Submitted:
23 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
We conclude that Hbr not only favored growth, but also colonization with R. intraradices and consequently biomass production, and N and P content in plant tissue. The growth response and nutrient exchange suggest interdependence between brassinosteroids and the endomycorrhizal fungus.

Keywords:
Arbuscular mycorrhizae
; Homobrassinolide
; Yield components.
1. Introduction
Cattle production systems in southern Mexico are mainly based on the grazing of introduced grasses of the genera Cynodon, Panicum, Digitaria and lately the genus Brachiaria, and within this genus, B. decumbens Stapf is one of the preferred grasses in this region. In general, grasslands are maintained with the minimum of inputs and over time they degrade due to the effects of seasonal distribution of precipitation, high temperatures and soil acidity. In addition, the absence of sustainable practices generates seasonal biomass production, with changes in the allocation of dry matter in the yield components and an increase in undesirable plants [1].
These abiotic factors, in combination with poor grassland management, become a threat to the sustainability of the system. In this regard, there is currently an increased interest in some mechanisms of action of various rhizosphere microorganisms in the sustainable management of grasslands, such as endomycorrhizal fungi [2] that establish symbiosis with the root system of plants in almost all ecosystems [3], favor the transport of nutrients and water [4] and have been shown to increase crop yields through nutritional effects and the regulation of plant hormones [5].
The symbiosis presents preference between some species and different endomycorrhizal fungi [6], however, B. decumbens presents high percentage of radical colonization and biomass in-growth in different environments [7,8].
In the case of brassinosteroids, which are essential steroid hormones for plant growth, they exert regulation of various processes in crop growth and yield, such as cell expansion and division, vascular differentiation [9,10], pollen and stamen development [10], and homeostasis and signaling [11]. In addition, they have been shown to positively affect symbiotic interaction with changes in plant hormone levels when symbiosis is established [12], as in solanaceae [13]. They also interact with plants in the presence of biotic and abiotic stresses [9,11].
Therefore, in order to improve the production and quality of biomass in Brachiaria de-cumbens Stapf, it was biofertilized with R. intraradices and/or A. brasilense in interaction with a homobrassinolide in Chiapas Mexico.
2. Materials and Methods
2.1. Study Area Location
The present investigation was carried out from March to July 2023 under nursery conditions in the Pichucalco Experimental Station, (Latitude 17° 30” N, Longitude 93° 07” W altitude and 36 m above sea level) of the National Institute of Forestry, Agricultural and Livestock Research (INIFAP) in Pichucalco, Chiapas Mexico.
2.2. Edaphoclimatic Conditions of the Study Area
The climate of the region is type Af, [14] warm humid with rain all year round, average annual precipitation of 3996 mm and average temperature of 25 °C.
The soil belongs to the Acrisol group, the substrate was made with the soil plus 50% washed river sand and it was solarized for 72 h with the following physical-chemical characteristics: Sandy crumb texture, sand 62.84%, silt 19.36% and clay 17.80% (Bayoucus), 3.72 % organic matter (Walkley-Black), pH 5.58 (1:2 H2O), Apparent density 1.47 g.ml-1, 0.16 % N total (%) (Kjelddhall), P 57.7 mg.kg-1 (colorimetry), 84 K+ int. (mg/kg−1), (atomic spectrophotometry), 0.3 mg/kg de Na++, 284.0 mg/kg de Ca++, 1.5 Meq.100g-1 cation exchange capacity (CEC) and 0.05 ds.m-1 electrical conductivity (conductometer).
Plastic bags (25x35 cm) with a capacity of 5.0 kg were filled with the substrate, previously perforated at the bottom to promote drainage. The bags were placed on wooden structures to avoid contact with the ground.
2.3. Biofertilizers and Homobrassinolide
The endomycorrhizal fungus Rhizophagus intraradices (Schenk et Sm) Walker et Schuessler, was reproduced in sterile soil in the root system of Sorghum bicolor L. At the time of packaging there were 40 spores per gram of soil plus propagules, and the level of colonization in the root system was 95% (Micorriza INIFAP® Rosario Izapa, Chiapas, México. Data indicated on the product).
Azospirillum brasilense Tarrand, Krieg et Döbereiner, was produced by the company Bio-fabrica Siglo XXI in Xochitepec, Morelos, México, under the trade name AzoFer Plus, having a concentration of 500 × 106 bacteria∙g−1 (Data indicated on the product).
Brassinosteroid CIDEF-4 (HBr) is a product of Natura del. Desierto, SA de CV in Mexico with 80% steroidal content, and 10% active ingredient. Its soluble presentation is not toxic and it is compatible for its application with agrochemicals.
2.4. Experiment Setup and Application of Microorganisms and Homobrassinolide
The seeds of Brachiaria decumbens Stapf were sown in each treatment and the microorganisms were adhered to 4% of the weight of the seed with carboxymethyl cellulose. The inoculated seeds were sown 3 cm deep.
Brassinosteroid CIDEF-4 (HBr) was used at 2 mgL-1, and the first foliar application of homobrassinolide was carried out 12 days after sowing (das), the time in which the emission of the first true leaves occurred in all treatments, subsequently the application foliar treatment of homobrassinolide was carried out every 28 days.
The treatments were: 1) Control, 2) Rhizophagus intraradices, 3) Azospirillum brasilense, 4) R. intraradices + A. brasilense, 5) Homobrassinolide CIDEF-4 (Hbr), 6) Hbr + R. intraradices, 7) Hbr + A. brasilense, 8) Hbr + R. intraradices + A. brasilense. In each treatment there were five repetitions, and they were distributed in a completely randomized design. The experimental unit was one container with one plant. Plants were irrigated with water drawn from a deep well.
2.5. Variables Evaluated
Destructive sampling of the plants was carried out at 28, 56, 84 and 112 (das). In addition, morphological variables (plant height, number of leaves, diameter of stem and number of stems) and physiological variables (dry weight of aerial and root components and leaf area) were recorded. The percentage of mycorrhizal colonization was determined in the root and the N and P content in the plant tissue.
The physiological yield components of aerial and root were weighed on a semi-analytical balance (Ohaus Adventurer Pro, USA) after drying in a forced air oven at 60-75°C to constant weight. Leaf area (cm2) was obtained using a leaf area integrator (LI-COR, LI 3000ª, USA).
The percentage of colonization was quantified at 28 das using the technique [15]. One hundred root segments of 1.5-1.6 cm in length were observed with an optical microscope with oil immersion objective (100 X). With the previous variables, the Relative Growth Rate (RGR) [16] was determined.
N and P content was obtained by foliar analysis. N was determined by microkjeldahl and phosphorus content with an Olsen/spectrophotometer (Thermo Fisher Scientific Model 400 ¼) at the soil and water laboratory of the Agricultural Sciences Faculty of the UNACH in Huehuetan, Chiapas, Mexico.
2.6. Statistical Analysis
A completely randomized design was used, performing an ANOVA analysis of variance, using the SAS System for Windows Ver. 8.1 (1999-2000) [17], When the ANOVA was significant, the parameters were compared by Tukey test (p ≤ 0.05). The data, expressed in percentages, were transformed by the formula arc.sin, before the ANOVA and the data were plotted using Sigma Plot version 11.0.
3. Results
3.1. Morphological and Physiological Yield Components
The average height of plants in interaction with the microorganisms and the homobrassinolide, alone or combined, increased 53.9 % more, compared to the control at 28 das and at the end of the evaluation the increase represented 16.6 % (p ≤ 0.05).
The maximum average height reached with the treatments where Hbr was applied was 8.8 % compared to the biofertilized treatments alone. On the other hand, the biofertilized treatments were 11.2 % taller than the control. The most contrasting effect was between the treatments with Hbr and the control with a difference of 20.9 cm in height (Table 1).
In contrast, the ratio was 24 % when measuring the height of plants with Hbr compared to biofertilized plants alone.
The most contrasting difference in leaf number was at 56 and 84 das and was between the control treatments and the application of Hbr alone or in combination with the microorganisms. This represented an increase of 90.9 % (56 das) and 112 % (84 das). The average of the four samples in the treatments biofertilized with R intraradices and/or A. brasilense was 35 % higher than the control. When applying Hbr in combination with the microorganisms, the number of leaves increased in different percentages compared to the same treatments without Hbr. The initial value at 28 das, the difference when applying Hbr was 8 %, at 56 das 40.7 %, and at 84 and 112 das it was 16 %. In all samplings there were statistical differences (p ≤ 0.05), the first statistical group was formed by the treatments where Hbr was applied alone and when biofertilization was also applied with R. intraradices.
