
Submitted:
29 August 2024
Posted:
30 August 2024
Read the latest preprint version here
Abstract
Anthocyanins are well known as colorants of flowers, but many other invisible functions might have been more important during the evolution of complex biosynthesis networks in plants. The number of anthocyanin decorating enzymes, the subtle control of structural genes by numerous transcription factors, and routes of intracellular anthocyanin transport are reviewed here. Various ecological functions of anthocyanins hold the key to understanding evolutionary trajectories that lead to the success of these pigments. Proposed functions include carbon sinks that prevent excessive sugar levels, sun blockers protecting the photosynthesis apparatus, antioxidants scavenging reactive oxygen species, providing camouflage, and attraction of pollinators and seed dispersers. Anthocyanins clearly fulfill different functions in different plant organs. It is currently believed that protective functions in leaves gave rise to the biosynthesis network and that flower and fruit coloration evolved later.
Keywords:
anthocyanin modification
; flavonoids
; specialized plant metabolism
; pigmentation
; transcriptional regulation
; ecological function
; metabolic diversity
Background
Anthocyanins are well known as a phylogenetically widespread group of plant pigments providing colorful patterns to flowers and fruits. Anthocyanins can provide a range of different colors including red, pink, purple, and blue. This turns anthocyanins into an economically relevant target in the engineering of horticultural plants (Gu et al., 2019; Lozoya-Gloria et al., 2023). Especially the rise of genome editing methods like CRISPR/Cas in plants (Cardi et al., 2023) opens new avenues for targeted modification of the anthocyanin biosynthesis to achieve a desired flower color. However, such endeavors require a comprehensive understanding of all genes involved in the biosynthesis pathway. Due to the clearly visible phenotype, the anthocyanin biosynthesis quickly emerged as a model system for research on biosynthesis pathways (Winkel-Shirley, 2001). First studies investigating genes of the anthocyanin biosynthesis date back to the 1950s, when Barbara McClintock studied transposons in maize (McClintock, 1950) that generated diverse pigmentation patterns by disrupting anthocyanin biosynthesis genes. First biochemical studies of anthocyanins in flowers are even older - ranging at least back to 1835 (Marquardt, 1835). Anthocyanins were also involved in classical genetic experiments performed by Gregor Mendel, as he worked on the flower color of peas (Mendel, 1865). However, it took almost 150 years until the underlying genes were discovered (Hellens et al., 2010; Moreau et al., 2012). Anthocyanins are relevant for studies in many different fields, because they are present in almost all plant lineages. Given this distribution, it is not surprising that numerous studies described the loss of anthocyanin pigmentation in flowers due to mutations (Ho & Smith, 2016; Del Valle et al., 2019; Wong et al., 2024; Marin-Recinos & Pucker, 2024). This lack of pigmentation in individual plants or at a species level can be identified visibly and thus received substantial attention. In many cases, not the anthocyanin biosynthesis genes themselves are mutated, but changes in their transcriptional regulators prevent an activation of the anthocyanin biosynthesis apparatus (Marin-Recinos & Pucker, 2024). A noteworthy exception to the almost ubiquitous distribution of anthocyanins is the flowering plant order Caryophyllales, in which anthocyanins have been replaced by betalains, another pigment type with partial functional redundancy (Timoneda et al., 2019). Anthocyanins and betalains appear mutually exclusive as some families within the Caryophyllales maintained their anthocyanin pigmentation, while others show only betalain pigmentation (Mabry & Turner, 1964; Timoneda et al., 2019). This complex pattern of pigment biosynthesis emergence and loss in the Caryophyllales provides an excellent system for evolutionary studies on biosynthesis pathways (Sheehan et al., 2020; Pucker et al., 2024). To fully understand why anthocyanins have been replaced by betalains or why anthocyanin loss might be neutral or even advantageous under specific conditions, it is important to fully understand the biochemical and ecological functions of anthocyanins. This is a challenging undertaking given the biochemical diversity of anthocyanins facilitated by species-specific differences in the anthocyanin biosynthesis pathway and promiscuous enzymes that form a complex network of interconnected pathways.
In this review, we will summarize the existing knowledge about the biosynthesis of anthocyanins, the transcriptional control of this biosynthesis pathway, the intracellular transport of anthocyanins, and their various ecological functions.
Biosynthesis of a Diverse Set of Anthocyanins
Anthocyanins comprise an aglycone that can be decorated by a range of sugar moieties, acids, and methyl groups (Figure 1). While the biosynthesis of the aglycone is well understood and largely conserved between plant species, the decoration is highly diverse and often differs between species or larger evolutionary lineages (Pucker et al., 2020). For example, cyanidin 3,7,3’-triglucosides appear to be specific to Epidendroideae (Hughes et al., 2021). Flavonoid patterns are even considered useful characters in phylogenetic studies (Murai et al., 2008). The current knowledge about genes involved in the decoration of anthocyanins is limited to a small number of plant species. The huge diversity of flower colors and hues seen in other parts of plants suggest an enormous variety of different anthocyanin derivatives and combinations of these. Therefore, it is likely that most anthocyanin modification reactions have not been discovered and characterized yet.
Anthocyanins are produced through one branch of the flavonoid biosynthesis pathway (Figure 2). Other branches of the flavonoid biosynthesis are competing for substrate with the anthocyanin biosynthesis and lead to biochemically different classes of metabolites including flavones, flavonols, or proanthocyanidins. Phenylalanine is considered as the substrate of the anthocyanin biosynthesis. The general phenylpropanoid pathway comprising the three enzymes phenylalanine ammonium lyase (PAL), cinnamate 4-hydroxylase (C4H), and 4-coumarate:CoA ligase (4CL) processes phenylalanine and provides p-coumaroyl-CoA as substrate to the flavonoid biosynthesis. The enzymes naringenin-chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), dihydroflavonol 4-reductase (DFR), anthocyanidin synthase (ANS), and anthocyanin-related glutathione S-transferase (arGST) produce anthocyanidins which can be converted into anthocyanins through the addition of a sugar moiety by an UDP-dependent anthocyanidin-3-O-glucosyltransferase (3GT) (Winkel-Shirley, 2001; Grotewold, 2006; Eichenberger et al., 2023). The diversity of anthocyanins produced through this pathway is increased by two enzymes that can add additional hydroxyl groups to the B ring of the molecule: flavonoid 3’-hydroxylase (F3’H) and flavonoid 3’,5’-hydroxylase (F3’5’H) (Seitz et al., 2007). With increasing numbers of hydroxyl groups on the B-ring, anthocyanins are classified as pelargonidin, cyanidin, or delphinidin derivatives. This hydroxyl group difference has important functional consequences, because the color of the molecules ranges from orange-red (pelargonidin derivatives) to blue (delphinidin derivatives). Since genes required for this anthocyanin core biosynthesis are well conserved across plant species, their identification based on orthology is straight forward (Pucker et al., 2020; Rempel et al., 2023) and resulted in a large number of studies reporting on just these genes in numerous species (Marin-Recinos & Pucker, 2024).
Catalytic promiscuity is a feature of the specialized plant metabolism that can help to explain the plethora of different metabolites produced by a comparatively small number of enzymes and forms the basis for the evolution of novel enzymes (Weng & Noel, 2012; Moghe & Last, 2015). One prominent example in the flavonoid biosynthesis is the first committed enzyme CHS, which catalyzes the production of naringenin chalcone, but also has the ability to produce other polyketides (Ni et al., 2020; Waki et al., 2020). The binding of CHI-like (CHIL) proteins to CHS rectifies this promiscuity and ensures the efficient production of precursors for the following step in the flavonoid biosynthesis, but does not substantially increase the activity of CHS as initially assumed (Morita et al., 2014; Ban et al., 2018; Ni et al., 2020; Waki et al., 2020; Wolf-Saxon et al., 2023). CHS duplication and subsequent loss of CHIL interaction ability resulted in the evolution of novel enzyme functions through neofunctionalization (Waki et al., 2020). The interaction of CHS and CHIL appears conserved across all land plants and CHILs show species-specific affinity for CHS (Waki et al., 2020). Another striking example for catalytic promiscuity are the members of the large 2-oxoglutarate-dependent dioxygenase family that are active in the flavonoid biosynthesis: F3H, FLS, and ANS (Martens et al., 2003; Gebhardt et al., 2007; Kawai et al., 2014; Li et al., 2020a; Busche et al., 2021; Schilbert et al., 2021). While these enzymes predominantly catalyze different reactions, they can partially catalyze each other's reaction with the extent of these side activities appearing species specific (Martens et al., 2003; Gebhardt et al., 2007; Kawai et al., 2014; Li et al., 2020a; Schilbert et al., 2021). In the Apiaceae, FNS I was identified as another member of this gene family that evolved through gene duplication from F3H (Gebhardt et al., 2007; Andersen et al., 2016; Pucker & Iorizzo, 2023). However, F3H is assumed to have evolved from a promiscuous ancestor that harbored flavone synthase activity (Li et al., 2020a). In summary, genes encoding promiscuous enzymes that undergo specialization after gene duplications appear to be a predominant theme in the evolution of the flavonoid biosynthesis.
Genes associated with the decoration of anthocyanins are less conserved and thus knowledge about them beyond A. thaliana is sparse in the literature. Studies in A. thaliana identified a range of genes that are associated with the anthocyanin decoration including UGT75C1/At4g14090 (Tohge et al., 2005), UGT79B1/At5g54060 (Yonekura-Sakakibara et al., 2012), 3AT1/At1g03940 and 3AT2/At1G03495 (Luo et al., 2007), BGLU10/At4g27830 (Kovinich et al., 2014), 5MAT/At3g29590 (Luo et al., 2007), and SCPL10/At2g23000 (Fraser et al., 2007). Glycosylation of anthocyanidins constitutes the final step of the anthocyanin biosynthesis. Usually, anthocyanidins are first glycosylated at the C3 position. Tohge et al. (2005) demonstrated that UGT78D2 (At5g17050) catalyzes this glycosylation of anthocyanidins at the C3 positions. Like many other UGTs, UGT78D2 also shows a high substrate promiscuity: In A. thaliana and Vitis vinifera, 3-O-glycosylation of flavonols is also catalyzed by UGT78D2 (Ford et al., 1998; Tohge et al., 2005). In Glycine max, GmUGT78K1 from black seed coat showed 3-O-glycosylation activity of anthocyanidins and flavonols (Kovinich et al., 2010). Furthermore, UGT78G1 (GT83F) was identified as another enzyme that can perform 3-O-glycosylation of pelargonidin and cyanidin, while mostly acting on other flavonoids (Modolo et al., 2007, 2009). Interestingly, this enzyme has been reported to also catalyze deglycosylation of anthocyanidin 3-O-glycosides, i.e., the removal of a sugar moiety from an anthocyanin (Modolo et al., 2009). Following the 3-O-glycosylation, an additional 5-O-glycosylation is possible. Several studies reported 5-O-glycosyltransferases in a range of plant species. The best studied UDP-glycosyltransferase adding a glucose at the 5-position of the anthocyanin A-ring is the A. thaliana UGT75C1/AAt4g14090 (Tohge et al., 2005). UGT75C1 is responsible for glycosylation of anthocyanidin 3-O-β-D-glucosides leading to a 3-O-5-O-diglucoside product. Anthocyanidin 3-O-5-O-diglucosides have been reported to be more stable and soluble, responsible for bright-purple flower coloration (Jonsson et al., 1984) and form the basis for further complex modifications, e.g. sinapoylation or coumaroylation. Multiple UDP-dependent 5GTs were identified in Dahlia variabilis (Ogata et al., 2001), Petunia hybrida (Yamazaki et al., 2002), Gentiana triflora (Nakatsuka et al., 2008), Freesia hybrida (Ju et al., 2018), Cyclamen purpurascens (Kang et al., 2021) and Paeonia (Li et al., 2022). Nakatsuka et al. (2008) showed that the Gt5GT accepts different anthocyanidin 3-O-beta-D-glucosides as substrates whereas the enzyme does not show activity using aglyconic anthocyanidins as substrates (Nakatsuka et al., 2008). As reported for multiple UGTs, UGT75C1 also has a high promiscuity. In A. thaliana, UGT75C1 is involved in the glycosylation of the plant hormone indole-3-acetic acid (IAA) (Nvsvrot et al., 2023), whereas SlUGT75C1 glycosylates abscisic acid (ABA) as well as IAA (Sun et al., 2017). This example shows additional roles of UGT75C1 in fruit ripening and drought resistance in Solanum lycopersicum thus supporting a broad substrate promiscuity even beyond anthocyanins. Despite this high promiscuity, AtUGT75C1 does not appear to participate noticeably in flavonol glycosylation (Yonekura-Sakakibara et al., 2008). To the best of our knowledge, there are no reports about UDP-dependent enzymes catalyzing a 7-O-glycosylation of anthocyanins. However, DgAA7GT from Delphinium grandiflorum, an acyl-glucose-dependent glycosyltransferase, is proposed to glycosylate anthocyanidin 3-O-glycosides and anthocyanidin 3-O-malylglycosides (Matsuba et al., 2010). In monocotyledones, AaAA7GT from Agapanthus africanus was found to 7-O-glycosylate anthocyanidin 3-O-glycosides, 3-O-galactosides and 3-O-rutinosides (Miyahara et al., 2012). Both AA7GTs do not accept aglyconic anthocyanidins as substrates. In Vitis amurensis, 3,5,7-O-triglycosides were reported thus suggesting that an enzyme for 7-O-glycosylation must exist (Zhu et al., 2021). Various galactosylated anthocyanidins were reported in different Actinidia species: cyanidin- and delphinidin 3-O-galactosides, as well as cyanidin- and delphinidin 3-[2-(xylosyl)galactosides], requiring the activity of galactosyltransferases (Montefiori et al., 2009). In Vigna mungo, an UF3GaT was described, showing high UDP-dependent 3-O-galactosylation activity for different flavonols and anthocyanidins (Mato et al., 1998). UCGalT1, also an UDP-dependent galactosyltransferase, was identified in purple carrots (Daucus carota) taproots and purple celery (Apium graveolens) (Xu et al., 2016; Feng et al., 2018). Recombinant expression of DcUCGalT1 as well as AgUCGalT1 in Escherichia coli led to the presence of 3-O-galactosidated anthocyanins. Both enzymes did not showed activity with either UDP-glucose or UDP-xylose as a sugar donor but had a high promiscuity regarding the sugar acceptor: Both UCGalT1 showed activity when incubated with flavonols (quercetin and kaempferol) and UDP-galactose, whereas DcUCGalT1 also catalyzed the galactose transfer of UDP-galactose to cyanidin, peonidin and pelargonidin (Xu et al., 2016; Feng et al., 2018).
3-O-glycosylation is the prerequisite for multiple further modification steps. The addition of another sugar to the 3-O-glycosyl residue of the anthocyanin leads to formation of disaccharide residues, e.g. the attachment of rhamnose to 3-O-glycosylated anthocyanins results in anthocyanin-rutinosides (Harborne, 1962). Multiple anthocyanin rutinosides in different plant species, especially Petunia and Solanum, were detected (Andersen & Jordheim, 2006). Among others, 6-hydroxycyanidin 3-rutinoside, cyanidin 3-rutinoside, pelargonidin 3-rutinoside were found in Alstroemeria cultivars (Tatsuzawa et al., 2003). The first rhamnosyltranserases were described in Silene dioica and Petunia, catalyzing the addition of a rhamnosyl group to 3-O-glycosylated and 3,5-diglycosylated anthocyanins (Kamsteeg et al., 1980; Brugliera et al., 1994; Kroon et al., 1994). In 2013, Cs1,6RhaT, encoding for an anthocyanin 3-O-glycoside 1,6-rhamnosyltransferase, was discovered, catalyzing the formation of peonidin- and cyanidin rutinosides in Citrus species with a high promiscuity. i.e. affinity for flavanones, flavones, and flavonols (Frydman et al., 2013). Interestingly, the gene is only common to non-bitter Citrus species, whereas bitter-tasting Citrus species have the gene Cm1,2RhaT, encoding a 1,2-rhamnosyltransferase that rhamnosylates flavanone-7-O-glycosides (Frydman et al., 2004). In Lobelia erinus, two UDP-dependent rhamnosyltransferases were found (ABRT2 and ABRT4) to rhamnosylate anthocyanin 3-O-glycosides (Hsu et al., 2017).
In contrast to UDP-dependent glycosylation, AAGTs are acyl-glucose-dependent anthocyanin glucosyltransferases belonging to the glycoside hydrolase family 1 (GH1) (Matsuba et al., 2010). Multiple BGLUs are shown to be responsible for encoding GH1-type glycosyltransferases using already substituted flavonoids as a substrate in A. thaliana (Miyahara et al., 2013; Kovinich et al., 2014; Frommann et al., 2024). However, only AtBGLU10 seems to play a role in anthocyanin modification (Miyahara et al., 2013; Kovinich et al., 2014), whereas BGLU1, BGLU3, BGLU4, and BGLU6 prefer flavonols over anthocyanins as substrates (Ishihara et al., 2016; Frommann et al., 2024).
The most common anthocyanins have a xylosyl-group attached to the 2’’-position of the 3-O-sugar. In A. thaliana, AtUGT79B1 is known to catalyze the sugar attachment to 3-O-glycosylated anthocyanins (Yonekura-Sakakibara et al., 2012). Different anthocyanins can act as sugar acceptors, i.e. cyanidin 3-O-glucoside, delphinidin 3-O-glucoside and pelargonidin 3-O-glucoside. Interestingly, no activity was shown with cyanidin 3-O-rhamnoside but with cyanidin 3-O-rhamnosyl(1→6)glucoside as a sugar donor (Yonekura-Sakakibara et al., 2012). The enzyme also has a high promiscuity as it shows a high activity with the flavonols kaempferol 3-O-glucoside and quercetin 3-O-glucoside. Nevertheless, UDP-xylose is the only accepted sugar donor (Yonekura-Sakakibara et al., 2012). In Actinidia chinensis, AcUFGT6b is proposed to encode an anthocyanin xylosetransferase (Liu et al., 2019a).
Addition of acyl groups to anthocyanins is another common modification of anthocyanins. In crops alone, various acylated anthocyanins were reported in 23 species (Jokioja et al., 2021). Acylation of anthocyanins leads to higher activity, increased stability, and altered polarity which enhances the antioxidant effect and enables the usage as dye in industry (Jokioja et al., 2021). In A. thaliana, the known anthocyanin-related acyltransferases belong to the BAHD superfamily. For At3AT1 (At1g03940) and At3AT2 (At1G03495), different acyl donors (p-coumaroyl-CoA, feruloyl-CoA and caffeoyl-CoA) and acyl acceptors (cyanidin 3-glucoside, pelargonidin 3-glucoside, malvidin 3-glucoside, quercetin 3-glucoside, kaempferol 3-glucoside and kaempferol 7-glucoside) are known (Luo et al., 2007). At5MAT (At3g29590) also has a high promiscuity, as it acylates multiple 3-O-glycosylated anthocyanins, i.e. cyanidin 3,5-diglucoside, cyanidin 3-coumaroylglucoside 5-glucoside, cyanidin 3-O-(xylosyl) 6’’-O-p-coumaroyl glucoside 5-O-glucoside (A3) and delphinidin 3-coumaroylrutinoside 5-glucoside (Luo et al., 2007).
Transcriptional Regulation of Anthocyanin Biosynthesis
Crucial for the metabolic flux control through the flavonoid biosynthesis is the transcriptional regulation of enzyme-encoding genes (Figure 3). Many MYB transcription factors, members of the largest transcription factor family in plants, play crucial roles in this regulation (Jaakola, 2013; Albert et al., 2014; LaFountain & Yuan, 2021; Marin-Recinos & Pucker, 2024). Genes of the anthocyanin biosynthesis branch are activated by a complex of multiple transcription factors including a MYB, a bHLH, and a WD40 protein leading to the name MBW complex (Ramsay & Glover, 2005; Gonzalez et al., 2008). It is generally assumed that the MYB component is determining the target gene specificity of this complex, because different MYB proteins can be incorporated into this complex leading to variations in the set of target genes (Hichri et al., 2011; Gonzalez et al., 2016; Marin-Recinos & Pucker, 2024). While the concept of one transcription factor complex regulating all anthocyanin biosynthesis genes seems simple, a large number of gene duplications at different taxonomic levels turn the regulation into a complex system.
Since anthocyanin activating MYBs have been investigated in a large number of plant species, a pattern emerged that implicated different MYB subgroup 6 (SG6) members in the activation of the anthocyanin biosynthesis. They are generally interacting with bHLHs of the IIIf subgroup. Initial studies in Zea mays discovered R and B as two copies of an anthocyanin-regulating bHLH gene (Chandler et al., 1989). COLORLESS1 (C1) is a MYB transcription factor that was also identified as a crucial factor in the anthocyanin biosynthesis regulation of maize (Cone et al., 1986; Paz-Ares et al., 1987). In A. thaliana the MYB75 (PAP1), MYB90 (PAP1), MYB113, and MYB114 are anthocyanin regulators and also very close homologs suggesting an evolutionary recent radiation (Borevitz et al., 2000; Stracke et al., 2001; Gonzalez et al., 2008). Anthocyanin activating MYBs from monocot species like banana do not efficiently interact with the same bHLHs as MYBs from dicots like A. thaliana (Lloyd et al., 1992; Busche et al., 2023).
There is also an additional component, the WRKY protein TTG2, that can be associated with the MBW complex (Pesch et al., 2014; Verweij et al., 2016; Lloyd et al., 2017). The current model suggests that TTG2 is activated by the MBW complex and that the addition of the WRKY component to the MBW complex increases the specificity for the vacuolar transport associated genes TT12 and TT13 (Gonzalez et al., 2016; Lloyd et al., 2017). PH3 is a petunia gene with a function similar to the Arabidopsis TTG2 (Verweij et al., 2016) thus suggesting that the involvement of WRKY is conserved across plant species. Anthocyanin biosynthesis activating WRKYs have also been described in pear and apple, but at least apples MdWRKY40 is reported to be functionally divergent from PH3 and TTG2 (An et al., 2019; Li et al., 2020b).
There are also reports that anthocyanin biosynthesis activation is possible without the canonical MBW complex. A recent study in blueberry revealed that proanthocyanidin biosynthesis regulators like TT2/MYB123 (subgroup 5, SG5) can contribute to the anthocyanin regulation (Karppinen et al., 2021). Several studies suggested that MYB5 orthologs are also able to activate the anthocyanin biosynthesis (Cavallini et al., 2014; Jiang et al., 2023; Li et al., 2024). In Vitis, VvMYB5a and VvMYB5b can activate specific genes in the anthocyanin biosynthesis with VvMYB5b being able to slightly trigger the accumulation of anthocyanins when heterologously expressed in unpigmented petunia (Cavallini et al., 2014). Additionally, it seems that only VvMYB5a and VvMYB5b, but not the MYB75/PAP1/AN2 ortholog VvMYBA1 are able to activate the HYDROXYLATION AT FIVE (HF2, F3’5’H) expression (Cavallini et al., 2014). It is important to note that Vitis also harbors VvMYBA1 and VvMYBA2 as orthologs of PhAN2 which seem to be the dominant activator of most anthocyanin biosynthesis genes (Walker et al., 2006; Azuma et al., 2007; Cavallini et al., 2014; Jiu et al., 2021). Studies in strawberries reported MYB5 as an anthocyanin regulator that is TTG1-independent, but forms a MBW complex with EGL3 (bHLH) and LWD1 (WD40) instead (Jiang et al., 2023). Reports in other species also associated MYB5 with anthocyanin biosynthesis: PhPH4, also belonging to the MYB5 lineage, regulates vacuolar acidification in petunia (Quattrocchio et al., 2006), SmMYB5 activates anthocyanin biosynthesis in eggplant (Li et al., 2024), and FhMYB5 expression in Freesia hybrida correlates with anthocyanin pigmentation (Li et al., 2019c). Additionally, MYB5a/NEGAN is responsible for the pigmentation pattern in Mimulus (Zheng et al., 2021). While studies in many species identified MYB5 orthologs as anthocyanin biosynthesis regulators CsMYB5a and CsMYB5e in Camellia sinensis activate proanthocyanidin biosynthesis, but not anthocyanin biosynthesis (Jiang et al., 2018) and A. thaliana MYB5 seems not tightly connected to the anthocyanin biosynthesis. In summary, a part of the MYB5 lineage seems to activate specific genes in the anthocyanin biosynthesis which could represent a mechanism to fine-tuned activation of specific branches within the complex anthocyanin metabolism.
