Submitted:
24 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
Various dietary supplements have been shown to extend the life span of Drosophila melanogaster, including several that promote autophagy, such as rapamycin and spermidine. The goal of the study presented here was to test numerous additional potential anti-aging supplements, primarily inhibitors of the target of rapamycin (TOR) and/or phosphatidylinositol 3-kinase (PI3K). Using a single, comparatively long-lived y w test strain, screening was performed in male flies supplemented either throughout adulthood or in a few cases beginning in middle or late adult life, with concentrations spanning 4-6 orders of magnitude in most cases. Supplementation with PP242 and deferiprone, an iron chelator, beginning in late adult life had no positive effect on life span. Lifelong supplementation with Ku-0063794, LY294002, PX-866-17OH, Torin2 and WYE-28 had no effect at any dose. Rapamycin, spermidine and wortmannin all had significant life-shortening effects at the highest doses tested. AZD8055, PI-103 hydrochloride and WYE-132 yielded slight beneficial effects at 1-2 doses, but only 100 nM AZD8055 was confirmed to have a minor (1.3%) effect in a replicate experiment, which was encompassed by other control groups within the same study. These compounds had no effect on fly fecundity (egg-laying) or fertility (development of progeny to adulthood), but equivalent doses of rapamycin abolished fertility. The solvent DMSO had no significant effect on life span at the concentrations used to solubilize most compounds in the fly medium, but it drastically curtailed both survival and fertility at higher concentrations. 2-hydroxypropyl-β-cyclodextrin also failed to extend the life span when provided throughout adulthood or beginning in mid-adult life. Collectively, the results suggest that inhibition of the TOR/PI3K pathway and autophagy through dietary intervention is not a straightforward anti-aging strategy in Drosophila, and that further extension of life is difficult in comparatively long-lived flies.
Keywords:
life span
; aging
; Drosophila
; rapamycin
; TOR inhibition
1. Introduction
Drosophila melanogaster is widely used as a model organism in which to test the effects of dietary supplements on aging and longevity [1]. Life extension has been reported at one or more concentrations for the following supplements, amongst many others: rapamycin (sirolimus), which promotes autophagy as an inhibitor of the target of rapamycin (TOR) signal transduction pathway downstream from phosphatidylinositol 3-kinase (PI3K) [2,3], the rapamycin analog (rapalog) everolimus and other kinase inhibitors [4], and spermidine, a natural polyamine that inhibits oxidative stress and also promotes autophagy [5,6].
The PI3K/AKT/TOR pathway is of particular interest because mutations in all components of the pathway are common in various solid tumors [7] and because rapamycin itself extends life and diminishes the incidence of cancer in numerous strains of mice [8,9,10]. The rapalogs everolimus and temsirolimus are used to treat tuberous sclerosis complex (TSC) [11,12], which arises from TSC1/2 mutations that up-regulate the pathway, and some human cancers [13,14,15]. Rapamycin is also used off-label by some healthy adults for potential extension of health span and even life span [16]. Nonetheless, numerous authors express varying levels of concern about known side effects for both rapamycin and rapalogs [9,11,12,13,14,15], and the limited effectiveness of rapalogs against cancer has led to their being superseded by other modes of treatment in some cases [15,17] and to investigation of alternative means of inhibiting TOR activity [13,18]. In the context of aging, low or intermittent dosing of rapamycin is an option to minimize side effects [9,16]. In female wDah Drosophila, treatment for 15-30 days in early adult life was sufficient to extend the median adult life span from 72 to 78-80 days [3].
The TOR protein is a serine/threonine kinase that functions as the catalytic subunit in two multiprotein complexes (TORC1 and TORC2) that were first identified from spontaneous mutations conferring rapamycin resistance in Saccharomyces cerevisiae [19]. TORC1 is activated by nutrients [9,20] and by growth factors acting through PI3K via AKT inhibition of TSC1/2 [18,21] or through mitogen activated protein kinase (MAPK) [22]. It phosphorylates ribosomal protein S6 kinase (S6K) [23,24] and eukaryotic initiation factor 4E-binding protein 1 (4E-BP1) [24,25] to promote translation and growth in nutrient-rich conditions [21]. TORC2 is likewise activated by increases in nutrient levels and by growth factors via PI3K, but it is additionally activated by decreases in nutrient levels and by various forms of stress [26]. It phosphorylates at least 26 targets and has more diverse functions, including (i) activation of AKT and PKC, which promote TORC1 signaling, (ii) activation of SGK, which also promotes TORC1 and additionally up-regulates ion channels, glucose and amino acid carriers, (iii) actin cytoskeletal remodeling and (iv) either positive or negative regulation of autophagy [26]. Both TORC1 and TORC2 also have roles in promoting lipid biosynthesis [27].
It has been proposed that inhibition of TORC1 is beneficial for life span, but inhibition of TORC2 is detrimental [9,28]. Rapamycin inhibits TORC1 acutely, which is thought to give rise to its life-extending effect, but prolonged exposure to rapamycin also interrupts the assembly of TORC2 to a cell type-specific extent [29], leading to side effects such as insulin resistance [28]. The best cost:benefit ratio might therefore be achieved using low doses, intermittent or transient exposure to rapamycin, or compounds that are more highly selective for TORC1 [9]. However, regulation of either complex in isolation is not straightforward, because of feedback loops whereby TORC1 inhibits the upstream insulin receptor substrate-1 and rapamycin relieves this inhibition [30,31], TORC2 activates TORC1 via phosphorylation of AKT, while TORC1 negatively regulates TORC2 via phosphorylation of its Rictor component by S6K [30,32]. Furthermore, although TORC1 action against S6K is highly sensitive to inhibition by rapamycin [28], in mammalian cells rapamycin only partly or temporarily suppresses phosphorylation of 4E-BP1 and activation of autophagy [33,34].
Alternative inhibitors of the TOR pathway that vary in their selectivity for TORC1, TORC2 and PI3K might therefore be of value to obtain or improve upon the benefits while avoiding side effects of rapamycin. Second-generation TOR kinase inhibitors block both complexes but not the feedback activation of PI3K, whereas dual kinase inhibitors block both TOR and PI3K [18]. Although neither category has yielded favorable outcomes in initial trials compared with rapalogs for cancer [13], preliminary results are somewhat more promising but not always consistent for longevity in Drosophila. The TOR selective inhibitor 1 (Torin1) caused a substantial but quite variable increase of once-mated Dahomey female longevity from a short (mean 22 d) baseline [35]. Torin2, which has comparable selectivity for mTOR over PI3K in cellular but not biochemical assays and ~10-fold greater bioavailability than Torin1 [36], with an effective dose ~1/1000 that of rapamycin against neuroblastoma cell lines [37], nonetheless increased Canton S male median life span by only 4% at the lowest levels of a 0.5 – 10 µM dose range and had no beneficial effect in females with a more optimal (median 55 d) baseline [38]. AZD8055 promotes autophagy and inhibits both TOR complexes with at least 1000× selectivity vs. PI3K and other kinases [39]. PP242 also inhibits both complexes with high selectivity relative to PI3K and other kinases [40]. In combination with the senolytic navitoclax (but not separately), either AZD8055 or PP242 increased survival times from a 6-day baseline in female w1118 flies exposed to hydrogen peroxide and sensitized senescent cells to navitoclax more effectively than rapamycin [41]. In contrast, in male flies of the same strain, lifelong supplementation with PP242 increased mean survival from ~46 to ~57 d, comparable to the effect of rapamycin [42]. LY294002, which was initially identified as a PI3K inhibitor [43] but subsequently found also to inhibit TOR [44] and numerous unrelated kinases [45], was reported to extend life span at 5 µM in Canton S flies of both sexes from a 51-54 d baseline [46], but it had no effect at 0.5-4.5 mM in Oregon R males under mass-screening conditions that yielded 20-30 d average life spans [4]. Similarly, wortmannin was found first to inhibit PI3K [47,48] and then TOR [44] and to extend life span slightly from a 33-35 d baseline at 0.5 µM in Canton S males but not females [46], while failing to extend life in the mass screen of Oregon R males at 0.5-4.5 mM concentrations [4]. It was separately reported to have no significant effect at 5 nM, while increasing Canton S male and decreasing female median life spans at 5 µM and benefitting both sexes in combination with either rapamycin (at 5 µM) or the NF-κB inhibitor pyrrolidine dithiocarbamate (at 5 nM) [49].
The primary goal of the experiments reported here was to retest longevity effects of rapamycin, spermidine, Torin2, AZD8055, LY294002 and wortmannin supplementation beginning in early adult life using a y w strain and environmental conditions associated with comparatively long life, and to test several additional kinase inhibitors and other potential life-extending supplements. Additional TORC1/TORC2 inhibitors included Ku-0063794 [50], WYE-28 [51] and WYE-132 [52], all of which are even more highly selective than PP242. Conversely, the wortmannin derivative PX-866 shows high selectivity for PI3K over TOR [53]. Its more potent primary biological metabolite PX-866-17OH [54,55] was tested, along with a soluble hydrochloride salt of PI-103, which inhibits PI3K class IA isoforms with >100× higher potency than LY294002 [56,57], while also inhibiting TORC1, TORC2 and DNA-PK [57]. Biochemical IC50 concentrations of the tested inhibitors of TOR and PI3K, where available, are summarized in Table 1 [58,59], while noting that cellular EC50 values and relative selectivity are substantially different for Torin2 [36] and are likely to differ for other compounds as well. A pilot test of supplementation beginning in late adult life was first performed for PP242 and the iron chelator deferiprone (DFP), which induces mitophagy and decreases production of reactive oxygen species [60]. DFP at a 163 µM dose partially rescued the life-shortening effect of frataxin knockdown in a y w background, although it had no notable effect on the control life span [61]. Supplementation was also initiated in early or mid-adult life for 2-hydroxypropyl-β-cyclodextrin (2-HP-β-CD), which inhibits AKT/TOR phosphorylation and facilitates initiation of autophagy, although in HepG2 cells it also blocks the later stages of autophagy and leads to apoptosis [62]. Lastly, the solvent dimethylsulfoxide (DMSO), which was used to solubilize many of the TOR/PI3K inhibitors, has itself been reported to shorten the life span of Drosophila at 0.5% but not 0.1% [63], and to extend life in Caenorhabditis elegans (in the presence of 5-fluorouracil) at 0.5-2.0% [64]. It was therefore tested for its own effects on life span that could mask or amplify the effects of the dissolved supplements. Given that the degree of oral uptake and bioavailability of the tested compounds in flies is not known, supplements were tested over the widest practical range of concentrations, spanning 4-6 orders of magnitude in most cases. The experiments were performed in male flies with the intention to repeat tests in females and to test for potential trade-offs related to fertility for any compound at any dose yielding consistent life extension.
2. Results
2.1. Longevity Effects of DMSO, PP242 and Deferiprone Supplementation Late in Adult Life
An initial test of longevity effects was performed in y w male flies on a Torula yeast-based medium, with DMSO, PP242 and DFP supplementation beginning at 64 days of adult age (Figure 1). Mortality prior to this time was ≤10% in all groups. A 17% decrease in remaining survival time was observed for water-supplemented (H2O) in comparison with regular control flies (Figure 1, Table 2), with elevated mortality coinciding with insufficient drying of vials on a few occasions. None of the supplemented flies surpassed the survival times of flies on the regular control medium. In comparison with the H2O controls, DMSO at final concentrations of 0.0002 – 0.2% had no significant effect on survival, but 2% DMSO cut the remaining survival time by 75%, from 21.0 to 4.4 days (Figure 1A). PP242 at the highest dose (649 nM) increased survival time by 10% relative to H2O controls, while the lowest doses decreased it by 9-14% (Figure 1B). Deferiprone at 7.19 – 719 µM did not affect survivorship vs. the H2O control; at 71.9 – 719 nM, it increased survival up to essentially the same length as the regular control group (Figure 1C).