The number of stems per plant showed significant statistical differences (p<0.05) in favor of the treatments applied only with Hbr and in combination with biofertilization of R. intraradices, as well as biofertilization alone of R. intraradices from the first sampling. The most notable difference between treatments occurred at 84 and 112 das. At 84 das, the number of stems increased 41% in the treatments with the biofertilization of the microorganisms plus Hbr compared to the same treatments with only the microorganisms. At 112 das, the Hbr treatment increased the number of stems by 92% in relation to the control, 65% in relation to the treatments with microorganisms and 29% with microorganisms plus Hbr. Stem thickness is more consistent when combining Hbr and R. intraradices from the beginning of the evaluation. The greatest increase occurred at 28, 84 and 112 das with the Hbr plus R. intraradices treatment and was statistically superior (p ≤ 0.05) to the rest of the treatments. The difference represented 13% more in the thickness of the stem for the treatment R. intraradices plus Hbr compared to the control.
In relation to the physiological components of yield (Table 2), the greatest increase in root biomass after 28 das occurred in the treatment where Hbr was applied and was statistically different from the other treatments (p<0.05). At this time, the lowest value was presented with the control.
On the other hand, at 56 and 84 das, the interaction of the application with Hbr and the microorganisms induced increases in root biomass of 19 and 28 % respectively in comparison with the biofertilized treatments but without Hbr. In the last evaluation at 112 das, the root biomass of the biofertilized treatments alone increased 43 % more in comparison with the same treatments plus Hbr.
Leaf biomass presented the greatest increase with biofertilization of R. intraradices plus Hbr at 28, 84 and 112 das, and was statistically different (p<0.05). The treatment with Hbr alone at 56 and 112 das was included in the same statistical group.
Stem dry weight presented the highest values in the first samples at 28 and 56 das with the Hbr alone treatment and statistically different (p<0.05). At 84 das, the individual biofertilization of R. intraradices and A. brasilense was included in the same statistical group. At the end of the study, the highest biomass allocation to stems was found with the Hbr treatment plus the two biofertilizers.
Leaf area presented differential induction in the response to biofertilization of microorganisms, alone or associated through time and in interaction with Hbr. In the first sampling at 28 das, there was a statistical difference and the greatest leaf area, at this time was in the treatments with Hbr and when biofertilized with the endomycorrhizal fungus. On the other hand, at 56 das, the greatest leaf area was found in plants biofertilized with the endomycorrhizal fungus and nitrogen-fixing bacteria separately. In the third sampling, at 84 das, the greatest increase occurred in the treatment with the biofertilization of R. intraradices plus the application of Hbr. At the end of the evaluation, the greatest increase in leaf area and statistically different (p<0.05) was in the treatments where Hbr was applied alone, in interaction with the biofertilization with R. intraradices, and in co-inoculation with A. brasilense (Table 2).
3.2. Biomass Allocation in Shoot and Root
The highest biomass allocation to the root system in relation to the shoot was found in the initial sampling with the symbiosis of the two microorganisms in the plant and when Hbr was included in the same treatment. This same effect was found in the plants only treated with Hbr. In the following samples, the highest value was found in the plants where Hbr was included (Figure 1).
The greatest initial increase in biomass allocation to the aerial part of B. decumbens Stapf occurred with the treatments biofertilized with R. intraradices, alone, in co-inoculation with A. brasilense or when Hbr was included. The application of Hbr alone showed the greatest biomass accumulation in the four samples.
In the aerial part, at 28, 56 and 84 das the highest biomass allocation was presented with the application of Hbr alone and in the treatment with biofertilization of R. intraradices plus Hbr. At 112 days, in addition to the previous treatments, the aerial biomass was higher with the co-inoculation of R. intraradices plus A. brasilense and the application of Hbr.
Relative Growth Rate
Microorganisms alone or in co-inoculation induce differential effect on the relative growth rate of B. decumbens Stapf. There are fluctuations in it over time, and it increases during the first evaluation period when Hbr is included (Figure 2).
In the period from 56 to 84 das, there was a significant increase in the relative growth rate with the biofertilization of A. brasilense, which is similar to the growth achieved when Hbr is included in the biofertilization. On the other hand, in the third period, there was a significant increase in the control, same as the induction of the growth rate of the treatment with the symbiosis of the microorganisms plus Hbr.
In general, after the initial high growth, its effect decreases in the following period or at the end of the evaluation. It should be noted that the most consistent relative growth rate was presented in the treatment with Hbr and the symbiosis of the two microorganisms.
In the case of A. brasilense, the effect is opposite to the growth induced by R. intraradices, which was in this case at the beginning, and decreases notably in the last two months of the evaluation. A. brasilense increases in the second period and decreases in the third period. The co-inoculation of the two microorganisms plus Hbr caused a slight initial decrease compared to the microorganisms alone plus Hbr, but it was higher in the second and third evaluation periods.
Mycorrhizal colonization
Mycorrhizal colonization increased 42 % more with R. intraradices alone and in co-inoculation with A. brasilense compared to the control and A. brasilense (Figure 3).
The application of Hbr alone was 37 % and with R. intrardices 43 %. Hbr in combination with A. brasilense showed colonization of 33 %. The highest value of 39.8 % was with the co-inoculation of the two microorganisms plus Hbr and represented 58 % more in relation to the control.
Content of N and P in plant tissue
The N content in plant tissue of B. decumbens Stapf increased significantly with biofertilizers plus Hbr applications (Figure 4).
4. Discussion
The increase in morphological components of B. decumbens Stapf biofertilized with R. intraradices and/or A. brasilense alone or in co-inoculation, indicates compatibility between the two, even though it is possible to find differences in the induction of host plant growth depending on the microorganisms [18]. The development of biofertilized plants at the beginning of root colonization seems to be related to photosynthate demand of the root system [19]. This same effect was higher in the same treatments with the biofertilizers plus Hbr. applications. The previous higher expression was when combining biofertilization of R. intraradices and exogenous Hbr applications.
The positive effect on plant height when applying Hbr alone or combined with R. intraradices biofertilization compared to A. brasilense has been demonstrated in solanaceae [13], tobacco, tomato [20] and in wheat where synthetic Br foliar was applied and mycorrhization was increased [21]. In addition, in vitro plants of great dwarf banana (Musa sp.) also increased the height of shoots with the application of Hbr (Cidef-4) [22]. In greenhouse, plant height was greater with the application of Hbr Biobrás (6-ABr) alone, in banana FHIA-18 subjected to high temperatures [23] and in field conditions biofertilization alone of R. intraradices induced more height of Brachiaria decumbens [24].
The separate contrasting effects of Hbr and endomycorrhizal fungi in inducing differential response when applied may be due, in the case of endocmycorrhizal fungi, to the stimulation of host plant growth [25], through the transport of nutrients [26] and water [27] and with the application of Hbr, the benefit in the improvement of plants is attributed to its influence on various growth processes [9], in addition to improving their permanence in environments with stress, biotic or abiotic [28].
In the case of R. intraradices, the contrasting effect between the number of leaves of B. decumbens from the first sampling coincides with high mycorrhization (40 %) compared to the control (19 %). This suggests, efficiency of symbiosis with R. intraradices. This fact has been cited in B. decumbens Stapf [29], and in B. brizantha (Hochst. ex A. Rich) cv insurgent with R. intraradices [8]. In other crops biofertilized with R. intraradices, leaf number has also been increased as in Coffea canephora (Pierre) ex Froehrner [30], and Theobroma cacao [31].
The application of brassinosteroid alone in various crops, such as in vitro plants, induces a positive response in the number of leaves, as in banana clone FHIA-18 with the addition of Hbr (Biobras-6) [32], in Lactuca sativa L. with the foliar spraying of Biobras-16 [33]. In contrast, in Stevia rebaudiana Bert the highest number of leaves (p<0.05) was found with the mixture of R. intraradices and Hbr [34]. The above response may be related to the extension of the mycelium of the fungus which allows it to act as an extension of the root absorption surface [26] and favors the persistence of the host plant in adverse environmental conditions [35], such as drought [27] and in general, the concomitant increase in photosynthetic activity and growth [36].
Additionally, these results with positive effects on plant growth also occur under biotic or abiotic stress conditions [9,28] (Gonzalez et al., 2008; Faridud-din et al., 2014).
In potato (Solanum tuberosum L.), vitro plants transplanted in greenhouse, Hbr (BB-6) was applied to the foliage and an increase number of minitubers per plant was observed [37], and banana clone FHIA-18 [38] reported an increase in the diameter of the pseudostem, to apply the Hbr (BB-6) by immersion and foliar spraying and when applying CIDEF 4 (hbr), The shoot height of Musa spp cv great dwarf is increased in vitro by growing in two culture media [22].
In the case of A. brasilense, there is a tendency for morphological variables to increase when it is associated with the endomycorrhizal fungus. It is likely that the contributions of radical exudates of the bacterium, such as indole acetic acid [39], favor communication with soil microorganisms [40,41]. Phytohormones have been considered as signaling compounds for endomycorrhizal fungi that can influence spore germination, hyphal growth and root colonization [42] (Larosse et al., 2002).
