Submitted:
25 August 2024
Posted:
27 August 2024
You are already at the latest version
Abstract
Bacillus amyloliquefaciens GL18 has been proven to have good resistance ability and can promote the growth of Avena sativa cultivar ‘Qingyan 1’. Herein, Illumina high-throughput transcriptome sequencing technology was used to analyze the differential expression of genes between the interaction group (LB) and the control group (LCK) when ‘Qingyan 1’ roots interacted with GL18 bacterial suspension or sterile water for 12 h under 4 °C low-temperature stress. The gene function annotation information was obtained using Gene Ontology (GO) functional classification and enrichment analysis. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis and annotation of differentially expressed genes was used to explore the related signal transduction pathway of ‘Qingyan 1’ response to cold stress induced by Bacillus GL18. The results showed that after ‘Qingyan 1’ seedlings interacted with GL18 bacterial suspension or sterile water for 12 h, comparing the LB with the LCK or CK (the control group at 26 °C), there were 12,061 and 25,489 differentially expressed genes, respectively. Their functions were mainly related to metabolic process, cellular process, catalytic activity, binding, and cellular parts, and they were significantly enriched in plant tissue growth and development, cell wall, organelle membrane synthesis, thiamine and its compounds, biosynthesis, polyamine decomposition, and other metabolic processes. Signal transduction was detected through KEGG pathway enrichment analysis, and corresponding genes, such as AUXI, PP2C, SNRK2, ABF, BRI1, and PIF4, were expressed. The expression of genes directly involved in plant growth was upregulated in the presence of GL18, as shown by real-time quantitative PCR analysis. It was speculated that the normal growth and development of ‘Qingyan 1’ under low-temperature stress promoted by GL18 is a result of the integration of multiple pathways in terms of regulating endogenous hormone metabolism, secondary metabolite synthesis, and photosynthesis.
Keywords:
low-temperature stress
; Bacillus amyloliquefaciens
; ‘Qingyan 1’
; transcriptome
; signal transduction pathway analysis
1. Introduction
Abiotic stress is the primary factor limiting agricultural development in alpine regions, and it inhibits plant growth by affecting plant photosynthesis, pigment content, and water relationships, thereby reducing yield and quality[1,2]. Among them, temperature stress severely inhibits plant life activities and even leads to the destruction of cell and tissue structures, which is not conducive to plant growth and development [3]. High temperature mainly leads to changes in enzyme activity by enhancing plant transpiration, thus affecting plant growth, photosynthesis, and metabolism of certain substances [4]. Under 45°C heat stress conditions, wheat (Triticum aestivum) seedlings have accelerated senescence, reduced photosynthesis, photosynthetic enzyme inactivation, and chloroplast oxidative damage [5]. However, low-temperature stimulation can damage plant cell structure and affect cell metabolism, thus affecting its growth. Studies have found that plant responses to low-temperature stimulation mainly involve the hormone signaling pathway ABC transporter, peroxisome, ubiquitination proteolysis, and the calcium regulatory signaling pathway [6,7].
The low temperatures, hypoxia, drought, and other conditions in the Qinghai-Tibet Plateau have limited the growth of vegetation in the alpine pastoral areas to a certain extent. The short growth period and low forage yield of grass in the extraction area, coupled with rodent damage, overgrazing, and other factors, further aggravate vegetation degradation and forage shortage in alpine grasslands [8,9]. Therefore, improving the resistance of alpine forage to extreme habitats and maintaining its normal growth and development have become important measures for solving forage shortage and ecological restoration in the alpine region. Avena sativa cultivar ‘Qingyan 1’ is endemic to the Qinghai-Tibet Plateau, and it was systematically bred from ‘Qingyongjiu 146’ (Denmark) and ‘Bayan No. 3’ (a hybrid bred by the Qinghai Academy of Animal Husbandry and Veterinary Sciences). A. sativa cultivar ‘Qingyan 1’ has the characteristics of early maturity, prolongation resistance, drought tolerance, and barren resistance. It can complete the whole growth period in different areas at an altitude of 1745–3103 m and has unique survival and adaptation abilities in the alpine environment of the Tibetan Plateau [10]. In addition, its stems and leaves are tender and juicy, with good palatability and high nutritional value. It can be used as a high-quality livestock feed and provides a renewable energy source, which is an important basis for stably promoting the development of animal husbandry in alpine pastoral areas [11]. Furthermore, ‘Qingyan 1’ has well-developed roots and good water retention ability, and it can be used as an important cultivated forage for ecological environment protection and soil erosion prevention on the Qinghai-Tibet Plateau [12]. Although ‘Qingyan 1’ prefers coolness and has strong adaptability to adversity, cold still affects its growth and development to a certain extent. Especially, cold inhibits seedling growth, causing leaf tips to turn yellow, wilt, and grow slowly. It also causes tissue necrosis of seedlings [13].
Special microbial resources are also bred in the special habitats of the Qinghai-Tibet Plateau. Bacillus, as an important biocontrol bacterium, can produce endophytic spores that can resist adverse environments, such as salt, drought, cold, and ultraviolet light, and once formed, spores can be preserved for a long time; thus, it tolerates various adverse conditions [14]. Bacillus directly alleviates the harm of stress on plant growth and development by improving the absorption rate of plant nutrients; water and mineral intake; and the formation of extracellular polysaccharides, biofilms, and many organic solutes [15]. Plant resistance to stress can also be indirectly improved by regulating plant chemotaxis, hormone synthesis, and levels, activating the antioxidant defense system, and regulating stress-response genes [16]. Zubair et al. [17] showed that B. psychrophilus CJCL2 and RJGP41 positively regulated plant response to stress by regulating abscisic acid, lipid peroxidation, and proline accumulation, thus significantly improving the growth of wheat (T. aestivum) under cold stress. It has also been found that B. atrophaeus TS1 can induce the expression of defense-related genes in wheat (T. aestivum) under low-temperature stress, regulate the upregulated expression of lipoxygenase-coding genes and mitogen-activated protein kinase, and regulate the normal growth of plants under low-temperature [18]. In addition, B. amyloliquefaciens GL18 has good resistance to the plateau environment and can withstand low temperatures at 4 °C, 11% NaCl concentration, and 15% PEG-6000 concentration, showing certain low temperature, salt, and drought tolerance. Moreover, it has a good growth-promoting effect [19]. Therefore, it is proposed that beneficial microorganisms interact with plants to improve plant growth by improving nutrient absorption, plant hormones, and osmotic substances and inhibiting the synthesis of pathogen biomolecules. In this study, the interaction system between Bacillus GL18 and ‘Qingyan 1’ was established under low-temperature stress, and the molecular mechanism of GL18 promoting its growth was explored using transcriptomic sequencing.
2. Materials and Methods
2.1. Bacterial Strain and Plant Material
Bacillus amyloliquefaciens strain GL18 was isolated from the rhizosphere of Kobresia myosuroides in an alpine meadow in Nianbaoyuze, Guoluo Prefecture, Qinghai Province.
Avena sativa cultivar “Qingyan 1” was provided by the Key Laboratory of Superior Forage Germplasm in the Qinghai-Tibetan Plateau.
2.2. Medium and Soil
LB medium was used for Bacillus culture [20]. Natural soil (taken from Qinghai University experimental field) and vermiculite (purchased from Qinghai Province Xining Flower Exhibition Center).
2.3. GL18 Bacteria Suspension
Strain GL18 was inoculated in liquid medium containing 100 mL LB and incubated in shaking table at 37 °C and 200 r/min for 24 h as seed liquid, then the seed solution was inoculated into a triangular bottle containing 300 mL LB liquid medium at 3% inoculation rate, and continued to culture for 24 h. After the culture, the bacterial solution was removed, centrifuged at 4 °C, 8000 r/min for 10 min, precipitated, washed the bacterial cells with sterile water twice, and prepared into bacterial suspension with sterile water at a concentration of 1×107 cfu/mL (OD600=1.0) [21].
2.4. Interaction between ‘Qingyan 1’ Root and GL18 Bacterial Suspension
Natural soil and vermiculite were mixed at a mass ratio of 2:1 and sterilized at 120 °C. The intact ‘Qingyan 1’ seeds were selected and put in 20% sodium hypochlorite solution, disinfected for 20 min, and rinsed off with sterile water. The disinfected seeds were planted in flowerpots at 25 seeds per pot and cultured in a light incubator (26 °C, photoperiod of 16/8 h) [22]. When the height of the ‘Qingyan 1’ seedlings reached approximately 8 cm, they were carefully taken out, the roots were rinsed with sterile water, and filter paper was used to absorb the water from the roots. Next, two temperatures and three treatments were set: LCK (sterile water control at 4 °C), LB (GL18 bacterial suspension at 4 °C), and CK (sterile water control at 26 °C). The roots of the seedlings in the three groups were completely immersed in 35 mL sterile water or GL18 bacterial suspension (cell concentration of 1 × 107 CFU/mL) to establish an interaction system between ‘Qingyan 1’ and Bacillus GL18, and then the interaction time was set to 12 h (Figure 1).
2.5‘. Qingyan 1’ Root for Transcriptome Sequencing
The ‘Qingyan 1’ with 12 h interaction was removed from the sterile water or GL18 bacterial suspension, and the root residual liquid was absorbed using filter paper, wrapped in tin foil, and placed in a 15 mL enzyme-free centrifuge tube for quick freezing in liquid nitrogen. One treatment was repeated three times, and five plants were taken from every repeat so that nine samples were placed in an incubator with dry ice and sent to Shanghai Meiji Biomedical Technology Co., Ltd., for transcriptome sequencing. Then, the sequencing was performed on the Illumina platform.
2.6. Transcriptome Data Analysis
SeqPrep (https://github.com/jstjohn/SeqPrep) software sequencing data quality control was used. TopHat2 [23] software was used to compare the sequencing data with the oat reference genome sequence for differential expression gene analysis. DESeq2 [24] software was used to analyze the differential expression of genes or transcripts among the samples. Software Blast2go [25] and Goatools [26] were used to conduct functional annotation and enrichment analysis of Gene Ontology (GO) of the differentially expressed genes in gene sets to analyze the functions of the differentially expressed genes after the interaction between ‘Qingyan 1’ and strain GL18. Using R language on the genes concentrated the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. The metabolic pathway of the KEGG pathway, a differentially expressed gene in the samples, was analyzed to explore the response of normal growth induced by Bacillus under low-temperature stress.
