
Submitted:
26 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
Reactive oxygen species (ROS) are intermediates in oxidation-reduction reactions with the capacity to modify biomolecules and temporarily or permanently alter cell behaviour through signalling pathways under physiological and pathophysiological conditions where there is an imbalance between oxidative factors and the antioxidant response of the organism, a phenomenon known as oxidative stress. Evidence suggests that differential modulation of ROS-mediated oxidative stress occurs in the pathogenesis and progression of melanoma, and that this imbalance in redox homeostasis appears to be functionally linked to microRNA-mediated non-mutational epigenetic reprogramming involving genes and transcription factors. The relationship between ROS-mediated stress control, tumour microenvironment and microRNAs expression in melanoma is not fully understood. The aim of this review is to analyze the involvement of miRNAs in the modulation of signalling pathways involved in ROS-mediated oxidative stress in melanoma. It is hoped that these considerations will contribute to the understanding of the mechanisms associated with a potential epigenetic network regulation, where the modulation of oxidative stress is consolidated as a common factor in melanoma and therefore a potential footprint poorly documented.
Keywords:
Melanoma
; ROS
; microRNAs
; Redox Homeostasis
; Oxidative Stress
Introduction
The term reactive oxygen species (ROS) refers to chemical structures such as superoxide anions, peroxides and hydroxyl radicals, which contain one or more unpaired electrons; they are intermediates in oxidation-reduction reactions with the capacity to modify biomolecules and temporarily or permanently alter cell behaviour through various signalling pathways in physiological and pathophysiological conditions where there is an imbalance between oxidative factors and the antioxidant response of the organism, a phenomenon known as oxidative stress [1]. While in a healthy cell, stress potentiates mechanisms that generally lead to cell death, in cancer, a strong survival capacity of tumour cells is observed at levels of oxidative stress increased by ROS, a condition that involves, among other things, genetic and epigenetic changes that affect the biochemistry and modulation of mitochondrial function [2,3].
Melanoma is a type of skin cancer that originates from melanocytes, cells specialized in melanin synthesis, and is characterized by increased cell proliferation, resistance to chemotherapeutic agents and metastasis. According to the World Health Organization (WHO), melanoma is one of the deadliest cancers in the world, with a survival rate of more than 80% of reported cases in the early stages of diagnosis, but falling to 5% in the metastatic stages [4,5]. Phenotypes associated with the pathogenesis and progression of this cancer include modulation of oxidative stress, which coincides with changes in gene expression and metabolic reprogramming involving microRNAs (miRNAs or miRs), small non-coding RNAs involved in post-transcriptional epigenetic regulation, which regulate the cellular response to changes in the microenvironment by acting as switches in pathway activation and gene repression in virtually all cancer pathways involved in adaptation and survival [6,7]. It is possible that this modulation of oxidative stress may emerge as a new hallmark underlying cancer.
The relationship between the control of ROS-mediated stress, the tumour microenvironment and the expression of miRNAs in melanoma is not fully understood due to the use of linear regulatory models that limit the integration of a dynamic molecular architecture and configuration associated with the modulation of oxidative stress as a survival strategy; Therefore, new approaches are required, possibly from a network biology that integrates signalling pathways, transcription factors, miRNAs and genes, which together will guide the development of new therapeutic strategies for early diagnosis and potentially effective treatments for more advanced stages of this cancer [8,9]. The aim of this review is to analyze the involvement of miRNAs in the modulation of signalling pathways in ROS-mediated oxidative stress environments in melanoma. The paper is divided into three sections: the first section deals with the described relationship between miRNAs and ROS in melanoma; the second section reviews the involvement of different signalling pathways modulated by ROS and their relationship with changes in miRNA expression; and the third section presents a bioinformatic and statistical functional enrichment approach for the construction of potential network models mentioned above, based on a dataset reported in the Gene Expression Omnibus (GEO).
miRNAs, ROS and Melanoma
Molecular mechanisms associated with melanoma tumour physiology in processes such as proliferation, growth, energy metabolism, migration, differentiation and cell death involve oxidative stress and thus genetic and epigenetic alterations of oncogenes and tumour suppressor genes [10,11,12,13]. Non-mutational epigenetic reprogramming is exerted by miRNAs, small non-coding RNAs of 18-22 nt, which play a role in the post-transcriptional regulation of up to 60% of mammalian protein-coding mRNAs [14,15,16,17]. Most miRNAs are derived from long intramolecular double-stranded RNAs; these RNAs are sequentially cleaved by type III RNases, first in the nucleus and then in the cytoplasm, to generate a miRNA duplex. In subsequent steps, one of the strands of the duplex associates with an RNA-induced silencing complex (RISC), which triggers a decrease in a specific mRNA by degrading the transcript or repressing mRNA-to-protein translation [18,19]. A single miRNA can target hundreds of mRNAs, and a single mRNA can target multiple miRNAs, so variations in the expression of thousands of mRNAs could be explained by the coordinated network expression patterns of a few miRNAs [20]. There are miRBase records of 2600 corresponding sequences for mature miRNAs in humans [16,21,22,23,24].
Potential relationships between ROS and miRNAs in cancer have been reported, e.g., miRNA-21 can inhibit the antioxidant pathway of superoxide dismutase (SOD), facilitating oxidative damage by superoxide anion (O2-) and other ROS, as well as enhancing oncogenic and fibrotic processes associated with carcinoma [24,25]. Alterations in miRNA-5096 increase hydroxyl radical (OH-) levels and suppress non-apoptotic cell proliferation in breast cancer cells; the mechanism of cell damage of this miRNA would be mediated by ROS and is reversible with the use of antioxidants such as N-acetyl cysteine (NAC) [26]. Meanwhile, it has been shown that the increase of miRNA-34 in human glioma cells, together with the suppression of nitrogen dioxide radicals, supports the increase of apoptosis rate and the decrease of cell viability in a glioma model [27].
In melanoma, miRNAs are also differentially expressed and their alteration may be related to changes in the oxidative environment and the activation of cancer imprints. For example, enrichment of extracellular vesicles with miRNA-214 secreted by melanoma cells stimulates over-activation of macrophages, which release nitric oxide (NO), a process that facilitates endothelial permeability and favors metastasis [19]. Nitric oxide alone is not capable of causing DNA damage, but it can inhibit enzymes such as DNA ligase and thus indirectly cause double helix breaks [28,29]. It has also been suggested that silencing miRNA-517a in melanoma patient samples results in overexpression of CDKN1C (cyclin-dependent kinase inhibitor 1C) and suppression of the JNK kinase-mediated survival and proliferation pathway, leading to increased ROS [8]. Furthermore, excessive levels of ROS, such as those caused by the administration of chemotherapeutic agents, induce the development of resistance mechanisms to the oxidative environment; such resistance has been studied in the human melanoma cell line A375, in which activation of the mitogen-activated protein kinase (MAPK) pathway (RAS/MEK/ERK) and the transcription factors hypoxia-inducible factor 1 (HIF-1α) and the master regulator of melanocyte differentiation, microphthalmia-associated transcription factor (MITF), were observed.
Indeed, other miRNAs with different molecular targets have been linked to the HIF-1α and MITF pathways. The miR-33a and miR-138 are thought to directly regulate HIF-1α in the transition from melanocyte to metastatic melanoma [30,31]. miR-182 induces increased invasion and metastasis by binding to the tumour suppressor FBXW, and its binding to FOXO3 and MITF appears to be associated with melanoma progression, and miR-182 is a target of epigenetic modulation with hypermethylated CpG islands in melanoma cells [14]. On the other hand, miR-211 modulates the expression of genes involved in cell cycle regulation and tumour suppression, such as p16INK4A, BRN2 and MITF [32,33]. Meanwhile, miR-203 has been suggested to act as a tumour suppressor by regulating melanosome transport and tyrosinase enzyme (TYR) expression through the kinesin 5 (kif5b) superfamily of proteins and appears to negatively modulate one of the major signalling pathways active in melanoma cells, the CREB1/MITF/Rab27a pathway [34].
In addition to promoting a tumorigenic environment, miRNAs can also act as regulators of p53-associated tumour suppressor conditions and molecular machinery [35,36,37,38]. Activation of p53, stimulated by cellular stresses such as ionizing radiation, hypoxia, carcinogens or oxidative stress, leads to cell cycle arrest and promotes DNA repair or induces apoptosis through various pathways [39]. The expression of miR-18b is significantly reduced in patient-derived melanomas and cell lines due to hypermethylation of the p53 pathway, while its stable overexpression results in potent tumour suppressor activity as measured by cell viability, induction of apoptosis and reduction of tumour growth in xenograft assays [40].
It is possible that oxidative stress in melanoma involves non-mutational epigenetic reprogramming in which networked sets of miRNAs, together with transcription factors such as MITF and HIF-1α, exert control over specific mRNAs whose protein products modulate the activity of various signalling pathways that ensure survival under these stress conditions, an aspect that will be discussed in the next section.
Oxidative stress Signalling Pathways and miRNA Expression in Melanoma
Growth suppressor evasion, resistance to cell death, replicative immortality, angiogenesis, invasiveness, metastasis and sustained proliferative signalling are hallmarks of cancer, coupled with energy dysregulation, genomic instability, release of phenotypic plasticity, non-mutational epigenetic reprogramming and sustained inflammation, with a characteristic stress-mediated environment [41]. These hallmarks coincide with the modulation of several melanoma-associated signalling pathways, such as the MAPK pathway and MITF, which stimulate growth. There is also activation of the phosphatidylinositol 3-kinase (PI3K/AKT) pathway, which is responsible for altering extracellular functions as a metabolic regulator of survival and a generator of cell growth signals [42,43].
HIF-1α, also, acts as a transcriptional mediator of tissue hypoxia; HIF-1α is hyperactivated in conditions of prolonged anoxia, as a consequence of possible adaptation to tumour microenvironment factors, the reduction of mitochondrial respiration in low-oxygen environments and the subsequent Warburg effect, conditions that alternate with the control of ROS and therefore the modulation of oxidative stress. Specifically, HIF-1α inactivates the enzyme pyruvate kinase, thereby redirecting cellular metabolism towards glycolysis [44]. Mutations in signalling pathways such as PI3K/AKT can stabilize HIF-1α even under normoxic conditions [45,46]. Other signalling pathways, such as NRF2/Keap1, regulate the antioxidant response in melanoma and have an ambivalent function, protecting the healthy melanocyte from stress but also promoting tumour cell survival against ROS-induced damage [47]. Other ROS-enriched environments may promote pathways such as the Wnt pathway, which is involved in melanogenesis through MITF [48]. Signalling interactions are not necessarily linear, but involve a complex multi-scale regulatory network that can be reprogrammed by miRNAs, even in the presence of genetic changes [49].