2.2. Longevity Effects of Spermidine and TOR/PI3K Inhibitors – Screening Study
Spermidine and the remaining TOR/PI3K inhibitors listed in Table 1 were tested in six nearly concurrent experiments commenced over an eight-day interval, with lifelong supplementation in continuous darkness beginning 1-2 days after collection of 0-1-day-old adults (8 days for LY294002). Independent regular and H2O control groups were included in each experiment except for Torin2 and PI-103 hydrochloride, which included only an H2O control group. Survivorship curves for all supplements are shown in Figure 2. Mean adult life spans and logrank test results are shown in Table 3 for each experiment consisting of 1-3 supplements and the corresponding control groups. No significant difference in survivorship was observed among the 11 control groups in the six experiments (p = 0.085). In the first experiment, spermidine (dissolved in water) was associated with a 10-11% reduction of life span at the highest dose (10 mM) and no effect at lower doses (10 µM – 1 mM). DMSO again had no effect at final concentrations of 0.02 – 0.2%, corresponding to the highest concentrations for dissolved TOR/PI3K inhibitors. WYE-28 had no effect at concentrations ranging from 10 pM – 1 µM. In the second experiment, WYE-132 was associated with a 5-6% increase in mean life span at 10 pM, the lowest dose tested, but no significant effect at 100 pM – 1 µM. LY294002 had no effect at any dose ranging from 100 nM to 10 µM. In the third experiment, AZD8055 extended life by 4-7% at 10 µM and 100 nM, while having no effect at 1 µM or 100 pM – 10 nM. Rapamycin at high concentrations (100 – 400 µM) caused a major, dose-dependent decrease in life span ranging from 26-45%. The rapamycin groups were also significantly shorter-lived (p < 0.0005) than flies supplemented with equivalent or near-equivalent concentrations of DMSO (400, 200 and 100 µM rapamycin vs. 0.2, 0.1 and 0.04% DMSO, respectively). In the fourth experiment, lower doses of rapamycin (100 nM – 10 µM) and Ku-0063794 (1 nM – 10 µM) also had no effect. Wortmannin had no effect at 1 nM – 1 µM, but at 10 µM it caused a catastrophic 69-70% decrease in mean survival. In the fifth experiment, PX-866-17OH (1 nM – 10 µM) did not alter the life span. In the final experiment, Torin2 (100 pM – 10 µM) had no effect, while PI-103 hydrochloride (10 µM) increased mean life span by 4% relative to the H2O control group and had no effect at lower doses (100 pM – 1 µM).
2.3. Longevity effects of TOR/PI3K inhibitors – confirmation study
An additional experiment was performed under 12 h light:12 h dark conditions for compounds at doses that yielded significant increases in life span in the screening study. Since in the cases of AZD8055 and PI 103-HCl the highest concentration (10 µM) increased the life span, here additional 50 µM treatment groups were added. Rapamycin was also tested at 10 and 50 µM, and a control group containing an identical concentration of DMSO was prepared for each treatment group (Figure 3, Table 4). The variation among control groups was somewhat greater than before, with 0.2% DMSO yielding the longest life span and 0.04% DMSO yielding the shortest life span of any group. AZD8055 (100 nM) was the only treatment that extended life in relation to both regular and H2O controls, albeit by only 1.3% vs. regular controls. It also extended life vs. the corresponding 0.0004% DMSO control, but not 0.2% DMSO (Figure 3A).
2.4. Fecundity and Fertility Effects of TOR/PI3K Inhibitors in the Confirmation Study
Early adult fecundity (egg laying) and fertility (numbers of eggs developing to adulthood) were determined over the first four days of adult life for the same treatment groups as in the life span confirmation study, except that 200 µM rapamycin and a corresponding 0.8% DMSO control group were substituted for the regular and H2O controls. Rapamycin at concentrations of 50–200 µM impaired egg laying (Figure 4A) and at 10 – 200 µM totally abolished development of progeny to adulthood (Figure 4B). All groups conformed to the model assumptions of normal distribution and equal variance for fecundity, and all except the rapamycin groups conformed for fertility. One-way analysis of variance (ANOVA) showed a significant overall difference in fecundity (p = 0.008). Pairwise comparisons between DMSO control and supplementation groups showed marginal decreases (p < 0.050) that were not below the Bonferroni-corrected threshold (p < 0.006) for 200 µM rapamycin (p = 0.007), 50 µM rapamycin (p = 0.042) and 10 µM AZD8055 (p = 0.046), and no difference for the other groups. For fertility, one-way ANOVA showed no difference among the groups excluding rapamycin (p = 0.27). At a Bonferroni-corrected threshold of p < 0.017 for three comparisons, Mann-Whitney U tests revealed significantly lower fertility in the 10 and 50 µM rapamycin groups (p = 0.014), but not for 200 µM rapamycin vs. 0.8% DMSO (p = 0.046), which itself almost abolished development to adulthood.
2.5. Longevity Effect of 2-Hydroxypropyl-β-Cyclodextrin (2-HP-β-CD)
The final compound tested was 2-HP-β-CD (Figure 5, Table 5). In an initial experiment, supplementation at concentrations of 0.14–0.56 g/L (corresponding to 96-384 µM based on an average molecular weight of 1460 g/mol) on standard medium beginning at 50 days of adult age had a positive but nonsignificant effect (+7%) on remaining survival time (Figure 5A). In a replicate experiment, late-life supplementation diminished the survival time by up to 10% at 0.56 g/L, with no significant effect at 0.14 g/L and a borderline significant decrease at 2.80 g/L (Figure 5B). Lifelong supplementation on standard medium had no effect at all (Figure 5C). On an alternate medium containing more concentrated yeast and sugar but lacking cornmeal, there was an increase in mean life span of up to 5% that was not significant after correction for multiple comparisons (Figure 5D).
3. Discussion
Results of this study show that none in a set of TOR and PI3K inhibitors supplemented over a wide dose range throughout adult life showed any consistent life-extending effect in male flies of the y w test strain, with the minor exception of 100 nM AZD8055, which increased mean life span by only 1.3% vs. the regular control group in the replicate experiment. In contrast with past reports, rapamycin, wortmannin and the polyamine spermidine all had life-shortening effects at the highest doses. In addition, deferiperone provided only in late adult life had no effect and 2-HP-β-CD administered either throughout adulthood or beginning in mid-adult life had no reproducible beneficial effect on longevity. Rapamycin at lower doses did not affect life span or fecundity, but it completely suppressed the development of eggs to adulthood, in contrast with other TOR/PI3K inhibitors at comparable doses. Results of follow-up experiments examining the effects of rapamycin on longevity and fertility in multiple strains and on multiple diets will be presented and discussed in a separate manuscript [Jackson et al., in preparation].
Several of the supplements in this study (Ku-0063794, PI-103 hydrochloride, PX-866-17OH, WYE-28, WYE-132 and 2-HP-β-CD) have not been tested previously for longevity effects in Drosophila, or for AZD8055 only in the presence of a strong exogenous stressor [41]. For supplements that were tested previously, the current findings confirm the absence of a beneficial effect of DFP in control flies [61]. Life extension was reported for rapamycin [2,3,42,46], LY294002 [46], Torin2 [38], wortmannin [46], PP242 [42] and spermidine [5], although for rapamycin, LY294002 and wortmannin, some investigators reported no change or even adverse effects [4,65]. In general, where differences were observed, plausible explanations include the use of male vs. female flies, genetic differences among fly strains and differences in supplement dose, food composition or other aspects of the environment. For LY294002, low but not high doses were reported to extend life, albeit in different strains under strikingly dissimilar housing conditions [4,46]. The present study introduces a third strain and shows no effect in y w males at the low dose (5 µM) that was beneficial in Canton S males and females [46]. Torin2 was reported to have a slight (4%) positive effect only for 2/4 concentrations in male and 0/4 concentrations in female flies [38], which is not drastically divergent from the 0 ± 1% effect in males in the current study that includes the same concentration range. Again for wortmannin, the positive effect previously reported for male mean life span was only 4% at 0.5 µM [46], which does not differ drastically from the 0 ± 2% effect at the same dose in the current study. For higher doses (0.5-4.5 mM), wortmannin was screened and not included on a list of compounds with beneficial effects [4], but it was not clear whether it had neutral or adverse effects. The severe toxicity of 10 µM wortmannin in older flies in the present study is consistent with cell killing by wortmannin in the 2.3–58.4 µM concentration range (levels vastly in excess of its IC50 for TOR and PI3K) [48], which suggests that millimolar concentrations would have an even greater detrimental effect. In the case of PP242, the supplement was only administered beginning in late adult life, so the observed life shortening is not directly at odds with the beneficial effect of lifelong supplementation with higher doses reported in w1118 males [42]. Given that only the lowest doses studied here had an adverse effect and considering the wide variation between control groups in the initial study, the finding of a reduction of life span by PP242 should be treated as very preliminary. Nonetheless, there was no sign of life extension for PP242, and after replicate experiments were performed, no life extension by lifelong supplementation with PI-103 hydrochloride, which is comparable to PP242 in its IC50 values for TORC1 and TORC2 (Table 1). For spermidine, published results show a large beneficial effect in w1118 females at doses of 10 µM – 1 mM and neutral or adverse effects in replicate experiments at 10 mM, whereas in males only 10 µM spermidine increased the mean adult life span from ~43 to ~50 days, with no effect observed for 100 µM, 1 mM or 10 mM spermidine [5]. In the current study, the lower three of these concentrations had no effect in y w males, while 10 mM spermidine was mildly detrimental. Thus, the relative trends for different doses are preserved, with lower doses being more beneficial than higher doses, but in the longer-lived y w strain the overall effect of spermidine is shifted in a negative direction. The variation between life spans in the published and present reports is also comparable in magnitude to the variation among replicate cohorts within each study, and it highlights the importance of repeated testing before reaching firm conclusions about longevity effects of any treatment in flies.
A limitation of this and some other studies of dietary supplements in Drosophila is the lack of a direct demonstration of uptake of the compounds that were tested. Consequently, the absence of an effect on life span might mean that the supplements are ineffective or it might result from low bioavailability due to degradation during storage or food preparation, metabolism in the food medium or gastrointestinal system of the flies or an absence of transport across the gut lining. Here, at least, the stability of most of the TOR and PI3K inhibitors and spermidine was demonstrated after prolonged storage of frozen aliquots and by heating in acid to mimic the conditions of food preparation. These results, and the toxicity observed at the highest doses of wortmannin, rapamycin and spermidine provide indirect evidence that the absence of effects on life span at other doses or for the other inhibitors is not simply due to low bioavailability. It should also be noted that AZD8055, Ku-0063794, rapamycin, Torin2, WYE-28 and WYE-132 were all supplemented at doses up to at least 1000× higher than their IC50 for TORC1 and wortmannin was supplemented up to >1000× the IC50 for PI3K; therefore, a bioavailability of 0.1% of the concentration added to the food should have been sufficient to inhibit the respective kinases. Based on the molecular weights of the inhibitors in Table 1, an average weight of ~0.7 mg for y w male flies in this laboratory and average food consumption of 2.5 µL/day for Drosophila on a comparable diet [66], 10 µM supplementation corresponds to at least 11 mg/kg/day, which would translate to daily intake of 0.77 g/day for a 70 kg human. For comparison, short-term human trials of rapamycin have used much lower doses, such as 1 mg/day [67].
2-HP-β-CD was tested in part because it had been shown to diminish the accumulation of lipofuscin [68], an age-pigment that may be both a cause and consequence of impaired autophagy [69]. Additionally, depletion of cellular cholesterol by the related compound, methyl-β-cyclodextrin, suppresses TORC1 activity [20] and 2-HP-β-CD partially rescues the life-shortening effect of the Niemann-Pick type C1 mutation (npc1-/-) in a mouse model while lowering elevated cholesterol levels in multiple organs [70]. Doses of 500 and 2000 mg/kg/day have no toxicological effects in dogs [71], so equivalent doses of 0.14 and 0.56 g/L (based on 2.5 µL/day food consumption by 0.7 mg flies) were used, along with a 5× higher dose of 2.80 g/L. All doses were well tolerated, but none had a significant, positive effect on life span when supplemented beginning either in early or middle adult life. Although the stability of the compound was not checked, it was heated only briefly to ~60 °C during food preparation, whereas thermodegradation of cyclodextrins typically occurs only at >250 °C [72]. The absence of a beneficial effect on life span is consistent with 2-HP-β-CD having no effect on cholesterol content in npc1+/+ mice and it provides no evidence for a reversible accumulation of lipofuscin or cholesterol imbalance as a cause of death in the y w fly strain. Efforts to quantify lipofuscin in organs of y w flies on the standard diet of this laboratory are in progress.
Finally, the fecundity and fertility study yielded two noteworthy findings. First, rapamycin completely suppressed the development of embryos to adulthood at concentrations where AZD8055 and PI-103 hydrochloride had no effect, even though rapamycin is intermediate between the other kinases in its potency against TORC1. These results might reflect either lower toxicity or faster inactivation of the other inhibitors in comparison with rapamycin. Second, the solvent DMSO also nearly completely abolished development at 0.8%, consistent with its severe, life-shortening effect at 2% final concentration. Although lower concentrations of DMSO used to dissolve TOR/PI3K inhibitors in this study did not significantly affect fertility or longevity (400 µM rapamycin was provided in a final concentration of 0.2% DMSO, 200 µM rapamycin was in 0.1% DMSO and all other supplements had ≤0.05% DMSO), potential effects of DMSO as solvent should be considered in longevity studies. Notably, Spindler and colleagues [4] also provided supplements in a final 0.2% DMSO concentration for their mass-screening experiments and reported average life spans of 20-30 days. Although they adjusted other conditions to achieve the shorter life spans for rapid screening, they noted the need for future experiments with longer control life spans; for such experiments, a control group with no DMSO would exclude an effect of the solvent overlaid on the effects of the supplements.