2.7. Expression of Growth-Related Genes in Plants by Real-Time Quantitative PCR Analysis
To validate the RNA-Seq data, the total RNA of the oat leaves was extracted from three samples using a Plant RNA Rapid Extraction Kit (Baiaoman Technology Co., Ltd., Beijing, China). The genes PP2C2, SNRK22, BRI1, PIF4 and MPK3 were selected for PCR analysis. The primers were designed by the Primer Quest tool of IDT3 (Supplementary material 1). The PCR was performed using the following cycling conditions: 95 °C for 30 s, 40 cycles at 95 °C for 15 s and 60 °C for 30 s, followed by an extension. Each PCR analysis was repeated three times, and the relative expression of the genes was calculated using the 2 −ΔΔ Ct relative quantification method.
3. Results
3.1. Sequencing Data Statistics and Evaluation
The Illumina sequencing platform was used to complete transcriptome sequencing, and the sequences measured in nine samples were statistically and qualitatively controlled. The quality of the data after quality control was compared with the reference genome. In addition, the Q20 and Q30 quality values of the comparison results of this sequencing were evaluated for base accuracy. The clean data of all of the samples were more than 6.1 GB, and the percentage of Q30 bases was more than 94.24%, indicating a low error rate and good sequencing quality. The nine samples were compared with the reference genome sequence of ‘Qingyan 1’; the comparison rate was 77.07%–81.72% and the GC content was 52.11%–54.36% (Supplementary material 2). A total of 130,655 unigenes were detected, comprising 90,333 known genes and 40,322 new genes, and 154,833 expressed transcripts, comprising 89,759 known transcripts and 65,074 new transcripts. The functional annotations of the unigenes were compared with six databases, and final numbers of 88,240 (NR: 97.68%), 78,596 (Swiss-Prot: 87.01%), 86,686 (COG: 95.96%), 50,752 (KEGG: 56.18%), 77,578 (GO: 85.88%), and 57,952 (Pfam: 64.15%) unigenes received functional annotations (Figure 2).
3.2. Statistics of Differentially Expressed Genes
‘Qingyan 1’ seedlings interacted with GL18 bacterial suspension or sterile water for 12 h to detect differentially expressed genes. The results showed that compared with the LCK control group at 4°C, a total of 12,061 differentially expressed genes were detected in the LB interaction group, among which 4,368 genes were upregulated and 7,698 genes were downregulated. There were 25,489 differentially expressed genes in the LB interaction group compared with the CK control group at 26°C, among which 14,515 genes were upregulated and 10,974 genes were downregulated. Compared with the CK control group at 26°C, there were 28,771 differentially expressed genes in the LCK interaction group, of which 17,361 genes were upregulated and 11,410 genes were downregulated (Figure 3).
Through differential expression Venn diagram analysis, it was found that ‘Qingyan 1’ seedlings interacting with GL18 bacterial suspension or sterile water for 12 h had a total of 200 differentially expressed genes in three different treatment groups, among which 718 differentially expressed genes were in the LB interaction and LCK control groups, and 3,886 differentially expressed genes were in the LB interaction and CK control groups. There were 3,987 differentially expressed genes between the LCK treatment and CK control groups (Figure 4).
3.3. Annotation Analysis of GO Function of Differentially Expressed Genes
GO functions are divided into three functional categories, namely, biological process, molecular function, and cellular component. GO annotation analysis was performed on the differential epigenetic genes in the three treatment groups, and the results showed that the differentially expressed genes of the three different treatment groups are mainly related to metabolic process, cellular process, catalytic activity, binding, and cell part.
After 12 h of interaction, the number of differentially expressed genes related to biological process, molecular function, and cell composition between the LB interaction and LCK control groups was 3054, 2464, and 3623, respectively. Among them, the number of differentially related genes of plant growth, response to stimulation, carbon utilization, antioxidant activity, and catalytic activity was 7, 316, 1, 17, and 1059, respectively. There were 12,986, 10,709, and 13,800 differentially expressed genes related to biological process, molecular function, and cell composition, respectively, between the LB interaction and CK control groups. Among them, the number of differential genes associated with plant growth, response to stimulus, carbon utilization, nitrogen utilization, antioxidant activity, and catalytic activity was 32, 1 245, 2, 17, 99, and 4 546, respectively (Figure 5).
3.4. GO Enrichment Analysis
The GO enrichment analysis of the differentially expressed genes in the gene sets of the three treatment groups showed that the differentially expressed genes in every group were significantly enriched in the biological process after 12 h of interaction. Interacting for 12 h, the differentially expressed genes between the LB interaction and LCK control groups were significantly enriched in cell wall polysaccharide catabolic process, negative regulation of phospholipid metabolic process, negative regulation of phospholipid biosynthetic process, regulation of meristem development, thiamine biosynthetic process, plant-type cell wall biogenesis, and other metabolic processes related to plant growth and development process. The differently expressed genes of the LB interaction and CK control groups were significantly enriched in the amine catabolic process, cellular biogenic amine catabolic process, polyamine catabolic process, branched-chain amino acid catabolic process, spermidine biosynthetic process, and other processes related to plant growth, metabolism, and catalytic activity (Figure 6).
In summary, Bacillus GL18 interacts with ‘Qingyan 1’ seedlings, and differential genes are significantly enriched in plant tissue growth and development, cell wall, and organelle membrane synthesis. It is speculated that GL18 can promote cell division, elongation, and expansion by increasing the level of endogenous hormones in plants and inducing hydrolase. Differential genes were significantly enriched in the biosynthesis of thiamine and its compounds, and it was speculated that GL18 activated the key enzymes in the thiamine synthesis pathway in ‘Qingyan 1’, regulated the thiamine synthesis in plants, affected its growth and development, enhanced the response of plants to biological and abiotic stresses, and improved the tolerance of oat plants to environmental stresses. In addition, different expression genes were significantly enriched in polyamine catabolism, which is closely related to plant growth and development and environmental stress response. It is speculated that GL18 can induce polyamine catabolism, thus promoting the formation of the flower organs and roots of oats and improving the adaptability to environmental stress.
3.5. KEGG Enrichment Analysis of Differentially Expressed Genes
To further explore the molecular mechanism of resistance to low-temperature stress induced by strain GL18, differential genes of three treatment groups were enriched in the KEGG database. The results showed that interacting for 12 h, the differentially expressed genes of the LB interaction and LCK control groups were significantly enriched in cyanoamino acid metabolism, phenylpropanoid biosynthesis, starch and sucrose metabolism, thiamine metabolism, porphyrin and chlorophyll metabolism, protein export, plant hormone signal transduction, beta-alanine metabolism, fatty acid degradation, alpha-linolenic acid metabolism, and other processes. The differentially expressed genes of arginine and proline metabolism, beta-alanine metabolism, glycolysis/gluconeogenesis, pentose and glucuronate interconversions, galactose metabolism, TCA cycle, as well as glycine, serine, and threonine metabolism, and other processes were enriched (Figure 7).
KEGG enrichment analysis showed that ‘Qingyan 1’ seedlings interacted with Bacillus GL18 under low-temperature stress, and the metabolic pathway of ‘Qingyan 1’ leaf tissues changed significantly. The differentially expressed genes are mainly concentrated in various amino acid metabolism, lipid metabolism, phenylpropane biosynthesis, and other related pathways. It is speculated that GL18 can promote the synthesis and metabolism of protein and lipid in ‘Qingyan 1’, promote its growth and development, and enhance its stress resistance. In addition, the genes were significantly enriched in the metabolic pathways closely related to plant growth and development, such as porphyrin and chlorophyll metabolism and starch and sucrose metabolism. It was speculated that GL18 promoted the photosynthesis of ‘Qingyan 1’; accumulated carbohydrates, organic acids, and other photosynthetic products; enhanced the resistance of ‘Qingyan 1’ to low temperature; and promoted its normal growth.
3.6. Analysis of GL18-Related Signal Transduction Pathways in ‘Qingyan 1’ Response
The interaction between Bacillus GL18 and ‘Qingyan 1’ under low-temperature stress revealed several plants hormone-mediated signaling and metabolic pathways, and several differentially expressed genes related to growth promotion and stress resistance were significantly upregulated.
3.6.1. Tryptophan Metabolism
Tryptophan is an essential amino acid for plant protein synthesis and an important precursor for auxin biosynthesis in plants [27,28]. Under low-temperature stress, after the interaction between ‘Qingyan 1’ and Bacillus GL18, GL18 induced the plants to participate in the tryptophan-dependent indole-3-acetic acid (IAA) synthesis pathway. The auxin-related factor ARF activated AUX/IAA, GH3, and SAUR proteins, and the protein-coding gene AUXI, which promotes cell enlargement and plant growth indirectly, was upregulated. GL18 can promote ‘Qingyan 1’ cell division, root elongation, and fruit development by inducing IAA formation.
3.6.2. Carotenoid Biosynthesis
Carotenoids is a general term for a class of important natural pigments, which play an important role in plant growth and development, photosynthesis, and biosynthesis of plant hormones and volatile substances [29,30]. Under low-temperature stress, after the interaction between Bacillus GL18 and A. sativa, GL18 induced carotenoid biosynthesis in the plants and indirectly activated abscisic acid receptor PYR/PYL, thereby inhibiting the upregulated expression of the protein phosphatase-coding genes PP2C and SNRK2, and finally led to the upregulated expression of transcription regulatory factor ABF. It then reduced seed stomatal closure and promoted seed break from dormancy, thus promoting germination. It was speculated that strain GL18 induced the carotenoid metabolic pathway in ‘Qingyan 1’ under low-temperature stress, thus promoting the conversion and absorption of light energy, promoting photosynthesis, and increasing biomass.
3.6.3. Brassinosteroid Biosynthesis
Brassinosteroid is a new type of plant endogenous hormone, which can regulate various processes, such as cell elongation, cell division, photomorphogenesis, and abiotic and biological stress responses during plant growth and development [31,32]. Under low-temperature stress, after the interaction between Bacillus GL18 and ‘Qingyan 1’, GL18 induced the biosynthesis of brassinosteroid in the seedlings, the upregulated expression of brassinosteroid insensitive protein kinase BRI1, and the activation of signal kinase gene BSK after phosphorylation, which inhibited the phosphorylation of insensitive protein BIN2. Eventually, TCH4 and CYCD3 genes were expressed, thereby indirectly promoting plant cell division and elongation. It was speculated that strain GL18 promoted brassinosteroid biosynthesis in ‘Qingyan 1’ under low-temperature stress, accelerated cell division and elongation, regulated photosynthesis, respiration, and transpiration of plants, enhanced cold tolerance, and maintained normal growth at low temperatures.
