
Submitted:
03 September 2024
Posted:
03 September 2024
You are already at the latest version
Abstract
Obesity and chronic inflammation are well-known risk factors for cancer. Interleukin-33 (IL-33) functions in inflammatory diseases, particularly those related to obesity and cancer. However, the correlation and molecular mechanisms linking IL-33, obesity, and cancer remain unclear. IL-33 is a nuclear cytokine released as an alarm signal when cells are damaged or stressed. It is expressed in epithelial, endothelial, and fibroblast-like cells during normal conditions and in-flammation. When IL-33 is present, immune cells expressing the IL-1RL1/ST2 receptor are alerted in response to tissue damage or cell injury. Research on IL-33 and its correlation with obesity has revealed improvements in inflammatory and metabolic changes associated with obesity. IL-33 can have both pro-tumorigenic and antitumorigenic effects, depending on the cellular content, in-flammatory environment, expression levels, and bioactivity of IL-33. This review will discuss the various functions of IL-33 in obesity-driven inflammation and cancers and how they intersect.
Keywords:
IL-33
; ST2
; Immune Cells
; Inflammation
; Obesity
; Cancer
1. Introduction
IL-33 is a nuclear cytokine from the IL-1 superfamily that has been extensively studied since its discovery nearly two decades ago [1]. Initially recognized for its role in inducing type 2 immune responses [2, 3], IL-33 is a tissue-derived cytokine with diverse funcitons. It acts as both a traditional cytokine and an intracellular nuclear factor [3, 4], playing a crucial role in cell communication and addressing inflammation within the cell [4]. Since its discovery, IL-33 has been associated with various health conditions, including asthma, airway inflammatory diseases [5], organ fibrosis [6], kidney injury [7], pediatric heart disease and transplantation [8], rheumatoid arthritis [9, 10], and neurodegenerative diseases[11]. Additionally, IL-33 is essential in obesity [12, 13] and various cancers such as hematologic malignancies [14] and solid cancers [15-19].
Chronic inflammation is long-lasting and can persist for several months to years [20]. It can occur due to various reasons [20]. Chronic inflammatory diseases are the leading cause of death worldwide, as identified by the World Health Organization (WHO), which ranks chronic diseases as the greatest threat to human health. The prevalence of specific chronic inflammation-mediated diseases includes diabetes, cardiovascular diseases, arthritis and joint diseases, allergies, and chronic obstructive pulmonary disease (COPD).
Obesity is a characterized by persistent low-grade inflammation, which can lead to various metabolic diseases such as insulin resistance, Type 2 diabetes, hypertension, cardiovascular disease, dyslipidemia, and even cancer [21]. During obesity, communication through cytokines of the TNF family triggers cell death and inflammation in the adipose tissue, causing an overflow of lipids, glucotoxicity, and insulin resistance [21]. Research has shown that approximately 4-8% of all cancers are linked to obesity [22]. Additionally, obesity has been associated with several types of cancer, including breast, colorectal, kidney, gallbladder, prostate, and liver. Studies have indicated promising findings related to IL-33 and these types of cancer [23]. Understanding the complex connections between obesity and cancer, as well as how IL-33 influences them, could be crucial for the future management of these conditions, offering hope in the fight against these diseases.
2. IL-33 and Its Receptor Biology
2.1. IL-33
The IL-33 gene is located on chromosome 9p24.1 in humans and chromosome 19qC1 in mice. The human IL-33 gene has eight exons, with exon one being non-coding and 2-8 coding for the IL-33 protein, covering over 42kb of genomic DNA [24]. The IL-33 protein consists of an N-terminal domain containing a chromatin-binding motif, a nuclear localization signal, a central domain, and a C-terminal IL-1-like cytokine domain that gives IL-33 cytokine-like properties [1, 3]. The nuclear localization signal is within amino acids 46-67 [25]. The binding capacity to DNA determines the functions of full-length IL-33 in the nucleus as a transcription factor [26]. In mice, two different promoters initiate IL-33 gene transcription, resulting in two IL-33 transcripts (Il33a and Il33b) with differing 5’-untranslated regions but encoding the same protein. The human IL-33 mRNA encodes a protein containing 270 amino acids, while the mouse IL-33 protein comprises 266 amino acids, and the two proteins are 55% similar. IL-33 possesses a three-dimensional structure similar to other family members, consisting of 12 β-strands arranged in a β-trefoil fold [27, 28]. IL-33 possesses a binding area with a combination of polar and nonpolar regions to create a specific binding between the receptor and ligand [28]. Research indicates that the IL-33 protein is highly conserved in mammals and closely related to IL-18 among the IL-1 family members.
2.2. IL-33 Receptor
The IL-33 receptor, also known as serum stimulation-2 (ST2), interleukin 1 receptor-like 1 (IL1RL1), or T1, was initially discovered in 1989 and was considered an orphan receptor until the identification of IL-33 [29, 30]. There are multiple isoforms of ST2, including membrane-bound (ST2L) and soluble form (sST2). ST2L consists of three immunoglobulin-like motifs responsible for extracellular ligand-binding (extracellular domain), an intracellular domain, and a transmembrane domain. ST2L’s extracellular domain binds IL-33 with the help of IL-1 receptor accessory protein (IL-1RAcP), which forms the transmembrane IL-33 receptor complex and recruits downstream signaling components via its Toll/interleukin-1 receptor (TIR) domain [2]. sST2 may act as a decoy receptor for IL-33 [31] due to its lack of intracellular and transmembrane domains but possesses a C-terminal sequence [32]. The IL-1R8 receptor, known as SIGIRR (single immunoglobulin IL-1R-related receptor) or TIR8 [33], is a negative regulator of the IL-33/ST2 signaling pathway [34]. It can dimerize with ST2L to reduce the biological effects of IL-33.
2.3. IL-33 and ST2 Distribution
2.3.1. IL-33 and ST2 Expression in Immune Cells
IL-33 is typically found in the stroma and is released by damaged or necrotic barrier cells such as endothelial and epithelial cells [35]. It is expressed at high levels in the steady-state nuclei of various cell types [24], including immune cells like resting dendritic cells (DCs) [36], activated macrophages [37], monocytes, regulatory T cells (Tregs), and mast cells [37], endothelial cells, epithelial cells of barrier tissues such as lung, intestine, skin, and fibroblasts, glial cells, astrocytes, smooth muscle cells and platelets. IL-33 is a ubiquitous and crucial immune modulator that influences various immune cell responses [38]. It acts as an alarm signal (alarmins or danger signals), rapidly released from producing cells in response to cellular stress and injury [39].
ST2 is expressed in T-helper 2 (Th2) cells, mast cells [32, 40, 41], cardiomyocytes [42], and other immune cells [35, 43], including innate lymphoid cell type 2 (ILC2)[44], Tregs [45], CD4+ and CD8+ T cells [46], NKT cells [47, 48], and myeloid cells, including basophils, eosinophils and macrophages [37]. Recently, ST2 was reported to be expressed in Th1 cells [49, 50], though the expression level is low and transient upon virus infection[51].
2.3.2. IL-33 and ST2 Immune Cell Distribution in Adipose Tissue
IL-33 and ST2 can be found abundantly in adipose tissue, including preadipocytes, adipocytes, and endothelial cells. Infiltrating immune cells positive for IL-33 and ST2 are also plentiful in adipose tissue. Adipose tissue is a crucial for regulating energy balance and is generally categorized as white adipose tissue (WAT) and brown adipose tissue (BAT) [52-54]. Additionally, a type of adipose tissue known as brown-in-white (brite)/beige adipose tissue (BeAT) shares characteristics with both WAT and BAT regarding morphology and function [55]. Immune cells and their mediators have been identified as significant regulators of metabolism, leading to a new paradigm in immune regulation of metabolism [56]. Innate immune cells present in adipose tissue include macrophages, neutrophils, eosinophils, dendritic cells (DCs), innate lymphoid cells (ILCs), and natural killer (NK) cells [57]. The most common immune cells in lean adipose tissue are alternatively activated macrophages (M2-like macrophages), type 2 innate lymphoid cells (ILC2s), eosinophils, Tregs, and Th2 cells [58, 59], all contributing to an anti-inflammatory environment. However, during the development of obesity adipocyte hypertrophy, the immune cell populations undergo dynamic changes. Most immune cells increase in adipose tissue, leading to an inflammatory condition supporting insulin resistance. Exceptions to this increase include Tregs, eosinophils [56], and Th2 cells [60] (Figure 1).
2.3.3. IL-33 and ST2 Immune Cell Distribution in the Tumor Microenvironment
The tumor microenvironment (TME) varies across different types of tumors, but typically includes immune cells, stromal cells, blood vessels, and extracellular matrix [61], and actively promotes cancer progression. Depending on the circumstances, immune cells play a crucial role in the TME and can either inhibit or support tumor growth [62]. ST2 is expressed in various immune cells, and IL-33 is released from damaged structural cells. IL-33/ST2 signaling is involved in various cancers, exerting both protumor and antitumor functions within the environment [63].
3. IL-33/ST2 Signaling Pathway
IL-33 has a dual role, acting as a nuclear factor by binding to chromatin to suppress inflammatory responses and as a cytokine released into the extracellular space in response to cell damage or mechanical injury [4]. This dual functionality underscores its crucial role in immune modulation. IL-33 exerts its cytokine activity by binding to the primary receptor ST2 and then recruiting the accessory receptor, IL-1RAcP [1, 41, 64, 65].
When IL-33 binds to the IL-33 receptor, forming the ST2L and IL-1RAcP heterodimeric complex, it initiates a signaling pathway. Following receptor binding, the TIR domain dimerization starts a pathway that includes the myeloid differentiation primary response protein 88 (MyD88), IL-1R-associated kinase 1 (IRAK1), IRAK4, tumor necrosis factor (TNF) receptor-associated factor 6 (TRAF6), mitogen-activated protein kinases (MAPKs), or nuclear factor-κB (NF-κB) [24].
The intracellular signaling pathways initiated by IL-33 activate a wide array of downstream effectors. These effectors play a significant role in immune modulation and inflammatory responses [38, 39], cell survival [66, 67], proliferation [17], differentiation [17], apoptosis[66], angiogenesis [68], migration [69], metabolic regulation [70], and metastasis [71] (Figure 2).
4. Effect of Il-33 in the Tumor Microenvironment
The cytokine IL-33 has a dual role in the tumor microenvironment (TME). It can both promote and inhibit tumor growth. Its pro-tumor effects involve the accumulation of immune-suppressive cells. On the other hand, it can inhibit tumor growth by working with the T cell receptor (TCR) and IL-12 signaling to enhance the effectiveness of CD8+ T cells [72]. Additionally, IL-33 can amplify Th1 CD4+ T cells and Th2-type responses through its activity on various immune cells such as human basophils, allergen-reactive Th2 cells, iNKT, NK cells [73], eosinophils, and ILC2 [74].