MITF Signalling Pathway
Studies have linked MITF function to cellular plasticity in melanoma, and several changes in its expression and activity as a suppressor and mediator of tumour progression have been documented [50]. These regulatory mechanisms include cellular extrinsic signals in combination with intrinsic post-transcriptional epigenetic changes that determine the status of tumour cells [51]. The microenvironmental signals generated drive a reversible phenotypic switch between a proliferative and a highly invasive phenotype.
Melanocytes are melanin-producing cells whose differentiation, proliferation and survival are highly dependent on MITF [52]. In normal cells, MIFT is critical for melanocyte survival through increased expression of BCL2 and BcL xL (anti-apoptotic factors) [53] and DICER, a factor that has been shown to be critical for melanocyte survival through its role in processing miRNAs [54]. In melanoma cells, depletion of these anti-apoptotic proteins, particularly Bcl-2, leads to cell death [52].
The expression of MITF is regulated by melanocyte stimulating hormone (α-MSH) and its action on the MC1R receptor; this interaction increases the transcription of genes related to pigmentation, in terms of melanin synthesis. The physiological response requires exposure to ultraviolet radiation (UVR), which increases the likelihood of DNA damage and activation of p53, leading to transcription of pro-opiomelanocortin (POMC) in keratinocytes. POMC is enzymatically cleaved to produce αMSH, which binds to MC1R in melanocytes. This triggers the rest of the pigment response, resulting in the transcription of MITF and its targets TYR, dopachrome tautomerase (DCT) and tyrosinase-related enzyme 1 (TYRP1). Melanin is produced in melanosomes and transported to keratinocytes, where they form protective layers over their nuclei [55].
Transcriptional control of MITF is provided by a number of transcription factors and regulators associated with signalling pathways such as LEF1 (lymphoid enhancer-binding factor 1) and β-catenin, both effectors of the Wnt pathway, discussed below [51]. It has also been shown that this αMSH/MC1R/cAMP/PKA signalling cascade can direct β-catenin to specific cAMP response element-binding protein (CREB) promoters to activate transcription of target genes, including MITF [56]. ZEB1 and ZEB2 are genes that determine cellular state by modulating MITF expression. ZEB2 promotes MITF expression and is associated with a differentiated and proliferative cell state. In addition to, ZEB1 is associated with low MITF expression and a more invasive cell state [57].
In addition to the transcription factors and post-translational regulators described above, MITF regulation also involves the regulation of several miRNAs. In fact, using TargetScan 7.2, more than 30 potential sites in the 3′UTR region of the MITF gene (ENST00000328528.6) were found to be regulated by miRNAs [58]. The miRNA-182 promotes cell migration and survival in melanoma through negative regulation of MITF [59]. It has also been reported that miRNA-137 harbors a melanoma susceptibility allele and is a down-regulator of MITF expression, apparently associated with cell cycle arrest in G1 [60]. Downregulation of Bcl-2 expression by small interfering RNA has been reported to modulate miRNA-211 expression [61,62]. In addition, immunofluorescence assays revealed a reduction in MITF following Bcl-2 overexpression, suggesting that Bcl-2 may be a regulator of MITF in the context of melanoma [63]. Indeed, miR-211, a melanocyte lineage-specific miRNA, localizes to an intron of TRPM1, a target gene of MITF, which would explain the above findings [62].
HIF-1α Signalling Pathway
Under normoxic physiological conditions, HIF-1α is regulated by its hydroxylation at proline residues for subsequent binding to the von Hippel-Lindau tumour suppressor protein (pVHL), which mediates its degradation by the ubiquitin-proteosome complex [64]. A fundamental premise of the hyperactivation of the HIF-1α pathway under normoxic conditions is the sustained stimulation of growth signalling pathways such as PI3K/AKT, mTOR, Wnt/β-catenin and NOTCH. Meanwhile, the HIF-1α pathway, which is activated under anoxic conditions and is sensitive to ROS, regulates the expression of miRNA-210, miRNA-421, miRNA-382 and miRNA-687, miRNAs with abnormal expression in cancer [65,66,67]. This suggests a link between ROS activation of HIF-1α and abnormal expression of miRNAs associated with tumour progression. This phenotype may also occur in melanoma, where miRNA-mediated changes in HIF-1α and MITF expression have been reported [68,69].
αMSH regulates HIF-1α expression through cAMP. Given this specificity, it is likely that there are specific molecular mechanisms responsible for HIF-1α regulation in melanoma cells involving the previously described MITF signaling cascade. Using chromatin immunoprecipitation assays, it was concluded that MITF is a transcriptional target of HIF-1α through stimulation by cAMP [69]. Furthermore, in vivo and in vitro hypoxia signalling was identified as a negative regulator of MITF expression by finding that HIF-1α and MITF signalling were mostly closely correlated [68].
Several studies establish a clear relationship between some miRNAs and the HIF-1α signalling pathway in melanoma [70]. In 2015, Zhou et al. over-expressed miRNA-33a in the A375 cell line (amelanotic and metastatic) and in parallel inhibited the expression of this miRNA in the non-metastatic WM35 and metastatic WM45 cell lines; in this study, the authors demonstrated that miRNA-33a can inhibit proliferation, invasion and metastasis of human melanoma cells via inhibition of HIF-1α. Similarly, in mouse experiments, miRNA-33a was shown to inhibit both growth and metastasis of melanoma xenografts. These results provide a novel experimental basis for miRNA-33a to act as a tumour suppressor by inhibition of HIF-1α in human melanoma cell lines [30]. In 2019, Qiu et al. performed a study in A375 and WM35 cell lines and found that miRNA-138 levels were negatively correlated with HIF-1α mRNA levels and that in cell tissues with advanced metastatic stages, there was a decrease in miRNA-138 along with corresponding increased expression of HIF-1α activity, with an apparent antagonistic function. Reducing HIF-1α activity by RNA interference inhibited E-cadherin expression, increased vimentin levels and appeared to reverse the epithelial-to-mesenchymal transition process [71].
MAPK Signalling Pathway
One of the most important signalling pathways in melanoma is the MAPK pathway, a pathway that centralizes aerobic cellular metabolism and affects mitochondrial metabolism where there is an accumulation of reducing species such as NADH; it is also an activator of ERK kinase and promotes the expression of MITF via MEK [72]. Different activating mutations have been identified in melanoma subtypes of molecules that are part of this pathway, such as N-RAS, BRAF, MEK and ERK; in fact, more than half of the reported melanoma mutations correspond to BRAF V600E [73,74].
Under physiological conditions, once the GDP-GTP exchange with the RAS protein occurs, its conversion to its active form is promoted, the PI3K/AKT pathway is potentiated and the activity of the serine/threonine kinase RAF is stimulated, which phosphorylates and activates the tyrosine/threonine kinases MEK1 and MEK2 [75]. Next, MEK activates ERK1 and ERK2, then ERK translocate to the nucleus and activates cyclin CCND1, which forms the CCND1-CDK4/6 complex responsible for phosphorylating the RB1 protein, which in turn inactivates E2F, a growth factor in cell proliferation and metabolism [76,77].
As mentioned above, mutations in the BRAF gene occur in 50-60% of all melanomas, of which approximately 90% are V600E [78]; in this mutation, the amino acid glutamic acid (E) is substituted by valine (V) at position 600 of the protein [79]. In BRAF V600E mutations, there is increased kinase activity due to the lack of physiological negative feedback, leading to permanent activation of the MAPK pathway. The MAPK signalling cascade can also be hyperactivated by defects in the p16 tumour suppressor, which is responsible for the negative feedback of CCND1, leading to the loss of negative regulation of retinoblastoma protein (RB1) phosphorylation and hence the release of E2F mentioned above [77]. The characteristics of the MAPK pathway also include the down-regulation of MITF activity through the ubiquitin-proteasome system. However, due to the subsequent phosphorylation of p38, the MEK cascade can promote MITF expression leading to melanocyte differentiation and thus pathological melanogenesis.
The MAPK pathway has been described to be affected not only by ligand-receptor interactions but also by various stressors in the cell. Oxidative stress caused by ROS can induce activation of this pathway by ERK, JNK or p38, but the mechanisms by which ROS can activate these kinases are not well defined [80]. Evidence in cancer suggests that oxidative stress induces activation of an epidermal growth factor (EGF) receptor via phosphorylation of receptor tyrosine kinases (RTKs), of which hydrogen peroxide is a mediator for independent phosphorylation. Some approaches focus on the molecular mechanisms underlying JNK and p38 activation by redox-sensitive proteins such as thioredoxin and glutaredoxin, following the link between the ability of ROS to oxidize thioredoxin to dissociate from ASK-1, leading to subsequent activation of the JNK and p38 pathways [81].
The involvement of the MAPK pathway in processes such as proliferation, invasion and migration in melanoma cells in parallel with the expression of specific miRNAs has been reported in several studies, and it has been suggested that miRNA-340 is able to regulate MAPK signalling by reducing the expression of phosphorylated Erk1/2. This suggests that miRNA-340 appears to be a modulator of this pathway [82]. There is also evidence that oncogenic MAPK signalling stimulates the miRNA-29 family, specifically p53-dependent miRNA-29b2/c transcription; however, it has also been proposed that levels of this miRNA decrease during melanoma progression. miRNA-29 and MAPK could act as tumour suppressors by targeting MAFG and MYBL2 [83]. At present, the role of miRNA-29 is not clear, although it could act as a tumour suppressor as its expression decreases in parallel with tumour progression, but it could also be a possible oncogene, being stimulated by MAPK and playing a role in melanogenesis [83].
PI3K- AKT Signalling Pathway
The PI3K/AKT signalling pathway is involved in cell survival and metabolic regulation. Hyperactivation of the pathway is present in more than 50% of melanomas as a result of AKT3 amplification and subsequent loss of the tumour suppressor PTEN through epigenetic silencing or deletion [84]. AKT signalling is initiated by PI3K upon stimulation by exogenous growth factors, together with PIP expression, which promotes translocation of AKT to the plasma membrane for activation by phosphorylation. The intracellular level of PIP3 is negatively regulated by the PTEN phosphatase, and deficiency of this tumour suppressor induces positive regulation of PIP3 and promotes AKT activation. Studies using RNA interference targeting AKT3 or PTEN mutations reduced the tumorigenic potential of melanoma cells [42]. It is possible that increased cellular metabolism may affect the production of free radicals, which cells then use as signalling molecules to regulate MAPK and PI3K-AKT pathways to adapt to increased ROS levels and subsequent oxidative stress.