3.6.4. Diterpenoid Biosynthesis
Diterpenoid compounds have been found to exist in organisms such as higher and lower plants, fungi, and bacteria, and the key enzyme of their biosynthesis is diterpene synthase [33]. Under low-temperature stress, after the interaction between Bacillus GL18 and ‘Qingyan 1’, GL18 induced diterpenoid biosynthesis in the plants, indirectly activated the receptor of gibberellins in plant somatic cells GIDI, directly activated the downregulated expression of the gene DELLA, and reduced its ubiquitination of the protein-coding gene GID2. In addition, the expression of PIF4, the encoding gene of photochromic interaction factor TF, was upregulated, and the two had a reciprocal inhibitory effect, thereby indirectly promoting plant seed germination and stem elongation. It was speculated that the stress strain, GL18, stimulated the synthesis of signal transducers, such as gibberellin, in ‘Qingyan 1’ under low temperatures, and caused the interaction of terpenoids with proline and betaine to promote plant growth and resist stress.
3.6.5. Abscisic Acid-Mediated Stress Resistance Signal Transduction
Abscisic acid, as a plant stress response hormone, is not only a signal of plant response to abiotic stress but also an important factor causing adaptive regulatory response and gene expression in plants and is involved in response regulation under abiotic stress [34]. After the interaction between Bacillus GL18 and ‘Qingyan 1’, GL18 induced the expression of multiple differential genes in the abiotic stress response pathway in the seedlings. When plants are subjected to abiotic stress, such as low temperature, salt, and drought, abscisic acid-mediated defense signal expression in ‘Qingyan 1’ causes the plant’s defense response, namely the receptor PYR/PYL, on the cell membrane to receive the signal and transduce it into the cell, thus inhibiting the upregulated expression of protein phosphatase coding gene PP2C. This reduces its dephosphorylation results in the upregulation of the serine/threonine protein kinase gene SNRK2, thereby inducing plant responses to abiotic stress and enhancing resistance. It was speculated that strain GL18 promoted ABA biosynthesis in ‘Qingyan 1’ under low-temperature stress, triggered abiotic stress response-related signaling pathway mediated by ABA, and enhanced stress resistance by inducing the transcription and expression of stress-related genes.
3.6.6. Ethylene-Mediated Stress Resistance Signal Transduction
Ethylene, as a gas hormone, not only plays an important role in physiological processes, such as seed germination, organ aging, and fruit ripening and shedding, but also plays a role in regulating plant response to abiotic stresses, such as low temperature and waterlogging [35]. Under low-temperature stress, after the interaction between Bacillus GL18 and ‘Qingyan 1’, the ethylene molecule receives the signal in the plant and causes the corresponding defense response; that is, MPK3/6 protein is directly activated after the phosphorylation of mitogen-activated protein kinase MKK9 in the ethylene-mediated MAPK signal transduction pathway. Under the influence of strain GL18, the expression of the MPK3 coding gene was upregulated, while the expression of the EBF1_2 binding protein coding gene was downregulated, thereby inducing plant defense response to abiotic stress. It is speculated that plant cells produce cold damage signals in low-temperature environments, accelerate the conversion of methionine to S-adenylyl methionine in the process of ethylene biosynthesis in plants, and then form the key enzyme ACC oxidase required for ethylene degradation, thus enhancing plant stress resistance and maintaining normal growth and development.
In summary, under low-temperature stress, Bacillus GL18 induced the synthesis of growth-promoting related plant hormones and the generation of stress-resistance related compounds in ‘Qingyan 1’ seedlings and the corresponding regulatory expression of functional genes in leaf tissues of the plants in the LB interaction group. AUX/IAA proteins participate in auxin signaling by interacting with ARF protein as a transcription suppressor, thus promoting plant root development, bud growth, and fruit ripening. PP2C, SNRK2, ABF, and BRI1 were significantly upregulated in the LB interaction group, and these proteins are associated with plant photosynthesis, antioxidant, and abiotic stress tolerance. It was speculated that the expression of these proteins regulated the responses of related genes and many transcription factors, thereby regulating plant growth and development, photomorphogenesis, and other metabolic processes. The PIF4 gene is related to seed germination, morphogenesis, chloroplast differentiation, and flowering and regulates plant growth by promoting plant hormone biosynthesis and signal transduction. The upregulated expression of PP2C, SNRK2, and MPK3 genes in ABA and the ethylene-mediated stress signal transduction pathway suggests that the interaction of these two hormones may enhance the adaptability of ‘Qingyan 1’ to environmental stress (Figure 8).
3.7. PCR Validation of GL18 on the Expression of Growth-Related Genes
We analyzed the expression patterns of the genes PP2C2, SNRK22, BRI1, PIF4 and MPK3 of strain GL18 under low-temperature were detected by real-time fluorescence quantitative PCR. The results revealed that at 4 ℃ stress, GL18 caused the differential expression of a set of genes involved in oat cold tolerance. It including seedlings inoculated with sterile water and grown in cold or room-temperature; seedlings inoculated with GL18 and grown in cold. It showed that the expression trends of these genes were generally consistent with the RNA-Seq results, indicating that the RNA-Seq data were authentic and reliable (Figure 9).
4. Discussion
In the process of growth and development, plants and plant growth-promoting rhizobacteria form symbiotic relationships to alleviate abiotic stress, while rhizosphere bacteria exert various positive effects on host plants through various mechanisms, thereby reducing the harm of stress and promoting plant growth. Furthermore, some plant root secretions act as microbial attractants and maintain the function and structure of microbial communities near the roots [36,37]. We found that B. amyloliquefaciens GL18 suspension had positive effects on endogenous hormone levels, antioxidant enzyme activity, reactive oxygen species (ROS) accumulation, membrane peroxidation degree, and osmotic substance content in ‘Qingyan 1’ seedlings under low-temperature stress of 4°C19. In addition, there were functional genes related to promoting biomass synthesis and stress tolerance in the GL18 genome (accession number: CP096033). To explore the molecular mechanism of Bacillus GL18 in promoting ‘Qingyan 1’ growth under low-temperature stress, we used transcriptome sequencing technology to analyze the response of the seedling leaves to GL18.
It has been reported that Bacillus can promote plant growth by producing plant growth hormones to supply to the host or regulate plant growth and development by regulating the expression of growth hormone-related genes in the host plant and inducing plant resistance [38]. The AUXI gene is a key factor regulating gene expression in plant auxin signal transduction [39]. Currently, 29 members of the Aux/IAA gene family have been isolated from Arabidopsis thaliana [40]. In addition, Aux/IAA family members are present in large numbers in a variety of plants, including Cucumis sativus [41], Oryza sativa [42], and Solanum lycopersicum [43]. In this study, after the interaction between strain GL18 and ‘Qingyan 1’, the Aux/IAA coding gene expression was upregulated in the tryptophan metabolism pathway. The Aux/IAA coding protein can bind with the auxin response factor (ARF), thereby regulating the expression of the plant auxin coding gene and then promoting the growth and development of plants. In plants, carotenoids are precursors of ABA synthesis, and their accumulation in response to osmosis, high salt, dehydration, and cold stress is consistent with changes in plant ABA levels [44,45]. Strain GL18 activates carotenoid metabolic pathways in ‘Qingyan 1’ leaf cells under low-temperature stress, promoting the absorption, transfer, and transformation of light energy in plants by synthesizing light and pigment. Moreover, it is speculated that the synthesis of ABA in plants may increase the soluble sugar content of plants, protect the plasma membrane from cold damage by regulating osmosis, and also provide energy for the synthesis of other organic matter. In the brassinosteroid biosynthesis pathway, its insensitive protein kinase gene BRI1 is significantly upregulated. The BRI protein is a receptor kinase localized on the cell surface in the brassinosteroid signaling pathway and can be expressed in most plant tissues or organs [46]. He et al. [47] fused the extracellular leucine repeat sequence and transmembrane domain of A. thaliana receptor kinase BRI1 protein with the serine/threonine kinase domain of O. sativa resistance receptor XA21 and found that the chimeric receptor participated in the brassinosteroid synthesis pathway in the plants. It was speculated that GL18 stimulated the extracellular domain of BRI1 to sense brassinosteroid, thereby regulating plant growth, development, and tolerance to low temperatures. The phytochrome-interacting factor PIF4 gene is a regulatory center in the process of growth and directly regulates the growth and development of plants, but its activity is limited to a short time with light. Meanwhile, it is involved in gibberellin biosynthesis and signal transduction [48]. In this study, the upregulated expression of gene PIF4 may regulate plant growth through photosynthesis and transpiration and may also change the sensitivity of plant hormones, such as gibberellin, to regulate the growth of plant hypocotyl, petiole, and leaf. It was found that rhizosphere bacteria 5C-2 containing ACC deaminase stimulated leaf growth and the flowering of A. thaliana through the ethylene signal transduction pathway [49]. It was also reported that inoculation of the rhizosphere bacteria STM196 promoted root hair elongation of A. thaliana seedlings, and the ethylene signal pathway was activated, suggesting that local ethylene release could help the strain induce plant growth [50]. Strain GL18 induced ethylene-mediated signal transduction in ‘Qingyan 1’, during which ethylene precursor ACC was degraded by ACC deaminase, thereby reducing ethylene level during plant growth and promoting plant growth. Meanwhile, GL18 synthesized and secreted IAA, which was absorbed by plants to form endogenous auxin, thus stimulating plant cell division and elongation. IAA also stimulates the activity of ACC synthase.
