incubated at 37°C overnight. Several colonies were picked and grown in duplicate in 5 mL of liquid LB/ampicillin medium (0.1 mg/mL) with shaking at 37°C until the optical density reached 0.6 AU at 600 nm (pre-inoculum). One group of the pre-inoculum was induced with IPTG (0.3 mM) and incubated for three hours. Finally, the induced bacteria were centrifuged at 14000 rpm for five minutes and washed twice with 50 mM phosphate-buffered saline (PBS). To verify EcFtsZ expression, the bacterial pellets were dissolved in loading buffer and boiled, followed by electrophoresis on 10% SDS-PAGE gels. The most promising colony was selected and inoculated in quadruplicate in 4 L of liquid LB/ampicillin (0.1 mg/mL) medium. The culture was incubated at 37 °C with shaking (200 rpm) until an optical density of 0.6 AU at 600 nm was reached. Subsequently, the bacteria were then induced for EcFtsZ expression with IPTG (0.5 mM) for period of four hours at 37 °C. The medium was then centrifuged at 10000 rpm for 10 minutes at 5°C. The resulting pellet was then resuspended in 40 mL of buffer A and previously filtered through MCE (mixed cellulose ester) membranes with a pore size of 0.22 μm. Buffer A contains 50 mM Tris-HCl, 50 mM KCl, 1 mM EDTA, and 10% glycerol.
Extraction of EcFtsZ
A homogenate of bacteria induced in buffer A was combined with a protease inhibitor cocktail tablet (Roche #1187358001), dissolved in 1 mL of buffer A and 400 μL of lysozyme (30 mg/mL) to extract EcFtsZ. The resulting mixture was maintained on ice and stirred periodically at regular intervals for 1.5 hours. Subsequently, it was added 200 μL of 10% w/v deoxycholic acid and incubated at 37 °C until it became viscous, then it was added 300 μL of DNAase I (10 mg/mL) until losing the viscosity. Finally, the homogenate was centrifuged at 100000xg for 60 minutes at 5 °C to recover the supernatant. The supernatant was treated with ammonium sulfate until a concentration of 30% w/v. The mixture was shaken at 10 °C for 20 minutes and centrifuged at 10000xg for 30 minutes at 5 °C. The precipitate was resuspended in 5 mL of buffer A and dialyzed in 2 L of buffer A at 10 °C overnight.
Glutamate Polymerization Cycles
The dialyzed EcFtsZ sample was centrifuged at 10000xg, for 30 minutes at 5 °C. Following this, 5 mL of the supernatant was mixed with 15 mL of polymerization buffer (Buffer P: 50 mM MES, pH 6.5, 1 M monosodium glutamate, 10 mM CaCl2, 10 mM MgCl2) by inversion. Subsequently, it was added 200 μL of 200 mM guanosine triphosphate (GTP), and the mixture was incubated at 37°C for 30 minutes without agitation. During this time, a white precipitate was observed, indicating that the polymerization was complete. The sample was centrifuged at 10000xg, for 30 minutes at 5 °C. The depolymerization cycle involved the resuspension of the precipitate in 5 mL of cold buffer A and subsequent incubation under ice for ten minutes. The sample was centrifuged at 10000xg, for 30 minutes at 5 °C. The procedure was repeated three times (three cycles). Finally, the precipitate sample was resuspended in 2 mL of cold buffer A and dialyzed in 2 L of buffer A at 10 °C overnight. The purification of the EcFtsZ protein was monitored by SDS-PAGE gels.
Turbidity Assay
The turbidity assay was assessed by spectroscopy in a solution containing 50 mM MES buffer (pH 6.5), 50 mM KCl, 10 mM MgCl2, and 15 μM EcFtsZ at 30°C. The sample was heated to 30 °C, and the polymerization reaction was initiated by the addition of 1 mM GTP. The turbidity was measured at an angle of 90° angle by illuminating it with 350 nm light in a Perkin-Elmer LS-50 Spectrofluorimeter, using a 2% transmission filter in the emission beam.
GTPase Activity
The GTPase activity assay was carried out in several stages: 1) A mixture of malachite green 0.045% w/v (VM solution) and sodium molybdate 4.2% w/v (M solution) was prepared in 4M hydrochloric acid. The VM solution was mixed with the M solution in a 3:1 ratio, allowed to stand for 20 minutes, and then filtered using Whatman paper number five. Five milliliters of the filtered VM-M mixture was combined with 100 µL of Triton X-100 0.02% v/v (VM-M-TX solution). 2) Sodium phosphate solutions with concentrations of 5, 10, 20, 50, 100, 170, and 230 µM were prepared to plot the calibration curve. A 50-μL aliquot of each phosphate solution was combined with 800 μL of VM-M-TX solution and vigorously shaken for one minute. Subsequently, 100 μL of 34% w/v sodium citrate was added, and the solutions were allowed to stand for 20 minutes. Finally, the absorbance was measured at 630 nm using a Spectrophotometer UV-Vis LKB 4050. 3) The GTPase reaction solution contained 50 mM HEPES, 100 mM KCl, 10 mM MgCl2, and 0.5 mM GTP sodium salt. The GTPase activity was quantified by incubating 98 μL of the reaction solution with 2 μL of EcFtsZ (Cf ~6 μM) at 37 °C for 10, 15, 30, and 45 minutes. After the reaction time, 70 μL of the reaction mixture and 12 μL of 70% v/v perchloric acid were added and allowed to stand for ten minutes. The supernatant (50 μL) was mixed with VM-M-TX solution (800 μL) and was centrifuged at 10.000xg for five minutes. Bovine serum albumin (BSA) was used as a negative control.
Size-Exclusion Chromatography
The EcFtsZ protein was separated using a FPLC (fast protein liquid chromatography) system, which consisted of a Merck-Hitachi L-6000 pump, Rheodyne injector, Pharmacia Biotech Superose 12 column (1.0 cm x 30 cm) and a UV-Vis L-4250 detector Merck-Hitachi. The column was equilibrated with 10 mM phosphate buffer at pH 7.5 and 100 mM NaCl. The column was calibrated using the following protein standards: α-Lactalbumin (14 kDa), Trypsin inhibitor (20 kDa), Carbonic anhydrase (29 kDa), Ovalbumin (45 kDa), Bovine serum albumin (BSA) (66 kDa), and Alcohol dehydrogenase (150 kDa). The separation was carried out at 25 °C, with a flow rate of 0.66 mL/min. Finally, the calibrated column was injected with 150 μL of EcFtsZ (~11 mg/mL).
The molecular size of proteins according to Irvine [11], was determined by the equation:
where Kav corresponds to the partition coefficient, Ve is the elution volume of each protein, Vo is the exclusion volume, Vt is the total volume of the estimated column a from the column dimensions (Vt = π ∙ r2 x length). The excluded and inclusion volume was determined by chromatographing dextran blue (2000 kDa) and potassium dichromate, respectively.
Kav = (Ve - Vo) / (Vt - Vo)
The molecular weight of EcFtsZ was estimated by extrapolating the calibration curve obtained from plotting the Kav values of each standard protein as a function of the logarithm of the molecular weight. The Stokes radius was also determined by representing (-log Kav)1/2 as a function of the Stokes radius of the standard proteins.
The friction quotient f/fo was calculated by the following equation:
where Rs is the Stokes radius and Rmin is the radius of a smooth non-hydrated sphere. Since the partial specific volume (υ) of an average globular protein is 0.73 cm3/g, the volume occupied by a spherical protein of molecular mass (M) was calculated in Daltons (Da) of the as follows:
f/fo = Rs/Rmin
V (nm3 ) = (0.73 cm3/g) (1021 nm/cm3) x M (Da)/(6.023 x 1023 Da/g)
V (nm3) = 1.212 x 10−3 (nm3/Da) x M (Da)
V = (1.212 x 10−3 x M) nm3
V (nm3) = 1.212 x 10−3 (nm3/Da) x M (Da)
V = (1.212 x 10−3 x M) nm3
The radius of a sphere is (3V/4π)1/3 and assuming that the protein is spherical, the R0 was calculated as follows:
Rmin = (3V/4π)1/3 = (0.066 x M1/3 ) nm
Chemical Crosslinking
A 0.1 M formaldehyde solution was prepared in 50 mM phosphate buffer (K2HPO4/KH2PO4) at pH 7.5. The EcFtsZ protein was dialyzed with phosphate buffer in a 0.5 mL Amicon tube, 3 kDa MWCO, before reacting with formaldehyde. The reaction mixture consisted of 20 μL of formaldehyde (Cf ~0.08 M) and 3 μL of EcFtsZ (Cf ~35 μM) incubated for 20 minutes at 25 °C. The reaction was stopped by adding 2 μL of 1.4 M glycine.