The IL-33/ST2 pathway is mainly involved in Th2 responses. When IL-33 activates Th2 receptors, it produces cytokines such as IL-4, IL-5, and IL-13 [84]. These cytokines from Th2 cells enhance chemokine production in epithelial cells [85-87]. The IL-33 gene and the cytokines are associated with Th2-related diseases, protecting against intracellular pathogens, promoting tissue repair, and assisting with chronic inflammatory diseases [84, 88]. IL-4, a critical Th2 cytokine, is primarily produced by activated T cells, mast cells, basophils, and eosinophils. It's crucial function is transforming naive CD4+T cells into Th2 cells, initiating allergic reactions, and triggering immune responses against extracellular parasites [89]. This role of IL-4 is particularly relevant to the development of atopic dermatitis and asthma [90]. IL-4's role in asthma is further highlighted by its promotion of airway inflammation and hyperresponsiveness within the cell, exacerbating the symptoms and severity of the condition [90]. Similarly, in the case of dermatitis, IL-4's promotion of IgE antibodies disrupts the skin barrier function, leading to eczematous lesions in affected individuals [87]. Il-4 also directly affects other diseases like inflammatory bowel disease (IBD), contributing to chronic inflammation and fibrosis by promoting tissue remodeling, scarring, and autoimmune diseases [87].
IL-4 also promotes the proliferation and survival of several cancer cells [91], being over-expressed by many human tumor types, including malignant glioma, ovarian, lung, breast, pancreatic, colon, and bladder carcinomas, which also overexpress its receptors (IL-4R)[92-94]. While several therapeutics have been developed for asthma and inflammatory disease treatment, their non-efficacy has stopped their development. However, most therapeutics showed good tolerance and high safety in humans, making developing clinical trials for cancer therapy possible [91].
IL-5, a significant player in developing allergic reactions and inflammation, regulates the production, maturation, activation, and survival of eosinophil progenitors in the blood [95]. Some T lymphocytes mainly make it. IL-5 causes B lymphocytes to make more antibodies and increases the number of eosinophils in the airways. Its primary role is to promote the activation and longevity of eosinophils, aiding in their migration from the blood to the airways [90]. By promoting the production of antibodies, particularly immunoglobulin A (IgA) antibodies, which are crucial for mucosal immunity, IL-5 can also influence B cell activity [90]. IL-5, made in the laboratory, is used as a biological response modifier to boost the immune system in cancer therapy.
IL-13, a crucial factor in developing allergic inflammation [96], closely interacts with IL-5 and significantly impacts airways by promoting mucus production [97]. IL-13 encourages goblet cells in the airway to produce mucus, leading to airway blockage and mucus plugging and directly contributing to asthma [97] [98]. Additionally, IL-13 is overexpressed in a variety of solid tumors and is correlated with poor prognosis in glioblastoma, colorectal cancer, adrenocortical carcinoma, pancreatic cancer, and breast cancer [99].
It is crucial to recognize the diverse effects of IL-33 on immune responses in the context of cancer [75]. For instance, IL-33 can activate ILC2 in the lung, promoting tumor burden [76]. This activation leads to innate type 2 inflammation and suppression of IFN-γ production and cytotoxic function of lung NK cells [76]. In melanoma, IL-33 overexpression has been found to inhibit lung metastases by activating CD8+ T cells and NK cells [77]. On the other hand, induced tumoral expression of IL-33 can promote anti-melanoma immune responses through IFN-γ-producing CD8+ T cells and NK cells [78], offering a ray of hope in cancer therapy. Eosinophils [79] and DCs [80] could also mediate the anti-melanoma effects of exogenous IL-33. In breast cancer, IL-33 secreted by cancer-associated fibroblasts enhances ILC2 and Th2 type responses, induces TCR-independent secretion of IL-13, and recruits immunosuppressive granulocytes [81]. Moreover, IL-33 activates intrinsic signaling in Treg cells, necessary for their immunosuppressive action in cancer [82]. When combined with the pro-inflammatory cytokine IL-12, IL-33 boosts the production of the Th1 cytokine IFN-γ [72]. Overall, the IL-33/ST2 signaling pathway significantly impacts the TME in cancer, impacting immune effector cells and controlling the recruitment of cells that either promote or inhibit tumor growth [83]. Due to its immunomodulatory activity, IL-33 can synergize with various cancer therapies, including immune checkpoint blockade and chemotherapy. This aspect has been recently reviewed [74]. This information focuses on the effects of IL-33/ST2 on immune cells and stroma cells in the TME (Figure 3).
4.1. Effect of Il-33 on Neutrophils, Eosinophils, Mast Cells, and Basophils
IL-33 exerts a range of effects on innate immune cells, such as neutrophils, eosinophils, mast cells, and basophils, depending on the specific disease context [100]. It can promote inflammatory or regulatory responses in neutrophils and eosinophils, contributing to tissue repair or immunosuppression [101]. Studies have shown that IL-33 directly acts on murine neutrophils, promoting their recruitment and activation in inflamed tissues [101]. In cancer, IL-33-driven neutrophils are associated with pro-inflammatory and regulatory functions.
Eosinophils, known for their role in host defense against parasites and infections, are activated by IL-33 for allergic reactions and tissue repair [101]. In the context of cancer, Il-33 has been linked to utilizing eosinophils for antitumor effects, such as inhibiting growth and preventing metastasis. IL-33-activated eosinophils express more effector molecules, eosinophil cationic protein (ECP), eosinophil peroxidase (EPX), and granzymes, as well as integrin CD11b/CD18 immune synapse, leading to the killing of tumor cells in vitro and in vivo [102, 103].
Mast cells, immune cells that respond to IL-33, accumulate in tumors and their microenvironment during disease progression[104]. They are involved in tumor growth, and their mediators exert both pro- and antitumorigenic roles in different human cancers [105]. IL-33 activation of mast cells and basophils may impact tumor progression by modulating cytokine release and triggering immune complex activation. According to recent studies, tumor-infiltrating mast cells activated by IL-33 secrete IL-2 and promote the expansion of ICOS+ Tregs, leading to gastric cancer progression [106].
Basophils, produced in the bone marrow, are white blood cells that play a crucial role in maintaining a healthy immune system. Abnormal levels of basophils may indicate inflammation, hyperthyroidism, or other health conditions. IL-33 can activate murine and human basophils, increasing histamine and cytokine production in vitro and promoting their expansion in vivo[107-111]. Notably, there is still a limited association between basophils and cancer and IL-33 regulation [101] [112].
4.2. Effect of Il-33 on Macrophages and Dendritic Cells
Macrophages and DCs play crucial roles in the innate immune system, particularly in tumor immunity and immunology [113]. IL-33 is a significant regulator of DC and macrophage activity within the tumor microenvironment, exhibiting both pro- and antitumorigenic actions [113]. Notably, IL-33 is crucial in promoting M2-like polarization within macrophages and is associated with immunosuppressive and tumor-promoting properties [113]. This process involves the release of anti-inflammatory cytokines and the stimulation of tumor angiogenesis, invasion, and metastasis [112, 114].
IL-33 influences DCs in addition to its impact on macrophages. DCs activated by IL-33 exhibit increased cytokine production and enhanced antigen presentation capability, promoting T cell activation and antitumor immune responses. However, the presence of conflicting studies regarding IL-33's influence on DC-mediated antitumor immunity suggests that its function in DCs may vary depending on its location within the body [112]. This discrepancy underscores the need for further research in this area.
Furthermore, with the assistance of dectin1-activated DC, IL-33 can induce Th9 cell differentiation and enhance antitumor efficacy [115, 116]. While some research indicates that IL-33 improves T cell activation and DC-mediated antigen presentation, other studies argue that IL-33 may cause DCs to acquire tolerogenic characteristics, inhibiting the immune system's ability to combat cancer [112].
4.3. Effect of Il-33 on CD8+ T, NK and NKT Cells
Several studies demonstrated that IL-33 expression positively correlated with CD8+ T cell and natural killer (NK) recruitment and activation in the TME. CD8+ T, NK, and natural killer T (NKT) cells are effective tumor-killer cells. They share many phenotypes and functions for antitumor immunity [117]. IL-33 directly activated NK and NKT cells, leading to the production of interferon-gamma (IFN-γ) and the promotion of Th1 immunity [118]. It also targets immunoregulatory invariant natural killer T (iNKT) cells to enhance their activation state [47]. This activation of NK and NKT cells induces the production of IFN-γ and promotes Th1 immunity in both human and mouse NK cells [119].
When combined with IL-12, IL-33 activates NK cells, enhancing their ability to destroy tumor cells through cytotoxicity. In preclinical models, IL-33 has been found to support the recruitment and activation of cytotoxic NK cells in the tumor microenvironment, hindering the formation of metastasis and suppressing tumor progression [112]. However, it is crucial to note that conflicting reports suggest that in specific tumor microenvironments, IL-33/ST2 signaling may hinder NK cell activation. This underscores the importance of considering the context-dependent effects of IL-33 on NK cell-mediated antitumor responses. This factor should be a key consideration in future research and clinical applications, highlighting the need for further investigation and careful application [120].
4.4. Effect of Il-33 on Myeloid-Derived Suppressor Cells
IL-33 significantly impacts myeloid-derived suppressor cells (MDSCs), potent mediators of immune suppression within the tumor [121]. MDSCs play an essential role in cancer by helping tumors evade the immune system and accelerating disease progression [121]. IL-33 partially controls the function and development of MDSC within the tumor [121]. In the TME, IL-33 reduces apoptosis and supports the survival of MDSCs by inducing autocrine secretion of GM-CSF, creating positive feedback for MDSC accumulation [121]. The accumulation and promotion of MDSCs are associated with tumor development and increased immunosuppression due to IL-33. Conversely, it has been demonstrated that in some situations, IL-33 inhibits the growth of MDSCs and reduces their immunosuppressive abilities, leading to enhanced antitumor immune responses and tumor regression [112, 121].
4.5. Effect of Il-33 on T helper Cells
IL-33 plays a significant role in regulating the development and function of CD4+ T cells, including CD4+ T helper (Th) cells and CD4+ Treg cells [100, 122]. Early in T cell activation, it stimulates Th1 differentiation and works with IL-12 to improve Th1 polarization [100, 122]. IL-33 may also promote Th1 cell differentiation depending on IL-12 and ST2 [122]. MyD88 is necessary for IL-33 and IL-12-induced Th1 cell development and IFN-γ production [122]. Additionally, IL-33 supports the proliferation of Th cells that produce IL-9 (Th9), aiding robust immune responses against tumors.
Conversely, IL-33 signaling is known to promote the proliferation of suppressive CD4+ Foxp3+ GATA3+ Treg cells, particularly in tumor-specific environments high in IL-33 [40, 122]. These Treg cells play a complex regulatory role, potentially limiting antitumor immunity and exhibiting vigorous suppressor activities [112]. In preclinical models, IL-9-producing Th9 cells are crucial for initiating strong immune responses against tumors and assisting in tumor regression. Research on Th-cell differentiation suggests that IL-33 has context-dependent effects on Th1- and Th2-skewing Th-cell differentiation, a complexity that underscores the intricacies of immune system regulation [112, 122]. The unique microenvironmental cues in the tumor milieu may affect the signaling pathway's function in Th cell development.
Th17 cells play a significant role in inflammation and are implicated in various autoimmune diseases [123]. They are influenced by different molecular signals within the TME [124]. This regulation affects the proliferation, differentiation, metabolic reprogramming, and phenotypic transformation of Th17 cells, leading to their dual impact on tumor progression [124]. A recent study indicates that during inflammation, Th17 cells accumulating in the small intestine express the IL-33 receptor (ST2), with intestinal epithelial cells (IEC) being the primary source of the alarm IL-33 [125]. When exposed to IL-33, both mouse and human Th17 cells show reduced expression of pro-inflammatory genes and increased IL-10. This anti-inflammatory cytokine can limit inflammation in the intestine and control previously committed pathogenic Th17 cells. The study showed that in response to IL-33, Th17 cells acquire an immunosuppressive property. The IL-33/ST2 axis is likely crucial in controlling pathogenic Th17 cells in the small intestine to sustain homeostasis [125].
