
Submitted:
26 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
Lipopeptides are bioactive compounds produced by Bacillus spp. This work provides insight into de antifungal activity of lipopeptides through a meta-analysis of published studies, in addition following PCR (polymerase chain reaction) technique, the presence of lipopeptides genes in 20 Bacillus strains were also analyzed. The meta-analysis of published studies showed that iturin, fengycin and surfactin significantly reduced the growth of phytopathogenic fungi. Similarly, when applied to plant tissues, the lipopeptides reduced the severity of damage in plant tissues caused by phytopathogenic fungi. As for the lipopeptides genes detection, we used 20 Bacillus strains reported to have antagonistic activity against various fungal plant pathogens. The PCR amplification study was carried out to detect the genes involved in the synthesis of lipopeptides: ituA for iturin A, fenD for fengycin and srfA for surfactin. Comparative sequence analysis showed a range of 98 to 100% identity with the corresponding genes in several Bacillus species. We observed at least one Bacillus gene in the strains: CBRF5, CBRF15, CBSN67, and CBRM9. In the strain CBRF6, we detected the presence of the fabD and srfAD genes. For the development of natural fungicides to control phytopathogenic fungi, lipopeptides produced by Bacillus spp. could be a useful tool.
Keywords:
lipopeptides
; Bacillus sp
; iturin A
; fengycin
; surfactin
1. Introduction
Microbial metabolism produces a wide arrange of bioactive molecules known as secondary metabolites, which include peptides, polyketides, carbohydrates, lipids, terpenoids, steroids and alkaloids that themselves are prepared from primary metabolites, linking these two branches of metabolism. The determination of the structure and function of microbial secondary metabolites has been studied for various decades owing their importance in pharmaceutical and food industry.[1]
There are many secondary metabolites, such as terpenes, flavonoids and peptides that are produced in some cases as protection against other microorganisms. One class of microbial secondary metabolites is the non-ribosomal peptides (NRPs), which are derived from proteinogenic and non-proteinogenic amino acids. NRPs are synthesized from large multidomain enzymes, the non-ribosomal peptide synthetases (NRPS).[2]
Cyclic lipopeptides are proteins synthesized from long multi-enzyme complexes. Their chemical structure includes a cyclic portion of a single fatty acid linked to 7 to 10 α-amino acids [3] Several species of Bacillus produce lipopeptides that function as antibiotics with a wide range of effectiveness against bacteria and fungi.[4] The lipopeptides produced by Bacillus spp. are categorized into three families comprising iturin, fengycin, and surfactin. The iturin family consists primarily of iturin A and C, bacillomycin D, F, L, and Lc, and mycosubtilin. These lipopeptides exhibit antifungal activity and hinder the growth of diverse pathogens in plants.[5] The fengycin family comprises mainly A and B fengycins that exhibit fungitoxic activity in filamentous fungi, while the surfactin family has low fungitoxic activity but shows synergistic activity with iturin or fengycin.[6] Biological activity studies have revealed conflicting outcomes with respect to the impact of lipopeptides on phytopathogenic fungi. E.[7] found that iturin suppresses the mycelium growth of Fusarium graminearum, whereas surfactin shows no influence on this variable.[8] discovered that surfactin hampers the expansion of the mycelium of the F. moniliforme fungus. The inconsistency in the antifungal efficacy results for the lipopeptides warrants a re-analysis of the compound’s impact via meta-analysis of published studies. Such an analysis could prove valuable in identifying general trends, specific patterns, and conclusive information regarding the effects on phytopathogenic fungi.[9]
The critical role of lipopeptides in suppressing fungal plant pathogens requires a distribution of the genes produced by certain Bacillus species.[10] The ability of B. subtilis to produce antibiotics is the result of the high percent (4 and 5 %) of its genome dedicated to biosynthesis, potentially giving rise to over two dozen antimicrobial peptides (AMPs).[11] Bacteria devote up to 20% of their genome to the synthesis of secondary metabolites with a broad spectrum of activity and diverse structures, as revealed by their genome sequencing.[12] These metabolites are inclusive of gene-encoded antibiotics as well as numerous non-ribosomal synthesized peptides.[13]
The iturin A operon consists of four open reading frames (ORFs): ituD, ituA, ituB and ituC. The ituD gene shares homology with the malonyl-CoA trans-acylase protein FAbD, which is involved in fatty acid synthesis in E. coli [14] and B. subtilis 168.[15] The second gene, ituA, encodes a 449 kDa protein with three functional modules homologous to fatty acid synthetase, amino acid transferase and peptide synthetase. The third gene, ituB, and the fourth gene, ituC, encode peptide synthetases of 609 and 297 kDa, respectively, which form peptide chains in the ituA precursor.
Five genes involved in the biosynthesis of fengycin have been reported in a 37 kb DNA fragment: fenC, fenD, fenE, fenA and fenB. Among these genes, fenC encodes a fengycin synthetase of 2560 amino acids with a mass of 287 KDa. This protein contains two amino acid activation modules, FenC1 and FenC2, which activate L-glutamic acid and L-ornithine. While FenB activates L-isoleucine. This enzyme also contains a thioesterase domain, suggesting that FenB is involved in the termination of fengycin synthesis.[16] Other genes are involved in the production of fengycin, such as the pps group. Among the five pps genes, ppsE encodes a protein with 74% sequence identity to fenE in fengycin synthetase.[17]
Surfactin biosynthesis in Bacillus is encoded by the srf operon. This operon comprises four open reading frames encoding the srfA and srfD protein components of the surfactin synthetase multienzyme system.[18] This lipopeptide is synthesized through the interaction of three enzymatic fractions, E1 and E2 (Surfactin synthetase), and E3 (Acyltransferase enzyme). Fraction E1 contains the surfactin synthetase enzymes SrfA and SrfB, each comprising three amino acid activation modules. SrfA thioesterifies L-Glu and two leucine residues, while SrfB incorporates L-Asp, L-Val, and L-Leu as thioesters; Likewise, srfB contains the genes comP and comA that comprise a signal translation system involved in the pathway of competence development and is necessary for the transcription of srfA.19 The SrfC enzyme contributes to the C-terminal L-Leucine residue. Regarding the role of the external thioesterase enzyme SrfD (SrfTE-II) is encoded by the fourth srfD open reading frame of the srf-operon,[18] suggesting that srfD functions as the thioesterase/acyltransferase enzyme in the initiation process of surfactin formation.[19]
The objective of the present study was to conduct a meta-analysis of published studies on the effect of lipopeptides on mycelial growth and damage suppression in plant tissues. In addition, we also detected the presence of biosynthetic genes such as ituA, iturin A, fenD, fengycin, and srfA surfactin in native strains of Bacillus spp., using specific primers for the polymerase chain reaction (PCR).
2. Materials and Methods
2.1. Data Collection for Lipopeptide Antifungal Activity Meta-Analysis
For the meta-analysis, we reviewed literature on the efficacy of lipopeptides (iturin, surfactin and fengycin) from Bacillus spp. on phytopathogenic fungi. We included studies that investigated at least one of the following aspects: 1) impacts of lipopeptides on phytopathogenic fungi in vitro and in vivo, 2) effectiveness of iturin, surfactin and fengycin against phytopathogenic fungal species.
