Submitted:
02 September 2024
Posted:
02 September 2024
You are already at the latest version
Abstract
Sect. Tuberculata in the genus Camellia, is named after the "tubercle-like projections on the surface of the capsule and ovary". Due to complex morphological variations in these taxon and insufficient field investigations, the interspecific relationships are unclear, the species definitions are vague, and the names are confusing. This is not conducive to the conservation and study of these species. Therefore, herein, we systematically explored the taxonomic status of five sect. Tuberculata species using morphological, pollen morphology, and molecular phylogenetic methods. The result showed that (1) morphological characteristics for the flower, fruit, and leaves of C. anlungensis and C. leyeensis were similar. Furthermore, the pollen characteristics and pollen wall ornamentation showed that it was no significant difference between the two species. (2) there were significant differences between C. acutiperulata and C. anlungensis in terms of leaf shape (elliptic vs. obovate), calyx characteristics (sepal apex pointed vs. sepal oblong), and fruit shape (subglobose folds with shallow verruculose vs. flat folds and verruculose protuberances with pronounced internal cleavage). (3) C. pyxidiacea and C. rubituberculata differed in flower color (white or few light vs. red), fruit verrucae (obviously deeply cleft vs. shallowly uncracked). (4) a phylogenetic tree based on the chloroplast genome showed that C. anlungensis and C. leyeensis form a single clade (BS = 100%, PP = 1.0) and were on a different branch with C. acutiperulata in clade II, (BS = 100%, PP = 1.0); C. pyxidiacea and C. rubituberculata were clustered on different branches of clade I (BS = 99%, PP = 1.00). Integrated above results, we consider that C. leyeensis should be treated as a homonym of C. anlungensis; C. acutiperulata, C. pyxidiacea, C. rubituberculata respectively as a separate species. Clarifying the taxonomic status of these five species not only advances our understanding of the significance and complexity of the systematic classification of the genus Camellia but also has important implications for diversity conservation and population genetics.
Keywords:
Sect. Tuberculata
; Morphology
; Pollen morphology
; Molecular systematics
; Taxonomic revision
1. Introduction
Section Tuberculata belongs to the genus Camellia, is named after its special morphological characteristics "fruits that show verrucose protuberances" [1]. A species was first discovered in Sichuan Province, China, in 1939 by the famous botanist professor Chien [2], who named it as "Camellia tuberculata Chien". Subsequently, some unknown species with verrucose protuberances were gradually discovered [3,4,5,6,7]. These species was divided into section Heterogenea Sealy in 1958 [8]. Chang Hongda, a famous plant taxonomist on the basis of morphology and field studies, have established sect. Tuberculata based on the tuberculation of the fruit in 1981 [1]; since then, the classification sect. Tuberculata is monophyletic group by molecular phylogenetic, morphological and micro-morphological studies [4,7,9]. However, at the species level, most of the taxonomic treatments were carried out only based on few plant specimens and insufficient field surveys, which resulted in confusing interspecies relationships, confuse interspecies definitions, and nomenclatural confusion. Therefore, to solve the taxonomic relationship for these species in sect. Tuberculata, it is necessary to conduct taxonomic research on the taxonomic groups with taxonomic conflicts.
In the last decade, leaves micromorphology and molecular studies were carried out on the plants of the sect. Tuberculata species (C. anlungensis Chang, C. leyeensis Chang & Y.C. Zhong, C. acutiperulata Chang & Ye, C. pyxidiacea Xu, F. P. Chen & C. Y. Deng and C. rubituberculata Chang & Yu), for example Jiang et al. (2009) were considered the merge application of C. leyeensis with C. anlungensis, and the treatment of C. pyxidiacea, C. rubituberculata, and C. acutiperulata as independent species based on leaves micromorphological studies [8]. Jiang et al. (2017) were constructed a Bayesian phylogenetic tree of Camellia genus based on some chloroplast genes (matK, rbcL, ycf1, trn L-F) union matrix showed that C. anlungensis was not in the same branch as C. acutiperulata [9]. Transcriptome phylogenetic studies of 116 species of Camellia genus showed that C. leyeensis, C. acutiperulata, C. rubituberculata, and C. anlungensis should be treated as different species [10]. Recently, Xiao conducted the first study on the pollen morphology of 13 species of sect. Tuberculata plants, and the results showed that there are interspecific differences in the pollen fraction of sect. Tuberculata plants, which may provide an auxiliary basis for the evolution of the sect. Tuberculata [11]. Ran et al (2024) reported the chloroplast genomes of 13 sect. Tuberculata plants showed that C. pyxidiacea, C. anlungensis, and C. leyeensis clustered on a clade (BS = 100%, PP = 1.00) [12]. However, there are still some problems in these species, such as unclear interspecific relationships, absence of key taxonomic traits, and so on, which is duo to the absence of insufficient field investigation and incomplete research for morphology and systematics at the population level. Therefore, it is necessary to adopt comprehensive methods and multi-disciplinary evidence for species with significant taxonomic conflicts and many taxonomic problems.
Five species of sect. Tuberculata are distribute in the Panjiang River Basin, China, all of which species are narrowly distribute. C. anlungensis was discovered in 1981 by Chang, Lekang township, Wangmo County, Guizhou Province, and its main morphological characteristics are obovate, leathery leaf, a cuneate base, and subglobose capsules [13]; C. rubituberculata was discovered in Qinglong County, Guizhou Province in 1984 and named by Chang (1984) based on its tuberculate pericarp and red flowers [5]; in the same year, another species as C. acutiperulata by Chang (1984) was found from Longlin County, Guangxi Province based on its long elliptical leaf blade and pointed sepal apex [5]. In 1987, a new species in Luoping County, Yunnan Province, it was named as C. pyxidiacea based on its pericarp surfaces with verrucous protuberances and folds as well as deep grooves in the transverse cleavage [14]. In 1991, a species was discovered in Leye County, Guangxi Province, named as C. leyeensis based on its leathery, elliptic leaves with broadly cuneate base, sometimes slightly rounded, globose capsule with pleated pericarp and pilose seed epidermis [15]. Due to difficulties related to the collection of the five plants in taxa belonging to sect. Tuberculata, previous studies on their species have relied mainly on incomplete specimens in the collections for taxonomic studies, and many specimens lacked key "flower and fruit" data, which resulted in unclear interspecific delimitation and still taxonomic problems have not been resolved yet. In this study, we conducted a comparative analysis of five species using traditional morphology, micro morphology, specimen and phylogenetic studies of chloroplast genomes to elucidate the taxonomic status of five species of sect. Tuberculata.
2. Materials and Methods
2.1 Collection of Materials
The study was conducted by field sampling five species of plants in sect. Tuberculata from the Panjiang River Basin, China. We collected fresh pollen preserved in electron microscope fixative and healthy leaves preserved in bags filled with silica gel and sent them to the laboratory for study. The specimens were deposited in the Tree Herbarium of the College of Forestry, Guizhou University (GZAC), the collected information and sample information in table 1 and figure 7.