A number of other transcription factors have been implicated in the regulation of anthocyanin biosynthesis genes including ERF, NAC, MADS-box, bZIP, BBX, and PIF3. In Malus domestica, ethylene response factor 109 (ERF109) activates the anthocyanin biosynthesis in early stages of apple coloration by binding promoters of MdCHS, MdUFGT, and MdbHLH3 (Ma et al., 2021). Another study in red-skinned pear reported the interaction of PyERF with the anthocyanin regulators PyMYB114 and PybHLH3 to co-regulate the anthocyanin biosynthesis (Yao et al., 2017). NAC proteins have been identified as activators of the anthocyanin biosynthesis in Arabidopsis thaliana and Prunus persica (Morishita et al., 2009; Zhou et al., 2015). NACs probably trigger anthocyanin pigmentation through up-regulation of the important anthocyanin biosynthesis activating MYB as this was observed for a BLOOD/PpNAC1 heterodimer in the coloration of blood-fleshed peach during fruit ripening (Zhou et al., 2015). Since the expression pattern of the SQUAMOSA-class MADS-box gene VmTDR4 is tightly correlated with the anthocyanin pigmentation in bilberry fruits, it was postulated that this transcription factor directly or indirectly controls the anthocyanin biosynthesis genes (Jaakola et al., 2010). The bZIP protein AtHY5 binds to the promoter of MYB75/PAP1 and triggers the anthocyanin biosynthesis through up-regulation of this MYB gene (Shin et al., 2013). This result is supported by a study in Malus domestica that identified MdHY5 as an activator of the anthocyanin regulator MdMYB10 (An et al., 2017). Another bZIP protein, PybZIPa, was reported as an activator of the anthocyanin activating MYB and UFGT, but not orthologous to HY5 (Liu et al., 2019b). The A. thaliana BBX protein BBX22/LZF1 was identified as an activator of the anthocyanin-regulating MYB75/PAP1 (Chang et al., 2008). HY5 was discovered as a factor contributing to the expression of BBX22/LZF1 (Chang et al., 2008). The apple BBX protein MdCOL11, ortholog of AtBBX22, is interacting with MdHY5 to activate the anthocyanin regulator MdMYBA (Bai et al., 2014). A study in rice identified that OsBBX14 and OsHY5 interact to activate the anthocyanin regulating MYB and bHLH gene (Kim et al., 2018). A study in red pear revealed that PpBBX16 interacts with PpHY5 to activate the expression of anthocyanin biosynthesis genes (Bai et al., 2019a). In summary, this suggests that HY5 usually acts in a complex with a BBX protein. In A. thaliana, PIF3 is another light-responsive transcription factor that can activate the anthocyanin biosynthesis, but requires the simultaneous binding of HY5 (Shin et al., 2007). Another positive regulator of the anthocyanin biosynthesis in A. thaliana, especially associated with up-regulation of the UFGT, is the phytochrome-interacting ankyrin repeat protein 2 (PIA2) (Yoo et al., 2011).
MYB112 was reported to trigger anthocyanin biosynthesis and block flavonol biosynthesis under salt and high light stress conditions, by activating the anthocyanin activator PAP1 and repressing the flavonol activators MYB12/MYB111 (Lotkowska et al., 2015).
Not all transcription factors of the anthocyanin biosynthesis are activators. A comprehensive review summarized the role of various factors that repress the expression of anthocyanin biosynthesis genes (LaFountain & Yuan, 2021). Examples for characterized MYB repressors connected to the anthocyanin biosynthesis are PhMYB27 in petunia (Mur, 1995), FaMYB1 in strawberry (Aharoni et al., 2001), CPC in Arabidopsis (Zhu et al., 2009), SPL9 group in Arabidopsis (Gou et al., 2011), MYBL2 in Arabidopsis (Dubos et al., 2008), MYBC2-L1 and MYBC2-L3 in grapevine (Cavallini et al., 2015), MYBF2 in ginkgo (Xu et al., 2014), and MaMYB4 in banana (Deng et al., 2021).
Displayed transcription factors are ETHYLENE RESPONSIVE FACTOR 109 (ERF109), BLOOD, NAM - ATAF1- CUC2 (NAC1), Vaccinium myrtillus TAPETUM DEGENERATION RETARDATION 4 (VmTDR4), phytochrome interacting factor 3 (PIF3), LONG HYPOCOTYL 5 (HY5), B-BOX DOMAIN PROTEIN 22 (BBX22), phytochrome-interacting ankyrin repeat protein 2 (PIA2), Pyrus pyrifolia bZIPa (PybZIPa). Interactions are displayed via arrows, dashed arrows indicate involvement in early anthocyanin biosynthesis (Ma et al., 2021, Zhou et al., 2015, Jaakola et al., 2010, Shin et al., 2007, Shin et al., 2013, An et al., 2017, Chang et al., 2008, Yoo et al., 2011, Liu et al., 2019b).
The myeoblastosis - basic helix-loop-helix – WD40 (MYB-bHLH-WD40, MBW) complex consists MYB75/PRODUCTION OF ANTHOCYANIN1 (PAP1), MYB90/PAP2, MYB113, MYB114, Vitis vinifera MYBA1,Petunia hybrida anthocyanin 2 (PhAN2), DEEP PURPLE (DPL), PURPLE HAZE (PHZ) or MYB5 and bHLH42/TRANSPARENT TESTA 8 (TT8), GLABRA 3 (GL3), ENHANCER OF GLABRA 3 (EGL3), myelocytomatosis 1 (MYC1), Petunia hybrida anthocyanin 1 (PhAN1) or JOHNANDFRANSCESCA 13 (JAF13) and TRANSPARENT TESTA GLABRA 1 (TTG1) and LIGHT-REGULATED WD1 (LWD1).
Displayed structural genes are chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), flavonoid 3’-hydroxylase (F3’H), dihydroflavonol 4-reductase (DFR), anthocyanidin synthase (ANS), anthocyanin-related glutathione S-transferase (arGST) and UDP-dependent flavonoid-3-O-glycosyltransferase (UFGT).
Activity of the anthocyanin biosynthesis is also controlled by regulatory RNAs. There are microRNAs (miRNAs) that suppress target genes and long non-coding RNAs (lncRNAs) that counteract the miRNAs by target mimicry, i.e., sequestering miRNAs without cleavage of the lncRNAs (Franco-Zorrilla et al., 2007; Zhang et al., 2018; Meng et al., 2023). Several studies associated expression patterns of certain lncRNAs with the accumulation of anthocyanins, but evidence for the molecular mechanism is missing (Zhang et al., 2022b; Yu et al., 2022; Tan et al., 2023). Anthocyanin accumulation in Arabidopsis is increased by miR156 that targets anthocyanin repressors of the SPL9 group (Gou et al., 2011). The authors demonstrated that SPL9 captures PAP1 proteins and prevents the formation of the MBW complex that is required for the activation of anthocyanin biosynthesis genes. A similar system comprising miR156a and SPL9 as negative regulators of the anthocyanin biosynthesis was reported as the mechanism explaining the coloring of the peel in red pear (Liu et al., 2021). Two long non-coding natural antisense transcripts (lncNATs) have been reported as repressors of the anthocyanin biosynthesis activators DcMYB6 and DcMYB7 (Chialva et al., 2021). A regulatory system identified in Malus spectabilis under nitrogen starvation comprises miR858 as a repressor of MYB62-like, which in turn represses the anthocyanin biosynthesis, thus high expression of miR858 leads to anthocyanin accumulation (Meng et al., 2023). This study also identified eTM858-1 and eTM858-2, two target mimics of miR858, which reduce the activity of miR858 on MYB62-like thus leading to a repression of the anthocyanin biosynthesis (Meng et al., 2023). In sea buckthorn, LNC1 and LNC2 (TCONS_00694050 and TCONS_00438839) were reported as endogenous target mimics of miR156a and miR828a, respectively (Zhang et al., 2018). While miR156a targets the anthocyanin biosynthesis repressor SPL9, miR828a targets the anthocyanin biosynthesis activator MYB114 (Zhang et al., 2018). Consequently, a high abundance of LNC1 leads to reduced anthocyanin accumulation in the fruit, while higher abundance of LNC2 results in a higher anthocyanin accumulation (Zhang et al., 2018).
In apple, MdLNC499 activates the expression of MdERF109, which encodes a transcriptional activator of the anthocyanin biosynthesis genes CHS, bHLH3, and UFGT (Ma et al., 2021).
Transport of Anthocyanins
Anthocyanins are produced at the endoplasmatic side of the ER and require transportation into the central vacuole for long term storage (Pucker & Selmar, 2022) (Figure 4). Details regarding the intracellular transport of anthocyanins have eluded researchers as results of previous studies seem to contradict each other. Different ABCC proteins have been implicated in the transport of anthocyanins across a membrane (Goodman et al., 2004; Francisco et al., 2013; Behrens et al., 2019). However, it is not clear whether anthocyanins are imported into the ER and transferred to the vacuole or transported through the cytoplasm and then imported into the central vacuole (Pucker & Selmar, 2022). The mechanism could differ between plant species or both routes could contribute to the anthocyanin transport. The discovery of TT9, a vesicle associated protein, as a crucial factor for proanthocyanidin pigmentation in A. thaliana (Ichino et al., 2014) could suggest that anthocyanin transport is also occurring through vesicles. MATE (TT12) transporters have also been proposed as potential anthocyanin transporters (Marinova et al., 2007). They would require a proton gradient that could be maintained by AHA10/TT13 (Appelhagen et al., 2015). However, it was also suggested that MATEs are not transporting anthocyanins, but proanthocyanidin precursors (Zhao & Dixon, 2009). Previously, it was postulated that TT19, a glutathione S-transferase (GST), serves a ‘ligandin’, an anthocyanin protection protein, during the transport through the cytoplasm (Mueller et al., 2000; Kitamura et al., 2004; Sun et al., 2012). However, a recent study demonstrated an enzymatic function of this anthocyanin-related GST (arGST) in the synthesis of cyanidin (Eichenberger et al., 2023). While this finding does not rule out an arGST function in the transport of anthocyanins, it provides an alternative explanation for previous observations that led to the postulation of the arGST function in the anthocyanin transport. Understanding the transport of anthocyanins is crucial as some modification reactions take place in the central vacuole, i.e., after the successful translocation (Nozue et al., 1997). Anthocyanin localization within the cell can influence the phenotypic appearance of the plant. Clusters of anthocyanins can be dispersed rapidly, altering the color of the plant organ (Irani & Grotewold, 2005). Changes in the vacuolar acidity can quickly change the appearance of anthocyanins as these are depending on the pH (Verweij et al., 2008). While there is substantial research on the intracellular transport of anthocyanins, not much is known about the transport of anthocyanins between plant cells or even different organs (Pucker & Selmar, 2022). Since all plant cells should be able to produce anthocyanins based on phenylalanine, transport between cells might not occur at a relevant level.
Cell-Specific Accumulation of Anthocyanins and Pigmentation Patterns
Pigmentation patterns mostly serve the attraction of pollinators by boosting the visibility of the flower to increase the number of visitation events. This can be achieved through high contrast patterns or so-called landing lights (Davies et al., 2012). These landing lights are usually colored in a shade or color contrasting the rest of the flower and provide information for the pollinator on the location and orientation for the landing on the flower (Richter et al., 2023). From there, spatially pigmented nectar guides can lead the way to the nectaries thus substantially increasing pollination success (Owen & Bradshaw, 2011; Davies et al., 2012). Anthocyanin pigmentation patterns can usually be attributed to the spatial expression of flavonoid and anthocyanin pathway related genes. Anthocyanin biosynthesis genes are regulated by an ensemble of transcription factors that are active in very specific flower parts and tissues (Davies et al., 2012; Albert et al., 2014; Ding et al., 2020). The anthocyanin biosynthesis promotion MYB is the most specific transcription factor in most plant species (Marin-Recinos & Pucker, 2024). Different tissue types of the flower petal can contribute to specific spatial pigmentation of the tissue or the adjacent cells. This can be seen in Antirrhinum in the phenotype Venosa where the transcription factor VENOSA from the vascular tissues leads to red coloration of the adjacent adaxial epidermis (Shang et al., 2011). In Petunia hybrida, the MADS-box gene DEFICIENS was reported as a cell layer-specific transcription factor in flowers that influences the pigmentation pattern through control of the expression of the major anthocyanin biosynthesis regulating MYB ANTHOCYANIN2 (Chopy et al., 2024). In Gossypium barbadense a MYB transcription factor is essential for the formation of one large spot at the base of the petal which attracts pollinators to the flowers more efficiently (Abid et al., 2022). In Gossypium arboreum a differentially expressed GST was reported as being responsible for an analogous petal spot (Zhang et al., 2022a). The petal spot formation in Gorteria diffusa is also controlled by an anthocyanin MYB that triggers the malonylation of anthocyanins specifically in this petal region (Fattorini et al., 2024).
Several plant species display intensely pigmented spots on their petals that appear to be randomly distributed. For example, in Mimulus lewisii, Mimulus guttatus, and Digitalis purpurea the lower petal contains an area with multiple spots contrasting the background (Glover et al., 2013; Ding et al., 2020; Wolff et al., 2024). This area is centered on the petal and forms nectar guides in Mimulus (Ding et al., 2020) and a putative landing site in Digitalis (Wolff et al., 2024). In Mimulus, it was shown that the basis of spot formation is an activator-repressor-relationship of two transcription factors (Figure 5), which is conforming with a reaction-diffusion model proposed by Turing and Gierer & Meinhardt (Turing, 1952; Gierer & Meinhardt, 1972; Ding et al., 2020). This model assumes a relationship between an activator and a repressor, in which the activator (in this case the R2R3-MYB NEGAN) enhances its own expression and, in this case, upregulates the anthocyanin biosynthesis genes (Ding et al., 2020). Through diffusion of the activator into adjacent cells, an anthocyanin-pigmented spot develops (Ding et al., 2020). At the same time the activator upregulates a repressor gene (here: R3-MYB MlRTO) which competes with the activator for binding with the bHLH and WD40 partners (Ding et al., 2020). Thereby it reduces the functionality of the activator and therefore the expression of anthocyanin biosynthesis genes (Ding et al., 2020). Both regulators diffuse over cell boundaries. The repressor diffuses faster and represses the anthocyanin biosynthesis gene activity in cells adjacent to the cells where the activator is active (Ding et al., 2020). The pigmented spot is confined to the specific boundaries being set by the diffusion- and degradation-properties of both transcription factors. White halos surrounding pigmented spots on otherwise differently pigmented backgrounds can be explained by the repressor activity in the spot-adjacent cells (Davies et al., 2012; Ding et al., 2020).
Another striking pigmentation pattern presents itself in the Petunia hybrida ‘Red Star’ variety. Flowers of this variety display a white star-like pattern on a red petal background (Koseki et al., 2005). It was shown that spatial post transcriptional gene silencing of the PhCHS A mRNA is responsible for the lack of pigmentation around the center vein of the petals (Koseki et al., 2005). The resulting lack of CHS A enzymes in this tissue leads to the disruption of the early flavonoid pathway and a subsequent block of anthocyanin production.
Depending on their localization in certain cell layers (spongy palisade mesophyll, upper epidermis, lower epidermis), the same anthocyanins can lead to different colors of plant structures as recently observed in leaves of Tipularia discolor (Hughes et al., 2021). A large study investigating hundreds of plant species reported that mesophyll is the most frequent location of anthocyanins and only 24% of the studied species displayed anthocyanins in the epidermis (Lee & Collins, 2001). The huge diversity of anthocyanin accumulation patterns reported is strong support for a broad range of different anthocyanin functions (Lee & Collins, 2001; Gould, 2004). In contrast, anthocyanins have been reported to accumulate in the epidermis of A. thaliana plants under nitrogen starvation (Shi & Xie, 2010). A study on various crucifers also reported high anthocyanin concentration in the epidermis with spread into the mesophyll occurring only at very high anthocyanin concentrations (Mark Hodges & Nozzolillo, 1996). Understanding the localization of anthocyanins inside plants is a remaining challenge that can help to understand their physiological function and evolutionary relevance.
Ecological Functions of Anthocyanins
Functions of anthocyanins might differ depending on developmental state, plant organ, or environmental conditions. For example, anthocyanins in leaves are likely protecting against high light intensities and other stress factors (Pringsheim, 1881; Wheldale, 1916; Landi et al., 2015), while anthocyanins in flowers and fruits are most likely attracting pollinators and seed dispersers (Weiss, 1991; Willmer et al., 2009; Ruxton & Schaefer, 2016; Garcia et al., 2022). Understanding the ecological functions of anthocyanins is crucial when generating hypotheses about evolutionary trajectories that lead to the anthocyanin biosynthesis as we see it today. For example, loss of anthocyanins could coincide with changes in the pollination system.
Protective Functions of Anthocyanins in Photosynthetically Active Plant Organs
Anthocyanins protect green plant parts from excessive light intensities and oxidative stress. While the leaves of many plants turn red in response to high light intensities (Albert et al., 2009; Araguirang & Richter, 2022), other species show constant red leaf pigmentation (Gould et al., 1995; Burger & Edwards, 1996; Chalker-Scott, 1999; Logan et al., 2015). The red color of anthocyanins is due to a strong absorbance of blue-green light which could damage cell components (Smillie & Hetherington, 1999; Neill & Gould, 2000). Anthocyanins can act as sunscreen shielding the leaf tissue from excess light that would otherwise be captured by chlorophyll b (Gould et al., 1995; Feild et al., 2001). The maximum photosynthesis of leaves with anthocyanin pigmentation was increased compared to green leaves and photoinhibition was reduced (Gould et al., 1995; Manetas et al., 2003). The red undersurface of understory plants or floating leaves of aquatic plants is probably not backscattering red light as initially hypothesized, but helps the plants to cope with high-intensity sun flecks (Gould et al., 1995; Hughes et al., 2008).
In addition to protection against excess light, anthocyanins have also been reported as antioxidants that quench reactive oxygen species (ROS) that could otherwise damage cell structures (Gould et al., 2002a; Nagata et al., 2003; Kytridis & Manetas, 2006). While ROS are mainly produced in chloroplasts, anthocyanins are transported into and stored in the central vacuole (Pucker & Selmar, 2022). This brings up the question how anthocyanins can counteract ROS without physical proximity. Photooxidative stress leads to an inactivation of the ascorbate peroxidase that normally degrades H2O2 (Yabuta et al., 2002; Kytridis & Manetas, 2006). H2O2 is the ROS type that temporarily accumulates in plastids under photooxidative conditions and can move into the vacuole either through passive diffusion or through proteins located in the tonoplast (Nakano & Asada, 1980; Bienert et al., 2006; Agati et al., 2013). The anthocyanin-filled vacuole would become a H2O2 sink which can explain the ROS scavenging through anthocyanins accumulated in the vacuole probably catalyzed by peroxidases (Yamasaki et al., 1997; Gould et al., 2002b; Kytridis & Manetas, 2006). This hypothesis aligned with the observation that ROS quenching by anthocyanins is substantially weaker in species like Rosa sp. and R. communis where anthocyanins are not located in the same cells as the plastid accumulating H2O2 (Kytridis & Manetas, 2006). A systematic study revealed that the photoprotective function of anthocyanins is especially important at low temperatures and high light intensities, when other photoprotection mechanisms are less effective (Gould et al., 2018). Anthocyanin levels might also be regulated in response to ROS to ensure that excess ROS is scavenged (Xu & Rothstein, 2018). The photoprotective role of anthocyanins is not restricted to leaves, but also extends to photosynthetically active stems (Gould et al., 2010). A protective function of anthocyanins could explain their presence in young leaves, seedlings, or developing fruits (Lee et al., 1987; Christie et al., 1994; Dodd et al., 1998; Merzlyak & Chivkunova, 2000; Chen et al., 2021). These emerging structures require a protective anthocyanin pigmentation, because light capture ability develops before CO2 assimilation capacity thus protection against photoinhibition is needed (Krause et al., 1995; Karageorgou & Manetas, 2006). The de novo synthesis of anthocyanins in senescent leaves during autumn did appear less obvious and turned into an intensely studied field with many hypotheses arising to explain this phenomenon (Matile et al., 1992; Matile, 2000; Feild et al., 2001; Archetti, 2009; Archetti et al., 2009; Renner & Zohner, 2019). Anthocyanins can delay senescence and ensure functionality of leaves which improves the resorption of nitrogen and phosphorus in autumn (Hoch et al., 2001; Feild et al., 2001; Lee et al., 2003). Anthocyanin formation during autumn leading to red leaves is more pronounced in species that grow in areas with shorter vegetation periods and are more often experiencing cold snaps (Renner & Zohner, 2019). In the light of the huge importance of anthocyanins in photoprotection, it is hypothesized that this function of anthocyanins pre-dates their visual cue functions (Gould et al., 1995).
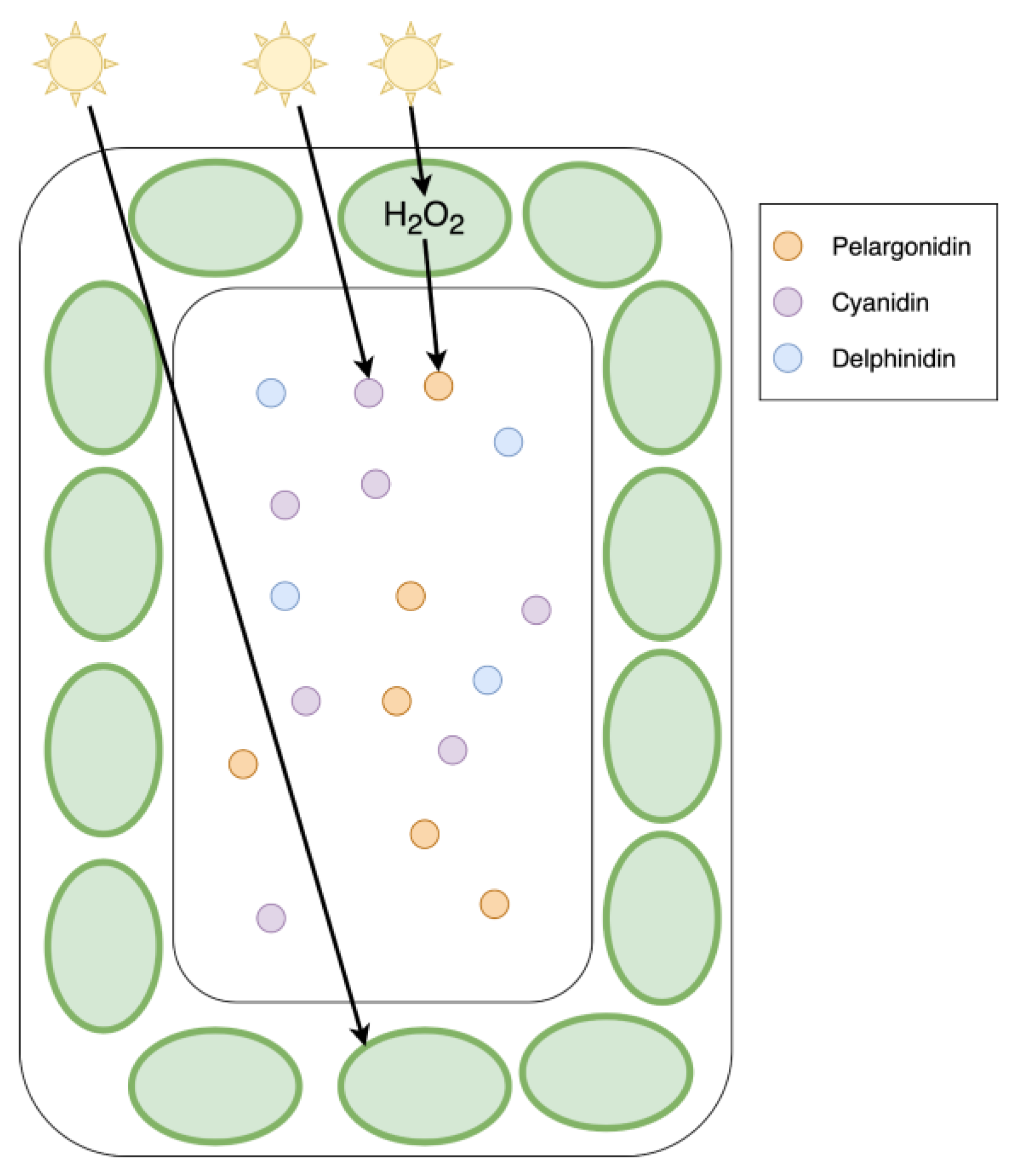
Figure 6.