Tumour cells can positively regulate the KEAP1/NRF2 pathway, a key pathway for sensing and responding to oxidative stress. Thus, high levels of ROS and antioxidant defense pathways are present in the cancer cell to ensure its survival and metabolic needs [43]. Evidence in cancer suggests that hydrogen peroxide-induced oxidative stress activates a PIP3-dependent signalling mechanism through inactivation of the previously described tumour suppressor PTEN [85].
Previous studies have shown that miRNA-425 is able to inhibit cell proliferation by inducing apoptosis in A375 and SK-MEL-28 cells. Subsequent luciferase assays together with bioinformatics analysis have identified IGF-1 as a target gene of miRNA-425, which in turn inhibits the IGF-1-activated PI3K-AKT pathway. This suggests that miRNA-425 is able to inhibit melanoma progression through IGF-1 [86]. PCB2 is an oncogene that promotes tumorigenesis and metastasis; experimental evidence shows that in melanoma there is negative regulation of miRNA-5195-3p and positive regulation of PCBP2. It has been shown that miRNA-5195-3p inhibits PI3K/AKT activation in melanoma by inhibiting this gene [87].
Wnt Signalling Pathway
The Wnt signalling pathway plays a fundamental role in the differentiation and proliferation of melanocytes in the embryonic stage and in colon, breast and prostate cancer [48]. In melanoma, its role is controversial because the levels of the pathway mediator, β-catenin, decrease as cell proliferation progresses in an inverse relationship [88]. This pathway is activated by a canonical axis linked to the growth and transformation of melanoma cells [48], through the specific binding of Wnt1a and 3a proteins to the FZD (Frizzled,)/LRP (low-density lipoprotein receptor) receptor complex, which after anchoring to the destruction complex formed by the Axin protein, Adenomatous polyposis coli (APC), glycogen synthase kinase-3 (GSK-3) and casein kinase-1 (CK1), allowing β-catenin to be released into the nucleus where it acts as a coactivator in the TCF (T-cell transcription factor/LEF (lymphocyte enhancer factor) complex, regulating the expression of genes involved in cell proliferation [88]. It has even been shown that β-catenin itself is a regulator, as is the MAPK pathway acting on MITF, which is fundamental in controlling cell proliferation, survival and differentiation in melanoma.
The non-canonical axis is associated with metastasis and acts independently of β-catenin. Wnt5a forms a receptor complex with FZD that activates JNK/PKC and calcium, which can transactivate Jun and NFAT response genes involved in cell cycle regulation and contribute to survival. Similarly, the Wnt a5 ligand can down-regulate LEF-1 by favoring cell invasion through retention of β-catenin and promoting its degradation by ubiquitination through up-regulation of Siah2 [89].
The association of this pathway with various miRNAs in processes such as cell proliferation, migration and apoptosis in melanoma has been investigated. Wang et al. in 2017 demonstrated that miRNA-136 is able to inhibit the aforementioned processes in the B16 cell line via downregulation of pre-melanosome protein (PMEL) through downregulation of the Wnt pathway, suggesting that Wnt is critical for tumour survival [90]. Oncomirs involved in the positive regulation of Wnt have also been reported, such as miRNA-25, which was overexpressed in the MV3 cell line and downregulated the tumour suppressor and Wnt regulator DKK3 [91]. On the other hand, Wnt suppression affects tumorigenesis; Shi and his group proposed in 2019 that miRNA-22 targets FMNL2, a gene whose protein would be related to metastasis and tumour progression, and by inhibiting it suppresses melanoma development and down-regulates Wnt [92].
The positive regulation of Wnt is not only due to the expression of specific miRNAs, but also to the presence of an environment with increased oxidative stress from ROS. Wu and co-workers suggest that Wnt activation requires NOX1, a protein that promotes an oxidative stress environment and inhibits nucleoredoxin, a suppressor of the Wnt pathway [93], suggesting that activation of the Wnt pathway is stimulated by ROS and by the expression of specific miRNAs that promote cell survival and proliferation in melanoma.
NRF2/KEAP1 Wnt Signalling Pathway
In melanoma development, proliferation, invasion and survival, there is evidence for increased expression of erythroid nuclear related factor 2 (NRF2) [94], a fundamental player in the antioxidant response that ensures tumour survival to ROS damage [47]. Under physiological conditions, it is negatively regulated by Kelch-like ECH-associated protein 1 (KEAP) to maintain its basal level in the cytoplasm through degradation by the ubiquitin-proteosome system [95]. This regulatory protein has three characteristic domains in which IVR triggers the response after sensing redox reactions by oxidizing, allowing NRF2 to dissociate from the regulatory complex and migrate to the nucleus where it dimerizes with the small protein homologue of the aponeurotic muscle fibrosarcoma oncogene (sMaf); This heterodimer then binds to the antioxidant response element (ARE) within the DNA, triggering transcription at target genes and recruitment of other transcriptional activators [94].
However, other mechanisms are known by which NRF2 contributes to melanogenesis, as it inhibits MITF, developing dedifferentiated and invasive melanoma [95], and exerts a function on the redox capacity of melanoma, being more expressed in advanced, metastatic and drug-resistant melanoma by nuclear accumulation without affecting KEAP1 levels in the cytoplasm, performing a positive regulation of NRF2 target genes [94,95,96]. A study in A375 cells and G361 cells showed that non-thermal plasma (NTP)-induced cell damage and caspase activity in G361 cells decreased in the presence of cytoglobin through activation of the NRF2 pathway, suggesting that cytoglobin expression and the presence of melanin increase the resistance of melanotic melanoma cells to oxidative stress damage by activation of antioxidant systems [97]. Nrf2/Keap1 upon exposure to ROS leads to translocation of Nrf2 to the nucleus, resulting in an increase in antioxidant gene expression and thus a decrease in ROS levels [98,99].
The role of the NRF2/KEAP1 pathway in melanoma development is twofold: on the one hand, its potential antioxidant function reduces the likelihood of cancer; however, there is evidence that this pathway is also used as a mechanism of resistance to ROS damage in melanoma. The pathway is known to be associated with the expression of miRNAs identified as potential tumour suppressors such as miRNA-29, miRNA-181c and miRNA-200c, but also with the expression of potential oncogenes such as miRNA-193b-365, miRNA-32 and miRNA-592 [100].
Expression Networks and Regulation under Oxidative Stress-Induced Melanoma
The study of signalling pathways provides insight into the molecular mechanisms involved in melanoma development and progression, including alterations in ROS-induced oxidative stress. However, more recent approaches integrate functional enrichment by downstream signalling pathways into the construction of gene regulatory networks between miRNAs, genes and transcription factors, facilitating the abstraction of information in a visual format that is often intuitive and interpretable of non-linear regulation exerted by miRNAs [101,102]. As an example of this approach, regulatory and co-expression networks were constructed from the GSE109378 dataset reported in the Gene Expression Omnibus - GEO [103], in which changes in gene expression were evaluated by next generation sequencing (NGS) in the human melanoma cell line (SKMEL29), with silencing of miR-211 (SK-P8-2) and xenografts in each line, and their potential association with changes in ROS levels, mitochondrial respiration, and cell growth and invasion processes [104].
From these data, differential expression analysis was performed using the SAM and LIMMA libraries (Bioconductor) in R code [105,106]. The data were normalized in the VSN library (Bioconductor) by stabilizing the means and making the samples comparable using a Log2 transformation. For log-fold change estimation, the expression of those mRNAs with a p-value, corrected for multiple testing, of less than 5% was defined as differential [106]. The comparison was first performed between the SKMEL29 and SK-P8-2 lines and between the respective xenografts, and then the differentially expressed mRNAs were compared between lines and xenografts. Initially, 1,300 mRNAs with differential expression were obtained and, after manual curation, 30 of them were related to oxidative stress processes; finally, 6 were found to be differentially expressed. The intensity values of the mRNAs were visualized in heatmaps using the “pheatmap” function in the R code, taking the average value for each of the selected mRNAs in the cell line samples (Cell line) and xenograft samples (Xeno). The color and intensity of the squares were used to represent changes (absolute values) in expression [107].
As shown in Figure 1, MAFF and TFEB mRNAs are upregulated in both cell line and Xeno samples. MAFF mRNA has been implicated in melanogenesis by forming a heterodimer with NRF3 in response to stress [108], whereas the TFEB gene would cross-regulate with MITF in cellular clearance pathways [109,110] and induce melanoma growth by participating in metabolic regulation and ERK1/2 activation [111]. ZEB1, NFE2 and DYRK1B mRNAs were found to be decreased in both conditions. The transcription factor ZEB1 is a known inducer of mesenchymal–epithelial transition and invasiveness; in melanoma, elevated ZEB1 levels are associated with resistance to treatment with MAPK inhibitors [112], while the ZEB1/ZEB2 ratio would mediate phenotypic plasticity [57,112,113]. The transcription factor NFE2 is involved in megakaryocyte production [114], although it has not been reported to be altered in melanoma, its regulation by Nrf2 would promote the accumulation of ROS on megakaryocytes [115].
Dual-specificity tyrosine phosphorylation-regulated kinase 1B-DYRK1B is activated by the RAF-MEK1/2-ERK1/2 signalling pathway in melanoma and would be related to the promotion of cell differentiation [116]. The only gene that showed differential expression in the conditions studied was the T-cell intracellular antigen 1 gene, which encodes the TIA1 protein, an RNA-binding protein associated with nucleolytic activity against cytotoxic lymphocyte target cells, which has been proposed to be an oncogene localized in the cytoplasm of esophageal squamous cell carcinoma cells and to promote the expression of the progression-related genes SKP2 and CCNA2 [117]. It is possible that the increase in TIA1 in xenografts derived from miR-211 deletion lines compared to deletion-only cell lines reflects the influence of media-influenced tumour growth on the expression of tumour progression-related genes.
Using the String platform, the co-expression network was constructed (Figure 2), where the proteins encoded by the genes over- and under-expressed in both conditions are related to signalling pathways associated with oxidative stress, response to ROS, response to cytoplasmic stress and some of the pathways mentioned above, such as the PI3K-AKT-mTOR signalling pathway.
Figure 2. shows several distinct nodes. The node shown in blue/purple, formed by TIA1, G3BP1 and TIAL1, is associated with the formation of stress granules (SGs), cytoplasmic structures induced in response to environmental stress, mainly viral infection [118]; the formation of SGs appears to play an important role in the progression of several types of cancer by inhibiting apoptosis in response to stress [119,120]; although in melanoma the presence of the retinoblastoma binding (RB) mRNA Rbfox2 in SGs was associated with progression and metastasis [121], the presence of TIA1, G3BP1 and TIAL1 in melanoma-derived SGs has not been reported, so new experimental approaches would be needed.