The search for articles was conducted using Google Scholar and the databases Science Direct, Springer Journal, Taylor & Francis, and Wiley Online Library. The keywords used individually or in combination were: lipopeptides, biological effectiveness, iturin, surfactin, fengycin, in vitro activity, in vivo activity; these were also combined with the terms Fusarium spp., Colletotrichum spp. and Alternaria spp.
Data were included if the studies met these criteria: 1) studies conducted after 2000, 2) studies assessing the effectiveness of iturin, surfactin and fengycin in vitro and in vivo on phytopathogenic fungi, and 3) studies evaluating lipopeptide iturin’s impact-related aspects. Data from the same study, which covered numerous lipopeptides, several species of phytopathogenic fungi, and various types of plants, evaluated at different times over multiple years, using different application methods and lipopeptide concentrations, were treated distinctly and regarded as independent cases. The analysis assessed the inhibition of fungal growth (%) using in vitro test of the radial mycelial growth in Petri dish containing solid growth medium and severity of damage to plant organs (leaves, roots, fruits, etc) was taken as score of area damaged or the degree of damage using specific diagrammatic scale for assessment the severity. A total of forty articles were included in the meta-analysis. Of these, only 10 met the criteria (Table S1). This resulted in twenty-seven studies on fungal growth inhibition (%) and thirty studies on severity of plant tissue damage. For the meta-analysis, the ability of lipopeptides to inhibit fungal growth in vitro is indicated by positive values in the variable fungal growth inhibition (%) and the ability of lipopeptides to reduce the severity of plant tissue damage is indicated by negative values in the variable plant tissue damage.
2.2. Effect Sizes
The effects of lipopeptides on inhibition of fungal growth and severity of damage to plant tissues caused by phytopathogenic fungi were evaluated using a ratio (response quotient) of the effect between the groups treated with lipopeptides (experimental group) and that of a group not exposed to lipopeptides (control group). As a result, negative values in the response ratio indicate a reduction in the severity of damage to plant tissue, whereas for inhibition of fungal growth (%), a positive value indicates an increase in the percentage of inhibition produced by the lipopeptides. To calculate the effect size for each variable, the mean, standard deviation, and sample size were obtained for the control and experimental groups. All standard error values were converted to standard deviation according to the equation SD = SE √ n, where SD is the standard deviation, SE is the standard error, and n is the sample size.
2.3. Data Analysis
A separate analysis was carried out to evaluate the effect of the lipopeptides iturin, surfactin, fengycin and their concentrations. The variables analyzed were inhibition of fungal growth (%) and severity of plant tissue damage. The effect size was calculated using Hedges: d = [(XO-XY) / s] J, where XO is the mean response of the control group, XY is the mean response of the experimental group, s is the pooled SD and J is the bias correction factor due to the small sample size. The analysis was performed in the Meta Win 2.1 statistical program using fixed effects models. Confidence intervals were generated at the 95% confidence level (bias-corrected bootstrap) for all effect sizes from 999 iterations. Effects will be considered significant when the 95% confidence intervals do not overlap with zero. To evaluate whether the categorical groups (inhibition of fungal growth (%) and severity of damage to plant tissue were homogeneous with respect to effect size, the heterogeneity within each group (QW) and between the groups (QB) were calculated and the importance was evaluated by X2.
The risk of bias in the dataset was assessed using the Rosenberg method[20] by the number of certainty (nfs). The nfs indicates the number of non-significant, unpublished, or missing studies that would have to be added to the meta-analysis to change the results from significant to non-significant. Results are considered robust if the nfs is greater than 5n + 10, where n is the number of studies.[21]
2.4. Bacillus Strains under Study for Detection of Lipopeptide Biosysntesis Genes
Bacillus strains from the microorganism collection of the Microbiology Laboratory of the Conkal Institute of Technology were used, which have been reported to have antifungal activity,[22] ten B. subtilis strains (CBRF4, CBRF12, CBRF15, CBCK36, CBCK47, CBMT2, CBMT51, CBMN22, CBCC2 and CBSN67), two B. amyloliquefaciens strains (CBRF6 and CBRF11), one B. cereus strain (CBCK41), one Peanibacillus sp. (CBRM17) and five Bacillus strains not identified to species level (CBCK44, CBCK46, CBRM9, CBRF5 and CBDG60). The Bacillus subtilis strain CBRF8 was used as a reference for the detection of genes encoding the production of lipopeptides such as iturin A and surfactin.[22] The strains were activated on nutrient agar and stored at 4 °C until use. In this work no confirmation of the lipopeptide production by the Bacillus strains was performed, as this task was not part of the study objective.
2.5. DNA Extraction of Bacillus Strains
Chromosomal DNA was extracted using the Wizard® Genomic DNA Purification Kit (Promega, USA). DNA quantification was performed in a Nanodrop Lite (Thermo Scientific®) and DNA integrity was visualized on 1% agarose gels. The extracted DNA samples were stored at -20°C until use.
2.6. Detection and Sequence Analysis of Lipopeptide Biosynthesis Genes
PCR amplifications were performed in a thermocycler (Techne TC-312) using primers specific for lipopeptide biosynthesis genes[23] (Table S2). The final volume of the reaction mixture was 50 μL, containing 50 ng DNA, 2.0 mM Mg+2 solution for ITUD1F/ITUD1R and FEND1F/FEND1R and 1.5 mM for SUR3F/SUR3R, 0.2 mM dNTP mixture, 0. 5 µM (for ITUD1F/ITUD1R and FEND1F/FEND1R) and 1.0 µM (for SUR3F/SUR3R) primers) and Taq DNA Polymerase 0.5 U (Invitrogen) for ITUD1F/ITUD1R and FEND1F/FEND1R) and 2.0 U (Invitrogen) for SUR3F/SUR3R. The amplification conditions were as follows: (i) ITUD1F/ITUD1R with an initial denaturation temperature of 3 min at 94 °C, 40 cycles each consisting of denaturation for 1 min at 94 °C, annealing for 1 min at 60 °C and extension for 1 min at 72 °C with a final extension step of 10 min at 72 °C. (ii) FEND1F/FEND1R with an initial denaturation temperature of 3 minutes at 94 °C, 45 cycles each consisting of denaturation for 1 minute at 94 °C, annealing for 1 minute at 62 °C and extension for 1.45 minutes at 72 °C with a final extension step of 6 minutes at 72 °C. (iii) SUR3F/SUR3R with an initial denaturation temperature of 3 minutes at 94 °C, 40 cycles each consisting of denaturation for 1 minute at 94 °C, annealing for 1 minute at 57 °C and extension for 1 minute at 72 °C with a final extension step of 10 minutes at 72 °C. PCR products were analyzed by SYBR™ Green staining on 1% agarose gels. (ii) FEND1F/FEND1R with an initial denaturation temperature of 3 minutes at 94 °C, 45 cycles each consisting of denaturation for 1 minute at 94 °C, annealing for 1 minute at 62°C and extension for 1.45 minutes at 72 °C with a final extension step of 6 minutes at 72°C. (iii) SUR3F/SUR3R with an initial denaturation temperature of 3 min at 94 °C, 40 cycles each consisting of denaturation for 1 min at 94 °C, annealing for 1 min at 57 °C and extension for 1 min at 72 °C with a final extension step of 10 min at 72 °C. PCR products were analyzed by SYBR™ Green staining on 1% agarose gels. (ii) FEND1F/FEND1R with an initial denaturation temperature of 3 minutes at 94 °C, followed by 45 cycles, each comprising denaturation for 1 minute at 94 °C, annealing for 1 minute at 62 °C and extension for 1.45 minutes at 72 °C, with a final extension step of 6 minutes at 72 °C. (iii) Use of SUR3F/SUR3R proceeded with an initial denaturation temperature of 3 minutes at 94 °C, followed by 40 cycles consisting of denaturation for 1 minute at 94 °C, annealing for 1 minute at 57 °C and extension for 1 minute at 72 °C, with a final extension step of 10 minutes at 72 °C. PCR products were analyzed on 1% agarose gels using SYBR™ Green staining. (iii) SUR3F/SUR3R with an initial denaturation temperature of 3 minutes at 94 °C, 40 cycles each consisting of denaturation for 1 minute at 94 °C, annealing for 1 minute at 57 °C and extension for 1 minute at 72 °C with a final extension step of 10 minutes at 72 °C. PCR products were examined using 1% agarose gels with SYBR™ Green staining. Once again, PCR products were analyzed on 1% agarose gels by SYBR™ Green staining. The amplifications were achieved using SUR3F/SUR3R with an initial denaturation temperature of 3 minutes at 94 °C, followed by 40 cycles of denaturation for 1 minute at 94 °C, annealing for 1 minute at 57 °C and extension for 1 minute at 72 °C with a final extension step of 10 minutes at 72 °C.