Although insufficient dosing or bioavailability cannot be excluded as possible reasons for the absence of beneficial effects of the TOR/PI3K inhibitors and other autophagy-related supplements in this study, a straightforward explanation is that this pathway and process are not the limiting factor in the aging and death of male flies of this lineage and dietary regimen. Consistent with these findings, genetic interventions in TOR signaling and autophagy that had been reported by others to extend the life spans of Drosophila had no beneficial effect in y w (or w1118) flies of either sex in this laboratory [73]. Although inhibition of TOR/PI3K or enhancement of autophagy might be beneficial in some fly strains or environmental conditions, dietary interventions intended to regulate aging by this mechanism do not appear to be universally effective methods for life extension in Drosophila.
4. Materials and Methods
4.1. Fly Strain and Media
The y w strain used in this study was generated in the laboratory of W.C. Orr (Southern Methodist University, Dallas, TX) [74] and has been used in previous studies in this laboratory [73]. Fly stocks were maintained and experiments were performed on the previously described medium [73], except for one experiment with 2-HP-β-CD that used a high yeast medium prepared by boiling 10.0 g/L agar in deionized water, then adding 100 g/L Torula yeast and 50.0 g/L sucrose, boiling again, cooling to ~60 °C, then adding 3.00 g/L methyl-4-hydroxybenzoate dissolved in 30 mL ethanol as a mold inhibitor and 3.00 mL/L propionic acid to inhibit bacterial growth.
4.2. Supplements
Rapamycin was purchased from LC Laboratories (Woburn, MA). Spermidine and 3-hydroxy-1,2-dimethyl-4(1H)-pyridone (deferiprone) were from Sigma Aldrich (St. Louis, MO, USA). Compounds AZD8055, Ku-0063794, LY294002, PI-103 hydrochloride, PP242, PX-866-17OH, Torin2, wortmannin, WYE-28 and WYE-132 were from Chemdea (Ridgewood, NJ). Dimethylsulfoxide (DMSO), ethanol and (2-hydroxypropyl)-β-cyclodextrin (2-HP-β-CD) were from Fisher Scientific (Waltham, MA, USA). Supplements other than spermidine and 2-HP-β-CD were dissolved in DMSO and stored immediately in 25 mM aliquots at -20 °C in a nondefrosting freezer except for rapamycin (200 mM), PP242 (32.4 mM), WYE-28 (2.5 mM), WYE-132 (2.5 mM) and deferiprone (35.9 mM in water). Spermidine was dissolved in deionized water, filter sterilized and stored in 1 M aliquots at -20 °C for no more than one month; however, aliquots of TOR/PI3K inhibitors were stored for up to 1 month for experiments shown in Figure 4, 2-3 months for Table 2 and Table 3 and up to 1 year for Table 4. 2-HP-β-CD was dissolved in deionized water, aliquots of other supplements were thawed, and all supplements were then serially diluted in deionized water just prior to mixing into freshly prepared food (1% v/v 2-HP-β-CD, 2% v/v for all others) at ~60 °C, immediately before or after dispensing the medium into individual vials. H2O control vials were prepared with an equivalent volume of water, and in most experiments regular control vials were also prepared without added water. The medium was then allowed to solidify and provided to flies 1-3 days after preparation.
To assess the stability of the supplements under conditions approximating those of the current study, Norman Arundel (Chemdea) performed ultra performance liquid chromatography (UPLC) with ultraviolet detection using aliquots of the TOR/PI3K inhibitors listed in Table 3 stored in DMSO at -20 °C for 0, 30 and 90 days. Separate aliquots were heated at 65 °C for 30 minutes in 0.33% propionic acid, 0.033% phosphoric acid, corresponding to the acid and heat exposures during preparation of standard fly medium. Concentrations ranged from 2.5 – 200 mM during storage and were lowered by dilution in DMSO to 1.25 – 18 mM before separation on a Waters HSS 1.8u column (2.1 × 50 mm) on a Shimadzu Nexera UPLC equipped with SPD-M40 photodiode array detector and LCMS2020 spectrometer. Gradient elution was performed with alternation between 99% of 0.1% formic acid in water and 80% acetonitrile, 20% methanol. In all cases, concentrations ranged from 91-106% of the 0-day baseline and no decomposition products or change in retention time were detected. Spermidine was stored as a 1 M solution in deuterated water at -20 °C for 0, 30 and 90 days or heated in acids at 65 °C for 30 minutes and detected with AdvanceCore Bruker 1H NMR at 400 MHz. There was no noticeable change in NMR peaks due to storage or heating in acid. Functionally, rapamycin stored in DMSO at -20 °C for 95 days maintained the same ability as at one day fully to suppress the development of embryos to adulthood at concentrations of 10 – 50 µM in standard medium.
4.3. Life Span
Male flies were isolated in four groups of 25/vial (25 × 95 mm) under light carbon dioxide anesthesia 0-1 day after eclosion, excluding immature individuals and any with visible signs of injury, sickness or genetic deformity. They were maintained at 25 ± 1 °C and kept in continuous darkness owing to the light sensitivity of some supplements, except the TOR/PI3K inhibitor confirmation study and experiments with 2-HP-β-CD were performed on a 12 h light:12 h dark cycle. Fresh vials were provided and deaths or escapes were recorded every 1-2 d, with supplementation beginning at age 64 d for the initial test of DMSO, PP242 and deferiprone. All other supplements were introduced 2 d after collection except LY294002 (8 d), PI-103 hydrochloride (1 d), Torin2 (1 d) and 2-HP-β-CD (either 1 d or 50 d).
4.4. Fertility
Groups of three female and three male flies were collected 0-19 hours after eclosion and provided with fresh vials containing media with supplements daily for four successive days at 25 ± 1 °C on a 12 h light:12 h dark cycle. Eggs laid were counted when flies were transferred, and adults that eclosed fully within 14 days were also counted.
4.5. Statistics
Logrank tests of survivorship were performed using SYSTAT 12 software for the time periods when supplements were provided. For each inhibitor, a test was performed comparing all doses with both regular and H2O control groups, where applicable; in cases where the comparison reached a significance threshold of p < 0.050 by the method of Mantel-Haenszel, comparisons were performed for each dose vs. both controls. For doses reaching the threshold of p < 0.050, paired comparisons were made for each control group separately. For the initial, late-life supplementation study, where a difference between control groups was detected, only the H2O control was included in the initial comparison, but supplement doses showing a significant difference vs. the H2O control group were additionally compared pairwise with the regular control group. In the screening study, to minimize the likelihood of false negative results, and owing to the complexity of the design with two control and multiple supplementation groups, no adjustment was made for multiple comparisons. Instead, any dose of a compound reaching the threshold of p < 0.050 vs. both controls in the screening study was retested in the confirmation study. In the latter case, groups showing significant effects vs. both regular and H2O controls were also compared with a control group treated with an identical concentration of the DMSO solvent. For the 2-HP-β-CD study, survival times for all doses were compared with that of the H2O control group in each experiment, and pairwise comparisons were made where the comparison reached a significance threshold of p < 0.050. Given that only one control group was used, a significance threshold of p < 0.017 was set based on Bonferroni correction for three comparisons within each experiment.
For the fertility study, total yields of embryos and subsequently eclosed adults over four days were compared by one-way analysis of variance after using Kolmogorov-Smirnov tests to validate the model assumption of normal distribution (at a significance threshold of p < 0.004 based on Bonferroni correction for 14 multiple comparisons) and Levene’s tests for equal variance. Where significant effects were observed, paired t tests were then performed for individual groups vs. control groups with equivalent volumes of DMSO solvent (with a significance threshold of p < 0.006 after Bonferroni correction for nine comparisons). For groups that did not conform to the model assumptions, Mann-Whitney U tests were used for the paired comparisons.
Author Contributions
Conceptualization, R.J.M.; methodology, A.C.N. and R.J.M.; validation, R.J.M.; formal analysis, R.J.M.; investigation, A.A.B., E.M.S., C.C.C., M.A.S., A.C.N. and R.J.M.; resources, R.J.M.; data curation, A.A.B., E.M.S., C.C.C., M.A.S. and R.J.M.; writing—original draft preparation, R.J.M.; writing—review and editing, A.A.B., A.C.N. and R.J.M.; visualization, A.A.B., A.C.N. and R.J.M.; supervision, R.J.M.; funding acquisition, R.J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by donations to the University of South Alabama from Alva LLC, Mr. Douglas R. Arends and Dr. Robin J. Mockett.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
Thanks are due to Jason Gaspar for suggesting the experiments with 2-HP-β-CD, Mehran Nisa for assistance with fly transfers in the screening study, Autumn McDaniel for counting embryos and adults in the fertility experiment and Madison Parfait for help with food preparation during the replication test of TOR/PI3K inhibitors. Sincere gratitude is extended to Norman Arundel (Chemdea) for testing of the stability of TOR inhibitors in frozen, stored aliquots. Results in Figure 2 and Table 3 were presented in a poster at conference Experimental Biology 2022 and the poster abstract was published [75]; copyright (2022) Wiley. The content is reported here in full with permission of the publisher, John Wiley & Sons, Inc.
Conflicts of Interest
