Submitted:
03 September 2024
Posted:
05 September 2024
You are already at the latest version
Abstract
Genetically diverse influenza A viruses (IAV) circulates in wild aquatic birds with relatively frequent spillover (cases of single infections) to other mammals, but only a small proportion of outbreaks. The spillover of IAV to marine mammals such as seals have been reported throughout the last 45 years. Majority of these viruses when reached seals population cased pneumonia and death of infected animals. Until 2016, all avian influenza viruses found in seals were of low pathotype (LP) IAV. The H5Nx clade 2.4.4.4b highly pathogenic avian influenza viruses (HPAIV) were found in seals for the first time in 2016/2017. Our review highlights 45 years of AIV history in seals population, showing interesting features of different AIV subtypes and emphasis on avian-to-mammals transmission. Furthermore, we also included phylogenetic characterisation of the representative seal isolates.
Keywords:
avian influenza virus
; marine mammals
; seals
; LPAIV
; HPAIV
; H5N1
Brief Introduction
Avian influenza viruses (AIVs) are classified as either low- pathogenicity (LP) or high-pathogenicity (HP). LPAIVs generally cause mild infections, whilst HPAIVs can cause high mortality in a wide range of avian species. AIV subtypes are defined based on their surface glycoproteins, haemagglutinin (HA; H1-H16) and neuraminidase (NA; N1-N9), and HPAIVs appear restricted to the H5 and H7 subtypes. These viruses sporadically were found in other animals, including marine mammals. Until 2016, none HPAIV was reported in seal population. Currently circulating strains of H5 HPAI viruses emerged in 1996 in Southern China as A/goose/Guangdong (GsGD) and then evolved rapidly, expanding the geographical locations and animal species infected. The cross-species transmission of current H5N1 HPAIV clade 2.3.4.4b was reported in over 43 mammalian species across Europe, North America, South America, and Asia (EFSA, April-June 2023). Seals were one of the massively impacted marine mammal species by the current H5N1 influenza outbreak. In this review we look at 45 years of AIV history in seals population. We also performed phylogenetic analysis of seal isolates obtained in the last 45 years and simple PB2 protein analysis of 72 isolates (seals and other marine mammals) available in GSAID with emphasis on two mutations (D701N and E627K) being know of enhancing replication of avian influenza viruses in mammalian cells (Briggs and Kapczynski 2023).
Seals along with walruses and sea lions are classified as pinnipeds (suborder Pinnipedia). There are 32 species of seals that belongs to 17 genera and 2 families (Otariidae and Phocidae). The harbour seals (Phoca vitulina) which were the most frequently infected seals species by avian influenza virus (AIV) belong to Genus Phoca (common seals) which represents Northern Hemisphere seals. They are nonmigratory, earless seal that can be found along coastlines and in a few freshwater lakes in Canada and Alaska (NOAA, 2024). The grey seals (Halichoerus grypus) are only found in the North Atlantic and there are three populations of grey seals that are recognized: the Northeast Atlantic, the Northwest Atlantic and Baltic Sea (Haug et al., 2007). A large colony of grey seals can be found in the UK (Stewart et al., 2014) with an estimate number of more than 120,000 animals, representing 40% of the world's population and 95% of the European population of grey seals (Nature Scotland, 2024; Figure 1). In U.S. waters, the number of pupping sites has increased from one in 1988 to nine in 2019 and are located in Maine and Massachusetts (Wood et al. 2019). From 1988–2019, the number of pups born increased however these numbers do not reflect changes in total population growth resulting from Canadian seals migrating to the USA regions (Wood et al. 2019). The other interesting seals representing Northern Hemisphere are Northern Elephant Seal (Mirounga angustirostris) with the largest colonies off southern California in the Channel Islands. They have one of the longest migrations of any mammal; some have been recorded traveling over 13,000 miles roundtrip. Those seals nearly extinct in 1892 (Townsend, 1912) but refilled its range within 70–80 years (Lowry et al., 2014; Steward et al., 1994). Those animals were found positive for H1N1 pandemic influenza virus twice (in 2011 and 2019) with no clinical sign of disease. The Southern Elephant Seal (M. leonina), in contrary, can be found throughout sub-Antarctic regions and they have greater body mass than their northern counterpart. They only occasionally can be found in Antarctica to mate but the breeding locations are distributed in subantarctic locations. The largest population is in the South Atlantic on South Georgia (Boyd et al., 1996) other breeding locations include Falkland Islands and Valdes Peninsula in Argentina. Two other big populations of southern elephant seals can be found is in the south Indian Ocean, mostly in the Kerguelen Islands and in the subantarctic islands of the Pacific Ocean south of Tasmania and New Zealand, mainly Macquarie Island. Recent 2.3.4.4b H5N1 virus cause massive mortality among southern elephant young pups that were born in 2023 across Argentina where 96% of total population was dead after the AIV viral infection (NEWS, Jan 2024).
The first Reported Cases of Avian Influenza Viruses (AIV) in Seals Were of Low Pathogenic Subtype (LPAIV)
LPAIV: H7N7 (1979), H4N5 (1982), H3N3 (1992), H4N6 (1992, 2002, 2012)
The first cases of avian influenza virus that manifested as a clinical disease (pneumonia with fatal outcome) in seals were of law pathogenic avian influenza virus pathotype (LPAIV). In the winter of 1979-1980, approximately 600 harbour seals (Phoca vitulina) were suffering from pneumonia that ended in a fatal outcome on Cape Cod Peninsula in USA (Lang et al., 1981, Webster et al. 1981; Geraci et al., 1982). The influenza A virus H7N7 (previously called serotype Hav1 Neq1) was confirmed in lung and brain tissues (Lang et al., 1981). Interestingly, this virus was harmless to chickens and turkeys in transmission experiments but previous human infection by H7N7 (Hav1 Neq1) was reported (Lang et al., 1981). Similarly, Webster et al. (1981) have shown that the seal virus replicates poorly, produce no disease signs, and was not shed in the feces of avian species, yet was able to replicate in ferrets, cats, and pigs with no clinical disease. No antibodies to this virus were detected in harbour seals other than New England coast, grey seals, or fur seals suggesting that this virus may be a new introduction to this species (Webster et al., 1981). The A/Seal/Mass/1/80 influenza virus provides the first evidence suggesting that a strain deriving all of its genes from one or more avian influenza viruses can be associated with severe disease in a mammalian population in nature (Webster et al., 1981). Following the first cases of avian influenza viruses in harbour seals, Hinshaw et al. (1981) designed the study to see whether there is a possibility of avian viruses to replicate in three mammals-pigs, ferrets, and cats and the avian strains replicated to high titers in the respiratory tract (10(5) to 10(7) 50% egg infective doses per ml of nasal wash), with peak titers at 2 to 4 days post-inoculation, similar to the pattern of human and other mammalian viruses in these animals. The infected mammals had no significant disease signs and produced low levels of humoral antibodies; however, challenge experiments in ferrets indicated that they were immune (Hinshaw et al, 1981). This virus was also able to replicate in squirrel monkeys (Murphy et al., 1983). Later, Li et al. (1990) have shown that this H7N7 virus can adopt to the CEC by introducing mutation close to the HA cleavage which resulted in cleavability by ubiquitous proteases and enhance pathogenicity for chickens whereas adaptation to MDCK cells leads to mutation that was distant from the cleavage side which increase cleavability, but the virus was still apathogenic for the chicken (Li et al, 1990). The second time when the harbour seals were reported dying of pneumonia on the New England coast was from June 1982 through March 1983 (Hinshaw et al., 1984). After performing serological and RNA-RNA assays it was shown that these viruses are closely antigenically and genetically related to circulating avian virus strains but were distinct from mammalian viruses, including H7N7 isolates recovered from seals in 1980 and were later classified as H4N5 subtype (A/seals/Mass/133/82) (Hinshaw et al., 1984). Interestingly, these seal viruses were able to replicate in the intestinal tracts of ducks as compared to the previous isolates (H7N7) that replicated very poorly or not at all (Hinshaw et al., 1984). The H4N5 seal virus was also used to experimentally infect harbour seals (N=1), ringed seals (N=2) and harp seals (N=3). Although the clinical sign of the disease was mild, the virus was recovered from nasal (5/6) and anal swab samples (2/6). The seals also produced antibodies to the challenge virus (3/3) at two weeks post-challenge that were still present at 30 days post-challenge starting to slowly decline (Hinshaw et al., 1984). This virus replicated to high titter of the virus was seen in lungs (10^5.5 EID50) as compared to 10^3.5 in brain and lymph nodes tissues. No previous evidence of contact with this virus was seen in seals population before 1982 (Hinshaw et al., 1984). We performed phylogenetic analysis of first seal isolate A/seals/Mass/133/82 H4N5 and compare it with other H4N5 viruses available in the database (N=22), a 99.7%-99.89% similarity in HA was shown, and the closest related strains were of wild birds isolates isolated from USA, Canada and Peru between 1990 and 2018 with exception to the strain A/duck/Alberta/2016 (99.27% similarity) (Figure 2). Following these two epizootics of influenza virus in seals in 1980s, the prevalence of influenza virus on Ross Island, Antarctic region was examined, using samples collected in November 1979 from Weddell seals, Adelie penguins (N=100) and Antarctic skuas (N=60) in addition to serum samples collected from October 1985 to January 1986 (Austin and Webster 1992). No influenza virus was isolated, and none of the seals sera were positive for AIV. However, the antibodies against AIV were detected (most probably of H10 serotype) in penguins and skuas at the Ross Sea Dependency, Antarctic region (Austin and Webster 1992).