Table 1.
plant collection information for the five species.
| Species name | Location | Specimen number | Elevation(m) | Longitude and latitude |
|---|---|---|---|---|
| C. anlungensis | Wangmo County, Guizhou, China | GZAC, LZ20211204 | 449 | 106°16′40.69″ E, 25°2′51.65″ N |
| C. leyeensis | Leye County, Guangxi, China | GZAC, LZ20210413 | 684 | 106°17′31.48″ E, 24°51′29.98″ N |
| C. acutiperulata | Longlin County, Guangxi, China | GZAC, LZ20221103 | 851 | 104°52′39.66″ E, 24°39′31.59″ N |
| C. pyxidiacea | Xingyi City, Guizhou, China | GZAC, LZ20211204 | 837 | 104°32′18.18″ E, 24°47′53.12″ N |
| C. rubituberculata | Xingren County, Guizhou, China | GZAC, LZ20210411 | 1297 | 105°21′08.10″ E, 25°43′50.08″ N |
2.2. Experimental Research Method
2.2.1. Data Statistics
During the field survey, data were collected by traditional morphological methods for morphological characteristics of these species, Includes observations of the species' habitat, branches, leaves, flowers, and fruits in their natural state. Texture and color of leaves; type of margin; leaf base and veins; size, number, and color of petals; shape, size, and color of fruit. At least ten specimens were selected from each population to measure the above morphological data were statistics and analysis in Excel 2010 (table 2).
2.2.2. Micromorphological Characteristics of Pollen
Selected FAA-fixed anthers were dehydrated with a gradient of 30%–100% ethanol, and full anthers were selected under a deconstructing microscope, glued to conductive double-sided adhesive tape on the sample stage, and punctured through the anther wall with a deconstructing needle to release pollen. Representative pollen grains were observed under a JSM-6490LV scanning electron microscope (SEM) by coating and spraying with gold, and photographs were taken to preserve the characteristics of the pollen grains, such as the overall view, polar view, equatorial view, and pollen wall ornamentation. Ten pollen grains were selected for each species. For pollen description terminology, refer to the Handbook of palynology, Introduction to sporology, Illustrated Pollen Terminology, and other related works [16,17,18].
2.2.3. Chloroplast DNA Acquisition and Genome Assembly
The collected fresh leaves were used by the modified CTAB method [19] was selected for the extraction of DNA from leaves of five species of sect. Tuberculata. DNA quality was detected by 1% agarose gel electrophoresis, and DNA concentration and purity were determined by a NanoDrop 2000 spectrophotometer. Fragments that passed quality tests were sequenced using Illumina high-throughput sequencing platform. The DNA is then randomly interrupted, end-repaired, and joined to construct sequencing libraries [20,21]. Tailoring low-quality data using tribology software. The qualified complete genome sequences after filtering were compared with those in the experimental group. A circular cp gene map was obtained by de novo splicing of the filtered data using SOAPdenovo 2 and NOVOPlasty [22]. Finally, using C. rubituberculata Chang & Yu (MZ424202) as the reference sequence. the cp DNA was obtained through online annotation, BLAST comparison and manual correction. The cp DNA was mapped using the Organellar Genome DRAW (OGDRAW) software (http:// chlorobox. mpimp-golm. mpg. de/ OGD raw. html) online tool (https:// irscope. shinyapps.io/ Chloroplot/) [23]. The annotated chloroplast genome was uploaded to NCBI to obtain the GenBank accession number.
2.2.4. Expansion and Contraction of the IR Boundary
The chloroplast genome sequences of the five species were selected and analyzed for expansion and contraction of IR region boundaries using IRscope (https://irscope.shinyapps.io/irapp/) [24] online software and the data were plotted for comparison.
2.2.5. Phylogenetic Tree
The chloroplast genome sequences of 19 (contains five new sequences were newly sequenced) of the genus Camellia were downloaded from NCBI. A total of 18 cp genomes of Camellia species and a sister species Stewartia sichuanensis (S. Z. Yan) J. Li & T. L. Ming (GenBank accession number: ON853833) as an outgroup for phylogenetic analysis. Sequence comparison was performed using MAFFT 7 [25]. Phylogenetic trees were reconstructed using the maximum likelihood (ML) method in IQ-TREE v1.6.12 [26] and manually corrected with MEGA X [27], and the best tree-building model was selected. The optimal model (GTR + I + G) was determined using MrModeltest v2.3. The Bayesian (BI) analysis was using MrBayes v3.2.7 software [28] and landscaped in the iTOL v4 (https://itol.embl.de/) online tool [29].
Results
3.1. Morphological Studies
Statistical analyses based on some morphological data (figure 1 and table 2) for the five species. In general, the annual branch were not ribbed; the leaf blades for these species were similar; while, the flowers were white except for C. rubituberculata with red; there were obvious differences in the petals, bracts, and calyx; the fruit diameter and pericarp thickness of C. rubituberculata were greater than those of the other four species; and the morphological characteristics of C. anlungensis and C. leyeensis were very similar. The differences in morphological features of five species in table 2.
Table 2.
morphological characteristics of five species.
| Species name | C.anlungensis | C.leyeensis | C.acutiperulata | pyxidiacea | C.rubituberculata |
|---|---|---|---|---|---|
| Leaf type | leaves thin, leathery, obovate | leaves thin, leathery, obovate | leaves thin, coriaceous, elliptic | leaves thin, coriaceous, elliptic | leaves thin coriaceous, elliptic |
| - length (cm) | 10.72 ± 1.05 | 10.73 ± 0.55 | 12.37 ± 1.09 | 10.88 ± 1.53 | 10.68 ± 0.46 |
| - width (cm) | 3.44 ± 0.55 | 3.32 ± 0.38 | 3.92 ± 0.48 | 4.54 ± 0.30 | 4.08 ± 0.49 |
| Flower type | obovate | obovate | elliptic | elliptic | elliptic |
| - color | white | white | white | with pink petal apexes | red |
| - length (cm) | 2.42 ± 0.26 | 2.55 ± 0.27 | 3.71 ± 0.18 | 2.66 ± 0.14 | 2.98 ± 0.17 |
| - width (cm) | 1.3 ± 0.17 | 1.42 ± 0.24 | 1.62 ± 0.16 | 1.33 ± 0.2 | 2.71 ± 0.39 |
| Number of petals | 11 - 13 | 10 - 12 | 9 - 12 | 6 - 9 | 6 - 8 |
| Sepal | ovate | ovate | ovate, apex pointed, margin membranous | ovate | semicircular |
| Number of calyces | 5 - 7 | 5 - 8 | 5 - 7 | 6 - 8 | 7 - 9 |
| Fruit shape | capsule verrucose, subglobose, epidermis with verrucose projections | capsule verrucose, subglobose, epidermis with verrucose projections | capsule verrucose, subglobose, testa transversely lobed | fruit compressed globose, skin with verrucose protuberance and obvious parting | capsule verrucose, globose, seed coat shallowly undulate |
| - diameter (cm) | 1.75 ± 0.23 | 1.63 ± 0.17 | 1.70 ± 0.29 | 3.29 ± 0.40 | 4.99 ± 0.11 |
| Shell thickness (mm) | 1.79 ± 0.18 | 1.72± 0.34 | 3.49 ± 0.61 | 5.06 ± 1.29 | 13.16 ± 0.7 |
Note: the data in the table are average value ± standard deviation.
Figure 1.
morphological characteristics of five species (A: C. anlungensis, B: C. leyeensis, C: C. acutiperulata, D: C. pyxidiacea and E: C. rubituberculata). 1. habitat, 2. flower, 3. fruit, 4. petals, 5. calyxs, 6. pistils, 7. capsules, 8. seeds, 9. leaves).