Sedimentation Gradient
The EcFtsZ protein crosslinked and uncrosslinked were separated by sedimentation gradient in 1.2 mL tubes using an Optima L-100XP ultracentrifuge with SW40Ti rotor. The sedimentation was performed using discontinuous sucrose gradients, comprising sucrose solutions with a concentration of 16, 20, 23, 26, 29, 32, and 36% w/v, respectively. These solutions were placed from the bottom to the top of the centrifuge tube with a Hamilton syringe. The samples were centrifuged at 100,000 g, for 24 hours, at 5 °C, without brake. The discontinuous sucrose gradients were calibrated using the following protein standards: α-Lactalbumin (14.2 kDa), Trypsin inhibitor (20 kDa), Bovine serum albumin (66 kDa) and Alcohol dehydrogenase (151 kDa). Fractions of 50 μL were carefully taken from the top to the bottom of the centrifuge tube. The sucrose concentration in each fraction was measured using an optical refractometer.
Determination of Hydrodynamic Parameters
The sedimentation coefficient S20,w of EcFtsZ was calculated using two different methods: the interpolation method according to Clark [12], and the concentration tables method according to McEwen [13]. The sedimentation coefficients in Svedberg units (S) were determined using the following protein standards: α-Lactoalbumin at 1.75 S [14], soybean trypsin inhibitor at 2.6 S [15], bovine serum albumin at 4.3 S, and alcohol dehydrogenase at 7.4 S [16].
The hydrodynamic properties of a rigid lateral and linear trimer were calculated using a theoretical model. Kirkwood, Bloomfield, et al. developed the following equations that were employed in this study [17].
Kirkwood-Bloomfield equations:
where n is the number of monomers, r: radius of the sphere, R: distance from the center between two spheres, fn/f1: friction quotient of the multimer of n monomers, f1: friction coefficient of a monomer, fn: friction coefficient of n monomers, Sn/S1: sedimentation quotient of the multimer of n monomers, S1: sedimentation coefficient of a monomer, Sn: sedimentation coefficient of the multimer of n monomers.
The sedimentation coefficient S20,w in Svedberg units was determined by McEwen’s method using the following equation [13]:
were (ΔI) is the time integral between two concentrations of sucrose (% w/w) for a given particle density.
Protein-Protein Interaction Prediction Program
The PEPPI (Pipeline for the Extraction of Predicted Protein-protein Interactions) program was utilized to forecast protein-protein interactions (PPI) among EcFtsZ subunits. The program predicted the likelihood of direct physical interaction between two amino acid sequences of EcFtsZ (GenBank: CAD6022189.1) using various independent prediction methods, including machine learning-based classification. The program can be accessed through the following link: https://seq2fun.dcmb.med.umich.edu/PEPPI/.
Other Procedures
The samples were separated by SDS-PAGE according to Laemmli [18] and by PAGE-native without stacking gel according to Anh et al. [19]. Gels were stained with Coomassie blue or silver. The Novex native protein standards (Life Technologies) #LC0725 and the molecular mass standard 26616 (Thermo Scientific) were utilized. The concentration of EcFtsZ was determined using BSA as the standard protein [20]. For the immunoblot analysis, the proteins were separated using SDS-PAGE and transferred to a nitrocellulose membrane [21]. The membranes were then blocked with 5% nonfat milk in TBS-T buffer (100 mM Tris-Base, 150 mM NaCl, and 0.1% Tween-20). Subsequently, the membranes were incubated with Abcam Rabbit anti-FtsZ polyclonal antibody (ab225991) (1:1000) in TBS-T, followed by incubation with the anti-IgG antibody conjugated to Rockland Rabbit alkaline phosphatase (800-656-7625) (1:10000) in TBS-T. The membranes were developed by incubating them with a mixture of 10 mL of alkaline phosphatase buffer (100 mM Tris-base, 100 mM NaCl, and 5mM MgCl2), 66 μL of NBT (nitro blue tetrazolium) 1mg/mL, and 33 μL of BCIP (5-bromo-4-chloro-3-indoyl phosphate) 5mg/mL, both substrates previously dissolved in dimethylformamide (DFA). The purity percentage and apparent molecular weight of the protein were determined by analyzing the electrophoresis gels using GelAnalyzer 2010a software (www.gelanalyzer.com) developed by Istvan Lazar Jr. and Istvan Lazar.
Results
Functional Properties of Purified EcFtsZ
The EcFtsZ protein, expressed in E. coli, was extracted by precipitation with ammonium sulfate and purified through successive polymerization cycles and depolymerization with potassium glutamate and GTP. The EcFtsZ protein exhibited an apparent molecular weight of approximately 39 kDa as determined by SDS-PAGE (Figure 1A). The purification process yielded 33 mg of total protein with a purity of 89% and a concentration of 11 mg/mL (~273 μM). Although the protein recovery percentage was not quantified, it was observed that there was a loss of EcFtsZ during the depolymerization step (data not shown). To test the integrity and functionality of the purified protein, both its enzymatic activity and the polymerization reaction were measured. The purified EcFtsZ exhibited a GTPase activity of 8.8 μM Pi∙min−1 ∙μM−1, which is comparable to the GTPase activities reported by Rivas et al. [9] 4.8 μM Pi∙min−1 ∙μM−1 and Beuria et al. [22] 13.8 μM Pi∙min−1∙μM−1 for EcFtsZ.
The polymerization reaction of EcFtsZ, induced by GTP and Mg2+, was measured using the turbidity of the solution through light scattering (Figure 1B) in 50 mM MES buffer (pH 6.5). The progress plot of the EcFtsZ polymerization reaction demonstrated a rapid increase in turbidity immediately after the addition of GTP and Mg2+, followed by a change in rate until 8 minutes, at which point turbidity began to decrease. This decrease is likely due to GTP depletion. The results were consistent with those reported in previous studies by Scheffers et al. [7], Rivas et al. [9], Chen et al. [23], and Diaz-Espinoza et al. [24].
Native PAGE of Purified EcFtsZ
Electrophoresis was conducted under native conditions without gel stacking, in accordance with the methodology described by Ahn et al. [19], to directly observe the oligomerization state of EcFtsZ, as reported by Sossong et al. [6] and Rivas et al. [9] using other analytical techniques. The polypeptide bands presented in the experiments were significantly broad, which initially made it difficult to identify the oligomers (Figure 1C). Two polypeptide bands were observed above 66 and 146 kDa, corresponding to the dimeric and tetrameric states of EcFtsZ, respectively. The concentration of EcFtsZ has a negative impact on the resolution of the oligomers in the gel, due to the broadening of the polypeptide bands. The addition of magnesium chloride (5 mM) and potassium chloride (50 mM) to the electrophoresis gel significantly decreased this band broadening (Figure 1D). In these conditions, three polypeptide bands were observed between 8 and 10 μM of protein, corresponding to the dimeric, tetrameric, and hexameric states. The trimer was observed at 15μM of protein and the resolution of the bands is lost from 30 μM of protein (Figure 1D). This result confirms that EcFtsZ exists in a multi-oligomeric state in solution, with the concentration and types of oligomers varying with protein concentration. To better see this variation, each of the polypeptide bands was measured by optical density and plotted as a function of EcFtsZ concentration (Figure 1E). The data demonstrate that the dimeric, trimeric, and tetrameric states were achieved at concentrations of 10, 15, and 30 μM EcFtsZ, respectively.
Chemical Cross-Linking
We employed chemical cross-linking with formaldehyde to identify the oligomeric states of EcFtsZ in the presence or absence of magnesium chloride and potassium chloride. The results revealed the presence of a polypeptide double band at 130 kDa, which belongs to two trimers of EcFtsZ in the absence of magnesium chloride and potassium chloride (Figure 2A). These polypeptide bands in the presence of the salts were not observed (Figure 2B). The results indicate that Mg2+ and K+ ions are not conducive to the formation of trimers during protein-protein interactions. It is noteworthy that the presence of these salts resulted in the detection of dimers that were resistant to the dissociating conditions of sodium dodecyl sulfate electrophoresis (Figure 2B). Finally, the hydrodynamic properties of EcFtsZ cross-linked with formaldehyde were analyzed using a sucrose gradient.
