
Submitted:
23 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
The aim of this study was to detect seasonal and species differences in serum 25-hydroxy vitamin D (25(OH)D) concentrations during summer and winter months in Asian (Elephas maximus) and African elephants (Loxodonta africana), black (Diceros bicornis) and Indian rhinoceroses (Rhinoceros unicornis) and Malayan tapirs (Tapirus indicus) - when kept in captivity in (Western) Europe. Both elephant species had a low circulating level of serum 25(OH)D while African elephants (median 33.2 nmol/l – range: 21.2-58.8 nmol/l,) did not show a seasonal variation. Asian elephants had significant higher circulating levels of serum 25(OH)D compared to their African counterparts, but also did not show a seasonal difference (median 68.7, – range 55.5-110.6 nmol/l). Both rhinoceros species investigated had higher serum 25(OH)D levels compared to both elephant species; the Indian rhinoceros had high circulating levels year around (median 107.9 – range: 106.3-132.8 nmol/l), while the black rhinoceroses showed significant lower 25(OH)D levels in winter (summer median 109.0– range 51.5-251.0 nmol/l; winter median 58.3 nmol/l - range 23.5-226.0 nmol). Malayan tapirs have very low levels of serum 25(OH)D (median < 20.3 – range < 20.3-33.5 nmol/l), which is comparable to horses. Higher levels of circulating 25(OH)D of elephants and rhinoceroses compared to horses, could indicate that elephants and rhinoceroses are capable of producing vitamin D. However, this requires further testing. If this is indeed the case, this might indicate that the Indian rhinoceros are capable of producing enough endogenous vitamin D year around at latitudes around 52°N, while both elephant species and the black rhinoceros are not. To the author’s knowledge, this is the first report of vitamin D levels in tapirs.
Keywords:
vitamin D
; tapir
; elephant
; rhino
; seasonal
; latitude
1. Introduction
Elephants, rhinoceroses and tapirs, also commonly known as pachyderms, are kept a lot in zoos all over the world predominantly for conservation efforts. Most of them are housed in institutions in Europe and North America at much higher latitude than their natural range. Therefore, there are many differences regarding the natural habitat and the UVb radiation and environmental temperature of the captive versus wild pachyderms. This might contribute to hypovitaminosis D in captivity, since both factors are crucial in endogenous vitamin D production, which can result in hypocalcaemia and/or hypophosphataemia in humans [1] and might also lead to hypocalcaemia and/or hypophosphataemia in captive pachyderms. In captivity, problems with hypocalcaemia, especially in relation to dystocia and musculoskeletal problems have been described for (captive) Asian elephants [2,3,4,5,6]. In captive Asian elephants, both in Europe and in current range countries, hypocalcaemia is reported [2,6,7], all associated with unbalanced nutrition/too low dietary calcium (Ca) levels [6,8,9]. Van Sonsbeek et al. (2013)[7] demonstrated a significant increase of Ca plasma levels after oral Ca supplementation in summer and oral cholecalciferol administration in summer and winter in captive Asian elephants in Europe, but not in African elephants. To the author’s knowledge, no reports are available on hypocalcaemia in rhinoceroses nor tapirs.
Based on the habitats and anatomy of the two elephant species it is expected that serum 25(OH)D levels follow the same trend as the UV index, meaning it fluctuates with seasons, having a high summer 25(OH)D level and lower winter vit D level. However, they are often compared to horses regarding their vitamin D metabolism [7,10,11]. If they are truly comparable with horses, serum 25(OH)D levels would be low or undetectable [12]. It was also suggested that African elephants might have a higher threshold to endogenously produce cholecalciferol in the skin compared with Asian elephants, like seen in humans which originates from living around the equator versus people living in the Northern Hemisphere [7]. Both temperature and UV radiation in Europe is much lower compared to Africa (see Figure 1), so it might also be possible that the threshold to endogenously produce cholecalciferol is not met in African elephants and that African elephants will not show a seasonal trend and low overall 25(OH)D values in their serum. Actually, this condition is not only the case for the African species, but for all pachyderms.
The aim of the study was to detect seasonal and species differences in the vitamin D levels during summer and winter months in Asian and African elephants, Malayan tapir, and black and Indian rhinoceros kept in captivity in (Western) Europe.
2. Materials and Methods
2.1. Samples
Serum collected for health monitoring of the captive pachyderms from the Rotterdam Zoo and stored at the biobank at the Rotterdam Zoo (-80°C), and serum sent to the Rotterdam Zoo for progesterone analyses from pachyderms at other institutions in the period of January 2020 to October 2023 were used for this study, with permission obtained for use of the samples. Details of individuals participating in the study can be found in Table 1.
For the African elephant, 97 unique samples of ten individual elephants housed at two different zoological institutions (Wuppertal Zoo, Hilvarenbeek Zoo) were available: age at the start of the monitoring period ranged from 1-35 years (years); eight females (age 4-35 years), two males (age 1-27 years). For the Asian elephant, 94 unique samples of five individual elephants housed at one institution (Rotterdam Zoo; age 0-50 years old) were analysed; four females (10-50 years) and 1 male (1 year). For the black rhinoceros, 90 unique samples of five individuals were available: age 0-19 years, three males (0-19 years) and two female (3-9 years) housed at one institution (Rotterdam Zoo) while for the Indian rhinoceros, four unique samples of one female rhinoceros housed at one institution (Rotterdam Zoo), age at the start of the period five years (only samples during 2022 and 2023 were used). For the Malayan tapir, 36 unique samples of two individual tapirs, age 7-9 years; one male (seven years, only samples in 2023 were used) and one female (nine years) housed at one institution (Rotterdam Zoo) were analysed.
All animals in all institutions had access to outside during summer for at least 8 hours during the daytime, in winter this was often reduced to a maximum of 6 hours during the daytime, depending on the temperature. No analyses were performed on the food items. Cholecalciferol intake was estimated based on the bodyweight of the animal, amount of concentrates and information on the amount of cholecalciferol on the package, this can be found in Table 1.