The differential response in the growth of the different organs of B. decumbens Stapf, as a consequence of biofertilization with the microorganisms, suggests compatibility between the two.
Physiological components
The increase in dry matter allocation to the different physiological components of yield in B. decumbens Stapf in most samples and plant structures, when Hbr was applied alone, and when combining the same treatment with R. intraradices, suggests interdependence between the two. The above response is expressed in the growth of mycorrhizal plants, through the increase in nutrient content of the host and biomass production [43]. In this regard, [21] when epibrassinolide was applied to wheat and rice, reported an increase in the growth of both mycorrhizal species established under salt stress conditions. Similar results were reported in cotton when exogenous applications of brassinosteroids were applied, with an increase in yield and greater accumulation of biomass [44].
The above response seems to be related to various phytohormones, such as auxins, ethylene, jasmonic acid, brassinosteroid and strigolactones involved in the growth and development of root hairs [12,45] as happens when biofertilizing trifoliate orange under drought stress [46].
In other crops, brassinosteroids are also cited as positively affecting the mycorrhizal symbiosis of tobacco and tomato plants [20] and auxins, brassinosteroids and strigolactones are responsible for the establishment of mycorrhizal associations [47]. In general, microbial communities are attracted by some signaling factors in the form of exudates released by plant roots [48] and lead to improved plant growth [49].
Hormonal homeostasis (in response to fungal associations) leads to signaling and induces various physiological changes to the benefit of the plant [47], such as the transport of nutrients and water through the hypha, which is thinner than plant roots and allows it to penetrate to places where root absorbing hairs normally do not [50].
Root colonization is high from the beginning (40 %) and the benefits of the symbiosis are reflected in this variable. It is likely that the initial establishment of the mycorrhizal symbiosis and its effect on the number of leaves favored the transport of nutrients to the plant [51].
The increase in root biomass of all biofertilized treatments alone or combined seems to be influenced by the amount of root exudates that favor root growth and this condition allows them to explore more soil surface [52]. The lowest root growth was in the control, possibly due to the low levels of native mycorrhization compared to the functioning of the R. intraradices-B. decumbens symbiosis that has been cited as effective in improving their productivity [53].
Higher root biomass allocation in response to Hbr applications is supposed to [54] stimulate the intrinsic potential of plants and consequently promote growth and yield [55]. Brassinosteroids have now been shown to play an important role in yield increase [44]. The increase in vegetative and reproductive development has been reported in different annual crops biofertilized with endomycorhysic fungi and A. brasilense [56].
The effect of R. intraradices in inducing greater biomass in the stem of biofertilized plants from the beginning of the evaluation is due to the nutrient and water supply to the host plant [57], and consequently changes in its structures are induced [58]. In Coffea canephora (Pierre) ex Froehner [30] and Tabebuia donnell-smithii Rose, the same response occurs when biofertilized with R. intraradices [59].
Foliar laminae biomass also increases. The above may be related with the establishment of the symbiosis, where the fungus receives photosynthates and the plant increases its access to mineral nutrition and water [60]. In general, biofertilization with endomycorrhizal fungi significantly increases the allocation of dry matter to the different morphological and physiological yield components, compared to the control.
The importance of the interaction of rhizosphere microbiota with plant root exudates such as indole acetic acid [39], cytokinins and gibberellins [61] has been demonstrated. In addition, it has been shown that the interaction of endomycorrhizal fungi with plants producing specialized metabolites such as flavonoids [57], which establish communication with other soil microorganisms [40,41] and are considered signaling compounds for endomycorrhizal fungi that can influence spore germination, hyphal growth and root colonization [42].
In addition to the above, plant growth is influenced by various environmental factors [62], which can be reflected in differential growth regulated by genetic traits with little variation in phenotypic plasticity through modular growth [63].
In the case of B. decumbens Stapf, its utilization depends on the proportion of leaves, stems and roots that are generated and these components result in forage yield. Knowledge of the influence of environmental seasonality on the growth and forage production of species of interest allows the identification of forage availability and, consequently, the adoption of differential management strategies for each species.
Mycorrhizal colonization
Colonization was observed in all B. decumbens plants, including the control (possible native strain), and was greater in plants biofertilized with R. intraradices compared to plants without biofertilization. The above confirms the presence of endomycorrhizal fungi in the regional substrate; however, they have a lower capacity for colonization and growth induction, even though B. decumbens has been considered to have high mycorrhizal dependence [7,8]. The average percentage of colonization in the control was 16% and with R. intraradices 41%. The above may be related to the supplementation of carbon sources from the host plant to the fungus [64]. The high colonization capacity of R. intraradices has been cited in other crops, annuals and perennials [19,56]. It should be noted that mycorrhizal colonization of plants is influenced, in addition to environmental factors, by variations in the host genotype [65].
In general, the results showed that the increase in physiological variables coincides with the increase in colonization levels.
The phosphorus and nitrogen content increased in all treatments with the microorganisms alone or in co-inoculation, and they statistically surpassed the control (P ≤ 0.05). The preference of endomycorrhizal fungi to transport phosphorus has been cited [66], in addition to raising the phosphorus content, iron and zinc are increased [20] (von Sivers, et al., 2019), and the transport of nitrogen [66].
5. Conclusions
The exogenous applications of Hbr favored the growth of B. decumbens in the morphological and physiological components of the yield and the effect was synergistic when combined with the biofertilization of R. intraradices. In addition to the production of biomass, the content of N and P in the tissue of the plant. The response in growth and nutrient exchange suggests interdependence between brassinosteroids and endomycorrhizal fungi.
Author Contributions
Conceptualization: J.F.A.M. Data acquisition: J.C.I.P. and R.O.C.A. Data analysis: J.F.A.M. and G.R.G. Design of methodology: J.F.A.M. and D.R.A. Writing and editing: J.C.I.P., J.F.A.M., R.P.R. and M.T.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Data Availability Statement
Data are contained within the article.
Acknowledgments
All authors thank the Faculty of Agricultural Sciences of the Autonomous University of Chiapas for the facilities provided in the Laboratory y al Campo Experimental Pichucalco (INIFAP-Pacífico Sur) por las instalaciones para el desarrollo de la Investigación.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ramírez, R.O.; Hernández-Garay, A.; Da Silva, S.C.; Pérez, P.J.; Enríquez, Q.J.F.; Quero, C.A.R.; et al. Acumulación de forraje, cre-cimiento y características estructurales del pasto Mombaza (Panicum maximum Jacq. Jacq.) cosechado a diferentes intervalos de corte. Téc Pecu Méx. 2009, 47(2), pp. 203-213.
- Redecker, D.; Kodner, R.; Graham, L.E. Glomalean fungi from the Ordovician. Science. 2000, 289 (5486):1920-1. [CrossRef] [PubMed]
- Strack, D.; Fester, T.; Hause, B.; Schliemann, W.; Walter, M.H. Arbuscular mycorrhiza: biological, chemical and molecular aspects, J. Chem. Ecol., 2003, 29, pp. 1955-1979. [CrossRef]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Plant drought tolerance enhancement by arbuscular mycorrhizal symbiosis. In Mycorrhizal Fungi, Fulton, S.M. Ed.; Nova Science Publishers lnc, New York, U.S.A. 2011; pp 229–240.
- Dohroo, A. Azospirillum- a potent Biofertilizer in Agriculture. Global Scientific and Academic Research Journal of Multidisciplinary Studies. 2024, 3(2), pp. 8-11. https://gsarpublishers.com/wp-content/uploads/2024/02/GSARJMS192024-Gelary-script.pdf.
- Aguirre-Medina, J.F.; Cadena-Iñiguez, J.; Aguirre-Cadena, J.F. Influence of Endomycorrhizal Fungi on the Growth of Tropical Plant Species. In Mycorrhizal Fungi-Utilization in Agriculture and Industry. R. Radhakrishnan (Ed) IntechOpen. London, England, 2020. [CrossRef]
- Prieto Benavides, O.; Belezaca Pinargote, C.; Mora Silva, W.; Vallejo Zambrano, E.; Gutiérrez Lara1, V.; Pinargote, Mendoza, E. Inoculación de Brachiaria decumbens con hongos formadores de micorriza arbuscular nativos del trópico húmedo ecuatoriano. Ciencia y Tecnología 2011, 4(2), pp. 9-18. file:///C:/Users/franc/Downloads/Dialnet-InoculacionDeBrachiariaDecumbensConHongosFormadore-4149411.pdf.
- Lozano-Contreras, M.G.; Rivas-Pantoja, F.; Castillo-Huchim, J. E. Crecimiento de plántulas de Brachiaria brizantha en respuesta a la aplicación de hongos micorrizógenos y bacterias diazotróficas Pastos y Forrajes, 2013, 36(2), 227-232. http://scielo.sld.cu/scielo.php?script=sci_abstract&pid=S0864-03942013000200007.