Size-Exclusion Chromatography
The EcFtsZ protein was separated by size-exclusion chromatography using a calibrated column with molecular mass markers. Three peaks during the elution period were observed. The EcFtsZ protein was detected in peak number 3 at 21.04 minutes with the highest absorbance (Figure 2C). The concentration of the EcFtsZ protein in the column was approximately 15 μM. At this concentration, dimers and trimers are the most common oligomers, according to Sonsog et al. [6]. When plotting the partition coefficient (Kav) of each standard against the logarithm of their molecular masses [25], it was observed that the molecular mass of the EcFtsZ protein in peak number 3 was 131 kDa (Figure 2D), which is 3.3 times higher than the molecular mass of the monomer (~39 kDa) observed by SDS-PAGE. These results suggest that peak number 3 is primarily composed of trimers. A plot of the (-logKav)1/2 versus the Stokes radius of each standard protein [16] (Figure 2E) revealed that the Stokes radius of EcFtsZ was 4.41 nm, in contrast to the Stokes radius of FtsZ monomer equal to 2.3 nm [9]. The theoretical molecular mass of a FtsZ monomer as proposed by Erickson (40.3 kDa) [17] was employed in conjunction with the Stokes radius (4.41 nm) to calculate the experimental friction quotient (f/fo) for the EcFtsZ protein, which yielded a value of 1.9 (Table 1). The FtsZ monomer has a f/fo value of 1.25, which is typical of a globular protein. A moderately elongated protein has a f/fo range between 1.5 and 1.9 [17]. We assume that the EcFtsZ protein, located at peak number 3 with a f/fo equal to 1.9, corresponds to a trimer and not to an elongated monomer. To test this hypothesis, we calculated the theoretical friction quotient (fn/f1) using the Kirkwood-Bloomfield equations for a dimer and linear trimer, which yielded values of 1.3 (Table S1) and 1.8 (Table 1 and S1) respectively. This last value of 1.8 closely matches the experimental value. Additionally, the theoretical friction quotient (fn/f1) for a lateral trimer was also calculated but yielded a value further from the experimental value (1.6) (Table 1 and S1).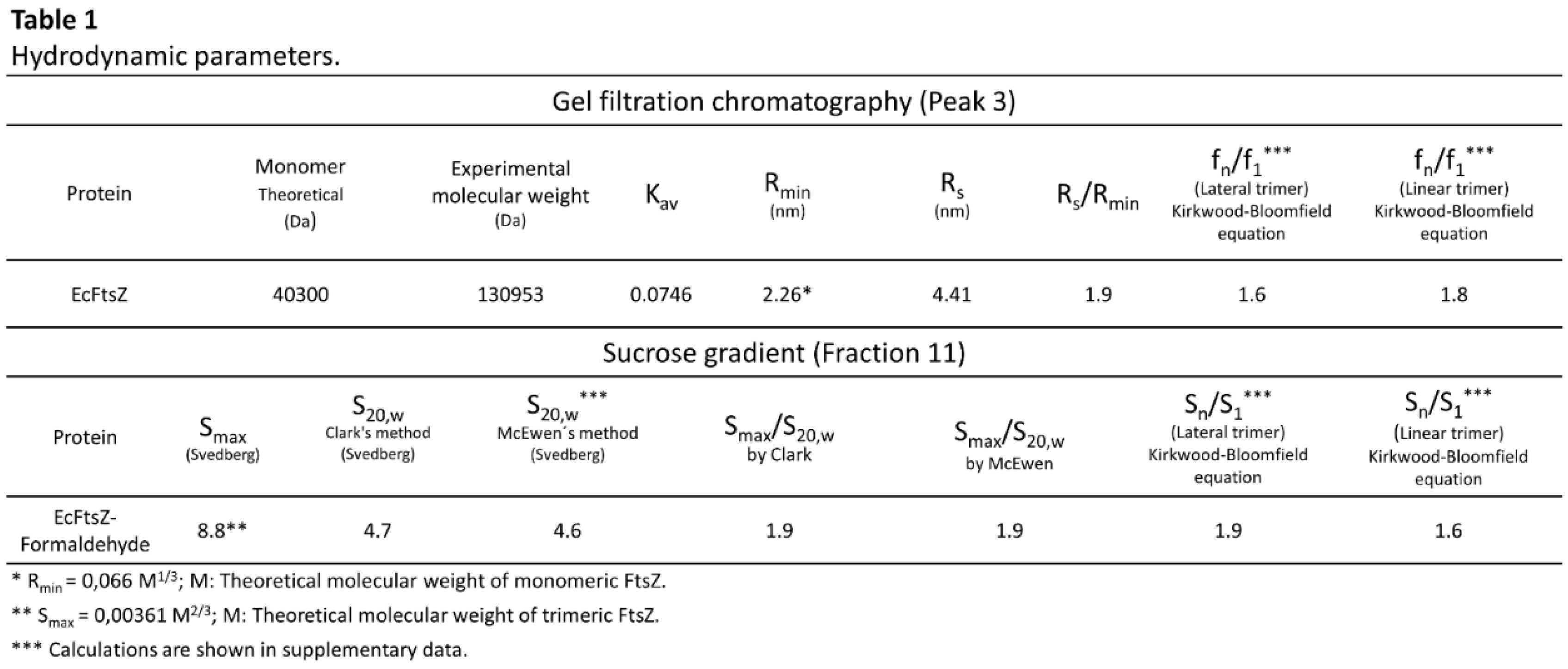
Sedimentation Gradient
Samples of EcFtsZ, with and without formaldehyde, were loaded onto a sucrose gradient and separated by ultracentrifugation (Figure 3A and B). The polypeptide profile observed by native-PAGE of the fractions of EcFtsZ without formaldehyde is shown in Figure 3A. This profile displays four distinct bands, corresponding to the dimeric, trimeric, tetrameric, and hexameric states of EcFtsZ. The polypeptide band corresponding to the trimeric state exhibited a greater width and intensity than the other polypeptide bands between fractions 7 and 14. Another characteristic is the polypeptide bands of the oligomers identified in the sedimentation gradient were symmetrically distributed around fractions 9 and 10 (Figure 3A). The sedimentation process was conducted for 24 hours, allowing for the establishment of new protein self-association-dissociation equilibria during the migration of proteins within the sucrose solution. This resulted in similar quantities of the oligomer types between fractions. The immunoblot performed on fraction 9 revealed the presence of two bands recognized by anti-EcFtsZ antibodies. The first band was observed at 39 kDa and the second at 75 kDa. These bands correspond to the monomer and dimer of EcFtsZ, respectively. Notably, the dimer demonstrated resistance to the dissociating conditions of SDS-PAGE electrophoresis (Figure 3A).
Figure 3B shows the polypeptide profile of the samples of EcFtsZ cross-linked with formaldehyde and separated by sedimentation. Samples from fractions 8 to 13 were combined to form the set (8-13) and analyzed by SDS-PAGE and immunoblotting. The set (8-13) contains a band of approximately 128 kDa recognized by anti-EcFtsZ antibodies. This band corresponds to a trimer of EcFtsZ that has been cross-linked with formaldehyde. The separation peak of the EcFtsZ protein was observed in fraction 11.
The sucrose gradient was calibrated using standard proteins (data not shown), and a linear relationship was observed between the sedimentation coefficient (S20,w) and the migration volume of each standard protein, according to Clark’s method [12]. The S20,w of the EcFtsZ protein in fraction 11 was of 4.68 S (Figure 3C) while by McEwen’s method was of 4.62 S (Supplementary data S3).
The coefficient Smax was calculated for an EcFtsZ trimer with a mass of 120.9 kDa and resulted in a value of 8.8 S (Table 1). The experimental sedimentation quotient (Smax/S20,w) of the EcFtsZ protein cross-linked was equal to 1.9. The theoretical sedimentation quotient (Sn/S1) of a lateral trimer was equal to the experimental value (1.9), while that of a linear trimer was equal to 1.6 (Table 1 and S2). The results of size-exclusion chromatography and sedimentation for sucrose gradient indicate that the EcFtsZ trimers can exist in two structural forms: linear and lateral.
