
Submitted:
03 September 2024
Posted:
04 September 2024
Read the latest preprint version here
Abstract
Groupers (Epinephelidae) are ecologically important mesopredators that support valuable fisheries across the globe. Many groupers display slow growth and maturity, high longevity, ontogenetic habitat shifts, spawning-related migrations and aggregations, and protogynous hermaphroditism, which make them susceptible to overexploitation. In this review, I synthesize available information related to the management of grouper fisheries across the southeastern and Caribbean U.S. I highlight current management challenges, such as managing multispecies reef fish fisheries with growing recreational fishing effort. I discuss management interventions with limited success, such as establishing marine protected areas to improve the populations of groupers that display protogynous hermaphroditism. I also highlight management successes, such as recovering historically depleted grouper stocks, and ecosystem-based considerations in grouper stock assessments. I discuss how climate change and anthropogenic effects are expected to affect groupers. Lastly, I provide examples of stakeholders involved in monitoring and management efforts directed at grouper stocks. The purposes of this review are to demonstrate the complexities of managing grouper fisheries and provide a road map for future research and conservation efforts into these economically and ecologically relevant fishes within and beyond the region.
Keywords:
Serranidae
; Epinephelidae
; Reef fish
; Hermaphroditism
; Fish spawning aggregations
; Sustainable fisheries
1. Introduction
Groupers (Epinephelidae) are highly prized reef-associated mesopredators that support recreational, commercial, and artisanal fisheries across their range. They can also have important ecological roles as ecosystem engineers shaping the physical and biogenic structure [1,2]. Many groupers display slow growth and maturity, high longevity, sequential hermaphroditism, ontogenetic habitat shifts, and spawning-related migrations and aggregations, which make them susceptible to overexploitation [3]. A recent global assessment of grouper species indicated that 30% of species are data deficient, 12% risk extinction under current conditions, and 13% are considered near threatened [3]. Their life history, data availability, and population trends highlight the need to carefully consider groupers in a fisheries management context.
In this review, I present information regarding the management of grouper fisheries in the southeastern U.S. (SEUS), which is composed of the Caribbean (CAB), Gulf of Mexico (GOM), and South Atlantic (SA) subregions. I provide information on the history of these fisheries across the region, evidence of the complexities of managing grouper fisheries, and case studies of management challenges and successes. Lastly, I discuss the potential effects of climate change and other anthropogenically-driven perturbations on grouper fisheries and highlight technological advances to monitor these stocks. Providing species profiles is not among the goals of this review, but rather to draw enough detail from different species to emphasize the complexities of managing grouper fisheries. This synthesis aims to direct future research and conservation efforts into grouper fisheries within and beyond the SEUS that could be applied to other taxa.
2. Relevance and Management of Grouper Fisheries in the SEUS
In the SEUS, groupers support some of the most valuable finfish fisheries with Red Grouper Epinephelus morio and Gag Mycteroperca microlepis among the most harvested reef fishes in recreational and commercial sectors of the GOM and SA [4,5,6,7]. Harvest of groupers in the GOM can be traced back to the 1800s when settlers from the northeast U.S. traveled south to harvest Red Snapper Lutjanus campechanus, and groupers quickly went from bycatch to target species when reductions of the Red Snapper population were evident [8,9]. In the CAB, Red Hind E. guttatus has a high importance in commercial fisheries [10], and Nassau Grouper E. striatus supported both recreational and commercial fisheries until the 1970s when severe population declines started being evident [11,12,13]. In 1980, groupers were among the 3 most landed (by weight) families across the CAB [14]. Evidently, groupers distributed across the SEUS (or within any of the included subregions) display varying fishery relevance. As such, their degree of management can range from nonexistent (e.g., Marbled Grouper Dermatolepis inermis; Figure 1) to complete harvest moratorium (e.g., Nassau Grouper).