4.6. Effect of Il-33 on CD4+ Treg Cells and in Tumor Immunity
IL-33 signaling affects effector Th cells and the growth and activity of regulatory T (Treg) cells in the TME. Research has shown that suppressive CD4+ Foxp3+ GATA3+ Treg cells proliferate in response to ST2/IL-33 signaling, both in vivo and in vitro, aiding immunosuppression and tumor immune evasion [112]. The generation of IL-2 by DCs and the promotion of transforming growth factor (TGF)-β1-mediated Treg cell development are two of the processes by which IL-33 signaling leads to the expansion of ST2+ Treg cells [112, 126]. These IL-33-expanded Treg cells are found in both immune and non-immune tissues, where they have a solid suppressor effect that helps create an immunosuppressive tumor microenvironment[127] .
In the lungs, CD4+Foxp3+ Treg cells expressed the IL-33 receptor ST2 [128]. When exposed to IL-33, Treg cells increased their canonical Th2 transcription factor GATA3 and ST2 expression and produced type 2 cytokines. This exposure to IL-33 led to a significant shift in the immune response, as Treg cells lost their ability to suppress effector T cells [128]. Moreover, IL-33 has been implicated in promoting the accumulation and maintenance of ST2+ Treg cells in inflamed tissues, further exacerbating immunosuppression and facilitating tumor progression [112].
The complex interaction between IL-33 signaling and CD4+ T cell subsets emphasizes its significance in supporting antitumor immune responses [112]. By stimulating the growth of suppressive Treg cells and modifying Th cell differentiation, IL-33 regulates adaptive immune responses inside the tumor. The crucial role of IL-33 in promoting Treg cell expansion not only highlights its potential as a target for therapeutic intervention in cancer immunotherapy but also underscores the urgency of understanding and utilizing IL-33 in cancer treatment [116].
4.7. IL-33-Expressing Cells and the Effects of IL-33 on Non-Immune Cells in the Tumor Microenvironment
The impact of IL-33 depends on the cells that produce it in the TME. These cells include immune cells, stromal cells, non-tumor epithelial cells, and transformed (epithelial) tumor cells, all of which are essential sources of IL-33 in the TME [35]. Fibroblasts play a significant role in the TME. The dysregulation of fibroblasts during cancer creates an environment that promotes tumor growth by remodeling the extracellular matrix and releasing pro-angiogenic factors [129, 130]. Studies have shown that cancer-associated fibroblasts can induce IL-33 [131].
Additionally, stromal cells have been identified as the primary non-immune cell source of IL-33, and IL-33 derived from stromal cells has been linked to tumor metastasis [75, 129, 132, 133]. Analysis of The Cancer Genome Atlas (TCGA) data and mRNA sequencing from TCGA revealed that IL-33, expressed in endothelial and epithelial cells, could also contribute to tumor immune responses [75, 132]. Furthermore, IL-33 derived from non-tumor epithelial cells has been found to drive colorectal cancer tumorigenesis in APCMin/+ mice [134]. The expression of IL-33 changes during tumor progression, while IL-33 is highly expressed in normal epithelial cells; tumor development leads to the downregulation of IL-33 in epithelial cells but upregulation of IL-33 in the tumor stroma and serum [63]. Therapeutic overexpression of IL-33 in tumor epithelial cells promotes type 1 antitumor immune responses through CD8+ T cells and NK cells, while tumor stromal IL-33 stimulates immune tolerance and suppression via Treg and MDSC [63]. Overall, IL-33 expression is context-dependent; in some tumors, IL-33 is highly expressed in normal epithelial cells but downregulated in tumor cells in advanced carcinoma [63, 135, 136], but in other tumors, levels of IL-33 or ST2 protein expression were increased in cancer lesions compared to matched normal breast tissues [83, 137, 138].
5. Effect of IL-33 on Adipose Tissue
Obesity is associated with chronic inflammation, hormonal imbalances, insulin resistance, high insulin levels, adipokines, and immune function changes [139]. Hormones like adipokines from adipose tissue contribute to low-grade inflammation, leading to conditions like insulin resistance, diabetes, high blood pressure, and asthma, all associated with obesity [140]. Adipokines can stimulate cell growth, angiogenesis, and apoptosis, all of which are directly linked to cancer progression. These hormonal functions also extend to regulating appetite, energy metabolism, and inflammation, collectively making individuals more susceptible to cancer onset and advancement [141].
IL-33 is produced by various cells in adipose tissue, including endothelial cells, fibroblasts, and adipocytes [142-144]. ILC2, a source of type 2 cytokines IL-5 and -13, helps regulate beige fat biogenesis and maintain tissue homeostasis in adipose tissue [145, 146]. IL-33 is essential for maintaining ILC2 in WAT and limiting mouse adiposity by increasing caloric expenditure [147]. Adiponectin limits activation through AMPK-mediated feedback inhibition of IL-33 signaling [148]. IL-33 also has protective effects on adipose tissue inflammation during obesity in mice by inducing Th2 cytokines in WAT and promoting the polarization of WAT macrophages toward an M2 alternatively activated phenotype, leading to reduced adipose mass and fasting glucose [147]. In adipose tissue, IL-33 is crucial for maintaining immune cell balance, which has consequences for inflammation and obesity-related problems. Stimulating ILC2s in adipose tissue encourages the "beiging" of white adipocytes and raises energy expenditure, potentially preventing obesity and its related conditions [149]. However, the function of IL-33 in both immunity and metabolism is complex, characterized by a duality of pro- and anti-inflammatory properties.[150] In atherosclerosis sites, IL-33 is expressed by murine cardiac fibroblasts in response to stress from immune signaling by endothelial cells [150-152].
Research has shown mixed results about its precise function in the development of metabolic disorders like diabetes, obesity, and cardiovascular disease [150]. The IL-33/ST2 pathway is considered protective during obesity and manipulating IL-33 expression to induce Th2 cytokines and promote macrophage polarization may be a beneficial therapeutic strategy for treating or preventing type 2 diabetes in obese patients.
Interestingly, in the context of obesity, white adipose tissue shows increased IL-33 expression, while local ILC2 exhibit a reduction in abundance [150]. This imbalance underscores the importance of understanding potential disparities in regulating IL-33 at both tissue-specific and systemic levels amid obesity [150]. IL-33 also plays a role within adipose tissue to aid in the activation activity with immune cells and moderate adipose microenvironments, impacting specific inflammatory properties and metabolic processes. Its influence shapes the composition of immune cells within adipose tissue and promotes the activation and recruitment of immune cells like macrophages and endothelial cells [152]. Clarifying these intricacies is essential in comprehending the significant role of IL-33 in obesity-related pathophysiology [150].
6. Discussion of Obesity and Cancer Correlation
The latest scientific research confirms a clear connection between a high body mass index (BMI) and the risk of various types of cancer [22, 23]. Although obesity itself may not directly cause cancer, it does increase the risk of developing an aggressive, fast-growing form of cancer that is more difficult to treat. Therefore, addressing obesity is essential for reducing the likelihood of cancer complications [22].
Research suggests that the IL-33 protein plays diverse roles in adipose tissue and the tumor microenvironment (Figure 4). Further research is needed to fully understand the potential of IL-33 in regulating obesity and reducing cancer risk. However, the findings fo far are promising, offering a potential avenue for disease management.
Future research should focus on how IL-33 influences fat tissue function, energy expenditure, and immune modulation to identify potential therapeutic targets. Additionally, further investigating the influence of immune cells on antitumor and pro-tumor processes in different types of obesity-related cancer is crucial. Targeting IL-33 signaling pathways and downstream effects could help regulate obesity and exhibit antitumor properties. Therefore, IL-33's anti-inflammatory properties offer another potential avenue for intervention, as chronic inflammation is a critical factor in obesity and a significant driver of cancer development. IL-33-based therapies could reduce obesity-induced inflammation in fat tissue and other affected organs, lowering cancer risk.
Furthermore, conducting clinical trials and employing personalized medicine approaches is essential to assess the safety and effectiveness of IL-33-based interventions. This involves developing personalized treatment plans tailored to patients’ unique risk factors and metabolic profiles. A comprehensive understanding of IL-33’s role in immune regulation and metabolism is crucial for using it as a therapeutic target to manage obesity and reduce cancer risk. Collaboration among scientists, clinicians, and industry partners is vital for expanding our understanding of IL-33 biology and translating scientific discoveries into applications [43].
7. Conclusions and Prospects
Our current investigation into the diverse functions of the IL-33/ST2 axis in immune modulation, inflammation, obesity, tumor growth, and tumor suppression is of significant importance. Our understanding of the relationship between the IL-33/ST2 axis, obesity, and cancer has advanced significantly, underscoring the importance of our work. The intricate interplay between the IL-33/ST2 axis, the tumor microenvironment (TME), and its immune cells components, such as tumor-infiltrating lymphocytes, macrophages, and dendritic cells requires thorough investigation, especially in the context of tumors associated with obesity. A comprehensive understanding of how the IL-33/ST2 axis contributes to obesity-related cancer necessitates both animal studies and human research. While some promising results have been observed in animal models, there is still much to learn about the diverse roles of IL-33/ST2 in different types of obesity-related cancer and their responses to treatment, particularly in obese individuals.
Author Contributions
Conceptualization, Q.Z.; writing—original draft preparation, C.K., Q.Z.; writing—review and editing, C.K., Q.Z.; funding acquisition, Q.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Cancer Institute of the National Institutes of Health (NIH), grant number R01CA255802; the National Institute of General Medical Sciences of the National Institutes of Health, grant number P20 GM103629; the University Senate Committee on Research Fellowship Program Award; the Carol Lavin Bernick Faculty Grant and The APC was funded by XXX. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented are available in the references cited in this article.
Acknowledgments
Figures created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schmitz J, Owyang A, Oldham E, Song Y, Murphy E, McClanahan TK, et al. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity 2005; 23(5):479-490.
- Garlanda C, Dinarello CA, Mantovani A. The interleukin-1 family: back to the future. Immunity 2013; 39(6):1003-1018.
- Carriere V RL, Ortega N, Lacorre DA, Americh L, Aguilar L, Bouche G, Girard JP. IL-1-like cytokine ligand for ST2 receptor, is a chromatin-associated nuclear factor in vivo. Proc Natl Acad Sci U S A 2007; 104(287-2).
- Cayrol C, & Girard, J. P.. Interleukin-33 (IL-33): A nuclear cytokine from the IL-1 family. Immunological reviews 2018; 281(1):154–168.
- Gaurav R, Poole JA. Interleukin (IL)-33 immunobiology in asthma and airway inflammatory diseases. J Asthma 2022; 59(12):2530-2538.
- Di Carmine S, Scott MM, McLean MH, McSorley HJ. The role of interleukin-33 in organ fibrosis. Discov Immunol 2022; 1(1):kyac006.
- Chen WY, Li LC, Wu YH, Yang JL, Tzeng HT. Emerging Roles of Interleukin-33-responsive Kidney Group 2 Innate Lymphoid Cells in Acute Kidney Injury. Int J Mol Sci 2020; 21(4).