Prediction of Protein-Protein Interactions
The PEPPI program predicted two tight interactions with a high final likelihood value involving the C-terminal domains of the subunits (C-C contact) (Figure 4A and B). These results suggest that EcFtsZ oligomers may form through protein-protein interactions in the C subdomain. One of the interactions occurs between amino acids Arg202 and Phe210 located in the T7 loop (Figure 4A and Figure 4C). The other interaction occurs between amino acids Lys319 and Thr327, which are located in the N-segment of a region described as an intrinsically disordered region (IDR) that is highly flexible in the C-terminal domain (Figure 4B and 4C) [39,40]. To determine whether these amino acids are conserved or not, sequence alignment was performed on FtsZ from E. coli, O. antarctica, and M. jannaschii, which are mesophilic, psychrophilic, and thermophilic bacteria, respectively. The alignment revealed that most of the amino acids in the T7 loop are conserved, while the IDR segment is not conserved in E. coli (Figure 4C).
Discussion
Purified EcFtsZ
The characterization of a protein necessitates the acquisition of the protein in a highly pure state. Beuria et al. [27] described a novel method for purifying EcFtsZ through glutamate-induced polymerization, achieving a purity of 95% after the second polymerization cycle but we achieved a purity of 89% after the third polymerization cycle. A fourth cycle did not result in a significant improvement in protein purity (data not shown). Although we did not achieve higher purity, the impurities did not affect the electrophoretic mobility on native gels, the GTPase activity, and the polymerization reaction of the EcFtsZ protein.
Native PAGE
The native PAGE is an efficient technique for studying oligomeric proteins and protein complexes [28,29,30], but the proteins are separated on the acrylamide gels with a lower resolution because the electrophoretic mobility is primarily influenced by the intrinsic charge negative of the proteins. The molecular masses and oligomeric states of proteins and complexes are not easily determined by native PAGE, unless the proteins have favorable physical parameters, especially an isoelectric point below 5.4 [29], as in the case of EcFtsZ, which has a theoretical isoelectric point of 4.6. The main characteristic of the native PAGE gels of the EcFtsZ protein is the broadness of the bands, suggesting that several distinct populations of the protein of similar size may be represented. This phenomenon of broadening of band has been observed in homologous FtsZ from plants [31]. When the magnesium and potassium ions are added to the separation gel, the broadening of the bands of EcFtsZ is reduced, improving the analytical separation. The possible cause of reducing of broadening of bands may be the reduction of non-specific protein-protein interactions due to the significant conformational changes induced for coordination of magnesium in the GTPase proteins such as FtsZ [32,33], the self-association of the FtsZ protein induced for magnesium [9]. Finally, the native PAGE has been used successfully in the oligomeric state study of proteins homologous to FtsZ from plants [31] and to analyze the constituents of protein complexes in the E. coli divisome [34].
Cross-Linking with Formaldehyde
Cross-linking is typically employed to capture and stabilize noncovalent, transient, or labile interactions between proteins [35]. Formaldehyde is a cross-linking agent that reacts when two reactive groups in two interacting proteins are near one another. Formaldehyde reacts with the lateral amino group of a lysine, forming a Schiff base on the protein or subunit. Then reacts with the lateral amido group of an glutamine or asparagine, or with the lateral guanidinium group of an arginine located on a second subunit or protein, producing a cross-linked secondary amine [36,37]. In the absence of magnesium and potassium, the cross-linking reaction between the EcFtsZ protein and formaldehyde produced trimers with a sedimentation quotient characteristic of a lateral trimer, while the results by size-exclusion chromatography showed a friction quotient characteristic of a linear trimer. The presence of lateral and linear oligomers would also be with the broad band observed in native PAGE (Figure 1C), indicating the presence of protein populations of similar mass but with different electrophoretic mobilities. Such differences are due to variations in the charge/mass ratio regimes and could be caused, for example, by lateral and linear in subunit interactions, which would support the existence of several types of oligomers. These lateral and linear interactions in the oligomers of FtsZ were observed in the FtsZ1 and FtsZ2 protein from Arabidopsis thaliana [31]. To elucidate the nature of the lateral and linear interactions in the EcFtsZ protein, we used the sequence of EcFtsZ (GenBank AJF449721) and subjected it to analysis with the Protein-Protein Interaction Prediction Program (PEPPI) available at https://zhanggroup.org/PEPPI/ [38]. The analysis entailed the prediction of Protein-Protein interactions between two Apo subunits of EcFtsZ.
Prediction of Protein-Protein Interactions
The T7 loop (tubulin loop number 7) of FtsZ plays a key role in the self-association of protein monomers [41]. The T7 loop is part of the active site for GTP hydrolysis and is formed by two FtsZ monomers [41]. In M. jannaschii protofilaments, the complete GTPase site of FtsZ was observed to be formed by the T7 loop of one subunit approaching the N-terminal GTP-binding domain of another subunit, resulting in an N-C contact between the subunits [41,42]. In addition, N-C contacts between dimers of EcFtsZ have been observed in molecular dynamics simulations [43,44]. The introduction of point mutations in loop T7 of EcFtsZ (M206C, N207C, D209C) has been observed to have a significant effect on the polymerization and GTPase activity of the protein. However, these mutations do not appear to affect the in vitro interaction with wild-type EcFtsZ, indicating that mutations in the T7 loop do not appear to abolish EcFtsZ self-association [41]. The PEPPI program was used to analyze the interaction between a wild-type EcFtsZ sequence and mutant EcFtsZ sequences (M206C, N207C, D209C) to observe the effect of these mutations in the T7 loop on C-C contacts. The prediction results indicated that these mutations did not affect the C-C contacts between the subunits (data not shown). This suggests that the self-association between wild-type and mutant EcFtsZ observed by Scheffers et al. [41] may occur through C-C contacts.
A lateral inter-protofilament contact interface has been observed in Mycobacterium tuberculosis FtsZ protofilaments near of the outer faces of the S7 and S10 strands in the C-terminal subdomain [45]. More recently, Guan et al. [46] identified the formation of two pairs of salt bridges in the C subdomain. These bridges are formed by Arg229 of one subunit and Asp301 of the other and are located at the lateral inter-protofilament contact interface. The interaction predicted by the PEPPI program in segment N of the IDR near of the S10 strand in the C subdomain of EcFtsZ suggests that this segment may be involved in lateral inter-protofilament contacts as well as lateral C-C contacts in EcFtsZ oligomers prior to polymerization.