Transcriptome sequencing showed that differentially expressed genes were significantly enriched in photosynthetic pathways such as chlorophyll biosynthesis and starch and sucrose metabolism. Therefore, it was speculated that strain GL18 could promote photosynthesis in ‘Qingyan 1’ leaves under low-temperature stress to achieve light energy conversion and convert inorganic matter into organic matter, thus maintaining plant growth and development. This is consistent with the reported conclusion that Bacillus can promote the changes in stomatal conductance, net photosynthetic rate, and transpiration rate of beet leaves [51]. Differential genes are significantly enriched in the biosynthesis process of thiamine, polyamines, and phenylpropane, which have growth promotion and anti-stress effects. Among them, studies have shown that plants can increase the expression level of the key enzyme of thiamine synthesis under stress, increase the content of thiamine in cells, enhance the antioxidant capacity of crops, and thus alleviate the harm of abiotic stress on plants [52]. Strain GL18 was speculated to possibly promote the metabolism of thiamine in ‘Qingyan 1’, maintain the REDOX state balance of plant cells, provide a possibility for the rapid release of ROS when cells are stimulated by foreign stimuli, and then induce downstream signaling molecules to make ‘Qingyan 1’ obtain resistance. Under stress, the putrescine, spermidine, and spermidine contents of plants will change. Studies have shown that polyamines can inhibit ethylene biosynthesis in plants, thereby regulating the defense process, and polyamines can induce proline accumulation and contribute to osmotic regulation [53,54]. It is speculated that GL18 promoted polyamine biosynthesis in ‘Qingyan 1’ and regulated the expression of corresponding genes, thus promoting plant growth. L-phenylalanine ammonia-lyase is a key enzyme involved in lignin synthesis, which plays an important role in preventing disease, maintaining plant standing, and transporting water and nutrients [55]. It is speculated that strain GL18 promoted the expression of the L-phenylalanine ammonia-lyase encoding gene in ‘Qingyan 1’ leaves under low-temperature stress and induced the production of flavonoids to alleviate oxidative damage.
5. Conclusions
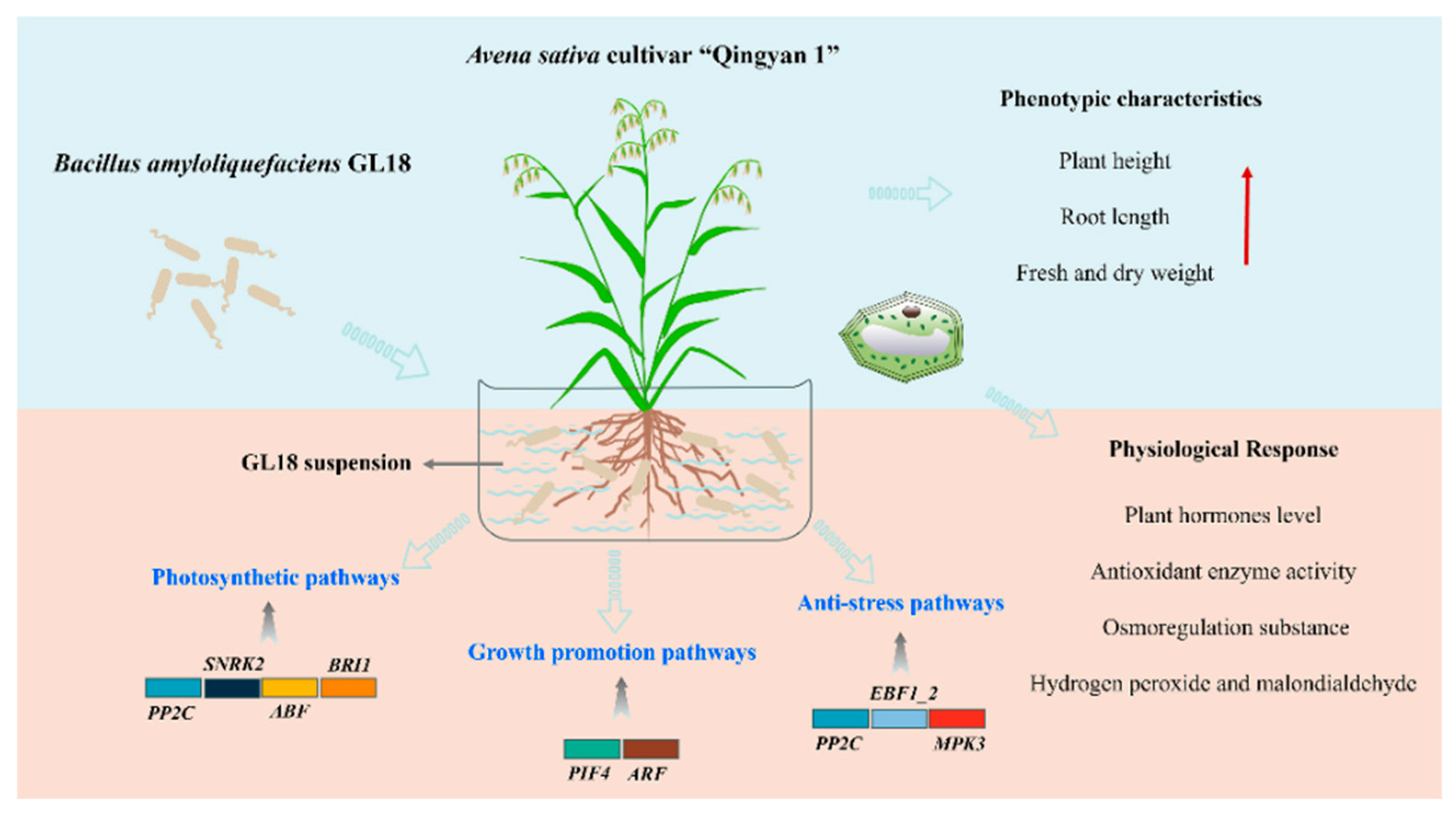
In the early stage, we found that strain GL18 is a plant rhizosphere growth promoting bacterium with good biocontrol characteristics, which has strong stress resistance abilities, especially low-temperature. Under low temperature stress, we found that GL18 has positive regulation on ‘Qingyan 1’ antioxidant enzyme activity, plant hormone level, osmoregulation substances and membrane peroxidation. Therefore, we further explore how GL18 plays its growth promoting effect on plant under low-temperature stress through transcriptome sequencing. Fortunately, it is found that differentially expressed genes regulate plant growth through photosynthesis, transpiration and plant hormones, which is a good verification for our previous experiments (Figure 10). Furthermore, we will use metabolomics to explore the induction of differential metabolites by Bacillus GL18 in ‘Qingyan 1’, it also includes the content of quick-acting phosphorus, quick-acting potassium, ammonium nitrogen, and nitrate nitrogen in the soil, thus improving soil fertility.
6. Patents
The invention patent named ‘1 Bacillus amyloliquefaciens strain for promoting the growth of oat (Avena sativa) and its application’ (the patent number: ZL 2022 1 0002973.3).
Author Contributions
Methodology, L.C., Y.Z. and Y.X.; software, T.W. and J.L.; formal analysis, L.C. and T.W.; data curation, F.C., L.W. and Y.G.; writing—original draft preparation, L.C. and T.W.; writing—review and editing, Y.Z. and Y.X.; visualization, Y.Z. and Y.X.; funding acquisition, Y.Z. and Y.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by China National Key Research and Development Program (2021YFC3201604); Independent Project in the direction of Ecosystem Succession and Management of a World-class Discipline of Ecology (2023-ZZ-01); National Natural Science Foundation of China (32160030); Applied Basic Research Project of Qinghai Provincial Department of Science and Technology (2023-ZJ-709).
Data Availability Statement
We uploaded the raw data to NCBI, and the accession number of these SRA data is PRJNA1036295.
Acknowledgments
Authors are grateful to the authorities of respective departments for support in doing this research. We thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pathak, M.R.; Teixeira da Silva, J.A.; Wani, S.H. Polyamines in response to abiotic stress tolerance through transgenic approaches. GM crops & food 2014, 5(2): 87-96. [CrossRef]
- Gong, Z.; Xiong, L., Shi, H.; et al. Plant abiotic stress response and nutrient use efficiency. Sci China Life Sci. 2020, 63(5):635-674. [CrossRef]
- Zhou, L.Y.; Li, Y.Y.; Wang, W.N.; et al. Research progress in the metabolomics for plants response to temperature stress. Journal of Shanxi Agricultural Sciences 2017,45(02):317-320.
- Wang, H.; Zhao, J.; Lai, M.; et al. Differential gene expression analysis and physiological response characteristics of passion fruit (Passiflora edulis) buds under high-temperature stress. Peer J 2023, 11: e14839. [CrossRef]
- Akter, N.; Rafiqul Islam, M. Heat stress effects and management in wheat. A review. Agronomy for sustainable development 2017, 37: 1-17. [CrossRef]
- Megha, S.; Basu, U.; Kav, N.N.V. Regulation of low temperature stress in plants by microRNAs. Plant, Cell & Environment 2018, 41(1): 1-15. [CrossRef]
- Zhang, D.W.; Wang, S.; Gao, Y.; et al. Role of microRNAs in response to low temperature stress in plants. Plant Physiol. J 2019, 55: 117-124.
- He, J.S.; Liu, Z.P.; Yao, T.; et al. Analysis of the main constraints and restoration techniques of degraded grassland on the Tibetan Plateau. Science & Technology Review 2020,38(17):66-80.
- Sun, Y.; Guo, R.; Liu, H.C.; et al. Evolutionary game of overgrazing and grassland degradation governance in Qinghai-Tibet Plateau. Science & Technology for Development 2022,18(05):665-673.
- Liang, G.L.; Zhou, Q.P.; Geng, F.; et al. Karyotype analysis of Avena sativa L. ‘Qingyan No.1’. Acta Agrestia Sinica 2016,24(02):389-392.
- Liu, K.Q.; Liu, W.H.; Jia, Z.F.; et al. Effects of drought stress on yield and dry matter accumulation and distribution of Avena sativa cv. Qingyan No.1. Acta Prataculturae Sinica 2021,30(03):177-188.
- Chen, G.; Zhang, Z.H.; Hu, Z.Z. Herbage yield dynamics and solar energy conversion of annual cultivated grassland in Xining city. Acta Prataculturae Sinica 2003, (01):69-73.
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: an overview. Archives of biochemistry and biophysics 2005, 444(2): 139-158.
- Ongena, M.; Jacques, P. Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16(3):115-25. [CrossRef]
- Goswami, M.; Suresh, D. Plant growth-promoting rhizobacteria—alleviators of abiotic stresses in soil: a review. Pedosphere 2020, 30(1): 40-61.