In January 1991 and January to February 1992, harbour seals were found dead along the Cape Cod peninsula of Massachusetts, two avian influenza viruses were identified at that time– H4N6 and H3N3, out of which, H3N3 was isolated for a first time from the seals (Callan et al., 1995). The antigenic reactivity of H3 viruses isolated from seals showed that three seal viruses (A/Seal/MA/3911/92 and A/Seal/MA/4007/92) had patterns similar to A/Duck/Ukraine/1/63 and one virus (A/Seal/MA/3984/92) had a pattern more similar to the human virus, A/Aichi/2/68 whereas genetically the HAs of these new viruses were closely related to the HAs of viruses recovered from North American birds (Callan et al., 1995). Callan et al., (1995) have shown that residues 226 and 228 in HA of the seal isolates are glutamine and glycine, respectively, which coincide with the consensus for the receptor binding site of most avian and equine H3 viruses. The H7N7 (A/Seal/MA/1/80) and the H4N5 (A/Seal/MA/133/82) viruses previously isolated from seals also have the same avian receptor binding sequence, G225-Q-S-G-R229 (Naeve and Webster, 1983; Donis et al., 1989). This suggests that the binding sequence may be well adapted for seals since mutations away from this sequence have not been observed in viruses isolated from seals (Callan et al., 1995). Here we performed genetic analysis of HA and NA segments of H3N3 avian viruses available in the database (N=47) along with A/seal/Mas/3911/1992 (Suppl. Figure 1 and Figure 2). The HA of seal H3N3 was 99.87 % similar to A/mallard duck/AL/1988 and A/ruddy turnstone/Dalaware bay/1994 and almost identical (99.9% similarity) to wild bird isolates isolated from USA, Canada and Guatemala between 1994 and 2015 and 2001 swine isolate from Canada. The NA segment clustered with A/mallard duck/AL/1988 and A/ruddy turnstone/Dalaware bay/1994 and like HA segment, also NA of seal isolates clustered together with other wild bird isolates isolated later from USA, Canada and Guatemala, and swine isolate from Canada (2001). Matrosovich et al. (2000) have used four H3N3 viruses isolated from the seals during the outbreak and assessed their affinity for sialylglycopolymers, 3′SL-PAA (avian receptor) and 6′SLN-PAA (human receptor), three out of four isolates had typical avian virus-like pattern of binding to sialylglycopolymers which was of about 50 μM−1 and 1 μM, respectively. One of the isolates, had a lower binding affinity for 3′SL-PAA with no increase in binding to 6′SLN-PAA, and carried two amino acids substitutions A138S and R220S (Matrosovich et al. 2000). This finding suggests that an avian influenza virus can infect seals without substantial changes in its receptor-binding specificity Matrosovich et al. (2000). An interesting study was done by Ramis et al., (2012) where the authors have assessed the patterns of attachment for different influenza virus strains (H4N5, H7N7 and human H1N1, H3N2 and Human B influenza) of harbour seals, grey seals (Halichoerus grypus), harbour porpoises (Phocoena phocoena), and bottlenose dolphins (Tursiops truncatus). It was shown that attachment of avian influenza A viruses to tracheal and bronchial epithelium was moderate in seals (harbour seal and gray seal) and absent in cetaceans (harbor porpoise and bottlenose dolphin) which suggested that seals are high susceptibility to these viruses and efficient transmission of these viruses can occur. However, scarce attachment of these viruses to bronchioles and alveoli of harbour seals is consistent with low pathogenicity of these viruses for harbour seals during experimental infection (Ramis et al., 2012). Lack of attachment of avian influenza A viruses tested to trachea and bronchi of harbour porpoises and bottlenose dolphins suggests low susceptibility and inefficient transmission in these species (Ramis et al., 2012). The H4N6 virus was also isolated from Caspian seal at Zhemchuzhny island (Astrakhan Region, Russia) in 2012, the seal did not show any clinical sign of disease (Gulyaeva, 2018). The genetic analysis of the virus showed that Caspian seal isolate is closely related to avian-derived influenza viruses of the classical Eurasian lineage circulating in wild birds and no mammalian adaptational mutations were found in PB2 protein (Gulyaeva, 2018). The virus could replicate in lungs of mice and cause severe disease in the experimental mice without prior adaptation (Gulyaeva, 2018). Serological evidence for infection of Caspian seals with influenza A and B viruses was shown using sera collected in 1993―2000 that reacted strongly with A/Bangkok/1/79 (H3N2) strain, suggesting that the seals were infected with human influenza A virus that circulated among humans in 1979―1981 but did not react at all with the A/Philippines/2/82 strain, which was prevalent in humans in 1982―1983 (Ohishi et al., 2002). The youngest seal with positive sera was 14.5 years old in 2000, implying that A/Bangkok/1/79-like viruses were maintained in the Caspian seal population at least until 1985 (Ohishi et al., 2002). None of 27 Caspian seals sera samples tested against H1, H3, H5 and H7 were positive in 2012 (Gulyaeva, 2018).