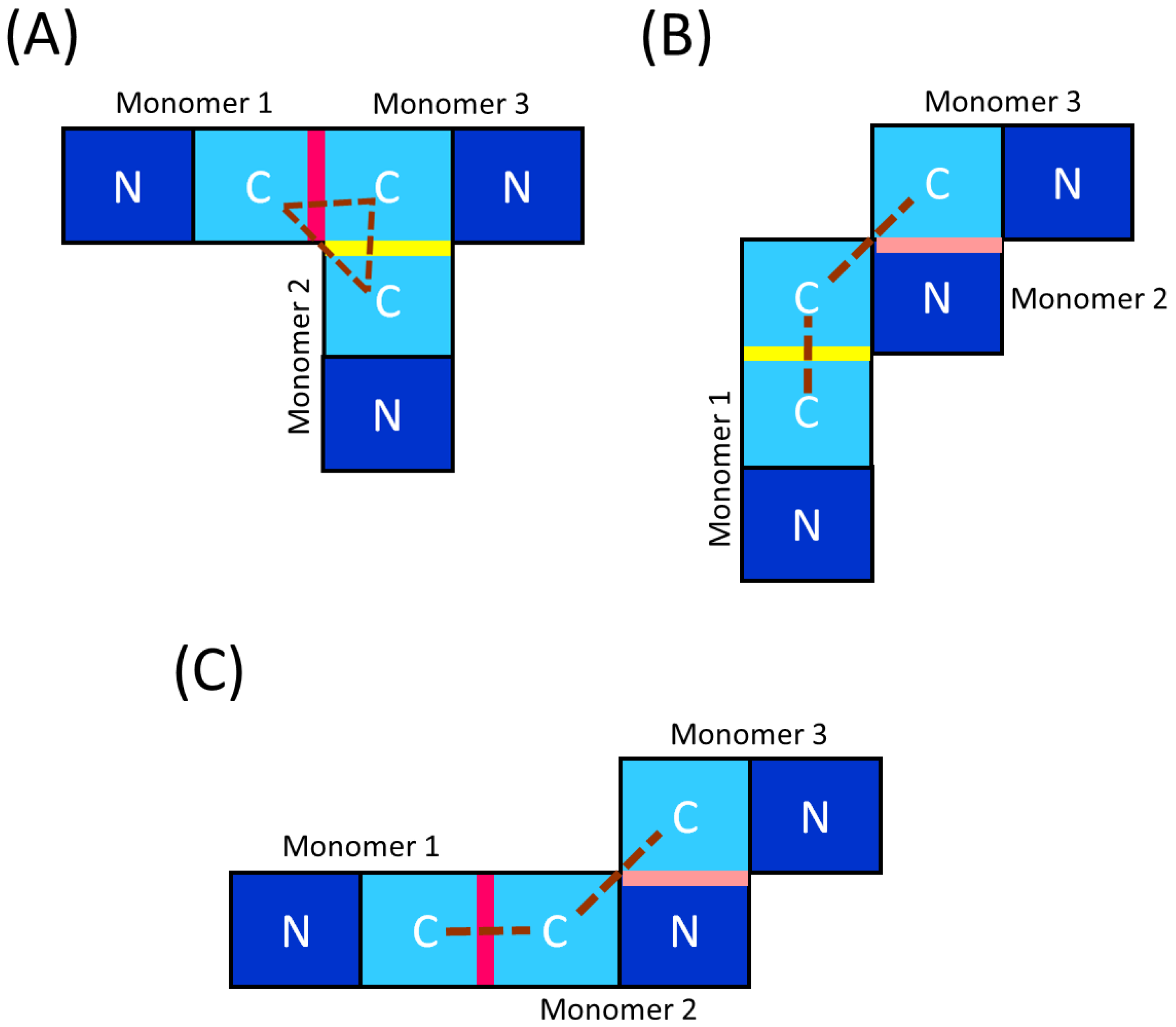
Following on from the results described in this paper, we propose that the trimer of the EcFtsZ protein can exist in three different forms, for example, two lateral forms and one linear form (Figure 5). One of the lateral form would be by C-C contacts between the N segments of the IDR (lateral contact) and C-C contacts between T7 loops (Figure 5A), while the other lateral form would be by C-C contacts between the N segments of the IDR (lateral contact) and N-C contacts between a T7 loop and the GTP-binding domain (Figure 5B). The linear form would be by the C-C contacts between the T7 loops and the N-C contacts between a T7 loop and the GTP-binding domain (Figure 5C). Finally, trimers are a structural motif important in nature, being the most common quaternary structure motif in the Protein Data Bank (PDB) after dimers and monomers. In particular, lateral trimers account for approximately two-thirds of all PDB trimers, whereas only one-third of trimers are linear trimers [47]. Future structural studies on the trimer of EcFtsZ are needed to determine the contact zones between the subunits and better understand the protein-protein interactions of the EcFtsZ subunits involved in the assembly process of larger oligomers.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Araujo N.A. and Pouchucq L. designed the study; Araujo N.A and Veloso M. performed the experiments; Araujo N.A. analyzed the data and wrote the manuscript. All authors read and approved the final manuscript.
Acknowledgments
Araujo N.A. thanks the Internationalization Project UCH1566 of the Chilean Ministry of Education for funding the postdoctoral fellowship with which this research was carried out. Araujo N.A. thanks Dr. Valentina Carrasco for the donation of recombinant EcFtsZ, the molecular mass standard for native gels and the anti-EcFtsZ polyclonal antibody. Araujo N.A. would like to express gratitude to Dr. José Bubis for their invaluable assistance in the review and correction of the text. All the authors thank Professor Dr. Octavio Monasterio for allowing the development of this research and for supporting it with materials and reagents in the Laboratory of Structural and Molecular Biology (BEM).
Conflicts of Interest
The authors declare that they have no conflicts of interest regarding the contents of this article.
References
- Nogales, E.; Downing, K.H.; Amos, L.A.; Löwe, J. Tubulin and FtsZ form a distinct family of GTPases. 1998, Nat. Struct. Biol. 5. [CrossRef]
- Addinall, S.G.; Holland, B. The Tubulin Ancester, FtsZ, Draughtsman, Designer and Driving Force for Bacterial Cytokinesis. J. Mol. Biol. 2002, 318, 219–236. [Google Scholar] [CrossRef] [PubMed]
- D. Bramhill, Bacterial cell division, Annu. Rev. Cell. Dev. Biol. 1997. [CrossRef]
- Lutkenhaus, J.; Addinall, S.G. BACTERIAL CELL DIVISION AND THE Z RING. Annu. Rev. Biochem. 1997, 66, 93–116. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Aportela, E.; López-Blanco, J.R.; Andreu, J.M.; Chacón, P. Understanding Nucleotide-Regulated FtsZ Filament Dynamics and the Monomer Assembly Switch with Large-Scale Atomistic Simulations. Biophys. J. 2014, 107, 2164–2176. [Google Scholar] [CrossRef] [PubMed]
- Sossong, T.M.; Brigham-Burke, M.R.; Hensley, P.; Pearce, K.H. Self-Activation of Guanosine Triphosphatase Activity by Oligomerization of the Bacterial Cell Division Protein FtsZ. Biochemistry 1999, 38, 14843–14850. [Google Scholar] [CrossRef]
- Scheffers, D.-J.; de Wit, J.G.; Blaauwen, T.D.; Driessen, A.J.M. GTP Hydrolysis of Cell Division Protein FtsZ: Evidence that the Active Site Is Formed by the Association of Monomers. Biochemistry 2001, 41, 521–529. [Google Scholar] [CrossRef]
- den Blaauwen, T.; Andreu, J.M.; Monasterio, O. Bacterial cell division proteins as antibiotic targets. Bioorg. Chem. 2014, 55, 27–38. [Google Scholar] [CrossRef]
- Rivas, G.; López, A.; Mingorance, J.; Ferrándiz, M.J.; Zorrilla, S.; Minton, A.P.; Vicente, M.; Andreu, J.M. Magnesium-induced Linear Self-association of the FtsZ Bacterial Cell Division Protein Monomer. 275, 1174; 9. [Google Scholar] [CrossRef]
- Bowman, G.R.; Perez, A.M.; Ptacin, J.L.; Ighodaro, E.; Folta-Stogniew, E.; Comolli, L.R.; Shapiro, L. Oligomerization and higher-order assembly contribute to sub-cellular localization of a bacterial scaffold. Mol. Microbiol. 2013, 90, 776–795. [Google Scholar] [CrossRef]
- Irvine, G.B. Determination of Molecular Size by Size-Exclusion Chromatography (Gel Filtration). Curr. Protoc. Cell Biol. 2000, 6, 5–5. [Google Scholar] [CrossRef]
- Clark, R.W. Calculation of s20,w values using ultracentrifuge sedimentation data from linear sucrose gradients, an improved, simplified method. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 1976, 428, 269–274. [Google Scholar] [CrossRef]
- McEwen, C. Tables for estimating sedimentation through linear concentration gradients of sucrose solution. Anal. Biochem. 1967, 20, 114–149. [Google Scholar] [CrossRef]
- Morris, G.A. The Self-Assembly and Structure of Caseins in Solution. Biotechnol. Genet. Eng. Rev. 2002, 19, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, E.; McLaren, A.D. Studies on Crystalline Trypsin, Soybean Trypsin Inhibitor and Inhibitor-Trypsin Compound with the Ultracentrifuge1,2. J. Am. Chem. Soc. 1953, 75, 2587–2591. [Google Scholar] [CrossRef]
- Siegel, L.M.; Monty, K.J. Determination of molecular weights and frictional ratios of proteins in impure systems by use of gel filtration and density gradient centrifugation. Application to crude preparations of sulfite and hydroxylamine reductases. Biochim. et Biophys. Acta (BBA) - Biophys. Incl. Photosynth. 1966, 112, 346–362. [Google Scholar] [CrossRef]
- Erickson, H.P. Size and Shape of Protein Molecules at the Nanometer Level Determined by Sedimentation, Gel Filtration, and Electron Microscopy. Biol. Proced. Online 2009, 11, 32–51. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Ahn, T.; Yim, S.-K.; Choi, H.-I.; Yun, C.-H. Polyacrylamide Gel Electrophoresis without a Stacking Gel: Use of Amino Acids as Electrolytes. Anal. Biochem. 2001, 291, 300–303. [Google Scholar] [CrossRef]
- M.M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding, Anal. Biochem. 1976. [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef]