Another highly interconnected node is the one shown in white, formed by DYRK1B, DCAF7 and CTBP1/2. DCAF7 and DYRK1B form a complex associated with the regulation of processes such as cell proliferation, differentiation and survival; it has been proposed that DCAF7 acts as an adaptor protein capable of mediating the binding and subsequent phosphorylation of E1A by DYRK1A/B, triggering a suppressive effect on proliferation through negative regulation of the transcriptional co-repressor CTBP [122]. In addition, elevated levels of CTBP1 in melanoma have been associated with increased proliferation and DNA damage [123], although by sequencing CTBP1 mRNA was not found to be differentially expressed in the two conditions evaluated, it is possible that by qPCR differences related to the decrease in DYRK1B could be found (Figure 2.).
The most highly connected node contains the proteins NFE2, MAF and NFE2L2, which have been implicated in chaperone-mediated autophagy, a process of selective degradation of cytosolic proteins in response to oxidative stress [124]; It has been reported that NRF2 has the function of regulating the antioxidant response, either by promoting tumor resistance to oxidative stress damage or by modulating carcinogenesis [100], while MAFF has been evaluated as a tumor suppressor and its interaction with NRF2 may suggest a protective antioxidant response of the healthy melanocyte.
Finally, proteins associated with PI3K-AKT-mTOR are shown in pink. In 2015, Hambright et al. demonstrated in different cell lines the key role of PI3K/AKT/mTOR signalling and upregulation of the antioxidant system in ensuring the survival of melanoma cells. Altering redox homeostasis by increasing oxidative stress in melanoma cells inhibited PI3K/AKT/mTOR signalling by disrupting mTORC1 formation, thereby reducing colony formation and cell proliferation [43]. However, the interaction between mTOR, ROS and the antioxidant response is not fully understood, as these ROS may play a dual role by possessing both activating and inhibitory functions [125].
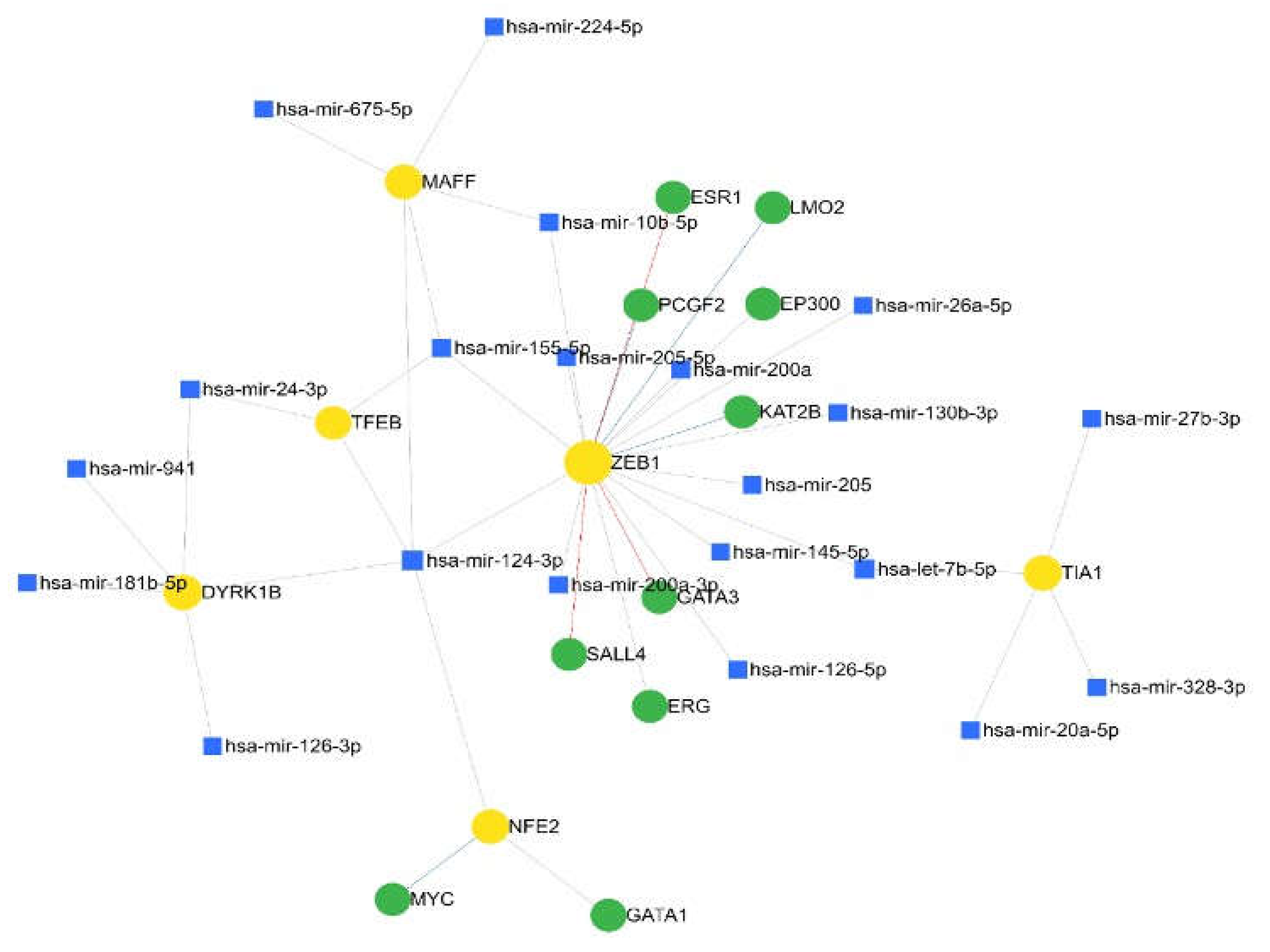
For the construction of regulatory network models, we used miRNet (https://www.mirnet.ca/), a freely available network-based web tool that integrates several statistical tools, data mining and visualization systems for the integrated study of miRNA-molecular target interaction [107,108]. In addition to implementing a flexible interface for filtering, refining and customizing data during network construction, miRNet includes a network visualization system with the possibility of functional enrichment analysis. The tool was fed with the IDs of the over- and under-expressed mRNAs from Figure 1 and for functional enrichment the KEGG and GO pathways were used with statistical analysis of a hypergeometric test.
In Figure 3, the transcription factor ZEB1 (zinc-fingered E-box binding homeobox 1), best known for its involvement in mesenchymal epithelial transition processes, cancer cell differentiation, progression and metastasis [56,126] through the regulation of proteins such as SALL4 and GATA3 [127,128], which were also found to be related in the network, is the hub with the highest number of interactions. Its involvement in modulating the oxidative stress environment has been described in breast cancer models through the promotion of MCT4 and GPX4 [129,130]; in melanoma, it has been associated with increased resistance to MAPK inhibitors, promoting immune evasion [112,131]. It is possible that the highly defined DNA- and protein-binding domains explain the large number of interactions represented in the hub and the high modularity exerted. MAFF has been postulated as a possible tumor suppressor regulated by miRNA-224-5p in hepatocellular carcinoma [132].
Among the miRNAs that potentially modulate ZEB1 in the proposed network, we can highlight some that have been reported in the literature to act on other molecular targets in melanoma. The miR-126, which inhibits invasion and migration in cervical cancer cells by binding to ZEB1 [117]; the miR-126-3p isoform, which was found to be associated with the transcription factor DYRK1B, is involved in acquired resistance to dabrafenib in melanoma cells by regulating ADAM9 and VEGF-A [133]. The miR-200a-3p has been reported as a potential regulator of melanogenesis through direct binding to MITF [134]. miR-205-5p functions as a negative suppressor of the PI3K/AKT pathway in renal cancer [133]. Another node described is miR-224-5p and MAFF, this miRNA has been reported as an oncomiR and biomarker in several types of cancer [135,136], additionally miR-224-5p binding to MAFF occurs in hepatocellular carcinoma [132].
In the network, one of the miRNAs that interacts with multiple targets is mir 24-3p, which has been associated with negative regulation in metastatic cancer, in B16F10 melanoma cells ectopic expression could be generated in melanoma, which would generate suppression in cell migration, mir-24-3p generates the suppression of cancer cell growth, the overexpression of this mir-24-3p could generate a decrease in cell viability, as well as the inhibition of cell migration and invasion. mir 24-3p could control cell proliferation by regulating hydrofolate reductase [137].
Meanwhile, exosomal mir-155-5p would be related to the pro-angiogenic onset of Caf (cancer-associated fibroblasts), as it could be delivered to fibroblasts and generate angiogenic factors such as vascular endothelial growth factor and fibroblast growth factor 2. The B16F10 cell line, which is a metastatic melanoma line, produces exosomal secretions with the ability to induce reprogramming of fibroblasts into Caf and the expression of tumour angiogenesis markers. These exosomes suppress SOSC1 expression, leading to activation of the JAK2/STAT3 pathway, which in turn regulates a pathogenic switch that increases vascular endothelial growth factor and fibroblast growth factor 2 expression in fibroblasts [138].
Likewise, TIA1 is modulated by miR-27b, among others, which is widely associated with melanoma development, as described in 2021 by Yi Tian and collaborators, where they determined expression levels in melanoma cells and normal tissues by immunoelectrotransfer, correlating a negative regulation with MYC, a gene related to the Wnt-β catenin pathway, a pathway of importance in melanogenesis described previously, as a target in this case [139]. Another modulator detected in the network is mir 20a- 5p, also described in recent years as a tumour suppressor in different tissues, demonstrated in 2019 by Ahred and his team in B16 melanoma cells, where a suppression of the same was detected in comparison with non-malignant keratinocytes [140].
Conclusion
Melanoma remains a cancer with high mortality and therapeutic resistance, necessitating the development of new strategies and effective diagnostic and therapeutic targets for diagnosis and treatment. Oxidative stress is a common factor in this and other cancers, not only providing an optimal environment for tumorigenesis, but also actively participating in the regulation of metabolic pathways and gene regulation at the post-transcriptional level through the expression of miRNAs, and possibly in the regulation of the entire tumour machinery in its progression and metastasis. As described in this paper, the role of oxidative stress is fundamental in the maintenance and regulation of tumors and is probably a common factor in all types of cancer. Carcinogenesis and tumour progression in melanoma involve cooperative and synergistic mechanisms linking genes, miRNAs and transcription factors. The lack of research confirming specific interactions of ROS through the regulation of miRNA expression or other factors in the activation of signalling pathways limits the projection and implications of oxidative stress modulation.