Figure 1.
morphological characteristics of five species (A: C. anlungensis, B: C. leyeensis, C: C. acutiperulata, D: C. pyxidiacea and E: C. rubituberculata). 1. habitat, 2. flower, 3. fruit, 4. petals, 5. calyxs, 6. pistils, 7. capsules, 8. seeds, 9. leaves).

3.2. Micromorphological Characteristics of Pollen
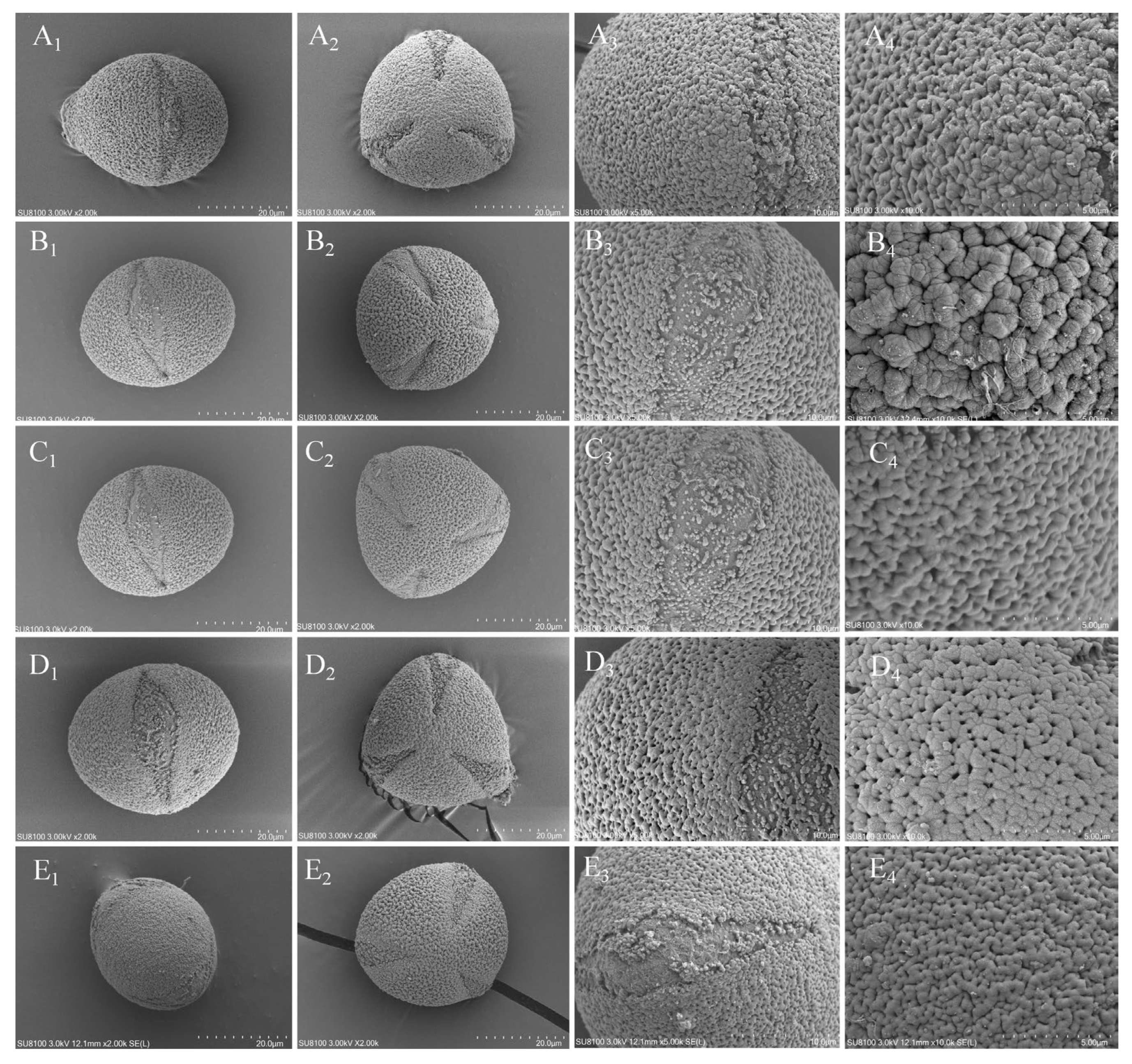
The experimental results of the present study showed that there were no significant differences in pollen shape, polar view, equatorial view, pollen aperture, pollen wall ornamentation, or ridge among the five sect. Tuberculata plants, which could not effectively provide a basis for species identification (figure 2 and table 3).
Table 3.
morphological characteristics of the pollen of five species.
| Species | Pollen Shape | Polar view | Equatorial view | Pollen aperture | Pollen wall ornamentation | Ridge | |
|---|---|---|---|---|---|---|---|
| C. anlungensis | oblate spherical | Triangular | Oval | Colpus | Uneven | Wavy | |
| C. leyeensis | oblate spherical | Triangular | Oval | Colpus | Uneven | Wavy | |
| C. acutiperulata | oblate spherical | Triangular | Oval | Colpus | Uneven | Wavy | |
| C. pyxidiacea | oblate spherical | Triangular | Oval | Colpus | Uneven | Wavy | |
| C. rubituberculata | oblate spherical | Oval | Oval | Colpus | Uneven | Wavy |
Note: the data in the table are average value ± standard deviation.
Figure 2.
morphological characteristics of the pollen of five species. (A: C. anlungensis, B: C. leyeensis, C: C. acutiperulata, D: C. pyxidiacea, E: C. rubituberculata). (1. equatorial view, 2. polar view, 3. Pollen aperture, 4. pollen wall ornamentation).
Figure 2.
morphological characteristics of the pollen of five species. (A: C. anlungensis, B: C. leyeensis, C: C. acutiperulata, D: C. pyxidiacea, E: C. rubituberculata). (1. equatorial view, 2. polar view, 3. Pollen aperture, 4. pollen wall ornamentation).
3.3. Features of Chloroplast Genomes
These whole chloroplast genomes of five species were submitted to the NCBI GenBank and accession numbers were obtained (table 4). A comparison of these chloroplast genomes of five species showed (figure 3 and table 4) that these chloroplast genome length ranged from 156,624 bp to 157,063 bp. It consisted of LSC regions (86,212 – 86,689 bp), an SSC regions (18,276 – 18,286 bp) and IR regions (51,986 – 52,130 bp). The total number of genes was 130 – 136 that which is including 87 – 91 CDSs, 35 – 37 tRNAs, and 8 rRNAs. The chloroplast GC content ranged from 37.30% to 37.33% and the GC content varied only slightly between regions with the IR region having a higher GC content than the other regions.