2.7. Analysis of Gene Sequences
Each of the PCR products were sequenced in sense and antisense by the company MacrogenUSA, (Maryland). Each of the sense and anti-sense sequences were aligned to obtain consensus, using the CAP3 program.[24] The BLASTn software (https://blast.ncbi.nlm.nih.gov/Blast.cgi, access 15 April 2024) and in UniProt (https://www.uniprot.org/blast/, access 15 April 2024), of the National Center for Biotechnology Information (NCBI)was used to analyze the percentage identity of the sequences.
3. Results
The results of this research are related first to the systematic search for published studies on the effect of lipopeptides on phytopathogenic fungi, followed by the detection of lipopeptides genes in Bacillus strains.
3.1. General Effects of Lipopeptides on Phytopathogenic Fungi
In general, lipopeptides were found to have significant effects on inhibiting fungal growth (%) and reducing the severity of damage to plant tissues caused by phytopathogenic fungi (Figure 1; Table 1). For the inhibition of fungal growth, exposure to lipopeptides resulted in an effect size of 4.04 (CI = 2.29 to 9.96, bias 95%), which represents a significant effect of the peptides compared to control. Additionally, it was found that the lipopeptides caused a significant decrease on the severity of damage to plant tissue, with an effect size of -1.10 (CI = -1.92 to -0.84, bias 95%).
3.2. Meta-Analysis of the Inhibition of Fungal Growth In Vitro and Severity of Damage to Plants
For the inhibition of fungal growth, the three lipopeptides studied (iturin, fengycin and surfactin) have significant effects. The size of the effect produced by surfactin is like that of fengycin, but greater than that of iturin (Figure 2). In the analysis of the effects by concentration ranges of the lipopeptides (data not shown), both at concentrations of 50 to 200 µg ml-1 and 200 to 400 µg ml-1 the lipopeptides have significant effects. The effect size of 3.95 (CI = 2.05 to 8.41, bias 95%) is not significantly different between the two concentrations analyzed. For the decrease in severity of damage to plant tissue, surfactin has the greater effect, but it was not significant from those of fengycin and iturin (Figure 3).
3.3. Detection and Analysis of Lipopeptide Biosynthesis Genes
Out of the 20 strains that were analyzed for the presence of the ituA, fenD and srfA genes, only strains CBRF5, CBRF6, CBSN67, CBRM9 and CBRF15 displayed the presence of lipopeptide biosynthesis genes in Table 2. The sizes of the PCR product were as expected when compared to the control strain B. subtilis CBRF8, with 647 bp for ituA, 964 bp for fenD, and 441 bp for srfA.
The fabD gene was identified for the biosynthesis iturin and fengycin, whereas the srf gene was identified for surfactin production. Sequences were analyzed using NCBI web page and UniProt blastx program, identifying the ACP-S malonyl transferase and surfactin synthetase proteins responsible for synthesizing the cyclic lipopeptides iturin A, fengycin, and surfactin. These findings are detailed in Table 3.
The CBRM9 strain displayed homology with the ituD gene, which codes for the protein named Iturin A synthetase D of B. amyloquefaciens. However, it also exhibited a 99.5% similarity with the ACP-malonyl transferase protein (fabD gene) from B. valezensis (UniProtKB-A0A6A8LHM2).
4. Discussion
The present study consisted of two sections, in the first section a meta-analysis of published studies of the effect of lipopeptides on phytopathogenic fungi was performed. Two variables were considered in the meta-analysis: in vitro inhibition of fungal growth and suppression of plant tissue damage caused by phytopathogenic fungi. In the second section, PCR analysis was conducted to detect the presence of the genes involved in the synthesis of lipopeptides (iturin, fengycin and surfactin) in native strains of Bacillus spp.
The meta-analysis of antifungal activity of lipopeptides revealed two key findings: Firstly, all three lipopeptides (iturin, fengycin and surfactin) inhibit in vitro mycelial growth, with a similar effect across a concentration range of 50 to 400 µg ml-1. Secondly, all three lipopeptides have suppressive effects on the severity of damage caused by phytopathogenic fungi to plants. It should be noted that in the meta-analysis, the effect size produced by surfactin was greater in general than those of fengycin and iturin. However, in various studies, the lipopeptides iturin and fengycin showed higher effects compared to surfactin on certain fungal species.[25,26] The meta-analysis in the present work may have overlooked this effect as it combined the average effect on various species of fungi. The lipopeptides then may have species specificity when targeting phytopathogenic fungi. Several fungal species from different classes were evaluated.[27] The results showed that iturin had effect on all species evaluated, whereas surfactin only had effect on the deuteromycetes Botrytis cinerea and Colletotrichum capsici, as well as the pseudofungi Phytophtora infestans. No effect of surfactin was observed in the basidiomycete Puccinia graminis or in the ascomycete Erysiphe graminis, indicating that surfactin selectively targets certain species.
Regarding the analysis of lipopeptide biosynthesis genes, of the 20 Bacillus spp. strains analyzed, the presence of these genes was detected in only five strains. Four strains showed a single gene, and one strain (CBRF6) showed two genes (fabD and srf). The above is consistent with results of other studies, where the presence of a single gene per strain was reported.[12,28,29] In the present work, the most frequently detected gene was that of iturin synthesis. Similar results were obtained by Gond et al.[30] and Sarangi et al.,[12] who performed gene and lipopeptide prospecting in strains of Bacillus spp. In contrast, Athukorala et al.[10] found that most strains produced surfactins, followed by iturin; Stankovic et al.[29] reported the bacillomycin synthesis gene as the most prevalent, followed by surfactin. Therefore, it is considered that there is no defined pattern of prevalence of any type of gene in Bacillus spp. strains.