- Beuria, T.K.; Shah, J.H.; Santra, M.K.; Kumar, V.; Panda, D. Effects of pH and ionic strength on the assembly and bundling of FtsZ protofilaments: A possible role of electrostatic interactions in the bundling of protofilaments. Int. J. Biol. Macromol. 2006, 40, 30–39. [Google Scholar] [CrossRef]
- Chen, Y.; Anderson, D.E.; Rajagopalan, M.; Erickson, H.P. Assembly Dynamics of Mycobacterium tuberculosis FtsZ. 282, 2773. [Google Scholar] [CrossRef]
- Díaz-Espinoza, R.; Garcés, A.P.; Arbildua, J.J.; Montecinos, F.; Brunet, J.E.; Lagos, R.; Monasterio, O. Domain folding and flexibility of Escherichia coli FtsZ determined by tryptophan site-directed mutagenesis. Protein Sci. 2007, 16, 1543–1556. [Google Scholar] [CrossRef]
- T.C. Laurent, J.A. T.C. Laurent, J.A. Killander, Theory of gel filtration and its experimental verification, J. Chromatogr A. 1964. [Google Scholar]
- V. Bloomfield, W.O. V. Bloomfield, W.O. Dalton, K.E. van Holde, Friction coefficients of multisubunit structures, I. Theory, Biopolymers. 1967. [Google Scholar]
- Beuria, T.K.; Krishnakumar, S.S.; Sahar, S.; Singh, N.; Gupta, K.; Meshram, M.; Panda, D. Glutamate-induced Assembly of Bacterial Cell Division Protein FtsZ. J. Biol. Chem. 2003, 278, 3735–3741. [Google Scholar] [CrossRef]
- C.N. Gallagher, R.E. C.N. Gallagher, R.E. Huber, Monomer-Dimer Equilibrium of Uncomplemented M15 β-Galactosidase from Escherichia coli, Biochem. 1997. [Google Scholar]
- Wittig, I.; Schägger, H. Advantages and limitations of clear-native PAGE. Proteomics 2005, 5, 4338–4346. [Google Scholar] [CrossRef] [PubMed]
- Wittig, I.; Karas, M.; Schägger, H. High Resolution Clear Native Electrophoresis for In-gel Functional Assays and Fluorescence Studies of Membrane Protein Complexes. Mol. Cell. Proteom. 2007, 6, 1215–1225. [Google Scholar] [CrossRef]
- Smith, A.G.; Johnson, C.B.; Vitha, S.; Holzenburg, A. Oligomerization of plant FtsZ1 and FtsZ2 plastid division proteins. Arch. Biochem. Biophys. 2011, 513, 94–101. [Google Scholar] [CrossRef]
- Lebbink, J.H.G.; Fish, A.; Reumer, A.; Natrajan, G.; Winterwerp, H.H.K.; Sixma, T.K. Magnesium Coordination Controls the Molecular Switch Function of DNA Mismatch Repair Protein MutS. J. Biol. Chem. 2010, 285, 13131–13141. [Google Scholar] [CrossRef]
- Hu, Z.; Marti, J. Discovering and Targeting Dynamic Drugging Pockets of Oncogenic Proteins: The Role of Magnesium in Conformational Changes of the G12D Mutated Kirsten Rat Sarcoma-Guanosine Diphosphate Complex. Int. J. Mol. Sci. 2022, 23, 13865. [Google Scholar] [CrossRef]
- Trip, E.N.; Scheffers, D.-J. A 1 MDa protein complex containing critical components of the Escherichia coli divisome. Sci. Rep. 2015, 5, 18190–18190. [Google Scholar] [CrossRef] [PubMed]
- X. Tang, J.E. X. Tang, J.E. Bruce, Chemical Cross-Linking for Protein–Protein Interaction Studies. In: Lipton, M.S., Paša-Tolic, L. (eds) Mass Spectrometry of Proteins and Peptides. 2009. [Google Scholar] [CrossRef]
- Fraenkel-Conrat, H.; Olcott, H.S. The Reaction of Formaldehyde with Proteins. V. Cross-linking between Amino and Primary Amide or Guanidyl Groups. J. Am. Chem. Soc. 1948, 70, 2673–2684. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.A.; Frey, B.L.; Smith, L.M.; Auble, D.T. Formaldehyde Crosslinking: A Tool for the Study of Chromatin Complexes. J. Biol. Chem. 2015, 290, 26404–26411. [Google Scholar] [CrossRef]
- Bell, E.W.; Schwartz, J.H.; Freddolino, P.L.; Zhang, Y. PEPPI: Whole-proteome Protein-protein Interaction Prediction through Structure and Sequence Similarity, Functional Association, and Machine Learning. J. Mol. Biol. 2022, 434, 167530–167530. [Google Scholar] [CrossRef]
- Huang, K.-H.; Mychack, A.; Tchorzewski, L.; Janakiraman, A. Characterization of the FtsZ C-Terminal Variable (CTV) Region in Z-Ring Assembly and Interaction with the Z-Ring Stabilizer ZapD in E. coli Cytokinesis. PLOS ONE 2016, 11, e0153337. [Google Scholar] [CrossRef]
- Muthukumar, V.C. Escherichia coli FtsZ molecular dynamics simulations. J. Biomol. Struct. Dyn. 2023, 42, 2653–2666. [Google Scholar] [CrossRef] [PubMed]
- Scheffers, D.-J.; de Wit, J.G.; Blaauwen, T.D.; Driessen, A.J.M. GTP Hydrolysis of Cell Division Protein FtsZ: Evidence that the Active Site Is Formed by the Association of Monomers. Biochemistry 2001, 41, 521–529. [Google Scholar] [CrossRef] [PubMed]
- A Oliva, M.; Cordell, S.C.; Löwe, J. Structural insights into FtsZ protofilament formation. Nat. Struct. Mol. Biol. 2004, 11, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- V.C. Muthukumar, Molecular Dynamics simulation of the E. 2802. [CrossRef]
- Muthukumar, V.C. The dynamics of Escherichia coli FtsZ dimer. J. Biomol. Struct. Dyn. 2023, 1–14. [Google Scholar] [CrossRef]
- Li, Y.; Hsin, J.; Zhao, L.; Cheng, Y.; Shang, W.; Huang, K.C.; Wang, H.-W.; Ye, S. FtsZ Protofilaments Use a Hinge-Opening Mechanism for Constrictive Force Generation. Science 2013, 341, 392–395. [Google Scholar] [CrossRef]
- Guan, F.; Yu, J.; Yu, J.; Liu, Y.; Li, Y.; Feng, X.-H.; Huang, K.C.; Chang, Z.; Ye, S. Lateral interactions between protofilaments of the bacterial tubulin homolog FtsZ are essential for cell division. eLife 2018, 7. [Google Scholar] [CrossRef]
- E.D. Levy, J.B. E.D. Levy, J.B. Pereira-Leal, C. Chothia, S.A. Teichmann, 3D complex: a structural classification of protein complexes. PLOS Comput Biol. 2006. [Google Scholar]
Figure 1.
Purification, polymerization and native PAGE of the EcFtsZ protein. (A) SDS-PAGE gel of purification of EcFtsZ. (B) Polymerization of EcFtsZ at 30°C. GTP addition is indicated by the arrow. (C) Native PAGE gel without KCl and MgCl2. (D) Native PAGE gel with KCl (50 mM) and MgCl2 (5 mM). (E) Optical density of polypeptide bands of oligomers versus protein concentration in gel with KCl and MgCl2. Isopropyl-β-D-thiogalactoside (IPTG).
Figure 1.
Purification, polymerization and native PAGE of the EcFtsZ protein. (A) SDS-PAGE gel of purification of EcFtsZ. (B) Polymerization of EcFtsZ at 30°C. GTP addition is indicated by the arrow. (C) Native PAGE gel without KCl and MgCl2. (D) Native PAGE gel with KCl (50 mM) and MgCl2 (5 mM). (E) Optical density of polypeptide bands of oligomers versus protein concentration in gel with KCl and MgCl2. Isopropyl-β-D-thiogalactoside (IPTG).