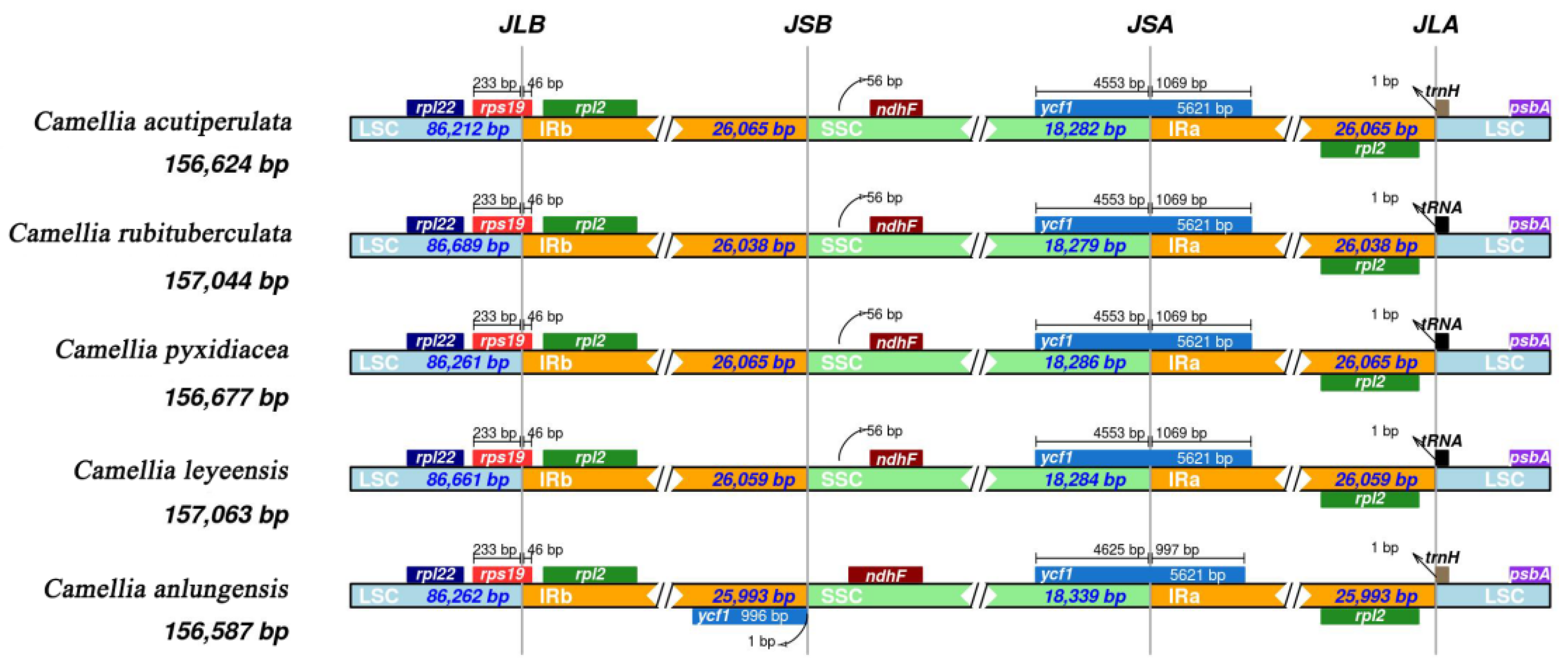
Although the size of IR regions is more highly conserved than other regions, contraction and expansion of IR boundaries are thought to play an important role in genome size. These contraction and expansion analysis of IR boundaries revealed (figure 4) that the rpl2 / rps19 genes in five species a high degree of conservation with identical gene length and location. However, ycf1 gene of C. anlungensis was found within the IR region and the distance between the ndhF gene and IRb region was less than 56 bp. The trnH gene in C. anlungensis and C. acutiperulata was located in the LSC region, whereas C. leyeensis, C. pyxidiacea and C. rubituberculata contained tRNA genes within the LSC region.
Table 4.
characterization of chloroplast genome of five species.
| acutiperulata | C. pyxidiacea | C. anlungensis | C. rubituberculata | C. leyeensis | |
|---|---|---|---|---|---|
| Genome size (bp) | 156,624 | 156,677 | 156,587 | 157,044 | 157,063 |
| GC (%) | 37.33 | 37.33 | 37.33 | 37.31 | 37.30 |
| LSC size (bp) | 86,212 | 86,261 | 86,262 | 86,689 | 86,661 |
| SSC size (bp) | 18,286 | 18,283 | 18,281 | 18,279 | 18,276 |
| IR size (bp) | 52,130 | 52,130 | 51,986 | 52,076 | 52,118 |
| GC in LSC (%) | 35.36 | 35.35 | 35.31 | 35.31 | 35.31 |
| GC in SSC (%) | 30.59 | 30.62 | 30.60 | 30.62 | 30.60 |
| GC in IR (%) | 42.95 | 42.95 | 42.96 | 42.98 | 42.96 |
| GC in CDS (%) | 37.61 | 37.53 | 37.54 | 37.65 | 37.55 |
| 1st position GC (%) | 45.37 | 45.19 | 45.24 | 45.42 | 45.26 |
| 2nd position GC (%) | 38.04 | 37.94 | 37.97 | 38.00 | 37.97 |
| 3rd position GC (%) | 29.43 | 29.43 | 29.40 | 29.53 | 29.40 |
| Length of CDS | 79,500 | 79,767 | 79,671 | 80,155 | 80,175 |
| Number of genes | 130 | 132 | 134 | 136 | 136 |
| Number of CDS | 87 | 87 | 89 | 91 | 91 |
| Number of tRNAs | 35 | 37 | 37 | 37 | 37 |
| Number of rRNAs | 8 | 8 | 8 | 8 | 8 |
| GenBank ID | OQ556869 | OP058659 | NC_050354 | MZ766253 | OK046127 |
Figure 3.
maps of cp gemomes of five species (A: C. anlungensis, B: C. leyeensis, C: C. acutiperulata, D: C. pyxidiacea, E: C. rubituberculata).
Figure 3.
maps of cp gemomes of five species (A: C. anlungensis, B: C. leyeensis, C: C. acutiperulata, D: C. pyxidiacea, E: C. rubituberculata).