In the U.S., federal marine fishery resources are governed by the Magnuson Stevens Fishery Conservation and Management Act (MSA) and managed by the National Oceanographic and Atmospheric Administration’s National Marine Fisheries Service (NMFS). The MSA was first passed in 1976, and among its many objectives are to prevent overfishing, rebuild overfished stocks, increase long-term economic and social benefits, and maintain a safe and sustainable seafood supply [15]. To do so, regional fishery management councils were created and tasked with constructing fishery management plans (FMPs) that comply with MSA guidelines. The MSA has undergone two revisions. First, the Sustainable Fisheries Act of 1996, which recognized the importance of healthy habitats for sustainable fisheries. Second, the MSA reauthorization of 2007, which further improved fisheries science, management, and conservation. For the purposes of this review, I will provide a generalized history of FMPs in the SEUS as it pertains to grouper fisheries. Any discussion regarding marine protected areas (MPAs) will be purposely saved for the case studies throughout the review.
In the SA, the Snapper-Grouper FMP was implemented in 1983 [16]. At the time, a substantial amount of reef fishes distributed across the SA were exhibiting growth overfishing [16], or harvesting below the size that would produce maximum sustainable yield (MSY) or maximize yield-per-recruit [17]. To alleviate this, the FMP introduced a 12” minimum size limit to a suite of reef fishes, including two groupers (Red Grouper and Nassau Grouper). An amendment implemented in 1999 brought minimum size limits up to ≥20” for many groupers. The amendments listed were selectively picked to highlight some of the management efforts directed at sustainably managing SA grouper stocks. For a comprehensive history of amendments to the SA Snapper-Grouper FMP visit https://www.fisheries.noaa.gov/action/south-atlantic-snapper-grouper-historical-amendments-and-rulemaking-1983-2017.
The GOM Reef Fish FMP was implemented in 1984 and included gear restrictions to assist in rebuilding declining reef fish stocks [18]. In 1994, a 20” minimum size limit was implemented for multiple groupers (Gag, Nassau Grouper, Red Grouper, Black Grouper M. bonaci, and Yellowfin Grouper M. venenosa). A regulatory amendment implemented in 2000 introduced increases to Gag minimum size limits (from 20” to 22” for recreational and 24” for commercial sectors) and implemented the first two MPAs directed at rebuilding the Gag stock (further discussed in section 3.2). To date, a substantial amount of amendments to the GOM Reef Fish FMP involve changes to grouper stocks, particularly Red Grouper and Gag. For a comprehensive history of the GOM Reef Fish FMP visit https://www.fisheries.noaa.gov/action/gulf-mexico-reef-fish-historical-amendments-and-rulemaking-1983-2017.
The CAB Reef Fish FMP was implemented in 1985 and focused on fish trap requirements and a minimum size limit on Nassau Grouper [14]. The first amendment was implemented in 1990 and included a prohibition on the take and possession of Nassau Grouper and introduced a seasonal spatial closure of a Red Hind spawning area, known as Red Hind Bank Marine Conservation District. This seasonal spatial closure became the first MPA specifically directed at improving population levels of a grouper species in the SEUS and was permanently closed to fishing in 1999 [19,20]. Additional seasonal spatial closures during the Red Hind spawning season were implemented across the CAB during 1993, 1996, and 2005 (further discussed in section 4.1). Seasonal harvest closures during the spawning season were implemented for a suite of grouper species in 2005. In 2022, the CAB-wide FMP transitioned to island-based (Puerto Rico, St. Thomas and St. John, and St. Croix) plans. For a comprehensive history of the CAB Reef Fish FMP visit https://www.caribbeanfmc.com/fishery-management/fishery-management-plans.