The discrete relationships cited throughout the paper, albeit of limited functional association, suggest the important role that miRNAs and ROS play through the activation of signalling pathways in melanoma tumour pathogenesis and progression. The network approaches presented in the third section facilitate the highly complex understanding of the tumour microenvironment and the imperative need to construct networks that integrate not only miRNAs, genes and signalling pathways, but also all the factors and cofactors that stimulate other as yet unreported and possibly important processes that condition the poorly documented modulation.
Acknowledgments
The authors thanks the Universidad del Bosque.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Piskounova E, Agathocleous M, Murphy MM, Hu Z, Huddlestun SE, Zhao Z, et al. Oxidative stress inhibits distant metastasis by human melanoma cells. Nature 2015;527:186–91. [CrossRef]
- Ralph SJ, Rodríguez-Enríquez S, Neuzil J, Moreno-Sánchez R. Bioenergetic pathways in tumor mitochondria as targets for cancer therapy and the importance of the ROS-induced apoptotic trigger. Mol Aspects Med 2010;31:29–59. [CrossRef]
- Venza I, Venza M, Visalli M, Lentini G, Teti D, d’Alcontres FS. ROS as Regulators of Cellular Processes in Melanoma. Oxid Med Cell Longev 2021;2021:1–19. [CrossRef]
- Pozzobon FC, Acosta ÁE, Castillo JS. Cáncer de piel en Colombia: cifras del Instituto Nacional de Cancerología. Revista de La Asociación Colombiana de Dermatología y Cirugía Dermatológica 2018;26. [CrossRef]
- IARC, Internacional Agency for Research on cancer. Cancer Today 2020. https://gco.iarc.fr/today/online-analysis-table?v=2020&mode=cancer&mode_population=continents&population=900&populations=904&key=asr&sex=0&cancer=39&type=0&statistic=5&prevalence=0&population_group=0&ages_group%5B%5D=0&ages_group%5B%5D=17&group_cancer=1&include_nmsc=1&include_nmsc_other=1#collapse-group-0-1 (accessed on 28 June 2023).
- Tormo, R. Estudio de los perfiles de expresión de micrornas en líneas celulares de cancer de mama triple negativo tratadas con doxorubicina: implicación de la familia mir-449 2017.
- Garzon R, Calin GA, Croce CM. MicroRNAs in Cancer. Annu Rev Med 2009;60:167–79. [CrossRef]
- Yang C, Yan Z, Hu F, Wei W, Sun Z, Xu W. Silencing of microRNA-517a induces oxidative stress injury in melanoma cells via inactivation of the JNK signaling pathway by upregulating CDKN1C. Cancer Cell Int 2020;20:32. [CrossRef]
- Schadendorf D, van Akkooi ACJ, Berking C, Griewank KG, Gutzmer R, Hauschild A, et al. Melanoma. The Lancet 2018;392:971–84. [CrossRef]
- Bennett, DC. Genetics of melanoma progression: the rise and fall of cell senescence. Pigment Cell Melanoma Res 2016;29:122–40. [CrossRef]
- Akbani R, Akdemir KC, Aksoy BA, Albert M, Ally A, Amin SB, et al. Genomic Classification of Cutaneous Melanoma. Cell 2015;161:1681–96. [CrossRef]
- Hodis E, Watson IR, Kryukov GV, Arold ST, Imielinski M, Theurillat J-P, et al. A Landscape of Driver Mutations in Melanoma. Cell 2012;150:251–63. [CrossRef]
- Shain AH, Yeh I, Kovalyshyn I, Sriharan A, Talevich E, Gagnon A, et al. The Genetic Evolution of Melanoma from Precursor Lesions. New England Journal of Medicine 2015;373:1926–36. [CrossRef]
- Liu S, Howell PM, Riker AI. Up-Regulation of miR-182 Expression after Epigenetic Modulation of Human Melanoma Cells. Ann Surg Oncol 2013;20:1745–52. [CrossRef]
- Glud M, Gniadecki R. MicroRNAs in the pathogenesis of malignant melanoma. Journal of the European Academy of Dermatology and Venereology 2013;27:142–50. [CrossRef]
- Kunz, M. MicroRNAs in Melanoma Biology, 2013, p. 103–20. [CrossRef]
- Mione M, Bosserhoff A. Micro RNA s in melanocyte and melanoma biology. Pigment Cell Melanoma Res 2015;28:340–54. [CrossRef]
- Ameres SL, Zamore PD. Diversifying microRNA sequence and function. Nat Rev Mol Cell Biol 2013;14:475–88. [CrossRef]
- Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol 2014;15:509–24. [CrossRef]
- Pritchard CC, Cheng HH, Tewari M. MicroRNA profiling: approaches and considerations. Nat Rev Genet 2012;13:358–69. [CrossRef]
- Price C, Chen J. MicroRNAs in cancer biology and therapy: Current status and perspectives. Genes Dis 2014;1:53–63. [CrossRef]
- Wang D, Qiu C, Zhang H, Wang J, Cui Q, Yin Y. Human MicroRNA Oncogenes and Tumor Suppressors Show Significantly Different Biological Patterns: From Functions to Targets. PLoS One 2010;5:e13067. [CrossRef]
- Luna Buitrago D, Lovering RC, Caporali A. Insights into Online microRNA Bioinformatics Tools. Noncoding RNA 2023;9:18. [CrossRef]
- Chao J, Guo Y, Li P, Chao L. Role of Kallistatin Treatment in Aging and Cancer by Modulating miR-34a and miR-21 Expression. Oxid Med Cell Longev 2017;2017:1–7. [CrossRef]
- Zhang X, Ng W-L, Wang P, Tian L, Werner E, Wang H, et al. MicroRNA-21 Modulates the Levels of Reactive Oxygen Species by Targeting SOD3 and TNF α. Cancer Res 2012;72:4707–13. [CrossRef]
- Yadav P, Sharma P, Sundaram S, Venkatraman G, Bera AK, Karunagaran D. SLC7A11/ xCT is a target of miR-5096 and its restoration partially rescues miR-5096-mediated ferroptosis and anti-tumor effects in human breast cancer cells. Cancer Lett 2021;522:211–24. [CrossRef]
- Li S-Z, Hu Y-Y, Zhao J, Zhao Y-B, Sun J-D, Yang Y, et al. MicroRNA-34a induces apoptosis in the human glioma cell line, A172, through enhanced ROS production and NOX2 expression. Biochem Biophys Res Commun 2014;444:6–12. [CrossRef]
- Díaz Arce, D. Revista cubana de investigaciones biomédicas. vol. 23. Editorial Ciencias Médicas; 2004.
- Andreucci E, Ruzzolini J, Bianchini F, Versienti G, Biagioni A, Lulli M, et al. miR-214-Enriched Extracellular Vesicles Released by Acid-Adapted Melanoma Cells Promote Inflammatory Macrophage-Dependent Tumor Trans-Endothelial Migration. Cancers (Basel) 2022;14:5090. [CrossRef]
- Zhou J, Xu D, Xie H, Tang J, Liu R, Li J, et al. miR-33a functions as a tumor suppressor in melanoma by targeting HIF-1α. Cancer Biol Ther 2015;16:846–55. [CrossRef]
- CHEN Y, CAO K, WANG S, CHEN J, HE B, HE G, et al. MicroRNA-138 suppresses proliferation, invasion and glycolysis in malignant melanoma cells by targeting HIF-1α. Exp Ther Med 2016;11:2513–8. [CrossRef]
- Boyle GM, Woods SL, Bonazzi VF, Stark MS, Hacker E, Aoude LG, et al. Melanoma cell invasiveness is regulated by miR-211 suppression of the BRN2 transcription factor. Pigment Cell Melanoma Res 2011;24:525–37. [CrossRef]
- Völler D, Ott C, Bosserhoff A. MicroRNAs in malignant melanoma. Clin Biochem 2013;46:909–17. [CrossRef]
- Noguchi S, Kumazaki M, Yasui Y, Mori T, Yamada N, Akao Y. MicroRNA-203 Regulates Melanosome Transport and Tyrosinase Expression in Melanoma Cells by Targeting Kinesin Superfamily Protein 5b. Journal of Investigative Dermatology 2014;134:461–9. [CrossRef]
- Shirvani H, Ghanavi J, Aliabadi A, Mousavinasab F, Talebi M, Majidpoor J, et al. MiR-211 plays a dual role in cancer development: From tumor suppressor to tumor enhancer. Cell Signal 2023;101:110504. [CrossRef]
- Donadelli M, Dando I, Fiorini C, Palmieri M. Regulation of miR-23b expression and its dual role on ROS production and tumour development. Cancer Lett 2014;349:107–13. [CrossRef]
- Kozak J, Jonak K, Maciejewski R. The function of miR-200 family in oxidative stress response evoked in cancer chemotherapy and radiotherapy. Biomedicine & Pharmacotherapy 2020;125:110037. [CrossRef]
- Cui Y, She K, Tian D, Zhang P, Xin X. miR-146a Inhibits Proliferation and Enhances Chemosensitivity in Epithelial Ovarian Cancer via Reduction of SOD2. Oncology Research Featuring Preclinical and Clinical Cancer Therapeutics 2016;23:275–82. [CrossRef]
- Pflaum J, Schlosser S, Müller M. p53 Family and Cellular Stress Responses in Cancer. Front Oncol 2014;4. [CrossRef]
- Dar AA, Majid S, Rittsteuer C, de Semir D, Bezrookove V, Tong S, et al. The Role of miR-18b in MDM2-p53 Pathway Signaling and Melanoma Progression. JNCI: Journal of the National Cancer Institute 2013;105:433–42. [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov 2022;12:31–46. [CrossRef]
- Irvine M, Stewart A, Pedersen B, Boyd S, Kefford R, Rizos H. Oncogenic PI3K/AKT promotes the step-wise evolution of combination BRAF/MEK inhibitor resistance in melanoma. Oncogenesis 2018;7:72. [CrossRef]
- Hambright HG, Meng P, Kumar AP, Ghosh R. Inhibition of PI3K/AKT/mTOR axis disrupts oxidative stress-mediated survival of melanoma cells. Oncotarget 2015;6:7195–208. [CrossRef]
- Fraga A, Ribeiro R, Medeiros R. Hipoxia tumoral: Papel del factor inducible por hipoxia. Actas Urol Esp 2009;33. [CrossRef]
- Joshi S, Singh AR, Durden DL. MDM2 Regulates Hypoxic Hypoxia-inducible Factor 1α Stability in an E3 Ligase, Proteasome, and PTEN-Phosphatidylinositol 3-Kinase-AKT-dependent Manner. Journal of Biological Chemistry 2014;289:22785–97. [CrossRef]
- Malekan M, Ebrahimzadeh MA, Sheida F. The role of Hypoxia-Inducible Factor-1alpha and its signaling in melanoma. Biomedicine & Pharmacotherapy 2021;141:111873. [CrossRef]
- Carpenter EL, Becker AL, Indra AK. NRF2 and Key Transcriptional Targets in Melanoma Redox Manipulation. Cancers (Basel) 2022;14:1531. [CrossRef]
- Xue G, Romano E, Massi D, Mandalà M. Wnt/β-catenin signaling in melanoma: Preclinical rationale and novel therapeutic insights. Cancer Treat Rev 2016;49:1–12. [CrossRef]
- Varrone F, Caputo E. The miRNAs Role in Melanoma and in Its Resistance to Therapy. Int J Mol Sci 2020;21:878. [CrossRef]
- Lister JA, Capper A, Zeng Z, Mathers ME, Richardson J, Paranthaman K, et al. A Conditional Zebrafish MITF Mutation Reveals MITF Levels Are Critical for Melanoma Promotion vs. Regression In Vivo. Journal of Investigative Dermatology 2014;134:133–40. [CrossRef]
- Hartman ML, Czyz M. MITF in melanoma: mechanisms behind its expression and activity. Cellular and Molecular Life Sciences 2015;72:1249–60. [CrossRef]
- Park HY, Kosmadaki M, Yaar M, Gilchrest BA. Cellular mechanisms regulating human melanogenesis. Cellular and Molecular Life Sciences 2009;66:1493–506. [CrossRef]
- Thompson JF, Scolyer RA, Kefford RF. Cutaneous melanoma. The Lancet 2005;365:687–701. [CrossRef]
- Levy C, Khaled M, Robinson KC, Veguilla RA, Chen P-H, Yokoyama S, et al. Lineage-Specific Transcriptional Regulation of DICER by MITF in Melanocytes. Cell 2010;141:994–1005. [CrossRef]
- Hsiao JJ, Fisher DE. The roles of microphthalmia-associated transcription factor and pigmentation in melanoma. Arch Biochem Biophys 2014;563:28–34. [CrossRef]
- Bruneel K, Verstappe J, Vandamme N, Berx G. Intrinsic Balance between ZEB Family Members Is Important for Melanocyte Homeostasis and Melanoma Progression. Cancers (Basel) 2020;12:2248. [CrossRef]
- Denecker G, Vandamme N, Akay Ö, Koludrovic D, Taminau J, Lemeire K, et al. Identification of a ZEB2-MITF-ZEB1 transcriptional network that controls melanogenesis and melanoma progression. Cell Death Differ 2014;21:1250–61. [CrossRef]
- TargetScanHuman 8.0 n.d. https://www.targetscan.org/vert_80/ (accessed 29 March 2024).