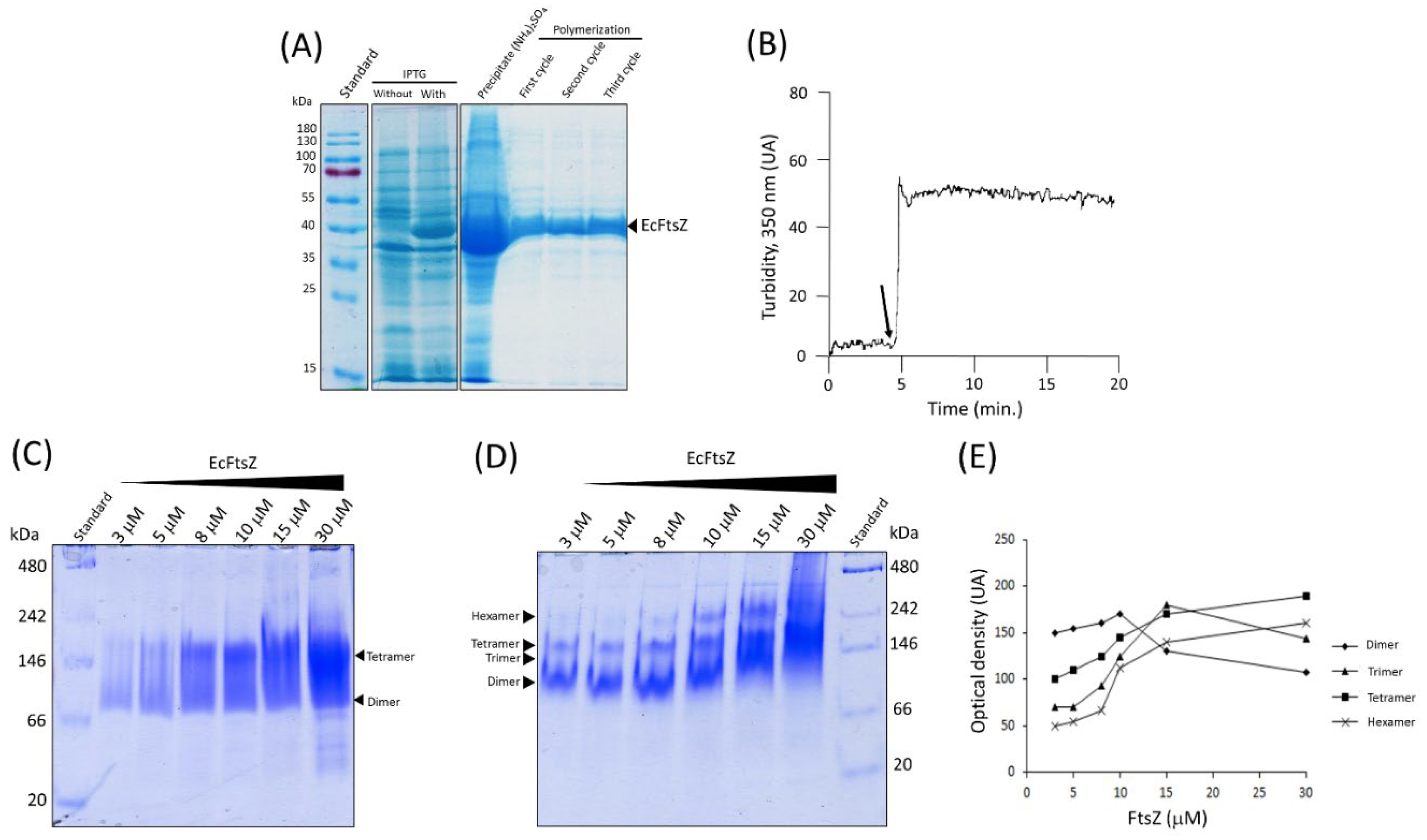
Figure 2.
Cross-linking and size-exclusion chromatography of EcFtsZ. (A) Cross-linking of EcFtsZ with formaldehyde without KCl and MgCl2. (B) Cross-linking of EcFtsZ with formaldehyde, KCl (5 μM) and MgCl2 (50 mM). (C) Gel filtration chromatography of EcFtsZ without Cross-linking. Immunoblotting of peak 3 with Anti-FtsZ within of the Figure 2C. (D) Plot of Partition coefficient (Kav) versus log10 molecular weight. (E) Plot of -(log Kav)1/2 versus Stokes radius. Ponceau red staining (RP). Immunoblot (IB). Retention time (tr).
Figure 2.
Cross-linking and size-exclusion chromatography of EcFtsZ. (A) Cross-linking of EcFtsZ with formaldehyde without KCl and MgCl2. (B) Cross-linking of EcFtsZ with formaldehyde, KCl (5 μM) and MgCl2 (50 mM). (C) Gel filtration chromatography of EcFtsZ without Cross-linking. Immunoblotting of peak 3 with Anti-FtsZ within of the Figure 2C. (D) Plot of Partition coefficient (Kav) versus log10 molecular weight. (E) Plot of -(log Kav)1/2 versus Stokes radius. Ponceau red staining (RP). Immunoblot (IB). Retention time (tr).
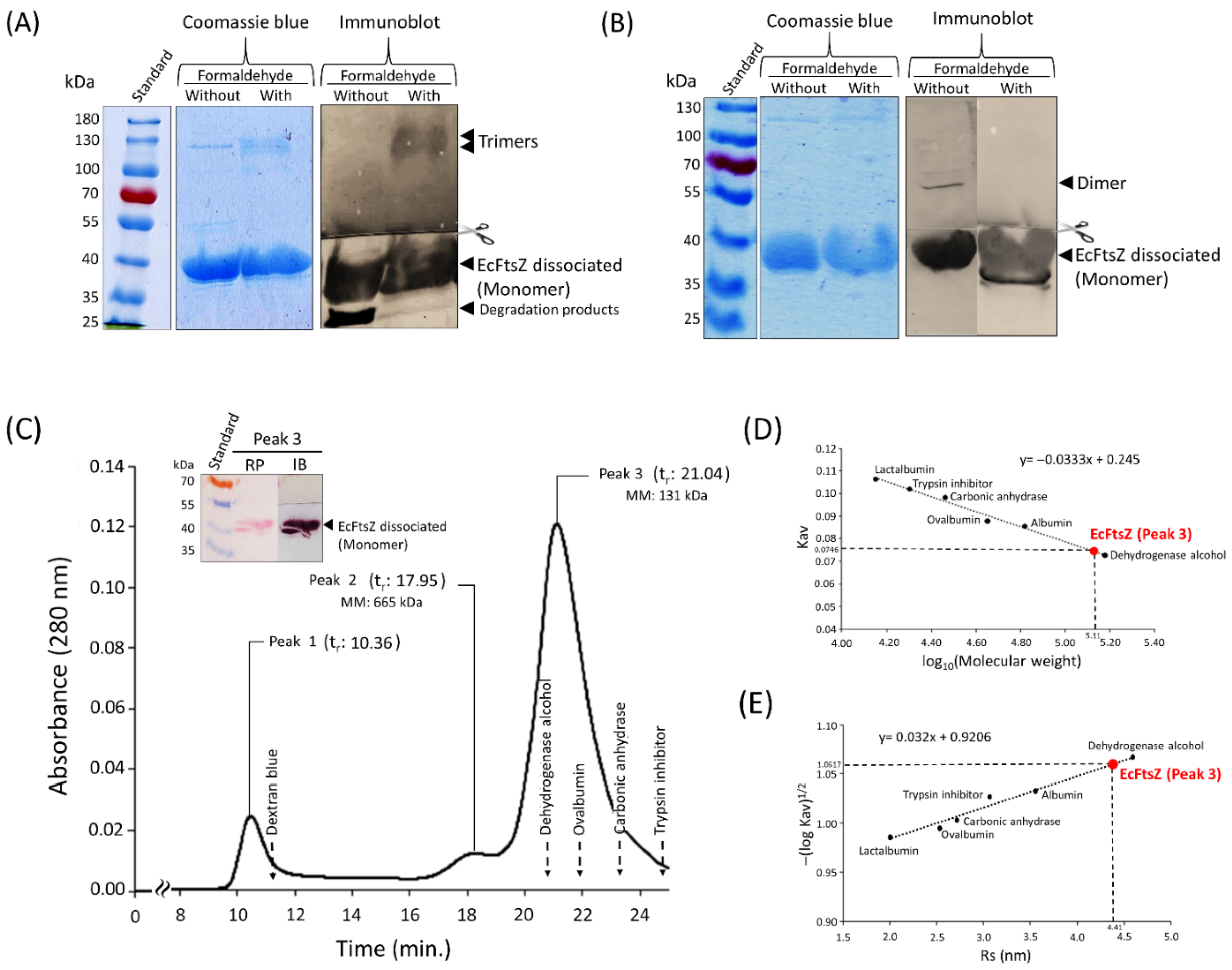
Figure 3.