- Brunetti G, Barile B, Nicchia GP, Onorati F, Luciani GB, Galeone A. The ST2/IL-33 Pathway in Adult and Paediatric Heart Disease and Transplantation. Biomedicines 2023; 11(6).
- De la Fuente M, MacDonald TT, Hermoso MA. The IL-33/ST2 axis: Role in health and disease. Cytokine Growth Factor Rev 2015; 26(6):615-623.
- Yi XM, Lian H, Li S. Signaling and functions of interleukin-33 in immune regulation and diseases. Cell Insight 2022; 1(4):100042.
- Jia Z, Guo M, Ge X, Chen F, Lei P. IL-33/ST2 Axis: A Potential Therapeutic Target in Neurodegenerative Diseases. Biomolecules 2023; 13(10).
- Hasan A, Kochumon, S., Al-Ozairi, E., Tuomilehto, J., & Ahmad, R. Association between Adipose Tissue Interleukin-33 and Immunometabolic Markers in Individuals with Varying Degrees of Glycemia. Disease markers, 2019.
- de Oliveira MFA, Talvani A, Rocha-Vieira E. IL-33 in obesity: where do we go from here?Inflamm Res 2019; 68(3):185-194.
- Allegra A, Innao V, Tartarisco G, Pioggia G, Casciaro M, Musolino C, et al. The ST2/Interleukin-33 Axis in Hematologic Malignancies: The IL-33 Paradox. Int J Mol Sci 2019; 20(20).
- Song M, Yang J, Di M, Hong Y, Pan Q, Du Y, et al. Alarmin IL-33 orchestrates antitumoral T cell responses to enhance sensitivity to 5-fluorouracil in colorectal cancer. Theranostics 2023; 13(5):1649-1668.
- Chatrabnous N, Jafarzadeh A, Ghaderi A, Ariafar A, Aminizadeh N, Ghassabi F, et al. Association of elevated interleukin-33 serum levels with tumorstages in patients with prostate cancer. Eur Cytokine Netw 2019; 30(4):144-150.
- Stojanovic B, Gajovic N, Jurisevic M, Stojanovic MD, Jovanovic M, Jovanovic I, et al. Decoding the IL-33/ST2 Axis: Its Impact on the Immune Landscape of Breast Cancer. Int J Mol Sci 2023; 24(18).
- Liu N, Chen J, Zhao Y, Zhang M, Piao L, Wang S, et al. Role of the IL-33/ST2 receptor axis in ovarian cancer progression. Oncol Lett 2021; 22(1):504.
- Borovcanin MM, Vesic K. Breast cancer in schizophrenia could be interleukin-33-mediated. World J Psychiatry 2021; 11(11):1065-1074.
- Pahwa, R. Chronic Inflammation. In. Edited by Goyal A. StatPearls: Treasure Island (FL); 2022.
- Hildebrandt X, Ibrahim M, Peltzer N. Cell death and inflammation during obesity: "Know my methods, WAT(son)". Cell Death Differ 2023; 30(2):279-292.
- Arnold M, Pandeya N, Byrnes G, Renehan PAG, Stevens GA, Ezzati PM, et al. Global burden of cancer attributable to high body-mass index in 2012: a population-based study. Lancet Oncol 2015; 16(1):36-46.
- Pati S, Irfan W, Jameel A, Ahmed S, Shahid RK. Obesity and Cancer: A Current Overview of Epidemiology, Pathogenesis, Outcomes, and Management. Cancers (Basel) 2023; 15(2).
- Liew FY, Girard JP, Turnquist HR. Interleukin-33 in health and disease. Nat Rev Immunol 2016; 16(11):676-689.
- Luzina IG, Clerman A, Fishelevich R, Todd NW, Lockatell V, Atamas SP. Identification of the IL-33 protein segment that controls subcellular localization, extracellular secretion, and functional maturation. Cytokine 2019; 119:1-6.
- Cayrol C, Girard JP. The IL-1-like cytokine IL-33 is inactivated after maturation by caspase-1. Proc Natl Acad Sci U S A 2009; 106(22):9021-9026.
- Liu X, Hammel M, He Y, Tainer JA, Jeng US, Zhang L, et al. Structural insights into the interaction of IL-33 with its receptors. Proc Natl Acad Sci U S A 2013; 110(37):14918-14923.
- Lingel A, Weiss TM, Niebuhr M, Pan B, Appleton BA, Wiesmann C, et al. Structure of IL-33 and its interaction with the ST2 and IL-1RAcP receptors--insight into heterotrimeric IL-1 signaling complexes. Structure 2009; 17(10):1398-1410.
- Klemenz R, Hoffmann S, Werenskiold AK. Serum- and oncoprotein-mediated induction of a gene with sequence similarity to the gene encoding carcinoembryonic antigen. Proc Natl Acad Sci U S A 1989; 86(15):5708-5712.
- Tominaga, S. A putative protein of a growth specific cDNA from BALB/c-3T3 cells is highly similar to the extracellular portion of mouse interleukin 1 receptor. FEBS Lett 1989; 258(2):301-304.
- Kakkar R, Lee RT. The IL-33/ST2 pathway: therapeutic target and novel biomarker. Nat Rev Drug Discov 2008; 7(10):827-840.
- Gächter T, Werenskiold AK, Klemenz R. Transcription of the interleukin-1 receptor-related T1 gene is initiated at different promoters in mast cells and fibroblasts. J Biol Chem 1996; 271(1):124-129.
- Mariotti FR, Supino D, Landolina N, Garlanda C, Mantovani A, Moretta L, et al. IL-1R8: A molecular brake of anti-tumor and anti-viral activity of NK cells and ILC. Semin Immunol 2023; 66:101712.
- Bulek K, Swaidani S, Qin J, Lu Y, Gulen MF, Herjan T, et al. The essential role of single Ig IL-1 receptor-related molecule/Toll IL-1R8 in regulation of Th2 immune response. J Immunol 2009; 182(5):2601-2609.
- Cayrol C, Girard JP. Interleukin-33 (IL-33): A critical review of its biology and the mechanisms involved in its release as a potent extracellular cytokine. Cytokine 2022; 156:155891.
- Göpfert C, Andreas N, Weber F, Häfner N, Yakovleva T, Gaestel M, et al. The p38-MK2/3 Module Is Critical for IL-33-Induced Signaling and Cytokine Production in Dendritic Cells. J Immunol 2018; 200(3):1198-1206.
- Ishiguro N, Moriyama M, Furusho K, Furukawa S, Shibata T, Murakami Y, et al. Activated M2 Macrophages Contribute to the Pathogenesis of IgG4-Related Disease via Toll-like Receptor 7/Interleukin-33 Signaling. Arthritis Rheumatol 2020; 72(1):166-178.
- He PY, Wu MY, Zheng LY, Duan Y, Fan Q, Zhu XM, et al. Interleukin-33/serum stimulation-2 pathway: Regulatory mechanisms and emerging implications in immune and inflammatory diseases. Cytokine Growth Factor Rev 2024; 76:112-126.
- Cayrol C, Girard JP. IL-33: an alarmin cytokine with crucial roles in innate immunity, inflammation and allergy. Curr Opin Immunol 2014; 31:31-37.
- Lohning M SA, Coyle AJ, Grogan JL, Lin S, Gutierrez-Ramos JC, et al. T1/ST2 is preferentially expressed on murine Th2 cells, independent of interleukin 4, interleukin 5, and interleukin 10, and important for Th2 effector function. Proc Natl Acad Sci USA 1998.
- Xu D, Chan WL, Leung BP, Huang F, Wheeler R, Piedrafita D, et al. Selective expression of a stable cell surface molecule on type 2 but not type 1 helper T cells. J Exp Med 1998; 187(5):787-794.
- Weinberg EO, Shimpo M, De Keulenaer GW, MacGillivray C, Tominaga S, Solomon SD, et al. Expression and regulation of ST2, an interleukin-1 receptor family member, in cardiomyocytes and myocardial infarction. Circulation 2002; 106(23):2961-2966.
- Griesenauer B, Paczesny S. The ST2/IL-33 Axis in Immune Cells during Inflammatory Diseases. Front Immunol 2017; 8:475.
- Neill DR, Wong SH, Bellosi A, Flynn RJ, Daly M, Langford TK, et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature 2010; 464(7293):1367-1370.
- Schiering C, Krausgruber T, Chomka A, Fröhlich A, Adelmann K, Wohlfert EA, et al. The alarmin IL-33 promotes regulatory T-cell function in the intestine. Nature 2014; 513(7519):564-568.
- Pastille E, Wasmer MH, Adamczyk A, Vu VP, Mager LF, Phuong NNT, et al. The IL-33/ST2 pathway shapes the regulatory T cell phenotype to promote intestinal cancer. Mucosal Immunol 2019; 12(4):990-1003.
- Bourgeois E, Van, L. P., Samson, M., Diem, S., Barra, A., Roga, S., Gombert, M., Schneider, E., Dy, M., Gourdy, P., Girard, P., & Herbelin, A. The pro-Th2 cytokine IL-33 directly interacts with invariant NKT and NK cells to induce IFN-γ production. European Journal of Immunology 2009; 39(4):1046-1055.
- Zou L, Dang W, Tao Y, Zhao H, Yang B, Xu X, et al. THE IL-33/ST2 AXIS PROMOTES ACUTE RESPIRATORY DISTRESS SYNDROME BY NATURAL KILLER T CELLS. Shock 2023; 59(6):902-911.
- Baumann C, Bonilla WV, Fröhlich A, Helmstetter C, Peine M, Hegazy AN, et al. T-bet- and STAT4-dependent IL-33 receptor expression directly promotes antiviral Th1 cell responses. Proc Natl Acad Sci U S A 2015; 112(13):4056-4061.
- Dwyer GK, D'Cruz LM, Turnquist HR. Emerging Functions of IL-33 in Homeostasis and Immunity. Annu Rev Immunol 2022; 40:15-43.
- Brunner TM, Serve S, Marx AF, Fadejeva J, Saikali P, Dzamukova M, et al. A type 1 immunity-restricted promoter of the IL-33 receptor gene directs antiviral T-cell responses. Nat Immunol 2024; 25(2):256-267.
- Gesta S, Tseng YH, Kahn CR. Developmental origin of fat: tracking obesity to its source. Cell 2007; 131(2):242-256.
- Rosen ED, Spiegelman BM. What we talk about when we talk about fat. Cell 2014; 156(1-2):20-44.
- Frontini A, Cinti S. Distribution and development of brown adipocytes in the murine and human adipose organ. Cell Metab 2010; 11(4):253-256.
- Lee MJ, Wu Y, Fried SK. Adipose tissue heterogeneity: implication of depot differences in adipose tissue for obesity complications. Mol Aspects Med 2013; 34(1):1-11.
- Ferrante, AW. The immune cells in adipose tissue. Diabetes Obes Metab 2013; 15 Suppl 3(0 3):34-38.
- Michailidou Z, Gomez-Salazar M, Alexaki VI. Innate Immune Cells in the Adipose Tissue in Health and Metabolic Disease. J Innate Immun 2022; 14(1):4-30.
- Man K, Kallies A, Vasanthakumar A. Resident and migratory adipose immune cells control systemic metabolism and thermogenesis. Cell Mol Immunol 2022; 19(3):421-431.
- Mathis, D. Immunological goings-on in visceral adipose tissue. Cell Metab 2013; 17(6):851-859.