- Segura MF, Hanniford D, Menendez S, Reavie L, Zou X, Alvarez-Diaz S, et al. Aberrant miR-182 expression promotes melanoma metastasis by repressing FOXO3 and microphthalmia-associated transcription factor. Proceedings of the National Academy of Sciences 2009;106:1814–9. [CrossRef]
- Bemis LT, Chen R, Amato CM, Classen EH, Robinson SE, Coffey DG, et al. MicroRNA-137 Targets Microphthalmia-Associated Transcription Factor in Melanoma Cell Lines. Cancer Res 2008;68:1362–8. [CrossRef]
- Mazar J, DeYoung K, Khaitan D, Meister E, Almodovar A, Goydos J, et al. The Regulation of miRNA-211 Expression and Its Role in Melanoma Cell Invasiveness. PLoS One 2010;5:e13779. [CrossRef]
- Margue C, Philippidou D, Reinsbach SE, Schmitt M, Behrmann I, Kreis S. New Target Genes of MITF-Induced microRNA-211 Contribute to Melanoma Cell Invasion. PLoS One 2013;8:e73473. [CrossRef]
- De Luca T, Pelosi A, Trisciuoglio D, D’Aguanno S, Desideri M, Farini V, et al. miR-211 and MITF modulation by Bcl-2 protein in melanoma cells. Mol Carcinog 2016;55:2304–12. [CrossRef]
- Mills CN, Joshi SS, Niles RM. Expression and function of hypoxia inducible factor-1 alpha in human melanoma under non-hypoxic conditions. Mol Cancer 2009;8:104. [CrossRef]
- Serocki M, Bartoszewska S, Janaszak-Jasiecka A, Ochocka RJ, Collawn JF, Bartoszewski R. miRNAs regulate the HIF switch during hypoxia: a novel therapeutic target. Angiogenesis 2018;21:183–202. [CrossRef]
- Seok J-K, Lee SH, Kim MJ, Lee Y-M. MicroRNA-382 induced by HIF-1α is an angiogenic miR targeting the tumor suppressor phosphatase and tensin homolog. Nucleic Acids Res 2014;42:8062–72. [CrossRef]
- Bhatt K, Wei Q, Pabla N, Dong G, Mi Q-S, Liang M, et al. MicroRNA-687 Induced by Hypoxia-Inducible Factor-1 Targets Phosphatase and Tensin Homolog in Renal Ischemia-Reperfusion Injury. Journal of the American Society of Nephrology 2015;26:1588–96. [CrossRef]
- Feige E, Yokoyama S, Levy C, Khaled M, Igras V, Lin RJ, et al. Hypoxia-induced transcriptional repression of the melanoma-associated oncogene MITF. Proceedings of the National Academy of Sciences 2011;108. [CrossRef]
- Buscà R, Berra E, Gaggioli C, Khaled M, Bille K, Marchetti B, et al. Hypoxia-inducible factor 1α is a new target of microphthalmia-associated transcription factor (MITF) in melanoma cells. J Cell Biol 2005;170:49–59. [CrossRef]
- Shen G, Li X, Jia Y, Piazza GA, Xi Y. Hypoxia-regulated microRNAs in human cancer. Acta Pharmacol Sin 2013;34:336–41. [CrossRef]
- Qiu H, Chen F, Chen M. MicroRNA-138 negatively regulates the hypoxia-inducible factor 1α to suppress melanoma growth and metastasis. Biol Open 2019;8. [CrossRef]
- Špaková I, Rabajdová M, Mičková H, Graier WF, Mareková M. Effect of hypoxia factors gene silencing on ROS production and metabolic status of A375 malignant melanoma cells. Sci Rep 2021;11:10325. [CrossRef]
- Ascierto PA, Kirkwood JM, Grob J-J, Simeone E, Grimaldi AM, Maio M, et al. The role of BRAF V600 mutation in melanoma. J Transl Med 2012;10:85. [CrossRef]
- Poulikakos PI, Zhang C, Bollag G, Shokat KM, Rosen N. RAF inhibitors transactivate RAF dimers and ERK signalling in cells with wild-type BRAF. Nature 2010;464:427–30. [CrossRef]
- McKay MM, Morrison DK. Integrating signals from RTKs to ERK/MAPK. Oncogene 2007;26:3113–21. [CrossRef]
- Held L, Eigentler TK, Meier F, Held M, Röcken M, Garbe C, et al. Oncogenetics of melanoma: basis for molecular diagnostics and therapy. JDDG: Journal Der Deutschen Dermatologischen Gesellschaft 2011;9:510–6. [CrossRef]
- Wang J, Shen WH, Jin YJ, Brandt-Rauf PW, Yin Y. A Molecular Link between E2F-1 and the MAPK Cascade. Journal of Biological Chemistry 2007;282:18521–31. [CrossRef]
- Cheng L, Lopez-Beltran A, Massari F, MacLennan GT, Montironi R. Molecular testing for BRAF mutations to inform melanoma treatment decisions: a move toward precision medicine. Modern Pathology 2018;31:24–38. [CrossRef]
- Colombino M, Capone M, Lissia A, Cossu A, Rubino C, De Giorgi V, et al. BRAF/NRAS Mutation Frequencies Among Primary Tumors and Metastases in Patients With Melanoma. Journal of Clinical Oncology 2012;30:2522–9. [CrossRef]
- Son Y, Cheong Y-K, Kim N-H, Chung H-T, Kang DG, Pae H-O. Mitogen-Activated Protein Kinases and Reactive Oxygen Species: How Can ROS Activate MAPK Pathways? J Signal Transduct 2011;2011:1–6. [CrossRef]
- Matsuzawa A, Ichijo H. Redox control of cell fate by MAP kinase: physiological roles of ASK1-MAP kinase pathway in stress signaling. Biochimica et Biophysica Acta (BBA) - General Subjects 2008;1780:1325–36. [CrossRef]
- Poenitzsch Strong AM, Setaluri V, Spiegelman VS. microRNA-340 as a modulator of RAS–RAF–MAPK signaling in melanoma. Arch Biochem Biophys 2014;563:118–24. [CrossRef]
- Vera O, Bok I, Jasani N, Nakamura K, Xu X, Mecozzi N, et al. A MAPK/miR-29 Axis Suppresses Melanoma by Targeting MAFG and MYBL2. Cancers (Basel) 2021;13:1408. [CrossRef]
- Madhunapantula S, V. , Robertson GP. The PTEN-AKT3 signaling cascade as a therapeutic target in melanoma. Pigment Cell Melanoma Res 2009;22:400–19. [CrossRef]
- Leslie, NR. Redox regulation of PI 3-kinase signalling via inactivation of PTEN. EMBO J 2003;22:5501–10. [CrossRef]
- Liu P, Hu Y, Ma L, Du M, Xia L, Hu Z. miR-425 inhibits melanoma metastasis through repression of PI3K-Akt pathway by targeting IGF-1. Biomedicine & Pharmacotherapy 2015;75:51–7. [CrossRef]
- Yang B, Wu Y, Chen Y, Li Y, Wang J, Cha X, et al. MiR-5195-3p targets the PCBP2/PI3K/AKT pathway to inhibit melanoma cell proliferation and migration. Heliyon 2023;9:e19227. [CrossRef]
- Atkinson JM, Rank KB, Zeng Y, Capen A, Yadav V, Manro JR, et al. Activating the Wnt/β-Catenin Pathway for the Treatment of Melanoma – Application of LY2090314, a Novel Selective Inhibitor of Glycogen Synthase Kinase-3. PLoS One 2015;10:e0125028. [CrossRef]
- Mao J, Wang J, Liu B, Pan W, Farr GH, Flynn C, et al. Low-Density Lipoprotein Receptor-Related Protein-5 Binds to Axin and Regulates the Canonical Wnt Signaling Pathway. Mol Cell 2001;7:801–9. [CrossRef]
- Wang J-J, Li Z-F, Li X-J, Han Z, Zhang L, Liu Z-J. Effects of microRNA-136 on melanoma cell proliferation, apoptosis, and epithelial–mesenchymal transition by targetting PMEL through the Wnt signaling pathway. Biosci Rep 2017;37. [CrossRef]
- Huo J, Zhang Y, Li R, Wang Y, Wu J, Zhang D. Upregulated MicroRNA-25 Mediates the Migration of Melanoma Cells by Targeting DKK3 through the WNT/β-Catenin Pathway. Int J Mol Sci 2016;17:1124. [CrossRef]
- L Shi, J-W Huo, S-S Chen, J-X Xue, W-Y Gao, X-Y Li, et al. MicroRNA-22 targets FMNL2 to inhibit melanoma progression via the regulation of the Wnt/β-catenin signaling pathway and epithelial-mesenchymal transition. Eur Rev Med Pharmacol Sci 2019.