Photoprotective and antioxidant functions of anthocyanins. H2O2 is generated in the chloroplasts by light exposure, but can move into the central vacuole, where antioxidant anthocyanins are located that can scavenge ROS.
Figure 6.
Photoprotective and antioxidant functions of anthocyanins. H2O2 is generated in the chloroplasts by light exposure, but can move into the central vacuole, where antioxidant anthocyanins are located that can scavenge ROS.
Importance of Anthocyanins in Drought and Salt Stress Response
Plants under drought stress caused by limited water availability or increased salt concentrations were often observed to turn red through the accumulation of anthocyanins. Studies exploring the transcriptomic or metabolomic changes report a generally increased activity of the flavonoid biosynthesis without specific explanations how the anthocyanin biosynthesis is increased. Therefore, it is plausible that ROS scavenging is performed by anthocyanins and other flavonoids, i.e., the enrichment of anthocyanins is the consequence of generally increased activity of the flavonoid biosynthesis. A specific function of anthocyanins might be capturing excess light as photosynthesis cannot take place in the absence of water. In Arabidopsis, the two glycosyltransferases UGT79B2 and UGT79B3 were identified as important targets for activation of the anthocyanin biosynthesis under drought and salt stress conditions, but also in response to cold stress (Li et al., 2017).The importance of flavonols in response to salt stress was investigated based on flavonol deficient mutants that were more adversely affected than the wild type (Li et al., 2019a). Given that the flavonol and anthocyanin biosynthesis branches are competing for shared substrates (Choudhary & Pucker, 2024), this observation suggests that generally antioxidants, but not specifically anthocyanins, are needed under these stress conditions. An investigation of carrot cell cultures under salt stress discovered an importance of MATE in the increased anthocyanin accumulation (Saad et al., 2021), which could indicate that a generally increased transport across membranes to counteract the osmotic challenges increases anthocyanin transport as a side effect. Anthocyanins account for less than 1% of the osmotic potential in a plant cell which makes a role as compensating salts unlikely (Manetas, 2006; Archetti et al., 2009). The sparsity of data about anthocyanin regulation in response to drought and salt stress suggests that their role in this context is rather minor and thus more challenging to resolve. This indicates that drought and salt stress response were not among the factors that contributed to the evolution of the anthocyanin biosynthesis.
Cold Stress Response
When exposed to low temperatures, many plants turn red due to accumulation of anthocyanins. This color change is caused by transcriptional up-regulation of the genes involved in the anthocyanin biosynthesis (Christie et al., 1994). Some evergreen species that maintain their leaves over the winter turn completely red due to intense anthocyanin accumulation (Oberbaueri & Starr, 2002; Hughes et al., 2005). Initially, it was postulated that anthocyanins could potentially turn light into heat thus increasing the plant temperature (Stiles, 1982), but presence in tropical plants makes this hypothesized function of foliar anthocyanins unlikely (Gould et al., 1995; Lee, 2002; Lee et al., 2003). Following the observation that light at low temperatures is particularly harmful for plants, it appears more likely that anthocyanins in evergreens protect against photoinhibition during winter (Gould et al., 1995; Oberbaueri & Starr, 2002; Hughes et al., 2005; Manetas, 2006). A role of anthocyanins in cold-stress mitigation was also reported in sweet potato (Wang et al., 2013) and tobacco (Naing et al., 2018). While a likely anthocyanin function of anthocyanins under cold stress is protection and potentially avoiding a sugar excess, the role of differently modified anthocyanins in cold response remains an open question. In Arabidopsis, high ambient temperatures lead to an inactivation of the anthocyanin biosynthesis genes through repression of the positive regulator HY5 (Kim et al., 2017). In plum fruit, the consumption of anthocyanins is increased at high temperatures due to high peroxidase activities resulting in lower anthocyanin levels (Niu et al., 2017). However, the balance between synthesis and consumption could be different between leaves and fruits or flowers.
Anthocyanin Accumulation as Sign of Nutritional Imbalance
An imbalance in the availability of sugar, nitrogen, phosphor, and other nutrients can trigger the accumulation of foliar anthocyanins (Jezek et al., 2023). Magnesium, sulfur, boron, copper, and sometimes potassium deficiencies can also result in the blushing of plants (Lawanson et al., 1972; Hajiboland & Farhanghi, 2010; D’Hooghe et al., 2013; Tavares et al., 2013; Kobayashi et al., 2013; Naya et al., 2014; Jezek et al., 2023), but results for some nutrients are restricted to individual species and limit general conclusions. While sugar and nitrogen are well investigated, little is known about most of the other factors. A positive impact of anthocyanins on performance under phosphorus starvation is suggested by an experiment with wheat seedlings (Kaur et al., 2022). ABI5 might be the interconnection of the anthocyanin boosting ABA signal, light signals, and the phosphor starvation signal in Arabidopsis (Wang et al., 2021; Lei et al., 2022). Results regarding the impact of sulfur starvation on the anthocyanin biosynthesis are inconsistent and might indicate species-specific differences, which have recently been reviewed by Jezek et al. (Jezek et al., 2023).
Sucrose and to a lower extent maltose can induce the anthocyanin biosynthesis when added to the growth media of Arabidopsis seedlings on agar plates (Teng et al., 2005; Solfanelli et al., 2006). Signals indicating the increased sugar availability or the sugar availability itself could lead to anthocyanin accumulation (Kranz et al., 1998; Weiss, 2000; Gollop et al., 2001; Lloyd & Zakhleniuk, 2004). There is no evidence for an osmotic effect of sucrose to explain this observation (Solfanelli et al., 2006). Instead, it appears that multiple signaling pathways are integrated, ultimately resulting in an activation of the anthocyanin biosynthesis. Previous studies investigated the impact of sucrose on the general flavonoid biosynthesis genes and the anthocyanin biosynthesis genes DFR, ANS, and UF3GT (Solfanelli et al., 2006; Loreti et al., 2008; Li et al., 2014). Additional investigations of the recently reported arGST (Eichenberger et al., 2023) and various anthocyanin transporters (Pucker & Selmar, 2022) would be important in the future and might help to understand the whole picture. While studies often report the transcriptional up-regulation of individual structural genes in the anthocyanin biosynthesis, it seems more plausible that this is the consequence of transcription factor activation. Given the possibility for extensive decoration with sugars, anthocyanins could represent sugar sinks that would delay the onset of a sugar-promoted leaf senescence by preventing excessive sugar levels (Waterman et al., 1984; Lo Piccolo et al., 2018). This aligns with the finding that anthocyanin accumulation during high light acclimation appears to depend mostly on the increased cellular sugar content (Zirngibl et al., 2023) and the importance of arogenate-derived phenylalanine levels for anthocyanin accumulation (Chen et al., 2016). Although stress-induced anthocyanins do not degrade immediately after the stress conditions are lifted (Kumar & Sharma, 1999), it is possible that individual sugar moieties might be released. However, based on data from Elatostema rugosum (Neill et al., 2002) and Zea mays (Mollier & Pellerin, 1999) the amount of carbon stored in anthocyanins was reported as insufficient to account for all accumulating sugar (Henry et al., 2012). A more comprehensive database could help to unravel the predominant function of anthocyanins and potential differences between plant species. SUC1 was proposed as a potential integrator of light, sugar, and ethylene signals in the control of the anthocyanin biosynthesis (Sivitz et al., 2008; Jeong et al., 2010) with ethylene having a negative influence on anthocyanin formation (Jeong et al., 2010; Meng et al., 2018). However, a more recent study suggested an intracellular sucrose detection following a SUC1-mediated import as part of the sucrose-induced anthocyanin formation (Lasin et al., 2020). PAP1 expression is up-regulated in response to sucrose treatments, while other sugars do not have the same effect (Kranz et al., 1998; Teng et al., 2005; Broeckling et al., 2016). The circadian rhythms of the PAP1 expression and the close connection of its expression pattern to intracellular sugar levels suggest that PAP1 expression might be controlled in vivo by sugar levels (Harmer et al., 2000; Thimm et al., 2004; Solfanelli et al., 2006). PAP1 specifically up-regulates structural genes of the anthocyanin biosynthesis (Kranz et al., 1998) thus other transcription factors must be involved in activating upstream genes (Solfanelli et al., 2006). In contrast to the PAP1 activation, the expression of its partner TT8 was reported to be repressed in the presence of sucrose through a signaling pathway comprising YDA-EIN3/EIL1 (Meng et al., 2018). MYB30 was recently reported as an inhibitor of MYB75/PAP1 under low sucrose conditions (Zhou et al., 2024). The postulated regulatory model involves the ubiquitin E3 ligase RHA2b as a MYB75 target gene which triggers the degradation of MYB30 under high sucrose levels resulting in higher MYB5 activity and anthocyanin formation (Zhou et al., 2024). SnRK1 is activated by carbon- and energy-depleting stress and dissociates the MBW complex thus preventing the carbon-intensive biosynthesis of anthocyanins (Broucke et al., 2023). The MYB75 protein is degraded and TTG1 is exported from the nucleus (Broucke et al., 2023). An inhibition of the flavonol biosynthesis in Arabidopsis seedlings was observed in the presence of high sucrose levels (Solfanelli et al., 2006), which aligns with the competition mitigation through almost mutually exclusive gene expression of the first committed genes in the anthocyanin and flavonol biosynthesis, respectively (Choudhary & Pucker, 2024).
Anthocyanins increase the survival rate under nitrogen starvation (Liang & He, 2018). On the molecular level, the decoration patterns of anthocyanins, i.e. the addition of sugar moieties and other functional groups, are altered under nitrogen deficiency (Mark Hodges & Nozzolillo, 1996; Shi & Xie, 2010; Zhou et al., 2012). This could be caused through differences in the activation of structural genes through stress-specific transcription factors. MYB90/PAP2 appears as the dominant anthocyanin activator under nitrogen starvation (Lea et al., 2007; Peng et al., 2007), while otherwise MYB75/PAP1 is considered the dominant anthocyanin activator (Teng et al., 2005; Gonzalez et al., 2008). Translocation of nitrogen into younger tissues and parallel breakdown of chlorophyll causes an initial reddening of blades, veins, and petioles of old leaves (Koeslin-Findeklee et al., 2015). The pigmentation patterns caused by different nutritional imbalances can be distinct: while nitrogen starvation leads to anthocyanin accumulation along the veins, a phosphorus limitation would trigger a reddening between leaf veins (Barker & Pilbeam, 2015; Koeslin-Findeklee et al., 2015). Such differences in blush patterns representative for certain stress conditions might have the potential to be developed into biomarkers (Kalaji et al., 2018; Jezek et al., 2023). Of particular economic importance are nutritional imbalances influencing the accumulation of anthocyanins in fruits which can alter the commercial value depending on consumer preferences (Jezek et al., 2018).
Pollinator Attraction
Anthocyanins are well known for their contribution to flower colors with a wide range of colors including orange, red, magenta, purple, blue, and black (Winkel-Shirley, 2001; Wolff & Pucker, 2024). Flower color is a central visual signal for communication between plants and pollinators that should make the flower standout from the rest of the plant (Miller et al., 2011). Evolutionary changes in flower colors, e.g., blue to red, are often associated with pollinator changes as different pollinator groups have different color perception and resulting preferences (Rausher, 2008; Van der Niet et al., 2014; Gervasi & Schiestl, 2017; Trunschke et al., 2021). Insects, birds, reptiles, and bats have been reported as potential pollinators. Many pollinators can generalize their foraging to an array of different flower colors (Lunau & Maier, 1995).
A wholesale loss of anthocyanin pigmentation within a plant species or genus has been frequently observed (Marin-Recinos & Pucker, 2024) and might suggest a transition from animal to wind pollination or from color-attracted pollinators to nocturnal insects (Hoballah et al., 2007; Smith, 2010). However, some communication between plant and pollinator is not visible to humans with the naked eye. For example, flavonol patterns signal in the UV range and are thus only visible to animals with a compatible vision like certain insects (Sheehan et al., 2016). In rare cases, the flower color can change following a visitation or pollination event to redirect pollinators to other flowers (Weiss, 1991; Willmer et al., 2009; Ruxton & Schaefer, 2016; Garcia et al., 2022). However, flower color change can also be independent of a visitation/pollination event as observed for Fuchsia excorticata (Delph & Lively, 1989), Pulmonaria collina (Oberrath & Böhning-Gaese, 1999), Pedicularis monbeigiana (Sun et al., 2005), and Victoria cruziana (Nowak et al., 2024).
Seed Disperser Attraction
Many fruits and berries like apples, pears, blackberries, blueberries, and grapes can be intensely pigmented by anthocyanins (Albert et al., 2023). The fruit coloration can be a signal of ripe fruits and attract frugivorous birds by providing strong contrast to the background (Turcek, 1963; Stiles, 1982). This attraction of birds seems to be important for the seed dispersal (Ridley, 1930; Stiles, 1982). Additional fruit flags that support the attraction of seed dispersers have been described (Stiles, 1982). The accumulation of anthocyanins in fruits like apple and pear is induced by light and involves several characteristic WRKY transcription factors including MdWRKY75 (Su et al., 2022), MdWRKY11 (Liu et al., 2019c), PpWRKY26 (Li et al., 2020b), and PpWRKY44 (Alabd et al., 2022). HY5 is a central light-responsive regulator that activates the anthocyanin biosynthesis promoting MYB genes and genes of the anthocyanin biosynthesis directly (Shin et al., 2013; Tao et al., 2018). It has been reported that the proteins BBX16 and BBX18 are interacting with HY5 in this light-dependent activation of the anthocyanin biosynthesis in pear (Bai et al., 2019a,b). To the best of our knowledge, there are no conclusive experiments showing the evolutionary benefit of light-induced formation of anthocyanins in fruits yet. It appears plausible that this is a derived property from the regulation of the anthocyanins in leaves, where light responsive formation of protective anthocyanins has obvious advantages, and could suggest that anthocyanins have also a protective function in fruits.
Herbivore Repellence and Pathogen Resistances
Plants are under constant thread by a range of different herbivore and parasite attacks directed against different plant structures and exposed to pathogen infections (Figure 7). Herbivores might target canopy, low branches, or seedlings while parasites might extract sugar and nitrogen from leaf veins or eat the leaf tissue (Karban & Baldwin, 1997; Lev-Yadun & Gould, 2009). Anthocyanins can contribute to numerous physical and chemical defense strategies that evolved in plants. As anthocyanins are usually not toxic to animals (Lee et al., 1987; Costa-Arbulú et al., 2001), their role can be considered as minor compared to many other defense compounds (Padmavati et al., 1997; Werlein et al., 2005; Lev-Yadun & Gould, 2009). However, the biosynthesis of anthocyanins is tightly connected to the biosynthesis of proanthocyanidins, which have been reported as herbivore repellents (Barbehenn & Constabel, 2011). A study investigating tropical trees observed lower herbivore damage on plants with high anthocyanin and tannin content (Gong et al., 2020). Another study reported that heterologous expression of the anthocyanin promoting MYB PAP1 in cotton increased the resistance to bollworm and spider mite (Li et al., 2019b). In light of the current knowledge about the coloration functions of anthocyanins, it appears more plausible that the pigments are involved in some kind of visual defense rather than direct repellence. A number of hypotheses have been formulated to explain the role of anthocyanins in defense against herbivores and parasites (Lev-Yadun & Gould, 2009).
Aposematism is the bright coloration of an unpalatable/toxic or thorny plant to scare herbivores. Animals associate the color with an unpleasant experience which prevents or at least reduces future feeding attempts. This has been reported for multiple toxin-rich or unpalatable plant parts including fruits (Harborne, 1982; Lev-Yadun et al., 2009), seeds (Cook et al., 1971; Williamson, 1982), flowers (Gerchman et al., 2012; Lev-Yadun, 2024), and thorns (Lev-Yadun, 2001, 2016). Given that multiple independent events of aposematism have been described in the context of anthocyanin pigmentation, an evolutionary benefit is highly likely (Lev-Yadun & Gould, 2009). Since plants can recover better than animals from losing part of their structures, an adaptive value is supported by the direct advantage of an individual plant that survives the encounter with a herbivore and lowers the chances of future feeding attempts. This could explain why aposematism spread faster in plants than in animals (Lev-Yadun & Gould, 2009). There are also anthocyanin-pigmented species like the carnivorous Drosera spatulata (Ichiishi et al., 1999), which obviously do not accumulate pigments to deter insects. Nevertheless, deterrence of herbivores by pigmentation and other means has been proposed as an important first line of plant defense (Lev-Yadun, 2021).
Bright coloration could also appear as mimicry i.e. non-toxic species showing bright colors to be mistaken with toxic plants occurring in the same habitat. It was postulated that mimicry in sessile plants would be less effective than in animals, because herbivores have more time to assess an individual and feeding attempts would not be detrimental for the herbivore (Williamson, 1982). Nevertheless, anthocyanin pigmentation could also contribute to cryptic mimicry by letting a plant structure appear dead or in a senescing stage (Stone, 1979; Juniper, 1993). The pigmentation could also generate the impression of defensive structures like thorns that are not actually sharp (Lev-Yadun, 2003). Since ants are often attracted by plants to defend against herbivores, some plant species display dark spots arranged in a way to mimic a column of ants that would pose a threat to herbivores (Lev-Yadun & Inbar, 2002; Lev-Yadun & Gould, 2009). Although the pigmentation composition of spots has not been tested, it is believed that they are anthocyanins (Lev-Yadun & Gould, 2009). Other reports describe aphid mimicry that makes a plant look infested in order to discourage grazers and insects from targeting this plant (Lev-Yadun & Inbar, 2002). Immature pods of several legume species appear to mimic aposematic poisonous caterpillars due to their shape, size, and pigmentation pattern, which might protect these structures against herbivores and also provide an advantage to the entire plant (Lev-Yadun & Inbar, 2002). Multiple functions of anthocyanins turn experimental investigations into a challenge. While high anthocyanin content in some plants could be the result of aposematism or mimicry, it might just indicate stress in other plant species. A study in Acer platanoides observed the highest anthocyanin production and thus red leaves in partially dead trees with the lowest defensive value (Sinkkonen, 2008).
Camouflage of plant structures due to pigmentation, especially dark pigmentation, has been postulated as another function of anthocyanins (Givnish, 1990; Gould, 2004; Wolff & Pucker, 2024). Variegation in understory plants in forest has the potential to disrupt leaf outlines and might protect against colour-blind herbivores (Givnish, 1990). The co-occurence of anthocyanins and chlorophyll in seedlings of Pseudopanax crassifolius provides the plant with a brownish appearance that resembles the background colors (Gould, 1993). Anthocyanins could mask the bright green color of chlorophyll-rich leaves thus making them less attractive to certain herbivores (Lev-Yadun & Gould, 2009).
Bright anthocyanin pigmentation could serve as anti camouflage, i.e., breaking the crypsis of herbivorous insects and making them more visible to predators or parasitoids (Lev-Yadun et al., 2004; Lev-Yadun & Silva, 2006; Ide, 2022). Red color of young leaves has been explained as undermining the camouflage of herbivorous insects (Karageorgou et al., 2008; Ide, 2022). This has the potential to deter such herbivores as they might generally avoid surfaces of unsuitable color (Lev-Yadun & Gould, 2009; Ide, 2022). This could explain the color differences observed between the adaxial and abaxial side of leaves as well as all other plant structures, because herbivores adapted to the color of one leaf surface would be exposed on all other surfaces (Lev-Yadun et al., 2004). However, this hypothesis is somewhat weakened by a simulation that showed that a visually complex surface has higher potential of hiding insects (Schaefer & Rolshausen, 2006).
Anthocyanins might serve an attraction function in the pollination or seed dispersal, while serving as herbivore defense against other species in the same plant. In Hypericum calycinum, dearomatized isoprenylated phloroglucinols have an attraction function in the petals, but a repellent function in stamens and ovaries (Gronquist et al., 2001). It is also feasible that the function of anthocyanins in a plant changes over time (Lev-Yadun & Gould, 2009). Such synergistic effects could lead to evolutionary advantages thus making plants that use anthocyanins in different ways more likely (Armbruster, 2002).
Anthocyanins have been reported to contribute to resistances or at least tolerances against pathogens. Resistance against Colletotrichum gloeosporioides was reported for anthocyanin-rich mango fruits compared to green fruits (Sivankalyani et al., 2016). Heterologous expression of the anthocyanin activator LEAF COLOUR from maize in apple resulted in an increased anthocyanin and proanthocyanidin level (Flachowsky et al., 2010). A stronger resistance against bacterial infection (Erwinia amylovora) and fungal infection (Venturia inaequalis) was observed (Flachowsky et al., 2010). High concentrations of anthocyanins in purple tomatoes conferred higher tolerance against Botrytis cinerea through perturbation of the ROS burst during infection resulting in an extended shelf life (Zhang et al., 2013). The susceptibility of the anthocyanin deficient Arabidopsis thaliana tt4 mutant against Verticillium dahliae supports the importance of anthocyanins in defense against fungal infections (Lei et al., 2018). Anthocyanin-rich potatoes have been reported to show better performance against Pectobacterium carotovorum when compared to unpigmented potatoes (Wegener & Jansen, 2007). In summary, multiple studies reported positive effects of high anthocyanin levels against pathogens, but the molecular mechanisms remained sometimes unknown. While an interference with ROS signals associated with infections has been identified as an important and potential universal mechanism (Zhang et al., 2013), it remains feasible that the increased survival of anthocyanin-rich plants is also partially due to better stress resilience due to high levels of antioxidants or the simultaneously increased levels of proanthocyanidins.
Conclusions
Moving beyond the core of the flavonoid biosynthesis and understanding the full diversity of various anthocyanin decorating reactions is the next big challenge. Interspecific diversity can be explored, but could also pose a powerful resource to understand the evolutionary, biochemical, and ecological importance of specific anthocyanin modifications through comparative analyses. Given the wide range of different functions, it is crucial to understand the relative importance of anthocyanins in any given context. Due to the various interactions of anthocyanin functions, this task is similar to solving a system of differential equations. There are already numerous plausible hypotheses about the ecological functions of anthocyanins, but experimental validations on a broad taxonomic level are still missing. This relative importance of different anthocyanin functions is likely to vary between plant lineages thus adding another layer of complexity.