- Rosier, A.; Medeiros, F.H.V.; Bais, H.P.; et al. Defining plant growth promoting rhizobacteria molecular and biochemical networks in beneficial plant-microbe interactions. Plant and Soil 2018, 428(1-2), 35–55. [CrossRef]
- Zubair, M.; Hanif, A.; Farzand, A.; et al. Genetic screening and expression analysis of psychrophilic Bacillus spp. reveal their potential to alleviate cold stress and modulate phytohormones in Wheat. Microorganisms 2019, 7(9):337. [CrossRef]
- Zubair, M.; Farzand, A.; Mumtaz, F.; et al. Novel genetic dysregulations and oxidative damage in Fusarium graminearum induced by plant defense eliciting psychrophilic Bacillus atrophaeus TS1. Int J Mol Sci. 2021, 22(22):12094. [CrossRef]
- Chen, L.; Xie, Y.L.; Wu, X.H.; et al. Physiological response of Avena sativa to low-temperature stress is promoted by Bacillus amyloliquefaciens GL18 and its functional genes. Russian Journal of Plant Physiology 2022, 69:161.
- Xie, Y.L.; Ma, L.Z.; Xu, Z.W.; et al. Molecular identification of Bacillus strains isolated from extreme dry-sand environment in Qinghai Chaidamu region and its lipopeptide compound analysis. Microbiology China 2012,39(08):1079-1086.
- Yang, X.; Xie, Y.L.; Chen, L.; et al. Biological activity of grass growth-promoting and genome analysis of Bacillus amyloliquefaciens DGL1 isolated from the rhizosphere of Nitraria tangutorum of sand soil in Qinghai Province. Acta Agrestia Sinica 2021, 29(8): 1637-1648.
- Ayaz, M.; Ali, Q.; Farzand, A.; et al. Nematicidal volatiles from Bacillus atrophaeus GBSC56 promote growth and stimulate induced systemic resistance in tomato against meloidogyne incognita. Int J Mol Sci. 2021, 22(9):5049. [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome biology 2013, 14(4): R36. [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome biology 2014, 15(12): 550.
- Conesa, A.; Götz, S.; García-Gómez, J.M.; et al. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21(18): 3674-3676. [CrossRef]
- Tang, H.; Klopfenstein, D.; Pedersen, B.; et al. GOATOOLS: tools for gene ontology. Zenodo., 2015.
- Wang, J.L.; Liu, D.C.; Guo, X.L.; et al. Research advances in auxin biosynthesis. Chinese Bulletin of Botany 2012,47(03):292-301.
- Yue, J.; Hu, X.; Huang, J. Origin of plant auxin biosynthesis. Trends Plant Sci. 2014, 19(12):764-70.
- Tian, Q.Y.; Yue, Y.Z.; Shen, H.M.; et al. Research progress in the regulation of carotenoid metabolism in plant ornamental organ. Biotechnology Bulletin 2022,38(12):35-46.
- Auldridge, M.E.; McCarty, D.R.; Klee, H.J. Plant carotenoid cleavage oxygenases and their apocarotenoid products. Current Opinion in Plant Biology 2006, 9(3), 315-321. [CrossRef]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; et al. Brassinosteroids: multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32(2):295-318. [CrossRef]
- Abe, H.; Marumo, S. Advance of new plant hormone brassinosteroid. ChemInform 2002,33(42). [CrossRef]
- Li, X.; Chen, M.; Chai, T.Y.; et al. Advances in structure-function relation of plant type Ⅲ polyketide synthases by site-directed mutagenesis. Chinese journal of biotechnology 2018,34(4).
- Thompson, A.J.; Jackson, A.C.; Symonds, R.C.; et al. Ectopic expression of a tomato 9-cis-epoxycarotenoid dioxygenase gene causes over-production of abscisic acid. The Plant Journal 2000, 23(3): 363-374.
- Pierik, R.; Tholen, D.; Poorter, H.; et al. The Janus face of ethylene: growth inhibition and stimulation. Trends Plant Sci. 2006, 11(4):176-83. [CrossRef]
- Gouda, S.; Kerry, R.G.; Das, G.; et al. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiological research 2018, 206: 131-140. [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; et al. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Frontiers in plant science 2017, 8: 537. [CrossRef]
- Luo, J.; Zhou, J.J.; Zhang, J.Z. Aux/IAA gene family in plants: molecular structure, regulation, and function. Int J Mol Sci. 2018, 19(1):259.
- Semeradova, H.; Montesinos, J.C.; Benkova, E. All roads lead to Auxin: post-translational regulation of Auxin transport by multiple hormonal pathways. Plant Commun. 2020, 22;1(3):100048. [CrossRef]
- Overvoorde, P.J.; Okushima, Y.; Alonso, J.M.; et al. Functional genomic analysis of the Auxin/Indole-3-Acetic acid gene family members in Arabidopsis thaliana. Plant Cell 2005, 17(12):3282-300.
- Gan, D.; Zhuang, D.; Ding, F.; et al. Identification and expression analysis of primary auxin-responsive Aux/IAA gene family in cucumber (Cucumis sativus). J Genet. 2013, 92(3):513-21. [CrossRef]
- Jain, M.; Kaur, N.; Garg, R.; et al. Structure and expression analysis of early auxin-responsive Aux/IAA gene family in rice (Oryza sativa). Funct Integr Genomics. 2006, 6(1):47-59. [CrossRef]
- Audran-Delalande, C.; Bassa, C.; Mila, I.; et al. Genome-wide identification, functional analysis and expression profiling of the Aux/IAA gene family in tomato. Plant Cell Physiol. 2012, 53(4):659-72. [CrossRef]
- Du, H.; Wu, N.; Chang, Y.; et al. Carotenoid deficiency impairs ABA and IAA biosynthesis and differentially affects drought and cold tolerance in rice. Plant Mol Biol. 2013, 83(4-5):475-88. [CrossRef]
- Liu, X.; Hu, Q.; Yan, J.; et al. ζ-Carotene isomerase suppresses tillering in rice through the coordinated biosynthesis of strigolactone and abscisic acid. Mol Plant. 2020, 13(12):1784-1801. [CrossRef]
- Lozano-Elena, F.; Caño-Delgado, A.I. Emerging roles of vascular brassinosteroid receptors of the BRI1-like family. Curr Opin Plant Biol. 2019, 51:105-113. [CrossRef]
- He, Z.; Wang, Z.Y.; Li, J.; et al. Perception of brassinosteroids by the extracellular domain of the receptor kinase BRI1. Science 2000, 288(5475): 2360-2363. [CrossRef]
- Filo, J.; Wu, A.; Eliason, E.; et al. Gibberellin driven growth in elf3 mutants requires PIF4 and PIF5. Plant Signal Behav. 2015, 10(3): e992707.
- Chen, L.; Dodd, I.C.; Theobald, J.C.; et al. The rhizobacterium Variovorax paradoxus 5C-2, containing ACC deaminase, promotes growth and development of Arabidopsis thaliana via an ethylene-dependent pathway. Journal of experimental botany 2013, 64(6): 1565-1573. [CrossRef]
- Galland, M.; Gamet, L.; Varoquaux, F.; et al. The ethylene pathway contributes to root hair elongation induced by the beneficial bacteria Phyllobacterium brassicacearum STM196. Plant Science 2012, 190: 74-81. [CrossRef]
- Shi, Y.W.; Lou, K.; Li, C.; et al. Effects of endogenous Bacillus polymyxoides S-7 on photosynthesis, yield and quality of sugar beet. Chinese Journal of Applied Ecology 2009, 20(03):597-602.
- Ferreira, S.; Hjernø, K.; Larsen, M.; et al. Proteome profiling of Populus euphratica Oliv. upon heat stress. Ann Bot. 2006, 98(2):361-77. [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; et al. Polyamine function in plants: metabolism, regulation on development, and roles in abiotic stress responses. Frontiers in plant science 2019, 9: 1945. [CrossRef]
- Pál, M.; Tajti, J.; Szalai, G.; et al. Interaction of polyamines, abscisic acid and proline under osmotic stress in the leaves of wheat plants. Sci Rep. 2018, 8: 12839. [CrossRef]
- Dong, C.J.; Li, L.; Cao, N.; et al. Roles of phenylalanine ammonia-lyase in low temperature tolerance in cucumber seedlings. Chinese Journal of Applied Ecology 2015,26(07):2041-2049.
Figure 1.
‘Qingyan 1’ root irrigated in GL18 bacterial suspension.

Figure 2.
Gene function annotation statistics.