2.2. Vitamin D Analysis
Total serum 25(OH)D concentrations were determined by the VIDAS® enzyme linked fluorescent assay (ELFA) (Biomérieux, Marcy-L’Etoile, France). The VIDAS 25 OH vitamin D TOTAL assay combines an enzyme immunoassay competition method with a final fluorescent detection (ELFA) [13] with a detection limit of 20.3 to 315 nmol/l.
Based on the UV index [14], the summer and winter can be divided into different months. A UV index of 1-2 means no UV radiation, a UV index of 3-4 represents very poor UV radiation, 5-6 a weak UV radiation, 7-8 is strong UV radiation, and 9-10 and higher represents a very strong UV radiation while > 11 is extreme UV radiation. When looking at the months in the year, during 2020-2023, January, November and December had a mean index of 0-1; May to July were graded 6-7. March and October showed always an index of two, while March, April, Augustus and September varied between three and five. Summer and Winter were divided based in UV index (< 2 and > 5, respectively) resulting in the winter being November, December, January and Summer as May, June, July. On the 21st of March Spring officially starts (longer and more sunny days in Western Europe) and ends the 21st of June when Summer is officially starting. The 21st of September Autumn officially starts (shorter and colder and more clouded days) with the 21st of December the start of the Winter. Here we define Summer as the period from April-September and Winter from October to March.
2.3. Statistical Methods
All tests were performed with R statistics (version 4.3.3). A Kolmogorov Smirnov test was performed on all data of the African elephants, Asian elephants and black rhinoceros. None of the animals had normally distributed data, therefore, a Kruskal Wallace rank sum test was performed to determine statistical differences between seasons in all species. As a post hoc test, a Dunn test was performed to determine the significant differences between the groups, with Holm-Bonferroni correction (ggstatsplot package in R).
To determine if there was a significant difference between African and Asian elephants, a Wilcoxon rank sum test was performed. Alpha error in the analysis was set at p < 0.05. Some individuals accounted for more samples compared to others and to correct for this, the mean was analysed per animal per season, which then counted for n = 1. The total mean was analysed by calculating the mean per individual, which accounted for n = 1. The animals which contributed only with one or two samples or did not have samples in all seasons were only used to calculate the total mean and median (individual number 5, 6, 7, 10, 15, 20; Table 1) but were not used in the Summer/Winter comparison.
No further testing was performed on the Indian rhinoceros, because of the low number of samples, and the Malayan tapir who turned out to have numerous samples for which values were under the detection level.
3. Results
As mentioned before, the data for both elephants and the black rhinoceroses were normally distributed. Therefore, next to the total mean and standard deviation, the median and range is provided for all species in Table 2. The median and range of Summer and Winter based on the two different division techniques (UV index vs Calendar) are also shown in Table 2. Figure 1 shows the 25(OH)D analyses in all African and Asian elephants and black rhinoceroses over time. Figure 2 is a ggplot, showing the distribution of the samples during the different seasons of the African and Asian elephants and black rhinoceroses.
No significant differences could be detected between the two seasons for African elephants (p = 0.71), Asian elephants (p = 0.09) by making use of the UV radiation index, nor when using the calendar. Comparing the mean of the Asian and African elephants, the latter showed a significant lower 25(OH)D serum level (p < 0.001).
For black rhinoceroses a significant difference in the 25(OH)D in Winter and Summer was observed when the division was made based on both UV index and calendar data (index: p < 0.001; calendar: p < 0.001) was found. The Indian rhinoceros data only contained four samples of one individual and only one of those samples originated in Winter, which had a high concentration compared with the black rhinoceroses.
The Malayan tapir 25(OH)D levels were almost all under the detection level of 20.3 nmol/l.
For the Indian rhinoceros and Malayan tapir, no additional statistics were performed.
4. Discussion
The samples originate only of a small number of individuals housed in only 1-2 institutions. Due to multiple missing samples and to reduce the impact of individuals on the mean, a mean was calculated per season per animal or a mean per animal reducing sample size even further. UV index in range countries is generally higher, with the exception for the Asian elephant and the Indian rhinoceros as shown in Figure 1. Therefore, it might not be a surprise that the African elephant serum 25(OH)D levels were significantly lower than that of their Asian counterparts supports the theory that the threshold for vitamin D production has not been met. However, another African megaherbivore, the black rhinoceros, did show significant higher levels of serum 25(OH)D in Summer while it also lives in areas with high UV radiation index and high ambient temperature, indicating that it might (at least partly) meet its threshold to endogenously produce vitamin D. The Indian rhinoceros showed higher mean vitamin D levels compared to black rhinoceroses although no statistics could be performed due to the low sample size. So it is unclear if the difference is statistically significant, which would require testing multiple Indian rhinoceroses. Furthermore, the Indian rhinoceros showed less variation in summer and winter compared to the black rhinoceroses, perhaps due to adaptations to their natural environment, where they live in range countries were UV radiation is not always very strong or extreme (Figure 1).