The authors declare that this study received funding from Alva LLC, Mr. Douglas R. Arends and Dr. Robin J. Mockett. The external funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Barardo, D.; Thornton, D.; Thoppil, H.; Walsh, M.; Sharifi, S.; Ferreira, S.; Anžič, A.; Fernandes, M.; Monteiro, P.; Grum, T.; et al. The DrugAge database of aging-related drugs. Aging Cell 2017, 16, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Bjedov, I.; Toivonen, J.M.; Kerr, F.; Slack, C.; Jacobson, J.; Foley, A.; Partridge, L. Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster. Cell Metab. 2010, 11, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Juricic, P.; Lu, Y.-X.; Leech, T.; Drews, L.F.; Paulitz, J.; Lu, J.; Nespital, T.; Azami, S.; Regan, J.C.; Funk, E.; et al. Long-lasting geroprotection from brief rapamycin treatment in early adulthood by persistently increased intestinal autophagy. Nat. Aging 2022, 2, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Spindler, S.R.; Li, R.; Dhahbi, J.M.; Yamakawa, A.; Sauer, F. Novel protein kinase signaling systems regulating lifespan identified by small molecule library screening using Drosophila. PLoS ONE 2012, 7, e29782. [Google Scholar] [CrossRef]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of autophagy by spermidine promotes longevity. Nat. Cell. Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef]
- Morselli, E.; Galluzzi, L.; Kepp, O.; Criollo, A.; Maiuri, M.C.; Tavernarakis, N.; Madeo, F.; Kroemer, G. Autophagy mediates pharmacological lifespan extension by spermidine and resveratrol. Aging (Albany NY) 2009, 1, 961–970. [Google Scholar] [CrossRef]
- Yuan, T.L.; Cantley, L.C. PI3K pathway alterations in cancer: variations on a theme. Oncogene 2008, 27, 5497–5510. [Google Scholar] [CrossRef]
- Harrison, D.E.; Strong, R.; Sharp, Z.D.; Nelson, J.F.; Astle, C.M.; Flurkey, K.; Nadon, N.L.; Wilkinson, J.E.; Frenkel, K.; Carter, C.S.; et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 2009, 460, 392–395. [Google Scholar] [CrossRef]
- Lamming, D.W. Rapamycin and rapalogs. In Anti-Aging Pharmacology; Koltover, V.K., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 89–118. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Cancer prevention with rapamycin. Oncotarget 2023, 14, 342–350. [Google Scholar] [CrossRef]
- Trelinska, J.; Dachowska, I.; Kotulska, K.; Fendler, W.; Jozwiak, S.; Mlynarski, W. Complications of mammalian target of rapamycin inhibitor anticancer treatment among patients with tuberous sclerosis complex are common and occasionally life-threatening. Anti-Cancer Drugs 2015, 26, 437–442. [Google Scholar] [CrossRef]
- Bissler, J.J.; Kingswood, J.C.; Radzikowska, E.; Zonnenberg, B.A.; Belousova, E.; Frost, M.D.; Sauter, M.; Brakemeier, S.; de Vries, P.J.; Berkowitz, N.; et al. Everolimus long-term use in patients with tuberous sclerosis complex: Four-year update of the EXIST-2 study. PLoS ONE 2017, 12, e0180939. [Google Scholar] [CrossRef]
- Pezzicoli, G.; Filoni, E.; Gernone, A.; Cosmai, L.; Rizzo, M.; Porta, C. Playing the devil’s advocate: Should we give a second chance to mTOR inhibition in renal clear cell carcinoma? - ie strategies to revert resistance to mTOR inhibitors. Cancer Manag. Res. 2021, 13, 7623–7636. [Google Scholar] [CrossRef]
- Kiesewetter, B.; Melhorn, P.; Macheiner, S.; Wolff, L.; Kretschmer-Chott, E.; Haug, A.; Mazal, P.; Raderer, M. Does the dose matter? Antiproliferative efficacy and toxicity of everolimus in patients with neuroendocrine tumors – Experiences from a tertiary referral center. J. Neuroendocrinol. 2023, 35, e13319. [Google Scholar] [CrossRef] [PubMed]
- Karatrasoglou, E.A.; Dimou, M.; Piperidou, A.; Lakiotaki, E.; Korkolopoulou, P.; Vassilakopoulos, T.P. The role of mTOR in B cell lymphoid malignancies: biologic and therapeutic aspects. Int. J. Mol. Sci. 2023, 24, 14110. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, T.L.; Green, A.S.; Haddad, G.; Hudson, J.; Isman, A.; Nyquist, A.; Rosen, B.S.; Suh, Y.; Zalzala, S.; Zhang, X.; et al. Evaluation of off-label rapamycin use to promote healthspan in 333 adults. GeroScience 2023, 45, 2757–2768. [Google Scholar] [CrossRef] [PubMed]
- Silkenstedt, E.; Dreyling, M. Mantle cell lymphoma – Update on molecular biology, prognostication and treatment approaches. Hematol. Oncol. 2023, 41(S1), 36–42. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, X.; Proud, C.G. mTOR inhibitors in cancer therapy. F1000Res. 2016, 5, F1000. [Google Scholar] [CrossRef]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef]
- Castellano, B.M.; Thelen, A.M.; Moldavski, O.; Feltes, M.; van der Welle, R.E.N.; Mydock-McGrane, L.; Jiang, X.; van Eijkeren, R.J.; Davis, O.B.; Louie, S.M.; et al. Lysosomal cholesterol activates mTORC1 via an SLC38A9-Niemann-Pick C1 signaling complex. Science 2017, 355, 1306–1311. [Google Scholar] [CrossRef]
- Hay, N.; Sonenberg, N. Upstream and downstream of mTOR. Genes Dev. 2004, 18, 1926–1945. [Google Scholar] [CrossRef]
- Minard, A.Y.; Tan, S.-X.; Yang, P.; Fazakerley, D.J.; Domanova, W.; Parker, B.L.; Humphrey, S.J.; Jothi, R.; Stöckli, J.; James, D.E. mTORC1 is a major regulatory node in the FGF21 signaling network in adipocytes. Cell Rep. 2016, 17, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.J.; Beal, P.A.; Keith, C.T.; Chen, J.; Shin, T.B.; Schreiber, S.L. Control of p70 S6 kinase by kinase activity of FRAP in vivo. Nature 1995, 377, 441–446. [Google Scholar] [CrossRef]
- Burnett, P.E.; Barrow, R.K.; Cohen, N.A.; Snyder, S.H.; Sabatini, D.M. RAFT1 phosphorylation of the translational regulators p70 S6 kinase and 4E-BP1. Proc. Natl. Acad. Sci. USA 1998, 95, 1432–1437. [Google Scholar] [CrossRef]
- von Manteuffel, S.R.; Gingras, A.-C.; Ming, X.-F.; Sonenberg, N.; Thomas, G. 4E-BP1 phosphorylation is mediated by the FRAP-p70s6k pathway and is independent of mitogen-activated protein kinase. Proc. Natl. Acad. Sci. USA 1996, 93, 4076–4080. [Google Scholar] [CrossRef]
- Ragupathi, A.; Kim, C.; Jacinto, E. The mTORC2 signaling network: targets and cross-talks. Biochem. J. 2024, 481, 45–91. [Google Scholar] [CrossRef] [PubMed]
- Lamming, D.W.; Sabatini, D.M. A central role for mTOR in lipid homeostasis. Cell Metab. 2013, 18, 465–469. [Google Scholar] [CrossRef]
- Lamming, D.W.; Ye, L.; Katajisto, P.; Goncalves, M.D.; Saitoh, M.; Stevens, D.M.; Davis, J.G.; Salmon, A.B.; Richardson, A.; Ahima, R.S.; et al. Rapamycin-induced insulin resistance is mediated by mTORC2 loss and uncoupled from longevity. Science 2012, 335, 1638–1643. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D.D.; Ali, S.M.; Sengupta, S.; Sheen, J.-H.; Hsu, P.P.; Bagley, A.F.; Markhard, A.L.; Sabatini, D.M. Prolonged rapamycin treatment inhibits mTORC2 assembly and Akt/PKB. Mol. Cell 2006, 22, 159–168. [Google Scholar] [CrossRef]
- Watanabe, R.; Wei, L.; Huang, J. mTOR signaling, function, novel inhibitors, and therapeutic targets. J. Nucl. Med. 2011, 52, 497–500. [Google Scholar] [CrossRef]
- O’Reilly, K.E.; Rojo, F.; She, Q.-B.; Solit, D.; Mills, G.B.; Smith, D.; Lane, H.; Hofmann, F.; Hicklin, D.J.; Ludwig, D.L.; et al. mTOR inhibition induces upstream receptor tyrosine kinase signaling and activates Akt. Cancer Res. 2006, 66, 1500–1508. [Google Scholar] [CrossRef]
- Julien, L.-A.; Carriere, A.; Moreau, J.; Roux, P.P. mTORC1-activated S6K1 phosphorylates Rictor on threonine 1135 and regulates mTORC2 signaling. Mol. Cell. Biol. 2010, 30, 908–921. [Google Scholar] [CrossRef]
- Thoreen, C.C.; Sabatini, D.M. Rapamycin inhibits mTORC1, but not completely. Autophagy 2009, 5, 725–726. [Google Scholar] [CrossRef] [PubMed]
- Choo, A.Y.; Yoon, S.-O.; Kim, S.G.; Roux, P.P.; Blenis, J. Rapamycin differentially inhibits S6Ks and 4E-BP-1 to mediate cell-type-specific repression of mRNA translation. Proc. Natl. Acad. Sci. USA 2008, 105, 17414–17419. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.S.; Wileman, T.; Chapman, T. Lifespan extension without fertility reduction following dietary addition of the autophagy activator Torin1 in Drosophila melanogaster. PLoS ONE 2018, 13, e0190105. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, J.; Kang, S.A.; Thoreen, C.C.; Hur, W.; Ahmed, T.; Sabatini, D.M.; Gray, N.S. Discovery of 9-(6-aminopyridin-3-yl)-1-(3-(trifluoromethyl)-phenyl)benzo[h][1,6]naphthyridin-2(1H)-one (Torin2) as a potent, selective, and orally available mammalian target of rapamycin (mTOR) inhibitor for treatment of cancer. J. Med. Chem. 2011, 54, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Waetzig, R.; Matthes, M.; Leister, J.; Penkivech, G.; Heise, T.; Corbacioglu, S.; Sommer, G. Comparing mTOR inhibitor rapamycin with Torin-2 within the RIST molecular-targeted regimen in neuroblastoma cells. Int. J. Med. Sci. 2021, 18, 137–149. [Google Scholar] [CrossRef]
- Vershinina, Y.S.; Krasnov, G.S.; Garbuz, D.G.; Shaposhnikov, M.V.; Fedorova, M.S.; Pudova, E.A.; Katunina, I.V.; Kornev, A.B.; Zemskaya, N.V.; Kudryavtsev, A.A.; et al. Transcriptomic analysis of the effect of Torin-2 on the central nervous system of Drosophila melanogaster. Int. J. Mol. Sci. 2023, 24, 9095. [Google Scholar] [CrossRef] [PubMed]
- Chresta, C.M.; Davies, B.R.; Hickson, I.; Harding, T.; Cosulich, S.; Critchlow, S.E.; Vincent, J.P.; Ellston, R.; Jones, D.; Sini, P.; et al. AZD8055 is a potent, selective, and orally bioavailable ATP-competitive mammalian target of rapamycin kinase inhibitor with in vitro and in vivo antitumor activity. Cancer Res. 2010, 70, 288–298. [Google Scholar] [CrossRef]
- Feldman, M.E.; Apsel, B.; Uotila, A.; Loewith, R.; Knight, Z.A.; Ruggero, D.; Shokat, K.M. Active-site inhibitors of mTOR target rapamycin-resistant outputs of mTORC1 and mTORC2. PLoS Biol. 2009, 7, e1000038. [Google Scholar] [CrossRef]
- Xu, W.; Zhao, T.; Chen, H.; Huang, N.; Gong, H.; Zhang, J.; Yang, Y.; Li, T.; Zhang, G.; Gong, C.; et al. Pan-mTOR inhibitors sensitize the senolytic activity of navitoclax via mTORC2 inhibition-mediated apoptotic signaling. Biochem. Pharmacol. 2022, 200, 115045. [Google Scholar] [CrossRef]