LPAIV: H3N8 (2011, 2017-2019)
In September 2011, a new outbreak of avian influenza virus was reported along the New England coast in the USA where harbor seals (Phoca vitulina) were found dead or moribund. This virus belonged to H3N8 subtype and was never isolated from seals prior to this event; however it was detected in harp seal in Northwest Atlantic once (Bogomolni et al., 2008; Anthony et al., 2012). “Unusual mortality event” was also declared in Maine, New Hampshire, and northern Massachusetts in November 2011 (Yang et al., 2015). Genetic characterisation of this viruses showed similarity to a waterfowl isolate circulating in North America since at least 2002 (Anthony et al., 2012). Interestingly, this new seal H3N8 viral isolate enquired an adaptational mutation D701N in the viral PB2 protein, however, maintained the avian phenotype at positions 226 (Q) and 228 (G) in HA protein, which correlates with a continued ability to use SAα-2,3 avian -type receptor (Anthony et al., 2012). Later, the binding properties and preferences of seal H3N8 virus to α2-3-linked and α2-6-linked sialosides were investigated in more details (Karlsson et al. 2014; Yang et al. 2015; Hussein et al., 2016). Karlsson et al. (2014) have demonstrated that seal H3N8 is unique amongst the avian H3N8 viruses tested by having enhanced α2,6 receptor binding, increased morbidity in mice and efficient respiratory droplet transmission in ferrets which was not seen in the other avian viruses within this clade (those only transmitted via contact). Later, Yang et al. (2015) have performed a detailed structural and biochemical analysis of the surface antigens of the A/harbor seal/New Hampshire/179629/2011 virus and shown that both the hemagglutinin and neuraminidase indicate a true avian receptor binding preference with a strong binding preference for the α2-3-linked sialosides, as well as mixed α2-3/α2-6 branched sialosides (glycans 65 to 66) but only little binding to human α2-6-linked sialosides. Hussein et al., (2016) have evaluated potential human transmissibility of seal H3N8. The binding of recombinant HA proteins of seal H3N8 and human-adapted H3N2 viruses to respiratory tissues of humans and ferrets were tested; showing that there was a strong tendency of the seal H3 to bind to lung alveoli in human tissue which was opposite to human-adapted H3 that bound mainly to the trachea and similar pattern was also seen in ferrets (Hussein et al., 2016). Furthermore, the binding of the recombinant HAs to a library of 610 glycans showed that the seal H3 bound preferentially to α-2,3 sialylated glycans which is in contrast to the human H3, which bound almost exclusively to α-2,6 sialylated glycans suggesting that seal H3N8 virus has retained its avian-like receptor binding specificity but could potentially establish infection in human lungs (Hussein et al., 2016).
The H3N8 virus was next found in grey seal pup in Cornwall, UK in 2017. The pup underwent medical treatment but died after two weeks (Venkatesh et al., 2020). Based on BLAST search and phylogenetic analysis this virus most probably originated from an unsampled, locally circulating (in Northern Europe) viruses, likely from wild Anseriformes. Furthermore, several mutations were detected, including D701N in the PB2 segment which is a rare mutation in this virus subtype and suggest on mammalian adaptation of bird viruses (Venkatesh et al., 2020).
Interestingly, although H3 influenza A virus is the most ubiquitous hemagglutinin subtype, and as we also showed here has a potential to replicate in human cells, no human infection was found until very recently where the person infected with H3N8 developed severe pneumonia in April 2022 followed by a second case in May 2022 in China (Bao et al., 2022, Tan et al., 2022). This human H3N8 strain was triple reassortant of avian influenza A-H3N8 virus (A/Henan/ZMD-22-2/2022(H3N8) with avian-origin HA and NA genes and six internal genes acquired from Eurasian lineage H9N2 viruses and it possessed E627K mutation in PB2 gene (Bao et al., 2022). Wang et al. (2023) have shown that H3N8 viruses circulating in wild migratory birds in China have evolved into different branches and have undergone complicated reassortment with viruses in waterfowl, they preferentially bind to avian-type receptor but they have acquired the ability to bind human-type receptors.
LPAIV: H10N7 (2014-2015 and 2021)
In 2014, the mass deaths among harbour seals (Phoca vitulina) occurred in Sweden (March 2014), Denmark (July 2014), and Germany (October 2014) where the seals were found dead being washed out to the shores (Zohari et al., 2014; Bodewes et al., 2015; Krog et al., 2015). Those were a first reported cases of dead seals outside the USA coast. From March through October 2014, 425 carcasses were detected in several seal colonies in the Kattegat and the Skagerrak seas in Sweden (Zohari et al., 2014). The H10N7 subtype was detected and based on HA characterisation the virus clustered with seal isolate from Germany (2014) and Swedish mallard H10 isolate from 2011 (Zohari et al., 2014). In Denmark, 152 harbour seals on the island of Anholt were found dead from severe pneumonia between June–August 2014 (Krog et al., 2015). In general, both the HA and NA segments showed high-level nucleotide sequence identity to AIVs from birds sampled in Scandinavia and the Republic of Georgia (Krog et al., 2015). In Germany, dead seal accounted for 12 % of the whole population of seals which at the time was approximately 12,000 animals in total (Bodewes et al., 2015). Influenza A virus (A/harbor seal/Germany/1/2014) of H10N7 subtype was isolated from lung and throat swab samples and replicated to the high level on 11-day-old embryonated chicken eggs and on MDCK cells. Genetic characterisation of the virus showed that seal H10N7 is closely related to the H10N7 viruses found in migratory ducks in Georgia, Egypt, and the Netherlands in 2009-2014 (Krog et al., 2015). Interestingly, at the amino acid level, the HA of the viruses from Denmark and Germany was 99.3% identical and the very first Swedish isolate was only around 97.5% identical to all the other strains (Krog et al., 2015). This low amino acid identity of HA was reflected by nucleotide mutations (dN/dS = 0.7) indicating that adaptation to seals was in progress (Krog et al., 2015). A limited number of seals (<180) were also found dead in the Netherlands from early November 2014 until early January 2015 (Bodewes et al. 2015). A serological investigation showed that antibodies against A(H10N7) was found in 41% (32 out of 78) pups, 10% (5 out of 52) weaners, and 58% (7 out of 12) subadults or adults harbor seals in 2015 (Bodewes et al. 2015). Although no cases of dead grey seals were reported, 26% (5 out of 19) were seropositive to influenza virus in 2015 (Bodewes et al. 2015). These findings indicate that, despite apparent low mortality, infection with seal influenza A(H10N7) virus was geographically widespread and also occurred in grey seals (Bodewes et al. 2015). Brand et al. (2016) have shown that seal/Germany/2014 H10N7 virus cause respiratory disease in harbor seals and experimentally infected ferrets. The lesions in both species were restricted to the respiratory tract with no evidence for spread to extra-respiratory tissues (Brand et al, 2016). The viral antigen was predominantly found in bronchial and submucosal glandular epithelial cells in harbour seals whereas in ferrets they were mainly found in bronchiolar and bronchial submucosal glandular epithelial cells and less frequently in type I and II pneumocytes, bronchial, bronchial glandular and tracheal epithelial cells which might suggest on slightly different cell-receptor distribution (Brand et al, 2016). Bodewes et al. (2016) have shown that the majority of the sequence variation collected from seals during the course of the outbreak between April 2014 and January 2015 occurred in HA genes. The highest variation in HA was observed at the beginning of the epidemic, and then the number of the mutations observed earlier had been fixed suggesting that when an avian influenza virus jumps the species barrier from birds to seals, amino acid changes in HA may occur rapidly and are important for virus adaptation to its new mammalian host (Bodewes et al., 2016). Later, Dittrich et al. (2018) by reverse genetics system generated recombinant avian H10N4 viruses that caried one of eight unique mutations or the complete HA from the seal virus were rescued. Wild-type recombinant H10N4 virus had high affinity to avian-type sialic acid receptors and no affinity to mammalian-type receptors whereas Q220L (H10 numbering) in the rim of the receptor binding pocket increased the affinity of the H10N4 virus to mammal-type receptors and completely abolished the affinity to avian-type receptors (Dittrich et al. 2018). Furthermore, all viruses, including the wild-type H10N7 virus, replicated at higher levels in chicken cells than in human cells suggesting that adaptive mutations (e.g., Q220L) enhanced replication in mammals and retained replication efficiency in the original avian host (Dittrich et al. 2018). Herfst et al. (2020) have shown that A/H10N7 viruses isolated from seals in Europe (A/harbour seal/NL/PV14–221_TS/2015 and A/harbor seal/S1047_14_L/Germany/2014) obtained changes in their HA segment that decreased avidity of the virus for avian-type receptors and an increase in its preference for human-type receptors. Those changes were caused by substitutions in the 220-loop that forms one edge of the receptor-binding pocket, in particular the amino acid substitution Q226L which showed stronger binding to the human-type sialic acid receptor (Herfst et al., 2020). Furthermore, these seal A/H10N7 viruses were aerosol or respiratory droplet-transmissible between ferrets (Herfst et al., 2020). Interestingly, Guan et al. (2019) have shown that H10N7 gull isolates found in Iceland in 2015, that were genetically related to H10 that cause influenza outbreaks and deaths among European seals in 2014, can also be transmitted between ferrets through the direct contact and aerosol routes, without prior adaptation.