- Fariduddin, Q.; Yusuf, M.; Ahmad, I.; Ahmad, A. Brassinosteroids and their role in response of plants to abiotic stresses. Biol. Plant. 2014, 58 (1), pp. 9-17. [CrossRef]
- Zhu, F., Yun, Z., Ma, Q., Gong, Q., Zeng, Y., Xu, J., Cheng, Y., Deng, X. Effects of exogenous 24-epibrassinolide treatment on posthar-vest quality and resistance of Satsuma mandarin (Citrus unshiu), Postharvest Biol. Technol, 2015, 100, pp. 8-15. [CrossRef]
- Zebosi, B.; Vollbrecht, E.; Bes, N.B. Conservation and diversification of genes regulating brassinosteroid biosynthesis and signal-ing. bioRxiv 2024 . [CrossRef]
- Hause, B.; Mrosk, C.; Isayenkov, S.; Strack, D. Jasmonates in arbuscular mycorrhizal interactions. Phytochemistry 2007, 68, pp.101–110. [CrossRef] [PubMed]
- Hansch, F.; Jaspar, H.; von Sivers, L.; Bitterlich, M.; Franken, P.; Kühn, C. Brassinosteroids and sucrose transport in mycorrhizal tomato plants, Plant Signal. Behav 2020, 15:2, 1714292. [CrossRef]
- García, A.E. Modificaciones al sistema de clasificación climática de Köppen (para adaptarlo a las condiciones de la República Mexicana). 5th Ed.; Instituto de Geografía and Universidad Nacional Autónoma de México, México. 2004; 90 p. http://www.publicaciones.igg.unam.mx/index.php/ig/catalog/book/83.
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Trans. Br. Mycol. Soc., 1970, 55, pp. 158-161. [CrossRef]
- Milthorpe, F.L.; Moorby, J. Introducción a la fisiología de los cultivos. Ed. Hemisferio Sur, Buenos Aires, Argentina; 1982. 259 p.
- SAS (Statistical Analysis System) (1999-2000) SAS/STAT User’S Guide: Ver 8.1. SAS Institute Inc., Cary NC.SAS 1999-2000.
- Jäderlund, L.; Arthurson, V.; Granhall, U.; Jansson, J.K. Specific interactions between arbuscular mycorrhizal fungi and plant growth-promoting bacteria: as revealed by different combinations. FEMS Microbiol. Lett. 2008, 287 (2), pp. 174–180. [CrossRef]
- Wright, S.F. Management of Arbuscular Mycorrhizal Fungi. In Roots and Soil Management: Interactions between Roots and the Soil, Zobel, R.W.; Wright, S.F. Eds., American Society of Agronomy, Crop Science Society of America, Soil Science Society of Ameri-ca, Madison, USA, 2005; pp. 183-197.
- von Sivers, L.; Jaspar, H.; Johst, B.; Roese, M.; Bitterlich, M.; Franken, P.; Kühn, C. Brassinosteroids Affect the Symbiosis Between the AM Fungus Rhizoglomus irregularis and Solanaceous Host Plants. Front. Plant Sci. 2019 10:571. [CrossRef]
- Tofighi, C.; Khavari-Nejad, R.A.; Najafi, F.; Razavi, K.; Rejali, F. Responses of wheat plants to interactions of 24-epibrassinolide and Glomus mosseae in saline condition. Physiol. Mol. Biol. Plants 2017, 23, pp. 557–564. [CrossRef] [PubMed]
- Herrera Aguilar, J.; Aguirre Medina, J. F.; Gálvez López, A. L.; Ley de Coss, A.; Martínez Solís, M. Efecto de reguladores de creci-miento en la reproducción in vitro de Musa spp cv gran enano. Agroproductividad 2017, 10 (9), pp. 20-25. http://www.colpos.mx/wb/index.php/agroproductividad#.WnoBa1TibIU.
- González-Olmedo, J.L.; Córdova, A.; Aragón, C. E.; Pina, D.; Rivas, M.; Rodríguez, R. Efecto de un análogo de brasinoesteroides sobre plántulas de FHIA-18 expuestas a un estrés térmico. InfoMusa. 2005, 14 (1), pp. 18-20. https://www.researchgate.net/publication/311736418_Efectos_de_un_analogo_de_brasinoesteroides_sobre_plantulas_de_FHIA-18_expuestas_a_un_estres_termico_InfoMusa_14_1_18-21_2005.
- Zamora Olivo, M.A.; Aguirre Medina, J.F.; Cano García. M.A.; Martínez-Tinajero, J.J. Productividad de Brachiaria brizantha (Hochst. ex A. Rich) y Clitoria ternatea L. con biofertilizantes. Agroproductividad 2013, 6(6), pp. 23-29. http://www.colpos.mx/wb/index.php/agroproductividad#.VB9GcpR5N8E.
- Artursson, V.; Finlay, R.D.; Jansson, J.K. Interactions between arbuscular mycorrhizal fungi and bacteria and their potential for stimulating plant growth. Environ. Microbiol. 2006, 8(1), pp. 1–10. [CrossRef]
- Leigh, J.; Hodge, A.; Fitter, A. H. Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material. New Phytol 2009, 181, pp. 199–207. [CrossRef] [PubMed]
- Posta, K.; Duc, H.N. Benefits of arbuscular mycorrhizal fungi application to crop production under water scarcity. In drought - detection and solutions. G. Ondrasek (Ed.) IntechOpen. London, England, 2020. [CrossRef]
- González, P.J.; Plana, R.; Rivera, R.; Fernández, F.; Arbola, J. Efectos de la inoculación de hongos micorrízicos arbusculares en pastos del género Brachiaria, cultivados en suelo Pardo Mullido. Rev. cuba. cienc. agríc, 2008, 42(1), pp. 101-106. https://www.redalyc.org/pdf/1930/193015413016.pdf.
- Howeler, R.H.; Sieverding, E.; Saif, S.R. Practical aspects of mycorrhizal technology in some tropical crops and pastures. Plant and Soil 1987, 100, pp. 249-283. https://www.jstor.org/stable/42939116.
- Ibarra-Puón, J.C.; Aguirre-Medina, J.F.; Ley-De Coss, A.; Cadena-Iñiguez, J.; Zavala-Mata A. Inoculación de Coffea canephora (Pierre) ex Froehner con Rhizophagus intraradices (Schenck et Sm.) Walker et Schuessler y Azospirillum brasilense Tarrand, Krieg et Döbereiner en vivero. Rev Chapingo Ser Hortic, 2014, 20(2), pp. 201-213. [CrossRef]
- Aguirre-Medina, J.F.; Mendoza-López, A.; Cadena-Iñiguez, J.; Avendaño-Arrazate, C.H. La Biofertilización del cacao (Theobroma cacao L.) en vivero con Azospirillum brasilense Tarrand, Krieg et Döbereiner y Glomus intraradices Schenk et Smith. Interciencia 2007, 32 (8), pp. 541-546. http://ve.scielo.org/scielo.php?pid=S0378-18442007000800010&script=sci_abstract&tlng=es.
- Izquierdo, H.; Núñez, M.; González, M.; Proenza, R. Efectos de la aplicación de un análogo espirostánico de brasinoesteroides en vitroplantas de banano (Musa spp.) durante la fase de aclimatización. Cult. trop. 2012, 33, pp. 71-76. https://www.medigraphic.com/cgi-bin/new/resumen.cgi?IDARTICULO=73722.
- Terry, A.E.; Ruiz, P.J.; Tejeda, P.T.; Reynaldo, E.I.; Díaz, M.M. Respuesta del cultivo de la lechuga (Lactuca sativa l.) a la aplicación de diferentes productos bioactivos. Cult. trop, 2011, 32(1), pp. 77-82. https://www.redalyc.org/pdf/1932/193222352010.pdf.
- Aguirre-Medina, J. F.; Gálvez-López, A. L.; Ibarra-Puón, J. C. Growth of Leucaena leucocephala (Lam.) de Wit biofertilized with arbuscular mycorrhizal fungi in the nursery. Rev. Chapingo, Ser. Cienc. For. y del Ambient, 2018, 24(1), pp. 49-58. [CrossRef]
- Doubková P, Vlasáková E, Sudová R. Arbuscular mycorrhizal symbiosis alleviates drought stress imposed on Knautia arvensis plants in serpentine soil. Plant Soil 2013, 370, pp. 149–161. [CrossRef]
- Cruz, R, de Sousa,; Vieira Araújo, F.H.; Cabral França, A.; Tadin Sardinha, L.; Miranda Machado, C.M. Physiological responses of Coffea arabica cultivars in association with arbuscular mycorrhizal fungi. Coffee Sci. 2020. 15, e151641. [CrossRef]
- Jiménez Terry, F.A.; Ramírez Aguilar, D.; Agramonte Peñalver, D. Empleo del BIOBRAS-6 en la micropropagación del cultivar de plátano FHIA21 (AAAB). Biotecnol. veg., 2002, 2 (3), pp. 131-136. https://revista.ibp.co.cu/index.php/BV/rt/printerFriendly/171/html.