3. Complexities of Managing Grouper Fisheries
3.1. Multispecies Reef Fish Fisheries
In the SEUS, groupers form part of diverse multispecies reef fish fisheries composed of species with varying life histories and exploitation histories. These fisheries provide year-round harvest opportunities for fishers as managed species have varying temporal management measures (e.g., seasonal closures). Due to non-selective (or low selectivity) fishing gear and the diversity of species sharing reef habitats, avoiding species closed to harvest is difficult, resulting in high levels of bycatch and mandatory discards. The latter induces discard mortality, which results from depredation, physical injury, or handling stress [21]. This source of mortality not only undermines minimum size limits but can also prevent meeting stock rebuilding targets. For example, the high discard mortality observed in GOM Gag has been linked to preventing stock rebuilding and potentially inducing recruitment overfishing [22].
Discards can be considered a significant source of mortality in groupers as many species are associated with deeper waters either throughout ontogeny (e.g., deepwater groupers) or during adult stages (e.g., Gag), which have been linked to increasing physical injuries (e.g., barotrauma) [23]. The MSA National Standard 9 mandates that bycatch and discard mortality be minimized, however, this has proven extremely difficult to achieve in the SEUS reef fish fisheries. It is worth noting that significant efforts have gone into understanding the mechanisms of discard mortality [24,25,26] and increasing public awareness of the utility of descender devices to mitigate discard mortality (https://returnemright.org/). Recent management changes have made it a requirement for GOM and SA reef fish fishers to carry a descending device on board [27,28].
A substantial increase in recreational effort has been observed across the GOM and Atlantic coasts in the last 40 years (Figure 2). In addition, increases in recreational landings have been reported for both the GOM and SA [29]. Due to recreational sector characteristics, such as large user group size and open access nature, recreational data contain much higher uncertainty than that of a commercial sector with mandatory reporting requirements, as observed in the GOM commercial grouper fisheries. Consequently, uncertainty in landings can increase as the recreational sector becomes more relevant unless recreational data collection programs continue to improve. Recent explorations of management strategies to rebuild the SA Red Snapper stock suggest that restricting recreational effort in the reef fish fishery could help meet stock rebuilding goals for Red Snapper and associated reef fishes [30]. Restricting recreational effort continues to be a promising avenue for rebuilding stocks, however, this must be weighed against the socioeconomic benefits that recreational fisheries provide.
Allocations of many SEUS reef fish stocks have changed over time. Increases and decreases in allocation are observed in the recreational and commercial sectors, respectively. This is especially the case for GOM Red Grouper and GOM Gag, which have recently undergone increases in the recreational sector [31,32]. As fisheries historically dominated by the commercial sector, these changes have steered commercial stakeholders to legally prosecute NMFS claiming that multiple MSA National Standards are not being met (https://www.nationalfisherman.com/gulf-south-atlantic/federal-appeals-court-orders-remand-of-red-grouper-allocations). Managing vulnerable groupers within an already complex multispecies reef fish fishery with multiple user groups, discard mortality rates, and growing uncertainty in recreational landings and effort, is a fundamental challenge faced across the region.
3.2. Protogynous Hermaphroditism
Marine reef fishes display diversity in reproductive systems represented by gonochorism, parthenogenesis, and hermaphroditism [33]. While a variety of these can be observed in groupers, hermaphroditism is most prevalent in species distributed in the SEUS (Table 1). Specifically, many of these groupers display monandric protogynous hermaphroditism (protogynous hermaphroditism hereafter), where individuals are born as females, reach sexual maturity, and then transition into males (Figure 3). Environmental variables, local demography, and mortality schedules have been linked to driving/facilitating sex change in this reproductive strategy [34]. In this section, I’ll discuss how groupers displaying protogynous hermaphroditism, in conjunction with other life history characteristics, are more conducive to being overexploited.