Later, the H10N7 virus was also isolated from 2 dead harbor seal in British Columbia, Canada in 2021, that died from bronchointerstitial pneumonia (Berhane et al., 2022). This new H10N7 virus isolated in Canada is a reassortant virus between North America and Eurasia lineages (Berhane et al., 2022). Interestingly, this virus carries a mutation at position 701 in PB2 segment (D701N) that was not seen in the other H10N7 isolates from seals (Berhane et al., 2022, Suppl. Figure 3). Based on HA and NA genetic analysis performed here the H10N7 viruses isolated from seals in Europe in 2014 clustered together and H10N7 Canadian isolate from 2021 were closely related to European H10N7 and H7N7 isolates from seals in 1980s in USA (Suppl. Fig 5 and 6). Interestingly, PB2 segment of H10N7 Canadian seal isolate from 2021 were closely related to 2.3.4.4b H5N1 seal isolates obtained in 2023 in USA (above 99.94% similarity) and current South America isolates from marine animals (such as sea lion, dolphin, porpoise) (Figure 3). This very high similarity was most probably due to D701N substitution in PB2 gene observed in the Canadian seal H10N7 and current H5N1 isolates (Suppl. Figure 3). Similarly to PB2, PB1 segment also clustered with current H5N1 2.3.4.4b isolates (Figure 3 and Figure 4). In addition, PB1 and MP segments were also closely related to H3N8 isolates obtained from seals in USA in 2011 and 2018 (Figure 4 and Figure 6). PA segment clustered with seal H4N5 isolate from 1982 USA and H1N1 isolates obtained from seals in California in 2011 but not with European H10N7 isolates (Figure 5). MP segment clustered with H3N8 isolates obtained from seals in USA in 2011 and 2018 (Figure 6). The phylogenetics analysis of NP, HA, NA and NS segments is shown in Suppl. Fig 4, Suppl. Figure 5, Suppl. Figure 6 and Suppl. Figure 7, respectively.
Pandemic H1N1 (2010, 2019)
In 2010, the pandemic H1N1 (pH1N1) in northern elephant seals on California coast was found which was the first report of pH1N1 in any marine mammal. None of the seals appeared ill (Goldstein et al., 2013). Genetic analysis showed greater than 99% homology for all segments to pandemic influenza A/California/04/2009 that circulated in people in California in 2009 (Goldstein et al., 2013). The antibodies to pH1N1 were detected in seals population in 2010 (and the virus was isolated). The antibodies to pH1N1 were also detected in following years 2011 and 2012 even though the PCR results were negative (Goldstein et al., 2013). Interestingly, antibodies to pandemic H1N1 were also detected in pups born in 2011 even though the seals were tested negative to the virus which could suggest on passive transfer of maternal antibodies to the pups (Goldstein et al., 2013). Between the 2011-2018 the surveillance in marine mammals along the California coast was conducted and the results shown frequent detection of antibodies against influenza virus in seals but rare PCR positive virus detection (Plancarte et al., 2023). In spring 2019, ten samples from northern elephant seals (Mirounga angustirostris) were PCR positive for pH1N1 avian influenza virus although the virus isolation was not successful. This represents the first report of human A(H1N1)pdm09 IAV in northern elephant seals since 2010, suggesting IAV continues to spill over from humans to pinnipeds (Plancarte et al., 2023).
The first reported Cases of Highly Pathogenic Avian Influenza Viruses (HPAIV) in Seals
HPAIV: H5N8 Clade 2.3.4.4b (2016/2017, 2021, 2022)
Since the first report of AIV in seals in 1979-1980s, it took over 30 years for the HPAIV to reach seal population and cause clinical disease. Two gray seals were found dead on the Baltic coast of Poland, first one on November 27, 2016 and the second one 5 months later (Dai-Lun Shin et al., 2019). The lung samples were collected and A/H5N8 clade 2.4.4.4 b virus was confirmed in both cases (Dai-Lun Shin et al., 2019). Only one virus was isolated, the isolation of the virus from the other animal failed; however, direct sequencing of the HA and NA genes was performed. This H5N8/seal virus 99.7%–100% similar to viruses that were circulating in aquatic wild bird species during the avian influenza outbreaks in 2016 and 2017. No mutations in PB2 segment of seal/H5N8 were found. This first findings of HPAIV in seals suggested that cross-species transmissions can occur sporadically, and possibility of seal-to-seal transmission should not be excluded (Dai-Lun Shin et al., 2019).
A few years later, H5N8 HPAIV was detected in harbour and grey seals in the UK (2021), harbour seals in Germany (2021) and one harbour seal in Denmark (2022). An unusual disease event occurred at Rehabilitation Center in the UK where few mammals, including red fox, gray seal and several common seals were dead due to H5N8 virus infection. The seals exhibited sudden-onset neurologic signs, including seizures before death or euthanasia. These events occurred roughly 1 week after 5 swans housed in the same quarantine unit died from infections with HPAI H5N8 virus (Floyd et al., 2021). Based on genetic and epidemiologic investigations the swans were most likely the source of infection for the fox and seals; virus transmission likely occurred by fomite transfer or aerosol spread (Floyd et al., 2021). The D701N amino acid substitution in PB2 gene identified in both sequences derived from the mammalian species and was absent from all avian sequences generated during this 2020–2021 outbreak in the United Kingdom (Floyd et al., 2021).
In mid-August 2021 three adults harbour seals (Phoca vitulina) were found dead at the German North Sea coast (Postel et al. 2022). Two variants of H5N8 virus were detected from the seal samples; genotype Ger-10-20-N8 that dominated the avian epizootic 2020/2021, and was found in Germany from October 2020 until July 2021 and the second, clustered with genotype Ger-02-21-N8, a much rarer genotype that was only detected three times in Germany (in February and March 2021) with a novel NP segment sharing the highest identity with Eurasian LPAIV strains found in wild birds (Postel et al., 2022). These findings suggest that there were most probably at least two independent entries of the H5N8 into seal population in Germany (Postel et al., 2022). Some of these viruses possess a mutation E637K in PB2 segment (Suppl. Figure 3).
HPAIV: H5N1 Clade 2.3.4.4b (2022-Current)
The current outbreak of 2.3.4.4b H5N1 viruses affected over 43 mammalian species, including marine mammals, across Europe, North America, South America, and Asia (EFSA, April-June 2023). The 2.3.4.4b H5N1 virus in seals was reported by North and South America (USA, Canada, Argentina, Chile, Uruguay, Brazil) and Europe (Germany, Russia, Denmark, South Georgia and the South Sandwich Islands, UK).
On 8 October 2023, the 2.3.4.4b H5N1 virus for a first-time reach to Antarctic and sub-Antarctic regions of South Georgia and the Falkland Islands (British overseas territory of South Georgia at Bird Island) (Bennison et. al, 2023). The main infected bird species were brown skuas where the mortality rate increased rapidly throughout the month. The virus was confirmed in skuas and kelp gulls across four different sampling locations in South Georgia and southern fulmar in the Falkland Islands (Bennison et. al, 2023). Clinical disease was also manifested in elephant and fur seals in South Georgia (Bennison et. al, 2023). Genetic assessment of the virus indicates spread from South America, likely through movement of migratory birds (Bennison et. al, 2023).