- Izquierdo Oviedo, H.; González Cepero, M.C.; Núñez Vázquez, M. de la C.; Proenza Llerena, R.; Cabrera Pino, J.C. Biological activity of biobras and Pectimorf-6 in each of the phases of micropropagation of banana (Musa sp.). J. Research Biol. 2017, 7(3), pp. 2231-2247. file:///C:/Users/franc/Downloads/Biological_activity_of_biobras_and_Pecti%20(1).pdf.
- Neetu, N.; Ashok, A.; Anju, T.; Alpa, A. Influence of arbuscular mycorrhizal fungi and Pseudomonas fluorescens at different su-perphosphate levels on linseed (Linum usitatissimum L.) growth response. Chil. J. Agric. Res. 2012, 72 (2), pp. 237-243.
- Singh, R.; Pandey, D. K.; Kumar, A.; Singh, M. PGPR isolates from the rhizosphere of vegetable crop Momordica charantia: char-acterization and application as biofertilizer. Int J Curr Microbiol Appl Sci, 2017, 6(3), pp. 1789-1802. [CrossRef]
- Desmet, S.; Saeys, Y.; Verstaen, K.; Dauwe, R.; Kim, H.; Niculaes, C.; Fukushima, A.; Goeminne, G.; Vanholme, R.; Ralph, J.; Boer-jan, W.; Morreel, K. Maize specialized metabolome networks reveal organ-preferential mixed glycosides. Comput Struct Biotechnol J. 2021, 19, pp. 1127–1144. [CrossRef]
- Larose, G.; Chênevert, R.; Moutoglis, P.; Gagné, S.; Piché, Y.; Vierheilig, H. Flavonoid levels in roots of Medicago sativa are mod-ulated by the developmental stage of the symbiosis and the root colonizing arbuscular mycorrhizal fungus. J. Plant Physiol. 2002, 159, pp. 1329–1339. [CrossRef]
- Ruiz-Lozano, J.M.; Perálvarez, M.C.; Aroca, R,; Azcón, R. The application of a treated sugar beet waste residue to soil modifies the responses of mycorrhizal and non mycorrhizal lettuce plants to drought stress. Plant Soil 2012, 346, pp. 153–166. [CrossRef]
- Lou, S.; Jiang, H.; Li, J.; Tian, L.; Du, M.; Ma, T.; Zhang, L.; Zhang, P. Effects of Exogenous Brassinosteroid and Reduced Leaf Source on Source–Sink Relationships and Boll Setting in Xinjiang Cotton. Agronomy 2024, 14, 1168. [CrossRef]
- Zhang, X.; Sun, S.; Nie, X.; Boutté, Y.; Grison, M.; Li, P.; Kuang, S.; Men, S. Sterol Methyl Oxidases Affect Embryo Development via Auxin-Associated Mechanisms. Plant Physiol. 2016, 171, pp. 468-482. [CrossRef]
- Liu, C.Y.; Zhang, F.; Zhang, D.J.; Srivastava, A. K.; Wu, Q-S.; Zou, Y-N. Mycorrhiza stimulates root-hair growth and IAA synthesis and transport in trifoliate orange under drought stress. Sci Rep 2018, 8, 1978. [CrossRef] [PubMed]
- Dhiman, M.; Sharma, L.; Kaushik, P.; Singh, A.; Sharma, M.M. Mycorrhiza: An Ecofriendly Bio-Tool for Better Survival of Plants in Nature. Sustainability 2022, 14, 10220. [CrossRef]
- Liao, D.; Wang, S.; Cui, M.; Liu, J.; Chen, A.; Xu, G. Phytohormones regulate the development of Arbuscular mycorrhizal symbio-sis. Int. J. Mol. Sci. 2018, 19, 3146. [CrossRef]
- Rao, A.V.; Tak, R. Growth of different tree species and their nutrient uptake in limestone mine spoil as influenced by Arbuscular mycorrhizal (AM)-fungi in Indian arid zone. J. Arid Environ. 2002, 51, pp. 113–119. [CrossRef]
- He J.D.; Chi G.G.; Zou Y.N.; Shu B.; Wu Q.S. Srivastava A.K., Kuča K. (2020): Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl. Soil Ecol., 2020, 154: 103592. [CrossRef]
- Powell, J.R.; Rillig, M.C. Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol. 2018, 220, pp. 1059–1075. [CrossRef]
- Altomare, C.; Tringovska, I. Beneficial Soil Microorganisms, an Ecological Alternative for Soil Fertility Management. In: Licht-fouse E. (eds). Genetics, Biofuels and Local Farming Systems. Sustainable Agriculture Reviews, Springer, Dordrecht, 2011, 7. pp. 161–214. [CrossRef]
- Kanno, T.; Saito, M.; Ando, Y.; Macedo, M. C. M.; Nakamura, T.; Miranda, C. H. B. Importance of indigenous arbuscular mycor-rhiza for growth and phosphorus uptake in tropical forage grasses growing on an acid, infertile soil from the Brazilian savannas. Trop. grassl, 2006, 40, pp. 94–101. Retrieved from http://www.tropicalgrasslands.info/public/journals/4/Historic/Tropical%20Grasslands%20Journal%20archive/PDFs/Vol_40_2006/Vol_40_02_20.
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved photosynthetic efficacy of maize (Zea mays) plants with Arbuscular mycorrhizal fungi (AMF) under high temperature stress. J. Photochem. Photobiol. Biol. 2018, 180, pp. 149–154. [CrossRef] [PubMed]
- Otie, V.; Ibrahim, A.; Udo, I.; Kashiwagi, J.; Matsuura, A.; Shao, Y.; Itam, M.; An, P.; Eneji, A.E. Foliarly applied 2,4- Epibrassino-steroid modulates the electrical conductivity of the saturated rhizospheric soil extracts of soybean under salinity stress. Plants 2022, 11, 2330. [CrossRef]
- Aguirre-Medina, J.F. 2006. Biofertilizantes microbianos: su aplicación en la agricultura. Folleto Técnico Núm. 1. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional Pacifico Sur. Campo Experimental Rosario Izapa. Chiapas, México. 1, 24 p.
- Devi, S. H.; Bhupenchandra, I.; Sinyorita, S.; Chongtham, S.; Devi, E. L. Mycorrhizal Fungi and Sustainable Agriculture. In Nitro-gen in Agriculture -Physiological, Agricultural and Ecological Aspects. T. Ohyama, & K. Inubushi (Eds.). IntechOpen, London, England. 2021. [CrossRef]
- França, A. C.; de Freitas A.N.; dos Santos, A. E.; Grazziotti, P.H.; de Andrade Júnio, V. C. Mycorrhizal fungi increase coffee plants competitiveness against Bidens pilosa interference. Pesqui. Agropecu. Trop, 2016, 46(2), pp.132-139. [CrossRef]
- Aguirre Medina, J.F.; Culebro Cifuentes, F.; Cadena Iñiguez, H.; Aguirre Cadena, J.F. Crecimiento de Tabebuia Donnell-Smithii (Rose) Inoculada con Hongos Micorrizicos y Azospirillum brasilense. Agrociencia. 2014, 48 (3), pp. 331-345. http://www.colpos.mx/agrocien/agrociencia.htm.
- Jacott, C.N.; Murray, J.D.; Ridout, C.J. Trade-offs in arbuscular mycorrhizal symbiosis: Disease resistance, growth responses and perspectives for crop breeding. Agronomy 2017, 7(4), 75. [CrossRef]
- Steenhoudt, O.; Vanderleyden, J. Azospirillum, a free living nitrogen-fixing bacterium closely associated with grasses: genetic, biochemical and ecological aspects. FEMS Microbiol. Rev. 2000, 24 (4), pp. 487–506. [CrossRef] [PubMed]
- Jefwa, J.M.; Sinclair, R.W.; Maghembe, J.A. Diversity of Glomale mycorrhizal fungi in Maize/Sesbania intercrops and maize monocrop systems in Southern Malawi. Agrofor. Syst. 2004, 67, pp. 107-114. [CrossRef]
- Perreta, M.G.; Vegetti, A.C. Patrones estructurales en las plantas vasculares: Una Revisión. Gayana Bot. 2005, 62 (1), pp. 9-19. [CrossRef]
- Adolfsson, L.; Solymosi, K.; Andersson, M. X.; Keresztes, Á.; Uddling, J.; Schoefs, B.; Spetea, C. (2015) Mycorrhiza symbiosis in-creases the surface for sunlight capture in Medicago truncatula for better photosynthetic production. PLoS ONE, 10:e0115314. [CrossRef]
- Baum, C.; Toljander, Y.K.; Eckhardt, K.U.; Weih, M. The significance of host-fungus combinations in ectomycorrhizal symbioses for the chemical quality of willow foliage. Plant Soil 2009, 323, pp. 213–224. [CrossRef]
- Tang, H.; Hassan, M.U.; Feng, L.; Nawaz, M.; Shah, A.N.; Qari, S.H.; Liu, Y.; Miao, J. The critical role of arbuscular mycorrhizal fungi to improve drought tolerance and nitrogen use efficiency in crops. Front. Plant Sci. 2022, 13, 919166. [CrossRef] [PubMed]
Figure 1.