It has long been understood that reproductive output (e.g., egg production) increases disproportionately with size/age in fish populations [35,36]. Consequently, fishing-induced size/age truncation has been shown to reduce stock resilience [37]. As such, significant management efforts are directed at conserving larger/older fish (e.g., using slot limits) to maintain sustainable spawning stock biomass levels. Protogynous hermaphrodites display dome-shaped egg production, meaning the oldest females show a reduction in egg production as energy is reallocated into sexual transition [38]. Consequently, protogynous hermaphrodites violate the hypothesis of the largest/oldest females having the highest contributions to reproductive output. Instead, size/age truncation in protogynous hermaphrodites severely depletes male sex ratios in naturally female-skewed populations, creating a unique situation of potential sperm limitation.
Decadal-scale declines in the male sex ratios of two SEUS protogynous hermaphrodites (Gag and Scamp M. phenax) have been documented [47,72]. Notably, both groupers display varying degrees of spawning aggregations, and this behavior in conjunction with increasing fishing pressure has been linked to the declines in male sex ratios [72]. Interestingly, another protogynous hermaphroditic grouper with significant fishing pressure (Red Grouper) has shown comparatively lighter declines in the male sex ratio, and this has been linked to its non-aggregating reproductive behavior [72]. In addition, a recent study observed low male gonadosomatic indices and milt reserves in Gag, suggesting they could actually be pair spawners, which would limit fertilization rates at low male sex ratios [47]. Whether this is a trend in other protogynous hermaphroditic groupers across the SEUS remains to be explored.
In response to the reduced GOM Gag population fitness, the Gulf of Mexico Fishery Management Council created two seasonal MPAs in 2000 (Madison-Swanson and Steamboat Lumps) to increase stock size, followed by a third in 2009 (The Edges). Almost 25 years later, the MPAs’ primary goal remains unattained. A recent study concluded that GOM Gag male sex ratio is at a historic low (5% in MPAs and 0% outside of protected areas) [47]. In addition, this study also reported increases in age at 50% transition to male, which accompanied by the reduced male sex ratio, suggests the male GOM Gag population is aging while experiencing limited recruitment [47]. Gag sexual transition does not require male presence, thus female groups traveling to shelf break spawning sites can include newly recruited males [47]. Consequently, fishing pressure inshore/mid-shelf can simultaneously remove females during peak egg production and bottleneck male recruitment. This case study highlights that MPA success can depend on its spatial extent, species’ life history, reproductive strategy, movement ecology, and surrounding context (e.g., fishery behavior and larval dispersal). It is important to note that measuring MPA effectiveness requires doing so at appropriate time frames [73], and success will be affected by enforcement and compliance [74].
Across the world, quantitative assessments are used to assess fish stocks and determine sustainable catch levels. A primary goal of these assessments is to determine reference points, such as MSY, fishing mortality at MSY (FMSY), and stock size at MSY (BMSY). These require knowledge of stock productivity, which is notoriously challenging to measure [75]. Stock-recruit relationships can be used to estimate BMSY. However, there tends to be a lack of reliable data to estimate steepness, the parameter that controls the shape of the stock-recruit relationship and has a strong influence on stock productivity [76,77]. Due to the uncertainty in reference points derived from the stock-recruit relationship, reference point proxies, such as quantities of the spawning potential ratio (SPR; the ratio of the fished stock size compared to unfished conditions), are used [75]. In the SEUS, a 30% SPR value is the most commonly used MSY proxy to derive reference points and is the current regulation for most GOM reef fishes [78]. However, a recent simulation study indicates that FMSY proxies of 40% and 50% SPRs are most probable in achieving long-term MSY for gonochoristic and hermaphroditic stocks, respectively [75]. Due to their complex life histories, protogynous hermaphrodites undoubtedly require conservative management approaches.