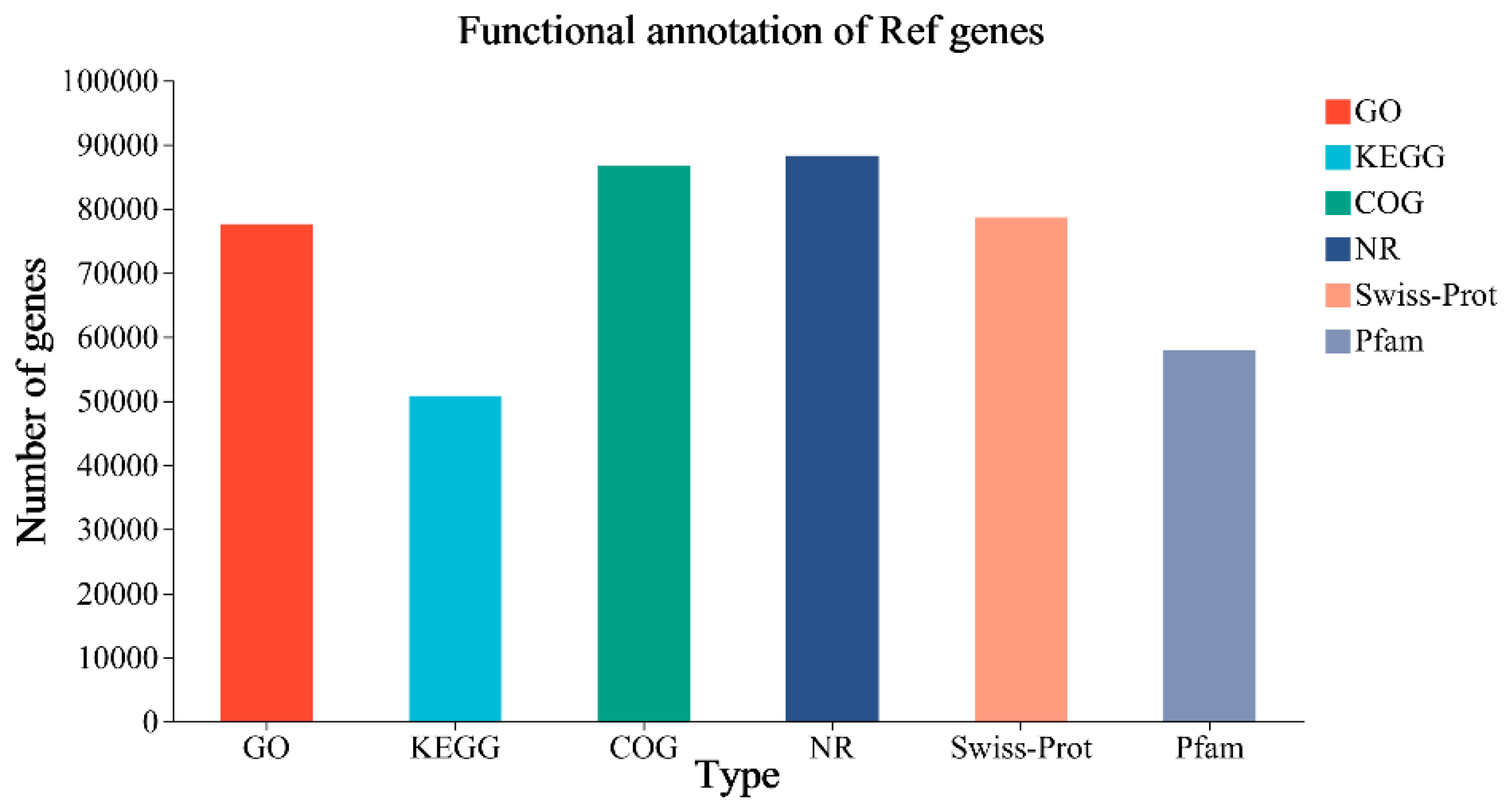
Figure 3.
Volcano map of differentially expressed genes. The horizontal coordinates represent the fold difference in gene expression, the vertical coordinates represent the statistical test value of the difference in gene expression change, gray circles, red circles, and green circles represented genes with no significant changes, upregulated genes, and downregulated genes, respectively.
Figure 3.
Volcano map of differentially expressed genes. The horizontal coordinates represent the fold difference in gene expression, the vertical coordinates represent the statistical test value of the difference in gene expression change, gray circles, red circles, and green circles represented genes with no significant changes, upregulated genes, and downregulated genes, respectively.
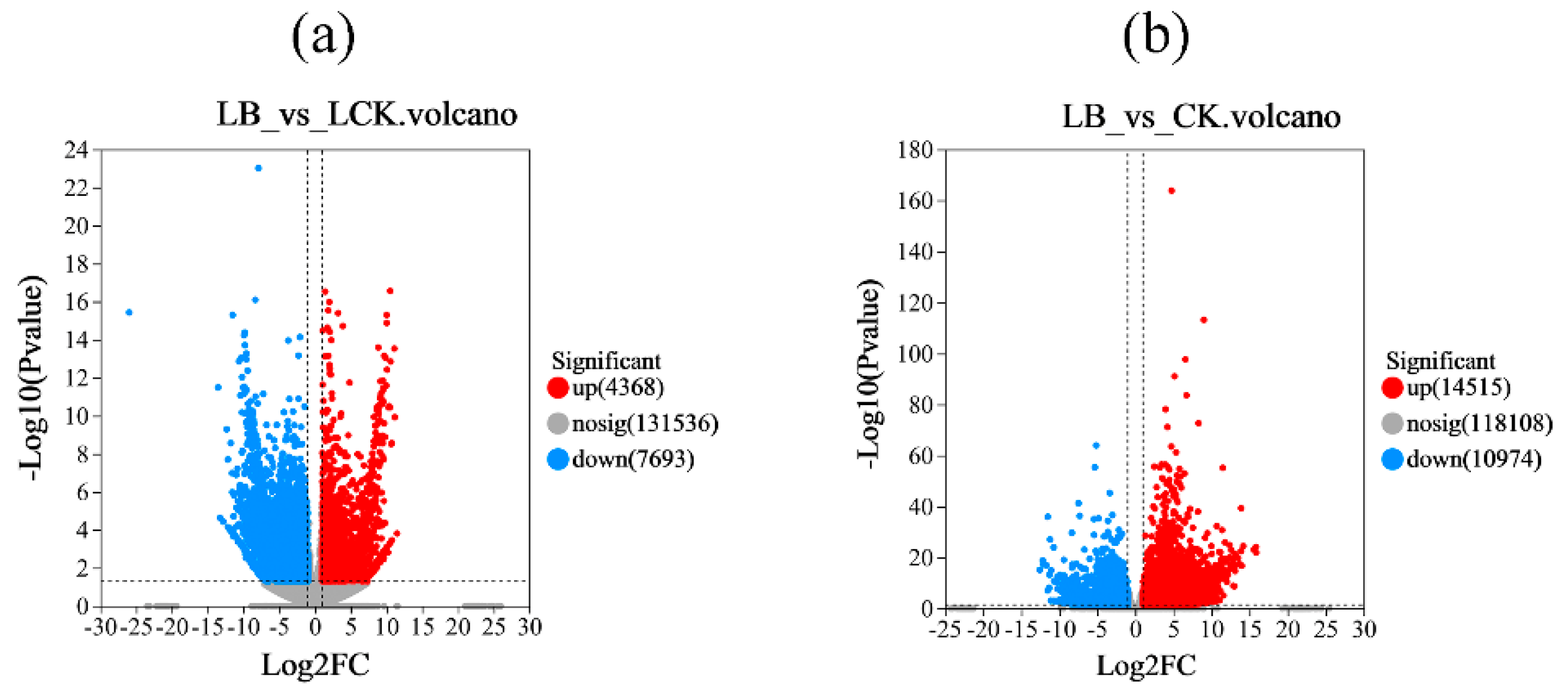
Figure 4.
Venn diagram of differently expressed genes.
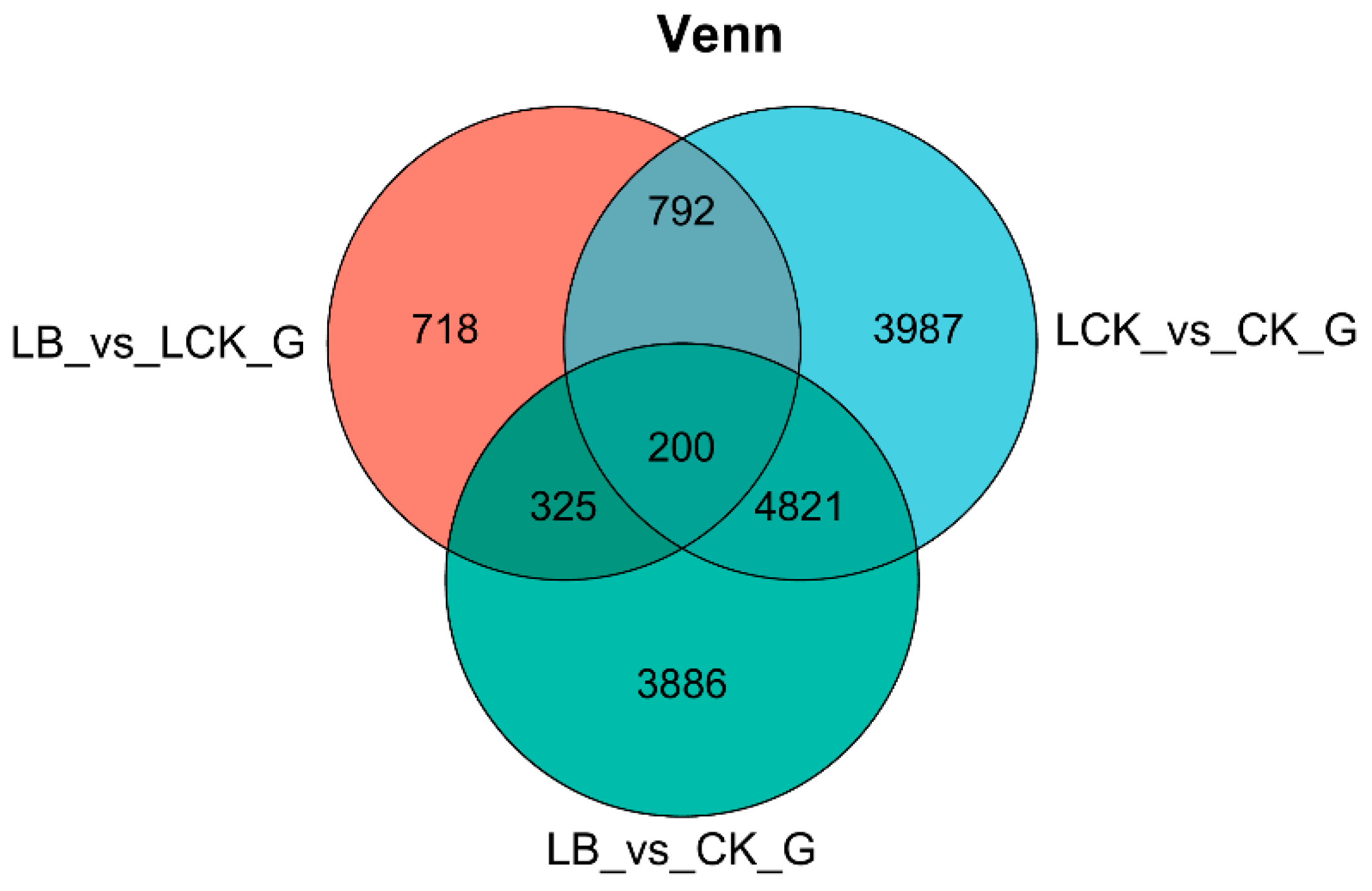
Figure 5.
Classification of GO function of differently expressed genes.