- Deiuliis J, Shah Z, Shah N, Needleman B, Mikami D, Narula V, et al. Visceral adipose inflammation in obesity is associated with critical alterations in tregulatory cell numbers. PLoS One 2011; 6(1):e16376.
- Anderson NM, Simon MC. The tumor microenvironment. Curr Biol 2020; 30(16):R921-R925.
- Truffi M, Sorrentino L, Corsi F. Fibroblasts in the Tumor Microenvironment. Adv Exp Med Biol 2020; 1234:15-29.
- Lu B, Yang M, Wang Q. Interleukin-33 in tumorigenesis, tumor immune evasion, and cancer immunotherapy. J Mol Med (Berl) 2016; 94(5):535-543.
- Ali S, Huber M, Kollewe C, Bischoff SC, Falk W, Martin MU. IL-1 receptor accessory protein is essential for IL-33-induced activation of T lymphocytes and mast cells. Proc Natl Acad Sci U S A 2007; 104(47):18660-18665.
- Chackerian AA, Oldham ER, Murphy EE, Schmitz J, Pflanz S, Kastelein RA. IL-1 receptor accessory protein and ST2 comprise the IL-33 receptor complex. J Immunol 2007; 179(4):2551-2555.
- Seki K, Sanada S, Kudinova AY, Steinhauser ML, Handa V, Gannon J, et al. Interleukin-33 prevents apoptosis and improves survival after experimental myocardial infarction through ST2 signaling. Circ Heart Fail 2009; 2(6):684-691.
- Zarezadeh Mehrabadi A, Shahba F, Khorramdelazad H, Aghamohammadi N, Karimi M, Bagherzadeh K, et al. Interleukin-1 receptor accessory protein (IL-1RAP): A magic bullet candidate for immunotherapy of human malignancies. Crit Rev Oncol Hematol 2024; 193:104200.
- Choi YS, Choi HJ, Min JK, Pyun BJ, Maeng YS, Park H, et al. Interleukin-33 induces angiogenesis and vascular permeability through ST2/TRAF6-mediated endothelial nitric oxide production. Blood 2009; 114(14):3117-3126.
- Artru F, Bou Saleh M, Maggiotto F, Lassailly G, Ningarhari M, Demaret J, et al. IL-33/ST2 pathway regulates neutrophil migration and predicts outcome in patients with severe alcoholic hepatitis. J Hepatol 2020; 72(6):1052-1061.
- Zeyda M, Wernly B, Demyanets S, Kaun C, Hämmerle M, Hantusch B, et al. Severe obesity increases adipose tissue expression of interleukin-33 and its receptor ST2, both predominantly detectable in endothelial cells of human adipose tissue. Int J Obes (Lond) 2013; 37(5):658-665.
- Kwon JW, Seok SH, Kim S, An HW, Choudhury AD, Woo SH, et al. A synergistic partnership between IL-33/ST2 and Wnt pathway through Bcl-xL drives gastric cancer stemness and metastasis. Oncogene 2023; 42(7):501-515.
- Yang Q, Li G, Zhu Y, Liu L, Chen E, Turnquist H, et al. IL-33 synergizes with TCR and IL-12 signaling to promote the effector function of CD8+ T cells. Eur J Immunol 2011; 41(11):3351-3360.
- Smithgall MD, Comeau MR, Yoon BR, Kaufman D, Armitage R, Smith DE. IL-33 amplifies both Th1- and Th2-type responses through its activity on human basophils, allergen-reactive Th2 cells, iNKT and NK cells. Int Immunol 2008; 20(8):1019-1030.
- Arrizabalaga L, Risson A, Ezcurra-Hualde M, Aranda F, Berraondo P. Unveiling the multifaceted antitumor effects of interleukin 33. Front Immunol 2024; 15:1425282.
- Peng L, Sun W, Wei F, Chen L, Wen W. Interleukin-33 modulates immune responses in cutaneous melanoma in a context-specific way. Aging (Albany NY) 2021; 13(5):6740-6751.
- Schuijs MJ, Png S, Richard AC, Tsyben A, Hamm G, Stockis J, et al. ILC2-driven innate immune checkpoint mechanism antagonizes NK cell antimetastatic function in the lung. Nat Immunol 2020; 21(9):998-1009.
- Gao K, Li X, Zhang L, Bai L, Dong W, Shi G, et al. Transgenic expression of IL-33 activates CD8(+) T cells and NK cells and inhibits tumor growth and metastasis in mice. Cancer Lett 2013; 335(2):463-471.
- Gao X, Wang X, Yang Q, Zhao X, Wen W, Li G, et al. Tumoral expression of IL-33 inhibits tumor growth and modifies the tumor microenvironment through CD8+ T and NK cells. J Immunol 2015; 194(1):438-445.
- Lucarini V, Ziccheddu G, Macchia I, La Sorsa V, Peschiaroli F, Buccione C, et al. IL-33 restricts tumor growth and inhibits pulmonary metastasis in melanoma-bearing mice through eosinophils. Oncoimmunology 2017; 6(6):e1317420.
- Dominguez D, Ye C, Geng Z, Chen S, Fan J, Qin L, et al. Exogenous IL-33 Restores Dendritic Cell Activation and Maturation in Established Cancer. J Immunol 2017; 198(3):1365-1375.
- Briukhovetska D, Dörr J, Endres S, Libby P, Dinarello CA, Kobold S. Interleukins in cancer: from biology to therapy. Nat Rev Cancer 2021; 21(8):481-499.
- Hatzioannou A, Banos A, Sakelaropoulos T, Fedonidis C, Vidali MS, Köhne M, et al. An intrinsic role of IL-33 in T. Nat Immunol 2020; 21(1):75-85.
- Yeoh WJ, Vu VP, Krebs P. IL-33 biology in cancer: An update and future perspectives. Cytokine 2022; 157:155961.
- Miller, AM. Role of IL-33 in inflammation and disease. Journal of Inflammation (London, England) 2011; 8(22).
- Milovanovic M, Volarevic V, Radosavljevic G, Jovanovic I, Pejnovic N, Arsenijevic N, et al. IL-33/ST2 axis in inflammation and immunopathology. Immunol Res 2012; 52(1-2):89-99.
- O'Donnell C, Mahmoud A, Keane J, Murphy C, White D, Carey S, et al. An antitumorigenic role for the IL-33 receptor, ST2L, in colon cancer. Br J Cancer 2016; 114(1):37-43.
- Kurowska-Stolarska M KP, Murphy G, Russo RC, Stolarski B, Garcia CC, Komai-Koma M, Pitman N, Li Y, Niedbala W. et al. IL-33 induces antigen-specific IL-5+ T cells and promotes allergic-induced airway inflammation independent of IL-4. J Immunol 2008:4780–4790.
- Kokubo K, Onodera, A., Kiuchi, M., Tsuji, K., Hirahara, K., & Nakayama, T. Conventional and pathogenic Th2 cells in inflammation, tissue repair, and fibrosis Frontiers in immunology 2022; 13.
- Stark JM, Tibbitt CA, Coquet JM. The Metabolic Requirements of Th2 Cell Differentiation. Frontiers in Immunology 2019; 10.
- Pelaia C, Paoletti, G., Puggioni, F., Racca, F., Pelaia, G., Canonica, G. W., & Heffler, E. Interleukin-5 in the Pathophysiology of Severe Asthma. Frontiers in Physiology 2019; 10.
- Setrerrahmane S, Xu H. Tumor-related interleukins: old validated targets for new anti-cancer drug development. Mol Cancer 2017; 16(1):153.
- Joshi BH, Leland P, Lababidi S, Varrichio F, Puri RK. Interleukin-4 receptor alpha overexpression in human bladder cancer correlates with the pathological grade and stage of the disease. Cancer Med 2014; 3(6):1615-1628.
- Prokopchuk O, Liu Y, Henne-Bruns D, Kornmann M. Interleukin-4 enhances proliferation of human pancreatic cancer cells: evidence for autocrine and paracrine actions. Br J Cancer 2005; 92(5):921-928.
- Todaro M, Lombardo Y, Francipane MG, Alea MP, Cammareri P, Iovino F, et al. Apoptosis resistance in epithelial tumors is mediated by tumor-cell-derived interleukin-4. Cell Death Differ 2008; 15(4):762-772.
- Pavord CEBaDDaID. Cytokine-Specific Therapy in Asthma. Middleton's Allergy (Eighth Edition) 2014:1491-1502.
- Gour N, & Wills-Karp, M. IL-4 and IL-13 signaling in allergic airway disease. Cytokine 2015; 75(1):68-78.
- A. SM. Interleukin-13 Stimulation Reveals the Cellular and Functional Plasticity of the Airway Epithelium. Annals of the American Thoracic Society 2018; 15(Suppl 2):S98–S102.
- Doran E, Cai, F., Holweg, C. T. J., Wong, K., Brumm, J., & Arron, J. R. Interleukin-13 in Asthma and Other Eosinophilic Disorders. Frontiers in medicine 2017; 4:139.
- Knudson KM, Hwang S, McCann MS, Joshi BH, Husain SR, Puri RK. Recent Advances in IL-13Rα2-Directed Cancer Immunotherapy. Front Immunol 2022; 13:878365.
- Alvarez F, Fritz, J. H., & Piccirillo, C. A. Pleiotropic Effects of IL-33 on CD4+ T Cell Differentiation and Effector Functions. rontiers in immunology 2019; 10(522).
- Zhou Z, Yan, F., & Liu, O. Interleukin (IL)-33: An orchestrator of immunity from host defence to tissue homeostasis. Clinical & Translational Immunology 2020; 9(6).
- Kang MH, Bae YS. IL-33 and IL-33-derived DC-based tumor immunotherapy. Exp Mol Med 2024; 56(6):1340-1347.
- Andreone S, Spadaro F, Buccione C, Mancini J, Tinari A, Sestili P, et al. IL-33 Promotes CD11b/CD18-Mediated Adhesion of Eosinophils to Cancer Cells and Synapse-Polarized Degranulation Leading to Tumor Cell Killing. Cancers (Basel) 2019; 11(11).
- Marichal T, Tsai M, Galli SJ. Mast cells: potential positive and negative roles in tumor biology. Cancer Immunol Res 2013; 1(5):269-279.
- Ribatti, D. Mast Cells and Resistance to Immunotherapy in Cancer. Arch Immunol Ther Exp (Warsz) 2023; 71(1):11.
- Lv Y, Tian W, Teng Y, Wang P, Zhao Y, Li Z, et al. Tumor-infiltrating mast cells stimulate ICOS. J Adv Res 2024; 57:149-162.
- Andreone S, Gambardella, A. R., Mancini, J., Loffredo, S., Marcella, S., La Sorsa, V., Varricchi, G., Schiavoni, G., & Mattei, F. Anti-Tumorigenic Activities of IL-33: A Mechanistic Insight. Frontiers in immunology 2020; 11, 571593.
- Afferni C, Buccione C, Andreone S, Galdiero MR, Varricchi G, Marone G, et al. The Pleiotropic Immunomodulatory Functions of IL-33 and Its Implications in Tumor Immunity. Front Immunol 2018; 9:2601.
- Schneider E, Petit-Bertron AF, Bricard R, Levasseur M, Ramadan A, Girard JP, et al. IL-33 activates unprimed murine basophils directly in vitro and induces their in vivo expansion indirectly by promoting hematopoietic growth factor production. J Immunol 2009; 183(6):3591-3597.