With the recent interest in the terrestrialization of plants and many genome sequencing efforts committed to early land plants, we can expect to gain a comprehensive understanding of the evolutionary trajectory that resulted in the anthocyanin biosynthesis. This will also help to understand in which order the anthocyanins acquired their various functions during evolution.
Transcriptional regulation is the central level for controlling anthocyanin accumulation and integrating environmental signals. Simultaneously, the transcriptional control poses an elegant system for metabolic engineering in plants. Purple tomatoes demonstrated the biotechnological potential and many genetic engineering and genome editing projects in crops and horticultural plants are going to follow. A detailed understanding of nuances in the anthocyanin biosynthesis enables the best selection of targets for such applications.
Author Contributions
N.G., J.M.H., and B.P. conceptualized this work, wrote the initial draft, and performed final editing. All authors have read and agreed to the published version of the manuscript.
Funding
Acknowledgments
Many thanks to all current and previous members of the Plant Biotechnology and Bioinformatics group for discussions. We acknowledge support by the Open Access Publication Funds of Technische Universität Braunschweig.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abid MA, Wei Y, Meng Z, Wang Y, Ye Y, Wang Y, He H, Zhou Q, Li Y, Wang P, et al.2022. Increasing floral visitation and hybrid seed production mediated by beauty mark in Gossypium hirsutum. Plant Biotechnology Journal 20: 1274–1284. [CrossRef]
- Agati G, Brunetti C; M, Di Ferdinando; F, Ferrini; S, Pollastri; M, Tattini. Functional roles of flavonoids in photoprotection: new evidence, lessons from the past. Plant physiology and biochemistry: PPB 2013, 72, 35–45. [Google Scholar] [CrossRef]
- Aharoni A, De Vos CHR; M, Wein; Z, Sun; R, Greco; A, Kroon; JNM, Mol; AP, O’Connell. The strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in transgenic tobacco. The Plant Journal 2001, 28, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Alabd A, Ahmad M; X, Zhang; Y, Gao; L, Peng; L, Zhang; J, Ni; S, Bai; Y, Teng. Light-responsive transcription factor PpWRKY44 induces anthocyanin accumulation by regulating PpMYB10 expression in pear. Horticulture Research 9: uhac199, 2022. [Google Scholar] [CrossRef]
- Albert NW, Davies KM; DH, Lewis; H, Zhang; M, Montefiori; C, Brendolise; MR, Boase; H, Ngo; PE, Jameson; KE, Schwinn. A conserved network of transcriptional activators and repressors regulates anthocyanin pigmentation in eudicots. The Plant Cell 2014a, 26, 962–980. [Google Scholar] [CrossRef] [PubMed]
- Albert NW, Davies KM; KE, Schwinn. Gene regulation networks generate diverse pigmentation patterns in plants. Plant Signaling & Behavior 2014b, 9, e29526. [Google Scholar] [CrossRef]
- Albert NW, Iorizzo M, Mengist MF, Montanari S, Zalapa J, Maule A, Edger PP, Yocca AE, Platts AE, Pucker B, et al.2023. Vaccinium as a comparative system for understanding of complex flavonoid accumulation profiles and regulation in fruit. Plant Physiology 192: 1696–1710. [CrossRef]
- Albert NW, Lewis DH; H, Zhang; LJ, Irving; PE, Jameson; KM, Davies. Light-induced vegetative anthocyanin pigmentation in Petunia. Journal of Experimental Botany 2009, 60, 2191–2202. [Google Scholar] [CrossRef]
- An J-P, Qu F-J; J-F, Yao; X-N, Wang; C-X, You; X-F, Wang; Y-J, Hao. The bZIP transcription factor MdHY5 regulates anthocyanin accumulation and nitrate assimilation in apple. Horticulture Research 2017, 4, 17023. [Google Scholar] [CrossRef]
- An J-P, Zhang X-W; C-X, You; S-Q, Bi; X-F, Wang; Y-J, Hao. MdWRKY40 promotes wounding-induced anthocyanin biosynthesis in association with MdMYB1 and undergoes MdBT2-mediated degradation. New Phytologist 2019, 224, 380–395. [Google Scholar] [CrossRef]
- Andersen TB, Hansen NB; T, Laursen; C, Weitzel; HT, Simonsen. Evolution of NADPH-cytochrome P450 oxidoreductases (POR) in Apiales – POR 1 is missing. Molecular Phylogenetics and Evolution 2016, 98, 21–28. [Google Scholar] [CrossRef]
- Andersen ØM, Jordheim M. The Anthocyanins. In: Flavonoids: Chemistry, Biochemistry and Applications; 2006. [Google Scholar]
- Appelhagen I, Nordholt N; T, Seidel; K, Spelt; R, Koes; F, Quattrochio; M, Sagasser; B, Weisshaar. TRANSPARENT TESTA 13 is a tonoplast P3A-ATPase required for vacuolar deposition of proanthocyanidins in Arabidopsis thaliana seeds. The Plant Journal 2015, 82, 840–849. [Google Scholar] [CrossRef]
- Araguirang GE, Richter AS. Activation of anthocyanin biosynthesis in high light – what is the initial signal? New Phytologist 2022, 236, 2037–2043. [Google Scholar] [CrossRef]
- Archetti M. 2009. Phylogenetic analysis reveals a scattered distribution of autumn colours. Annals of Botany 103: 703–713. [CrossRef]
- Archetti M, Döring TF, Hagen SB, Hughes NM, Leather SR, Lee DW, Lev-Yadun S, Manetas Y, Ougham HJ, Schaberg PG, et al.2009. Unravelling the evolution of autumn colours: an interdisciplinary approach. Trends in Ecology & Evolution 24: 166–173. [CrossRef]
- Armbruster WS. 2002. Can indirect selection and genetic context contribute to trait diversification? A transition-probability study of blossom-colour evolution in two genera. Journal of Evolutionary Biology 15: 468–486. [CrossRef]
- Azuma A, Kobayashi S; H, Yakushui; M, Yamada; N, Mitani; A, Sato. VvmybA1 genotype determines grape skin color. VITIS - Journal of Grapevine Research 2007, 46, 154–154. [Google Scholar] [CrossRef]
- Bai S, Saito T; C, Honda; Y, Hatsuyama; A, Ito; T, Moriguchi. An apple B-box protein, MdCOL11, is involved in UV-B- and temperature-induced anthocyanin biosynthesis. Planta 2014, 240, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Bai S, Tao R, Tang Y, Yin L, Ma Y, Ni J, Yan X, Yang Q, Wu Z, Zeng Y, et al.2019a. BBX16, a B-box protein, positively regulates light-induced anthocyanin accumulation by activating MYB10 in red pear. Plant Biotechnology Journal 17: 1985–1997. [CrossRef]
- Bai S, Tao R; L, Yin; J, Ni; Q, Yang; X, Yan; F, Yang; X, Guo; H, Li; Y, Teng. Two B-box proteins, PpBBX18 and PpBBX21, antagonistically regulate anthocyanin biosynthesis via competitive association with Pyrus pyrifolia ELONGATED HYPOCOTYL 5 in the peel of pear fruit. The Plant Journal: For Cell and Molecular Biology 2019b, 100, 1208–1223. [Google Scholar] [CrossRef]
- Ban Z, Qin H; AJ, Mitchell; B, Liu; F, Zhang; J-K, Weng; RA, Dixon; G, Wang. Noncatalytic chalcone isomerase-fold proteins in Humulus lupulus are auxiliary components in prenylated flavonoid biosynthesis. Proceedings of the National Academy of Sciences of the United States of America 2018, 115, E5223–E5232. [Google Scholar] [CrossRef]
- Barbehenn RV, Constabel CP. Tannins in plant-herbivore interactions. Phytochemistry 2011, 72, 1551–1565. [Google Scholar] [CrossRef] [PubMed]
- Barker A, Pilbeam D. Handbook of plant nutrition. 2015. [Google Scholar]
- Behrens CE, Smith KE; CV, Iancu; J, Choe; JV, Dean. Transport of Anthocyanins and other Flavonoids by the Arabidopsis ATP-Binding Cassette Transporter AtABCC2. Scientific Reports 2019, 9, 437. [Google Scholar] [CrossRef]
- Bienert GP, Schjoerring JK; TP, Jahn. Membrane transport of hydrogen peroxide. Biochimica Et Biophysica Acta 2006, 1758, 994–1003. [Google Scholar] [CrossRef]
- Bloor SJ, Abrahams S. The structure of the major anthocyanin in Arabidopsis thaliana. Phytochemistry 2002, 59, 343–346. [Google Scholar] [CrossRef]
- Borevitz JO, Xia Y; J, Blount; RA, Dixon; C, Lamb. Activation Tagging Identifies a Conserved MYB Regulator of Phenylpropanoid Biosynthesis. The Plant Cell 2000, 12, 2383–2393. [Google Scholar] [CrossRef]
- Broeckling BE, Watson RA; B, Steinwand; DR, Bush. Intronic Sequence Regulates Sugar-Dependent Expression of Arabidopsis thaliana Production of Anthocyanin Pigment-1/MYB75. PLOS ONE 2016, 11, e0156673. [Google Scholar] [CrossRef]
- Broucke E, Dang TTV; Y, Li; S, Hulsmans; J, Van Leene; G, De Jaeger; I, Hwang; E, Wim V den; F, Rolland. SnRK1 inhibits anthocyanin biosynthesis through both transcriptional regulation and direct phosphorylation and dissociation of the MYB/bHLH/TTG1 MBW complex. The Plant Journal 2023, 115, 1193–1213. [Google Scholar] [CrossRef] [PubMed]
- Brugliera F, Holton TA; TW, Stevenson; E, Farcy; C-Y, Lu; EC, Cornish. Isolation and characterization of a cDNA clone corresponding to the Rt locus of Petunia hybrida. The Plant Journal 1994, 5, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Burger J, Edwards GE. Photosynthetic Efficiency, and Photodamage by UV and Visible Radiation, in Red versus Green Leaf Coleus Varieties. Plant and Cell Physiology 1996, 37, 395–399. [Google Scholar] [CrossRef]
- Busche M, Acatay C; S, Martens; B, Weisshaar; R, Stracke. Functional Characterisation of Banana (Musa spp.) 2-Oxoglutarate-Dependent Dioxygenases Involved in Flavonoid Biosynthesis. Frontiers in Plant Science 12, 2021. [Google Scholar] [CrossRef]
- Busche M, Pucker B; B, Weisshaar; R, Stracke. Three R2R3-MYB transcription factors from banana (Musa acuminata) activate structural anthocyanin biosynthesis genes as part of an MBW complex. BMC Research Notes 2023, 16, 103. [Google Scholar] [CrossRef]
- Cardi T, Murovec J, Bakhsh A, Boniecka J, Bruegmann T, Bull SE, Eeckhaut T, Fladung M, Galovic V, Linkiewicz A, et al.2023. CRISPR/Cas-mediated plant genome editing: outstanding challenges a decade after implementation. Trends in Plant Science 28: 1144–1165. [CrossRef]
- Cavallini E, Matus JT; L, Finezzo; S, Zenoni; R, Loyola; F, Guzzo; R, Schlechter; A, Ageorges; P, Arce-Johnson; GB, Tornielli. The Phenylpropanoid Pathway Is Controlled at Different Branches by a Set of R2R3-MYB C2 Repressors in Grapevine. Plant Physiology 2015, 167, 1448–1470. [Google Scholar] [CrossRef]
- Cavallini E, Zenoni S; L, Finezzo; F, Guzzo; A, Zamboni; L, Avesani; GB, Tornielli. Functional Diversification of Grapevine MYB5a and MYB5b in the Control of Flavonoid Biosynthesis in a Petunia Anthocyanin Regulatory Mutant. Plant and Cell Physiology 2014, 55, 517–534. [Google Scholar] [CrossRef]
- Chalker-Scott L. 1999. Environmental Significance of Anthocyanins in Plant Stress Responses. Photochemistry and Photobiology 70: 1–9. [CrossRef]
- Chandler VL, Radicella JP; TP, Robbins; J, Chen; D, Turks. Two regulatory genes of the maize anthocyanin pathway are homologous: isolation of B utilizing R genomic sequences. The Plant Cell 1989, 1, 1175–1183. [Google Scholar] [CrossRef]
- Chang CJ, Li Y-H, Chen L-T, Chen W-C, Hsieh W-P, Shin J, Jane W-N, Chou S-J, Choi G, Hu J-M, et al.2008. LZF1, a HY5-regulated transcriptional factor, functions in Arabidopsis de-etiolation. The Plant Journal: For Cell and Molecular Biology 54: 205–219. [CrossRef]
- Chen Y, Huang Z; L, Tang. Invisible red in young leaves: Anthocyanin likely plays a defensive role in some other way beyond visual warning. Flora 2021, 280, 151833. [Google Scholar] [CrossRef]
- Chen Q, Man C; D, Li; H, Tan; Y, Xie; J, Huang. Arogenate Dehydratase Isoforms Differentially Regulate Anthocyanin Biosynthesis in Arabidopsis thaliana. Molecular Plant 2016, 9, 1609–1619. [Google Scholar] [CrossRef]
- Chialva C, Blein T; M, Crespi; D, Lijavetzky. Insights into long non-coding RNA regulation of anthocyanin carrot root pigmentation. Scientific Reports 2021, 11, 4093. [Google Scholar] [CrossRef]
- Chopy M, Cavallini-Speisser Q; P, Chambrier; P, Morel; J, Just; V, Hugouvieux; S, Rodrigues Bento; C, Zubieta; M, Vandenbussche; M, Monniaux. Cell layer–specific expression of the homeotic MADS-box transcription factor PhDEF contributes to modular petal morphogenesis in petunia. The Plant Cell 2024, 36, 324–345. [Google Scholar] [CrossRef] [PubMed]
- Choudhary N, Pucker B. Conserved amino acid residues and gene expression patterns associated with the substrate preferences of the competing enzymes FLS and DFR. : 2023.11.05.565693; 2023, 2024. [Google Scholar] [CrossRef]
- Christie PJ, Alfenito MR; V, Walbot. Impact of low-temperature stress on general phenylpropanoid and anthocyanin pathways: Enhancement of transcript abundance and anthocyanin pigmentation in maize seedlings. Planta 1994, 194, 541–549. [Google Scholar] [CrossRef]
- Cone KC, Burr FA; B, Burr. Molecular analysis of the maize anthocyanin regulatory locus C1. Proceedings of the National Academy of Sciences of the United States of America 1986, 83, 9631–9635. [Google Scholar] [CrossRef]
- Cook AD, Atsatt PR; CA, Simon. Doves and Dove Weed: Multiple Defenses against Avian Predation. BioScience 1971, 21, 277–281. [Google Scholar] [CrossRef]
- Costa-Arbulú C, Gianoli E; WL, Gonzáles; HM, Niemeyer. Feeding by the aphid Sipha flava produces a reddish spot on leaves of Sorghum halepense: an induced defense? Journal of Chemical Ecology 2001, 27, 273–283. [Google Scholar] [CrossRef]
- Davies KM, Albert NW; KE, Schwinn; KM, Davies; NW, Albert; KE, Schwinn. From landing lights to mimicry: the molecular regulation of flower colouration and mechanisms for pigmentation patterning. Functional Plant Biology 2012, 39, 619–638. [Google Scholar] [CrossRef] [PubMed]
- Del Valle JC, Alcalde-Eon C; MT, Escribano-Bailón; ML, Buide; JB, Whittall; E, Narbona. Stability of petal color polymorphism: the significance of anthocyanin accumulation in photosynthetic tissues. BMC Plant Biology 2019, 19, 496. [Google Scholar] [CrossRef]
- Delph LF, Lively CM. THE EVOLUTION OF FLORAL COLOR CHANGE: POLLINATOR ATTRACTION VERSUS PHYSIOLOGICAL CONSTRAINTS IN FUCHSIA EXCORTICATA. Evolution; International Journal of Organic Evolution 1989, 43, 1252–1262. [Google Scholar] [CrossRef]
- Deng G-M, Zhang S, Yang Q-S, Gao H-J, Sheng O, Bi F-C, Li C-Y, Dong T, Yi G-J, He W-D, et al.2021. MaMYB4, an R2R3-MYB Repressor Transcription Factor, Negatively Regulates the Biosynthesis of Anthocyanin in Banana. Frontiers in Plant Science 11.
- D’Hooghe P, Escamez S; J, Trouverie; J-C, Avice. Sulphur limitation provokes physiological and leaf proteome changes in oilseed rape that lead to perturbation of sulphur, carbon and oxidative metabolisms. BMC Plant Biology 2013, 13, 23. [Google Scholar] [CrossRef]
- Ding B, Patterson EL, Holalu SV, Li J, Johnson GA, Stanley LE, Greenlee AB, Peng F, Bradshaw HD, Blinov ML, et al.2020. Two MYB Proteins in a Self-Organizing Activator-Inhibitor System Produce Spotted Pigmentation Patterns. Current biology: CB 30: 802-814.e8. [CrossRef]
- Dodd IC, Critchley C; GS, Woodall; GR, Stewart. Photoinhibition in differently coloured juvenile leaves of Syzygium species. Journal of Experimental Botany 1998, 49, 1437–1445. [Google Scholar] [CrossRef]
- Dubos C, Le Gourrierec J; A, Baudry; G, Huep; E, Lanet; I, Debeaujon; J-M, Routaboul; A, Alboresi; B, Weisshaar; L, Lepiniec. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. The Plant Journal: For Cell and Molecular Biology 2008, 55, 940–953. [Google Scholar] [CrossRef]
- Eichenberger M, Schwander T; S, Hüppi; J, Kreuzer; PRE, Mittl; F, Peccati; G, Jiménez-Osés; M, Naesby; RM, Buller. The catalytic role of glutathione transferases in heterologous anthocyanin biosynthesis. Nature Catalysis 2023, 6, 927–938. [Google Scholar] [CrossRef]
- Fattorini R, Khojayori FN; G, Mellers; E, Moyroud; E, Herrero; RT, Kellenberger; R, Walker; Q, Wang; L, Hill; BJ, Glover. Complex petal spot formation in the Beetle Daisy (Gorteria diffusa) relies on spot-specific accumulation of malonylated anthocyanin regulated by paralogous GdMYBSG6 transcription factors. New Phytologist n/a, 2024. [Google Scholar] [CrossRef]
- Feild TS, Lee DW; NM, Holbrook. Why Leaves Turn Red in Autumn. The Role of Anthocyanins in Senescing Leaves of Red-Osier Dogwood. Plant Physiology 2001, 127, 566–574. [Google Scholar] [CrossRef]
- Feng K, Xu Z-S; J-X, Liu; J-W, Li; F, Wang; A-S, Xiong. Isolation, purification, and characterization of AgUCGalT1, a galactosyltransferase involved in anthocyanin galactosylation in purple celery (Apium graveolens L.). Planta 2018, 247, 1363–1375. [Google Scholar] [CrossRef]
- Flachowsky H, Szankowski I, Fischer TC, Richter K, Peil A, Höfer M, Dörschel C, Schmoock S, Gau AE, Halbwirth H, et al.2010. Transgenic apple plants overexpressing the Lc gene of maize show an altered growth habit and increased resistance to apple scab and fire blight. Planta 231: 623–635. [CrossRef]
- Ford CM, Boss PK; PB, Høj. Cloning and Characterization of Vitis viniferaUDP-Glucose:Flavonoid 3-O-Glucosyltransferase, a Homologue of the Enzyme Encoded by the Maize Bronze-1Locus That May Primarily Serve to Glucosylate Anthocyanidins in Vivo *. Journal of Biological Chemistry 1998, 273, 9224–9233. [Google Scholar] [CrossRef]
- Francisco RM, Regalado A, Ageorges A, Burla BJ, Bassin B, Eisenach C, Zarrouk O, Vialet S, Marlin T, Chaves MM, et al.2013. ABCC1, an ATP binding cassette protein from grape berry, transports anthocyanidin 3-O-Glucosides. The Plant Cell 25: 1840–1854. [CrossRef]
- Franco-Zorrilla JM, Valli A; M, Todesco; I, Mateos; MI, Puga; I, Rubio-Somoza; A, Leyva; D, Weigel; JA, García; J, Paz-Ares. Target mimicry provides a new mechanism for regulation of microRNA activity. Nature Genetics 2007, 39, 1033–1037. [Google Scholar] [CrossRef]
- Fraser CM, Thompson MG; AM, Shirley; J, Ralph; JA, Schoenherr; T, Sinlapadech; MC, Hall; C, Chapple. Related Arabidopsis Serine Carboxypeptidase-Like Sinapoylglucose Acyltransferases Display Distinct But Overlapping Substrate Specificities. Plant Physiology 2007, 144, 1986–1999. [Google Scholar] [CrossRef]
- Frommann J-F, Pucker B; LM, Sielmann; C, Müller; B, Weisshaar; R, Stracke; R, Schweiger. Metabolic fingerprinting reveals roles of Arabidopsis thaliana BGLU1, BGLU3 and BGLU4 in glycosylation of various flavonoids. : 2024.01.30.577901. [CrossRef]
- Frydman A, Liberman R; DV, Huhman; M, Carmeli-Weissberg; M, Sapir-Mir; R, Ophir; L, W. Sumner; Y, Eyal. The molecular and enzymatic basis of bitter/non-bitter flavor of citrus fruit: evolution of branch-forming rhamnosyltransferases under domestication. The Plant Journal 2013, 73, 166–178. [Google Scholar] [CrossRef]
- Frydman A, Weisshaus O; M, Bar-Peled; DV, Huhman; LW, Sumner; FR, Marin; E, Lewinsohn; R, Fluhr; J, Gressel; Y, Eyal. Citrus fruit bitter flavors: isolation and functional characterization of the gene Cm1,2RhaT encoding a 1,2 rhamnosyltransferase, a key enzyme in the biosynthesis of the bitter flavonoids of citrus. The Plant Journal: For Cell and Molecular Biology 2004, 40, 88–100. [Google Scholar] [CrossRef]
- Garcia JE, Hannah L; M, Shrestha; M, Burd; AG, Dyer. Fly pollination drives convergence of flower coloration. New Phytologist 2022, 233, 52–61. [Google Scholar] [CrossRef]
- Gebhardt YH, Witte S; H, Steuber; U, Matern; S, Martens. Evolution of Flavone Synthase I from Parsley Flavanone 3β-Hydroxylase by Site-Directed Mutagenesis. Plant Physiology 2007, 144, 1442–1454. [Google Scholar] [CrossRef]
- Gerchman Y, Dodek I; R, Petichov; Y, Yerushalmi; A, Lerner; T, Keasar. Beyond pollinator attraction: extra-floral displays deter herbivores in a Mediterranean annual plant. Evolutionary Ecology 2012, 26, 499–512. [Google Scholar] [CrossRef]
- Gervasi DDL, Schiestl FP. Real-time divergent evolution in plants driven by pollinators. Nature Communications 2017, 8, 14691. [Google Scholar] [CrossRef]
- Gierer A, Meinhardt H. A theory of biological pattern formation. Kybernetik 1972, 12, 30–39. [Google Scholar] [CrossRef]
- Givnish TJ. 1990. Leaf Mottling: Relation to Growth Form and Leaf Phenology and Possible Role as Camouflage. Functional Ecology 4: 463–474. [CrossRef]
- Glover BJ, Walker RH; E, Moyroud; SF, Brockington. How to spot a flower. New Phytologist 2013, 197, 687–689. [Google Scholar] [CrossRef]
- Gollop R, Farhi S; A, Perl. Regulation of the leucoanthocyanidin dioxygenase gene expression in Vitis vinifera. Plant Science 2001, 161, 579–588. [Google Scholar] [CrossRef]
- Gong W-C, Liu Y-H; C-M, Wang; Y-Q, Chen; K, Martin; L-Z, Meng. Why Are There so Many Plant Species That Transiently Flush Young Leaves Red in the Tropics? Frontiers in Plant Science 11, 2020. [Google Scholar] [CrossRef]
- Gonzalez A, Brown M, Hatlestad G, Akhavan N, Smith T, Hembd A, Moore J, Montes D, Mosley T, Resendez J, et al.2016. TTG2 controls the developmental regulation of seed coat tannins in Arabidopsis by regulating vacuolar transport steps in the proanthocyanidin pathway. Developmental Biology 419: 54–63. [CrossRef]
- Gonzalez A, Zhao M; JM, Leavitt; AM, Lloyd. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. The Plant Journal 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Goodman CD, Casati P; V, Walbot. A Multidrug Resistance–Associated Protein Involved in Anthocyanin Transport in Zea mays. The Plant Cell 2004, 16, 1812–1826. [Google Scholar] [CrossRef]
- Gou J-Y, Felippes FF; C-J, Liu; D, Weigel; J-W, Wang. Negative Regulation of Anthocyanin Biosynthesis in Arabidopsis by a miR156-Targeted SPL Transcription Factor. The Plant Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef]
- Gould KS. 1993. Leaf Heteroblasty in Pseudopanax crassifolius: Functional Significance of Leaf Morphology and Anatomy. Annals of Botany 71: 61–70. [CrossRef]
- Gould KS. 2004. Nature’s Swiss Army Knife: The Diverse Protective Roles of Anthocyanins in Leaves. Journal of Biomedicine and Biotechnology 2004: 314–320. [CrossRef]
- Gould KS, Dudle DA; HS, Neufeld. Why some stems are red: cauline anthocyanins shield photosystem II against high light stress. Journal of Experimental Botany 2010, 61, 2707–2717. [Google Scholar] [CrossRef]
- Gould KS, Jay-Allemand C; BA, Logan; Y, Baissac; LPR, Bidel. When are foliar anthocyanins useful to plants? Re-evaluation of the photoprotection hypothesis using Arabidopsis thaliana mutants that differ in anthocyanin accumulation. Environmental and Experimental Botany 2018, 154, 11–22. [Google Scholar] [CrossRef]
- Gould KS, Kuhn DN; DW, Lee; SF, Oberbauer. Why leaves are sometimes red. Nature 1995, 378, 241–242. [Google Scholar] [CrossRef]
- Gould KS, McKelvie J; KR, Markham. Do anthocyanins function as antioxidants in leaves? Imaging of H2O2 in red and green leaves after mechanical injury. Plant, Cell & Environment 2002a, 25, 1261–1269. [Google Scholar] [CrossRef]
- Gould KS, Neill SO; TC, Vogelmann. A unified explanation for anthocyanins in leaves? In: Advances in Botanical Research. Academic Press, 167–192; 2002b. [Google Scholar] [CrossRef]
- Gronquist M, Bezzerides A; A, Attygalle; J, Meinwald; M, Eisner; T, Eisner. Attractive and defensive functions of the ultraviolet pigments of a flower (Hypericum calycinum). Proceedings of the National Academy of Sciences of the United States of America 2001, 98, 13745–13750. [Google Scholar] [CrossRef]
- Grotewold E. 2006. The genetics and biochemistry of floral pigments. Annual Review of Plant Biology 57: 761–780. [CrossRef]
- Gu K-D, Wang C-K; D-G, Hu; Y-J, Hao. How do anthocyanins paint our horticultural products? Scientia Horticulturae 2019, 249, 257–262. [Google Scholar] [CrossRef]
- Hajiboland R, Farhanghi F. Remobilization of boron, photosynthesis, phenolic metabolism and anti-oxidant defense capacity in boron-deficient turnip (Brassica rapa L.) plants. Soil Science and Plant Nutrition 2010, 56, 427–437. [Google Scholar] [CrossRef]
- Harborne JB. 1962. Anthocyanins and their sugar components. Fortschritte Der Chemie Organischer Naturstoffe = Progress in the Chemistry of Organic Natural Products. Progres Dans La Chimie Des Substances Organiques Naturelles 20: 165–199. [CrossRef]
- Harborne JB. 1982. Introduction to Ecological Biochemistry.