In the analysis carried out on each of the three genes studied, in the case of iturin A, the amplification of the region of the fabD gene responsible for the formation of the ACP-malonyl transferase protein was observed. This is an enzyme of the transferase class (EC:2.3.1.39) and its sequence is reported as the genes bmyD (Uni-ProtKB - Q5KN8 and Q70JZ7), bamD (A0A1B1SP49) and fabD (UniProtKB-A7Z5A6). In the case of the protein identified in the CBRM9 strain, which showed 100% homology with iturin A synthetase D from B. amyloquefaciens, it also showed 99.5% similarity with the protein ACP-malonyltransferase (gene fabD) from B. valezensis (Uni-ProtKB-A0A6A8LHM2). This protein has been reported in B. velenzensis (WP_060561613.1), B. subtilis (WP_088110826.1) and B. amyloquefaciens (WP_079004866.1). Other Bacillus species that harbors lipopeptide synthesis genes include B. mycoides,[10] B. cereus, B. tequilensis, B. weihenstephanensis and B. licheniformis.[12] New iturin W with potent antifungal activity have been reported.[31]
Regarding the synthesis of fengycin, the presence of the fabD gene was detected in the CBRF15 strain, as already mentioned, which is responsible for the production of the ACP-malonyl transferase protein reported in B. thuringiensis (WP_102956566.1). Similar genes have been reported in B. weihenstephanensis, B. licheniformis, B. thuringiensis, B. cereus [12] and B. amyloliquefaciens.[32]
In the case of surfactin synthesis, the srf gene was identified, which was present in the strain B. amyloliquefaciens CBRF6.[28] The SrfD protein functions as a thioesterase/acyltransferase enzyme in the initiation process of surfactin synthesis.[33] This protein has been reported in Bacillales (WP_012116775.1) and biosynthetic genes in isolates of B. weihenstephanensis, B. tequilensis, B. thuringiensis, B. subtilis and B. cereus.[12]
The above findings highlight the diversity of Bacillus species, harboring lipopeptide biosynthesis genes responsible for producing iturin A, fengycin and surfactin. Additionally, certain strains possess multiple lipopeptides, as reported by wang et al.[32]
To elucidate the phylogenetic relationships between the deduced protein sequence of iturin A, surfactin, and fengycin of Bacillus strains and the described homologous sequences, a phylogenetic analysis was performed. Distinctive clades of the proteins iturin A, surfactin and fengycin were observed (Figure S1). Interestingly iturin A of CBRF5 strain is closely related to iturin A of CBRF6 (B. amyloliquefaciens) strain and forms an outgroup. Iturin A of CBRM9 strain appears to be more closely related to iturin A of CBRF6, whereas fengycin of CBRF15 (B. subtilis) clusters more closely to surfactin of CBRF6 (B. amyloliquefaciens).
The antifungal properties of the lipopeptides iturin, fengycin and surfactin have been evaluated in numerous phytopathogenic fungi species. Notably, evaluations on fungi of the deuteromycetes class are prevalent; some of the economically significant species include Colletotrichum gloeosporioides,[34] Fusarium spp.[7] and Botrytis cinerea.[27] Evaluations have been conducted on Basidiomycetes species including Mycosphaerella graminicola.[35] Generally, the evidence suggests that surfactin has lower or no activity in comparison to the activity reported by iturin and fengycin.[27,36,37] Therefore, a meta-analysis of published studies concerning the effectiveness of these lipopeptides against plant pathogenic fungi was conducted.
5. Conclusions
After a meta-analysis of published studies on the antifungal properties of the lipopeptides iturin, fengycin and surfactin, it was concluded that the three lipopeptides inhibited in vitro fungal growth of various phytopathogenic fungi and decreased the severity of fungal damage to plant tissues. Fengycin and surfactin showed the greatest effect. When analyzed 20 Bacillus strains for the presence of the lipopeptides’genes, lipopeptide biosynthesis genes were found in five out of 20 Bacillus spp. strains. PCR detected fabD genes responsible to produce iturin A and fengycin, as well as srf genes for surfactin.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Oligonucleotides used for detection of lipopeptide biosynthesis genes. Table S2. General summary of data sources for Meta-analysis. Figure S1. Phylogenetic tree of the deduced amino acid sequence of iturin A (ituA), surfactin (srfA), and fengycin (FenD) from Bacillus strains (CBRF5, CBRF6, CBRM9 CBRF15) and their homologous protein sequences. The accession number of the proteins used is indicated. Protein sequences were retrieved from the NCBI and UniProt database.
Author Contributions
Conceptualization, N.N.C-.C.; E.R.-S, E.H.-N. and A.R.-R.; methodology, J.C.-A.; N.N.C.-.C. and M.E.C.-B; software, E.H.-N. and D.A.D.-C.; validation, W.H and D.A.D.-C.-M., E.H.-N. and N.N.C.-.C.; formal analysis, N.N.C-.C. and D.A.D.-C.; investigation, M.E.C.-B.; N.N.C-.C.; resources, J.C.-A. and D.A.D.-C.; E.R.-S.; data curation, A.R.-R. and D.A.D.-C.; writing-original draft preparation, A.R.-R. and N.N.C-.C; writing—review and editing, W.H.-M., E.H.-N; J.C.-A. and N.N.C-.C; visualization, M.E.C.-B. and N.N.C-.C.; supervision, M.E.C.-B., E.H.-N and A.R.-R.; project administration, E.R.-S.; funding acquisition, A.R.-R. and E.R.-S. All authors have read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.”