- Pecaric-Petkovic T, Didichenko SA, Kaempfer S, Spiegl N, Dahinden CA. Human basophils and eosinophils are the direct target leukocytes of the novel IL-1 family member IL-33. Blood 2009; 113(7):1526-1534.
- Rivellese F, Suurmond J, de Paulis A, Marone G, Huizinga TW, Toes RE. IgE and IL-33-mediated triggering of human basophils inhibits TLR4-induced monocyte activation. Eur J Immunol 2014; 44(10):3045-3055.
- Afferni C BC, Andreone S, Galdiero MR, Varricchi G, Marone G, Mattei F, Schiavoni G. The Pleiotropic Immunomodulatory Functions of IL-33 and Its Implications in Tumor Immunity. Frontiers in immunology 2018; 9(2601).
- Prete AD, Salvi, V., Soriani, A., Laffranchi, M., Sozio, F., Bosisio, D., & Sozzani, S. Dendritic cell subsets in cancer immunity and tumor antigen sensing. Cellular and Molecular Immunology 2023; 20(5):432-447.
- Xu H, Li, D., Ma, J., Zhao, Y., Xu, L., Tian, R., Liu, Y., Sun, L., & Su, J. The IL-33/ST2 axis affects tumor growth by regulating mitophagy in macrophages and reprogramming their polarization. Cancer Biology & Medicine 2021; 18(1):172-183.
- Chen J ZY, Jiang Y, et al. Interleukin-33 Contributes to the Induction of Th9 Cells and Antitumor Efficacy by Dectin-1-Activated Dendritic Cells. Front Immunol 2018.
- Zhang X, Chen, W., Zeng, P., Xu, J., & Diao, H. The Contradictory Role of Interleukin-33 in Immune Cells and Tumor Immunity. Cancer management and research 2020; 12:7527-7537.
- Liu X, Li L, Si F, Huang L, Zhao Y, Zhang C, et al. NK and NKT cells have distinct properties and functions in cancer. Oncogene 2021; 40(27):4521-4537.
- Choi MR, Sosman, J. A., & Zhang, B.. The Janus Face of IL-33 Signaling in Tumor Development and Immune Escape. Cancers 2021; 13(13):3281.
- Zhang X CW, Zeng P, Xu J, Diao H. The Contradictory Role of Interleukin-33 in Immune Cells and Tumor Immunity. 2020.
- Stojanovic B, Gajovic, N., Jurisevic, M., Stojanovic, M. D., Jovanovic, M., Jovanovic, I., Stojanovic, B. S., & Milosevic, B. Decoding the IL-33/ST2 Axis: Its Impact on the Immune Landscape of Breast Cancer. International Journal of Molecular Sciences 2023; 24(18).
- Xiao P, Wan, X., Cui, B., Liu, Y., Qiu, C., Rong, J., Zheng, M., Song, Y., Chen, L., He, J., Tan, Q., Wang, X., Shao, X., Liu, Y., Cao, X., & Wang, Q. Interleukin 33 in tumor microenvironment is crucial for the accumulation and function of myeloid-derived suppressor cells. Oncoimmunology 2016; 5(1).
- Komai-Koma M, Wang, E., Kurowska-Stolarska, M., Li, D., McSharry, C., & Xu, D. Interleukin-33 promoting Th1 lymphocyte differentiation dependents on IL-12. Immunobiology 2016; 221(3):412–417.
- Wu X, Tian J, Wang S. Insight Into Non-Pathogenic Th17 Cells in Autoimmune Diseases. Front Immunol 2018; 9:1112.
- Xing J, Man C, Liu Y, Zhang Z, Peng H. Factors impacting the benefits and pathogenicity of Th17 cells in the tumor microenvironment. Front Immunol 2023; 14:1224269.
- Pascual-Reguant A, Bayat Sarmadi J, Baumann C, Noster R, Cirera-Salinas D, Curato C, et al. T. Mucosal Immunol 2017; 10(6):1431-1442.
- Guo H BE, Ma X, Zhao C, Zhao Y. Dual Immune Regulatory Roles of Interleukin-33 in Pathological Conditions. Cells 2022.
- Lei S, Jin J, Zhao X, Zhou L, Qi G, Yang J. The role of IL-33/ST2 signaling in the tumor microenvironment and Treg immunotherapy. Exp Biol Med (Maywood) 2022; 247(20):1810-1818.
- Chen CC, Kobayashi, T., Iijima, K., Hsu, F. C., & Kita, H. IL-33 dysregulates regulatory T cells and impairs established immunologic tolerance in the lungs. The Journal of allergy and clinical immunology 2017; 140(5):1351–1363.e1357.
- Shani O, Vorobyov T, Monteran L, Lavie D, Cohen N, Raz Y, et al. Fibroblast-Derived IL33 Facilitates Breast Cancer Metastasis by Modifying the Immune Microenvironment and Driving Type 2 Immunity. Cancer Res 2020; 80(23):5317-5329.
- Monteran L, Erez N. The Dark Side of Fibroblasts: Cancer-Associated Fibroblasts as Mediators of Immunosuppression in the Tumor Microenvironment. Front Immunol 2019; 10:1835.
- Chen SF, Nieh S, Jao SW, Wu MZ, Liu CL, Chang YC, et al. The paracrine effect of cancer-associated fibroblast-induced interleukin-33 regulates the invasiveness of head and neck squamous cell carcinoma. J Pathol 2013; 231(2):180-189.
- Peng L, Sun W, Chen L, Wen WP. The Role of Interleukin-33 in Head and Neck Squamous Cell Carcinoma Is Determined by Its Cellular Sources in the Tumor Microenvironment. Front Oncol 2020; 10:588454.
- Andersson P, Yang Y, Hosaka K, Zhang Y, Fischer C, Braun H, et al. Molecular mechanisms of IL-33-mediated stromal interactions in cancer metastasis. JCI Insight 2018; 3(20).
- He Z, Chen L, Souto FO, Canasto-Chibuque C, Bongers G, Deshpande M, et al. Epithelial-derived IL-33 promotes intestinal tumorigenesis in Apc. Sci Rep 2017; 7(1):5520.
- Chen L, Sun R, Xu J, Zhai W, Zhang D, Yang M, et al. Tumor-Derived IL33 Promotes Tissue-Resident CD8. Cancer Immunol Res 2020; 8(11):1381-1392.
- Yang M, Feng Y, Yue C, Xu B, Chen L, Jiang J, et al. Lower expression level of IL-33 is associated with poor prognosis of pulmonary adenocarcinoma. PLoS One 2018; 13(3):e0193428.
- Liu J, Shen JX, Hu JL, Huang WH, Zhang GJ. Significance of interleukin-33 and its related cytokines in patients with breast cancers. Front Immunol 2014; 5:141.
- Kim JY, Kim G, Lim SC, Choi HS. IL-33-Induced Transcriptional Activation of LPIN1 Accelerates Breast Tumorigenesis. Cancers (Basel) 2021; 13(9).
- Kirichenko TV, Markina, Y. V., Bogatyreva, A. I., Tolstik, T. V., Varaeva, Y. R., & Starodubova, A. V. The Role of Adipokines in Inflammatory Mechanisms of Obesity. International journal of molecular sciences 2022; 23(23).
- Schmidt FM, Weschenfelder, J., Sander, C., Minkwitz, J., Thormann, J., Chittka, T., Mergl, R., Kirkby, K. C., Faßhauer, M., Stumvoll, M., Holdt, L. M., Teupser, D., Hegerl, U., & Himmerich, H.. Inflammatory Cytokines in General and Central Obesity and Modulating Effects of Physical Activity. PLoS ONE 2015; 10(3).
- Kim JW, Kim, J. H., & Lee, Y. J. The Role of Adipokines in Tumor Progression and Its Association with Obesity. Biomedicines 2024; 12(1):97.
- Molofsky AB, Van Gool F, Liang HE, Van Dyken SJ, Nussbaum JC, Lee J, et al. Interleukin-33 and Interferon-γ Counter-Regulate Group 2 Innate Lymphoid Cell Activation during Immune Perturbation. Immunity 2015; 43(1):161-174.
- Odegaard JI, Lee MW, Sogawa Y, Bertholet AM, Locksley RM, Weinberg DE, et al. Perinatal Licensing of Thermogenesis by IL-33 and ST2. Cell 2016; 166(4):841-854.
- Li Q, Li D, Zhang X, Wan Q, Zhang W, Zheng M, et al. E3 Ligase VHL Promotes Group 2 Innate Lymphoid Cell Maturation and Function via Glycolysis Inhibition and Induction of Interleukin-33 Receptor. Immunity 2018; 48(2):258-270.e255.
- Brestoff JR, Kim BS, Saenz SA, Stine RR, Monticelli LA, Sonnenberg GF, et al. Group 2 innate lymphoid cells promote beiging of white adipose tissue and limit obesity. Nature 2015; 519(7542):242-246.
- Lee MW, Odegaard JI, Mukundan L, Qiu Y, Molofsky AB, Nussbaum JC, et al. Activated type 2 innate lymphoid cells regulate beige fat biogenesis. Cell 2015; 160(1-2):74-87.
- Miller AM, Asquith DL, Hueber AJ, Anderson LA, Holmes WM, McKenzie AN, et al. Interleukin-33 induces protective effects in adipose tissue inflammation during obesity in mice. Circ Res 2010; 107(5):650-658.
- Wang L, Luo Y, Luo L, Wu D, Ding X, Zheng H, et al. Adiponectin restrains ILC2 activation by AMPK-mediated feedback inhibition of IL-33 signaling. J Exp Med 2021; 218(2).
- Brestoff JR, Kim, B. S., Saenz, S. A., Stine, R. R., Monticelli, L. A., Sonnenberg, G. F., Thome, J. J., Farber, D. L., Lutfy, K., Seale, P., & Artis, D. Group 2 innate lymphoid cells promote beiging of white adipose tissue and limit obesity. Nature 2015; 519(7542):242-246.
- de Oliveira MFA, Talvani, A., & Rocha-Vieira, E. IL-33 in obesity: where do we go from here? Inflammation research : official journal of the European Histamine Research Society [et al] 2019; 68(3):185-194.
- Sanada S HD, Higgins LJ, Schreiter ER, McKenzie AN, Lee RT. IL-33 and ST2 comprise a critical biomechanically induced and cardioprotective signaling system. Clin Investig 2007:1538-1549.
- Miller AM XD, Asquith DL, Denby L, Li Y, Sattar N, Baker AH, McInnes IB, Liew FY. IL-33 reduces the development of atherosclerosis. J Exp Med 2008:339-346.
Figure 1.
IL-33 and ST2 distribution. Various immune cells release IL-33 in response to cell stress and injury. It is also released from the endothelial and epithelial cells of barrier tissues, glial cells and astrocytes, smooth muscle cells, and platelets. IL-33 signals through ST2, expressed in different types of immune cells, including innate lymphoid cells and adaptive immune cells, enhance their function. The population of immune cells that IL-33/ST2 targets in adipose tissue changes with obesity, leading to obesity-related inflammation. The lean adipose tissue predominantly contains non-inflammatory cells, such as activated M2 macrophages, eosinophils, ILC2 cells, Tregs, and Th2 cells. In obesity, the immune profile of the adipose tissue shifts to a pro-inflammatory state through recruiting macrophages (M1), NK cells, neutrophils, CD4+ T cells (Th1, Th17), and CD8+ T cells. DC, dendritic cell; M, macrophages; ILC2, innate lymphoid cell type 2; iNKT, invariant natural killer T; NK, natural killer cells; NKT, natural killer T cells. MDSC, myeloid-derived suppressor cells. This figure was created with BioRender.