Sedimentation gradient of the EcFtsZ protein. (A) Native PAGE of the sucrose gradient of EcFtsZ without formaldehyde. Peak of gradient EcFtsZ (∗). Immunoblotting of fraction 9 with Anti-FtsZ within of the Figure 3A. (B) SDS-PAGE of the sucrose gradient of EcFtsZ with formaldehyde. Peak of EcFtsZ-Formaldehyde (∗∗). Immunoblotting of fractions set (8-13) with Anti-FtsZ within of the Figure 3B. (C) Plot of Sedimentation coefficient (S20,w) versus protein migration volume (Clark’s method) [12]. Ponceau red staining (RP). Immunoblot (IB).
Figure 3.
Sedimentation gradient of the EcFtsZ protein. (A) Native PAGE of the sucrose gradient of EcFtsZ without formaldehyde. Peak of gradient EcFtsZ (∗). Immunoblotting of fraction 9 with Anti-FtsZ within of the Figure 3A. (B) SDS-PAGE of the sucrose gradient of EcFtsZ with formaldehyde. Peak of EcFtsZ-Formaldehyde (∗∗). Immunoblotting of fractions set (8-13) with Anti-FtsZ within of the Figure 3B. (C) Plot of Sedimentation coefficient (S20,w) versus protein migration volume (Clark’s method) [12]. Ponceau red staining (RP). Immunoblot (IB).
Figure 4.
Prediction of protein-protein interactions in the EcFtsZ protein and alignment of FtsZ sequences from mesophilic, psychrophilic and thermophilic bacteria. (A and B) Left: Ribbon molecular model of the crystal structure of truncated EcFtsZ-GDP (11-316) (PDB: 6UMK). (A) Right: C-C contact between the T7 loops of two EcFtsZ subunits predicted with the PEPPI program [39]. (B) Right: C-C contact between the N segments of the IDR of two EcFtsZ subunits predicted with the PEPPI program [39]. The GenBank sequence AJF449721 was used for prediction of protein-protein interactions. The molecular models were visualized with the RasMol program. Intrinsically Disordered Region (IDR). (C) Alignment of FtsZ protein sequences of E. coli (mesophilic) (GenBank AJF449721), O. antarctica (psychrophilic) (NCBI WP046046011201.1) and M. jannaschii (thermophilic) (NCBI WP_010869869.1). The alignment was conducted using the Clustal Omega program on the EBI website (https://www.ebi.ac.uk/jdispatcher/msa/clustalo) with the default parameters. The secondary structures for E. coli FtsZ are shown at the top. The red lines indicate the amino acids involved in the C-C contacts predicted by the PEPPI program.
Figure 4.
Prediction of protein-protein interactions in the EcFtsZ protein and alignment of FtsZ sequences from mesophilic, psychrophilic and thermophilic bacteria. (A and B) Left: Ribbon molecular model of the crystal structure of truncated EcFtsZ-GDP (11-316) (PDB: 6UMK). (A) Right: C-C contact between the T7 loops of two EcFtsZ subunits predicted with the PEPPI program [39]. (B) Right: C-C contact between the N segments of the IDR of two EcFtsZ subunits predicted with the PEPPI program [39]. The GenBank sequence AJF449721 was used for prediction of protein-protein interactions. The molecular models were visualized with the RasMol program. Intrinsically Disordered Region (IDR). (C) Alignment of FtsZ protein sequences of E. coli (mesophilic) (GenBank AJF449721), O. antarctica (psychrophilic) (NCBI WP046046011201.1) and M. jannaschii (thermophilic) (NCBI WP_010869869.1). The alignment was conducted using the Clustal Omega program on the EBI website (https://www.ebi.ac.uk/jdispatcher/msa/clustalo) with the default parameters. The secondary structures for E. coli FtsZ are shown at the top. The red lines indicate the amino acids involved in the C-C contacts predicted by the PEPPI program.
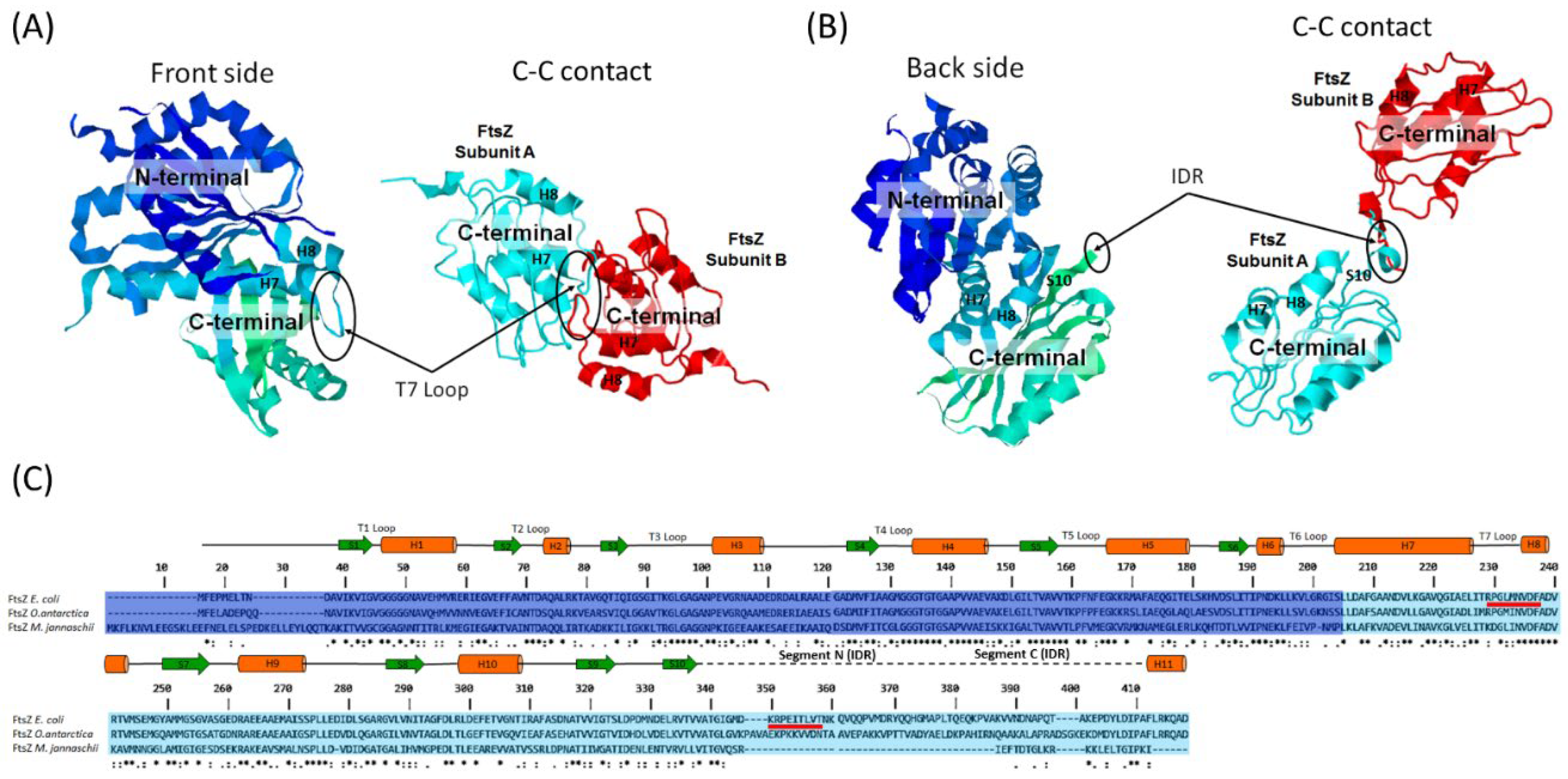
Figure 5.
Molecular representation of the trimers of EcFtsZ. (A) Lateral trimer with two C-C contacts. (B) Lateral trimer with one C-C contact. (C) Linear trimer with one C-C contact. The dashed line represents the arrangement between the C subunits of the protein. The fuchsia line indicates the lateral C-C contacts between the T7 loops. The yellow line indicates the lateral C-C contacts between the N segments from IDR. The pink line indicates the lineal N-C contacts between the GTP-binding N-domain of one subunit and the T7 loop of the C-domain of another subunit.
Figure 5.
Molecular representation of the trimers of EcFtsZ. (A) Lateral trimer with two C-C contacts. (B) Lateral trimer with one C-C contact. (C) Linear trimer with one C-C contact. The dashed line represents the arrangement between the C subunits of the protein. The fuchsia line indicates the lateral C-C contacts between the T7 loops. The yellow line indicates the lateral C-C contacts between the N segments from IDR. The pink line indicates the lineal N-C contacts between the GTP-binding N-domain of one subunit and the T7 loop of the C-domain of another subunit.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