The 25(OH)D levels in this study are considerably higher compared to the findings in other studies performed by the author [7,15]. Perhaps this might be due to a different test procedure, since the VIDAS analyses total 25(OH)D instead of only D3 metabolites. Here, a mean 25(OH)D of 34.5 ± 9.0 nmol/l in African elephants was found versus a mean (± SD) 25(OH)D3 of 11 ± 5 nmol/l by van Sonsbeek et al. (2011)[15] and 15.6 ± 7.7 nmol/l by van Sonsbeek et al. (2013)[7]. A study conducted in Florida, North America in African elephants [11] found a 25(OH)D level of 39.4 ± 18.7 nmol/l, which is actually more comparable to the values in the present study, despite that there is still the same difference in latitude between the Netherlands (52°N) and Florida (28°N). Asian elephants are more studied with regards to their vitamin D metabolism in comparison with their African counterparts. Mean 25(OH)D of Asian elephants reported here was 75.3 ± 21.1 nmol/l, versus 25(OH)D3 of 36 ± 11 nmol/l by van Sonsbeek et al. (2011)[15] and 35.6 ± 11.7 nmol/l by van Sonsbeek et al. (2013)[7]. This finding of much higher circulating levels of 25(OH)D compared to 25(OH)D3 in previous studies could indicate that both elephant species have high levels of circulating 25(OH)D2, which is supported by Childs-Sanford et al. (2020, 2023, 2024)[10,16,17]. Additionally, this can also indicate that (Asian) elephants can absorb cholecalciferol added to their diet, which has been proven in Asian elephants in a recent study by Childs-Sanford et al. (2023)[16]. In the study of Childs-Sanford et al. (2020, 2023)[10,16], captive Asian elephants sera samples were analysed discriminating 25(OH)D2 and D3 (43°N). They showed no detectable 25(OH)D3 levels in sera, 25(OH)D2 levels were detected and were on average 17.5 ± 2.2 nmol/l, which is much lower compared to the previous studies by van Sonsbeek et al. (2011, 2013)[7,15] and values reported here. A recent study analysed the 25(OH)D levels in serum on captive Asian elephants in India (Tamil Nadu; latitude 8-14°N). They found a mean of 27.7 ± 4.9 nmol/l (range: 21.2-35.4 nmol/l) [18] which in considerably lower compared to our findings and also compared to previous studies [7,15], except for the studies by Childs-Sanford et al. (2020, 2023, 2024)[10,16,17], unfortunately no details were provided on the diet or sunlight exposure by Perumbilly et al. (2024)[18] except that this was found to be sufficient. As mentioned above, this can be due to use of total versus differentiated 25(OH)D tests, or differences in cholecalciferol supplementation/daily vitamin D intake. The current study was unable to demonstrate significant seasonal differences in circulating 25(OH)D levels in both elephant species, which is in comparison to previous mentioned studies [7,10,11,15,17]. However, in van Sonsbeek et al. (2013)[7], a significant decrease in 1,25(OH)2D3 in African elephants was noticed during winter. This was negated when adding cholecalciferol to the diet which indicates that African elephants might not be able to endogenously produce sufficient vitamin D in winter months in the Northern Hemisphere. In the same study in Asian elephants, a significant change in bone marker concentration, indicative of higher levels of bone resorption during the winter time were found, which might also suggest that Asian elephants do not seem capable of producing sufficient amounts of cholecalciferol in winter months at the latitude of the study in the Northern Hemisphere and that Ca absorption might be (at least partly) depended on vitamin D. However, it should be mentioned that it is still unknown whether both elephants species are capable of producing endogenous cholecalciferol at all.
In a study by Olds et al. (2018)[19] seasonal variation of 25(OH)D levels was detected in two captive Eastern black rhinoceros in America with the mean serum 25(OH)D level of 100.6 nmol/l. This is slightly lower compared to the mean of 126.2 ± 70.2 nmol/l reported here. However, our median is much lower, namely 86.3 nmol/l (range 72.9-237.0 nmol/l). A study in wild black rhinoceroses shows higher levels: a mean of 139.0 nmol/l [20] which is higher compared to our summer median value of 109.0 nmol/l (range 51.5-251.0 nmol/l) and winter median 58.3 nmol/l (range 23.5 – 226.0 nmol/l), which might indicate that some black rhinoceroses are not capable of producing enough endogenous vitamin D. However, these data should be interpreted with care like the elephants, since the data includes D2 and D3 and like elephants, it is currently unknown if they are capable of producing endogenous cholecalciferol. However, the study performed by Clauss et al. (2002)[20] analysed 25(OH)D3 in wild black rhinoceroses, in which it is highly unlikely they receive oral cholecalciferol. This makes it very plausible that black rhinoceroses are capable of endogenous cholecalciferol production. Horses, which are also a member of the Perissodactyla, like rhinoceroses and tapirs, are depending on vitamin D2 in their diet and not able to endogenously produce vitamin D3 [12,21,22]. However, they also show seasonal variation in 25(OH)D levels which might be caused by the plants they consume, due to increasing levels of D2 due to UVb radiation of the roughage [12]. Additionally, the black rhinoceroses received a different amount of cholecalciferol during the course of the study, due to change of pellet and amount of pellets they received. This could be the reason why the levels of 25(OH)D in these animals are higher in 2023 compared to the previous years (Figure 1). This finding, makes it also very plausible that black rhinoceroses are capable of absorbing cholecalciferol in their digestive tract. Which is supported by Olds et al. (2018)[19], which demonstrated a correlation between levels of cholecalciferol in the diet and plasma 25(OH)D levels in captive black rhinoceroses.
To the author’s knowledge, no information is available on wild Indian rhinoceroses regarding serum vitamin D values. There is one study from North America in captive Indian rhinoceroses which shows a significant difference in seasons regarding 25(OH)D levels [23]. Serum 25(OH)D in Bapodra et al. (2014)[23] ranged from 22.4 (± 2.93) in winter to 32.8 (± 7.44) nmol/l in summer, which is much lower compared to our findings of 106.3-132.8 nmol/l. An explanation for the high levels of 25(OH)D in the present study could be that the animals in the study of Bapodra et al. (2014)[23] did not receive any cholecalciferol in their diet, which could indicate that Indian rhinoceroses are capable of absorption of cholecalciferol in their diet. The significant higher levels of 25(OH)D in summer found by Bapodra et al. (2014)[23] might indicate Indian rhinoceroses are capable of endogenously produce of vitamin D, however, it could also be caused by increasing levels of vitamin D2 in their diet as mentioned above.
If both Asian elephant and rhinoceros species show higher 25(OH)D levels compared to the African species, this could suggest that the Malayan tapirs should also have high values in summer or overall of serum 25(OH)D. However, they show very low vit D levels overall. Perhaps this is due to the fact they are mainly nocturnal and they are housed inside (out of fear for sunburn) or hide in the shade on very sunny days [24]. Some other nocturnal species like the Egyptian fruit bat show also undetectable levels of 25(OH)D in both captive and wild specimens [25]. However, when exposed actively to sunlight they are able to increase their vitamin D levels, indicating that they are able of cholecalciferol production in their skin (Southworth et al., 2013)[26]. Or perhaps their vitamin D physiology resembles that of horses, where the main vitamin D source is vitamin D2 and not cholecalciferol production in the skin [21]. This might also be the case for African elephants, which do have very low levels circulating serum levels of 25(OH)D.