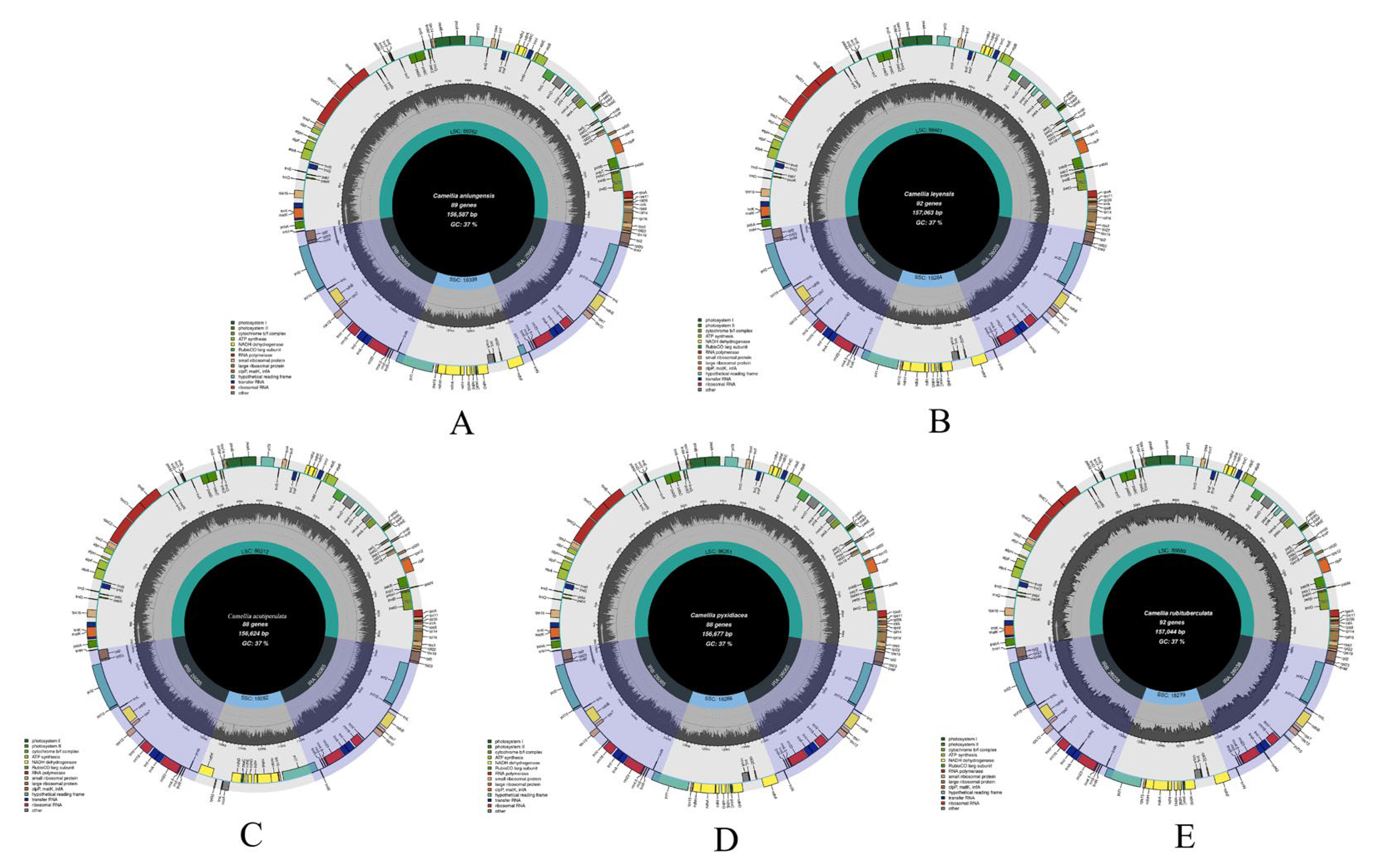
Figure 4.
analysis of the cp genome IR region boundaries of five species.
3.4. Phylogenetic Analysis
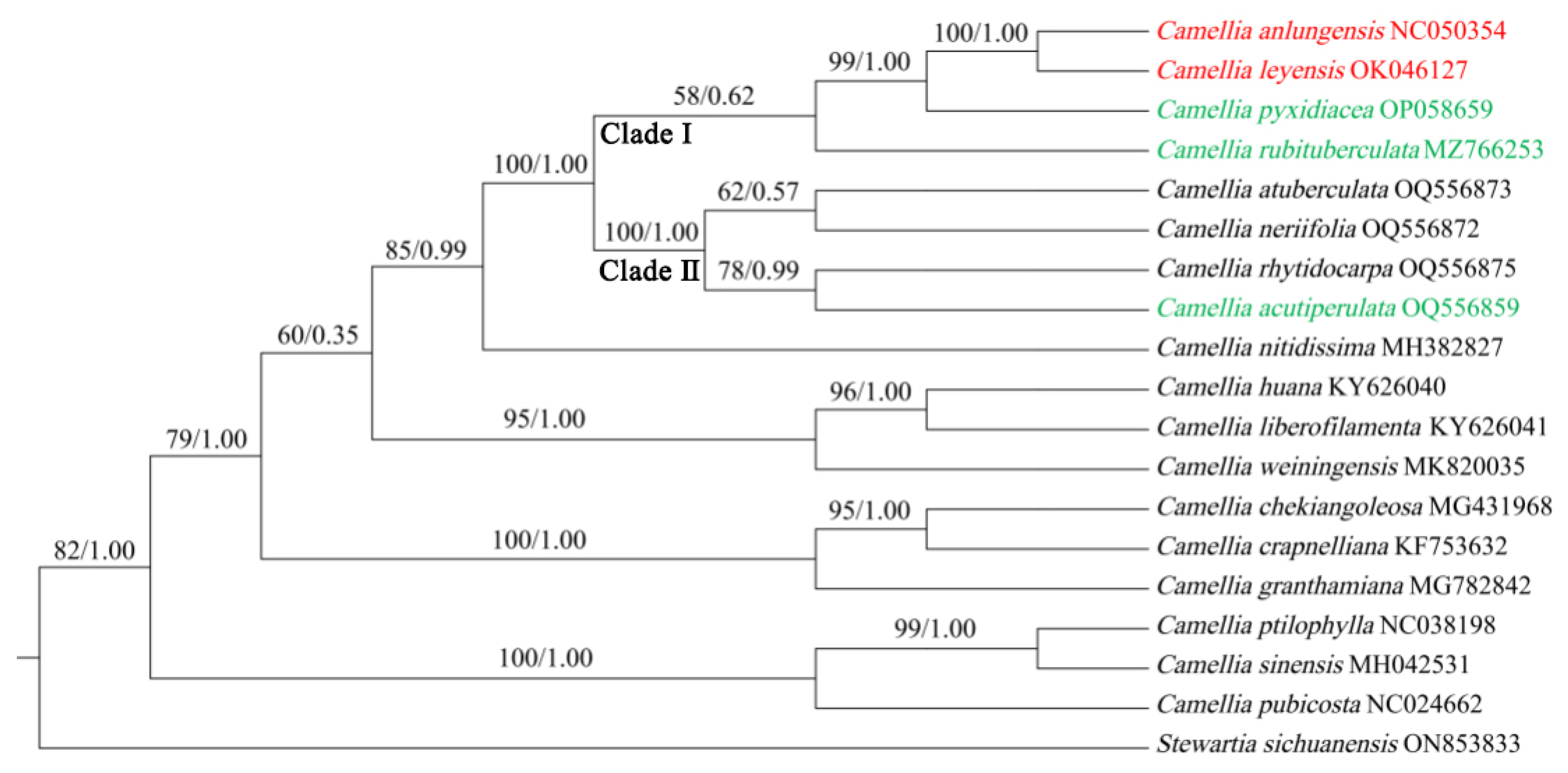
The phylogenetic tree was constructed using maximum likelihood and Bayesian methods by nineteen cp genomes that revealed five species were clustered on the mainbranch consisting of clade I and clade II with high support values (BS = 100%,PP = 1.0)(figure 5). C. anlungensis, C. leyeensis, C. pyxidiacea and C. rubituberculata are on a the clade I (BS = 58%, PP = 0.62), while C. atuberculata, C. neriifolia, C. acutiperulata, and C. rhytidocarpa were located in the clade II. C. anlungensis and C. leyeensis formed a independent clade with a significantly high bootstrap value (BS = 100%, PP = 1.0).
Figure 5.
phylogenetic tree obtained using the Maximum Likelihood (ML) and Bayesian inference (BI) methods for five species based on cp genomes.
Figure 5.
phylogenetic tree obtained using the Maximum Likelihood (ML) and Bayesian inference (BI) methods for five species based on cp genomes.
3.5. Taxonomic Treatment
In this study, we classified five species of sect. Tuberculata with distinct taxonomic positions based on field studies, employing comprehensive approaches encompassing morphology, specimen study, pollen morphology, and molecular systematics research. These result of research showed that C. leyeensis with C. anlungensis should merge and categorized as C. anlungensis, while C. acutiperulata, C. pyxidiacea, and C. rubituberculata are regarded as separate species.