- Harmer SL, Hogenesch JB; M, Straume; HS, Chang; B, Han; T, Zhu; X, Wang; JA, Kreps; SA, Kay. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science (New York, N.Y.) 2000, 290, 2110–2113. [Google Scholar] [CrossRef]
- Hellens RP, Moreau C, Lin-Wang K, Schwinn KE, Thomson SJ, Fiers MWEJ, Frew TJ, Murray SR, Hofer JMI, Jacobs JME, et al.2010. Identification of Mendel’s White Flower Character. PLOS ONE 5: e13230. [CrossRef]
- Henry A, Chopra S; DG, Clark; JP, Lynch. Responses to low phosphorus in high and low foliar anthocyanin coleus (Solenostemon scutellarioides) and maize (Zea mays). Functional Plant Biology 2012, 39, 255–265. [Google Scholar] [CrossRef]
- Hichri I, Barrieu F; J, Bogs; C, Kappel; S, Delrot; V, Lauvergeat. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. Journal of Experimental Botany 2011, 62, 2465–2483. [Google Scholar] [CrossRef]
- Ho WW, Smith SD. Molecular evolution of anthocyanin pigmentation genes following losses of flower color. BMC Evolutionary Biology 2016, 16, 98. [Google Scholar] [CrossRef]
- Hoballah ME, Gübitz T; J, Stuurman; L, Broger; M, Barone; T, Mandel; A, Dell’Olivo; M, Arnold; C, Kuhlemeier. Single Gene–Mediated Shift in Pollinator Attraction in Petunia. The Plant Cell 2007, 19, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Hoch WA, Zeldin EL; BH, McCown. Physiological significance of anthocyanins during autumnal leaf senescence. Tree Physiology 2001, 21, 1–8. [Google Scholar] [CrossRef]
- Hsu Y-H, Tagami T; K, Matsunaga; M, Okuyama; T, Suzuki; N, Noda; M, Suzuki; H, Shimura. Functional characterization of UDP-rhamnose-dependent rhamnosyltransferase involved in anthocyanin modification, a key enzyme determining blue coloration in Lobelia erinus. The Plant Journal: For Cell and Molecular Biology 2017, 89, 325–337. [Google Scholar] [CrossRef]
- Hughes NM, Connors MK; MH, Grace; MA, Lila; BN, Willans; AJ, Wommack. The same anthocyanins served four different ways: Insights into anthocyanin structure-function relationships from the wintergreen orchid, Tipularia discolor. Plant Science 2021, 303, 110793. [Google Scholar] [CrossRef]
- Hughes NM, Neufeld HS; KO, Burkey. Functional role of anthocyanins in high-light winter leaves of the evergreen herb Galax urceolata. New Phytologist 2005, 168, 575–587. [Google Scholar] [CrossRef]
- Hughes NM, Vogelmann TC; WK, Smith. Optical effects of abaxial anthocyanin on absorption of red wavelengths by understorey species: revisiting the back-scatter hypothesis. Journal of Experimental Botany 2008, 59, 3435–3442. [Google Scholar] [CrossRef]
- Ichiishi S, Nagamitsu T; Y, Kondo; T, Iwashina; K, Kondo; N, Tagashira. Effects of Macro-components and Sucrose in the Medium on in vitro Red-color Pigmentation in Dionaea muscipula Ellis and Drosera spathulata Laill. Plant Biotechnology 1999, 16, 235–238. [Google Scholar] [CrossRef]
- Ichino T, Fuji K, Ueda H, Takahashi H, Koumoto Y, Takagi J, Tamura K, Sasaki R, Aoki K, Shimada T, et al.2014. GFS9/TT9 contributes to intracellular membrane trafficking and flavonoid accumulation in Arabidopsis thaliana. The Plant Journal: For Cell and Molecular Biology 80: 410–423. [CrossRef]
- Ide J-Y. 2022. Why do red/purple young leaves suffer less insect herbivory: tests of the warning signal hypothesis and the undermining of insect camouflage hypothesis. Arthropod-Plant Interactions 16: 567–581. [CrossRef]
- Irani NG, Grotewold E. Light-induced morphological alteration in anthocyanin-accumulating vacuoles of maize cells. BMC Plant Biology 2005, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Ishihara H, Tohge T; P, Viehöver; AR, Fernie; B, Weisshaar; R, Stracke. Natural variation in flavonol accumulation in Arabidopsis is determined by the flavonol glucosyltransferase BGLU6. Journal of Experimental Botany 2016, 67, 1505–1517. [Google Scholar] [CrossRef]
- Jaakola L. 2013. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends in Plant Science 18: 477–483. [CrossRef]
- Jaakola L, Poole M, Jones MO, Kämäräinen-Karppinen T, Koskimäki JJ, Hohtola A, Häggman H, Fraser PD, Manning K, King GJ, et al.2010. A SQUAMOSA MADS Box Gene Involved in the Regulation of Anthocyanin Accumulation in Bilberry Fruits. Plant Physiology 153: 1619–1629. [CrossRef]
- Jeong S-W, Das PK, Jeoung SC, Song J-Y, Lee HK, Kim Y-K, Kim WJ, Park YI, Yoo S-D, Choi S-B, et al.2010. Ethylene Suppression of Sugar-Induced Anthocyanin Pigmentation in Arabidopsis1[C][W][OA]. Plant Physiology 154: 1514–1531. [CrossRef]
- Jezek M, Allan AC; JJ, Jones; C-M, Geilfus. Why do plants blush when they are hungry? New Phytologist 2023, 239, 494–505. [Google Scholar] [CrossRef]
- Jezek M, Zörb C; N, Merkt; C-M, Geilfus. Anthocyanin Management in Fruits by Fertilization. Journal of Agricultural and Food Chemistry 2018, 66, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Jiang X, Huang K, Zheng G, Hou H, Wang P, Jiang H, Zhao X, Li M, Zhang S, Liu Y, et al.2018. CsMYB5a and CsMYB5e from Camellia sinensis differentially regulate anthocyanin and proanthocyanidin biosynthesis. Plant Science 270: 209–220. [CrossRef]
- Jiang L, Yue M, Liu Y, Zhang N, Lin Y, Zhang Y, Wang Y, Li M, Luo Y, Zhang Y, et al.2023. A novel R2R3-MYB transcription factor FaMYB5 positively regulates anthocyanin and proanthocyanidin biosynthesis in cultivated strawberries (Fragaria × ananassa). Plant Biotechnology Journal 21: 1140–1158. [CrossRef]
- Jiu S, Guan L, Leng X, Zhang K, Haider MS, Yu X, Zhu X, Zheng T, Ge M, Wang C, et al.2021. The role of VvMYBA2r and VvMYBA2w alleles of the MYBA2 locus in the regulation of anthocyanin biosynthesis for molecular breeding of grape (Vitis spp.) skin coloration. Plant Biotechnology Journal 19: 1216–1239. [CrossRef]
- Jokioja J, Yang B; KM, Linderborg. Acylated anthocyanins: A review on their bioavailability and effects on postprandial carbohydrate metabolism and inflammation. Comprehensive Reviews in Food Science and Food Safety 2021, 20, 5570–5615. [Google Scholar] [CrossRef] [PubMed]
- Jonsson LMV, Aarsman MEG; J, van Diepen; P, de Vlaming; N, Smit; AW, Schram. Properties and genetic control of anthocyanin 5-O-glucosyltransferase in flowers of Petunia hybrida. Planta 1984, 160, 341–347. [Google Scholar] [CrossRef]
- Ju Z, Sun W; X, Meng; L, Liang; Y, Li; T, Zhou; H, Shen; X, Gao; L, Wang. Isolation and functional characterization of two 5-O-glucosyltransferases related to anthocyanin biosynthesis from Freesia hybrida. Plant Cell, Tissue and Organ Culture (PCTOC) 2018, 135, 99–110. [Google Scholar] [CrossRef]
- Juniper B. 1993. Flamboyant flushes: a reinterpretation of non-green flush colours in leaves. International dendrology society yearbook: 49–57.
- Kalaji HM, Bąba W, Gediga K, Goltsev V, Samborska IA, Cetner MD, Dimitrova S, Piszcz U, Bielecki K, Karmowska K, et al.2018. Chlorophyll fluorescence as a tool for nutrient status identification in rapeseed plants. Photosynthesis Research 136: 329–343. [CrossRef]
- Kamsteeg J, Brederode J van; van, Nigtevecht G. Identification, Properties and Genetic Control of UDP-ʟ-Rhamnose: Anthocyanidin 3-O-Glucoside, 6″-O-Rhamnosyltransferase Isolated from Retals of the Red Campion (Silene dioica). Zeitschrift für Naturforschung C 1980, 35, 249–257. [Google Scholar] [CrossRef]
- Kang X, Mikami R; Y, Akita. Characterization of 5-O-glucosyltransferase involved in anthocyanin biosynthesis in Cyclamen purpurascens. Plant Biotechnology 2021, 38, 263–268. [Google Scholar] [CrossRef]
- Karageorgou P, Buschmann C; Y, Manetas. Red leaf color as a warning signal against insect herbivory: Honest or mimetic? Flora - Morphology, Distribution, Functional Ecology of Plants 2008, 203, 648–652. [Google Scholar] [CrossRef]
- Karageorgou P, Manetas Y. The importance of being red when young: anthocyanins and the protection of young leaves of Quercus coccifera from insect herbivory and excess light. Tree Physiology 2006, 26, 613–621. [Google Scholar] [CrossRef]
- Karban R, Baldwin IT. Induced Responses to Herbivory. University of Chicago Press. 1997. [Google Scholar]
- Karppinen K, Lafferty DJ; NW, Albert; N, Mikkola; T, McGhie; AC, Allan; BM, Afzal; H, Häggman; RV, Espley; L, Jaakola. MYBA and MYBPA transcription factors co-regulate anthocyanin biosynthesis in blue-coloured berries. New Phytologist 2021, 232, 1350–1367. [Google Scholar] [CrossRef]
- Kaur S, Kumari A; N, Sharma; AK, Pandey; M, Garg. Physiological and molecular response of colored wheat seedlings against phosphate deficiency is linked to accumulation of distinct anthocyanins. Plant physiology and biochemistry: PPB 2022, 170, 338–349. [Google Scholar] [CrossRef]
- Kawai Y, Ono E; M, Mizutani. Evolution and diversity of the 2–oxoglutarate-dependent dioxygenase superfamily in plants. The Plant Journal 2014, 78, 328–343. [Google Scholar] [CrossRef]
- Kim S, Chen J, Cheng T, Gindulyte A, He J, He S, Li Q, Shoemaker BA, Thiessen PA, Yu B, et al.2023. PubChem 2023 update. Nucleic Acids Research 51: D1373–D1380. [CrossRef]
- Kim S, Hwang G; S, Lee; J-Y, Zhu; I, Paik; TT, Nguyen; J, Kim; E, Oh. High Ambient Temperature Represses Anthocyanin Biosynthesis through Degradation of HY5. Frontiers in Plant Science 2017, 8, 1787. [Google Scholar] [CrossRef]
- Kim D-H, Park S; J-Y, Lee; S-H, Ha; J-G, Lee; S-H, Lim. A Rice B-Box Protein, OsBBX14, Finely Regulates Anthocyanin Biosynthesis in Rice. International Journal of Molecular Sciences 2018, 19, 2190. [Google Scholar] [CrossRef]
- Kitamura S, Shikazono N; A, Tanaka. TRANSPARENT TESTA 19 is involved in the accumulation of both anthocyanins and proanthocyanidins in Arabidopsis. The Plant Journal: For Cell and Molecular Biology 2004, 37, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi NI, Saito T; N, Iwata; Y, Ohmae; R, Iwata; K, Tanoi; TM, Nakanishi. Leaf senescence in rice due to magnesium deficiency mediated defect in transpiration rate before sugar accumulation and chlorosis. Physiologia Plantarum 2013, 148, 490–501. [Google Scholar] [CrossRef]
- Koeslin-Findeklee F, Rizi VS; MA, Becker; S, Parra-Londono; M, Arif; S, Balazadeh; B, Mueller-Roeber; R, Kunze; WJ, Horst. Transcriptomic analysis of nitrogen starvation- and cultivar-specific leaf senescence in winter oilseed rape (Brassica napus L.). Plant Science: An International Journal of Experimental Plant Biology 2015, 233, 174–185. [Google Scholar] [CrossRef]
- Koseki M, Goto K; C, Masuta; A, Kanazawa. The Star-type Color Pattern in Petunia hybrida ‘Red Star’ Flowers is Induced by Sequence-Specific Degradation of Chalcone Synthase RNA. Plant and Cell Physiology 2005, 46, 1879–1883. [Google Scholar] [CrossRef] [PubMed]
- Kovinich N, Kayanja G; A, Chanoca; K, Riedl; MS, Otegui; E, Grotewold. Not all anthocyanins are born equal: distinct patterns induced by stress in Arabidopsis. Planta 2014, 240, 931–940. [Google Scholar] [CrossRef]
- Kovinich N, Saleem A; JT, Arnason; B, Miki. Functional characterization of a UDP-glucose:flavonoid 3-O-glucosyltransferase from the seed coat of black soybean (Glycine max (L.) Merr.). Phytochemistry 2010, 71, 1253–1263. [Google Scholar] [CrossRef]
- Kranz HD, Denekamp M, Greco R, Jin H, Leyva A, Meissner RC, Petroni K, Urzainqui A, Bevan M, Martin C, et al.1998. Towards functional characterisation of the members of the R2R3-MYB gene family from Arabidopsis thaliana. The Plant Journal: For Cell and Molecular Biology 16: 263–276. [CrossRef]
- Krause GH, Virgo A; K, Winter. High susceptibility to photoinhibition of young leaves of tropical forest trees. Planta 1995, 197, 583–591. [Google Scholar] [CrossRef]
- Kroon J, Souer E; A, De Graaff; Y, Xue; J, Mol; R, Koes. Cloning and structural analysis of the anthocyanin pigmentation locus Rt of Petunia hybrida: characterization of insertion sequences in two mutant alleles. The Plant Journal 1994, 5, 69–80. [Google Scholar] [CrossRef]
- Kumar V, Sharma SS. Nutrient Deficiency-dependent Anthocyanin Development in Spirodela Polyrhiza L. Schleid. Biologia Plantarum 1999, 42, 621–624. [Google Scholar] [CrossRef]
- Kytridis V-P, Manetas Y. Mesophyll versus epidermal anthocyanins as potential in vivo antioxidants: evidence linking the putative antioxidant role to the proximity of oxy-radical source. Journal of Experimental Botany 2006, 57, 2203–2210. [Google Scholar] [CrossRef] [PubMed]
- LaFountain AM, Yuan Y-W. Repressors of anthocyanin biosynthesis. New Phytologist 2021, 231, 933–949. [Google Scholar] [CrossRef]
- Landi M, Tattini M; KS, Gould. Multiple functional roles of anthocyanins in plant-environment interactions. Environmental and Experimental Botany 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Lasin P, Weise A; A, Reinders; JM, Ward. Arabidopsis Sucrose Transporter AtSuc1 introns act as strong enhancers of expression. Plant & Cell Physiology 2020, 61, 1054–1063. [Google Scholar] [CrossRef]
- Lawanson AO, Akindele BB; PB, Fasalojo; BL, Akpe. Time-course of anthocyanin formation during deficiencies of nitrogen, phosphorus and potassium in seedlings of zea mays Linn. var. E.S. 1. Zeitschrift für Pflanzenphysiologie 1972, 66, 251–253. [Google Scholar] [CrossRef]
- Lea US, Slimestad R; P, Smedvig; C, Lillo. Nitrogen deficiency enhances expression of specific MYB and bHLH transcription factors and accumulation of end products in the flavonoid pathway. Planta 2007, 225, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Lee DW. 2002. Anthocyanins in autumn leaf senescence. In: Advances in Botanical Research. Academic Press, 147–165. [CrossRef]
- Lee DW, Brammeier S; AP, Smith. The Selective Advantages of Anthocyanins in Developing Leaves of Mango and Cacao. Biotropica 1987, 19, 40–49. [Google Scholar] [CrossRef]
- Lee DW, Collins TM. Phylogenetic and Ontogenetic Influences on the Distribution of Anthocyanins and Betacyanins in Leaves of Tropical Plants. International Journal of Plant Sciences 2001, 162, 1141–1153. [Google Scholar] [CrossRef]
- Lee DW, O’Keefe J; NM, Holbrook; TS, Feild. Pigment dynamics and autumn leaf senescence in a New England deciduous forest, eastern USA. Ecological Research 2003, 18, 677–694. [Google Scholar] [CrossRef]
- Lei KJ, Zhang L; XY, Du; Y, An; GH, Chang; GY, An. A chalcone synthase controls the verticillium disease resistance response in both Arabidopsis thaliana and cotton. European Journal of Plant Pathology 2018, 152, 769–781. [Google Scholar] [CrossRef]
- Lei KJ, Zhou H; DL, Gu; GY, An. The involvement of abscisic acid-insensitive mutants in low phosphate stress responses during rhizosphere acidification, anthocyanin accumulation and Pi homeostasis in Arabidopsis. Plant Science: An International Journal of Experimental Plant Biology 2022, 322, 111358. [Google Scholar] [CrossRef]
- Lev-Yadun S. 2001. Aposematic (warning) Coloration Associated with Thorns in Higher Plants. Journal of Theoretical Biology 210: 385–388. [CrossRef]
- Lev-Yadun S. 2003. Why do some thorny plants resemble green zebras? Journal of Theoretical Biology 224: 483–489. [CrossRef]
- Lev-Yadun S. 2016. Biochemical Evidence of Convergent Evolution of Aposematic Coloration in Thorny, Spiny and Prickly Plants. In: Lev-Yadun S, ed. Defensive (anti-herbivory) Coloration in Land Plants. Cham: Springer International Publishing, 183–183. [CrossRef]
- Lev-Yadun S. 2021. Avoiding rather than resisting herbivore attacks is often the first line of plant defence. Biological Journal of the Linnean Society 134: 775–802. [CrossRef]
- Lev-Yadun S. 2024. Visual-, Olfactory-, and Nectar-Taste-Based Flower Aposematism. Plants 13: 391. [CrossRef]
- Lev-Yadun S, Dafni A; MA, Flaishman; M, Inbar; I, Izhaki; G, Katzir; G, Ne’eman. Plant coloration undermines herbivorous insect camouflage. BioEssays: News and Reviews in Molecular, Cellular and Developmental Biology 2004, 26, 1126–1130. [Google Scholar] [CrossRef] [PubMed]
- Lev-Yadun S, Gould KS. Role of Anthocyanins in Plant Defence. In: Winefield C, Davies K, Gould K, eds. Anthocyanins: Biosynthesis, Functions, and Applications. New York, NY: Springer, 22–28. [CrossRef]
- Lev-Yadun S, Inbar M. Defensive ant, aphid and caterpillar mimicry in plants? Biological Journal of the Linnean Society 2002, 77, 393–398. [Google Scholar] [CrossRef]
- Lev-Yadun S, Ne’eman G; I, Izhaki. Unripe red fruits may be aposematic. Plant Signaling & Behavior 2009, 4, 836–841. [Google Scholar] [CrossRef]
- Lev-Yadun S, Silva JA. Defensive coloration in plants: a review of current ideas about anti-herbivore coloration strategies. 2006. [Google Scholar]
- Li S, Dong Y; D, Li; S, Shi; N, Zhao; J, Liao; Y, Liu; H, Chen. Eggplant transcription factor SmMYB5 integrates jasmonate and light signaling during anthocyanin biosynthesis. Plant Physiology 2024, 194, 1139–1165. [Google Scholar] [CrossRef] [PubMed]
- Li B, Fan R, Guo S, Wang P, Zhu X, Fan Y, Chen Y, He K, Kumar A, Shi J, et al.2019a. The Arabidopsis MYB transcription factor, MYB111 modulates salt responses by regulating flavonoid biosynthesis. Environmental and Experimental Botany 166: 103807. [CrossRef]
- Li Y, Kong F; Z, Liu; L, Peng; Q, Shu. PhUGT78A22, a novel glycosyltransferase in Paeonia ‘He Xie’, can catalyze the transfer of glucose to glucosylated anthocyanins during petal blotch formation. BMC Plant Biology 2022, 22, 405. [Google Scholar] [CrossRef]
- Li P, Li Y-J; F-J, Zhang; G-Z, Zhang; X-Y, Jiang; H-M, Yu; B-K, Hou. The Arabidopsis UDP-glycosyltransferases UGT79B2 and UGT79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. The Plant Journal 2017, 89, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Li D-D, Ni R; P-P, Wang; X-S, Zhang; P-Y, Wang; T-T, Zhu; C-J, Sun; C-J, Liu; H-X, Lou; A-X, Cheng. Molecular Basis for Chemical Evolution of Flavones to Flavonols and Anthocyanins in Land Plants. Plant Physiology 2020a, 184, 1731–1743. [Google Scholar] [CrossRef]
- Li X, Ouyang X, Zhang Z, He L, Wang Y, Li Y, Zhao J, Chen Z, Wang C, Ding L, et al.2019b. Over-expression of the red plant gene R1 enhances anthocyanin production and resistance to bollworm and spider mite in cotton. Molecular Genetics and Genomics 294: 469–478. [CrossRef]