Comparing the very low serum 25(OH)D levels of the African elephants and Malayan tapirs with the intake in cholecalciferol (Table 1), might indicate that they either do not resorb cholecalciferol from the digestive tract or cholecalciferol is not their main metabolite in vitamin D metabolism. This would support the hypothesis that they have a similar vitamin D metabolism to horses. Another option could be that the amount of cholecalciferol given was too low for these species, at least in all African elephants, and one of the Malayan tapirs, which were given lower amounts than current recommendations (Table 1). Also, Asian elephants in the study here received a much lower amount of cholecalciferol than recommended. However, it should be noted that this interpretation might not be correct (lower than the recommendations), since it is unknown what amount of ergosterol the animals received in their diet and current recommendations are based on vitamin D and not cholecalciferol solely. As described above, levels of 25(OH)D in this study were much higher than in previous studies and in the captive population in Asia.
Future research should be done to gain further insight into the normal vitamin D metabolism, for instance in vivo and in vitro testing of UVb radiation would be highly recommended by the author to investigate the capacity of producing endogenous vitamin D, especially since UV index in range countries is much higher compared to places where most animals live in captivity (Europe and North America). However, it is known that (Malayan) tapirs, are very sensible for sunburn, so in vivo testing for these animals needs to be conducted with great care. There are virtually no referential values of serum 25(OH)D preferably with discrimination of 25(OH)D2 and 25(OH)D3 in these species in the wild, with the exception of the black rhinoceroses, especially presence of circulating 25(OH)D3 levels in wild animals might give an idea that these animals are capable of endogenous vitamin D production. Additionally, differentiation between pregnant animals, lactating animals, age and sex, time spent outside and the amount of vitamin D (both cholecalciferol and ergosterol) in their diet to investigate the effect of those variables on serum 25(OH)D should be investigated.
5. Conclusions
African elephants have significantly lower levels of serum 25(OH)D compared to their Asian counterparts when held in zoological institutions in the Northern Hemisphere. Circulating serum 25(OH)D levels in Asian elephants were found to be higher compared to previous studies despite low levels of cholecalciferol in their diet. Both elephant species have a low circulating level of serum 25(OH)D compared to the rhinoceroses in this study and do not show a seasonal variation. This might indicate they are uncapable of producing sufficient endogenous vitamin D whole year round or part of the year, or they receive insufficient vitamin D in their diet.
Both rhinoceros species showed much higher levels of 25(OH)D compared to all other pachyderms which makes it more likely that they are able to endogenously produce cholecalciferol. The Indian rhinoceros had high circulating levels year round, while the black rhinoceroses show a significant lower 25(OH)D level in the winter. This indicates that Indian rhinoceroses are capable of endogenously producing enough vitamin D year round, while black rhinoceros are not.
Malayan tapirs have very low circulating levels of serum 25(OH)D, which might be due to inability of endogenously produce of cholecalciferol or insufficient amount of vitamin D in offered in their diet, or might be normal for this species.
Author Contributions
Conceptualization, L.G.R.B. and R.J.C.; Data curation, L.G.R.B.; Formal analysis, L.G.R.B. and R.J.C.; Funding acquisition, L.G.R.B. and R.J.C.; Methodology, L.G.R.B. and R.J.C.; Project administration, L.G.R.B. and R.J.C.; Resources, L.G.R.B. and R.J.C.; Supervision, R.J.C.; Visualization, L.G.R.B.; Writing—original draft, L.G.R.B.; Writing—review and editing, L.G.R.B. and R.J.C.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Utrecht University Faculty of Veterinary Medicine and Rotterdam zoo.
Institutional Review Board Statement
This study was conducted in accordance with the Dutch law and required no ethical approval as surplus material has been used.
Data Availability Statement
The data presented in this study are available in the article.
Acknowledgments
We acknowledge the keepers, curators, vets and staff of the Rotterdam Zoo, Hilvarenbeek Zoo and Wüppertal Zoo for collecting/donating the samples as well as Christine Krüger-Velema, Carola Feijt and Rens Lindeboom for the analyses of the samples. The authors would like to thank Professor Wouter Hendriks for his comments and input.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Roach, J.O.; Benyon, S. Nutrition. In Metabolism and nutrition, 2nd ed.; Roach, J.O., Ed.; Elsevier Ltd.: London, NY, USA, 2003; pp. 154–156. [Google Scholar]
- Thitaram, C.; Pongsopawijit, P.; Thongtip, N.; Angkavanich, T.; Chansittivej, S.; Wongkalasin, W. Dystocia following prolonged retention of a dead fetus in an Asian elephant (Elephas maximus). Theriogenology 2006, 66, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Mahasawangkul, S.; Angkawanich, T. Elephant health status in Thailand—the role of elephant hospitals and mobile elephant clinics. Eu-Asia link project symposium. In Managing the health and reproduction of elephant populations in Asia; Elephant Hospital, National Elephant Institute, Forest Industry Organization: Lampang, Thailand, 2007; pp. 32–37. [Google Scholar]
- Sanyathitisaeree, P.; Yartbantoong, N.; Thongthipsiridej, S.; Theeraphan, W. Elephant health problems: an accumulative case report from the Kasetsart university veterinary teaching hospital– Kamphaengsaen. Eu-Asia link project symposium. In Managing the Health and Reproduction of Elephant Populations in Asia; Faculty of Veterinary Medicine, Kasetsart University: Bangkok, Thailand, 2007; pp. 41–48. [Google Scholar]
- Hermes, R.; Saragusty, J.; Schaftenaar, W.; Göritz, F.; Schmitt, D.L.; Hildebrandt, T.B. Obstetrics in elephants. Theriogenology 2008, 70, 131–144. [Google Scholar] [CrossRef] [PubMed]
- van der Kolk, J.H.; van Leeuwen, J.P.T.M.; van den Belt, A.J.M.; van Schaik, R.H.N.; Schaftenaar, W. Subclinical hypocalcaemia in captive Asian elephants (Elephas maximus). Veterinary Record 2008, 162, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Van Sonsbeek, G.R.; van der Kolk, J.H.; van Leeuwen, J.P.T.M.; Everts, H.; Marais, J.; Schaftenaar, W. Effect of calcium and cholecalciferol supplementation on several parameters of calcium status in plasma and urine of captive Asian (Elephas maximus) and African elephants (Loxodonta africana). Journal of Zoo and Wildlife Medicine 2013, 44, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Ensley, P.T.; Anderson, M.; Osborn, K.; Bissonnette, S.; Deftos, L. Osteodystrophy in an orphan Asian elephant. In Proceedings of the AAZV, Pittsburgh, Pennsylvania; 1994; pp. 142–143. [Google Scholar]
- Sach, F.; Dierenfeld, E.S.; Langley-Evans, S.C.; Watts, M.J.; Yon, L. African savanna elephants (Loxodonta africana) as an example of a herbivore making movement choices based on nutritional needs. PeerJ 2019, 7, e6260. [Google Scholar] [CrossRef] [PubMed]
- Childs-Sanford, S.E.; Makowski, A.J.; Wakshlag, J.J. The vitamin D status of Asian elephants (Elephas maximus) managed in a northern temperate climate. Journal of Zoo and Wildlife Medicine 2020, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Chen, T.C.; Holick, M.F.; Mikota, S.; Dierenfeld, E. Serum concentrations of calcium, phosphorus, and 25-hydroxyvitamin D in captive African elephants (Loxodonta africana). Journal of Zoo and Wildlife Medicine 2009, 40, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Dosi, M.C.M.C.; McGorum, B.C.; Kirton, R.D.; Cillán-García, E.; Mellanby, R.J.; Keen, J.A.; Hurst, E.A.; Morgan, R.A. The effect of season, management and endocrinopathies on vitamin D status in horses. Equine Veterinary Journal 2022, 55, 672–680. [Google Scholar] [CrossRef] [PubMed]
- BioMérieux. Vidas® 25 OH vitamin D total (vitD). 2016, pp. 1–9.