Root and aerial biomass of B. decumbens Stapf biofertilized with R. intraradices, A. brasilensis in interaction with an homobrassinolide. The values are averages of five repetitions per treatment.
Figure 1.
Root and aerial biomass of B. decumbens Stapf biofertilized with R. intraradices, A. brasilensis in interaction with an homobrassinolide. The values are averages of five repetitions per treatment.
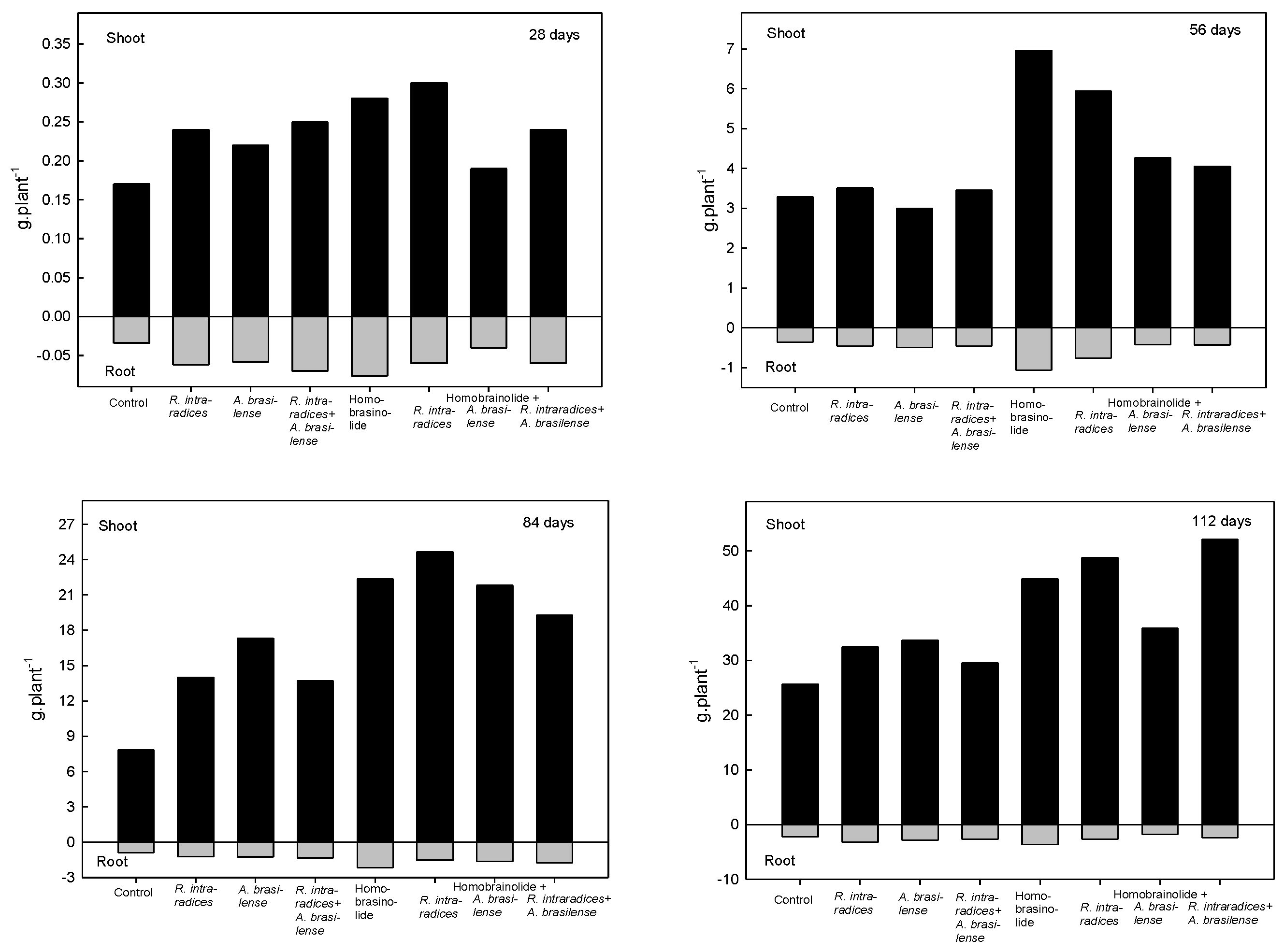
Figure 2.
Relative growth rate of B. decumbens Stapf plants biofertilized with A. brasilense and/or R. intraradices alone and/or combined in interaction with a homobrassinolide. Values are averages of five plants per treatment and sampling.
Figure 2.
Relative growth rate of B. decumbens Stapf plants biofertilized with A. brasilense and/or R. intraradices alone and/or combined in interaction with a homobrassinolide. Values are averages of five plants per treatment and sampling.
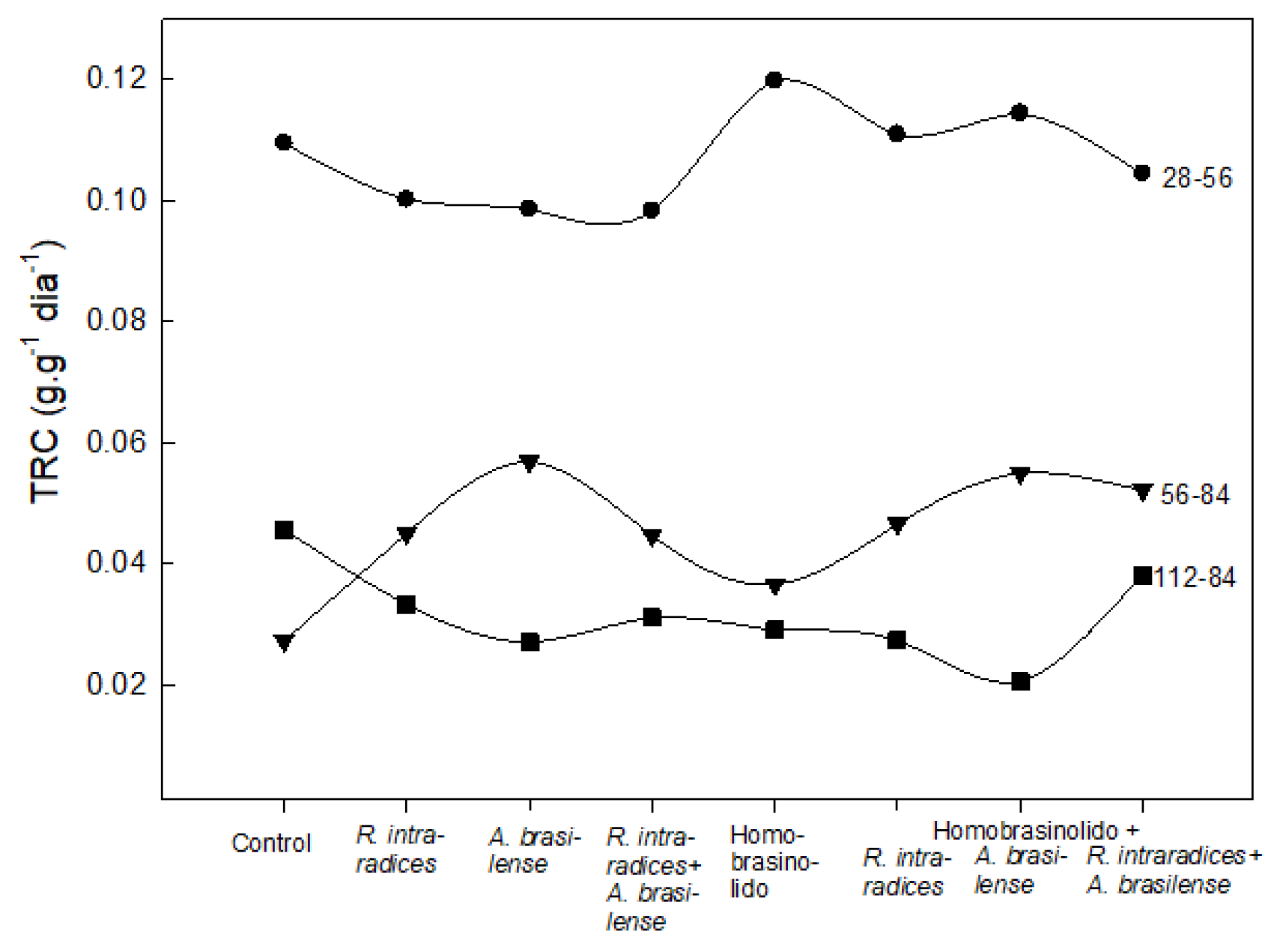
Figure 3.