Figure 6.
information on the specimens of C. anlungensis, C. leyeensis, C. acutiperulata, C. pyxidiacea and C. rubituberculata.
Figure 6.
information on the specimens of C. anlungensis, C. leyeensis, C. acutiperulata, C. pyxidiacea and C. rubituberculata.

Camellia Anlungensis Chang
Camellia anlungensis H. T. Chang, Tax. Gen. Camellia 48. 1981 et in Act. Scl. Nat. Univ. Sunyats. 30 (4): 88, 1991; H. T. Chang et B. Bartholomew. Camellias 69. 1984; Flora of Guizhou 5: 16,. pl. 3: 3 - 4. 1988.
Type: China, Guizhou, Anlong, Guizhou Team, Botanical Institute, Zhang Zhisong, Zhang Yongtian 5952; Ceheng, Luxiong Logging Farm, Cao Ziyu 1227, 1259; Wangmu, Guizhou Team 1757 (Type in Bot. Inst. Acad. Sin Peking) (PE, GZBI) (figure 6A).
Additional specimens examined:—China, Guizhou, Wangmo, Lekang, 400 – 500 m, 4 November 2021, LZ20211204 (GZAC).
= Camellia leyeensis H. T. Chang & Y. C.; Zhong in Act. Scl. Nat. Univ. Sunyats 30 (4): 89, 1991.
Type: China, Guangxi, Leye, Y. C. Zhong 890806 (SYS, GXFS, KUN) (figure 6B).
Additional specimens examined:—China, Guangxi, Leye, Yachang, 600 – 750 m, 13 April 2021, LZ20210413, LZ20210414, LZ20210415 (GZAC).
Morphological description: perennial shrub to small tree, plant height 2 – 4 m, diameter at breast height 4 – 10 cm, trunk yellow‒brown, height below branches 1.5 – 3 m. Leaves alternate, shoots without ribs, glabrous, leaves are thin and leathery, obovate, 9.14 – 12.54 cm long, 2.78 – 4.45 cm wide, apex sharply acute, base broadly cuneate, light green above, dark obscure, light yellowish green below, no black glandular dots, lateral veins 6 – 9 pairs, curved to the tip of the leaf near the margin, obvious on both the upper and lower surfaces, the margins sparsely serrulate, serration almost symmetrical on both sides, serration 20 – 31 pairs, teeth 2 – 4 mm apart, petiole length 0.4 – 0.8 cm. Flower white, calyx segments ovate and hairy, single at the top of the branch, nearly sessile. Petals 9 – 13, obovate–oblong, 2.08 – 2.95 cm long, 0.97 – 1.54 cm wide; outer filament whorl-free, anthers deep yellow, filaments creamy-white, coherent at base; ovary glabrous, 3 – 4 locular, 3 – 4 styles, 2.4 – 2.8 cm long. Capsule subsessile, carpophore 0.26 – 0.32 mm long, fruit 1.4 – 2.5 cm in diameter. Pericarp with many folds and verrucose projections, 1.2 – 2.3 mmthickest. Seed 1 per locule, dark brown and lanceolate, semiellipsoid, 13 – 16 mm. Flowering period, October – November.
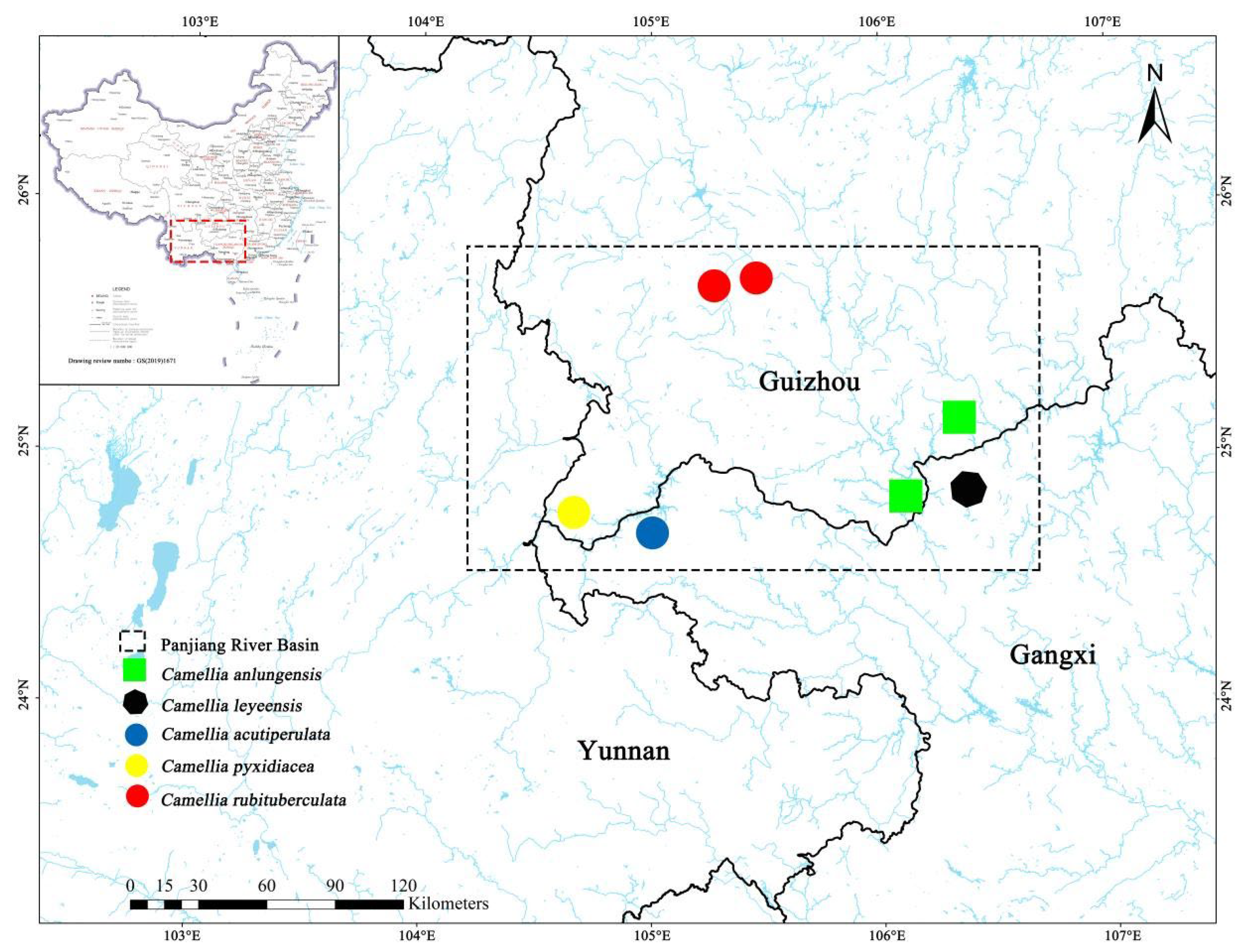
Distribution and habitat: distributed in Lekang village, Wangmo, Guizhou, China (figure 7). Most species grow on slopes and river valleys at 400 – 500 m above sea level, under mixed woods forests in the mountains, sometimes forming groups. It is also distributed in Yachang village, Leye, Guangxi, China. Most species grow on slopes at 600 – 750 m above sea level, under mixed woods forests in the mountains, sometimes forming groups.