- Li Y, Shan X; L, Zhou; R, Gao; S, Yang; S, Wang; L, Wang; X, Gao. The R2R3-MYB Factor FhMYB5 From Freesia hybrida Contributes to the Regulation of Anthocyanin and Proanthocyanidin Biosynthesis. Frontiers in Plant Science 9, 2019c. [Google Scholar]
- Li Y, Van den Ende W; F, Rolland. Sucrose induction of anthocyanin biosynthesis is mediated by DELLA. Molecular Plant 2014, 7, 570–572. [Google Scholar] [CrossRef]
- Li C, Wu J; K-D, Hu; S-W, Wei; H-Y, Sun; L-Y, Hu; Z, Han; G-F, Yao; H, Zhang. PyWRKY26 and PybHLH3 cotargeted the PyMYB114 promoter to regulate anthocyanin biosynthesis and transport in red-skinned pears. Horticulture Research 2020b, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liang J, He J. Protective role of anthocyanins in plants under low nitrogen stress. Biochemical and Biophysical Research Communications 2018, 498, 946–953. [Google Scholar] [CrossRef]
- Liu Y, Liu J; Y, Qi; A, Zhang; Z, Liu; X, Ren. Identification and characterization of AcUFGT6b, a xylosyltransferase involved in anthocyanin modification in red-fleshed kiwifruit (Actinidia chinensis). Plant Cell, Tissue and Organ Culture (PCTOC) 2019a, 138, 257–271. [Google Scholar] [CrossRef]
- Liu H, Shu Q; K, Lin-Wang; AC, Allan; RV, Espley; J, Su; M, Pei; J, Wu. The PyPIF5-PymiR156a-PySPL9-PyMYB114/MYB10 module regulates light-induced anthocyanin biosynthesis in red pear. Molecular Horticulture 2021, 1, 14. [Google Scholar] [CrossRef]
- Liu H, Su J; Y, Zhu; G, Yao; AC, Allan; C, Ampomah-Dwamena; Q, Shu; K, Lin-Wang; S, Zhang; J, Wu. The involvement of PybZIPa in light-induced anthocyanin accumulation via the activation of PyUFGT through binding to tandem G-boxes in its promoter. Horticulture Research 2019b, 6, 134. [Google Scholar] [CrossRef] [PubMed]
- Liu W, Wang Y, Yu L, Jiang H, Guo Z, Xu H, Jiang S, Fang H, Zhang J, Su M, et al.2019c. MdWRKY11 Participates in Anthocyanin Accumulation in Red-Fleshed Apples by Affecting MYB Transcription Factors and the Photoresponse Factor MdHY5. Journal of Agricultural and Food Chemistry 67: 8783–8793. [CrossRef]
- Lloyd A, Brockman A; L, Aguirre; A, Campbell; A, Bean; A, Cantero; A, Gonzalez. Advances in the MYB–bHLH–WD Repeat (MBW) Pigment Regulatory Model: Addition of a WRKY Factor and Co-option of an Anthocyanin MYB for Betalain Regulation. Plant and Cell Physiology 2017, 58, 1431–1441. [Google Scholar] [CrossRef]
- Lloyd AM, Walbot V; RW, Davis. Arabidopsis and Nicotiana Anthocyanin Production Activated by Maize Regulators R and C1. Science 1992, 258, 1773–1775. [Google Scholar] [CrossRef] [PubMed]
- Lloyd JC, Zakhleniuk OV. Responses of primary and secondary metabolism to sugar accumulation revealed by microarray expression analysis of the Arabidopsis mutant, pho3. Journal of Experimental Botany 2004, 55, 1221–1230. [Google Scholar] [CrossRef]
- Lo Piccolo E, Landi M, Pellegrini E, Agati G, Giordano C, Giordani T, Lorenzini G, Malorgio F, Massai R, Nali C, et al.2018. Multiple Consequences Induced by Epidermally-Located Anthocyanins in Young, Mature and Senescent Leaves of Prunus. Frontiers in Plant Science 9. [CrossRef]
- Logan BA, Stafstrom WC; MJL, Walsh; JS, Reblin; KS, Gould. Examining the photoprotection hypothesis for adaxial foliar anthocyanin accumulation by revisiting comparisons of green- and red-leafed varieties of coleus (Solenostemon scutellarioides). Photosynthesis Research 2015, 124, 267–274. [Google Scholar] [CrossRef]
- Loreti E, Povero G; G, Novi; C, Solfanelli; A, Alpi; P, Perata. Gibberellins, jasmonate and abscisic acid modulate the sucrose-induced expression of anthocyanin biosynthetic genes in Arabidopsis. New Phytologist 2008, 179, 1004–1016. [Google Scholar] [CrossRef] [PubMed]
- Lotkowska ME, Tohge T; AR, Fernie; G-P, Xue; S, Balazadeh; B, Mueller-Roeber. The Arabidopsis Transcription Factor MYB112 Promotes Anthocyanin Formation during Salinity and under High Light Stress. Plant Physiology 2015, 169, 1862–1880. [Google Scholar] [CrossRef]
- Lozoya-Gloria E, Cuéllar-González F; N, Ochoa-Alejo. Anthocyanin metabolic engineering of Euphorbia pulcherrima: advances and perspectives. Frontiers in Plant Science 14, 2023. [Google Scholar]
- Lunau K, Maier EJ. Innate colour preferences of flower visitors. Journal of Comparative Physiology A 1995, 177, 1–19. [Google Scholar] [CrossRef]
- Luo J, Nishiyama Y, Fuell C, Taguchi G, Elliott K, Hill L, Tanaka Y, Kitayama M, Yamazaki M, Bailey P, et al.2007. Convergent evolution in the BAHD family of acyl transferases: identification and characterization of anthocyanin acyl transferases from Arabidopsis thaliana. The Plant Journal 50: 678–695. [CrossRef]
- Ma H, Yang T; Y, Li; J, Zhang; T, Wu; T, Song; Y, Yao; J, Tian. The long noncoding RNA MdLNC499 bridges MdWRKY1 and MdERF109 function to regulate early-stage light-induced anthocyanin accumulation in apple fruit. The Plant Cell 2021, 33, 3309–3330. [Google Scholar] [CrossRef] [PubMed]
- Mabry TJ, Turner BL. Chemical Investigations of the Batidaceae. TAXON 1964, 13, 197–200. [Google Scholar] [CrossRef]
- Manetas Y. 2006. Why some leaves are anthocyanic and why most anthocyanic leaves are red? Flora 3: 163–177. [CrossRef]
- Manetas Y, Petropoulou Y; GK, Psaras; A, Drinia. Exposed red (anthocyanic) leaves of Quercus coccifera display shade characteristics. Functional plant biology: FPB 2003, 30, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Marinova K, Pourcel L; B, Weder; M, Schwarz; D, Barron; J-M, Routaboul; I, Debeaujon; M, Klein. The Arabidopsis MATE transporter TT12 acts as a vacuolar flavonoid/H+ -antiporter active in proanthocyanidin-accumulating cells of the seed coat. The Plant Cell 2007, 19, 2023–2038. [Google Scholar] [CrossRef]
- Marin-Recinos MF, Pucker B. Genetic factors explaining anthocyanin pigmentation differences. BMC Plant Biology 2024, 24, 627. [Google Scholar] [CrossRef]
- Mark Hodges D, Nozzolillo C. Anthocyanin and Anthocyanoplast Content of Cruciferous Seedlings Subjected to Mineral Nutrient Deficiencies. Journal of Plant Physiology 1996, 147, 749–754. [Google Scholar] [CrossRef]
- Marquardt C. 1835. Die Farben der Blüthen. Habicht.
- Martens S, Forkmann G; L, Britsch; F, Wellmann; U, Matern; R, Lukačin. Divergent evolution of flavonoid 2-oxoglutarate-dependent dioxygenases in parsley 1. FEBS Letters 2003, 544, 93–98. [Google Scholar] [CrossRef]
- Matile P. 2000. Biochemistry of Indian summer: physiology of autumnal leaf coloration. Experimental Gerontology 35: 145–158. [CrossRef]
- Matile Ph, Flach BM-P; BM, Eller. Autumn Leaves of Ginkgo biloba L.: Optical Properties, Pigments and Optical Brighteners. Botanica Acta 1992, 105, 13–17. [Google Scholar] [CrossRef]
- Mato M, Ozeki Y; Y, Itoh; D, Higeta; K, Yoshitama; S, Teramoto; R, Aida; N, Ishikura; M, Shibata. Isolation and Characterization of a cDNA Clone of UDP-Galactose: Flavonoid 3-0-Galactosyltransferase (UF3GaT) Expressed in Vigna mungo Seedlings. Plant and Cell Physiology 1998, 39, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Matsuba Y, Sasaki N, Tera M, Okamura M, Abe Y, Okamoto E, Nakamura H, Funabashi H, Takatsu M, Saito M, et al.2010. A Novel Glucosylation Reaction on Anthocyanins Catalyzed by Acyl-Glucose–Dependent Glucosyltransferase in the Petals of Carnation and Delphinium[C][W]. The Plant Cell 22: 3374–3389. [CrossRef]
- McClintock B. 1950. The Origin and Behavior of Mutable Loci in Maize. Proceedings of the National Academy of Sciences of the United States of America 36: 344–355.
- Mendel G. 1865. Versuche uber pflanzen-hybriden. Vorgelegt in den Sitzungen.
- Meng J, Wang H; R, Chi; Y, Qiao; J, Wei; Y, Zhang; M, Han; Y, Wang; H, Li. The eTM–miR858–MYB62-like module regulates anthocyanin biosynthesis under low-nitrogen conditions in Malus spectabilis. New Phytologist 2023, 238, 2524–2544. [Google Scholar] [CrossRef] [PubMed]
- Meng L-S, Xu M-K, Wan W, Yu F, Li C, Wang J-Y, Wei Z-Q, Lv M-J, Cao X-Y, Li Z-Y, et al.2018. Sucrose Signaling Regulates Anthocyanin Biosynthesis Through a MAPK Cascade in Arabidopsis thaliana. Genetics 210: 607–619. [CrossRef]
- Merzlyak MN, Chivkunova OB. Light-stress-induced pigment changes and evidence for anthocyanin photoprotection in apples. Journal of Photochemistry and Photobiology. B, Biology 2000, 55, 155–163. [Google Scholar] [CrossRef]
- Miller R, Owens SJ; B, Rørslett. Plants and colour: Flowers and pollination. Optics & Laser Technology 2011, 43, 282–294. [Google Scholar] [CrossRef]
- Miyahara T, Sakiyama R; Y, Ozeki; N, Sasaki. Acyl-glucose-dependent glucosyltransferase catalyzes the final step of anthocyanin formation in Arabidopsis. Journal of Plant Physiology 2013, 170, 619–624. [Google Scholar] [CrossRef]
- Miyahara T, Takahashi M; Y, Ozeki; N, Sasaki. Isolation of an acyl-glucose-dependent anthocyanin 7-O-glucosyltransferase from the monocot Agapanthus africanus. Journal of Plant Physiology 2012, 169, 1321–1326. [Google Scholar] [CrossRef]
- Modolo LV, Blount JW; L, Achnine; MA, Naoumkina; X, Wang; RA, Dixon. A functional genomics approach to (iso)flavonoid glycosylation in the model legume Medicago truncatula. Plant Molecular Biology 2007, 64, 499–518. [Google Scholar] [CrossRef] [PubMed]
- Modolo LV, Li L; H, Pan; JW, Blount; RA, Dixon; X, Wang. Crystal Structures of Glycosyltransferase UGT78G1 Reveal the Molecular Basis for Glycosylation and Deglycosylation of (Iso)flavonoids. Journal of Molecular Biology 2009, 392, 1292–1302. [Google Scholar] [CrossRef]
- Moghe GD, Last RL. Something Old, Something New: Conserved Enzymes and the Evolution of Novelty in Plant Specialized Metabolism. Plant Physiology 2015, 169, 1512–1523. [Google Scholar] [CrossRef]
- Mollier A, Pellerin S. Maize root system growth and development as influenced by phosphorus deficiency. Journal of Experimental Botany 1999, 50, 487–497. [Google Scholar] [CrossRef]
- Montefiori M, Comeskey DJ; M, Wohlers; TK, McGhie. Characterization and Quantification of Anthocyanins in Red Kiwifruit (Actinidia spp.). Journal of Agricultural and Food Chemistry 2009, 57, 6856–6861. [Google Scholar] [CrossRef]
- Moreau C, Ambrose MJ; L, Turner; L, Hill; THN, Ellis; JMI, Hofer. The B gene of pea encodes a defective flavonoid 3’,5’-hydroxylase, and confers pink flower color. Plant Physiology 2012, 159, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Morishita T, Kojima Y; T, Maruta; A, Nishizawa-Yokoi; Y, Yabuta; S, Shigeoka. Arabidopsis NAC transcription factor, ANAC078, regulates flavonoid biosynthesis under high-light. Plant & Cell Physiology 2009, 50, 2210–2222. [Google Scholar] [CrossRef]
- Morita Y, Takagi K, Fukuchi-Mizutani M, Ishiguro K, Tanaka Y, Nitasaka E, Nakayama M, Saito N, Kagami T, Hoshino A, et al.2014. A chalcone isomerase-like protein enhances flavonoid production and flower pigmentation. The Plant Journal 78: 294–304. [CrossRef]
- Mueller LA, Goodman CD; RA, Silady; V, Walbot. AN9, a Petunia Glutathione S-Transferase Required for Anthocyanin Sequestration, Is a Flavonoid-Binding Protein. Plant Physiology 2000, 123, 1561–1570. [Google Scholar] [CrossRef] [PubMed]
- Mur LA. 1995. Characterization of Members of the myb Gene Family of Transcription Factors from Petunia hybrida.
- Murai Y, Kokubugata G; M, Yokota; J, Kitajima; T, Iwashina. Flavonoids and anthocyanins from six Cassytha taxa (Lauraceae) as taxonomic markers. Biochemical Systematics and Ecology 2008, 36, 745–748. [Google Scholar] [CrossRef]
- Nagata T, Todoriki S; T, Masumizu; I, Suda; S, Furuta; Z, Du; S, Kikuchi. Levels of active oxygen species are controlled by ascorbic acid and anthocyanin in Arabidopsis. Journal of Agricultural and Food Chemistry 2003, 51, 2992–2999. [Google Scholar] [CrossRef]
- Naing AH, Ai TN; KB, Lim; IJ, Lee; CK, Kim. Overexpression of Rosea1 From Snapdragon Enhances Anthocyanin Accumulation and Abiotic Stress Tolerance in Transgenic Tobacco. Frontiers in Plant Science 9, 2018. [Google Scholar] [CrossRef]
- Nakano Y, Asada K. Spinach chloroplasts scavenge hydrogen peroxide on illumination. Plant and Cell Physiology 1980, 21, 1295–1307. [Google Scholar] [CrossRef]
- Nakatsuka T, Sato K; H, Takahashi; S, Yamamura; M, Nishihara. Cloning and characterization of the UDP-glucose:anthocyanin 5-O-glucosyltransferase gene from blue-flowered gentian. Journal of Experimental Botany 2008, 59, 1241–1252. [Google Scholar] [CrossRef]
- Naya L, Paul S; O, Valdés-López; AB, Mendoza-Soto; B, Nova-Franco; G, Sosa-Valencia; JL, Reyes; G, Hernández. Regulation of copper homeostasis and biotic interactions by microRNA 398b in common bean. PloS One 2014, 9, e84416. [Google Scholar] [CrossRef]
- Neill S, Gould KS. Optical properties of leaves in relation to anthocyanin concentration and distribution. Canadian Journal of Botany 2000, 77, 1777–1782. [Google Scholar] [CrossRef]
- Neill SO, Gould KS; PA, Kilmartin; KA, Mitchell; KR, Markham. Antioxidant activities of red versus green leaves in Elatostema rugosum. Plant, Cell & Environment 2002, 25, 539–547. [Google Scholar] [CrossRef]
- Ni R, Zhu T-T; X-S, Zhang; P-Y, Wang; C-J, Sun; Y-N, Qiao; H-X, Lou; A-X, Cheng. Identification and evolutionary analysis of chalcone isomerase-fold proteins in ferns. Journal of Experimental Botany 2020, 71, 290–304. [Google Scholar] [CrossRef] [PubMed]
- Niu J, Zhang G; W, Zhang; V, Goltsev; S, Sun; J, Wang; P, Li; F, Ma. Anthocyanin concentration depends on the counterbalance between its synthesis and degradation in plum fruit at high temperature. Scientific Reports 2017, 7, 7684. [Google Scholar] [CrossRef]
- Nowak MS, Harder B; SN, Meckoni; R, Friedhoff; K, Wolff; B, Pucker. Genome sequence and RNA-seq analysis reveal genetic basis of flower coloration in the giant water lily Victoria cruziana. : 2024.06.15.599162. [CrossRef]
- Nozue M, Yamada K; T, Nakamura; H, Kubo; M, Kondo; M, Nishimura. Expression of a Vacuolar Protein (VP24) in Anthocyanin-Producing Cells of Sweet Potato in Suspension Culture. Plant Physiology 1997, 115, 1065–1072. [Google Scholar] [CrossRef]
- Nvsvrot T, Yang X; Y, Zhang; L, Huang; G, Cai; Y, Ding; W, Ren; N, Wang. The PdeWRKY65-UGT75L28 gene module negatively regulates lignin biosynthesis in poplar petioles. Industrial Crops and Products 2023, 191, 115937. [Google Scholar] [CrossRef]
- Oberbaueri SF, Starr G. The role of anthocyanins for photosynthesis of Alaskan arctic evergreens during snowmelt. In: Advances in Botanical Research. Academic Press, 129–145. [CrossRef]
- Oberrath R, Böhning-Gaese K. Floral color change and the attraction of insect pollinators in lungwort (Pulmonaria collina). Oecologia 1999, 121, 383–391. [Google Scholar] [CrossRef]
- Ogata J u. n., Sakamoto T; M, Yamaguchi; S, Kawanobu; K, Yoshitama. Isolation and characterization of anthocyanin 5-O-glucosyltransferase from flowers of Dahlia variabilis. Journal of Plant Physiology 2001, 158, 709–714. [Google Scholar] [CrossRef]
- Owen CR, Bradshaw HD. Induced mutations affecting pollinator choice in Mimulus lewisii (Phrymaceae). Arthropod-Plant Interactions 2011, 5, 235–244. [Google Scholar] [CrossRef]
- Padmavati M, Sakthivel N; KV, Thara; AR, Reddy. Differential sensitivity of rice pathogens to growth inhibition by flavonoids. Phytochemistry 1997, 46, 499–502. [Google Scholar] [CrossRef]
- Paz-Ares J, Ghosal D; U, Wienand; PA, Peterson; H, Saedler. The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators. The EMBO Journal 1987, 6, 3553–3558. [Google Scholar] [CrossRef] [PubMed]
- Peng M, Bi Y-M; T, Zhu; SJ, Rothstein. Genome-wide analysis of Arabidopsis responsive transcriptome to nitrogen limitation and its regulation by the ubiquitin ligase gene NLA. Plant Molecular Biology 2007, 65, 775–797. [Google Scholar] [CrossRef] [PubMed]
- Pesch M, Dartan B; R, Birkenbihl; IE, Somssich; M, Hülskamp. Arabidopsis TTG2 Regulates TRY Expression through Enhancement of Activator Complex-Triggered Activation. The Plant Cell 2014, 26, 4067–4083. [Google Scholar] [CrossRef]
- Pringsheim N. 1881. Untersuchungen über Lichtwirkung und Chlorophyllfunction in der Pflanze. DMW - Deutsche Medizinische Wochenschrift 7: 245–248. [CrossRef]
- Pucker B, Iorizzo M. Apiaceae FNS I originated from F3H through tandem gene duplication. PLOS ONE 2023, 18, e0280155. [Google Scholar] [CrossRef]
- Pucker B, Reiher F; HM, Schilbert. Automatic Identification of Players in the Flavonoid Biosynthesis with Application on the Biomedicinal Plant Croton tiglium. Plants 2020, 9, 1103. [Google Scholar] [CrossRef]
- Pucker B, Selmar D. Biochemistry and Molecular Basis of Intracellular Flavonoid Transport in Plants. Plants (Basel, Switzerland) 2022, 11, 963. [Google Scholar] [CrossRef] [PubMed]
- Pucker B, Walker-Hale N; J, Dzurlic; WC, Yim; JC, Cushman; A, Crum; Y, Yang; SF, Brockington. Multiple mechanisms explain loss of anthocyanins from betalain-pigmented Caryophyllales, including repeated wholesale loss of a key anthocyanidin synthesis enzyme. New Phytologist 2024, 241, 471–489. [Google Scholar] [CrossRef]
- Quattrocchio F, Verweij W; A, Kroon; C, Spelt; J, Mol; R, Koes. PH4 of Petunia is an R2R3 MYB protein that activates vacuolar acidification through interactions with basic-helix-loop-helix transcription factors of the anthocyanin pathway. The Plant Cell 2006, 18, 1274–1291. [Google Scholar] [CrossRef]
- Ramsay NA, Glover BJ. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends in Plant Science 2005, 10, 63–70. [Google Scholar] [CrossRef]
- Rausher MD. 2008. Evolutionary Transitions in Floral Color. International Journal of Plant Sciences 169: 7–21. [CrossRef]
- Rempel A, Choudhary N; B, Pucker. KIPEs3: Automatic annotation of biosynthesis pathways. PLOS ONE 2023, 18, e0294342. [Google Scholar] [CrossRef]
- Renner SS, Zohner CM. The occurrence of red and yellow autumn leaves explained by regional differences in insolation and temperature. New Phytologist 2019, 224, 1464–1471. [Google Scholar] [CrossRef]
- Richter R, Dietz A; J, Foster; J, Spaethe; A, Stöckl. Flower patterns improve foraging efficiency in bumblebees by guiding approach flight and landing. Functional Ecology 2023, 37, 763–777. [Google Scholar] [CrossRef]
- Ridley HN. 1930. The dispersal of plants throughout the world.