- Wu Y, Antony S, Meitzler JL, Doroshow JH. Molecular mechanisms underlying chronic inflammation-associated cancers. Cancer Lett 2014;345:164–73. [CrossRef]
- Malakoutikhah Z, Mohajeri Z, Dana N, Haghjooy Javanmard S. The dual role of Nrf2 in melanoma: a systematic review. BMC Mol Cell Biol 2023;24:5. [CrossRef]
- Raghunath A, Sundarraj K, Arfuso F, Sethi G, Perumal E. Dysregulation of Nrf2 in Hepatocellular Carcinoma: Role in Cancer Progression and Chemoresistance. Cancers (Basel) 2018;10:481. [CrossRef]
- Jessen C, Kreß JKC, Baluapuri A, Hufnagel A, Schmitz W, Kneitz S, et al. The transcription factor NRF2 enhances melanoma malignancy by blocking differentiation and inducing COX2 expression. Oncogene 2020;39:6841–55. [CrossRef]
- De Backer J, Lin A, Berghe W Vanden, Bogaerts A, Hoogewijs D. Cytoglobin inhibits non-thermal plasma-induced apoptosis in melanoma cells through regulation of the NRF2-mediated antioxidant response. Redox Biol 2022;55. [CrossRef]
- Wu S, Lu H, Bai Y. Nrf2 in cancers: A double-edged sword. Cancer Med 2019;8:2252–67. [CrossRef]
- Taguchi K, Yamamoto M. The KEAP1–NRF2 System in Cancer. Front Oncol 2017;7. [CrossRef]
- Shah NM, Rushworth SA, Murray MY, Bowles KM, MacEwan DJ. Understanding the role of NRF2-regulated miRNAs in human malignancies. Oncotarget 2013;4:1130–42. [CrossRef]
- Liu Y-Y, Slotine J-J, Barabási A-L. Controllability of complex networks. Nature 2011;473:167–73. [CrossRef]
- MacNeil LT, Walhout AJM. Gene regulatory networks and the role of robustness and stochasticity in the control of gene expression. Genome Res 2011;21:645–57. [CrossRef]
- Home - GEO - NCBI, n.d. https://www.ncbi.nlm.nih.gov/geo/ (accessed on 28 March 2024).
- Sahoo A, Sahoo SK, Joshi P, Lee B, Perera RJ. MicroRNA-211 Loss Promotes Metabolic Vulnerability and BRAF Inhibitor Sensitivity in Melanoma. Journal of Investigative Dermatology 2019;139:167–76. [CrossRef]
- Efron B, Tibshirani R, Storey JD, Tusher V. Empirical Bayes Analysis of a Microarray Experiment. J Am Stat Assoc 2001;96:1151–60. [CrossRef]
- Schwender H, Ickstadt K. Empirical Bayes analysis of single nucleotide polymorphisms. BMC Bioinformatics 2008;9:1–15. [CrossRef]
- Grant GR, Manduchi E, Stoeckert CJ. Analysis and Management of Microarray Gene Expression Data. Curr Protoc Mol Biol 2007;77. [CrossRef]
- Waku T, Nakada S, Masuda H, Sumi H, Wada A, Hirose S, et al. The CNC-family transcription factor Nrf3 coordinates the melanogenesis cascade through macropinocytosis and autophagy regulation. Cell Rep 2023;42:111906. [CrossRef]
- Ballesteros-Álvarez J, Dilshat R, Fock V, Möller K, Karl L, Larue L, et al. MITF and TFEB cross-regulation in melanoma cells. PLoS One 2020;15:e0238546. [CrossRef]
- Möller K, Sigurbjornsdottir S, Arnthorsson AO, Pogenberg V, Dilshat R, Fock V, et al. MITF has a central role in regulating starvation-induced autophagy in melanoma. Sci Rep 2019;9:1055. [CrossRef]
- Ariano C, Costanza F, Akman M, Riganti C, Corà D, Casanova E, et al. TFEB inhibition induces melanoma shut-down by blocking the cell cycle and rewiring metabolism. Cell Death Dis 2023;14:314. [CrossRef]
- Richard G, Dalle S, Monet M, Ligier M, Boespflug A, Pommier RM, et al. ZEB 1-mediated melanoma cell plasticity enhances resistance to MAPK inhibitors. EMBO Mol Med 2016;8:1143–61. [CrossRef]
- Durand S, Tang Y, Pommier RM, Benboubker V, Grimont M, Boivin F, et al. ZEB1 controls a lineage-specific transcriptional program essential for melanoma cell state transitions. BioRxiv 2023:2023.02.10.526467. [CrossRef]
- Gasiorek JJ, Blank V. Regulation and function of the NFE2 transcription factor in hematopoietic and non-hematopoietic cells. Cellular and Molecular Life Sciences 2015;72:2323–35. [CrossRef]
- Motohashi H, Kimura M, Fujita R, Inoue A, Pan X, Takayama M, et al. NF-E2 domination over Nrf2 promotes ROS accumulation and megakaryocytic maturation. Blood 2010;115:677–86. [CrossRef]
- Ashford AL, Dunkley TPJ, Cockerill M, Rowlinson RA, Baak LM, Gallo R, et al. Identification of DYRK1B as a substrate of ERK1/2 and characterisation of the kinase activity of DYRK1B mutants from cancer and metabolic syndrome. Cellular and Molecular Life Sciences 2016;73:883–900. [CrossRef]
- Hamada J, Shoda K, Masuda K, Fujita Y, Naruto T, Kohmoto T, et al. Tumor-promoting function and prognostic significance of the RNA-binding protein T-cell intracellular antigen-1 in esophageal squamous cell carcinoma. Oncotarget 2016;7:17111–28. [CrossRef]
- Garaigorta U, Heim MH, Boyd B, Wieland S, Chisari F V. Hepatitis C Virus (HCV) Induces Formation of Stress Granules Whose Proteins Regulate HCV RNA Replication and Virus Assembly and Egress. J Virol 2012;86:11043–56. [CrossRef]
- Takayama K, Suzuki T, Fujimura T, Takahashi S, Inoue S. Association of USP10 with G3BP2 Inhibits p53 Signaling and Contributes to Poor Outcome in Prostate Cancer. Molecular Cancer Research 2018;16:846–56. [CrossRef]
- Legrand N, Dixon DA, Sobolewski C. Stress granules in colorectal cancer: Current knowledge and potential therapeutic applications. World J Gastroenterol 2020;26:5223–47. [CrossRef]
- Choi S, Sa M, Cho N, Kim KK, Park S-H. Rbfox2 dissociation from stress granules suppresses cancer progression. Exp Mol Med 2019;51:1–12. [CrossRef]
- Glenewinkel F, Cohen MJ, King CR, Kaspar S, Bamberg-Lemper S, Mymryk JS, et al. The adaptor protein DCAF7 mediates the interaction of the adenovirus E1A oncoprotein with the protein kinases DYRK1A and HIPK2. Sci Rep 2016;6:28241. [CrossRef]
- Deng H, Liu J, Deng Y, Han G, Shellman YG, Robinson SE, et al. CtBP1 Is Expressed in Melanoma and Represses the Transcription of p16INK4a and Brca1. Journal of Investigative Dermatology 2013;133:1294–301. [CrossRef]
- Pajares M, Rojo AI, Arias E, Díaz-Carretero A, Cuervo AM, Cuadrado A. Transcription factor NFE2L2/NRF2 modulates chaperone-mediated autophagy through the regulation of LAMP2A. Autophagy 2018;14:1310–22. [CrossRef]
- Sagwal SK, Bekeschus S. ROS Pleiotropy in Melanoma and Local Therapy with Physical Modalities. Oxid Med Cell Longev 2021;2021:1–21. [CrossRef]
- Wu H-T, Zhong H-T, Li G-W, Shen J-X, Ye Q-Q, Zhang M-L, et al. Oncogenic functions of the EMT-related transcription factor ZEB1 in breast cancer. J Transl Med 2020;18:51. [CrossRef]
- Wei S, Zhong L, Wang X, Zhang W. Low expression of GATA3 promotes cell proliferation and metastasis in gastric cancer. Cancer Manag Res 2017;Volume 9:769–80. [CrossRef]
- Chen T, Tsang JYS, Su X, Li P, Sun W, Wong ILK, et al. SALL4 promotes tumor progression in breast cancer by targeting EMT. Mol Carcinog 2020;59:1209–26. [CrossRef]
- Han X, Duan X, Liu Z, Long Y, Liu C, Zhou J, et al. ZEB1 directly inhibits GPX4 transcription contributing to ROS accumulation in breast cancer cells. Breast Cancer Res Treat 2021;188:329–42. [CrossRef]
- Han X, Long Y, Duan X, Liu Z, Hu X, Zhou J, et al. ZEB1 induces ROS generation through directly promoting MCT4 transcription to facilitate breast cancer. Exp Cell Res 2022;412:113044. [CrossRef]
- Plaschka M, Benboubker V, Grimont M, Berthet J, Tonon L, Lopez J, et al. ZEB1 transcription factor promotes immune escape in melanoma. J Immunother Cancer 2022;10:e003484. [CrossRef]
- Wu M, Deng X, Zhong Y, Hu L, Zhang X, Liang Y, et al. MafF Is Regulated via the circ-ITCH/miR-224-5p Axis and Acts as a Tumor Suppressor in Hepatocellular Carcinoma. Oncology Research Featuring Preclinical and Clinical Cancer Therapeutics 2020;28:299–309. [CrossRef]
- Caporali S, Amaro A, Levati L, Alvino E, Lacal PM, Mastroeni S, et al. miR-126-3p down-regulation contributes to dabrafenib acquired resistance in melanoma by up-regulating ADAM9 and VEGF-A. Journal of Experimental & Clinical Cancer Research 2019;38:272. [CrossRef]
- Itoh T, Fukatani K, Nakashima A, Suzuki K. MicroRNA-141-3p and microRNA-200a-3p regulate α-melanocyte stimulating hormone-stimulated melanogenesis by directly targeting microphthalmia-associated transcription factor. Sci Rep 2020;10:2149. [CrossRef]
- Li A, Wu N, Sun J. E2F1-induced microRNA-224-5p expression is associated with hepatocellular carcinoma cell migration, invasion and epithelial-mesenchymal transition via MREG. Oncol Lett 2022;23:82. [CrossRef]
- Rana S, Valbuena GN, Curry E, Bevan CL, Keun HC. MicroRNAs as biomarkers for prostate cancer prognosis: a systematic review and a systematic reanalysis of public data. Br J Cancer 2022;126:502–13. [CrossRef]
- Kang H, Rho JG, Kim C, Tak H, Lee H, Ji E, et al. The miR-24-3p/p130Cas: a novel axis regulating the migration and invasion of cancer cells. Sci Rep 2017;7:44847. [CrossRef]
- Zhou X, Yan T, Huang C, Xu Z, Wang L, Jiang E, et al. Melanoma cell-secreted exosomal miR-155-5p induce proangiogenic switch of cancer-associated fibroblasts via SOCS1/JAK2/STAT3 signaling pathway. Journal of Experimental & Clinical Cancer Research 2018;37:242. [CrossRef]
- Tian Y, Zeng J, Yang Z. MicroRNA-27b inhibits the development of melanoma by targeting MYC. Oncol Lett 2021;21:370. [CrossRef]
- Stope M, Ahrend H, Daeschlein G, Grove E, Paditz M, Mustea A, et al. MicroRNA-20a-3p and microRNA-20a-5p exhibit anti-proliferative activities in a melanoma in vitro model. SDRP Journal of Cellular and Molecular Physiology 2019;3:1–10. [CrossRef]
Figure 1.