Camellia acutiperulataChang & C. X. Ye
Camellia acutiperulata H. T. Chang & C. X. Ye in Act. Scl. Nat. Univ. Sunyats. 1984(2): 79, 1984. 30(4): 88, 1991, in clavi; Flora of Guangxi 1: 771, 1991.
Type: China, Guangxi, Longlin, Zeng Pei, Xie Qingjian 17023 (SYS) (figure 6C).
Additional specimens examined:— China, Gangxi, Longlin, Jinzhongshan, 800 - 900 m, 4 November 2022, LZ20221103, LZ20221104, LZ20221105 (GZAC).
Morphological description: perennial shrub to small tree, shoots glabrous. Leaves thin and leathery, elliptic with apex acute or slightly obtuse, 0.28 – 13.69 cm long and 3.32 – 4.73 cm wide, base cuneate to rounded, drying dark green above, slightly shiny, green below, glabrous, 6 – 7 pairs of lateral veins, both slightly elevated on the abaxial surface, margins sparsely serrulate or subentire in the lower half, 5–10 mm petiole length. Flower white, 1 – 2 axillary, sessile; 5 – 6 bracts, 3 – 5 mm long, apex pointed, glabrous or midrib pilose; sepals 5 – 7, broadly ovate, 1.4 – 1.7 cm long, margin membranous, apex pointed, glabrous or long velutinous at tip; petals 9-12, elliptic with 3.4 – 4.0 cm long, 1.3 – 1.9 cm wide, glabrous, filaments, outer whorl nearly free or slightly connate, glabrous, ovary 3 – 4 loculed, glabrous; styles 3 – 4, free, 2.0 – 2.3 cm long. Capsule subglobose, 1.3 – 2.1 cm in diameter, pericarp approximately with 3.1 – 4.3 mm thick, verruculose, raised. Seeds 1 – 2 per locule, orbicular, bald and glabrous. The flowering period is November – December.
Distribution and habitat: distributed in Longlin, Guangxi, China (figure 7). Most species grow on steep slopes at 800 – 900 m above sea level, under mixed woods forests in the mountains, sometimes forming small groups.
Camellia pyxidiaceaXu, F. P. Chen & C. Y. Deng
Camellia pyxidiacea Xu, F. P. Chen & C. Y. Deng in Guihaia 7(1): 19 - 21, 1987; H. T. Chang in Act. Sci. Nat. Univ. Sunyats. 30 (4): 88. 1991, in clavi.
Type: China, Yunnan, Luoping, Xu Zhaoran SJ5225 (SYS) (figure 6D).
Additional specimens examined: — China, Yunnan, Luoping, Yehou valley, 800 – 900 m, 4 December 2021, LZ20211204, LZ20211205, LZ20211206 (GZAC).
Morphological description: shrub to small tree; shoots glabrous, brown. Leaves leathery, elliptic, 8.14 – 13.08 cm long, 3.94 – 4.88 cm wide. apex neo-acute, margins serrulate, upper drying olive-green, slightly glossy, glabrous, underneath yellowish-green, glabrous; primary veins slightly convex above, distinctly elevated below; lateral veins 8 – 10 pairs, conspicuous above, slightly convex below, reticulate veins conspicuous on leaf surface with petiole length 6 – 12 mm, glabrous. Single flower axillary near tip, white or petal apex pink; calyx larger, 7 – 9, hairy at apex, semiorbicular or ovate, 3 – 6 mm long, 4 – 9 mm wide; petals 6 – 8, obovate, apically lobed, basally coherent, 2.43 – 2.88 cm long, 0.95 – 1.58 cm wide, petal apex hairy, filaments white, yellow; glabrous; styles 3 – 4, slender, 2.5 – 3.4 cm long. Capsule subglobose, 4.84 – 5.18 cm in diameter; ovary 3 – 4 loculed, pericarp longitudinally lobed, surface densely tuberculate and wrinkled, as well as transverse deep grooves, pericarp often dehiscent along these transverse grooves. Seeds hairy, triangular-orbicular or subsemi-orbicular. Flowering period in October. Fruit ripening in the following year in October – November.
Distribution and habitat: Distributed in Yehou valley, Luoping, Yunnan, China (figure 7). Most species grow on steep slopes at 800 – 900 m above sea level, under mixed woods forests in the mountains, sometimes forming groups.
Camellia rubituberculataChang
Camellia rubituberculata Chang, Lin & Liu, in Act. Scl. Nat. Univ. Sunyatseni, 23(2): 82, 1984; H. T. Chang. 30 (4): 88. 1991, in clavi.; Flora of Guizhou 5: 10. 1988.
Type: China, Guizhou, Qinglong, Lin Mengjia and Lu Qiming 155710 (SYS, GZTI) (figure 6E).
Additional specimens examined:— China, Guizhou, Qinglong, Longtou village , 1200 - 130 m, 11 April 2021, LZ20210411, LZ20210412, LZ20210413 (GZAC).
Morphological description: perennial shrub to subtree 5 – 10 m, annual shoots glabrous. Leaves thin and leathery, long ovate, 10.1 cm – 11.4 cm long, 3.5 – 4.8 cm wide; blade apex slightly pointed or acute mucronate, base broadly cuneate, The overall appearance of the leaves blade is olive green, glabrous, dark green adaxially with 7 – 8 pairs of lateral veins clearly visible on both sides. Flowers red, terminal, or axillary; 4 – 7 cm in diameter; bracts persistent with imbricate; 6 – 8, semiorbicular or suborbicular; 2.79 – 3.25 cm long, 2.15 – 3.24 cm wide; basally connate; petal apices shallowly columnar; outer filament whorl free, anthers dark yellow, filaments creamy white, basally connate, united with petals; outer whorl free, anthers dark yellow, filaments creamy white, base connate, united with petals. Capsule with verrucose globose, seed coat 3-4 cleft, thickness 1.22 cm; fruit diameter 4.84 – 5.18 cm; ovary 3 – 4 locular, containing 3 – 11 seeds, mostly triangular, ovate, obovate or trapezoidal-obovate, seeds brown, triangular-orbicular or subsemi-orbicular. Flowering period in October – November.
Distribution and habitat: distributed in Longtou village, Qinglong, Guizhou, China (figure 7). Most species grow on slopes at 1200 – 1300 m above sea level, under mixed woods forests in the mountains, sometimes forming groups.
Figure 7.
distribution map of five species.
Discussion
4.1. Taxonomic processing
Many taxonomic confictes are caused by differences in definition or incomplete taxonomic stduy. We conducted extensive filed surverys and specimen studies for the sect. Tuberculata, A series of morphological data were measuered at population level, including leaves, fruit and other morphological characteristics. The results showed that C. pyxidiacea and C. rubituberculata are differences in flower color, number of petals and stigmas, capsule morphology, etc., whcih supporting Min (1993; 1996) and Chang et al (1996) [3,4,5] views that there are significant morphological differences between the two species. At the same time, the above morphological forms are very similar in C. anlungensis and C. leyeensis. It is noteworthy that we also found that some other morphological shapes that it had not been paid attention to before have taxonomic, such as stigmas, trunk, and pericarp thickness also have certain taxonomic value among species.