Figure 1.
IL-33 and ST2 distribution. Various immune cells release IL-33 in response to cell stress and injury. It is also released from the endothelial and epithelial cells of barrier tissues, glial cells and astrocytes, smooth muscle cells, and platelets. IL-33 signals through ST2, expressed in different types of immune cells, including innate lymphoid cells and adaptive immune cells, enhance their function. The population of immune cells that IL-33/ST2 targets in adipose tissue changes with obesity, leading to obesity-related inflammation. The lean adipose tissue predominantly contains non-inflammatory cells, such as activated M2 macrophages, eosinophils, ILC2 cells, Tregs, and Th2 cells. In obesity, the immune profile of the adipose tissue shifts to a pro-inflammatory state through recruiting macrophages (M1), NK cells, neutrophils, CD4+ T cells (Th1, Th17), and CD8+ T cells. DC, dendritic cell; M, macrophages; ILC2, innate lymphoid cell type 2; iNKT, invariant natural killer T; NK, natural killer cells; NKT, natural killer T cells. MDSC, myeloid-derived suppressor cells. This figure was created with BioRender.
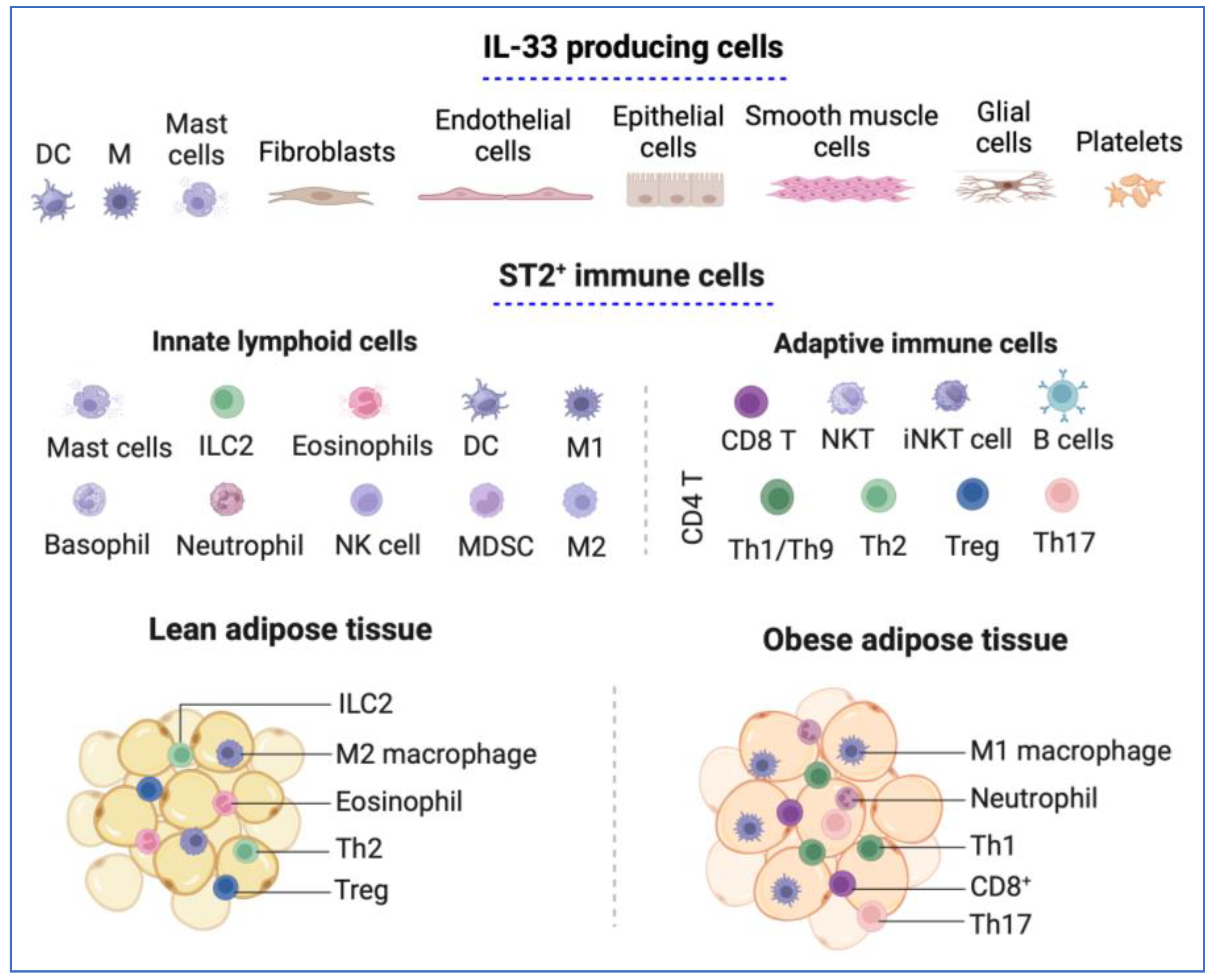
Figure 2.
IL-33/ST2 signaling pathway. IL-33 binds to the ST2 receptor, ST2L, and IL-1RAcP complex and then MyD88, similar to how other interleukin 1 subfamily members, such as Il-1α and IL-1β, bind. TIR initiates the pathway by first recruiting MyD88, which induces recruitment of IRAK1 and IRAK4, which further recruits TRAF6 to activate MAPK and NF-κB signaling pathways. The activity of MAPK pathways and canonical NF-κB is regulated at multiple levels. IL-33 can also bind to a decoy receptor formed by a soluble form of ST2, sST2, or SIGIRR for inactivation. ST2L, suppression of tumorigenicity 2 ligand; IL-1RAcP, IL-1 receptor accessory protein; MyD88, myeloid differentiation primary response 88; IRAK1, interleukin receptor-associated kinase 1; IRAK4, interleukin receptor-associated kinase 4; TRAF6, tumor necrosis factor receptor-associated factor 6; MAPK, mitogen-activated protein kinase; ERK1/2, extracellular signal-regulated kinases 1/2; JNK, c-Jun N-terminal kinase; p38, the subgroup of MAP kinases; AP-1, transcription factor; NF-κB, Nuclear Factor κB; TAK1, TGF-β-activated protein kinase 1; NEMO, NF-κB essential modulator; IKKβ, inhibitor of κB (IκB) kinase β. This figure was created with BioRender.
Figure 2.
IL-33/ST2 signaling pathway. IL-33 binds to the ST2 receptor, ST2L, and IL-1RAcP complex and then MyD88, similar to how other interleukin 1 subfamily members, such as Il-1α and IL-1β, bind. TIR initiates the pathway by first recruiting MyD88, which induces recruitment of IRAK1 and IRAK4, which further recruits TRAF6 to activate MAPK and NF-κB signaling pathways. The activity of MAPK pathways and canonical NF-κB is regulated at multiple levels. IL-33 can also bind to a decoy receptor formed by a soluble form of ST2, sST2, or SIGIRR for inactivation. ST2L, suppression of tumorigenicity 2 ligand; IL-1RAcP, IL-1 receptor accessory protein; MyD88, myeloid differentiation primary response 88; IRAK1, interleukin receptor-associated kinase 1; IRAK4, interleukin receptor-associated kinase 4; TRAF6, tumor necrosis factor receptor-associated factor 6; MAPK, mitogen-activated protein kinase; ERK1/2, extracellular signal-regulated kinases 1/2; JNK, c-Jun N-terminal kinase; p38, the subgroup of MAP kinases; AP-1, transcription factor; NF-κB, Nuclear Factor κB; TAK1, TGF-β-activated protein kinase 1; NEMO, NF-κB essential modulator; IKKβ, inhibitor of κB (IκB) kinase β. This figure was created with BioRender.
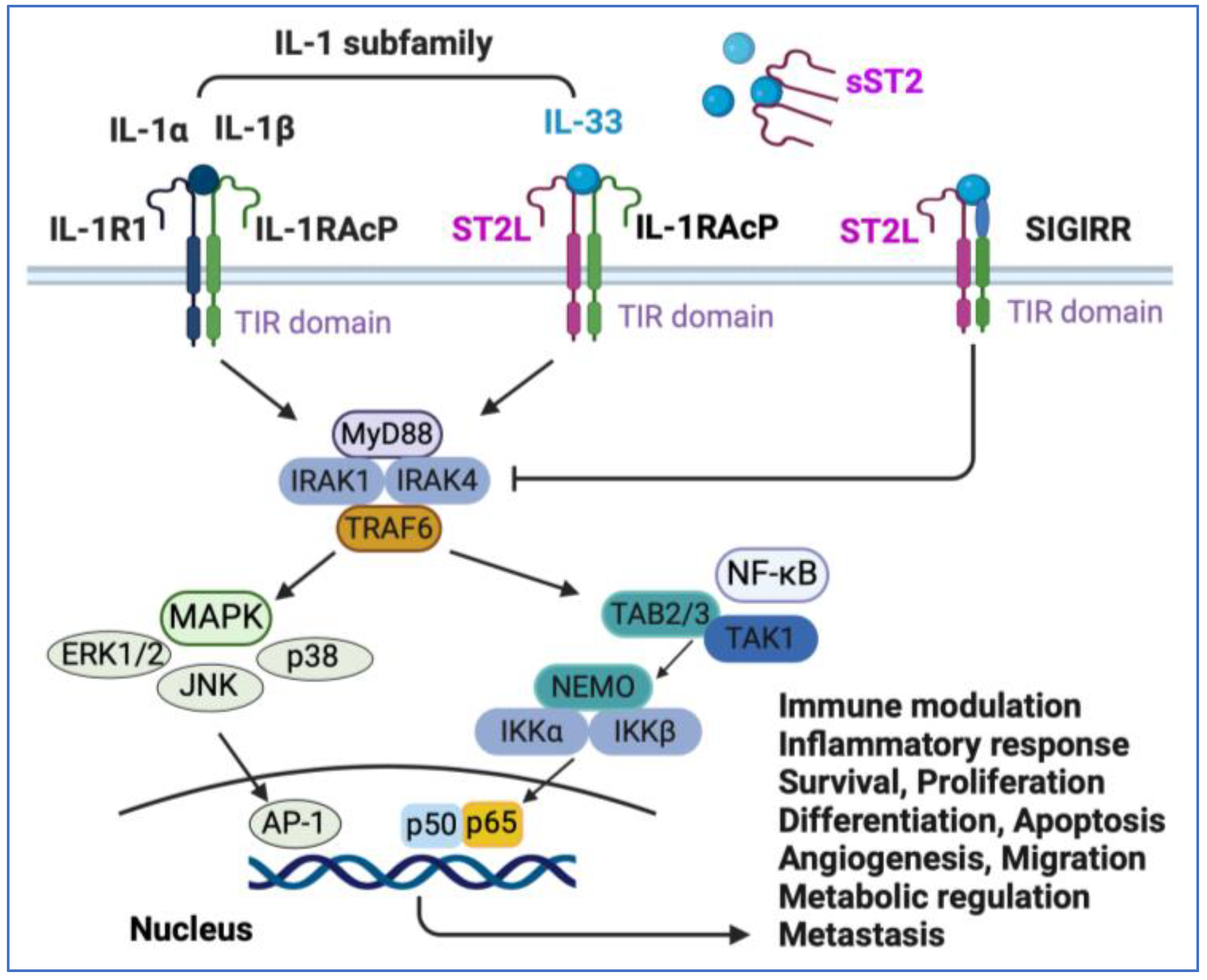
Figure 3.