- Chen, T.; Shen, L.; Yu, J.; Wan, H.; Guo, A.; Chen, J.; Long, Y.; Zhao, J.; Pei, G. Rapamycin and other longevity-promoting compounds enhance the generation of mouse induced pluripotent stem cells. Aging Cell 2011, 10, 908–911. [Google Scholar] [CrossRef] [PubMed]
- Vlahos, C.J.; Matter, W.F.; Hui, K.Y.; Brown, R.F. A specific inhibitor of phosphatidylinositol 3-kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002). J. Biol. Chem. 1994, 269, 5241–5248. [Google Scholar] [CrossRef] [PubMed]
- Brunn, G.J.; Williams, J.; Sabers, C.; Wiederrecht, G.; Lawrence, J.C.; Abraham, R.T. Direct inhibition of the signaling functions of the mammalian target of rapamycin by the phosphoinositide 3-kinase inhibitors, wortmannin and LY294002. EMBO J. 1996, 15, 5256–5267. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, S.I.; Zvelebil, M.J.; Shuttleworth, S.J.; Hancox, T.; Saghir, N.; Timms, J.F.; Waterfield, M.D. Exploring the specificity of the PI3K family inhibitor LY294002. Biochem. J. 2007, 404, 15–21. [Google Scholar] [CrossRef]
- Moskalev, A.A.; Shaposhnikov, M.V. Pharmacological inhibition of phosphoinositide 3 and TOR kinases improves survival of Drosophila melanogaster. Rejuv. Res. 2010, 13, 246–247. [Google Scholar] [CrossRef]
- Arcaro, A.; Wymann, M.P. Wortmannin is a potent phosphatidylinositol 3-kinase inhibitor: the role of phosphatidylinositol 3,4,5-trisphosphate in neutrophil responses. Biochem. J. 1993, 296, 297–301. [Google Scholar] [CrossRef]
- Powis, G.; Bonjouklian, R.; Berggren, M.M.; Gallegos, A.; Abraham, R.; Ashendel, C.; Zalkow, L.; Matter, W.F.; Dodge, J.; Grindey, G.; et al. Wortmannin, a potent and selective inhibitor of phosphatidylinositol-3-kinase. Cancer Res. 1994, 54, 2419–2423. [Google Scholar]
- Danilov, A.; Shaposhnikov, M.; Plyusnina, E.; Kogan, V.; Fedichev, P.; Moskalev, A. Selective anticancer agents suppress aging in Drosophila. Oncotarget 2013, 4, 1507–1526. [Google Scholar] [CrossRef]
- García-Martínez, J.M.; Moran, J.; Clarke, R.G.; Gray, A.; Cosulich, S.C.; Chresta, C.M.; Alessi, D.R. Ku-0063794 is a specific inhibitor of the mammalian target of rapamycin (mTOR). Biochem. J. 2009, 421, 29–42. [Google Scholar] [CrossRef]
- Zask, A.; Kaplan, J.; Verheijen, J.C.; Richard, D.J.; Curran, K.; Brooijmans, N.; Bennett, E.M.; Toral-Barza, L.; Hollander, I.; Ayral-Kaloustian, S.; et al. Morpholine derivatives greatly enhance the selectivity of mammalian target of rapamycin (mTOR) inhibitors. J. Med. Chem. 2009, 52, 7942–7945. [Google Scholar] [CrossRef]
- Yu, K.; Shi, C.; Toral-Barza, L.; Lucas, J.; Shor, B.; Kim, J.E.; Zhang, W.-G.; Mahoney, R.; Gaydos, C.; Tardio, L.; et al. Beyond rapalog therapy: preclinical pharmacology and antitumor activity of WYE-125132, an ATP-competitive and specific inhibitor of mTORC1 and mTORC2. Cancer Res. 2010, 70, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Wipf, P.; Minion, D.J.; Halter, R.J.; Berggren, M.I.; Ho, C.B.; Chiang, G.G.; Kirkpatrick, L.; Abraham, R.; Powis, G. Synthesis and biological evaluation of synthetic viridins derived from C(20)-heteroalkylation of the steroidal PI-3-kinase inhibitor wortmannin. Org. Biomol. Chem. 2004, 2, 1911–1920. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.S.; Bowles, D.W.; Falchook, G.S.; Messersmith, W.A.; George, G.C.; O’Bryant, C.L.; Vo, A.C.H.; Klucher, K.; Herbst, R.S.; Eckhardt, S.G.; et al. A multicenter phase I trial of PX-866, an oral irreversible phosphatidylinositol 3-kinase inhibitor, in patients with advanced solid tumors. Clin. Cancer Res. 2012, 18, 4173–4182. [Google Scholar] [CrossRef] [PubMed]
- Klucher, K.M.; Vo, A.; Walker, C.; Rosler, R.; Taylor, J.; Millard, J.; Peterson, S. 17-hydroxy-PX-866, the primary metabolite of PX-866, an irreversible, pan-isoform inhibitor of phosphatidylinositol-3 (PI3) kinase, has increased activity in biochemical and cellular assays. In Proceedings of the AACR Special Conference Targeting PI3K/mTOR signaling in cancer, San Francisco, CA, USA, 24-27 February 2011. [Google Scholar]
- Hayakawa, M.; Kaizawa, H.; Moritomo, H.; Koizumi, T.; Ohishi, T.; Yamano, M.; Okada, M.; Ohta, M.; Tsukamoto, S.-i.; Raynaud, F.I.; et al. Synthesis and biological evaluation of pyrido[3’,2’:4,5]furo[3,2-d]pyrimidine derivatives as novel PI3 kinase p110α inhibitors. Bioorg. Med. Chem. Lett. 2007, 17, 2438–2442. [Google Scholar] [CrossRef]
- Knight, Z.A.; Gonzalez, B.; Feldman, M.E.; Zunder, E.R.; Goldenberg, D.D.; Williams, O.; Loewith, R.; Stokoe, D.; Balla, A.; Toth, B.; et al. A pharmacological map of the PI3-K family defines a role for p110α in insulin signaling. Cell 2006, 125, 733–747. [Google Scholar] [CrossRef]
- Toral-Barza, L.; Zhang, W.-G.; Lamison, C.; LaRocque, J.; Gibbons, J.; Yu, K. Characterization of the cloned full-length and a truncated human target of rapamycin: activity, specificity, and enzyme inhibition as studied by a high capacity assay. Biochem. Biophys. Res. Commun. 2005, 332, 304–310. [Google Scholar] [CrossRef]
- Lehman, J.A.; Calvo, V.; Gomez-Cambronero, J. Mechanism of ribosomal p70S6 kinase activation by granulocyte macrophage colony-stimulating factor in neutrophils. J. Biol. Chem. 2003, 278, 28130–28138. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y.; Yanatori, I.; Tanaka, A.; Kishi, F.; Lemasters, J.J.; Nishina, S.; Sasaki, K.; Hino, K. Iron loss triggers mitophagy through induction of mitochondrial ferritin. EMBO Rep. 2020, 21, e50202. [Google Scholar] [CrossRef]
- Soriano, S.; Llorens, J.V.; Blanco-Sobero, L.; Gutiérrez, L.; Calap-Quintana, P.; Morales, M.P.; Moltó, M.D.; Martínez-Sebastián, M.J. Deferiprone and idebenone rescue frataxin depletion phenotypes in a Drosophila model of Friedreich’s ataxia. Gene 2013, 521, 274–281. [Google Scholar] [CrossRef]
- Sun, H.; Zong, H.; Wu, G. 2-Hydroxypropyl-β-cyclodextrin blocks autophagy flux and triggers caspase-8-mediated apoptotic cascades in HepG2 cells. Mol. Med. Rep. 2020, 22, 1901–1909. [Google Scholar] [CrossRef]
- Massie, H.R.; Williams, T.R.; Iodice, A.A. Influence of anti-inflammatory agents on the survival of Drosophila. J. Gerontol. 1985, 40, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, X.; Li, L.; Wang, D. Lifespan extension in Caenorhabditis elegans by DMSO is dependent on sir-2.1 and daf-16. Biochem. Biophys. Res. Commun. 2010, 400, 613–618. [Google Scholar] [CrossRef]
- Harrison, B.; Tran, T.T.; Taylor, D.; Lee, S.-D.; Min, K.-J. Effect of rapamycin on lifespan in Drosophila. Geriatr. Gerontol. Int. 2010, 10, 110–112. [Google Scholar] [CrossRef]
- Carvalho, G.B.; Kapahi, P.; Benzer, S. Compensatory ingestion upon dietary restriction in Drosophila melanogaster. Nat. Methods 2005, 2, 813–815. [Google Scholar] [CrossRef]
- Kraig, E.; Linehan, L.A.; Liang, H.; Romo, T.Q.; Liu, Q.; Wu, Y.; Benavides, A.D.; Curiel, T.J.; Javors, M.A.; Musi, N.; et al. A randomized control trial to establish the feasibility and safety of rapamycin treatment in an older human cohort: Immunological, physical performance, and cognitive effects. Exp. Gerontol. 2018, 105, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, J.; Mathieu, J.; Alvarez, P. 2-Hydoxypropyl-beta-cyclodextrin (HPβCD) reduces age-related lipofuscin accumulation through a cholesterol-associated pathway. Sci. Rep. 2017, 7, 2197. [Google Scholar] [CrossRef] [PubMed]
- Brunk, U.T.; Terman, A. The mitochondrial-lysosomal axis theory of aging. Eur. J. Biochem. 2002, 269, 1996–2002. [Google Scholar] [CrossRef]
- Ramirez, C.M.; Liu, B.; Taylor, A.M.; Repa, J.J.; Burns, D.K.; Weinberg, A.G.; Turley, S.D.; Dietschy, J.M. Weekly cyclodextrin administration normalizes cholesterol metabolism in nearly every organ of the Niemann-Pick type C1 mouse and markedly prolongs life. Pediatr. Res. 2010, 68, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.; Scott, R.C. 2-Hydroxypropyl-β-cyclodextrin (HP-β-CD): A toxicology review. Food Chem. Toxicol. 2005, 43, 1451–1459. [Google Scholar] [CrossRef]
- Trotta, F.; Zanetti, M.; Camino, G. Thermal degradation of cyclodextrins. Polym. Degrad. Stab. 2000, 69, 373–379. [Google Scholar] [CrossRef]
- Mockett, R.J.; Nobles, A.C. Lack of robustness of life extension associated with several single-gene P element mutations in Drosophila melanogaster. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Orr, W.C.; Sohal, R.S. The effects of catalase gene overexpression on life span and resistance to oxidative stress in transgenic Drosophila melanogaster. Arch. Biochem. Biophys. 1992, 297, 35–41. [Google Scholar] [CrossRef]
- Mockett, R.J.; Bearden, A.A.; Nobles, A.C. Longevity effects of DMSO-solubilized rapamycin and other compounds in y w male Drosophila melanogaster. FASEB J. 2022, 36S1. [Google Scholar] [CrossRef]
Figure 1.
Survival times of flies supplemented with DMSO, PP242 or DFP beginning in late adult life (64 days). Red: life shortening vs. both regular and H2O controls (p < 0.050 based on logrank tests).
Figure 1.
Survival times of flies supplemented with DMSO, PP242 or DFP beginning in late adult life (64 days). Red: life shortening vs. both regular and H2O controls (p < 0.050 based on logrank tests).
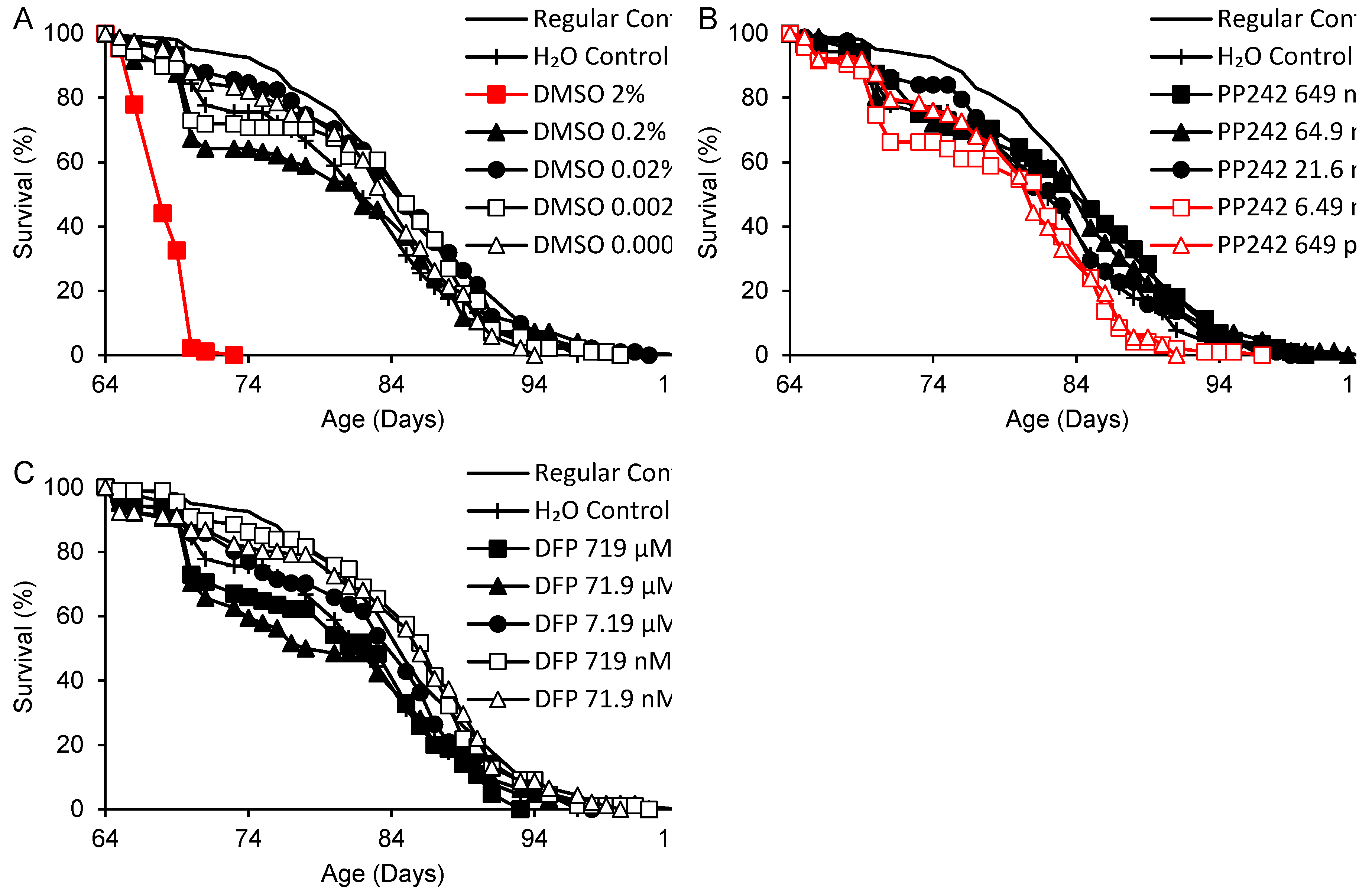
Figure 2.
Life spans of flies supplemented with spermidine, DMSO or TOR/PI3K inhibitors: AZD8055, Ku-0063794, LY294002, PI-103 hydrochloride (HCl), PX-866-17OH, rapamycin (Rapa), Torin2, wortmannin (Wort), WYE-28 and WYE-132. Supplements were provided beginning in early adult life (arrows). Green: life extension and red: life shortening vs. both regular and H2O controls, or vs. H2O controls alone for PI-103 hydrochloride (p < 0.050 based on logrank tests).
Figure 2.
Life spans of flies supplemented with spermidine, DMSO or TOR/PI3K inhibitors: AZD8055, Ku-0063794, LY294002, PI-103 hydrochloride (HCl), PX-866-17OH, rapamycin (Rapa), Torin2, wortmannin (Wort), WYE-28 and WYE-132. Supplements were provided beginning in early adult life (arrows). Green: life extension and red: life shortening vs. both regular and H2O controls, or vs. H2O controls alone for PI-103 hydrochloride (p < 0.050 based on logrank tests).