In South America, Argentina reported a mas mortality of young pups born in 2023 across Argentina, which represented almost 96% of all southern elephant (Mirounga leonina) seal pups born across Argentina (NEWS, 24 January 2024). Previously, a mass mortality event of more than 3,000 sea lions (Otaria flavescens) was observed in January and February 2023 in Peru (Gamara-Toledo et al., 2023). These viruses belong to the HPAI A/H5N1 lineage 2.3.4.4b and are 4:4 reassortants, where PA, HA, NA and MP belong to Eurasian lineage that initially entered North America from Eurasia and remaining PB2, PB1, NP and NS came from American lineage that was already circulating in North America (Leguia et al 2023). As reported by Leguia et al (2023) these viruses do not enquire mutations linked to mammalian host adaptation and enhanced transmission (such as PB2 E627K or D701N), but at least 8 novel polymorphic sites were found in their genome. Occurrence of the H5 virus in unusual host, sea lions (Otaria flavescens) was also reported in Chile on 10 February 2023 (WOAH report, March 2023). Genetic characterisation of isolated obtained from birds and marine mammals viruses revealed that all Chilean H5N1 viruses belong to lineage 2.3.4.4b and cluster monophyletically with viruses from Peru, indicating a single introduction from North America into Peru/Chile (Pardo-Roa et al, 2023). There were D701N (in two sea lions, one human, and one shorebird) and Q591K (human and one sea lion) mutations identified in PB2 segments (Pardo-Roa et al, 2023 BiorxV). Interesting, minor population of viruses carrying D701N mutation was present in 52.9% – 70.9% of sequence reads obtained from the samples tested suggesting on mix population of viruses within the sample (Pardo-Roa et al, 2023).
Previously, HPAIV H5N1 infection was reported in seals in USA (New England). After sequencing of 71 avian- and 13 seal-derived virus genomes from New England, in contrary to what was reported by Leguia et al (2023), all but 1 virus represented non reassortant Eurasia 2.3.4.4b viruses (Puryear et al. 2023). The authors concluded that virus outbreak among New England harbour and grey seals was concurrent with a wave of avian infections in the region and the evidence of mammal adaptation existed in a small subset of seals (PB2 E627K or D701N mutations) (Puryear et al. 2023). Many years before the HPAIV outbreak in seals, Puryear et al. 2016 have shown that North Atlantic grey seals from Cape Cod, MA, USA and Nova Scotia, Canada were consistently for IAV infection and positive sera showed a broad reactivity to diverse influenza subtypes suggesting that seals might possibly representing an endemically infected wild reservoir of IAV.
Here we performed a simple analysis of PB2 proteins of 72 isolates available in GSAID that were isolated from seals or other marine mammals (58 seals isolates of different subtype isolated between 1980-2023 along with recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise) (Suppl. Figure 3). A mutation D701N was found in H3N8 seal isolates isolated in 2011 and 2018 in USA and H10N7 seal isolate obtained from Canadian case in 2021. However, no mutation D701N was found in H10N7 from 2014 and 2015 isolated during epizootic in Europe. Several mutations D701N in PB2 were also found in recent 2.3.4.4b H5N8 and H5N1 seals isolates, and sea lions, dolphins and porpoise isolated from South America in 2023 (Suppl. Figure 3). Interestingly, none of the LPAIV seal isolate carried mutation E627K in PB2 and this mutation was only found in some of H5Nx viruses of 2.4.4.4b clade isolated recently (eg. H5N8 from Germany, Denmark or H5N1 from Scotland, USA) (Suppl. Figure 3).
Summary
In this short review we have shown 45 years of AIV history in seal population. The very first cases of AIV were reported in harbour seal population (Phoca vitulina) in late 1970s. Later, the virus was also isolated from grey seals (Halichoerus grypus). All AIV viruses between 1979-2016/17 were of low pathogenic pathotype (LPAIV). Some of LPAIV showed interesting features that allowed them to adapt to a new species (such as enquired mutation in PB2 segment) but not all of them, suggesting that the virus is able to replicate in the seal population with or without clinical sign of disease. This was later confirmed by seroprevalence and virus isolation. The H5N8 HPAIV clade 2.3.4.4b was found for a first time in a seal population in 2016/17. Current 2.3.4.4b H5N1 virus is causing massive mortality of seals and sea lions in America drastically reducing its population. Importantly, the AIV HPAIV H5N1 virus reached Antarctic region which is an important breeding ground for many key Antarctic species. This is alarming and along with a climate change might have impair ecology of seabirds, penguins, and marine mammals in this remote region. Previously the evidence of influenza A virus in various animals was reported in Arctic regions and 26 unique low and highly pathogenic subtypes have been characterized in the scientific literature (Gass et al., 2022). Furthermore, the papers cited here are also strong evidence for interspecies transmission of AIV between birds and marine mammals and shows that seals might be an important wild reservoir of influenza and may contribute to mammalian adaptation of avian variants.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- EFSA. Avian influenza overview April – June 2023. EFSA journal . [CrossRef]
- Briggs K, Kapczynski DR. Comparative analysis of PB2 residue 627E/K/V in H5 subtypes of avian influenza viruses isolated from birds and mammals. Front Vet Sci 2023, 10, 1250952. [CrossRef]
- National Oceanic and AtmosNOAA 2024. Available at https://www.fisheries.noaa.gov/.
- Haug, T., M.O. Hammill and D. Ólafsdóttir (Eds.). 2007. Grey seals in the North Atlantic and the Baltic. NAMMCO Scientific Publications. 6:7–12.
- Stewart BS, Yochem PK, Le Boeuf BJ, Huber HR, DeLong RL, Jameson RJ, et al. Population recovery and status of the northern elephant seal, Mirounga angustirostris. In: Le Boeuf BJ, Laws RM, editors. Elephant Seals: Population Ecology, Behavior, and Physiology. University of California Press; 1994. p. 29–48.
- Nature Scotland 2024. Available at hps://www.nature.scot/.
- Wood SA, Murray KT, Josephson E., Gilbert J. Rates of increase in gray seal (Halichoerus grypus atlantica) pupping at recolonized sites in the United States, 1988–2019. Journal of Mammalogy, Volume 101, Issue 1, 21 February 2020, Pages 121–128 . [CrossRef]
- Townsend, CH. The Northern Elephant Seal. New York Zoological Society. 1912. [Google Scholar]
- Lowry MS, Condit R, Hatfield B, Allen SG, Berger R, Laake J, et al. Abundance, distribution, and growth of the northern elephant seal, Mirounga angustirostris, in the United States in 2010. Aquatic Mammals. 2014; 40(1):20–31. [CrossRef]
- Stewart, J.E.; et al. (2014). "Finescale ecological niche modeling provides evidence that lactating grey seals (Halichoerus grypus) prefer access to fresh water in order to drink" (PDF). Marine Mammal Science. 30 (4): 1456–1472. [CrossRef]
- Boyd, I. L.; Walker, T. R.; Poncet, J. (1996). Walton, David W. H.; Vaughan, Alan P. M.; Hulbe, Christina L. (eds.). "Status of Southern Elephant seals at South Georgia". Antarctic Science. 8 (3): 237–244.
- NEWS. Available at htps://www.telegraph.co.uk/global-health/science-and-disease/bird-flu-kills-17000-elephant-seal-pups-in-argentina/. 1700.
- Lang G, Gagnon A, Geraci JR. Isolation of an influenza A virus from seals. Arch Virol. 1981;68(3-4):189-95. [CrossRef]
- Webster RG, Hinshaw VS, Bean WJ, Van Wyke KL, Geraci JR, St Aubin DJ, Petursson G. Characterization of an influenza A virus from seals. Virology. 1981 Sep;113(2):712-24. [CrossRef]
- Geraci JR, St Aubin DJ, Barker IK, Webster RG, Hinshaw VS, Bean WJ, Ruhnke HL, Prescott JH, Early G, Baker AS, Madoff S, Schooley RT. Mass mortality of harbor seals: pneumonia associated with influenza A virus. Science. 1982 Feb 26;215(4536):1129-31. [CrossRef]
- Hinshaw VS, Webster RG, Easterday BC, Bean WJ Jr. Replication of avian influenza A viruses in mammals. Infect Immun. 1981 Nov;34(2):354-61. [CrossRef]
- Murphy BR, Harper J, Sly DL, London WT, Miller NT, Webster RG. Evaluation of the A/Seal/Mass/1/80 virus in squirrel monkeys. Infect Immun. 1983 Oct; 42(1): 424–426. [CrossRef]
- Li SQ, Orlich M, Rott R. Generation of seal influenza virus variants pathogenic for chickens, because of hemagglutinin cleavage site changes. J Virol. 1990 Jul;64(7):3297-303. [CrossRef]
- Hinshaw VS, Bean WJ, Webster RG, Rehg JE, Fiorelli P, Early G, Geraci JR, St Aubin DJ. Are seals frequently infected with avian influenza viruses? J Virol. 1984 Sep;51(3):863-5. [CrossRef]
- Austin FJ, Webster RG. Evidence of ortho- and paramyxoviruses in fauna from Antarctica. J Wildl Dis. 1993 Oct;29(4):568-71. [CrossRef]
- Naeve C. W., Webster R. G. Sequence of the hemagglutinin gene from influenza virus A/Seal/Mass/1 /80. 1983Virology 129:298–308.