Funding
This work was funded Tecnológico Nacional de México (grant nos. 18872.23-PD and 14092.22-PD) and grant CONAHCYT, “Fortalecimiento de Infraestructura y Desarrollo de Capacidades Científicas” Number 322133. V.A.C.P, L.Y.C. and K.A.A. thanks CONAHCYT for support through her bachelor’s degree fellowships.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The first author would like to express their gratitude to PRODEP (DSA/103.5/16/10039) for their support of the NNCA through the “Conventional National Support for Doctoral Studies”.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- O’Brien, J.; Wright, G.D. An Ecological Perspective of Microbial Secondary Metabolism. Curr. Opin. Biotechnol. 2011, 22, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Cramer, R.A.; Stajich, J.E.; Yamanaka, Y.; Dietrich, F.S.; Steinbach, W.J.; Perfect, J.R. Phylogenomic Analysis of Non-Ribosomal Peptide Synthetases in the Genus Aspergillus. Gene 2006, 383, 24–32. [Google Scholar] [CrossRef]
- Akpa, E.; Jacques, P.; Wathelet, B.; Paquot, M.; Fuchs, R.; Budzikiewicz, H.; Thonart, P. Influence of Culture Conditions on Lipopeptide Production by Bacillus Subtilis. Appl. Biochem. Biotechnol. - Part A Enzym. Eng. Biotechnol. 2001, 91–93, 551–561. [Google Scholar] [CrossRef]
- Munakata, Y.; Heuson, E.; Daboudet, T.; Deracinois, B.; Duban, M.; Hehn, A.; Coutte, F.; Slezack-deschaumes, S. Screening of Antimicrobial Activities and Lipopeptide Production of Endophytic Bacteria Isolated from Vetiver Roots. Microorganisms 2022, 10, 209. [Google Scholar] [CrossRef] [PubMed]
- Romero, D.; De Vicente, A.; Olmos, J.L.; Dávila, J.C.; Pérez-García, A. Effect of Lipopeptides of Antagonistic Strains of Bacillus Subtilis on the Morphology and Ultrastructure of the Cucurbit Fungal Pathogen Podosphaera Fusca. J. Appl. Microbiol. 2007, 103, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Maget-Dana, R.; Thimon, L.; Peypoux, F.; Ptak, M. Surfactin/Iturin A Interactions May Explain the Synergistic Effect of Surfactin on the Biological Properties of Iturin A. Biochimie 1992, 74, 1047–1051. [Google Scholar] [CrossRef]
- Gong, A.D.; Li, H.P.; Yuan, Q.S.; Song, X.S.; Yao, W.; He, W.J.; Zhang, J.B.; Liao, Y.C. Antagonistic Mechanism of Iturin A and Plipastatin A from Bacillus Amyloliquefaciens S76-3 from Wheat Spikes against Fusarium Graminearum. PLoS One 2015, 10, e0116871. [Google Scholar] [CrossRef]
- Krishnan, N.; Velramar, B.; Velu, R.K. Investigation of Antifungal Activity of Surfactin against Mycotoxigenic Phytopathogenic Fungus Fusarium Moniliforme and Its Impact in Seed Germination and Mycotoxicosis. Pestic. Biochem. Physiol. 2019, 155, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.A.; McMullen, M.P.; Hershman, D.E.; Madden, L.V. Meta-Analysis of the Effects of Triazole-Based Fungicides on Wheat Yield and Test Weight as Influenced by Fusarium Head Blight Intensity. Disease Control and Pest Management 2010, 100, 160–171. [Google Scholar] [CrossRef]
- Athukorala, S.N.P.; Fernando, W.G.D.; Rashid, K.Y. Identification of Antifungal Antibiotics of Bacillus Species Isolated from Different Microhabitats Using Polymerase Chain Reaction and MALDI-TOF Mass Spectrometry. Can. J. Microbiol. 2009, 55, 1021–1032. [Google Scholar] [CrossRef]
- Stein, T. Bacillus Subtilis Antibiotics: Structures, Syntheses and Specific Functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Sarangi, T.; Ramakrishnan, S.; Nakkeeran, S. Antimicrobial Peptide Genes Present in Indigenous Isolates of Bacillus Spp. Exhibiting Antimicrobical Properties. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1361–1369. [Google Scholar] [CrossRef]
- Moyne, A.L.; Cleveland, T.E.; Tuzun, S. Molecular Characterization and Analysis of the Operon Encoding the Antifungal Lipopeptide Bacillomycin D. FEMS Microbiol. Lett. 2004, 234, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Verwoert, I.I.G.S.; Verbree, E.C.; Van der Linden, K.H.; Nijkamp, H.J.J.; Stuitje, A.R. Cloning, Nucleotide Sequence, and Expression of the Escherichia Coli FabD Gene, Encoding Malonyl Coenzyme A-Acyl Carrier Protein Transacylase. J. Bacteriol. 1992, 174, 2851–2857. [Google Scholar] [CrossRef]
- Morbidoni, H.R.; De Mendoza, D.; Cronan, J.E. Bacillus Subtilis Acyl Carrier Protein Is Encoded in a Cluster of Lipid Biosynthesis Genes. J. Bacteriol. 1996, 178, 4794–4800. [Google Scholar] [CrossRef]
- Lin, G.H.; Chen, C.L.; Tschen, J.S.M.; Tsay, S.S.; Chang, Y.S.; Liu, S.T. Molecular Cloning and Characterization of Fengycin Synthetase Gene FenB from Bacillus Subtilis. J. Bacteriol. 1998, 180, 1338–1341. [Google Scholar] [CrossRef]
- Tosato, V.; Albertini, A.M.; Zotti, M.; Sonda, S.; Bruschi, C.V. Sequence Completion, Identification and Definition of the Fengycin Operon in Bacillus Subtilis 168. Microbiology 1997, 143, 3443–3450. [Google Scholar] [CrossRef] [PubMed]
- Cosmina, P.; Rodriguez, F.; de Ferra, F.; Grandi, G.; Perego, M.; Venema, G.; van Sinderen, D. Sequence and Analysis of the Genetic Locus Responsible for Surfactin Synthesis in Bacillus Subtilis. Mol. Microbiol. 1993, 8, 821–831. [Google Scholar] [CrossRef]
- Menkhaus, M.; Ullrich, C.; Kluge, B.; Vater, J.; Vollenbroich, D.; Kamp, R.M. Structural and Functional Organization of the Surfactin Synthetase Multienzyme System. J. Biol. Chem. 1993, 268, 7678–7684. [Google Scholar] [CrossRef]
- Rosenberg, M.S. The File-Drawer Problem Revisited: A General Weighted Method for Calculating Fail-Safe Numbers in Meta-Analysis. Evolution (N. Y). 2005, 59, 464–468. [Google Scholar] [CrossRef]