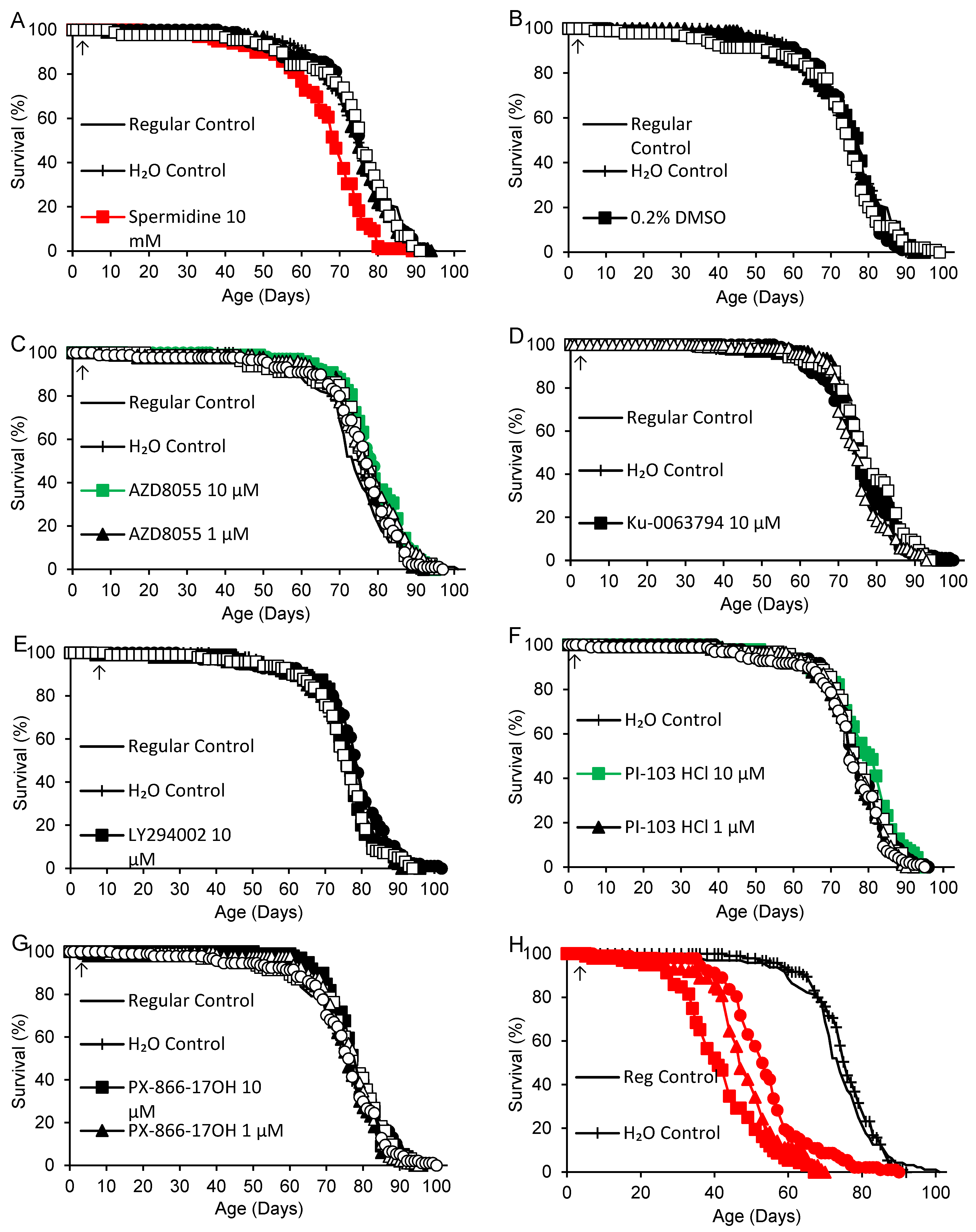
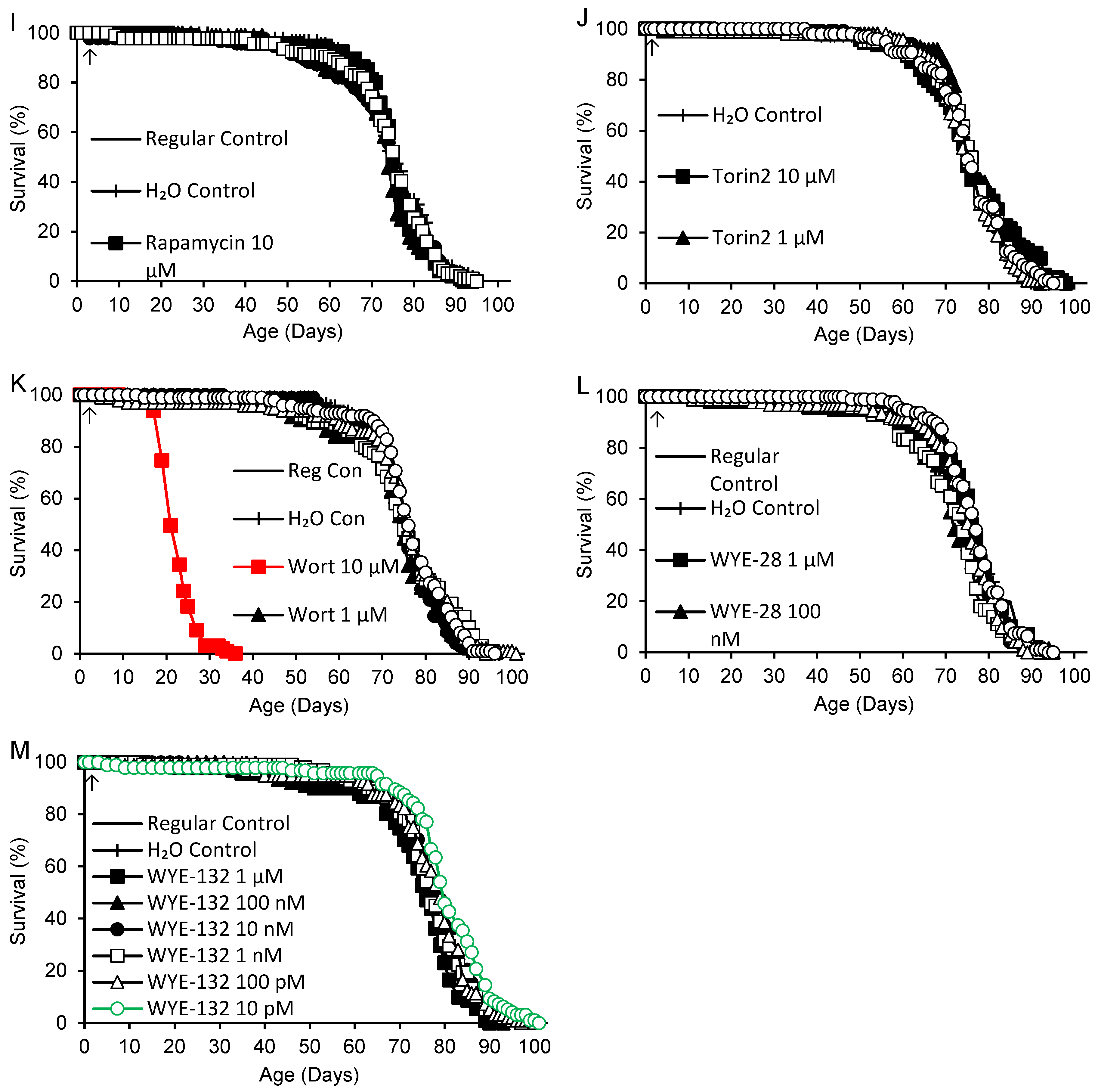
Figure 3.
Life spans of flies supplemented with AZD8055, PI-103 hydrochloride (HCl), rapamycin, WYE-132 or DMSO control beginning at 2 days of adult life (arrows). Green: life extension vs. regular, H2O and DMSO controls (p < 0.050 based on logrank tests).
Figure 3.
Life spans of flies supplemented with AZD8055, PI-103 hydrochloride (HCl), rapamycin, WYE-132 or DMSO control beginning at 2 days of adult life (arrows). Green: life extension vs. regular, H2O and DMSO controls (p < 0.050 based on logrank tests).
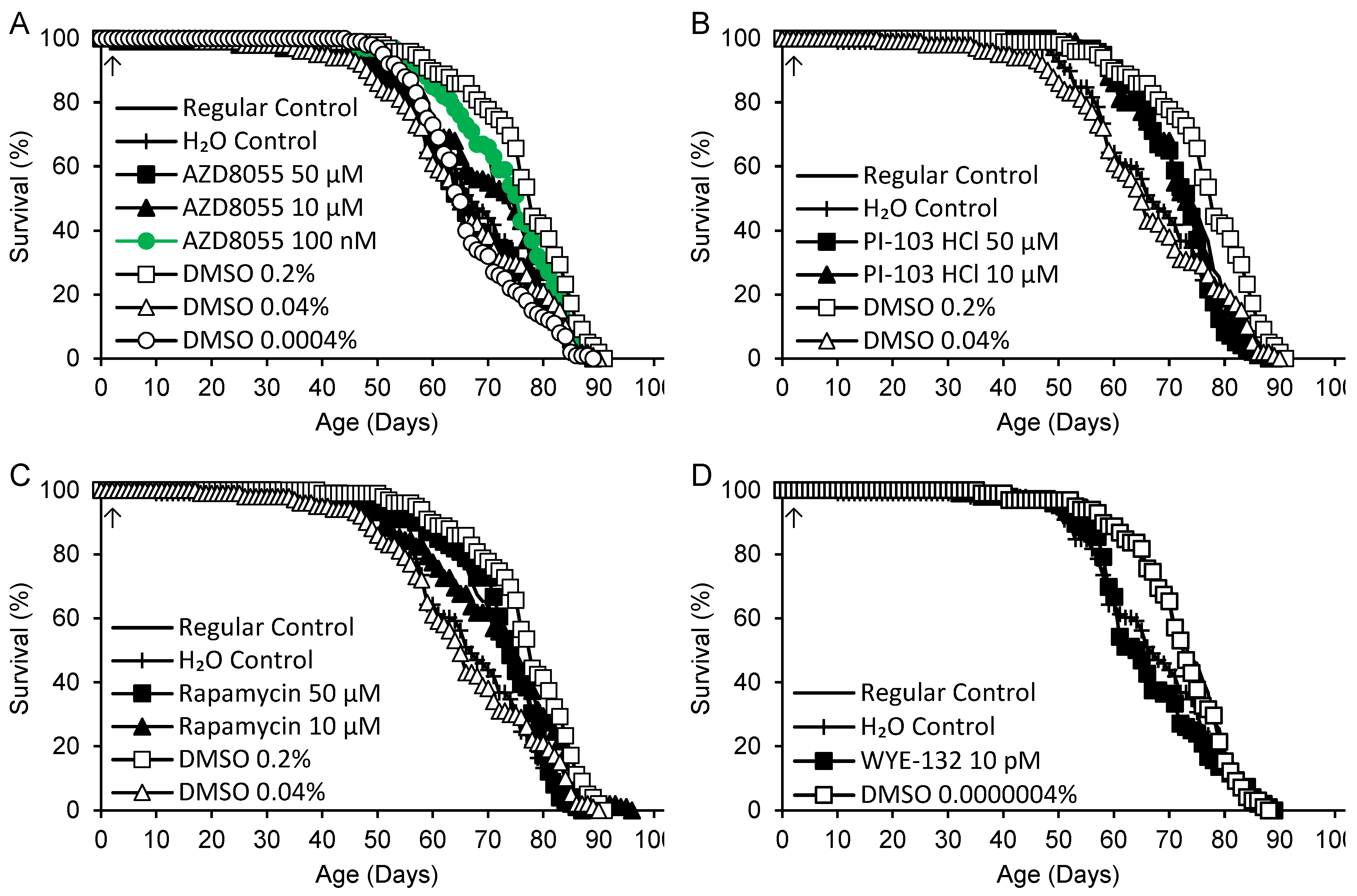
Figure 4.
Fecundity (A) and fertility (B) of flies supplemented with AZD8055, PI-103 hydrochloride (HCl), rapamycin, WYE-132 or DMSO control. Results (mean ± SEM) are numbers of eggs laid per female per day during the first four days of adulthood (A) and the corresponding numbers of adults developed from those eggs within 14 days at 25 °C (B). *p < 0.017.
Figure 4.
Fecundity (A) and fertility (B) of flies supplemented with AZD8055, PI-103 hydrochloride (HCl), rapamycin, WYE-132 or DMSO control. Results (mean ± SEM) are numbers of eggs laid per female per day during the first four days of adulthood (A) and the corresponding numbers of adults developed from those eggs within 14 days at 25 °C (B). *p < 0.017.