Heatmap of differentially expressed genes (DEGs) between the cell line depleted of miR-211 and the xenografts derived from these cell lines. The color and intensity of the squares represent changes (absolute values) in expression. Red (over expression) and green (under expression).
Figure 1.
Heatmap of differentially expressed genes (DEGs) between the cell line depleted of miR-211 and the xenografts derived from these cell lines. The color and intensity of the squares represent changes (absolute values) in expression. Red (over expression) and green (under expression).
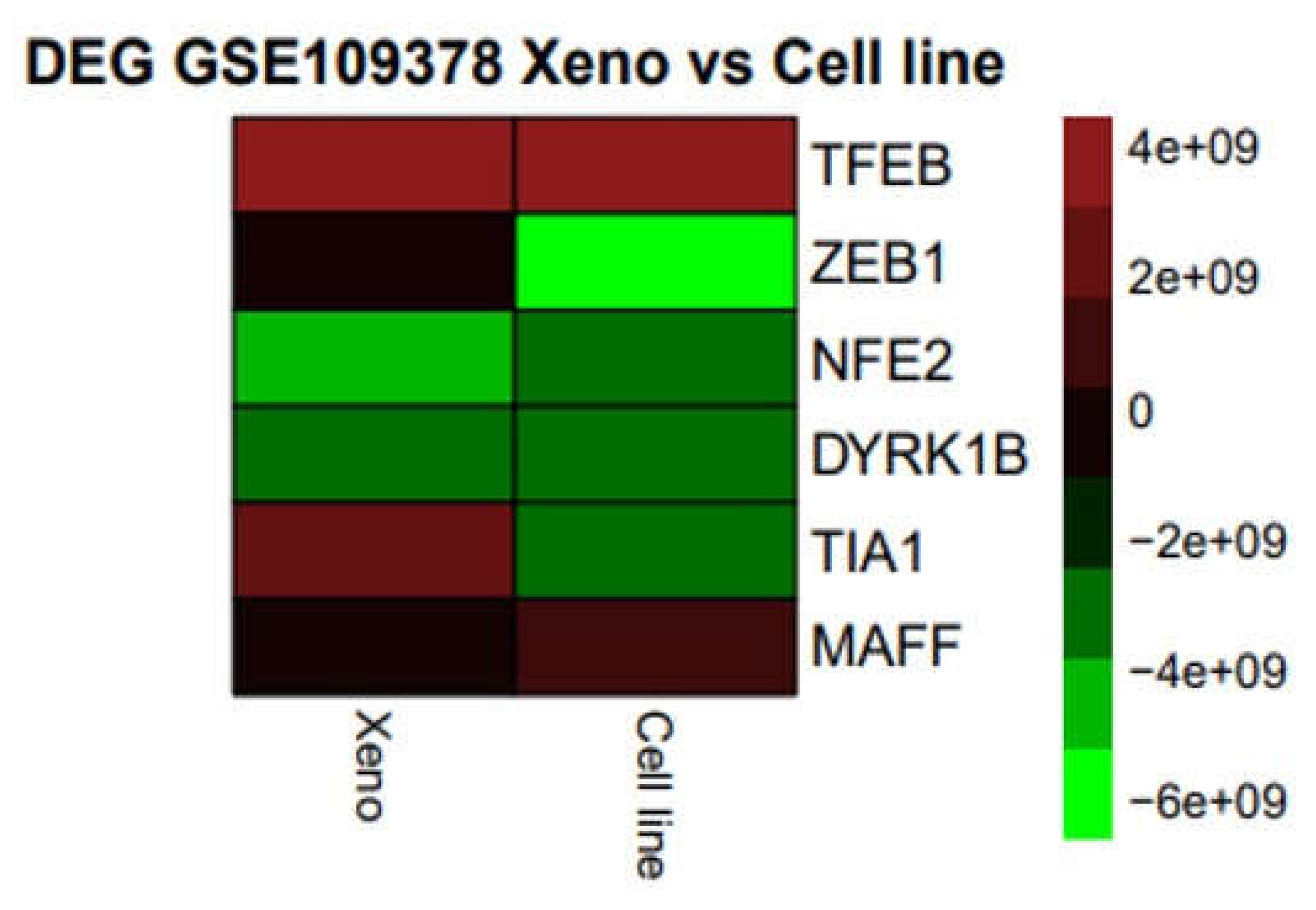
Figure 2.
Protein regulatory network associated with oxidative stress in melanoma.
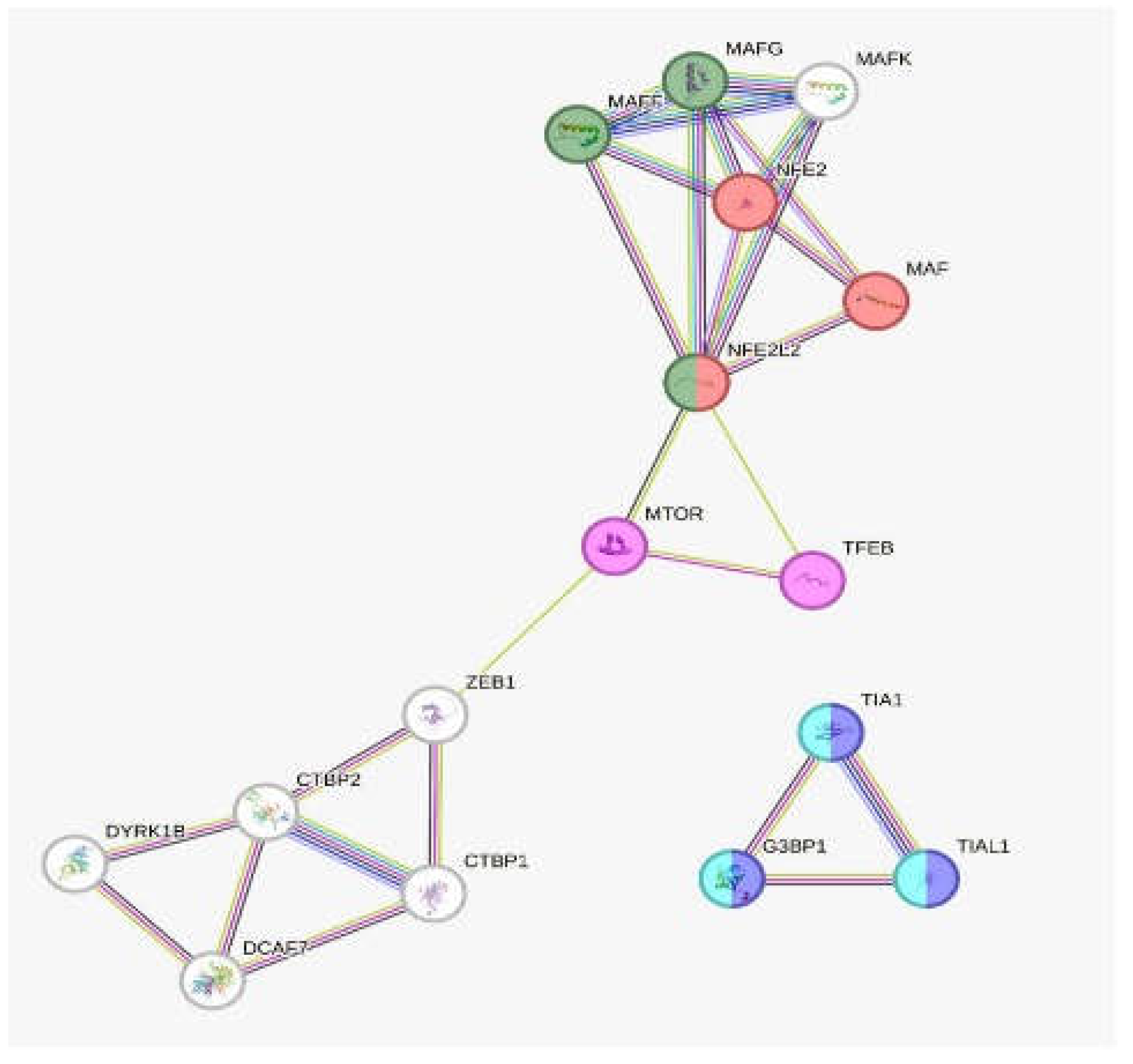
Figure 3.
Regulatory network of miRNAs, genes and transcription factors associated with oxidative stress in melanoma.
Figure 3.
Regulatory network of miRNAs, genes and transcription factors associated with oxidative stress in melanoma.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