- Rosenthal, R. The File Drawer Problem and Tolerance for Null Results. Psychol. Bull. 1979, 86, 638–641. [Google Scholar] [CrossRef]
- Mejía-Bautista, M.A.; Cristóbal-Alejo, J.; Tun-Suárez, J.M.; Reyes-Ramírez, A. Actividad in Vitro de Bacillus Spp. En La Inhibición de Crecimiento Micelial de Fusarium Equiseti y Fusarium Solani Aislado de Chile Habanero (Capsicum Chinense Jacq.). Agrociencia 2016, 50, 1123–1135. [Google Scholar]
- Ramarathnam, R.; Bo, S.; Chen, Y.; Fernando, W.G.D.; Xuewen, G.; De Kievit, T. Molecular and Biochemical Detection of Fengycin- and Bacillomycin D-Producing Bacillus Spp., Antagonistic to Fungal Pathogens of Canola and Wheat. Can. J. Microbiol. 2007, 53, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Madan, A. CAP3: A DNA Sequence Assembly Program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Bie, X. mei; Lv, F. xia; Zhao, H. zhen; Lu, Z. xin Antifungal Activity and Mechanism of Fengycin in the Presence and Absence of Commercial Surfactin Against Rhizopus Stolonifer. J. Microbiol. 2011, 49, 146–150. [Google Scholar] [CrossRef]
- Ye, Y. feng; Li, Q. qin; Fu, G.; Yuan, G. qing; Miao, J. hua; Lin, W. Identification of Antifungal Substance (Iturin A2) Produced by Bacillus Subtilis B47 and Its Effect on Southern Corn Leaf Blight. J. Integr. Agric. 2012, 11, 90–99. [Google Scholar] [CrossRef]
- Kong, H.G.; Lee, H.J.; Bae, J.Y.; Kim, N.H.; Moon, B.J.; Lee, S.W. Spatial and Temporal Distribution of a Biocontrol Bacterium Bacillus Licheniformis N1 on the Strawberry Plants. Plant Pathol. J. 2010, 26, 238–244. [Google Scholar] [CrossRef]
- Mora, I.; Cabrefiga, J.; Montesinos, E. Antimicrobial Peptide Genes in Bacillus Strains from Plant Environments. Int. Microbiol. 2011, 14, 213–223. [Google Scholar] [CrossRef]
- Stanković, S.; Mihajlović, S.; Draganić, V.; Dimkić, I.; Vukotić, G.; Berić, T.; Fira, D. Screening for the Presence of Biosynthetic Genes for Antimicrobial Lipopeptides in Natural Isolates of Bacillus Sp. Arch. Biol. Sci. 2012, 64, 1425–1432. [Google Scholar] [CrossRef]
- Gond, S.K.; Bergen, M.S.; Torres, M.S.; White, J.F. Endophytic Bacillus Spp. Produce Antifungal Lipopeptides and Induce Host Defence Gene Expression in Maize. Microbiol. Res. 2015, 172, 79–87. [Google Scholar] [CrossRef]
- Zhou, S.; Liu, G.; Zheng, R.; Sun, C.; Wu, S. Structural and Functional Insights into Iturin W, a Novel Lipopeptide Produced by the Deep-Sea Bacterium Bacillus Sp. Strain Wsm-1. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Wang, X.; Liang, L.; Shao, H.; Ye, X.; Yang, X.; Chen, X.; Shi, Y.; Zhang, L.; Xu, L.; Wang, J. Isolation of the Novel Strain Bacillus Amyloliquefaciens F9 and Identification of Lipopeptide Extract Components Responsible for Activity against Xanthomonas Citri Subsp. Citri. Plants 2022, 11, 457. [Google Scholar] [CrossRef] [PubMed]
- Steller, S.; Sokoll, A.; Wilde, C.; Bernhard, F.; Franke, P.; Vater, J. Initiation of Surfactin Biosynthesis and the Role of the SrfD-Thioesterase Protein. Biochemistry 2004, 43, 11331–11343. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Shiraishi, S.; Suzuki, S. Are Cyclic Lipopeptides Produced by Bacillus Amyloliquefaciens S13-3 Responsible for the Plant Defence Response in Strawberry against Colletotrichum Gloeosporioides? Lett. Appl. Microbiol. 2015, 60, 379–386. [Google Scholar] [CrossRef]
- Mejri, S.; Siah, A.; Coutte, F.; Magnin-Robert, M.; Randoux, B.; Tisserant, B.; Krier, F.; Jacques, P.; Reignault, P.; Halama, P. Biocontrol of the Wheat Pathogen Zymoseptoria Tritici Using Cyclic Lipopeptides from Bacillus Subtilis. Environ. Sci. Pollut. Res. 2018, 25, 29822–29833. [Google Scholar] [CrossRef]
- Dang, Y.; Zhao, F.; Liu, X.; Fan, X.; Huang, R.; Gao, W.; Wang, S.; Yang, C. Enhanced Production of Antifungal Lipopeptide Iturin A by Bacillus Amyloliquefaciens LL3 through Metabolic Engineering and Culture Conditions Optimization. Microb. Cell Fact. 2019, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.T.; Park, B.K.; Kim, S.E.; Lee, W.J.; Moon, J.S.; Cho, M.S.; Park, H.Y.; Hwang, I.; Kim, S.U. Organization and Characterization of Genetic Regions in Bacillus Subtilis Subsp. Krictiensis ATCC55079 Associated with the Biosynthesis of Iturin and Surfactin Compounds. PLoS One 2017, 12, e0188179. [Google Scholar] [CrossRef]
Figure 1.
Effect size (mean and 95% confidence interval) of lipopeptides on the in vitro inhibition of fungal growth and the suppression of the severity of foliar damage caused by phytopathogenic fungi. The number of sample points used to calculate each mean is shown for each analysis. Means with confidence intervals not overlapping zero were considered significant. n: sample size; nfs: number of the certainty; *: indicates the statistical robustness of nfs. The mean effect size E++ = -0.9829 (CI = -1.7811 to -0.6883, bias 95%).
Figure 1.
Effect size (mean and 95% confidence interval) of lipopeptides on the in vitro inhibition of fungal growth and the suppression of the severity of foliar damage caused by phytopathogenic fungi. The number of sample points used to calculate each mean is shown for each analysis. Means with confidence intervals not overlapping zero were considered significant. n: sample size; nfs: number of the certainty; *: indicates the statistical robustness of nfs. The mean effect size E++ = -0.9829 (CI = -1.7811 to -0.6883, bias 95%).
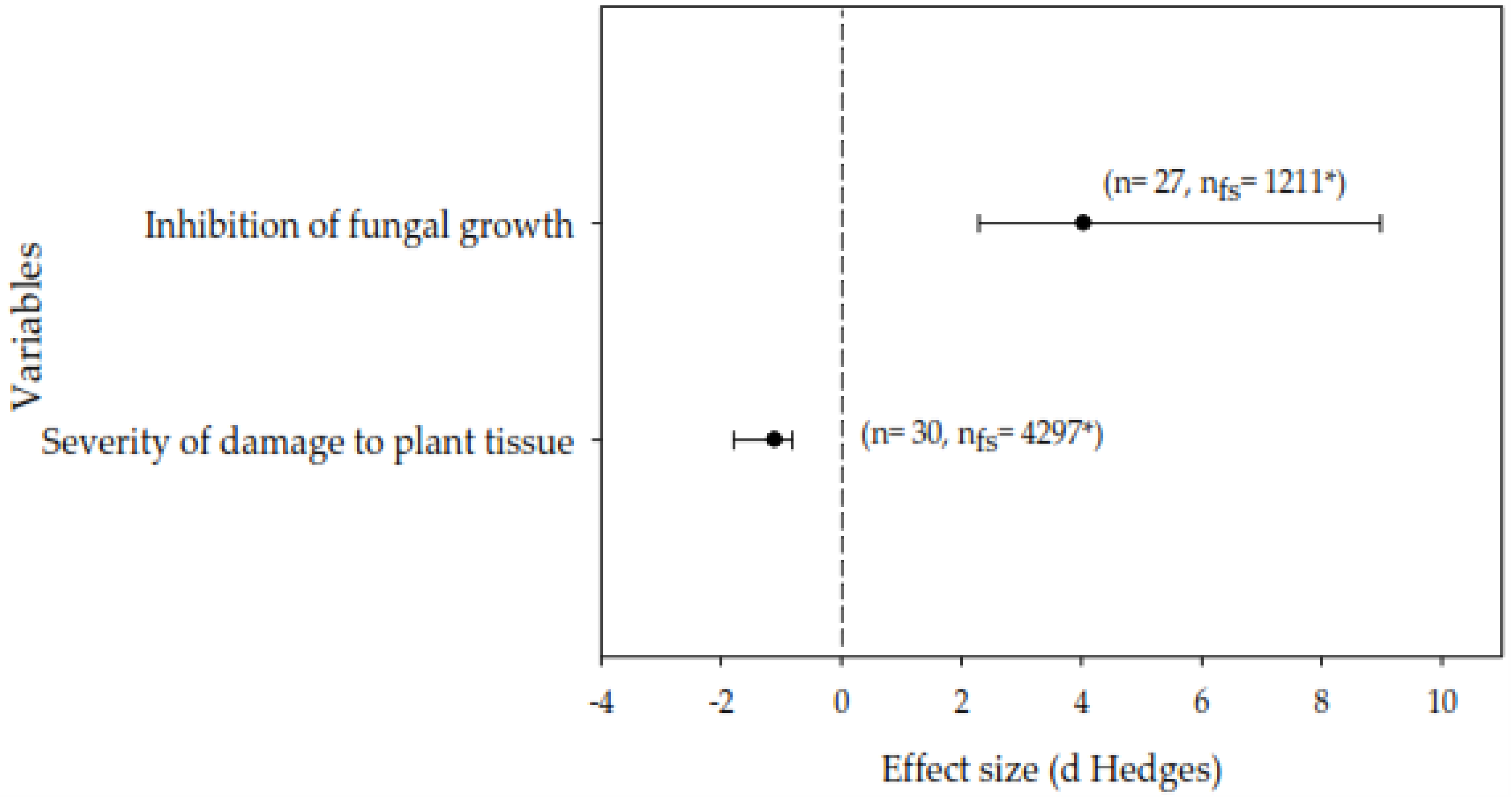
Figure 2.