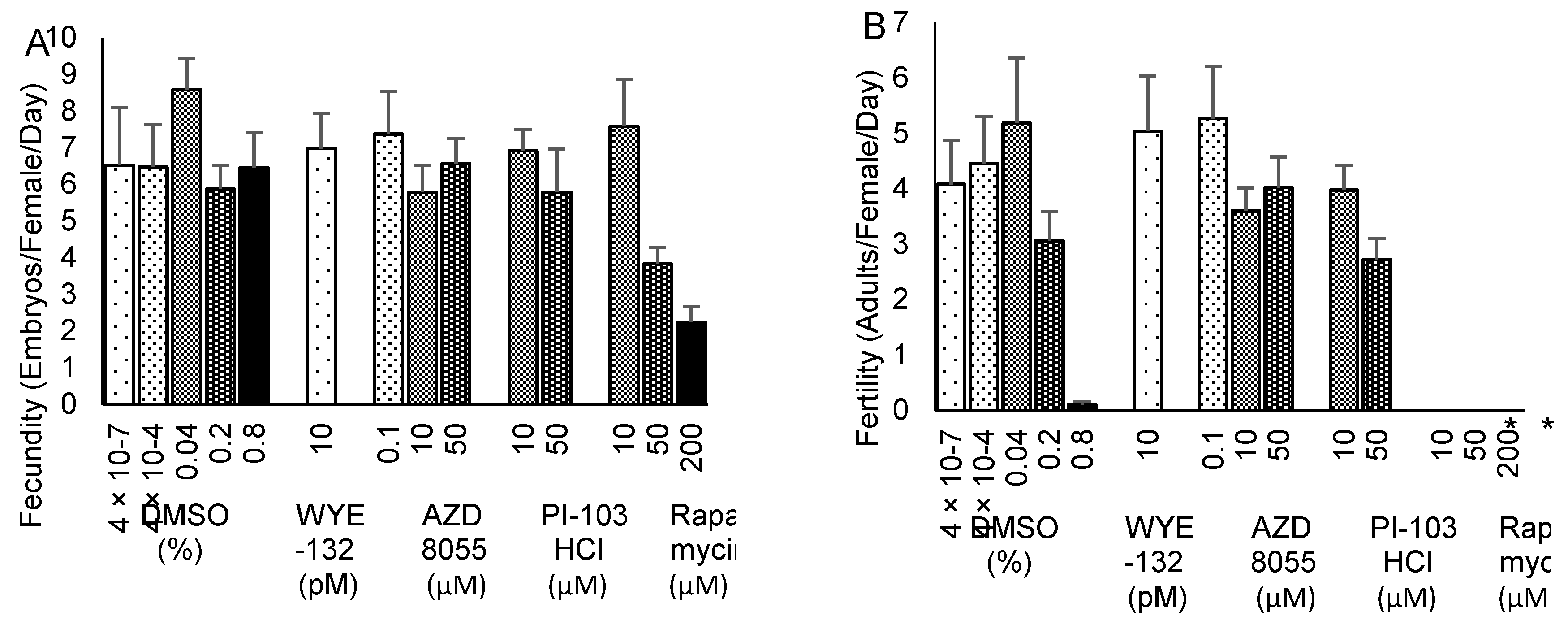
Figure 5.
Survival times of flies supplemented with 2-HP-β-CD beginning either at 50 days (A,B) or 1 day of adult life (arrows) (C,D). (A-C) Standard medium. (D) High yeast-high sugar medium. Red: life shortening vs. H2O control (p < 0.017 based on logrank tests with Bonferroni correction for three comparisons).
Figure 5.
Survival times of flies supplemented with 2-HP-β-CD beginning either at 50 days (A,B) or 1 day of adult life (arrows) (C,D). (A-C) Standard medium. (D) High yeast-high sugar medium. Red: life shortening vs. H2O control (p < 0.017 based on logrank tests with Bonferroni correction for three comparisons).
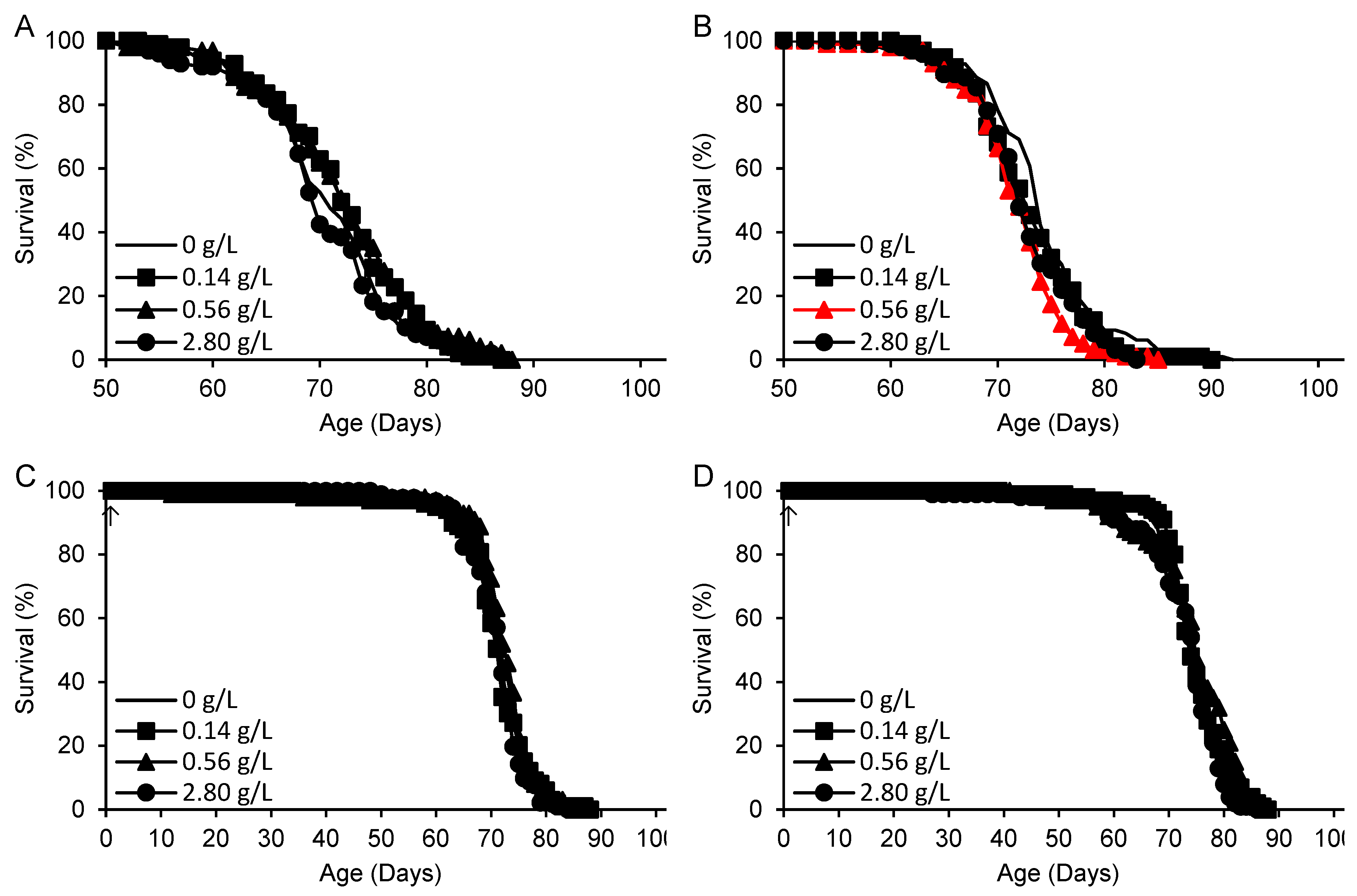

Table 1.
IC50 of TOR/PI3 kinase inhibitors.1
| Inhibitor | TORC1 (nM) 2 | TORC2 (nM) 3 | PI3Kinase (nM) 4 | Reference |
|---|---|---|---|---|
| AZD8055 | 0.13 ± 0.05 5 | 3200-18 900 6 | 39 | |
| Ku-0063794 | ~10 | ~10 | >10 000 | 50 |
| LY294002 | 1500 5 ~5000 7 . | 1400 | 43,44,58 | |
| PI-103 hydrochloride 8 | 20 | 83 | 3-250 6 | 56,57 |
| PP242 | 30 9 | 58 | 102-2200 6 | 40 |
| PX-866-17OH | 10 | 14-57 6,10 | 54 | |
| Rapamycin | 2 5,11 | — | — | 58 |
| Torin2 | 0.25 12 | 200 12 | 36 | |
| Wortmannin | 200 5 ~200 7, 300 13 . | 0.3-4 | 44,47,48,53,58 | |
| WYE-28 | 0.22 ± 0.06 5 | 4271 | 51 | |
| WYE-132 | 0.19 ± 0.07 5 | 3 | 1179->10 000 6 | 52 |
1 — = not an inhibitor of the listed kinase. Blank spaces indicate no known test data. Values may not be directly comparable between inhibitors in cases where different substrates were used for IC50 determinations. 2 Substrate biotinylated S6K for AZD8055, S6K1 T389 for Ku-0063794 and Torin2, His6-S6K T389 for LY294002, rapamycin, wortmannin, WYE-28 and WYE-132, except as noted in footnotes below, 4E-BP1 for PI-103 and PP242. 3 Substrate Akt for PI-103, Akt S473 for Ku-0063794 and WYE-132 (IC50 not stated for WYE-132 but comparable to TORC1 IC50 based on immunoblot [52]), mTOR labeling in presence of PKB for PP242. 4 Substrate PIP2 for AZD8055, Ku-0063794, WYE-28 and WYE-132, phosphatidylinositol for LY294002 [43], PI-103, PP242 [40 Ref. 21] and wortmannin [47,48], Akt T308 for Torin2. Substrate not specified for PX-866-17OH. 5 Values based on truncated, FLAG-tagged mTOR. For AZD8055, full-length mTOR IC50 = 0.8 ± 0.2 nM with 4E-BP-1 substrate. Values for AZD8055 also varied with ATP concentration, up to ~15 nM at 200 µM ATP. 6 Multiple values represent tests in >1 PI3K class I isoform (PI3Kα, β, γ or δ). 7 Values reported for TOR autophosphorylation without differentiating TORC1 vs. TORC2 [44]. 8 Values for PI-103. 9 8 nM for mTOR (Invitrogen). 10 For parent compound PX-866, TOR IC50 >10 000 nM, PI3K IC50 = 0.5 nM [53] or PI3K IC50 = 39-183 nM [54]; assay methods not specified. 11 0.2 nM for inhibition of neutrophil S6K enzymatic activity, not linked conclusively to TOR [59]. 12 Cellular EC50 values. In vitro mTOR IC50 = 2.1 nM with substrate GFP-4E-BP1. 13 Method not specified [53].
Table 2.
Mean survival times of flies supplemented with DMSO, PP242 or DFP beginning at 64 days of adult life.
Table 2.
Mean survival times of flies supplemented with DMSO, PP242 or DFP beginning at 64 days of adult life.
| Supplement (Concentration) 1 | p 2 | Survival (Days) | % vs. H2O (p) | % vs. Regular (p) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control (Regular) | 21.0 | ||||||||
| Control (H2O) | 17.4 | -17.0 | (0.002) | ||||||
| DMSO | <0.0005 | ||||||||
| DMSO (2%) | 4.4 | -74.7 | (< 0.0005) | -79.0 | (< 0.0005) | ||||
| DMSO (0.2%) | 16.1 | -7.4 | (0.92) | -23.1 | |||||
| DMSO (0.02%) | 19.8 | +13.7 | (0.055) | -5.6 | |||||
| DMSO (0.002%) | 18.1 | +3.8 | (0.21) | -13.8 | |||||
| DMSO (0.0002%) | 18.5 | +6.3 | (0.79) | -11.8 | |||||
| PP242 | <0.0005 | ||||||||
| PP242 (649 nM) | 19.1 | +9.7 | (0.048) | -8.9 | (0.60) | ||||
| PP242 (64.9 nM) | 18.5 | +6.0 | (0.15) | -12.0 | |||||
| PP242 (21.6 nM) | 18.2 | +4.3 | (0.63) | -13.4 | |||||
| PP242 (6.49 nM) | 15.0 | -13.7 | (0.030) | -28.3 | (< 0.0005) | ||||
| PP242 (649 pM) | 15.9 | -8.8 | (0.028) | -24.3 | (< 0.0005) | ||||
| DFP | 0.003 | ||||||||
| DFP (719 µM) | 16.2 | -6.9 | (0.45) | -22.7 | |||||
| DFP (71.9 µM) | 15.9 | -8.9 | (0.99) | -24.3 | |||||
| DFP (7.19 µM) | 18.6 | +6.4 | (0.19) | -11.6 | |||||
| DFP (719 nM) | 20.9 | +19.6 | (0.012) | -0.7 | (0.98) | ||||
| DFP (71.9 nM) | 20.2 | +16.0 | (0.006) | -3.7 | (0.82) | ||||
1 Concentrations are final values after diluting 100 µL supplement in 4.9 mL fly medium. 2 p values for each supplement are for logrank tests including the H2O control group and all concentrations of the supplement. Where p < 0.050, tests were performed for each dose individually. Where p < 0.050 again, comparisons were also made with the regular control group.
Table 3.
Adult mean life spans of flies supplemented with DMSO, spermidine or TOR/PI3K inhibitors beginning in early adult life.
Table 3.