Mycorrhizal colonization on B. decumbens Stapf biofertilized with A. brasilense and/or R. intraradices alone and/or combined in interaction with a homobrassinolide at 28 das. Values are averages of four samples per treatment.
Figure 3.
Mycorrhizal colonization on B. decumbens Stapf biofertilized with A. brasilense and/or R. intraradices alone and/or combined in interaction with a homobrassinolide at 28 das. Values are averages of four samples per treatment.
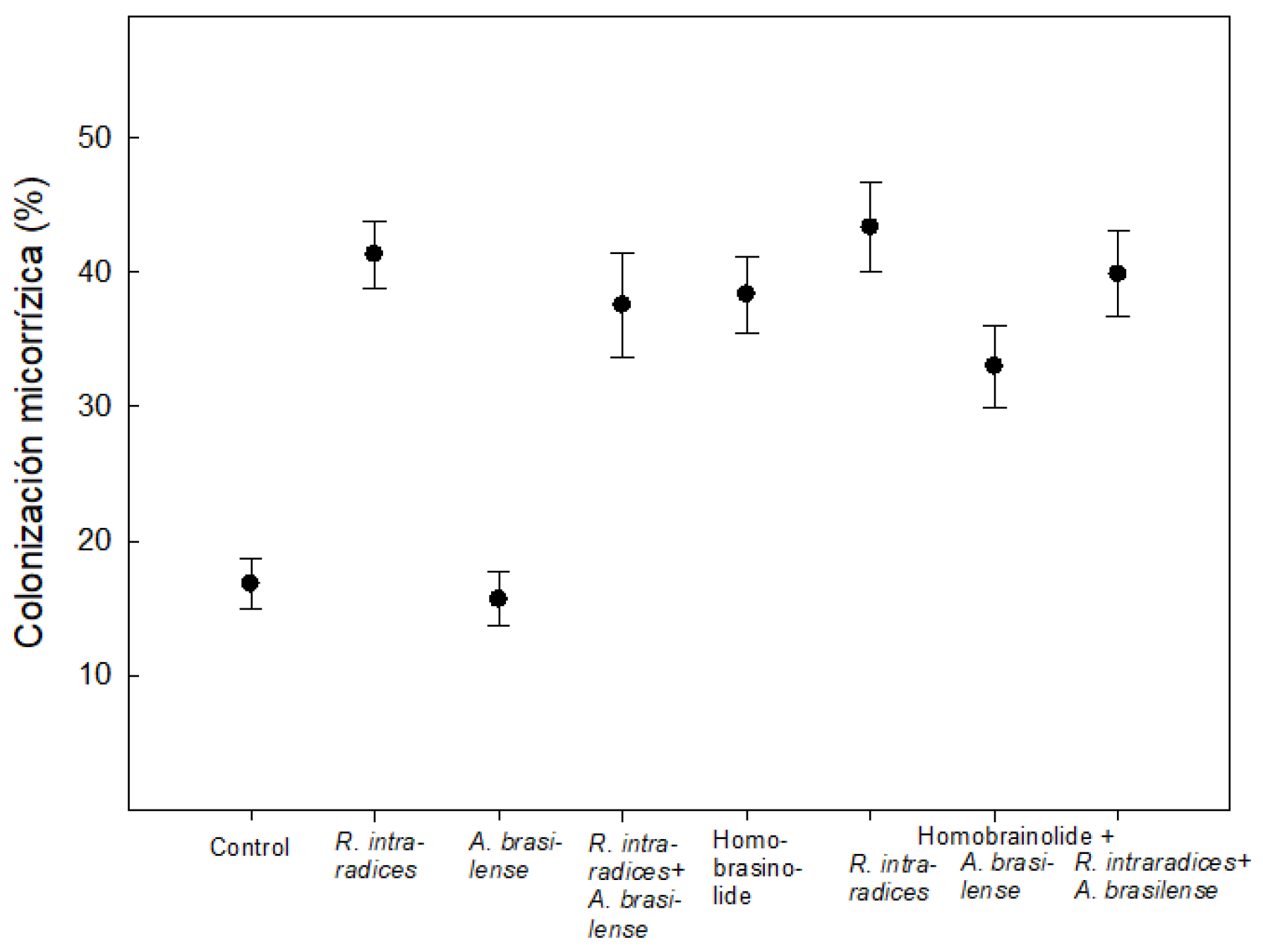
Figure 4.
Variation in nitrogen and phosphorus content in the plant tissue of B. decumbens Stapf biofertilized with R. intraradices, A. brasilense in interaction with homobrassinolide. Different letters indicate statistically significant differences according to the Tukey test (P ≤ 0.05). The vertical bars indicate the standard error of the mean (n = 4). Coefficient of variation = 14.9 %N y 15.1 %P.
Figure 4.
Variation in nitrogen and phosphorus content in the plant tissue of B. decumbens Stapf biofertilized with R. intraradices, A. brasilense in interaction with homobrassinolide. Different letters indicate statistically significant differences according to the Tukey test (P ≤ 0.05). The vertical bars indicate the standard error of the mean (n = 4). Coefficient of variation = 14.9 %N y 15.1 %P.
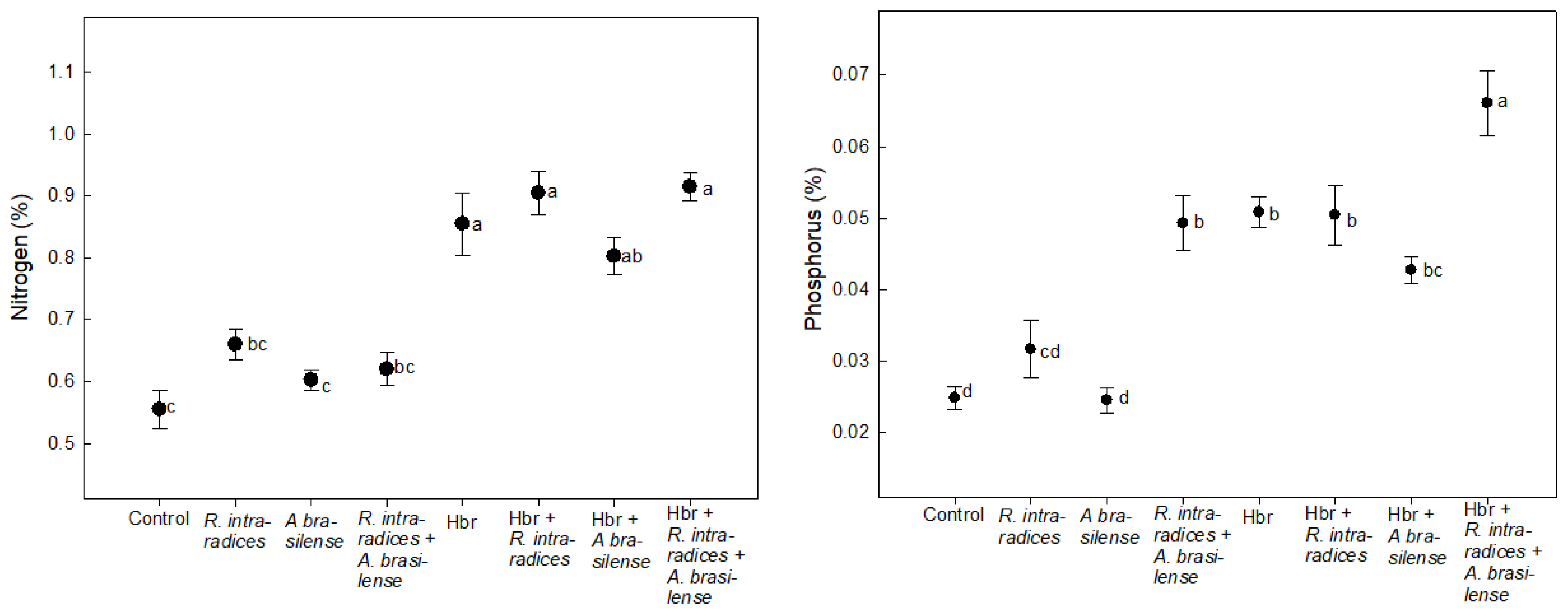

Table 1.