- Donis R. O., Bean W. J., Kawaoka Y., Webster R. G. Distinct lineages of influenza virus H4 hemagglutinin genes in different regions of the world. 1989; Virology 169:408–417. 1989.
- Callan RJ, Early G, Kida H, Hinshaw VS. The appearance of H3 influenza viruses in seals. J Gen Virol. 1995 Jan;76 ( Pt 1):199-203. [CrossRef]
- Matrosovich M, Tuzikov A, Bovin N, Gambaryan A, Klimov A, Castrucci MR, Donatelli I, Kawaoka Y. Early alterations of the receptor-binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals. J Virol. 2000 Sep;74(18):8502-12. [CrossRef]
- Ramis AJ, van Riel D, van de Bildt MW, Osterhaus A, Kuiken T. Influenza A and B virus attachment to respiratory tract in marine mammals. Emerg Infect Dis. 2012 May;18(5):817-20. [CrossRef]
- Gulyaeva M, Sobolev I, Sharshov K, Kurskaya O, Alekseev A, Shestopalova L, Kovner A, Bi Y, Shi W, Shchelkanov M, Shestopalov A. Characterization of Avian-like Influenza A (H4N6) Virus Isolated from Caspian Seal in 2012. Virol Sin. 2018 Oct;33(5):449-452. [CrossRef]
- Ohishi K, Ninomiya A, Kida H, Park C-H, Maruyama T, Arai T, Katsumata E, Tobayama T, Boltunov AN, Khuraskin LS, Miyazaki N. Serological evidence of transmission of human influenza A and B viruses to Caspian seals (Phoca caspica). Microbiol Immunol 2002;46(9):639-44. [CrossRef]
- Bogomolni AL, Gast RJ, Ellis JC, Dennett M, Pugliares KR, Lentell BJ, Moore MJ. Victims or vectors: a survey of marine vertebrate zoonoses from coastal waters of the Northwest Atlantic. Dis Aquat Organ 2008. 81:13–38.
- Anthony SJ, St Leger JA, Pugliares K, Ip HS, Chan JM, Carpenter ZW, Navarrete-Macias I, Sanchez-Leon M, Saliki JT, Pedersen J, Karesh W, Daszak P, Rabadan R, Rowles T, Lipkin WI. Emergence of fatal avian influenza in New England harbor seals. mBio. 2012 Jul 31;3(4):e00166-12. [CrossRef]
- Yang H, Nguyen HT, Carney PJ, Guo Z, Chang JC, Jones J, Davis CT, Villanueva JM, Gubareva LV, Stevens J. Structural and functional analysis of surface proteins from an A(H3N8) influenza virus isolated from New England harbor seals. J Virol. 2015 Mar;89(5):2801-12. [CrossRef]
- Karlsson EA, Ip HS, Hall JS, Yoon SW, Johnson J, Beck MA, Webby RJ, Schultz-Cherry S. Respiratory transmission of an avian H3N8 influenza virus isolated from a harbour seal. Nat Commun. 2014 Sep 3;5:4791. [CrossRef]
- Hussein IT, Krammer F, Ma E, Estrin M, Viswanathan K, Stebbins NW, Quinlan DS, Sasisekharan R, Runstadler. New England harbor seal H3N8 influenza virus retains avian-like receptor specificity. J. Sci Rep. 2016 Feb 18;6:21428. [CrossRef]
- Venkatesh D, Bianco C, Núñez A, Collins R, Thorpe D, Reid SM, Brookes SM, Essen S, McGinn N, Seekings J, Cooper J, Brown IH, Lewis NS. Detection of H3N8 influenza A virus with multiple mammalian-adaptive mutations in a rescued Grey seal (Halichoerus grypus) pup. Virus Evol. 2020 Mar 18;6(1):veaa016. [CrossRef]
- Bao P, Liu Y, Zhang X, Fan H, Zhao J, Mu M, Li H, Wang Y, Ge H, Li S, Yang X, Cui Q, Chen R, Gao L, Sun Z, Gao L, Qiu S, Liu X, Horby PW, Li X, Fang L, Liu W. Human infection with a reassortment avian influenza A H3N8 virus: an epidemiological investigation study. Nat Commun. 2022 Nov 10;13(1):6817. [CrossRef]
- Tan X, Yan XT, Liu Y, Wu Y,Liu JY, Mu M,Zhao J, Wang XY 1, Li JQ, Wen L, Guo P, Zhou ZG, Li XB, Bao PT. A case of human infection by H3N8 influenza virus. Emerg Microbes Infect. 2022 Dec;11(1):2214-2217. [CrossRef]
- Wang Y, Wang M, Zhang H, Zhao C, Zhang Y, Shen J, Sun X, Xu H, Xie Y, Gao X, Cui P, Chu D, Li Y, Liu W, Peng P, Deng G, Guo J, Li X. Prevalence, evolution, replication and transmission of H3N8 avian influenza viruses isolated from migratory birds in eastern China from 2017 to 2021. Emerg Microbes Infect. 2023 Dec;12(1):2184178. [CrossRef]
- Zohari S, Neimanis A, Härkönen T, Moraeus C, Valarcher JF. Avian influenza A(H10N7) virus involvement in mass mortality of harbour seals (Phoca vitulina) in Sweden, March through October 2014. Euro Surveill. 2014 Nov 20;19(46):20967. 20 October. [CrossRef]
- Bodewes R, Bestebroer TM, van der Vries E, Verhagen JH, Herfst S, Koopmans MP, Fouchier RA, Pfankuche VM, Wohlsein P, Siebert U, Baumgärtner W, Osterhaus AD. Avian Influenza A(H10N7) virus-associated mass deaths among harbor seals. Emerg Infect Dis. 2015 Apr;21(4):720-2. [CrossRef]
- Krog JS, Hansen MS, Holm E, Hjulsager CK, Chriél M, Pedersen K, O. Andresen L, Abildstrøm M, Jensen TH, Larsen LE. Influenza A(H10N7) Virus in Dead Harbor Seals, Denmark. Emerg Infect Dis. 2015 Apr; 21(4): 684–687. [CrossRef]
- Bodewes R, Zohari S, Krog JS, Hall MD, Harder TC, Bestebroer TM, van de Bildt MWG, Spronken MI, Larsen LE, Siebert U, Wohlsein P, Puff C, Seehusen F, Baumgärtner W, Härkönen T, Smits SL, Herfst S, Osterhaus ADME, Fouchier RAM, Koopmans MP, Kuiken T. Spatiotemporal Analysis of the Genetic Diversity of Seal Influenza A(H10N7) Virus, Northwestern Europe. J Virol. 2016 Apr 14;90(9):4269-4277. [CrossRef]
- van den Brand JM, Wohlsein P, Herfst S, Bodewes R, Pfankuche VM, van de Bildt MW, Seehusen F, Puff C, Richard M, Siebert U, Lehnert K, Bestebroer T, Lexmond P, Fouchier RA, Prenger-Berninghoff E, Herbst W, Koopmans M, Osterhaus AD, Kuiken T, Baumgärtner W. Influenza A (H10N7) Virus Causes Respiratory Tract Disease in Harbor Seals and Ferrets. PLoS One. 2016 Jul 22;11(7):e0159625. [CrossRef]