Effect size (mean and 95% confidence interval) of lipopeptides on the inhibition of fungal growth in vitro for the iturin, surfactin and fengycin categories. Effect sizes were considered significant if the 95% confidence intervals did not overlap zero. Effect sizes within analyzes were considered different from each other if their 95% confidence intervals did not overlap. n: sample size; nfs: number of the certainty; *: indicates the statistical robustness of nfs. The mean effect size E++ = 4.0395 (= 2.2811 to 9.3650, bias 95%).
Figure 2.
Effect size (mean and 95% confidence interval) of lipopeptides on the inhibition of fungal growth in vitro for the iturin, surfactin and fengycin categories. Effect sizes were considered significant if the 95% confidence intervals did not overlap zero. Effect sizes within analyzes were considered different from each other if their 95% confidence intervals did not overlap. n: sample size; nfs: number of the certainty; *: indicates the statistical robustness of nfs. The mean effect size E++ = 4.0395 (= 2.2811 to 9.3650, bias 95%).
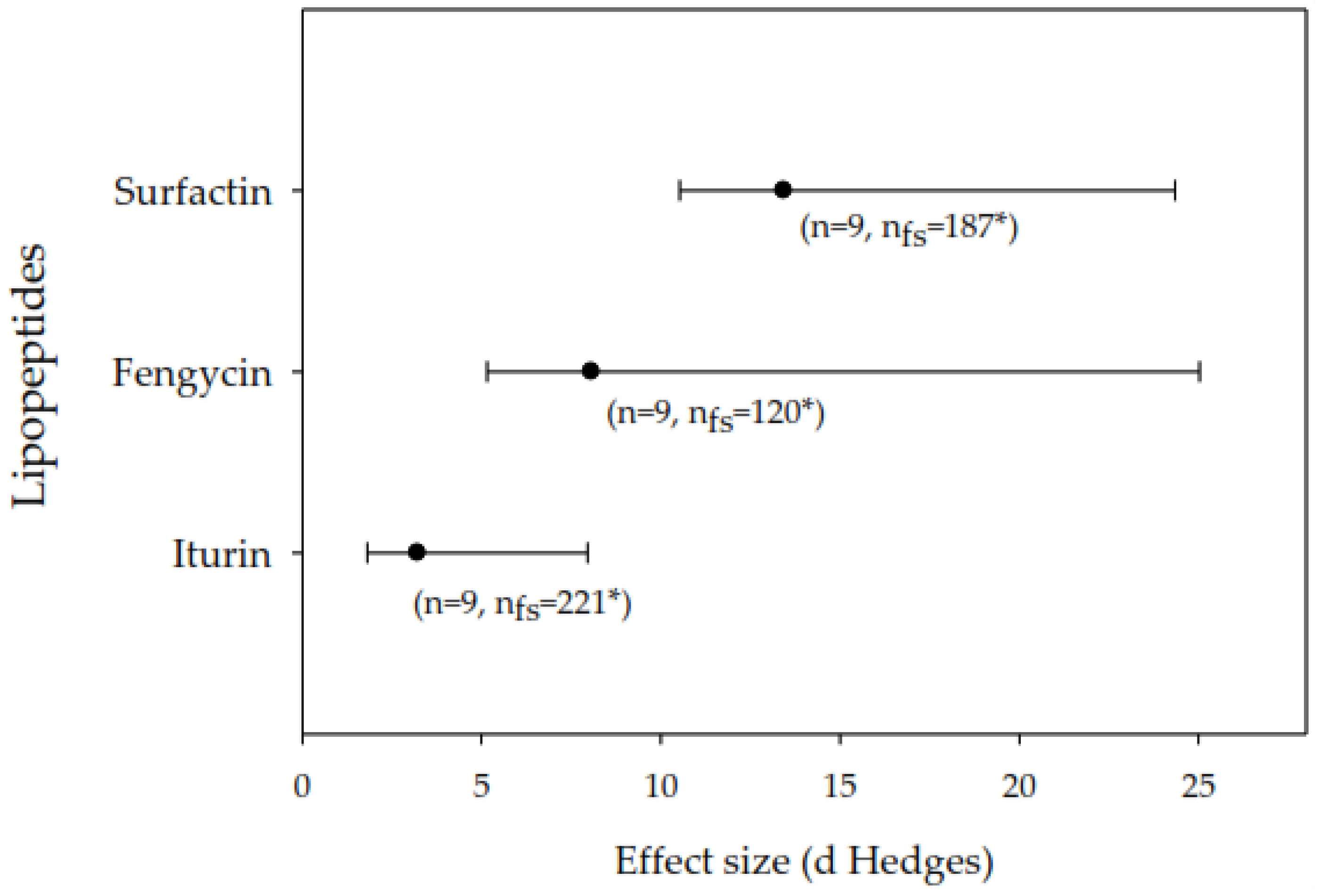
Figure 3.
Effect size (mean and 95% confidence interval) of lipopeptides on suppression of damage to plant tissue for the iturin, surfactin and fengycin categories. Effect sizes were considered significant if the 95% confidence intervals did not overlap zero. Effect sizes within analyzes were considered different from each other if their 95% confidence intervals did not overlap. n: sample size; nfs: number of the certainty; *: indicates the statistical robustness of nfs. The mean effect size E++ = -1.1017 (= -1.8703 to -0.8299, bias 95%).
Figure 3.
Effect size (mean and 95% confidence interval) of lipopeptides on suppression of damage to plant tissue for the iturin, surfactin and fengycin categories. Effect sizes were considered significant if the 95% confidence intervals did not overlap zero. Effect sizes within analyzes were considered different from each other if their 95% confidence intervals did not overlap. n: sample size; nfs: number of the certainty; *: indicates the statistical robustness of nfs. The mean effect size E++ = -1.1017 (= -1.8703 to -0.8299, bias 95%).