- KNMI/ESA UV index. Available online: https://www.esa.int/Applications/Observing_the_Earth/Thanks_to_ESA_KNMI_offers_a_UV_forecasting_Service (accessed on 22 November 2023).
- Van Sonsbeek, G.R.; van der Kolk, J.H.; van Leeuwen, J.P.T.M.; Schaftenaar, W. Preliminary validation of assays to measure parameters of calcium metabolism in captive Asian and African elephants in Western Europe. Journal of Veterinary Diagnostic Investigation 2011, 23, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Childs-Sanford, S.E.; Makowski, A.J.; Hilliard, R.L.; Wakshlag, J.J. Experimental cholecalciferol supplementation in a herd of managed Asian elephants (Elephas maximus). Journal of Zoo and Wildlife Medicine 2023, 54, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Childs-Sanford, S.E.; Kiso, W.K.; Schmitt, D.L. Serum vitamin D and selected biomarkers of calcium homeostasis in Asian elephants (Elephas maximus) managed at a low latitude. Journal of Zoo and Wildlife Medicine 2024, 55, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Perumbilly, A.J.; Balagangatharathilagar, M.; Kalaivanan, M.; Vishnurahav, R.B.; Begum, M.M. Quantitaive determination of 25-hydroxyvitamin D in adult captive Asian elephants (Elephas maximus) from Southern tropical regions of Tamil Nadu. International Journal of advanced biochemistry research 2024. [Google Scholar] [CrossRef]
- Olds, J.; Oltman, W.; Makowski, M.; Householder, H.; Keeley, L.L. Seasonal variation of serum 25-hydroxy-vitamin D in two captive Eastern black rhinoceros (Diceros bicornis michaeli) housed in a North American zoo. Journal of Zoo and Wildlife Medicine 2018, 49, 943–951. [Google Scholar] [PubMed]
- Clauss, M.; Jessup, D.A.; Norkus, E.B.; Chen, T.C.; Holick, M.F.; Streich, J.; Dierenfeld, E.S. Fat soluble vitamins in blood and tissues of free-ranging and captive rhinoceros. Journal of Wildlife Diseases 2002, 38, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Hurst, E.A.; Homer, N.Z.; Mellanby, J. Vitamin D metabolism and profiling in veterinary species. Metabolites 2020, 10, 371. [Google Scholar] [CrossRef] [PubMed]
- Azarpeykan, S.; Gee, E.K.; Thompson, K.G.; Dittmer, K.E. Undetectable vitamin D3 in equine skin irradiated with ultraviolet light. Journal of Equine Science 2022, 33, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Bapodra, P.; Dierenfeld, E.; Wolfe, B.A. Evaluation of season-related dietary changes on the serum profiles of fat soluble vitamins, mineral, fatty acids, and lipids in the captive greater-one horned rhinoceros (Rhinoceros unicornis). Zoo Biology 2014, 33, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Bruins-van Sonsbeek, L.G.R. Personal observations, 2024.
- Cavaleros, M.; Buffenstein, R.; Ross, F.P.; Pettifor, J.M. Vitamin D metabolism in frugivorous nocturnal mammal, the Egyptian fruit bat (Rousettus aegyptiacus). General and Comparative Endocrinology 2003, 133, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Southworth, L.O.; Holick, M.F.; Chen, T.C.; Kunz, T.H. Effects of sunlight on behaviour and 25-hydroxyvitamin D levels in two species of Old World fruit bats. Dermato-Endocrinology 2013, 5, 192–198. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Distribution in the natural range on a map with UV index and temperature. The African elephant in yellow, black rhinoceros in light green, Asian elephant in orange, Indian rhinoceros in blue and Malayan tapir in purple. Range countries all species current know how of the different IUCN specialist groups (Accessed multiple times during September 2023-January 2024). ∙ is indicating the current position of the location of the animals in the study. (This image was based on Martini,2014; KMNI/ESA, 2023).
Figure 1.
Distribution in the natural range on a map with UV index and temperature. The African elephant in yellow, black rhinoceros in light green, Asian elephant in orange, Indian rhinoceros in blue and Malayan tapir in purple. Range countries all species current know how of the different IUCN specialist groups (Accessed multiple times during September 2023-January 2024). ∙ is indicating the current position of the location of the animals in the study. (This image was based on Martini,2014; KMNI/ESA, 2023).