Adult mean life spans of flies supplemented with DMSO, spermidine or TOR/PI3K inhibitors beginning in early adult life.
| Supplement (Concentration) 1 | p 2 | Life Span (Days) | % vs. H2O (p) | % vs. Regular (p) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control (Regular) | 73.9 | ||||||||
| Control (H2O) | 74.1 | 0.2 | |||||||
| Spermidine | <0.0005 | ||||||||
| Spermidine (10 mM) | <0.0005 | 66.2 | -10.6 | (<0.0005) | -10.4 | (<0.0005) | |||
| Spermidine (1 mM) | 0.87 | 73.6 | -0.7 | -0.5 | |||||
| Spermidine (100 µM) | 0.92 | 74.3 | +0.2 | +0.5 | |||||
| Spermidine (10 µM) | 0.93 | 73.5 | -0.8 | -0.6 | |||||
| DMSO | 0.89 | ||||||||
| DMSO (0.2%) | 73.3 | -1.1 | -0.9 | ||||||
| DMSO (0.1%) | 73.2 | -1.2 | -1.0 | ||||||
| DMSO (0.04%) | 73.0 | -1.5 | -1.3 | ||||||
| DMSO (0.02%) | 72.0 | -2.9 | -2.6 | ||||||
| WYE-28 | 0.14 | ||||||||
| WYE-28 (1 µM) | 74.6 | +0.7 | +0.9 | ||||||
| WYE-28 (100 nM) | 72.4 | -2.3 | -2.1 | ||||||
| WYE-28 (10 nM) | 74.0 | -0.1 | +0.1 | ||||||
| WYE-28 (1 nM) | 71.8 | -3.0 | -2.8 | ||||||
| WYE-28 (100 pM) | 74.2 | +0.1 | +0.4 | ||||||
| WYE-28 (10 pM) | 76.6 | +3.4 | +3.6 | ||||||
| Control (Regular) | 76.2 | ||||||||
| Control (H2O) | 75.7 | -0.6 | |||||||
| WYE-132 | <0.0005 | ||||||||
| WYE-132 (1 µM) | 0.083 | 73.5 | -3.0 | -3.6 | |||||
| WYE-132 (100 nM) | 0.53 | 76.1 | +0.4 | -0.2 | |||||
| WYE-132 (10 nM) | 0.57 | 76.6 | +1.1 | +0.5 | |||||
| WYE-132 (1 nM) | 0.60 | 76.5 | +1.0 | +0.4 | |||||
| WYE-132 (100 pM) | 0.31 | 77.3 | +2.0 | +1.4 | |||||
| WYE-132 (10 pM) | 0.001 | 80.3 | +6.0 | (<0.0005) | +5.3 | (0.012) | |||
| LY294002 | 0.12 | ||||||||
| LY294002 (10 µM) | 75.6 | -0.2 | -0.8 | ||||||
| LY294002 (5 µM) | 75.3 | -0.5 | -1.1 | ||||||
| LY294002 (1 µM) | 76.9 | +1.5 | +0.9 | ||||||
| LY294002 (100 nM) | 74.2 | -2.0 | -2.6 | ||||||
| Control (Regular) | 73.3 | ||||||||
| Control (H2O) | 75.3 | +2.7 | |||||||
| AZD8055 | 0.036 | ||||||||
| AZD8055 (10 µM) | 0.009 | 78.1 | +3.7 | (0.006) | +6.6 | (0.006) | |||
| AZD8055 (1 µM) | 0.29 | 76.0 | +0.9 | +3.7 | |||||
| AZD8055 (100 nM) | 0.013 | 78.1 | +3.7 | (0.011) | +6.5 | (0.008) | |||
| AZD8055 (10 nM) | 0.38 | 75.4 | +0.1 | +2.8 | |||||
| AZD8055 (1 nM) | 0.078 | 76.7 | +1.8 | +4.6 | |||||
| AZD8055 (100 pM) | 0.45 | 75.1 | -0.3 | +2.4 | |||||
| Rapamycin | <0.0005 | ||||||||
| Rapamycin (400 µM) | <0.0005 | 41.8 | -44.5 | (<0.0005) | -42.9 | (<0.0005) | |||
| Rapamycin (200 µM) | <0.0005 | 47.7 | -36.7 | (<0.0005) | -34.9 | (<0.0005) | |||
| Rapamycin (100 µM) | <0.0005 | 54.2 | -28.0 | (<0.0005) | -26.1 | (<0.0005) | |||
| Control (Regular) | 73.4 | ||||||||
| Control (H2O) | 75.7 | +3.2 | |||||||
| Rapamycin | 0.22 | ||||||||
| Rapamycin (10 µM) | 74.2 | -2.0 | +1.1 | ||||||
| Rapamycin (1 µM) | 72.5 | -4.3 | -1.3 | ||||||
| Rapamycin (500 nM) | 73.1 | -3.4 | -0.3 | ||||||
| Rapamycin (100 nM) | 73.5 | -3.0 | +0.1 | ||||||
| Ku-0063794 | 0.062 | ||||||||
| Ku-0063794 (10 µM) | 75.8 | +0.1 | +3.3 | ||||||
| Ku-0063794 (1 µM) | 77.2 | +1.9 | +5.1 | ||||||
| Ku-0063794 (100 nM) | 75.6 | -0.1 | +3.0 | ||||||
| Ku-0063794 (10 nM) | 77.3 | +2.1 | +5.3 | ||||||
| Ku-0063794 (1 nM) | 74.7 | -1.4 | +1.8 | ||||||
| Wortmannin | <0.0005 | ||||||||
| Wortmannin (10 µM) | <0.0005 | 22.6 | -70.2 | (<0.0005) | -69.2 | (<0.0005) | |||
| Wortmannin (1 µM) | 0.21 | 72.6 | -4.1 | -1.0 | |||||
| Wortmannin (500 nM) | 0.27 | 74.7 | -1.4 | +1.8 | |||||
| Wortmannin (100 nM) | 0.11 | 73.9 | -2.4 | +0.7 | |||||
| Wortmannin (10 nM) | 0.11 | 74.4 | -1.7 | +1.4 | |||||
| Wortmannin (1 nM) | 0.092 | 76.2 | +0.7 | +3.8 | |||||
| Control (Regular) | 75.5 | ||||||||
| Control (H2O) | 75.4 | -0.1 | |||||||
| PX-866-17OH | 0.26 | ||||||||
| PX-866-17OH (10 µM) | 78.0 | +3.5 | +3.4 | ||||||
| PX-866-17OH (1 µM) | 74.2 | -1.6 | -1.7 | ||||||
| PX-866-17OH (500 nM) | 76.7 | +1.7 | +1.6 | ||||||
| PX-866-17OH (100 nM) | 75.6 | +0.2 | +0.1 | ||||||
| PX-866-17OH (10 nM) | 76.7 | +1.7 | +1.6 | ||||||
| PX-866-17OH (1 nM) | 74.5 | -1.2 | -1.3 | ||||||
| Control (H2O) | 75.4 | ||||||||
| PI-103 HCl | 0.029 | ||||||||
| PI-103 HCl (10 µM) | 0.010 | 78.4 | +4.0 | ||||||
| PI-103 HCl (1 µM) | 0.95 | 75.2 | -0.3 | ||||||
| PI-103 HCl (100 nM) | 0.41 | 76.6 | +1.5 | ||||||
| PI-103 HCl (10 nM) | 0.36 | 76.4 | +1.3 | ||||||
| PI-103 HCl (1 nM) | 0.91 | 75.6 | +0.3 | ||||||
| PI-103 HCl (100 pM) | 0.75 | 74.4 | -1.3 | ||||||
| Torin2 | 0.40 | ||||||||
| Torin2 (10 µM) | 75.3 | -0.1 | |||||||
| Torin2 (1 µM) | 76.3 | +1.2 | |||||||
| Torin2 (100 nM) | 75.4 | -0.1 | |||||||
| Torin2 (10 nM) | 74.9 | -0.7 | |||||||
| Torin2 (1 nM) | 75.0 | -0.6 | |||||||
| Torin2 (100 pM) | 75.3 | -0.1 | |||||||
1 Concentrations are final values after diluting 100 µL supplement in 4.9 mL fly medium. 2 p values for each supplement are for logrank tests including both control groups and all concentrations of the supplement. Where p < 0.050, tests were performed for each dose and both control groups. Where p < 0.050 again, comparisons were also made for each control group separately.
Table 4.
Adult mean life spans of flies supplemented with AZD8055, PI-103 hydrochloride (HCl), rapamycin (Rapa) or WYE-132, and DMSO control groups – replication experiment.
Table 4.
Adult mean life spans of flies supplemented with AZD8055, PI-103 hydrochloride (HCl), rapamycin (Rapa) or WYE-132, and DMSO control groups – replication experiment.
| Supplement (Concentration) 1 | p 2 | Life Span (Days) | % vs. H2O (p) | % vs. Regular (p) | % vs. DMSO (p) 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control (Regular) | 72.5 | ||||||||||
| Control (H2O) | 66.8 | -7.8 | (0.065) | ||||||||
| DMSO | <0.0005 | ||||||||||
| DMSO (0.2%) | <0.0005 | 76.5 | +14.5 | (<0.0005) | +5.6 | (<0.0005) | |||||
| DMSO (0.04%) | 0.20 | 65.6 | -1.9 | -9.5 | |||||||
| DMSO (0.0004%) | 0.022 | 66.6 | -0.3 | (0.45) | -8.1 | (0.005) | |||||
| DMSO (0.0000004%) | 0.12 | 71.8 | +7.4 | -1.0 | |||||||
| AZD8055 | 0.001 | ||||||||||
| AZD8055 (50 µM) | 0.16 | 66.6 | -0.4 | -8.1 | -13.0 | ||||||
| AZD8055 (10 µM) | 0.030 | 69.9 | +4.6 | (0.020) | -3.6 | (0.39) | +6.6 | (0.12) | |||
| AZD8055 (100 nM) | 0.001 | 73.4 | +9.8 | (<0.0005) | +1.3 | (0.030) | +10.2 | (<0.0005) | |||
| PI-103 HCl | 0.10 | ||||||||||
| PI-103 HCl (50 µM) | 71.8 | +7.5 | -0.9 | -6.1 | |||||||
| PI-103 HCl (10 µM) | 72.2 | +8.1 | -0.3 | +10.2 | |||||||
| Rapamycin | 0.003 | ||||||||||
| Rapamycin (50 µM) | 0.12 | 72.2 | +8.0 | -0.4 | -5.6 | ||||||
| Rapamycin (10 µM) | 0.002 | 71.4 | +6.9 | (0.002) | -1.4 | (0.069) | +9.0 | (0.007) | |||
| WYE-132 (10 pM) | 0.033 | 66.0 | -1.2 | (0.59) | -8.9 | (0.008) | -8.0 | (0.016) | |||
1 Concentrations are final values after diluting 100 µL supplement in 4.9 mL fly medium. 2 p values for each supplement are for logrank tests including regular and H2O control groups and all concentrations of the supplement. Where p < 0.050, tests were performed for each dose and both control groups. Where p < 0.050 again, comparisons were made for each control group separately. 3 50 µM supplement vs. 0.2% DMSO, 10 µM supplement vs. 0.04% DMSO, 100 nM supplement vs. 0.0004% DMSO, 10 pM supplement vs. 0.0000004% DMSO.
Table 5.
Survival times of flies supplemented with 2-HP-β-CD beginning at 50 days and life spans of flies supplemented with 2-HP-β-CD beginning at 1 day of adult life.
Table 5.
Survival times of flies supplemented with 2-HP-β-CD beginning at 50 days and life spans of flies supplemented with 2-HP-β-CD beginning at 1 day of adult life.
| Supplement (Concentration) 1 | p 2 | Survival (Days) | % vs. H2O (p) | Medium (Figure) | Supplement Onset (Days) | |
|---|---|---|---|---|---|---|
| Control (H2O) | 0.079 | 20.8 | Standard | 50 | ||
| 2-HP-β-CD (0.14 g/L) | 22.1 | +6.5 | (5A) | |||
| 2-HP-β-CD (0.56 g/L) | 22.3 | +7.1 | ||||
| 2-HP-β-CD (2.80 g/L) | 20.5 | -1.6 | ||||
| Control (H2O) | 0.002 | 24.2 | Standard | 50 | ||
| 2-HP-β-CD (0.14 g/L) | 23.1 | -4.8 | (0.12) | (5B) | ||
| 2-HP-β-CD (0.56 g/L) | 21.8 | -10.2 | (<0.0005) | |||
| 2-HP-β-CD (2.80 g/L) | 22.6 | -6.6 | (0.017) | |||
| Control (H2O) | 0.31 | 71.1 | Standard | 1 | ||
| 2-HP-β-CD (0.14 g/L) | 70.9 | -0.2 | (5C) | |||
| 2-HP-β-CD (0.56 g/L) | 72.0 | +1.3 | ||||
| 2-HP-β-CD (2.80 g/L) | 71.0 | -0.0 | ||||
| Control (H2O) | 0.040 | 71.0 | High | 1 | ||
| 2-HP-β-CD (0.14 g/L) | 74.6 | +5.1 | (0.27) | Sugar/ | ||
| 2-HP-β-CD (0.56 g/L) | 74.3 | +4.6 | (0.032) | Yeast | ||
| 2-HP-β-CD (2.80 g/L) | 72.7 | +2.3 | (0.65) | (5D) | ||
1 Concentrations are final values after diluting 1 mL supplement in 99 mL fly medium. 2 p values for each experiment are for logrank tests including the H2O control group and all concentrations of 2-HP-β-CD. Where p < 0.050, tests were performed for each dose vs. the H2O control group. The Bonferroni-adjusted significance threshold for the multiple comparisons was p < 0.017.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