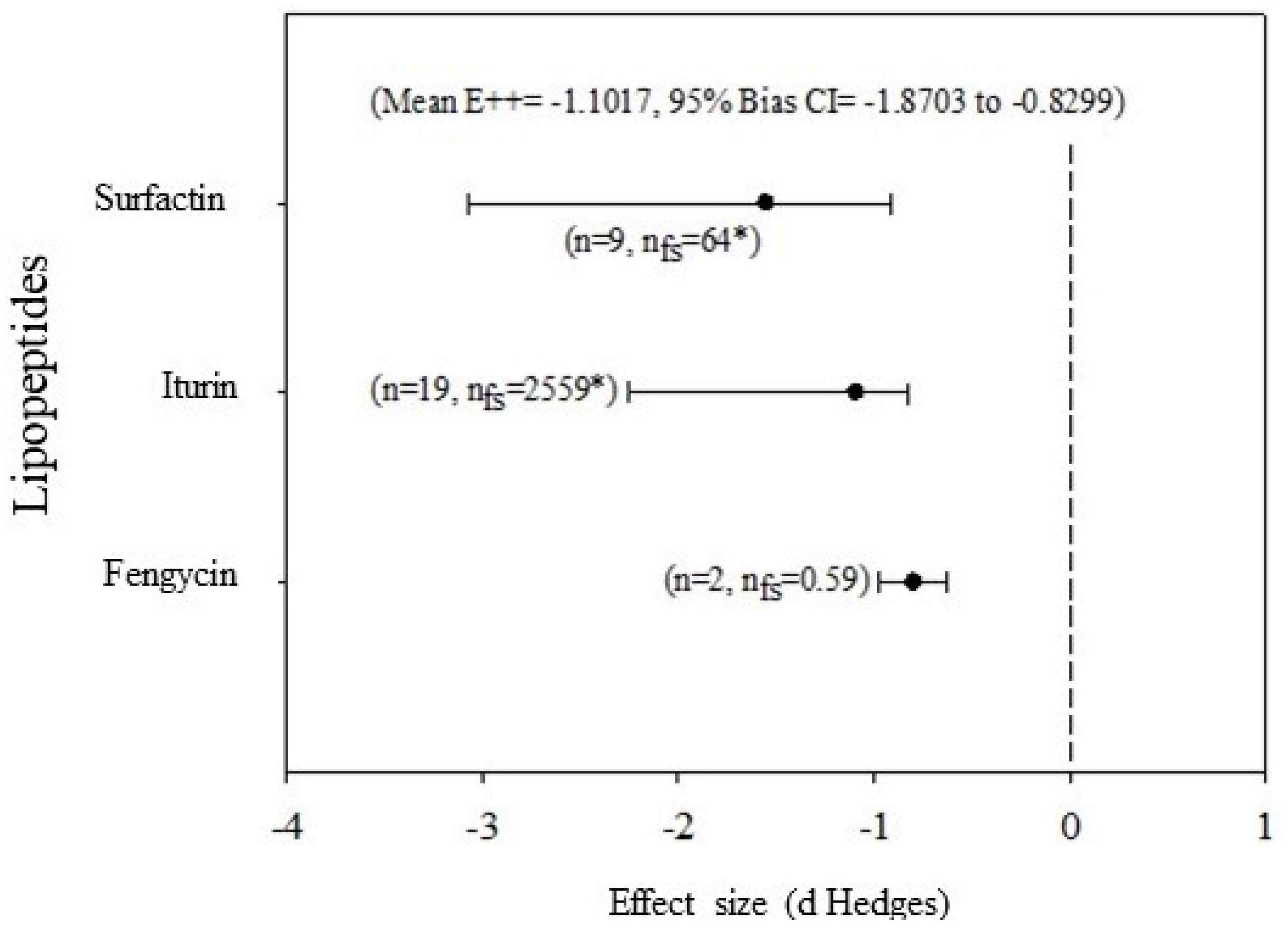

Table 1.
Heterogeneity statistics for each model in the analysis of frequency of inhibition of fungal growth in vitro and severity of damage to plant tissue by phytopathogenic fungi exposed to the lipopeptides iturin, fengycin and surfactin.
Table 1.
Heterogeneity statistics for each model in the analysis of frequency of inhibition of fungal growth in vitro and severity of damage to plant tissue by phytopathogenic fungi exposed to the lipopeptides iturin, fengycin and surfactin.
| Inhibition of fungal growth | Severity of damage to plant tissue | ||||||
|---|---|---|---|---|---|---|---|
| Df | Q.B. | Q | Df | Q.B. | Q | ||
| Complete model | 26 | 210.9195 | <0.0000 | 29 | 465.4113 | <0.0000 | |
| Lipopeptides | 2 | 43.1895 | <0.0000 | 2 | 2.3683 | <0.3060 | |
| Concentration | 1 | 31.9641 | <0.0000 | 3 | 23.6973 | <0.0000 | |
Df, degrees of freedom; QB, variation in effect size explained by the model.
Table 2.
Detection of lipopeptide genes from Bacillus spp. native.
| Strains | Genes for antimicrobial lipopeptides | ||
|---|---|---|---|
| ituA | fenD | srfA | |
| Bacillus sp. CBRF5 | + | - | - |
| B. amyloliquefaciens CBRF6 | + | - | + |
| B. subtilis CBRF15 | - | + | - |
| Bacillus sp. CBRM9 | + | - | - |
| B. subtilis CBSN67 | + | - | - |
Table 3.
Analysis of the deduced amino acid sequences of the lipopeptide synthesis genes.
| Homology | |||||||
|---|---|---|---|---|---|---|---|
| Cepa | Lipopeptide /Gene1 | GenBank | UniProt | ||||
| Protein | Identity (%) | Function | Protein | Identity (%) | Protein | ||
| CBRF5 | Iturin A/ fabD |
malonyl-CoA transacylase | 100 | WP_253612942.1 | ACP-S-m Transferase | 100 | Q5NKN8 |
| CBRF6 | Iturin A/ fabD |
malonyl-CoA transacylase | 100 | WP_253612942.1 | ACP-S-m Transferase | 100 | Q5NKN8 |
| CBSN67 | Iturin A/ fabD |
ACP-S-m transferase | 100 | WP_268428834.1 | Malonyl-CoA transacylase | 99.5 | A0A9W5LIK4 |
| CBRM9 | Iturin A/ ituD |
Iturin A synthetase D |
100 | AVN84851.1 | Iturin A synthetase D | 100 | A0A2P1IM41 |
| CBRF15 | Fengycin/ fabD | ACP-S- m transferase | 84.43 | MDA2462581.1 | ACP-S- m transferase | 84.4 | A0A9X7HYE2 |
| CBRF6 | Surfactin/ srf |
Surfactin synthetase |
100 | AEU04020.1 | Surfactin synthetase | 100 | G9JLX1 |
1 Gene name assigned by DNA sequence homology (Blastn-GenBank). ACP-Sm transferase; Acyl-S-malonyl transferase Carrier Protein.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