- Dittrich A, Scheibner D, Salaheldin AH, Veits J, Gischke M, Mettenleiter TC, Abdelwhab EM. Impact of Mutations in the Hemagglutinin of H10N7 Viruses Isolated from Seals on Virus Replication in Avian and Human Cells. Viruses. 2018 Feb; 10(2): 83. [CrossRef]
- Herfst S, Zhang J, Richard M, McBride R, Lexmond P, Bestebroer TM, Spronken MIJ, de Meulder D, van den Brand JM, Rosu ME, Martin SR, Gamblin SJ, Xiong X, Peng W, Bodewes R, van der Vries E, Osterhaus ADME, Paulson JC, Skehel JJ, Fouchier RAM. Hemagglutinin Traits Determine Transmission of Avian A/H10N7 Influenza Virus between Mammals. Cell Host Microbe. 2020 Oct 7;28(4):602-613.e7. [CrossRef]
- Guan M, Hall JS, Zhang X, Dusek RJ, Olivier AK, Liu L, Li L, Krauss S, Danner A, Li T, Rutvisuttinunt W, Lin X, Hallgrimsson GT, Ragnarsdottir SB, Vignisson SR, TeSlaa J, Nashold SW, Jarman R, Wan X-F. Aerosol Transmission of Gull-Origin Iceland Subtype H10N7 Influenza A Virus in Ferrets. J Virol. 2019 Jul 1; 93(13): e00282-19. [CrossRef]
- Berhane Y, Joseph T, Lung O, Embury-Hyatt C, Xu W, Cottrell P, Raverty S. Isolation and Characterization of Novel Reassortant Influenza A(H10N7) Virus in a Harbor Seal, British Columbia, Canada. Emerg Infect Dis. 2022 Jul; 28(7): 1480–1484. [CrossRef]
- Goldstein T, Mena I, Anthony SJ, Medina R, Robinson PW, Greig DJ, Costa DP, Lipkin WI, Garcia-Sastre A, Boyce WM. Pandemic H1N1 influenza isolated from free-ranging Northern Elephant Seals in 2010 off the central California coast. PLoS One. 2013 ;8(5):e62259. 15 May. [CrossRef]
- Plancarte M, Kovalenko GG, Baldassano J, Ramírez AL, Carrillo S, Duignan PG, Goodfellow I, Bortz E, Dutta J, van Bakel H, Coffey LL. Human influenza A virus H1N1 in marine mammals in California, 2019. PLoS One 2023 Mar 30;18(3):e0283049. [CrossRef]
- Shin DL, Siebert U, Lakemeyer J, Grilo M, Pawliczka I, Wu NH, Valentin-Weigand P, Haas L, Herrler G. Highly Pathogenic Avian Influenza A(H5N8) Virus in Gray Seals, Baltic Sea. Emerg Infect Dis. 2019 Dec;25(12):2295-2298. [CrossRef]
- Floyd T, Banyard AC, Lean FZX, Byrne AMP, Fullick E, Whittard E, Mollett BC, Bexton S, Swinson V, Macrelli M, Lewis NS, Reid SM, Núñez A, Duff JP, Hansen R, Brown IH. Encephalitis and Death in Wild Mammals at a Rehabilitation Center after Infection with Highly Pathogenic Avian Influenza A(H5N8) Virus, United Kingdom. Emerg Infect Dis. 2021 Nov;27(11):2856-2863. [CrossRef]
- Postel A, King J, Kaiser FK, Kennedy J, Lombardo MS, Reineking W, de le Roi M, Harder T, Pohlmann A, Gerlach T, Rimmelzwaan G, Rohner S, Striewe LC, Gross S, Schick LA, Klink JC, Kramer K, Osterhaus ADME, Beer M, Baumgärtner W, Siebert U, Becher P. Infections with highly pathogenic avian influenza A virus (HPAIV) H5N8 in harbor seals at the German North Sea coast, 2021. Emerg Microbes Infect. 2022 Dec;11(1):725-729. [CrossRef]
- Bennison A, Byrne AMP, Reid SM, Lynton-Jenkins JG, Mollett B, De Sliva D, Peers-Dent J, Finlayson K, Hall R, Blockley F, Blyth M, Falchieri M, Fowler Z, Fitzcharles EM, Brown IH, James J, Banyard AC. Detection and spread of high pathogenicity avian influenza virus H5N1 in the Antarctic Region. bioRxiv 2023. [CrossRef]
- Gamarra-Toledo V, Plaza PI, Gutiérrez R, Inga-Diaz G, Saravia-Guevara P, Pereyra-Meza O, Coronado-Flores E, Calderón-Cerrón A, Quiroz-Jiménez G, Martinez P, Huamán-Mendoza D, Nieto-Navarrete JC, Ventura S, Lambertucci SA. Mass Mortality of Sea Lions Caused by Highly Pathogenic Avian Influenza A(H5N1) Virus Emerg Infect Dis 2023 Dec;29(12):2553-2556. [CrossRef]
- Leguia M, Garcia-Glaessner A, Muñoz-Saavedra B, Juarez D, Barrera P, Calvo-Mac C, Jara J, Silva W, Ploog K, Amaro L, Colchao-Claux P, Johnson CK, Uhart MM, Nelson MI, Lescano J. Highly pathogenic avian influenza A (H5N1) in marine mammals and seabirds in Peru. Nat Commun. 2023; 14: 5489. Published online 2023 Sep 7. [CrossRef]
- World Animal Health Information System (WOAH-WAHIS) report, 23. Available at https://www.woah. 20 March.
- Pardo-Roa C, Nelson MI, Ariyama N, Aguayo C, Almonacid LI, Munoz G, Navarro C, Avila C, Ulloa M, Reyes R, Luppichini EF, Mathieu C, Vergara R, González A, González CG, Araya H, Fernández J, Fasce R, Johow M, Medina RA, Neira V. Cross-species transmission and PB2 mammalian adaptations of highly pathogenic avian influenza A/H5N1 viruses in Chile. bioRxiv. Preprint. 2023 Jun 30. [CrossRef]
- Puryear W, Sawatzki K, Hill N, Foss A, Stone JJ, Doughty L, Walk D, Gilbert K, Murray M, Cox E, Patel P, Mertz Z, Ellis S, Taylor J, Fauquier D, Smith A, DiGiovanni Jr RA, van de Guchte A, Gonzalez-Reiche AS, Khalil Z, van Bakel H, Torchetti MK, Lantz K, Lenoch JB, Runstadler J. Highly Pathogenic Avian Influenza A(H5N1) Virus Outbreak in New England Seals, United States. Emerg Infect Dis. 2023 Apr;29(4):786-791. [CrossRef]
- Puryear W, Keogh M, Hill N, Moxley J, Josephson E, Davis KR, Bandoro C, Lidgard D, Bogomolni A, Levin M, Lang S, Hammill M, Bowen D, Johnston DW, Romano T, Waring G, Runstadler J. Prevalence of influenza A virus in live-captured North Atlantic gray seals: a possible wild reservoir. Emerg Microbes Infect 2016 Aug 3;5(8):e81. [CrossRef]
- Gass Jr JD, Hunter K Kellogg HK, Hill NJ, Puryear WB, Nutter FB, Runstadler JA. Epidemiology and Ecology of Influenza A Viruses among Wildlife in the Arctic. Viruses 2022 Jul 13;14(7):1531. [CrossRef]
Figure 1.
The Horsey Grey Seals Colony, United Kingdom, February 2024. Photography by Klaudia Chrzastek.
Figure 1.
The Horsey Grey Seals Colony, United Kingdom, February 2024. Photography by Klaudia Chrzastek.

Figure 2.