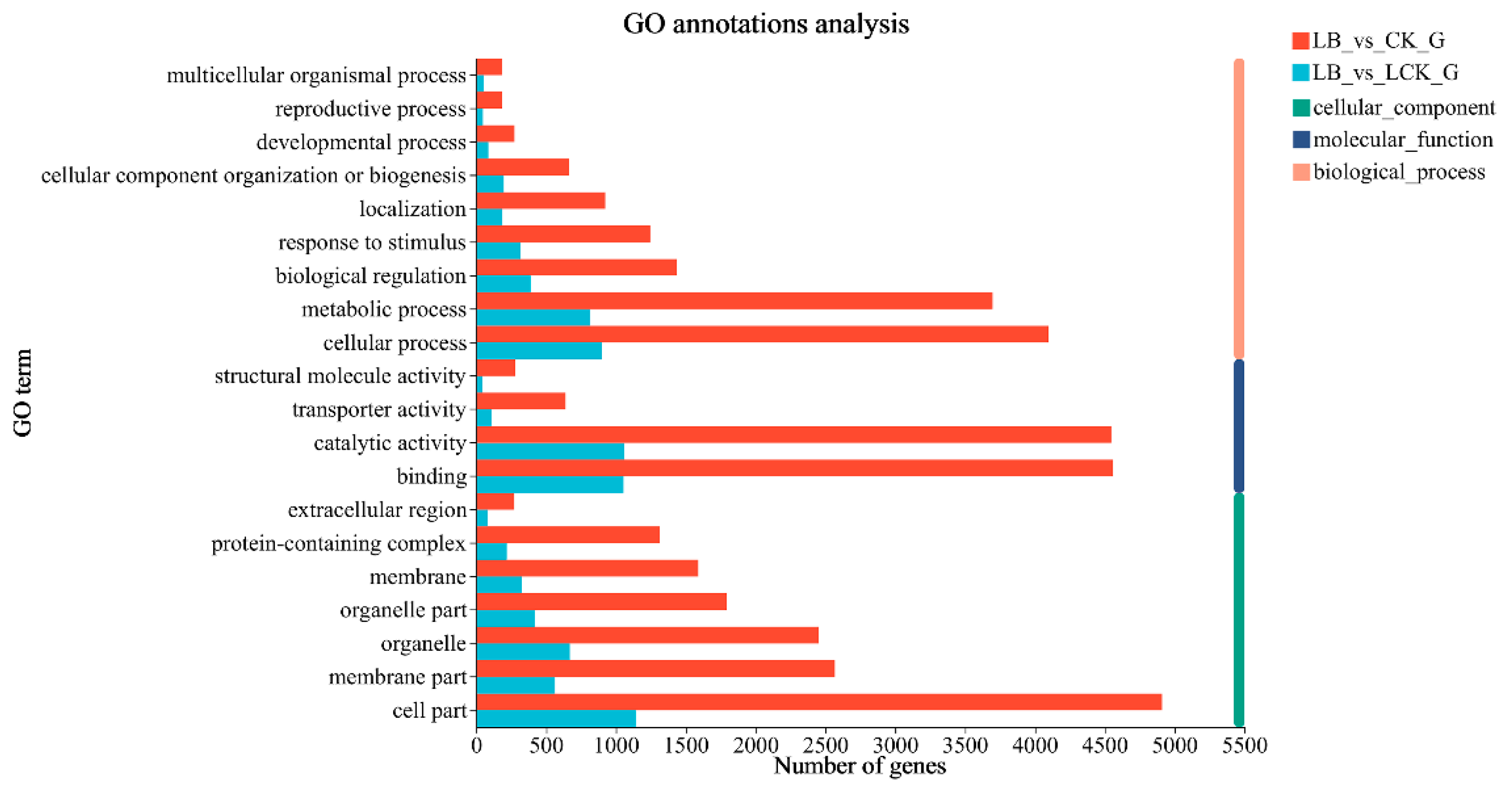
Figure 6.
Differential expressed gene GO function enrichment result. Rich factor, that is, the ratio of genes enriched in GO term to annotated genes. The greater the Rich factor, the greater the degree of enrichment, and the size of the dot represents the number of genes in this GO Term.
Figure 6.
Differential expressed gene GO function enrichment result. Rich factor, that is, the ratio of genes enriched in GO term to annotated genes. The greater the Rich factor, the greater the degree of enrichment, and the size of the dot represents the number of genes in this GO Term.
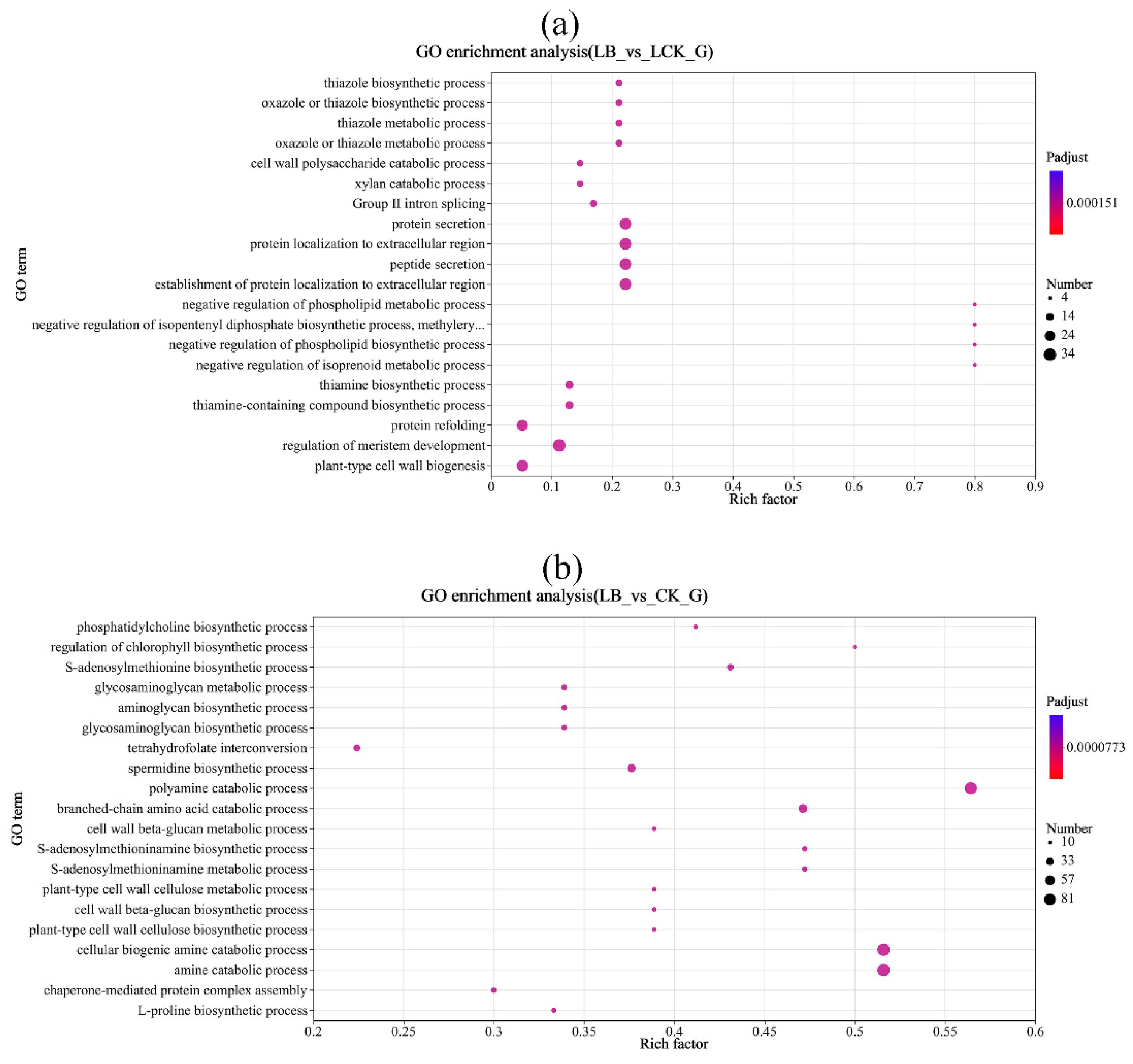
Figure 7.
Differential expressed gene KEGG pathway enrichment result. The greater the -log10 value, the more significant the enrichment of the KEGG pathway.
Figure 7.
Differential expressed gene KEGG pathway enrichment result. The greater the -log10 value, the more significant the enrichment of the KEGG pathway.
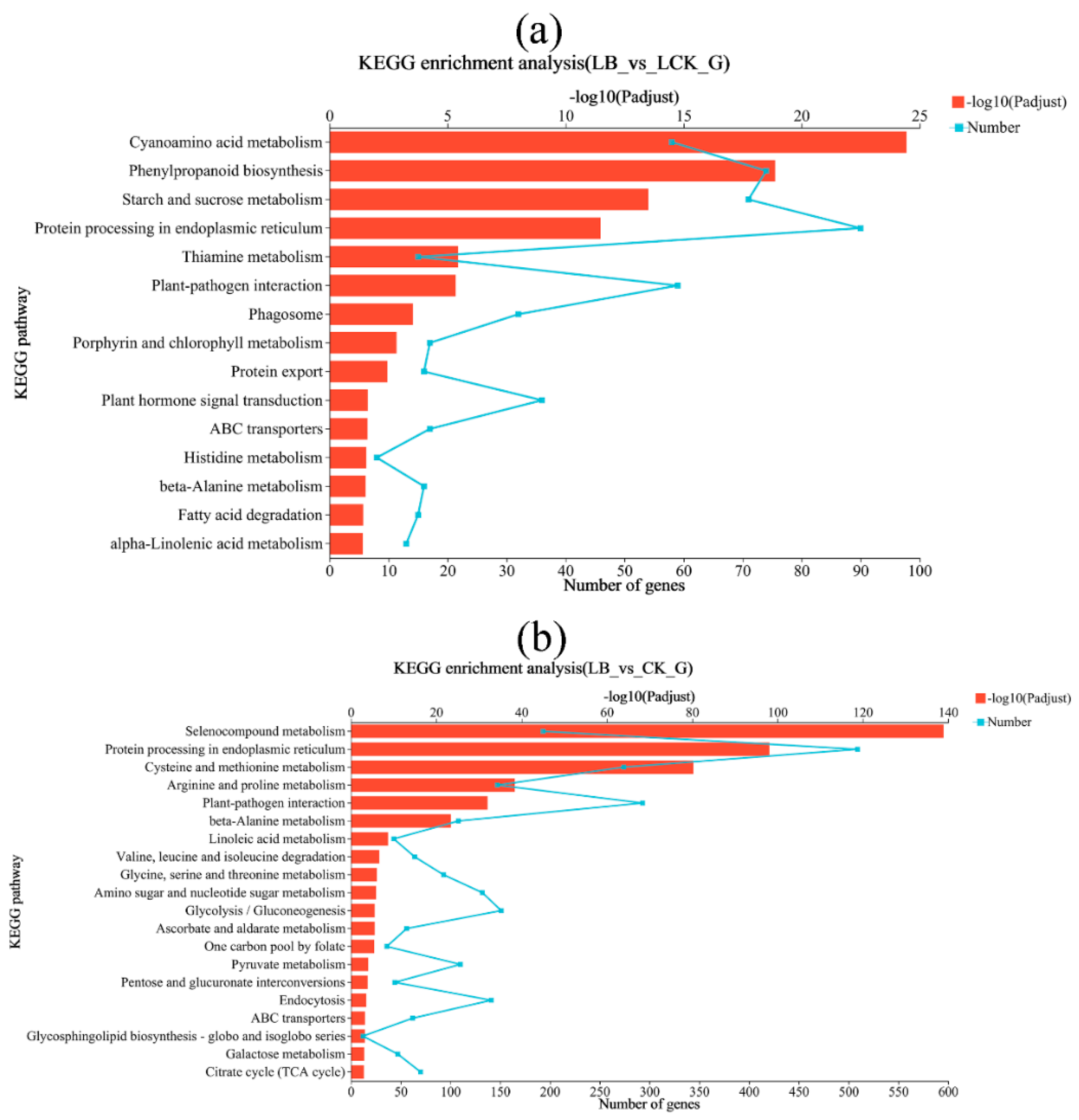
Figure 8.