Although the study of pollen morphology is becoming more common, most researchers have only studied the external morphology of pollen and seldom studied its internal. The results of this study showed that the pollen shape, polar view, equatorial view, pollen aperture, pollen wall ornamentation, and ridge of five sect. Tuberculata plants did not differ much. This is the same as with the results of the previous study [30,31]. However, the results of the present study can deepen the dissection of species morphology, although they do not provide strong evidence for species delimitation.
The whole chloroplast genome not only enriches the molecular phylogenetic database of wild plant resources but also plays an important role in evaluating the genetics and evolution of species and their origins [11]. The result of chloroplast genetic phyloenetic trees showed C. anlungensis and C. leyeensis were merged (BS = 100%, PP = 1.0). but C. pyxidiacea, C. rubituberculata and C. acutiperulata were located on different branches, further supporting that they were treated as independent species. In addition, in the field investigation, we found that some species with close geographical distribution maybe have natural hybridization. Therefore, we will increase the study of nuclear gene fragments in the next study to analysis their network relationships.
Based on the above research, our consider that C. leyeensis and C. anlungensis should be merged supporting the view of Min et al. (1993). C. acutiperulata, C. pyxidiacea, and C. rubituberculata should be restored to the status of separate species and supporting the view of Chang et al. (1996) [2,5].
5. Conclusions
In this study, morphology, pollen morphology and chloroplast genomes evidences were employed to investigate five species of sect. Tuberculata in the Panjiang Basin of China. Consequently, C. leyeensis should be treated as a single name of C. anlungensis; while C. acutiperulata, C. pyxidiacea, and C. rubituberculata were regarded as independent species. The study not only increases us understanding for Camellia species within the distribution area of the Panjiang Basin but also clarify the confusion of the classification of these plants in sect. Tuberculata to a foundation for resource development and protection.
Author Contributions
Conceptualization,.X.X.and Z.L.; methodology, X.X. and Z.R.; software, L.H. and X.X.; validation, Z.L. and M.T.; investigation, Z.R. and X.X., resources, C.Y.; data processing, Z.R.; writing—original draft preparation, X.X. and Z.R.; writing—review and editing, Z.L., Z.R. and X.X.; visualization, Z.L.; supervision, M.T. and Z.L.; project administration, Z.L.; funding acquisition, Z.L.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32400179), This study was supported by the Guizhou Provincial Basic Research Program (Natural Science) 2022 (072).
Data Availability Statement
GenBank accession numbers: OQ556869, OP0586859, NC_050354, MZ766253, and OK046127. All data and materials used in this study are included in this paper.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chang, H. T. A taxonomy of the genus Camellia. Editorial department of journal of Sun Yatsen University. 1981, 47–52. [Google Scholar]
- Chien, S.S. Four new ligneous plants of Szechuan. Contrib. Biol. Lab. Sci. Soc. China Bot. 1939, 12, 89–100. [Google Scholar]
- Min, T.L.; Zhong, Y.C. A revision of genus Camellia sect. Tuberculata. Acta Botanica Yunnanica. 1993, 15, 123–130. [Google Scholar]
- Min, T.L. A systematic synopsis of the genus Camellia. Acta Botanica Yunnanica. 1999, 21, 149–159. [Google Scholar]
- Chang, H.T.; Ren, S.X. Diagnosis on the systematic development of Camellia Ⅵ. revised on sect. Tuberculata of Camellia. supplement to the journal of Sun Yatsen University. 1996, 2: 55-60.
- Chang, H.T. New record of Camellia from South China. Acta Scientiarum Naturaliun Universitatis Sun Yatseni. 1984, 23, 77–82. [Google Scholar]
- Sealy. A revision of the genus Camellia. London: the Royal Horticulture Society. 1958, 1-239.
- Jiang, B.; Peng, Q.F.; Shen, Z.G.; Moller, M.; Pi, E.X.; Lu, H.F. Taxonomic treatments of Camellia (Theaceae) species with secretory structures based on integrated leaf characters. Plant Syst Evol. 2010, 290, 1–20. [Google Scholar] [CrossRef]
- Jiang, Z. D. Preliminary study of molecular phylogenetics and biogeography of the genus Camellia L. based on chloroplast DNA. Zhejiang Sci-Tech University. 2017.
- Wu, Q.; Tong, W.; Zhao, H.J.; Ge, R.H.; Li, R.P.; Huang, J.; Li, F.D. Comparative transcriptomic analysis unveils the deep phylogeny and secondary metabolite evolution of 116 Camellia plants. The Plant Journal. 2022, 111, 406–421. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X. Revision of the classification and comprehensive evaluation of the ornamental value of plants in the Camellia L. sect. Tuberculate. Guizhou University. 2023.
- Ran, Z.H.; Li, Z.; Xiao, X.; An, M.T.; Yan, C. Complete chloroplast genomes of 13 species of sect. Tuberculata Chang (Camellia L.): genomic features, comparative analysis, and phylogenetic relationships. BMC Genomics. 2024, 25, 108. [Google Scholar] [CrossRef]
- Lin, M.J.; Lu, Q.M. New record of Guizhou Camellia. Act. Sci. Nat. Univ. Sunyats. 1984, 83–85. [Google Scholar]
- Xu, Z.R.; Chen, F.; Deng, C.Y. A new species of sect. Tuberculata. Guihaia. 1987, 19–21. [Google Scholar]
- Chang, H.T.; Ren, S.X. A classification on the section Tuberculata of Camellia. Act. Sci. Nat. Univ. Sunyats. 1991, 30, 86–91. [Google Scholar]
- Erdtman, G. Handbook of palynology. Munksgaard, Copenhagen. 1978.
- Wang, K.F. & Wang X.Z. Introduction to sporology. Beijing: Peking University Press. 1983.
- Halbritter, D.D., Ulrich, D.S., Grímsson, D.F., Weber, P.D., Zetter, P.D., Hesse, P.D., Buchner, D.R., Svojtka, M.M., & Frosch-Radivo, A. Illustrated Pollen Terminology. Cambridge International Law Journal. 2018.
- Li, J.L.; Wang, S.; Yu, J.; Wang, L.; Zhou, S.L. A modified CTAB protocol for plant DNA extraction. Chinese Bulletin of Botany. 2013, 48, 72–78. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y. , Li, D.Z. GetOrganelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Bock, R. Organellar Genome DRAW (OGDRAW): a tool for the easy generation of highquality custom graphical maps of plastid and mitochondrial genome. Current Genetics 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Amyiryousefi, A.; Hyvonen, J.; Poczai, P. IRscope: an online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT: iterative refinement and additional methods. Methods Mol Biol. 2014, 1079, 131–46. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.; Minh, B.Q. IQTREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754-755. [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Research. 2021, 49, 293–296. [Google Scholar] [CrossRef]
- Efroni, I.; Eshed, Y.; Lifschitz, E. Morphogenesis of simple and compound leaves: a critical review. Plant Cell. 2010, 22, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.X.; Zavada, M.S.; Min, T.L. Pollen morphology of Camellia (Theaceae) and ITS taxonomic significance. Acta Botanica Yunnanica. 1992, 14, 275–282. [Google Scholar]
- Hu, Z.M.; Zhao, C.H.; Zhao, Y.Y.; Liu, J.X. Pollen morphology of Liliaceae and its systematic significance. Palynology 2021, 45, 531–568. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