Recent efforts have gone into considering MSY proxy values >30% SPR for hermaphroditic grouper stocks across the SEUS. The most recent SA Scamp stock assessment, in which the stock was found to be overfished, used an FMSY proxy of 40% SPR [58]. While 30% SPR continues to be the status quo across the region, management is also considering assessment results under higher SPR values by including them in assessment terms of reference. For example, the latest GOM Yellowedge Grouper H. flavolimbatus assessment used an MSY proxy of 30% SPR, and while the stock was deemed not overfished nor experiencing overfishing, the terminal year displayed the lowest spawning stock biomass value across the assessed period [79]. Additional projections using an MSY proxy of 40% SPR resulted in Yellowedge Grouper experiencing multiple years of overfishing, and 50% SPR resulted in the stock experiencing overfishing during the terminal year [80]. While there is no “one size fits all” answer to MSY proxy values, management should consider using SPR values ≥40% for hermaphroditic stocks and completely abandon considering 30%.
Protogynous hermaphrodites provide unique case studies where traditional fisheries management techniques can fall short of providing reliable estimates of stock status. Maintaining female-specific spawning stock biomass is of primary interest when managing fish stocks. However, careful consideration must be given to male-specific spawning stock biomass when assessing protogynous hermaphrodites, as is the case with many groupers across the SEUS [81]. Many of these groupers are showing historic declines in male sex ratios, which can result in sperm limitation and stock collapse. Efforts are underway to rebuild these stocks by using MPAs and considering conservative MSY proxies in stock assessments. However, these stocks remain among the most challenging to manage and rebuild. Many SEUS hermaphroditic grouper stocks have experienced historic population declines (Figure 4). Beyond groupers, many reef fishes that support fisheries display protogynous hermaphroditism (e.g., Sparidae and Labridae). Thus, conservative management strategies should be considered for these species while balancing socioeconomic benefits.
3.3. Deepwater Groupers
A significant proportion of SEUS groupers can be considered deepwater species occupying habitats at depths >100m. In the GOM and SA, Yellowedge Grouper and Snowy Grouper are the most harvested deepwater groupers (Figure 5). Deepwater groupers are especially susceptible to overexploitation as many of them show comparatively higher longevities, which are associated with slower growth, maturity, and transition rates [59,60,62,66,71]. Consequently, their lower natural mortality rates suggest comparatively lower productivity and fishing pressure can quickly cause severe population declines [82]. Additionally, most of these groupers have been confirmed to display protogynous hermaphroditism [60,63,67]. While little is known about their movement ecology, available information suggests many deepwater groupers display high site fidelity and disproportionate densities of individuals can be observed over small spatial footprints [83].
Like the Red Grouper and Gag fisheries, The SEUS deepwater grouper fisheries have historically been dominated by the commercial sector but are thought to have started during the late 1900s [5,79]. Due to their similarity in habitat use, four species are managed under a combined deepwater grouper stock in both the GOM and SA (Yellowedge Grouper, Snowy Grouper, Speckled Hind E. drummondhayi, and Warsaw Grouper H. nigritus). Consequently, species-specific landings can vary from year to year without causing any management concerns as long as the stock annual catch limits are not exceeded (Figure 6). A recent assessment of the GOM Yellowedge Grouper, the most landed species in the GOM commercial deepwater grouper fishery (Figure 6), found the species not to be overfished nor experiencing overfishing, but to have the lowest observed spawning stock biomass during the terminal year [79]. Additionally, stakeholder feedback reporting population declines in GOM Yellowedge Grouper suggests it could be due to increased fishing power (e.g., mapping and electric reels) and recreational effort [84]. This leads to speculate if both of these variables in conjunction with increased harvest restrictions in comparatively “shallower water” reef fishes (e.g., Red Snapper, Greater Amberjack Seriola dumerili, Gray Triggerfish Balistes capriscus, and Gag) could be leading to increased deepwater grouper recreational landings and effort. As recreational allocations continue to increase in other groupers (e.g., Red Grouper and Gag), should we expect the same patterns for the deepwater grouper complex? Commercial grouper landings are more accurate and precise, leading to smaller buffers between annual catch limits and annual catch targets than the recreational sector [32]. Thus, increases in the recreational deepwater grouper landings would likely be associated with increased uncertainty.