Morphological yield components of Brachiaria decumbnes Stapf. biofertilized with R. intraradices, A. brasilense and foliar application of homobrassinolide in nursery.
Table 1.
Morphological yield components of Brachiaria decumbnes Stapf. biofertilized with R. intraradices, A. brasilense and foliar application of homobrassinolide in nursery.
| Time (days) | Treatment | Height (cm.plant-1) |
Leaves (Number.plant-1) |
Stems (Number.plant-1) |
Stem diameter*** (mm.plant-1) |
| 28 | Control | 14.2 c** | 3.8 c | 2.4 b | 2.0 ab |
| R. intraradices | 15.8 bc | 4.6 abc | 3.2 ab | 2.0 ab | |
| A. brasilense | 15.0 bc | 4.8 ab | 2.8 b | 1.9 ab | |
| R. intraradices+A. brasilense | 16.8 ab | 4.4 bc | 3.0 b | 2.0 ab | |
| Hbr* | 15.8 bc | 5.4 a | 3.8 a | 2.2 a | |
| Hbr + R. intraradices | 18.4 a | 5.4 a | 3.0 ab | 2.3 a | |
| Hbr + A. brasilense | 18.2 a | 4.8 ab | 2.8 b | 1.7 b | |
| Hbr + R. intraradices + A. brasilense | 15.2 bc | 5.0 ab | 2.6 b | 2.0 ab | |
| CV (%) | 5.5 | 9.9 | 14.1 | 8.7 | |
| 56 | Control | 36.6 e | 14.4 f | 3.8 bc | 2.4 a |
| R. intraradices | 47.2 d | 21.0 d | 3.6 c | 2.5 a | |
| A. brasilense | 49.6 bcd | 16.2 ef | 3.8 bc | 2.4 a | |
| R. intraradices+A. brasilense | 46.2 d | 18.0 e | 4.0 bc | 2.5 a | |
| Hbr | 47.6 cd | 32.2 a | 5.0 a | 2.6 a | |
| Hbr + R. intraradices | 58.4 a | 29.2 b | 4.6 ab | 2.5 a | |
| Hbr + A. brasilense | 55.2 abc | 23.8 c | 4.2 abc | 2.6 a | |
| Hbr + R. intraradices + A. brasilense | 56.4 ab | 24.8 c | 3.8 c | 2.6 a | |
| CV (%) | 7.7 | 4.5 | 11.8 | 7.0 | |
| 84 | Control | 61.6 c | 29.6 d | 3.8 d | 2.5 b |
| R. intraradices | 66.8 b | 47.0 c | 4.6 cd | 2.6 b | |
| A. brasilense | 68.8 b | 55.2 bc | 4.6 cd | 2.5 b | |
| R. intraradices+ A. brasilense | 67.6 b | 53.6 bc | 5.4 bc | 2.5 b | |
| Hbr | 72.6 a | 70.2 a | 6.8 a | 2.6 b | |
| Hbr + R. intraradices | 69.4 b | 68.8 a | 5.6 bc | 2.9 a | |
| Hbr + A. brasilense | 69.4 b | 57.6 b | 6.4 ab | 2.7 b | |
| Hbr + R. intraradices + A. brasilense | 72.6 a | 55.0 cb | 6.0 ab | 2.9 a | |
| CV (%) | 2.2 | 7.9 | 10.1 | 2.9 | |
| 112 | Control | 65.4 d | 82.0 c | 5.0 d | 2.6 b |
| R. intraradices | 71.8 c | 98.4 b | 5.8 cd | 2.6 ab | |
| A. brasilense | 73.6 c | 99.2 b | 5.6 cd | 2.7 ab | |
| R. intraradices+ A. brasilense | 76.2 bc | 94.0 b | 6.0 bcd | 2.6 b | |
| Hbr | 82.4 a | 114.8 a | 9.6 a | 2.8 ab | |
| Hbr + R. intraradices | 81.6 a | 124.4 a | 8.8 a | 2.9 a | |
| Hbr + A. brasilense | 82.6 a | 95.4 b | 6.6 bc | 2.7 ab | |
| Hbr + R. intraradices + A. brasilense | 80.8 ab | 122.0a.6 | 7.2 b | 2.8 ab | |
| CV (%) | 3.4 | 5.2 | 8.9 | 5.2 |
*Hbr= Homobrassinolide. **Values with the same letter within each factor and column are equal according to Tukey’s test at p ≤ 0.05. CV = coefficient of variation (%). *** average of three stems per plant.
Table 2.
Physiological yield components of Brachiaria decumbnes Stapf. biofertilized with R. intraradices, A. brasilense and foliar application of homobrasinolide in nursery.
Table 2.
Physiological yield components of Brachiaria decumbnes Stapf. biofertilized with R. intraradices, A. brasilense and foliar application of homobrasinolide in nursery.
| dry weight (g.plant-1) | Leaf area (cm2.plant-1) | |||||
| Time (days) | Treatment | Root | Leaves | Stem | ||
| 28 | Control | 0.034 c** | 0.092 d | 0.044 e | 27.2 c | |
| R. intraradices | 0.062 b | 0.122 b | 0.068 abc | 38.6 b | ||
| A. brasilense | 0.064 b | 0.104 cd | 0.054 de | 41.3 b | ||
| R. intraradices+A. brasilense | 0.070 ab | 0.120 cd | 0.066 bc | 41.6 b | ||
| Hbr* | 0.076 a | 0.128 b | 0.078 a | 53.8 a | ||
| Hbr + R. intraradices | 0.060 b | 0.154 a | 0.076 ab | 61.3 a | ||
| Hbr + A. brasilense | 0.040 c | 0.090 d | 0.064 cd | 35.3 b | ||
| Hbr + R. intraradices + A. brasilense | 0.060 b | 0.112 bc | 0.068 abc | 27.2 c | ||
| CV (%) | 9.7 | 7.1 | 8.8 | 8.9 | ||
| 56 | Control | 0.35 d | 1.92 de | 1.37 d | 285.2 d | |
| R. intraradices | 0.40 cd | 2.00 cde | 1.50 cd | 301.8 d | ||
| A. brasilense | 0.49 c | 1.65 e | 1.34 d | 537.4 a | ||
| R. intraradices+A. brasilense | 0.46 c | 2.00 cde | 1.45 cd | 581.0 a | ||
| Hbr | 1.06 a | 3.91 a | 3.02 a | 469.9 b | ||
| Hbr + R. intraradices | 0.77 b | 3.12 b | 2.61 b | 445.2 bc | ||
| Hbr + A. brasilense | 0.42 cd | 2.53 c | 1.73 c | 456.7 bc | ||
| Hbr + R. intraradices + A. brasilense | 0.42 cd | 2.38 cd | 1.67 cd | 399.1 c | ||
| CV (%) | 8.1 | 11.1 | 9.8 | 6.9 | ||
| 84 | Control | 0.88 e | 4.18 e | 3.65 d | 537.38 f | |
| R. intraradices | 1.23 d | 7.07 d | 6.93 c | 621.13 ef | ||
| A. brasilense | 1.24 d | 8.69 c | 8.63 b | 928.17 d | ||
| R. intraradices+ A. brasilense | 1.32 cd | 7.06 e | 6.64 c | 719.11 e | ||
| Hbr | 2.15 a | 10.00 b | 11.55 a | 1055.43 c | ||
| Hbr + R. intraradices | 1.52 bcd | 12.14 a | 12.52 a | 1427.05 a | ||
| Hbr + A. brasilense | 1.63 bc | 10.51 b | 11.29 a | 1254.99 b | ||
| Hbr + R. intraradices + A. brasilense | 1.75 b | 10.01 b | 9.28 b | 1093.19 c | ||
| CV (%) | 11.3 | 6.0 | 9.2 | 5.7 | ||
| 112 | Control | 2.20 e | 10.73 c | 14.94 ed | 151.18 c | |
| R. intraradices | 3.21 b | 12.92 b | 19.47 d | 174.73 c | ||
| A. brasilense | 2.86 c | 13.41 b | 20.26 cd | 169.85 c | ||
| R. intraradices+ A. brasilense | 2.61 cd | 12.71 bc | 16.79 e | 171.86 c | ||
| Hbr | 3.64 a | 18.26 a | 26.63 b | 288.16 a | ||
| Hbr + R. intraradices | 2.69 cd | 19.85 a | 28.92 b | 274.60 a | ||
| Hbr + A. brasilense | 1.80 f | 13.58 b | 22.26 c | 206.41 c | ||
| Hbr + R. intraradices + A. brasilense | 2.44 de | 19.32 a | 32.79 a | 2812.44 a | ||
| CV (%) | 6.0 | 6.4 | 4.9 | 6.4 | ||
*Hbr= Homobrassinolide. **Values with the same letter within each factor and column are equal according to Tukey’s test at p ≤ 0.05. CV = coefficient of variation (%).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