Phylogenetic analysis of HA segment of H4N5 avian influenza viruses isolates obtained from seal in 1982 (marked in red) and wild bird isolates available in database (N=22).
Figure 2.
Phylogenetic analysis of HA segment of H4N5 avian influenza viruses isolates obtained from seal in 1982 (marked in red) and wild bird isolates available in database (N=22).

Figure 3.
Phylogenetic analysis of PB2 segment of avian influenza viruses found in seal population over the last 45 years. The analysis includes 72 isolates available in GSAID (as of March 2024): 58 seals isolate of different subtype isolated between 1980-2023 and recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise. The nucleotide sequences of PB2 segment were aligned using MUSCLE software and GTR nucleotide substitution model, with among-site rate variation model using a discrete gamma distribution. Bootstrap support values were generated using 500 rapid bootstrap replicates.
Figure 3.
Phylogenetic analysis of PB2 segment of avian influenza viruses found in seal population over the last 45 years. The analysis includes 72 isolates available in GSAID (as of March 2024): 58 seals isolate of different subtype isolated between 1980-2023 and recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise. The nucleotide sequences of PB2 segment were aligned using MUSCLE software and GTR nucleotide substitution model, with among-site rate variation model using a discrete gamma distribution. Bootstrap support values were generated using 500 rapid bootstrap replicates.

Figure 4.
Phylogenetic analysis of PB1 segment of avian influenza viruses found in seal population over the last 45 years. The analysis includes PB1 segments that are available in GSAID (as of March 2024) isolated between 1980-2023 and recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise. The nucleotide sequences of PB1 segment were aligned using MUSCLE software and GTR nucleotide substitution model, with among-site rate variation model using a discrete gamma distribution. Bootstrap support values were generated using 500 rapid bootstrap replicates.
Figure 4.
Phylogenetic analysis of PB1 segment of avian influenza viruses found in seal population over the last 45 years. The analysis includes PB1 segments that are available in GSAID (as of March 2024) isolated between 1980-2023 and recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise. The nucleotide sequences of PB1 segment were aligned using MUSCLE software and GTR nucleotide substitution model, with among-site rate variation model using a discrete gamma distribution. Bootstrap support values were generated using 500 rapid bootstrap replicates.

Figure 5.
Phylogenetic analysis of PA segment of avian influenza viruses found in seal population over the last 45 years. The analysis includes PA segments that are available in GSAID (as of March 2024) isolated between 1980-2023 and recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise. The nucleotide sequences of PA segment were aligned using MUSCLE software and GTR nucleotide substitution model, with among-site rate variation model using a discrete gamma distribution. Bootstrap support values were generated using 500 rapid bootstrap replicates.
Figure 5.
Phylogenetic analysis of PA segment of avian influenza viruses found in seal population over the last 45 years. The analysis includes PA segments that are available in GSAID (as of March 2024) isolated between 1980-2023 and recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise. The nucleotide sequences of PA segment were aligned using MUSCLE software and GTR nucleotide substitution model, with among-site rate variation model using a discrete gamma distribution. Bootstrap support values were generated using 500 rapid bootstrap replicates.
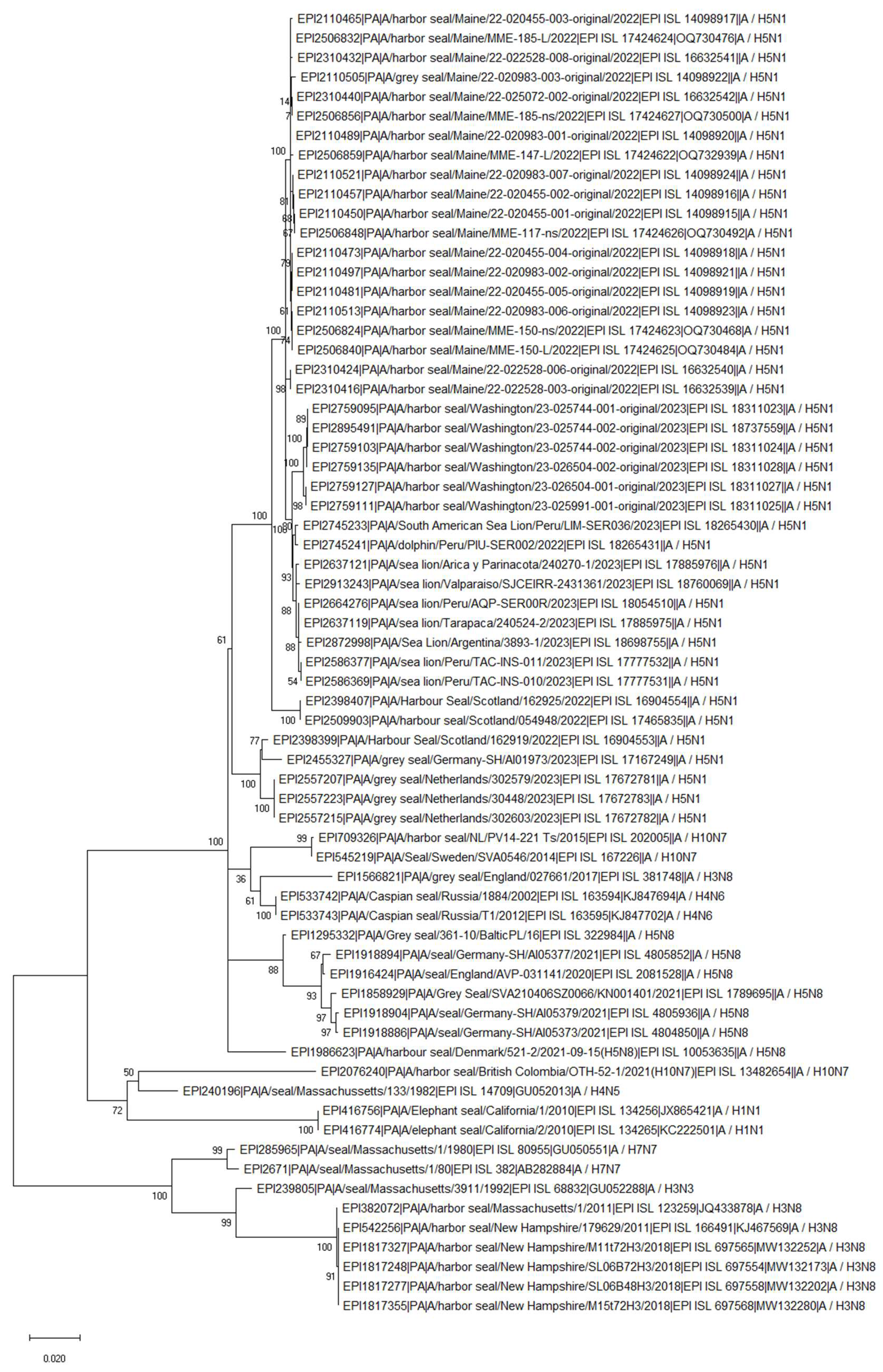
Figure 6.
Phylogenetic analysis of MP segment of avian influenza viruses found in seal population over the last 45 years. The analysis includes MP segments that are available in GSAID (as of March 2024) isolated between 1980-2023 and recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise. The nucleotide sequences of MP segment were aligned using MUSCLE software and GTR nucleotide substitution model, with among-site rate variation model using a discrete gamma distribution. Bootstrap support values were generated using 500 rapid bootstrap replicates.
Figure 6.
Phylogenetic analysis of MP segment of avian influenza viruses found in seal population over the last 45 years. The analysis includes MP segments that are available in GSAID (as of March 2024) isolated between 1980-2023 and recently isolated H5N1 clade 2.3.4.4b viruses from 8 sea lions, 3 dolphins and 3 porpoise. The nucleotide sequences of MP segment were aligned using MUSCLE software and GTR nucleotide substitution model, with among-site rate variation model using a discrete gamma distribution. Bootstrap support values were generated using 500 rapid bootstrap replicates.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