- Ruxton GD, Schaefer HM. Floral colour change as a potential signal to pollinators. Current Opinion in Plant Biology 2016, 32, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Saad KR, Kumar G; SN, Mudliar; P, Giridhar; NP, Shetty. Salt Stress-Induced Anthocyanin Biosynthesis Genes and MATE Transporter Involved in Anthocyanin Accumulation in Daucus carota Cell Culture. ACS Omega 2021, 6, 24502–24514. [Google Scholar] [CrossRef] [PubMed]
- Schaefer HM, Rolshausen G. Plants on red alert: do insects pay attention? BioEssays: News and Reviews in Molecular, Cellular and Developmental Biology 2006, 28, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Schilbert HM, Schöne M; T, Baier; M, Busche; P, Viehöver; B, Weisshaar; D, Holtgräwe. Characterization of the Brassica napus Flavonol Synthase Gene Family Reveals Bifunctional Flavonol Synthases. Frontiers in Plant Science 12, 2021. [Google Scholar]
- Seitz C, Ameres S; G, Forkmann. Identification of the molecular basis for the functional difference between flavonoid 3’-hydroxylase and flavonoid 3’,5’-hydroxylase. FEBS letters 2007, 581, 3429–3434. [Google Scholar] [CrossRef]
- Shang Y, Venail J; S, Mackay; PC, Bailey; KE, Schwinn; PE, Jameson; CR, Martin; KM, Davies. The molecular basis for venation patterning of pigmentation and its effect on pollinator attraction in flowers of Antirrhinum. New Phytologist 2011, 189, 602–615. [Google Scholar] [CrossRef]
- Sheehan H, Feng T, Walker-Hale N, Lopez-Nieves S, Pucker B, Guo R, Yim WC, Badgami R, Timoneda A, Zhao L, et al.2020. Evolution of l-DOPA 4,5-dioxygenase activity allows for recurrent specialisation to betalain pigmentation in Caryophyllales. New Phytologist 227: 914–929. [CrossRef]
- Sheehan H, Moser M; U, Klahre; K, Esfeld; A, Dell’Olivo; T, Mandel; S, Metzger; M, Vandenbussche; L, Freitas; C, Kuhlemeier. MYB-FL controls gain and loss of floral UV absorbance, a key trait affecting pollinator preference and reproductive isolation. Nature Genetics 2016, 48, 159–166. [Google Scholar] [CrossRef]
- Shi M-Z, Xie D-Y. Features of anthocyanin biosynthesis in pap1-D and wild-type Arabidopsis thaliana plants grown in different light intensity and culture media conditions. Planta 2010, 231, 1385–1400. [Google Scholar] [CrossRef]
- Shin DH, Choi M; K, Kim; G, Bang; M, Cho; S-B, Choi; G, Choi; Y-I, Park. HY5 regulates anthocyanin biosynthesis by inducing the transcriptional activation of the MYB75/PAP1 transcription factor in Arabidopsis. FEBS Letters 2013, 587, 1543–1547. [Google Scholar] [CrossRef]
- Shin J, Park E; G, Choi. PIF3 regulates anthocyanin biosynthesis in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis. The Plant Journal 2007, 49, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Sinkkonen A. 2008. Red Reveals Branch Die-back in Norway Maple Acer platanoides. Annals of Botany 102: 361–366. [CrossRef]
- Sivankalyani V, Feygenberg O; S, Diskin; B, Wright; N, Alkan. Increased anthocyanin and flavonoids in mango fruit peel are associated with cold and pathogen resistance. Postharvest Biology and Technology 2016, 111, 132–139. [Google Scholar] [CrossRef]
- Sivitz AB, Reinders A; JM, Ward. Arabidopsis Sucrose Transporter AtSUC1 Is Important for Pollen Germination and Sucrose-Induced Anthocyanin Accumulation. Plant Physiology 2008, 147, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Smillie RM, Hetherington SE. Photoabatement by Anthocyanin Shields Photosynthetic Systems from Light Stress. Photosynthetica 1999, 36, 451–463. [Google Scholar] [CrossRef]
- Smith SD. 2010. Using phylogenetics to detect pollinator-mediated floral evolution. New Phytologist 188: 354–363. [CrossRef]
- Solfanelli C, Poggi A; E, Loreti; A, Alpi; P, Perata. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiology 2006, 140, 637–646. [Google Scholar] [CrossRef]
- Stiles EW. 1982. Fruit Flags: Two Hypotheses. The American Naturalist 120: 500–509.
- Stone BC. 1979. Protective Coloration of Young Leaves in Certain Malaysian Palms. Biotropica. [CrossRef]
- Stracke R, Werber M; B, Weisshaar. The R2R3-MYB gene family in Arabidopsis thaliana. Current Opinion in Plant Biology 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Su M, Zuo W; Y, Wang; W, Liu; Z, Zhang; N, Wang; X, Chen. The WKRY transcription factor MdWRKY75 regulates anthocyanins accumulation in apples (Malus domestica). Functional plant biology: FPB 2022, 49, 799–809. [Google Scholar] [CrossRef]
- Sun Y, Ji K, Liang B, Du Y, Jiang L, Wang J, Kai W, Zhang Y, Zhai X, Chen P, et al.2017. Suppressing ABA uridine diphosphate glucosyltransferase (SlUGT75C1) alters fruit ripening and the stress response in tomato. The Plant Journal 91: 574–589. [CrossRef]
- Sun Y, Li H; J-R, Huang. Arabidopsis TT19 Functions as a Carrier to Transport Anthocyanin from the Cytosol to Tonoplasts. Molecular Plant 2012, 5, 387–400. [Google Scholar] [CrossRef]
- Sun SG, Liao K; J, Xia; YH, Guo. Floral colour change in Pedicularis monbeigiana (Orobanchaceae). Plant Systematics and Evolution 2005, 255, 77–85. [Google Scholar] [CrossRef]
- Tan H, Luo X; J, Lu; L, Wu; Y, Li; Y, Jin; X, Peng; X, Xu; J, Li; W, Zhang. The long noncoding RNA LINC15957 regulates anthocyanin accumulation in radish. Frontiers in Plant Science 14, 2023. [Google Scholar]
- Tao R, Bai S; J, Ni; Q, Yang; Y, Zhao; Y, Teng. The blue light signal transduction pathway is involved in anthocyanin accumulation in ‘Red Zaosu’ pear. Planta 2018, 248, 37–48. [Google Scholar] [CrossRef]
- Tatsuzawa F, Saito N; N, Murata; K, Shinoda; A, Shigihara; T, Honda. 6-Hydroxypelargonidin glycosides in the orange–red flowers of Alstroemeria. Phytochemistry 2003, 62, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Tavares S, Vesentini D; JC, Fernandes; RB, Ferreira; O, Laureano; JM, Ricardo-Da-Silva; S, Amâncio. Vitis vinifera secondary metabolism as affected by sulfate depletion: diagnosis through phenylpropanoid pathway genes and metabolites. Plant physiology and biochemistry: PPB 2013, 66, 118–126. [Google Scholar] [CrossRef]
- Teng S, Keurentjes J; L, Bentsink; M, Koornneef; S, Smeekens. Sucrose-Specific Induction of Anthocyanin Biosynthesis in Arabidopsis Requires the MYB75/PAP1 Gene. Plant Physiology 2005, 139, 1840–1852. [Google Scholar] [CrossRef] [PubMed]
- Thimm O, Bläsing O; Y, Gibon; A, Nagel; S, Meyer; P, Krüger; J, Selbig; LA, Müller; SY, Rhee; M, Stitt. MAPMAN: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. The Plant Journal: For Cell and Molecular Biology 2004, 37, 914–939. [Google Scholar] [CrossRef]
- Timoneda A, Feng T; H, Sheehan; N, Walker-Hale; B, Pucker; S, Lopez-Nieves; R, Guo; S, Brockington. The evolution of betalain biosynthesis in Caryophyllales. New Phytologist 2019, 224, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Tohge T, Nishiyama Y, Hirai MY, Yano M, Nakajima J, Awazuhara M, Inoue E, Takahashi H, Goodenowe DB, Kitayama M, et al.2005. Functional genomics by integrated analysis of metabolome and transcriptome of Arabidopsis plants over-expressing an MYB transcription factor. The Plant Journal: For Cell and Molecular Biology 42: 218–235. [CrossRef]
- Trunschke J, Lunau K; GH, Pyke; Z-X, Ren; H, Wang. Flower Color Evolution and the Evidence of Pollinator-Mediated Selection. Frontiers in Plant Science 2021, 12, 617851. [Google Scholar] [CrossRef]
- Turcek F. 1963. Color preference in fruit- and seed-eating birds. Proc Int Ornithol Congr 13: 285–292.
- Turing AM. 1952. The chemical basis of morphogenesis. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 237: 37–72. [CrossRef]
- Van der Niet T, Peakall R; SD, Johnson. Pollinator-driven ecological speciation in plants: new evidence and future perspectives. Annals of Botany 2014, 113, 199–212. [Google Scholar] [CrossRef]
- Verweij W, Spelt CE; M, Bliek; M, de Vries; N, Wit; M, Faraco; R, Koes; FM, Quattrocchio. Functionally Similar WRKY Proteins Regulate Vacuolar Acidification in Petunia and Hair Development in Arabidopsis. The Plant Cell 2016, 28, 786–803. [Google Scholar] [CrossRef]
- Verweij W, Spelt C; G-P, Di Sansebastiano; J, Vermeer; L, Reale; F, Ferranti; R, Koes; F, Quattrocchio. An H+ P-ATPase on the tonoplast determines vacuolar pH and flower colour. Nature Cell Biology 2008, 10, 1456–1462. [Google Scholar] [CrossRef]
- Waki T, Mameda R, Nakano T, Yamada S, Terashita M, Ito K, Tenma N, Li Y, Fujino N, Uno K, et al.2020. A conserved strategy of chalcone isomerase-like protein to rectify promiscuous chalcone synthase specificity. Nature Communications 11: 870. [CrossRef]
- Walker AR, Lee E; SP, Robinson. Two new grape cultivars, bud sports of Cabernet Sauvignon bearing pale-coloured berries, are the result of deletion of two regulatory genes of the berry colour locus. Plant Molecular Biology 2006, 62, 623–635. [Google Scholar] [CrossRef]
- Wang H, Fan W; H, Li; J, Yang; J, Huang; P, Zhang. Functional characterization of Dihydroflavonol-4-reductase in anthocyanin biosynthesis of purple sweet potato underlies the direct evidence of anthocyanins function against abiotic stresses. PloS One 2013, 8, e78484. [Google Scholar] [CrossRef] [PubMed]
- Wang T-J, Huang S; A, Zhang; P, Guo; Y, Liu; C, Xu; W, Cong; B, Liu; Z-Y, Xu. JMJ17-WRKY40 and HY5-ABI5 modules regulate the expression of ABA-responsive genes in Arabidopsis. The New Phytologist 2021, 230, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Waterman PG, Ross JA; DB, McKey. Factors affecting levels of some phenolic compounds, digestibility, and nitrogen content of the mature leaves ofBarteria fistulosa (Passifloraceae). Journal of Chemical Ecology 1984, 10, 387–401. [Google Scholar] [CrossRef]
- Wegener CB, Jansen G. Soft-rot Resistance of Coloured Potato Cultivars (Solanum tuberosum L.): The Role of Anthocyanins. Potato Research 2007, 50, 31–44. [Google Scholar] [CrossRef]
- Weiss MR. 1991. Floral colour changes as cues for pollinators. Nature 354: 227–229. [CrossRef]
- Weiss D. 2000. Regulation of flower pigmentation and growth: Multiple signaling pathways control anthocyanin synthesis in expanding petals. Physiologia Plantarum 110: 152–157. [CrossRef]
- Weng J-K, Noel JP. The Remarkable Pliability and Promiscuity of Specialized Metabolism. Cold Spring Harbor Symposia on Quantitative Biology 2012, 77, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Werlein H-D, Kütemeyer C; G, Schatton; EM, Hubbermann; K, Schwarz. Influence of elderberry and blackcurrant concentrates on the growth of microorganisms. Food Control 2005, 16, 729–733. [Google Scholar] [CrossRef]
- Wheldale M. 1916. The anthocyanin pigments of plants.
- Williamson GB. 1982. Plant mimicry: evolutionary constraints. Biological Journal of the Linnean Society 18: 49–58. [CrossRef]
- Willmer P, Stanley DA; K, Steijven; IM, Matthews; CV, Nuttman. Bidirectional flower color and shape changes allow a second opportunity for pollination. Current biology: CB 2009, 19, 919–923. [Google Scholar] [CrossRef]
- Winkel-Shirley B. 2001. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiology 126: 485–493. [CrossRef]
- Wolff K, Friedhoff R; JM, Horz; B, Pucker. Genome sequence of the medicinal and ornamental plant Digitalis purpurea reveals the molecular basis of flower color variation. : 2024.02.14.580303. [CrossRef]
- Wolff K, Pucker B. Dark Side of Anthocyanin Pigmentation. 2024. [Google Scholar] [CrossRef]
- Wolf-Saxon ER, Moorman CC; A, Castro; A, Ruiz-Rivera; JP, Mallari; JR, Burke. Regulatory ligand binding in plant chalcone isomerase-like (CHIL) proteins. The Journal of biological chemistry 2023, 299, 104804. [Google Scholar] [CrossRef]
- Wong DCJ, Wang Z; J, Perkins; X, Jin; GE, Marsh; EG, John; R, Peakall. Molecular Ecology; e17334: n/a, 2024. [Google Scholar] [CrossRef]
- Xu Z-S, Ma J; F, Wang; H-Y, Ma; Q-X, Wang; A-S, Xiong. Identification and characterization of DcUCGalT1, a galactosyltransferase responsible for anthocyanin galactosylation in purple carrot (Daucus carota L.) taproots. Scientific Reports 2016, 6, 27356. [Google Scholar] [CrossRef]
- Xu F, Ning Y; W, Zhang; Y, Liao; L, Li; H, Cheng; S, Cheng. An R2R3-MYB transcription factor as a negative regulator of the flavonoid biosynthesis pathway in Ginkgo biloba. Functional & Integrative Genomics 2014, 14, 177–189. [Google Scholar] [CrossRef]
- Xu Z, Rothstein SJ. ROS-Induced anthocyanin production provides feedback protection by scavenging ROS and maintaining photosynthetic capacity in Arabidopsis. Plant Signaling & Behavior 2018, 13, e1451708. [Google Scholar] [CrossRef]
- Yabuta Y, Motoki T; K, Yoshimura; T, Takeda; T, Ishikawa; S, Shigeoka. Thylakoid membrane-bound ascorbate peroxidase is a limiting factor of antioxidative systems under photo-oxidative stress. The Plant Journal 2002, 32, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki H, Sakihama Y; N, Ikehara. Flavonoid-Peroxidase Reaction as a Detoxification Mechanism of Plant Cells against H2O2. Plant Physiology 1997, 115, 1405–1412. [Google Scholar] [CrossRef]
- Yamazaki M, Yamagishi E; Z, Gong; M, Fukuchi-Mizutani; Y, Fukui; Y, Tanaka; T, Kusumi; M, Yamaguchi; K, Saito. Two flavonoid glucosyltransferases from Petunia hybrida: molecular cloning, biochemical properties and developmentally regulated expression. Plant Molecular Biology 2002, 48, 401–411. [Google Scholar] [CrossRef]
- Yao G, Ming M, Allan AC, Gu C, Li L, Wu X, Wang R, Chang Y, Qi K, Zhang S, et al.2017. Map-based cloning of the pear gene MYB114 identifies an interaction with other transcription factors to coordinately regulate fruit anthocyanin biosynthesis. The Plant Journal 92: 437–451. [CrossRef]
- Yonekura-Sakakibara K, Fukushima A, Nakabayashi R, Hanada K, Matsuda F, Sugawara S, Inoue E, Kuromori T, Ito T, Shinozaki K, et al.2012. Two glycosyltransferases involved in anthocyanin modification delineated by transcriptome independent component analysis in Arabidopsis thaliana. The Plant Journal 69: 154–167. [CrossRef]
- Yonekura-Sakakibara K, Tohge T; F, Matsuda; R, Nakabayashi; H, Takayama; R, Niida; A, Watanabe-Takahashi; E, Inoue; K, Saito. Comprehensive Flavonol Profiling and Transcriptome Coexpression Analysis Leading to Decoding Gene–Metabolite Correlations in Arabidopsis. The Plant Cell 2008, 20, 2160–2176. [Google Scholar] [CrossRef] [PubMed]
- Yoo J, Shin DH; M-H, Cho; T-L, Kim; SH, Bhoo; T-R, Hahn. An ankyrin repeat protein is involved in anthocyanin biosynthesis in Arabidopsis. Physiologia Plantarum 2011, 142, 314–325. [Google Scholar] [CrossRef]
- Yu J, Qiu K; W, Sun; T, Yang; T, Wu; T, Song; J, Zhang; Y, Yao; J, Tian. A long noncoding RNA functions in high-light-induced anthocyanin accumulation in apple by activating ethylene synthesis. Plant Physiology 2022, 189, 66–83. [Google Scholar] [CrossRef]
- Zhang Y, Butelli E, De Stefano R, Schoonbeek H, Magusin A, Pagliarani C, Wellner N, Hill L, Orzaez D, Granell A, et al.2013. Anthocyanins Double the Shelf Life of Tomatoes by Delaying Overripening and Reducing Susceptibility to Gray Mold. Current Biology 23: 1094–1100. [CrossRef]
- Zhang S, Chen J; T, Jiang; X, Cai; H, Wang; C, Liu; L, Tang; X, Li; X, Zhang; J, Zhang. Genetic mapping, transcriptomic sequencing and metabolic profiling indicated a glutathione S-transferase is responsible for the red-spot-petals in Gossypium arboreum. TAG. Theoretical and applied genetics. Theoretische und angewandte Genetik 2022a, 135, 3443–3454. [Google Scholar] [CrossRef]
- Zhang G, Chen D; T, Zhang; A, Duan; J, Zhang; C, He. Transcriptomic and functional analyses unveil the role of long non-coding RNAs in anthocyanin biosynthesis during sea buckthorn fruit ripening. DNA Research 2018, 25, 465–476. [Google Scholar] [CrossRef]
- Zhang M, Zhang X, Wang H, Ye M, Liu Y, Song Z, Du T, Cao H, Song L, Xiao X, et al.2022b. Identification and Analysis of Long Non-Coding RNAs Related to UV-B-Induced Anthocyanin Biosynthesis During Blood-Fleshed Peach (Prunus persica) Ripening. Frontiers in Genetics 13.
- Zhao J, Dixon RA. MATE Transporters Facilitate Vacuolar Uptake of Epicatechin 3′-O-Glucoside for Proanthocyanidin Biosynthesis in Medicago truncatula and Arabidopsis. The Plant Cell 2009, 21, 2323–2340. [Google Scholar] [CrossRef]
- Zheng X, Om K, Stanton KA, Thomas D, Cheng PA, Eggert A, Simmons E, Yuan Y-W, Conradi Smith GD, Puzey JR, et al.2021. The regulatory network for petal anthocyanin pigmentation is shaped by the MYB5a/NEGAN transcription factor in Mimulus. Genetics 217. [CrossRef]
- Zhou H, He J; Y, Zhang; H, Zhao; X, Sun; X, Chen; X, Liu; Y, Zheng; H, Lin. RHA2b-mediated MYB30 degradation facilitates MYB75-regulated, sucrose-induced anthocyanin biosynthesis in Arabidopsis seedlings. Plant Communications 2024, 5, 100744. [Google Scholar] [CrossRef] [PubMed]
- Zhou H, Lin-Wang K; H, Wang; C, Gu; AP, Dare; RV, Espley; H, He; AC, Allan; Y, Han. Molecular genetics of blood-fleshed peach reveals activation of anthocyanin biosynthesis by NAC transcription factors. The Plant Journal 2015, 82, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Zhou L-L, Shi M-Z; D-Y, Xie. Regulation of anthocyanin biosynthesis by nitrogen in TTG1-GL3/TT8-PAP1-programmed red cells of Arabidopsis thaliana. Planta 2012, 236, 825–837. [Google Scholar] [CrossRef]
- Zhu H-F, Fitzsimmons K; A, Khandelwal; RG, Kranz. CPC, a single-repeat R3 MYB, is a negative regulator of anthocyanin biosynthesis in Arabidopsis. Molecular Plant 2009, 2, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Zhu L, Li X; X, Hu; X, Wu; Y, Liu; Y, Yang; Y, Zang; H, Tang; C, Wang; J, Xu. Quality Characteristics and Anthocyanin Profiles of Different Vitis amurensis Grape Cultivars and Hybrids from Chinese Germplasm. Molecules 2021, 26, 6696. [Google Scholar] [CrossRef]
- Zirngibl M-E, Araguirang GE; A, Kitashova; K, Jahnke; T, Rolka; C, Kühn; T, Nägele; AS, Richter. Triose phosphate export from chloroplasts and cellular sugar content regulate anthocyanin biosynthesis during high light acclimation. Plant Communications 2023, 4, 100423. [Google Scholar] [CrossRef]
Figure 1.
A11 (cyanidin 3-O-[2-O(2-O-(sinapoyl)-β-D-xylopyranosyl)-6-O- (4-O-(β-D-glucopyranosyl)-p-coumaroyl-β-D-glucopyranoside] 5-O-[6-O-(malonyl) β-D-gluco-pyranoside]) is the major anthocyanin in the leaves and stems of Arabidopsis thaliana (Bloor & Abrahams, 2002). The IUPAC name was retrieved from PubChem (Kim et al., 2023). The visualization of stereochemistry might deviate slightly due to spatial organization.
Figure 1.
A11 (cyanidin 3-O-[2-O(2-O-(sinapoyl)-β-D-xylopyranosyl)-6-O- (4-O-(β-D-glucopyranosyl)-p-coumaroyl-β-D-glucopyranoside] 5-O-[6-O-(malonyl) β-D-gluco-pyranoside]) is the major anthocyanin in the leaves and stems of Arabidopsis thaliana (Bloor & Abrahams, 2002). The IUPAC name was retrieved from PubChem (Kim et al., 2023). The visualization of stereochemistry might deviate slightly due to spatial organization.