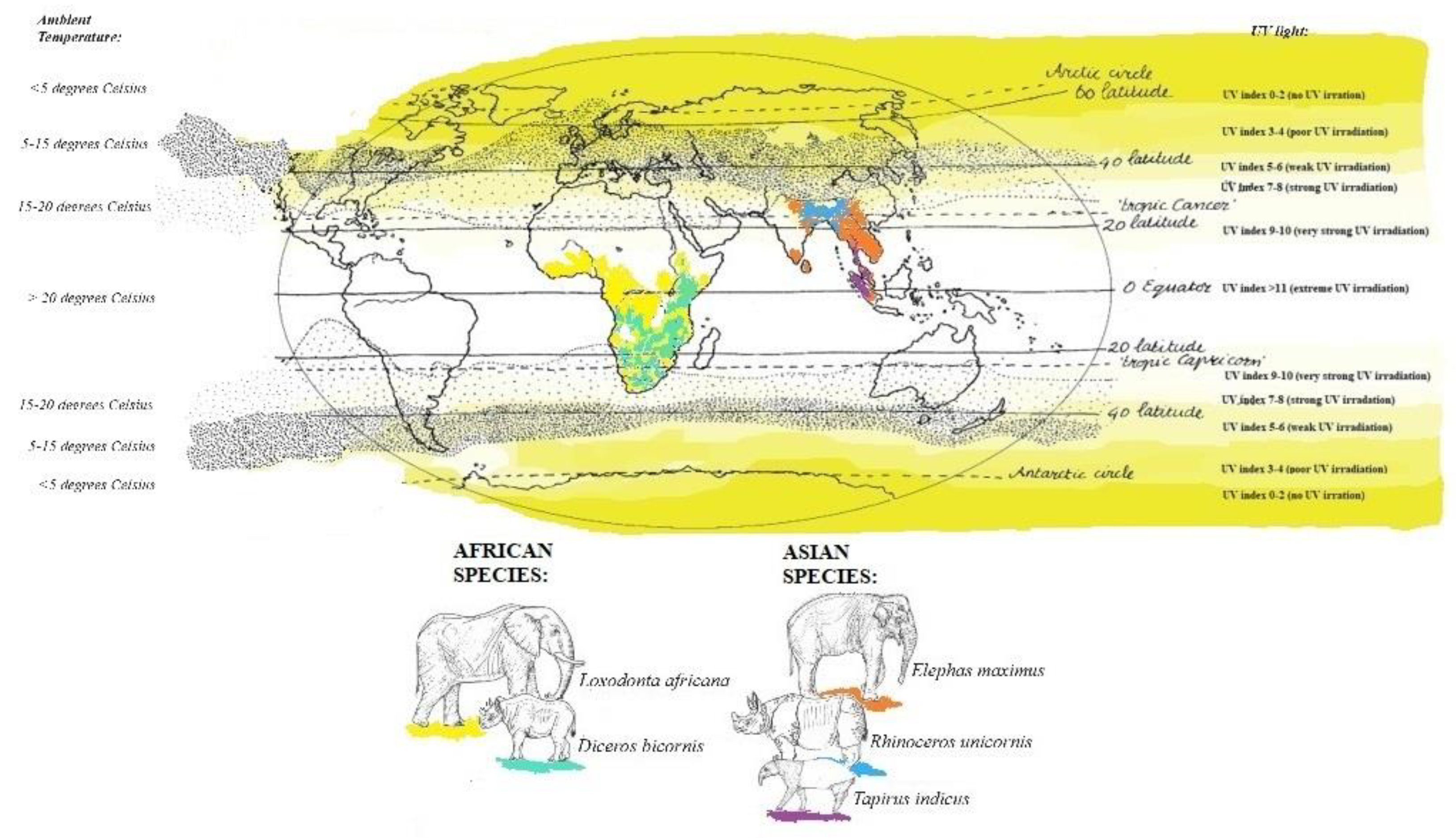
Figure 2.
Serum 25(OH)D concentrations in nmol/l from individual African elephant (A), Asian elephant (B) and black rhinoceros (C) during 2020-2023. The numbers of the individuals in the legenda correspond with the individuals in Table 1.
Figure 2.
Serum 25(OH)D concentrations in nmol/l from individual African elephant (A), Asian elephant (B) and black rhinoceros (C) during 2020-2023. The numbers of the individuals in the legenda correspond with the individuals in Table 1.
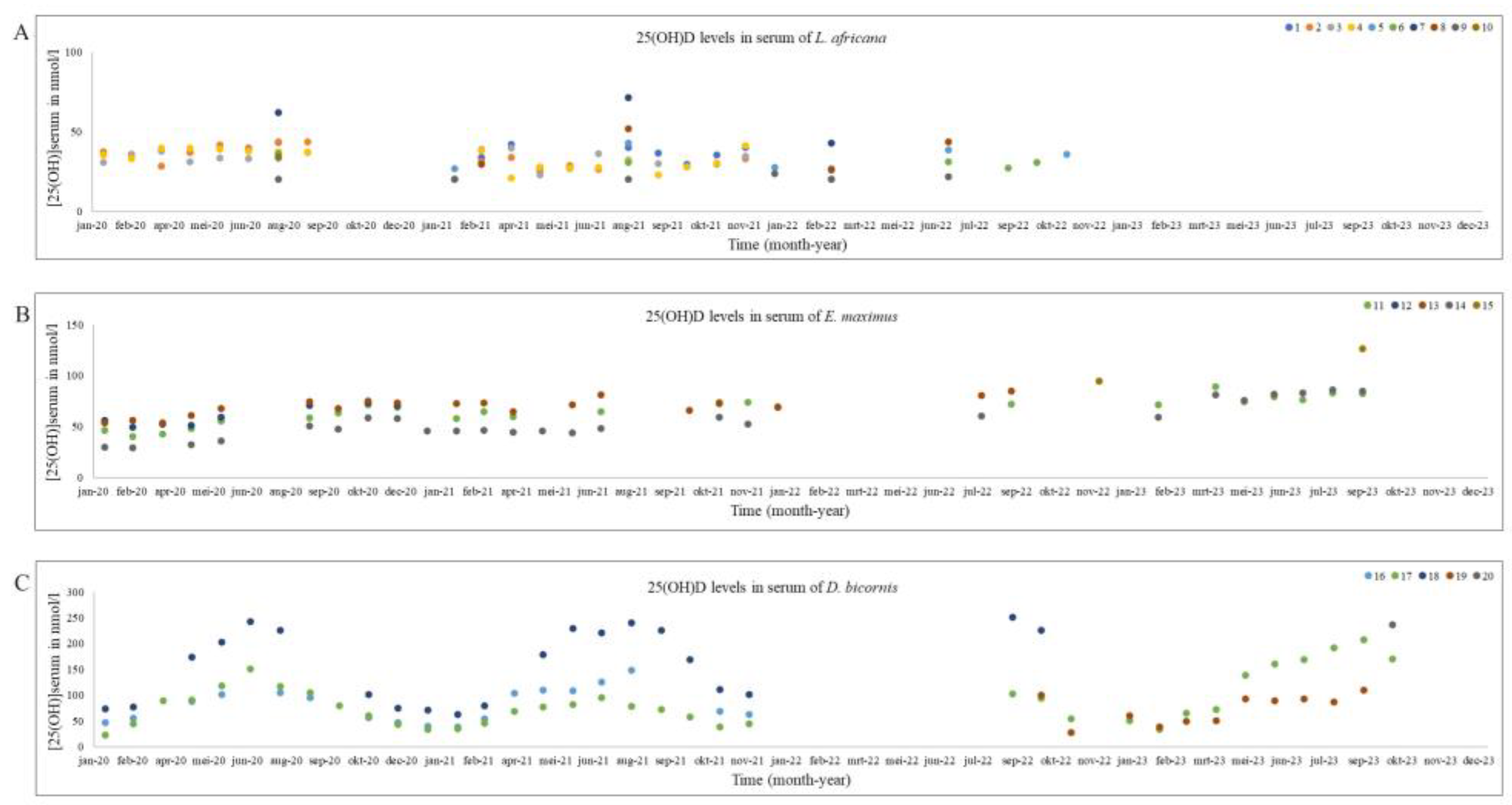
Figure 3.
Distribution of serum 25(OH)D concentrations (nmol/L) in African elephants (A), Asian elephants (B) and black rhinoceros (C) per season. The bars indicate the siginificant