Differential expressed gene in KEGG pathway enriched result. The color represents the expression level of the differential gene, the red represents the high expression level, and the blue indicates the low expression level.
Figure 8.
Differential expressed gene in KEGG pathway enriched result. The color represents the expression level of the differential gene, the red represents the high expression level, and the blue indicates the low expression level.
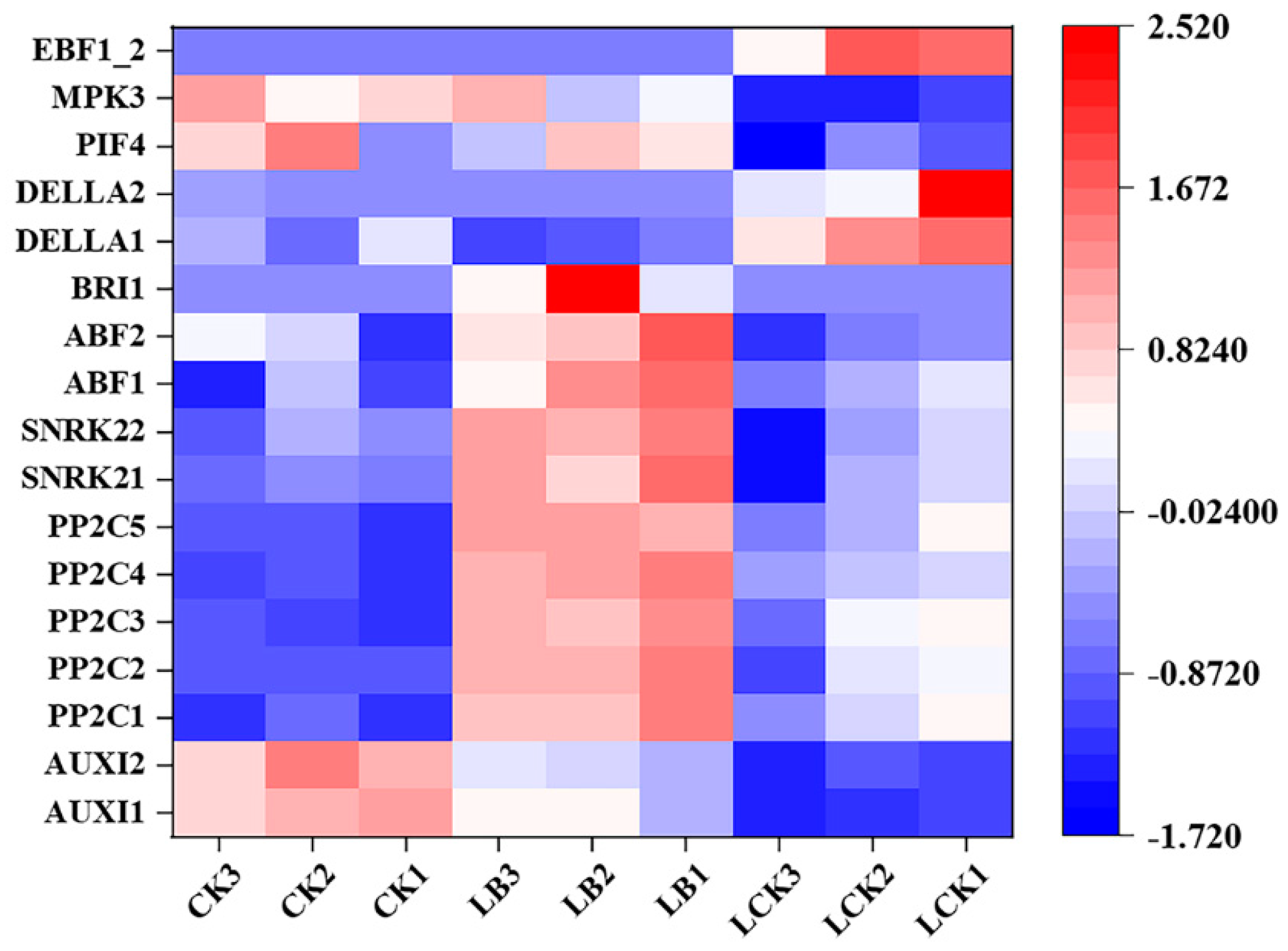
Figure 9.
Effects of Bacillus GL18 application on the expression of growth-related genes in oat plants under low-temperature stress. Different letters at each treatment indicate significance between inoculated and uninoculated conditions at the P ≤ 0.05 level based on a t-test.
Figure 9.
Effects of Bacillus GL18 application on the expression of growth-related genes in oat plants under low-temperature stress. Different letters at each treatment indicate significance between inoculated and uninoculated conditions at the P ≤ 0.05 level based on a t-test.
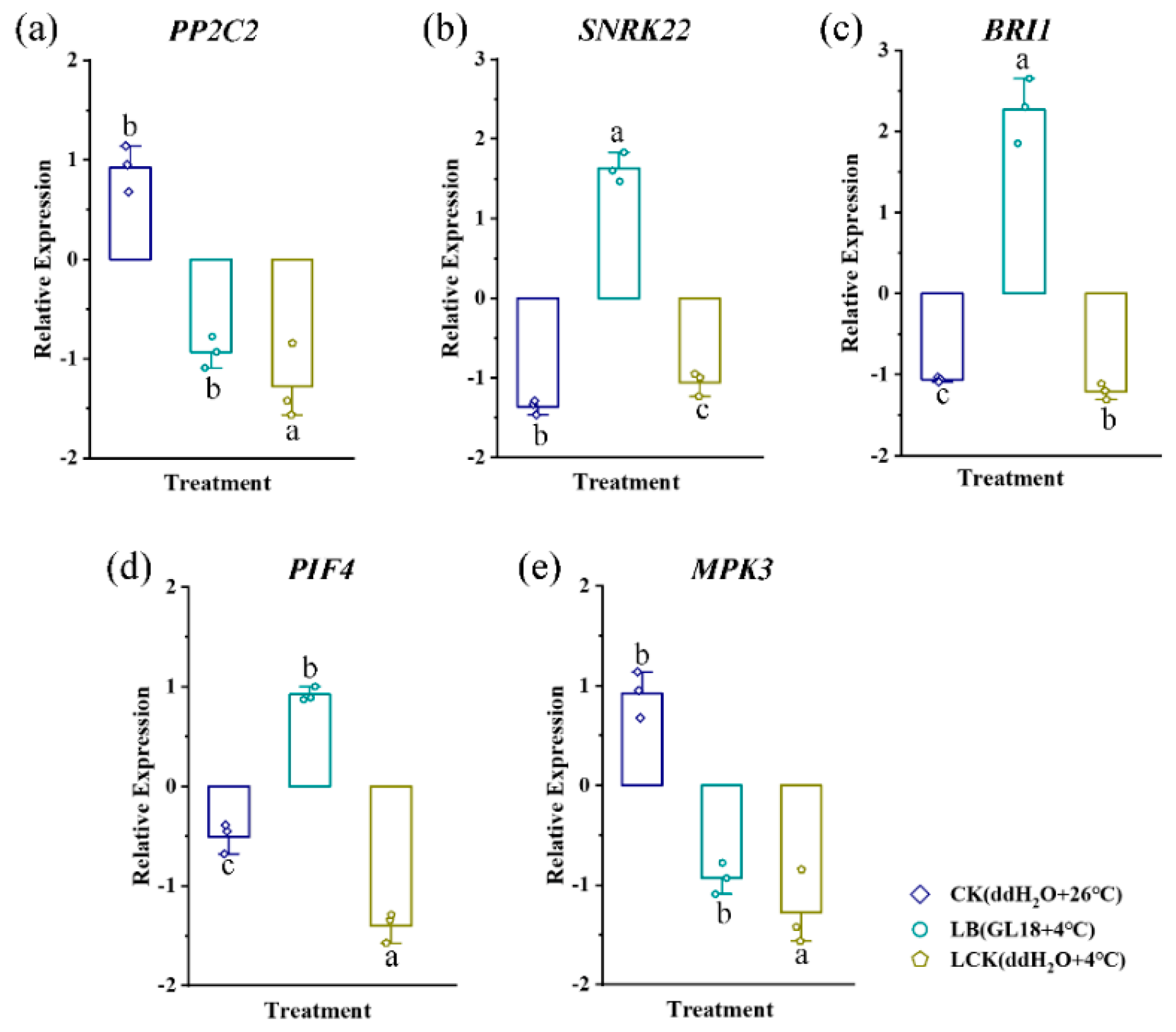
Figure 10.
Effect of Avena sativa inoculation with GL18.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