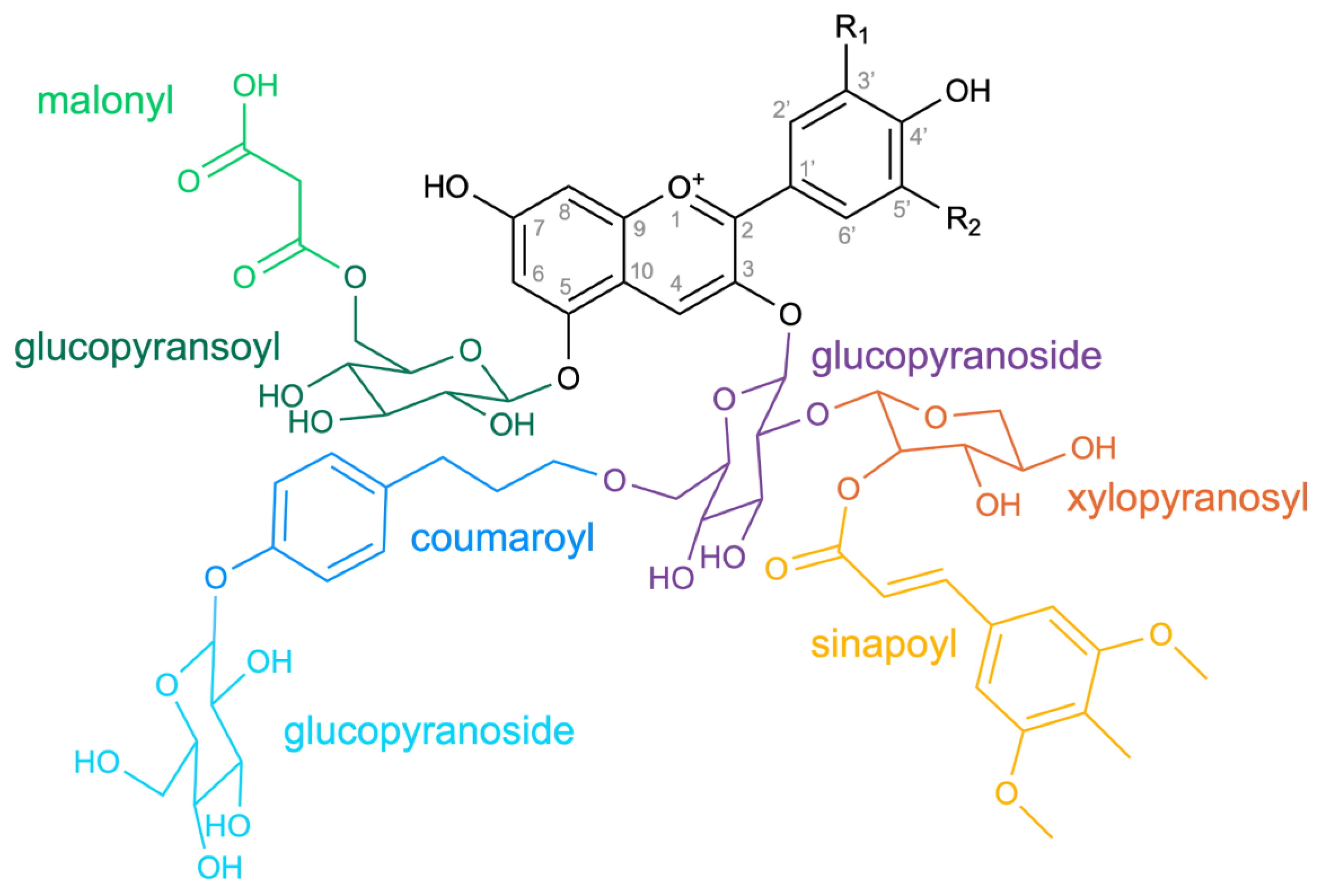
Figure 2.
Simplified illustration of the flavonoid biosynthesis with focus on the anthocyanin branch (A). Enzymes involved in the modification of anthocyanins (B). Displayed enzymes are chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), flavonoid 3’-hydroxylase (F3’H), flavonoid 3’,5’-hydroxylase (F3’5’H), dihydroflavonol 4-reductase (DFR), anthocyanidin synthase (ANS), anthocyanin-related glutathione S-transferase (arGST), UDP-dependent anthocyanidin-3-O-glycosyltransferase (3GT), UDP-glucosyltransferase 75C1 (UGT75C1), UDP-glucosyltransferase 78D2 (UGT78D2), UDP-glucosyltransferase 79B1 (UGT79B1), Malonyl-CoA:anthocyanidin 5-O-glucoside-6''-O-malonyltransferase (5MAT), Coumaroyl-CoA:anthocyanidin 3-O-glucoside-6''-O-coumaroyltransferase (3AT), sinapoyl-β-D-glucose:anthocyanin sinapoyl transferase 10 (SCPL10), and β-glucosidase 10 (BGLU10).
Figure 2.
Simplified illustration of the flavonoid biosynthesis with focus on the anthocyanin branch (A). Enzymes involved in the modification of anthocyanins (B). Displayed enzymes are chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), flavonoid 3’-hydroxylase (F3’H), flavonoid 3’,5’-hydroxylase (F3’5’H), dihydroflavonol 4-reductase (DFR), anthocyanidin synthase (ANS), anthocyanin-related glutathione S-transferase (arGST), UDP-dependent anthocyanidin-3-O-glycosyltransferase (3GT), UDP-glucosyltransferase 75C1 (UGT75C1), UDP-glucosyltransferase 78D2 (UGT78D2), UDP-glucosyltransferase 79B1 (UGT79B1), Malonyl-CoA:anthocyanidin 5-O-glucoside-6''-O-malonyltransferase (5MAT), Coumaroyl-CoA:anthocyanidin 3-O-glucoside-6''-O-coumaroyltransferase (3AT), sinapoyl-β-D-glucose:anthocyanin sinapoyl transferase 10 (SCPL10), and β-glucosidase 10 (BGLU10).
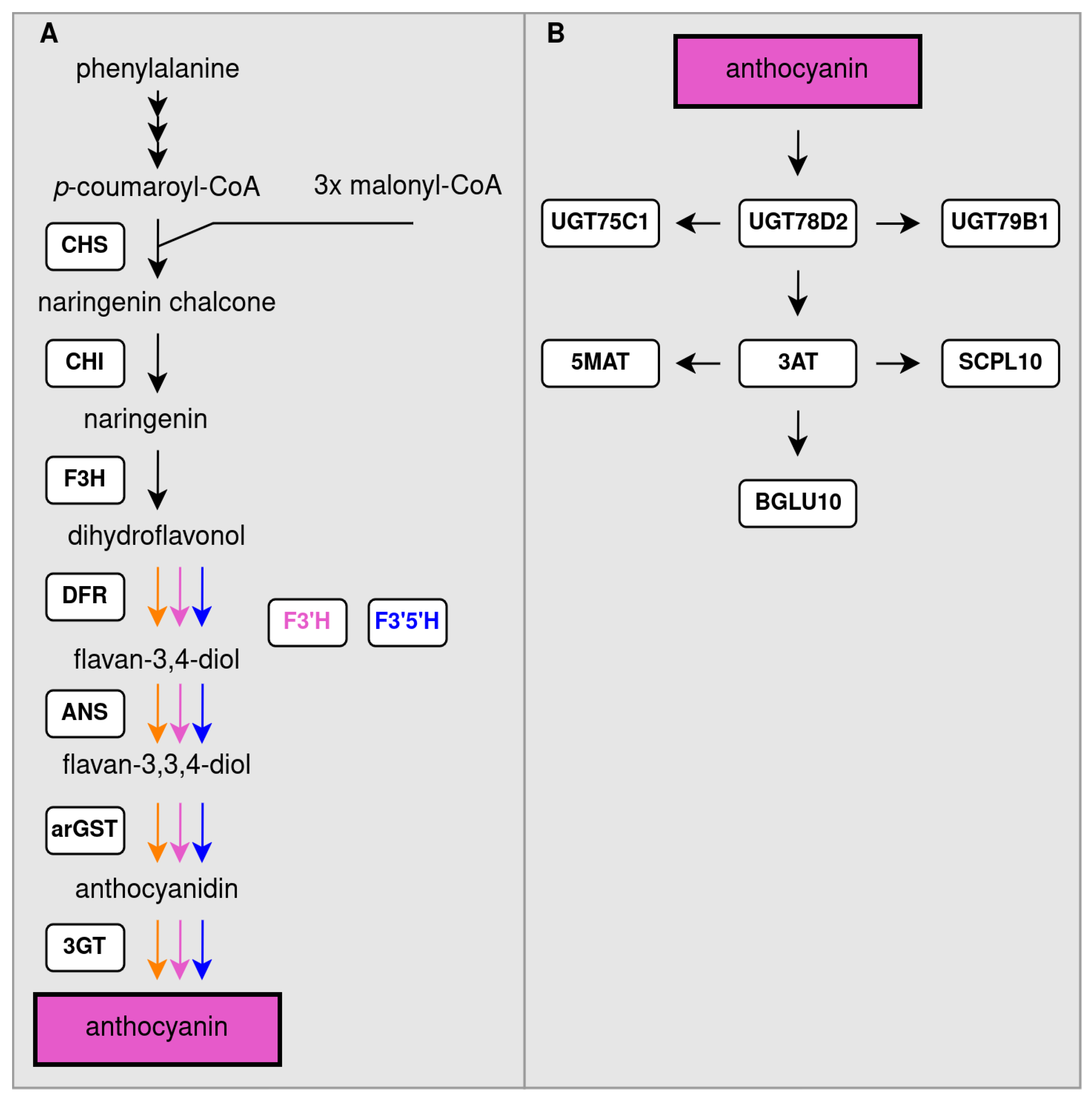
Figure 3.
Transcription factors influencing the activity of anthocyanin biosynthesis genes. Multiple transcription factors are promoting the anthocyanins biosynthesis through the activation of the anthocyanin biosynthesis regulating MYB.
Figure 3.
Transcription factors influencing the activity of anthocyanin biosynthesis genes. Multiple transcription factors are promoting the anthocyanins biosynthesis through the activation of the anthocyanin biosynthesis regulating MYB.
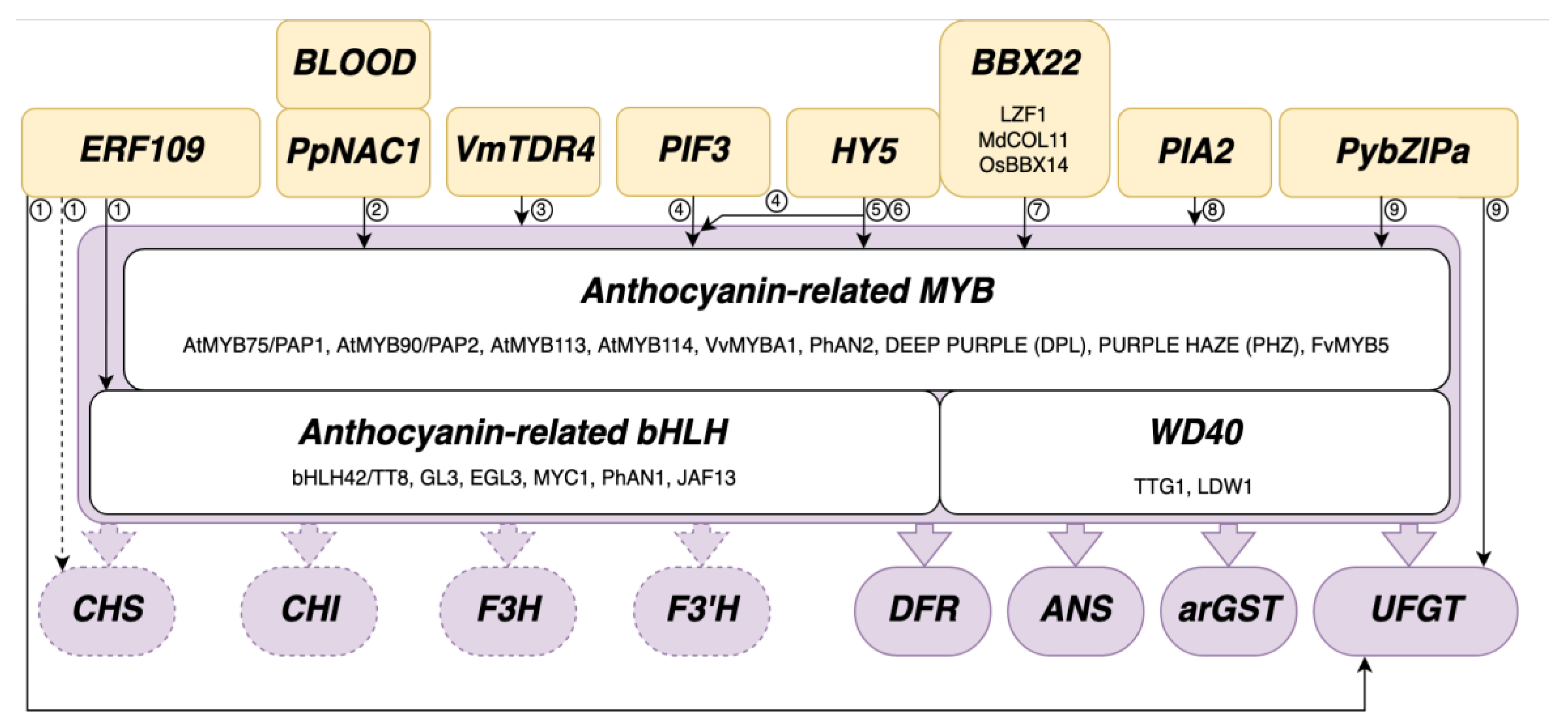
Figure 4.
Simplified illustration of proposed anthocyanin transport routes from the endoplasmic reticulum (ER) to the central vacuole. Figure is based on a previous version published in Pucker & Selmar, 2022.
Figure 4.
Simplified illustration of proposed anthocyanin transport routes from the endoplasmic reticulum (ER) to the central vacuole. Figure is based on a previous version published in Pucker & Selmar, 2022.
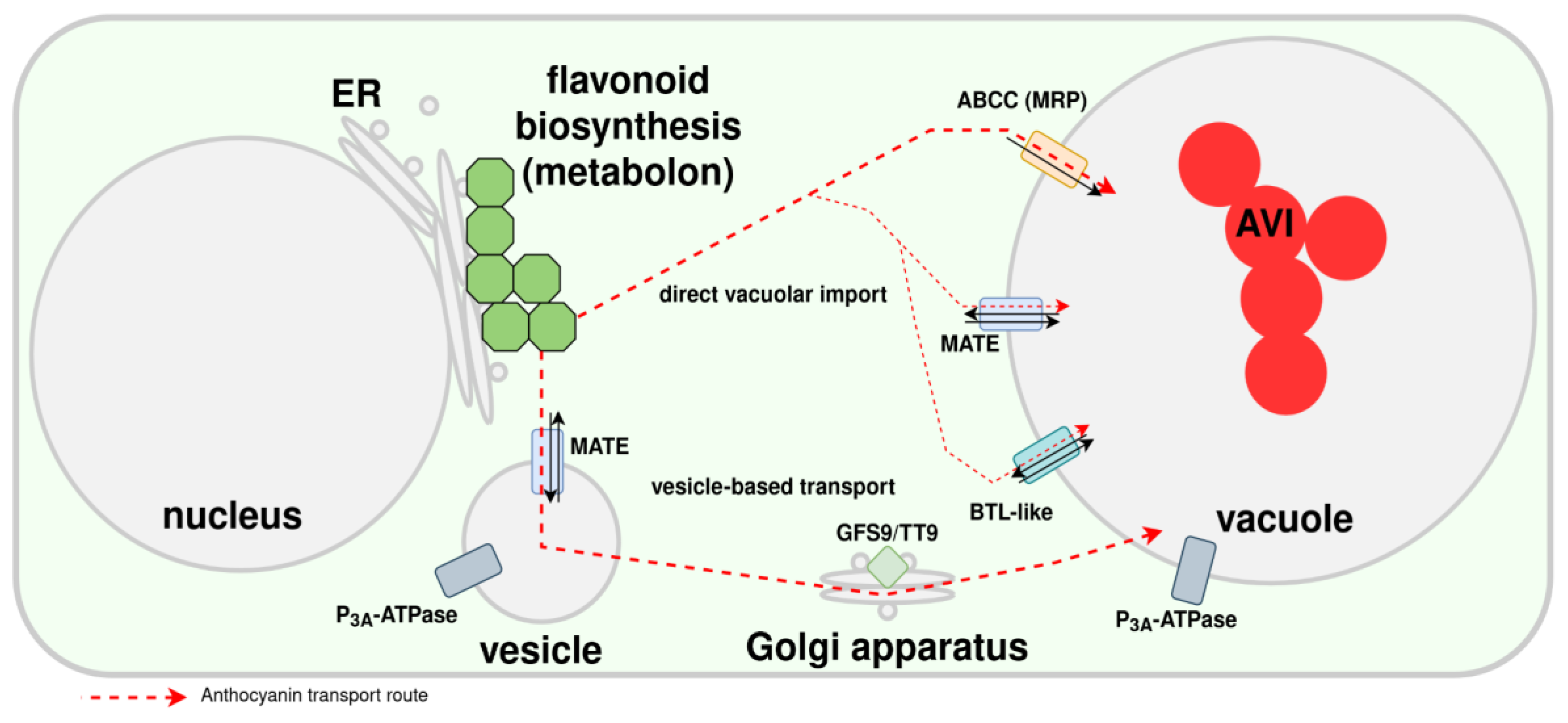
Figure 5.
Anthocyanin pigmentation patterns. Individual cells in specific layers are colored by anthocyanins, while adjacent cells do not accumulate anthocyanins (A). Patterns on flowers emerge due to anthocyanin accumulation in restricted areas like a bull’s eye in the center of a flower or intensely pigmented spots on the petals (B). The formation of pigmented spots can be explained by an activator-repressor model as proposed by Ding et al., 2020 (C).
Figure 5.
Anthocyanin pigmentation patterns. Individual cells in specific layers are colored by anthocyanins, while adjacent cells do not accumulate anthocyanins (A). Patterns on flowers emerge due to anthocyanin accumulation in restricted areas like a bull’s eye in the center of a flower or intensely pigmented spots on the petals (B). The formation of pigmented spots can be explained by an activator-repressor model as proposed by Ding et al., 2020 (C).
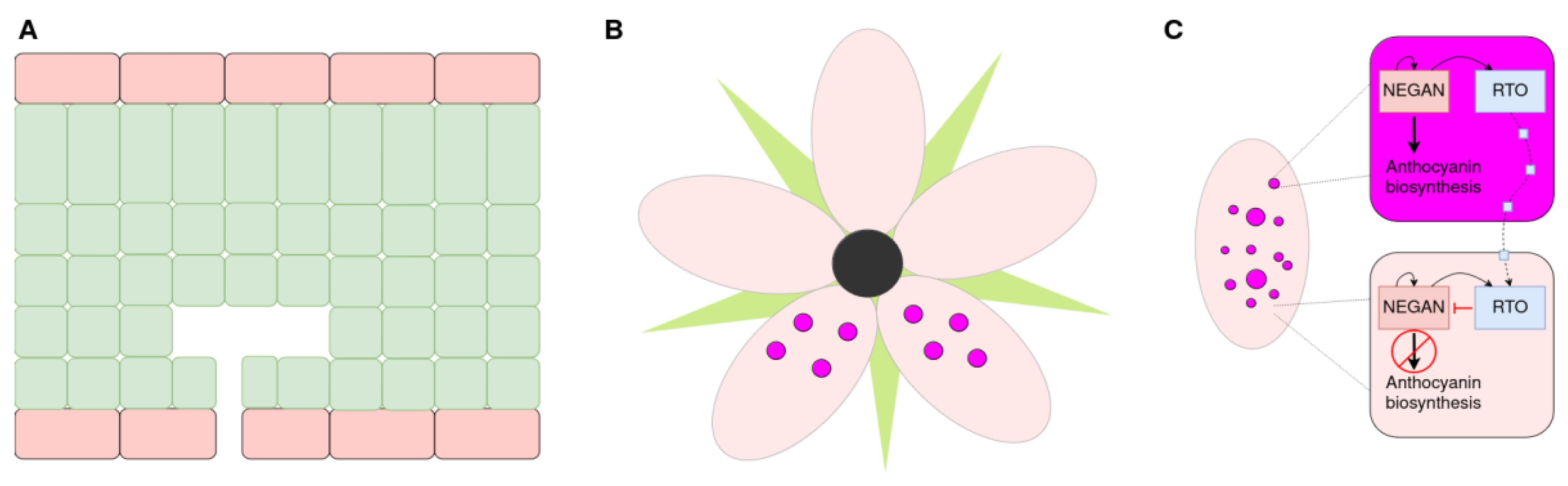
Figure 7.
Ecological functions of anthocyanins in herbivore repellence and pathogen resistances. The camouflage part of this figure is based on Wolff & Pucker, 2024.
Figure 7.
Ecological functions of anthocyanins in herbivore repellence and pathogen resistances. The camouflage part of this figure is based on Wolff & Pucker, 2024.
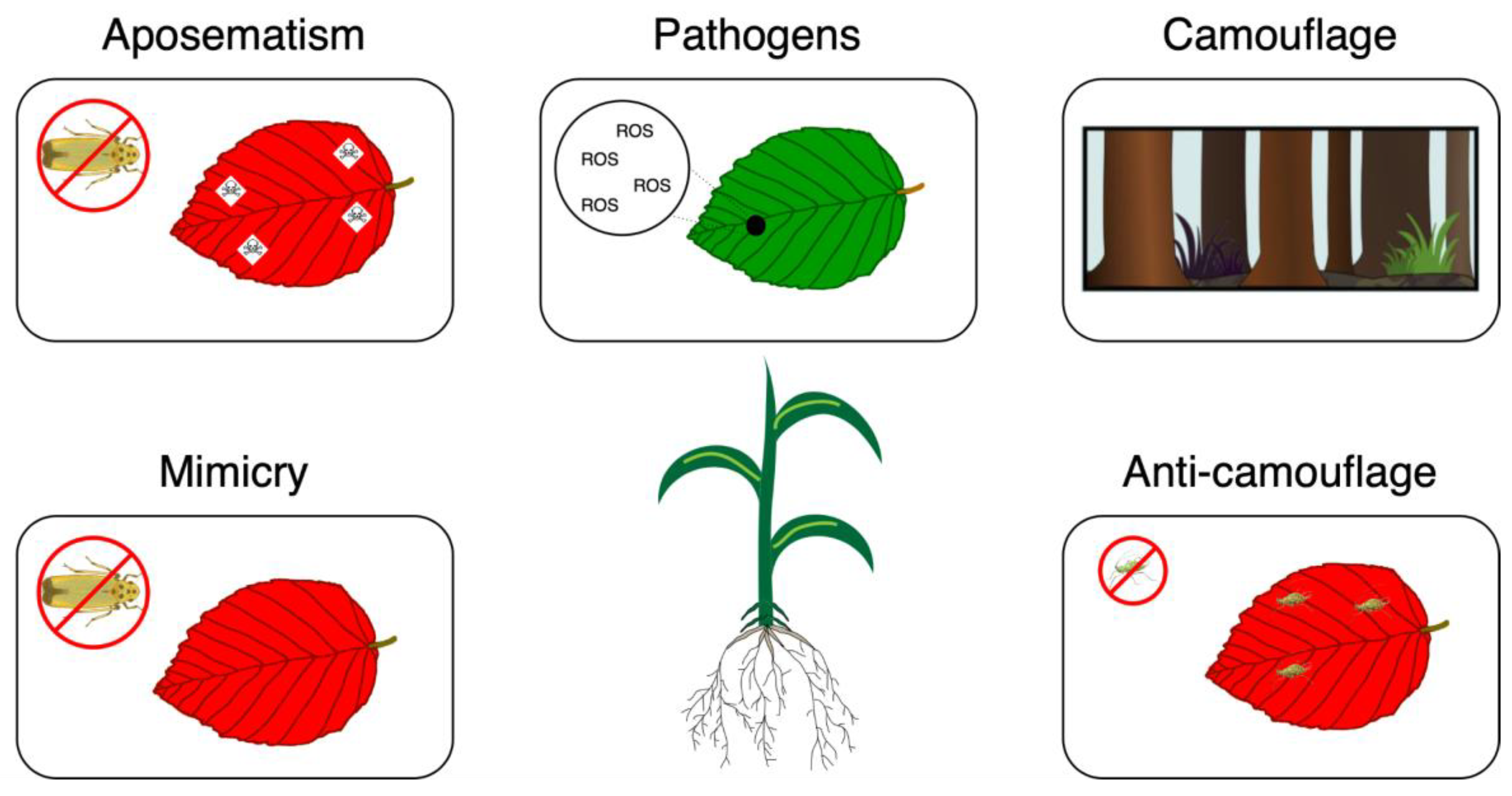
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