difference between Summer and Winter values. Suv is summer according to UV index (green), sc is summer according to calendar months (orange), wuv is winter according to UV index (purple) and wc is winter according to calendar months (pink); vitD is 25(OH)D.
Figure 3.
Distribution of serum 25(OH)D concentrations (nmol/L) in African elephants (A), Asian elephants (B) and black rhinoceros (C) per season. The bars indicate the siginificant difference between Summer and Winter values. Suv is summer according to UV index (green), sc is summer according to calendar months (orange), wuv is winter according to UV index (purple) and wc is winter according to calendar months (pink); vitD is 25(OH)D.
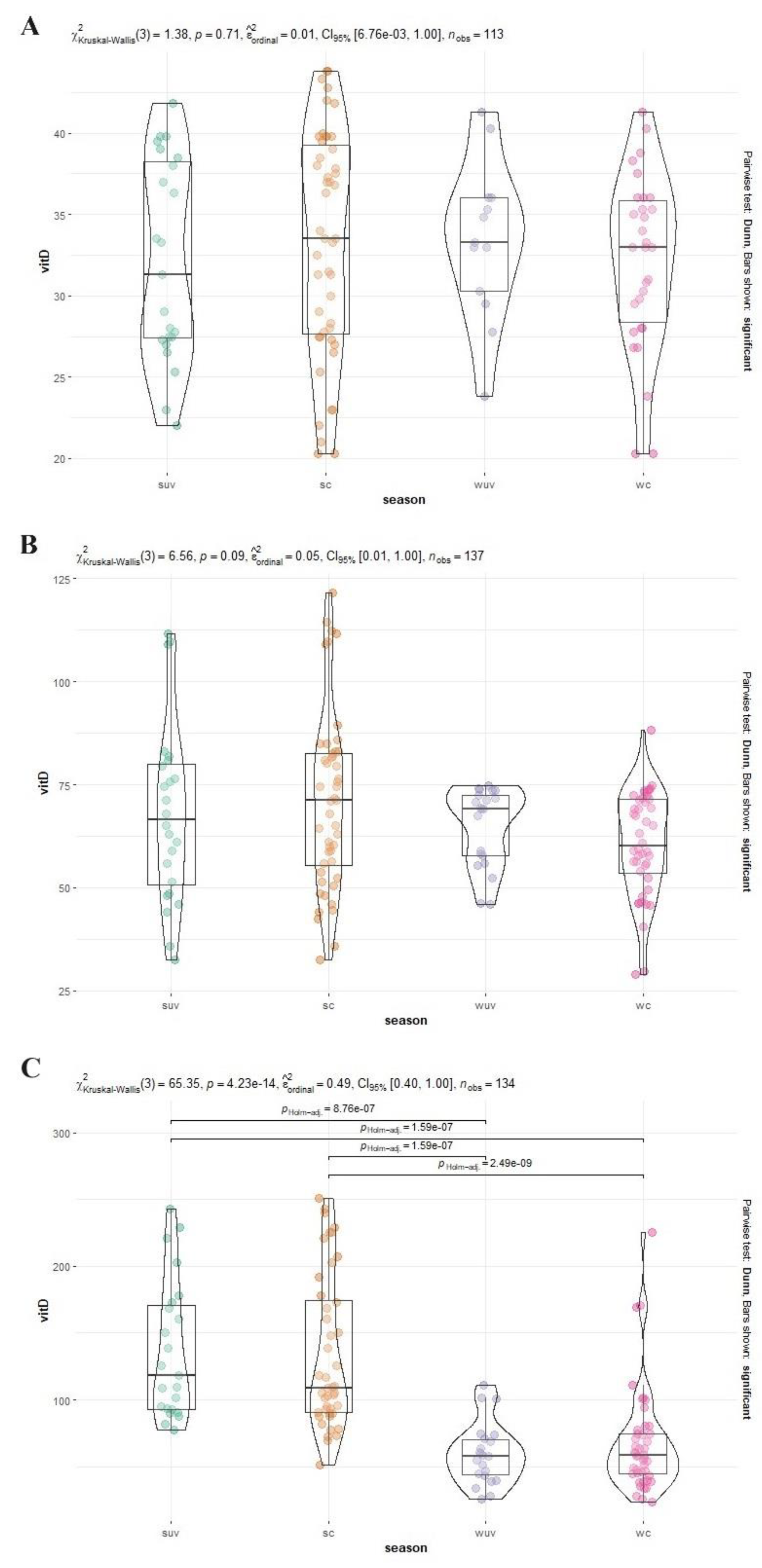

Table 1.