Role of IL-33 in the tumor microenvironment: IL-33 influences immune regulation and can have both pro-tumor and antitumor effects. The population of immune cells targeted by IL-33/ST2 in the TME changes depending on the context. This results in opposing effects in different tumors. Along with tumor development, IL-33 is likely downregulated in epithelial cells but upregulated in the tumor microenvironment. The increased IL-33 expression in stroma either maintains or activates suppressor immune cells such as macrophages, Tregs, and CD4+ Th2 or Th17 cells, thus contributing to tumor growth and metastasis. However, in contrast, IL-33 may also have an antitumor effect by activating innate natural kill (NK) cells and adaptive (CD4+ Th1 or CD8+ T cells) immune responses. IL-33/ST2 signaling can lead to a dual role on other cell types such as eosinophils, basophils, group 2 innate lymphoid cells (ILC2s), and myeloid-derived suppressor cells (MDSCs) either directly or through interaction with other cell types, depending on cancer type [83]. Cell names in red indicate their primary function is a pro-tumor effect, blue indicates their primary function is an antitumor effect, and green indicates dual roles. This figure was created with BioRender.
Figure 3.
Role of IL-33 in the tumor microenvironment: IL-33 influences immune regulation and can have both pro-tumor and antitumor effects. The population of immune cells targeted by IL-33/ST2 in the TME changes depending on the context. This results in opposing effects in different tumors. Along with tumor development, IL-33 is likely downregulated in epithelial cells but upregulated in the tumor microenvironment. The increased IL-33 expression in stroma either maintains or activates suppressor immune cells such as macrophages, Tregs, and CD4+ Th2 or Th17 cells, thus contributing to tumor growth and metastasis. However, in contrast, IL-33 may also have an antitumor effect by activating innate natural kill (NK) cells and adaptive (CD4+ Th1 or CD8+ T cells) immune responses. IL-33/ST2 signaling can lead to a dual role on other cell types such as eosinophils, basophils, group 2 innate lymphoid cells (ILC2s), and myeloid-derived suppressor cells (MDSCs) either directly or through interaction with other cell types, depending on cancer type [83]. Cell names in red indicate their primary function is a pro-tumor effect, blue indicates their primary function is an antitumor effect, and green indicates dual roles. This figure was created with BioRender.
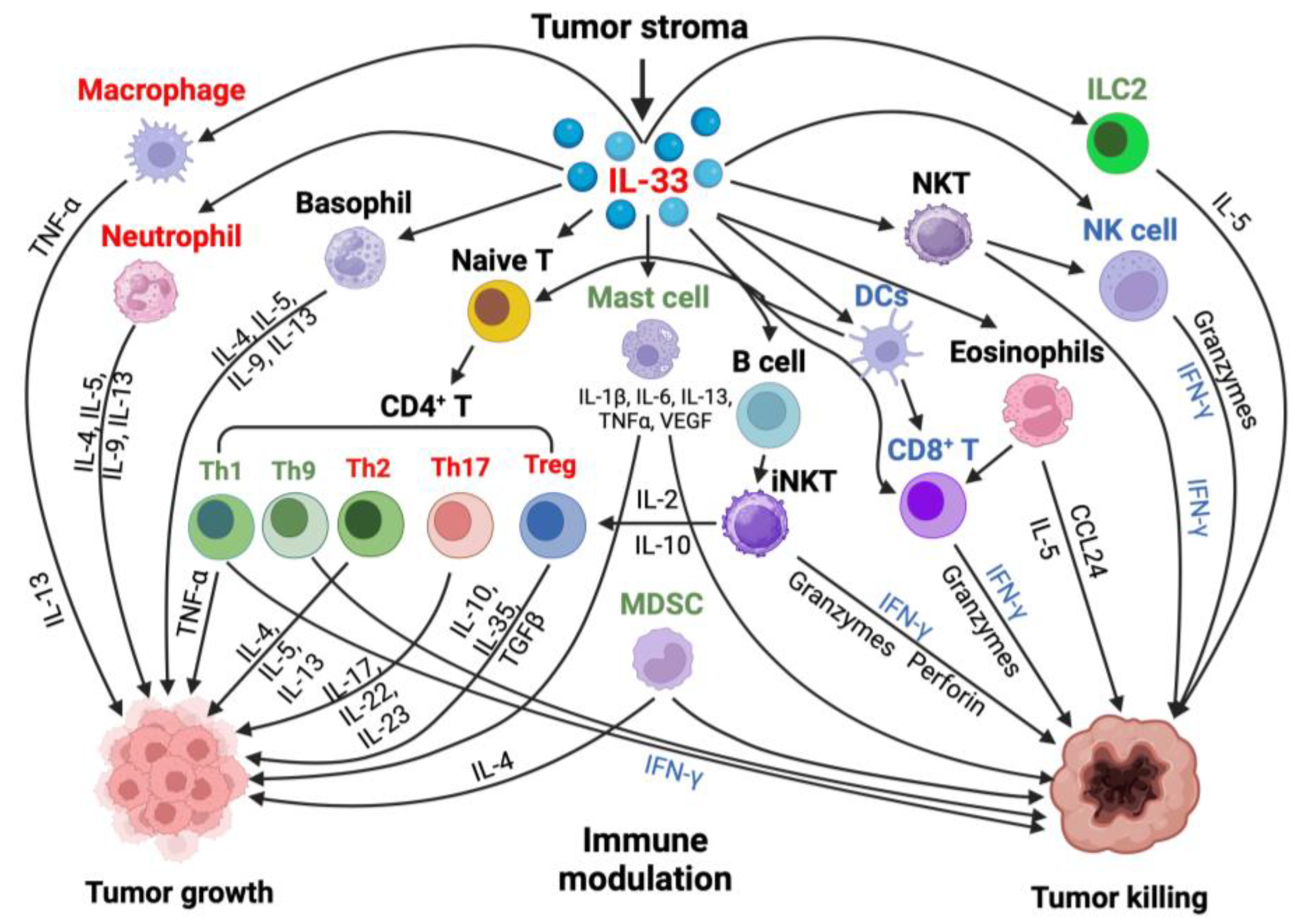
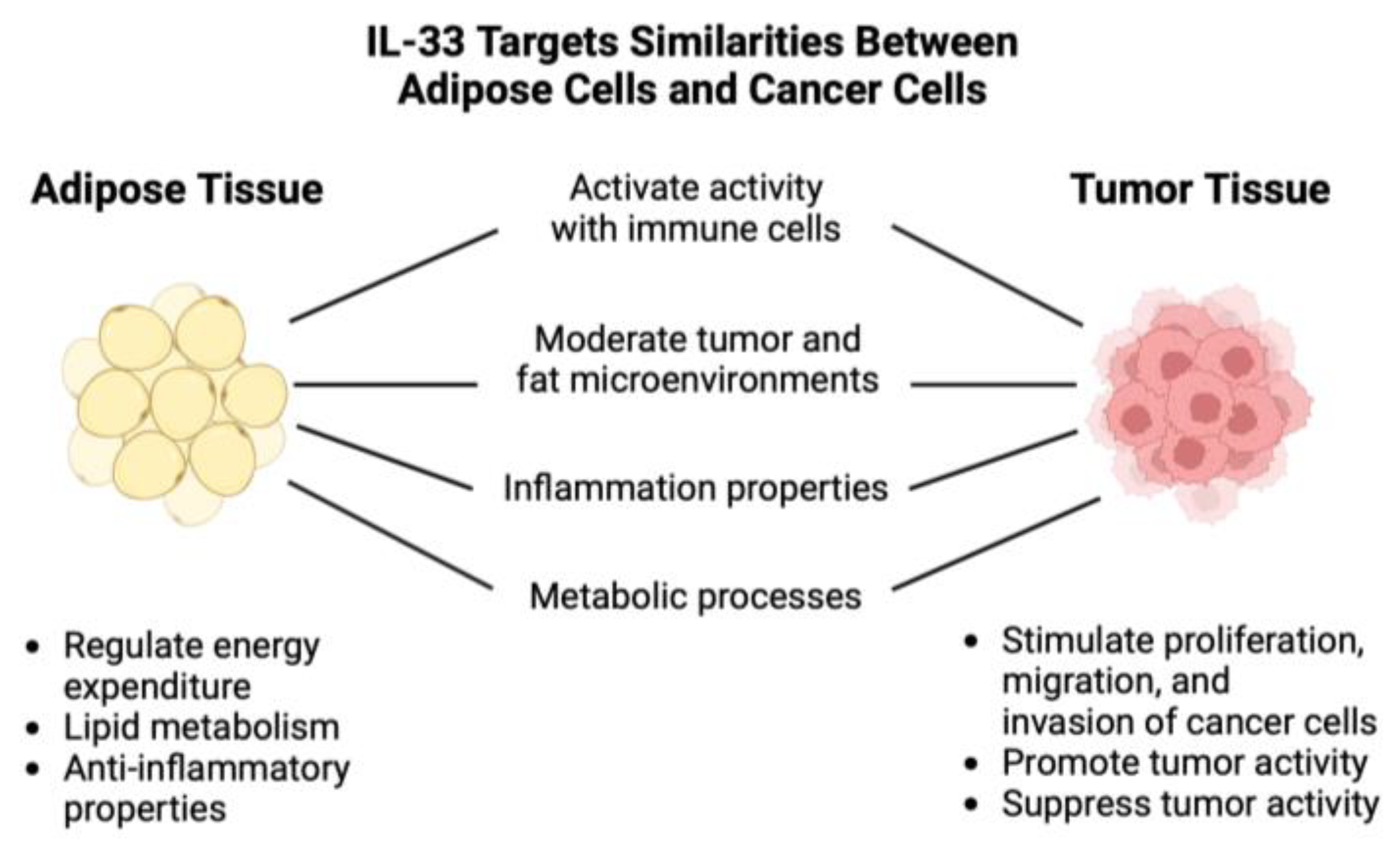
Figure 4.
Versatile Roles of IL-33 in Adipose Tissue and Tumor Microenvironment [12]. This diagram illustrates the versatile roles of IL-33 in both adipose tissue and cancer cells. It plays a crucial role in activating immune responses, moderating the microenvironments of tumors and fat, regulating inflammatory properties, and assisting in regulating metabolic processes in these contexts. The diagram highlights the stark contrast in IL-33's effects in these two settings, from stimulating cancer cells to regulating energy expenditure in adipose tissue.
Figure 4.
Versatile Roles of IL-33 in Adipose Tissue and Tumor Microenvironment [12]. This diagram illustrates the versatile roles of IL-33 in both adipose tissue and cancer cells. It plays a crucial role in activating immune responses, moderating the microenvironments of tumors and fat, regulating inflammatory properties, and assisting in regulating metabolic processes in these contexts. The diagram highlights the stark contrast in IL-33's effects in these two settings, from stimulating cancer cells to regulating energy expenditure in adipose tissue.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