Participating institutions (Zoo), animal data and calculated cholecalciferol intake of the animals through provided pellets.
Table 1.
Participating institutions (Zoo), animal data and calculated cholecalciferol intake of the animals through provided pellets.
| Zoo* | Individual | Species | Sex | Year of birth (years) |
n | Cholecalciferol intake (IU/kg BW)** |
Recommended vitamin D intake (IU/kg BW) |
| A | 1 | L. Africana | f | 2015 | 9 | 6.5 | 8–121 |
| 2 | L. Africana | f | 1985 | 18 | 6.5 | ||
| 3 | L. Africana | f | 1986 | 18 | 6.5 | ||
| 4 | L. Africana | f | 1992 | 19 | 6.5 | ||
| B | 5 | L. Africana | f | 1993 | 6 | 3.6 | |
| 6 | L. Africana | f | 2007 | 3 | 3.6 | ||
| 7 | L. Africana | f | 2016 | 5 | 3.6 | ||
| 8 | L. Africana | m | 2019 | 6 | 3.6 | ||
| 9 | L. Africana | m | 1993 | 5 | 3.6 | ||
| 10 | L. Africana | f | 1992 | 1 | 3.6 | ||
| C | 11 | E. maximus | f | 1970 | 25 | 5 | 12–151 |
| 12 | E. maximus | f | 2000 | 22 | 5 | ||
| 13 | E. maximus | f | 2003 | 20 | 5 | ||
| 14 | E. maximus | f | 2010 | 25 | 5 | ||
| 15 | E. maximus | m | 2021 | 2 | 5 | ||
| 16 | D. bicornis | m | 2001 | 19 | 2.5-6.6*** | 3-92 | |
| 17 | D. bicornis | f | 2011 | 37 | 2.5-6.6*** | ||
| 18 | D. bicornis | f | 2017 | 22 | 2.5-6.6*** | ||
| 19 | D. bicornis | m | 2020 | 11 | 2.5-6.6*** | ||
| 20 | D. bicornis | m | 2019 | 1 | 2.5-6.6*** | ||
| 21 | R. unicornis | f | 2017 | 4 | 1.5 | - | |
| 22 | T. indicus | f | 2011 | 32 | 10-30**** | 10.53 | |
| 23 | T. indicus | m | 2016 | 8 | 5 |
* Corresponding Zoos: A) Hilvarenbeek Zoo; B) Wuppertal Zoo; C) Rotterdam Zoo. ** Cholecalciferol was present in concentrates (pellet or bricks), and intake was estimated based on the amount on the package (in international units (IU)) and the amount of pellet/bricks given and the bodyweight (BW) of the animals. *** In the beginning of 2022, the type and amount of pellet had changed, 6.6 IU/kg BW is the maximum they received when eating all pellet offered per day, which was not always the case. **** In June 2021 the diet has changed in order to reduce the BCS of the female, the concentrates changed from horse to rhinoceros pellets and amount of pellets were reduced with 50%, therefore, she has started with an amount of 30 IU/kg BW to 10 IU/kg BW in 2023. Recommendations: 1Ullrey et al. 1997; 2advised in Pilgrim and Biddle, 2020 – based on Dierenfeld, 1996; 3advised in AZA Tapir TAG, 2013 – based on Padilla and Dowler, 1994), to the author’s knowledge there are no current recommendations of cholecalciferol in Indian rhinoceroses.
Table 2.
Serum 25(OH)D concentration (nmol/l) in captive pachyderms in Western Europe.
| Species*** | Mean (±SD) |
Median (range) | Median per season (range) | ||||
| Summer | Winter | ||||||
| Calendar (Apr-Sept) |
UV index >6 (May-July) |
Calendar (Oct-Mar) |
UV index <2 (Nov-Jan) |
||||
|
L. africana* n = 90, 10+ |
34.5 (± 9.0) |
33.2 (21.2-58.8) |
33.5 n = 47, 6 (20.3-43.8) |
31.3 n = 23, 6 (22.0-41.8) |
33.0 n = 30, 6 (20.3-41.3) |
33.3 n = 13, 6 (23.8-41.3) |
|
|
E. maximus n = 94, 5 |
75.3 (± 21.1) |
68.7 (55.5-110.6) |
71.2 n = 48, 4 (32.5-122.0) |
66.5 n = 24, 4 (32.5-112.0) |
60.2 n = 44, 4 (29.0-88.3) |
69.0 n = 21, 4 (46.0-74.8) |
|
|
D. bicornis n = 90, 5 |
126.2 (± 70.2) |
86.3 (72.9-237.0) |
109.0† n = 44, 4 (51.5-251.0) |
118.0† n = 23, 4 (77.5-243.0) |
58.3 n = 45, 4 (23.5-226.0) |
57.9 n = 22, 4 (26.0-111.0) |
|
|
R. unicornis n = 4, 1 |
- | 107.9 (106.3-132.8) |
109.5 (105.3-132.8) |
121.2 (109.5-132.8) |
106.3 |
106.3 | |
|
T. indicus** n = 26, 2 |
- | < 20.3 (< 20.3-33.5) |
< 20.3 (< 20.3-< 20.3) |
< 20.3 (< 20.3-< 20.3) |
20.3 (< 20.3-33.5) |
20.3 (< 20.3-33.5) |
|
+ First value is the number of values, second values the number of individuals. * Four values were below 20.3 nmol/l. ** Many values were below 20.3 nmol/l, for the statistics 20.3 nmol/l was used as the actual value, but this is most likely inaccurate. *** Mean per individual was used to calculate the total mean and median; in Table 1 the number of samples provided per individual are reported. For the seasons only the individuals with a complete dataset were used. † Value is statistically different compared to the corresponding Winter value.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