As reductions in the GOM Yellowedge Grouper population are evident, how possible is it to observe increased landings in other deepwater groupers, and to what scale? Many of these species have poorly understood life histories, highly uncertain recreational landings, and display decadal-scale overexploitation. Misty Grouper H. mystacinus and Marbled Grouper are two species distributed across the SEUS for which their population trends are unknown and landings data are scarce. The SA Speckled Hind stock has shown signs of overexploitation since the 1970s, which was followed by reduced bag limits in 1994 [63]. However, evidence suggests that the stock continued to be overexploited through the 2000s [63]. Additionally, a recent study reported low population connectivity, limited movement, and high numbers of harvested young fish in the GOM Snowy Grouper stock [85]. In attempts to help rebuild deepwater reef fishes (including groupers), the South Atlantic Fishery Management Council implemented eight deepwater MPAs in 2009 [86]. To date, the investigations conducted demonstrate no evidence of the MPAs improving the status of grouper stocks [87,88]. In fact, one of these studies found that community composition within the largest of these MPAs has shifted away from groupers to other reef fishes [88]. Thus, long-term monitoring of MPAs will be important to account for long generation times observed in these long-lived species and to investigate how different fishes respond to the MPA effect.
4. Recovery of Historically Exploited Stocks?
4.1. Nassau Grouper and Red Hind
During the 1900s, Nassau Grouper was considered the most important commercial grouper across the Caribbean [89]. Due to their aggregating behavior, where 1000s of individuals can be observed at a time, the spawning stock suffered severe fishing mortality, resulting in dramatic population declines [89]. By the 1970s, there was evidence of reductions in the density of individuals at aggregation sites and the number of aggregation sites, followed by reduced landings [89]. To facilitate recovery across the CAB, the Caribbean Fishery Management Council implemented a moratorium on Nassau Grouper in 1990. The species was also listed as threatened under the U.S. Endangered Species Act in 2016 [90,91].
In the decades following the moratorium, population trends of CAB Nassau Grouper are poorly understood, but much effort has gone into investigating population dynamics. A genetic study reported evidence of genetic differentiation in Nassau Grouper subpopulations across the Caribbean region, suggesting subpopulations’ spawning aggregations may be responsible for self-recruitment [92]. Juvenile Nassau Grouper habitat use has been described across the CAB [93,94,95], which can assist in the designation of critical habitats to maximize juvenile recruitment and survival. While current population trends of CAB Nassau Grouper are not well described, subpopulations in other Caribbean jurisdictions are showing signs of recovery. For example, Nassau Grouper spawning aggregation sites in the Cayman Islands have displayed positive responses to >15 years of conservation efforts [13]. Unfortunately, Nassau Grouper has been reported in Puerto Rico recreational landings after the commonwealth’s local moratorium, implemented in 2004 [96]. Thus, population recovery efforts must include stakeholder involvement to increase compliance and provide local ecological knowledge [97]. Designation under the U.S. Endangered Species Act provides funds to conduct applied research into the species, which can facilitate efforts to conduct an updated population assessment of the CAB Nassau Grouper subpopulation.
Population assessments indicated that the CAB Red Hind was experiencing overexploitation during the 1980s [98,99]. An assessment in 2000 concluded that the Puerto Rico subpopulation continued to experience overexploitation throughout the 1990s [100]. Similar declines were observed in a neighboring jurisdiction [101]. Monitoring efforts during the 30 years following the implementation of the Red Hind Bank Marine Conservation District concluded that the MPA has successfully recovered its Red Hind spawning aggregation [19,102]. While current CAB-wide Red Hind population trends are unknown, a subpopulation of the species is scheduled to undergo an assessment in 2025 (https://sedarweb.org/).
CAB Nassau Grouper and Red Hind populations have suffered historic overexploitation. Management efforts, which have included the use of MPAs, have demonstrated potential in rebuilding these stocks. Telemetry studies report that both these species undergo dynamic sex-specific movements between protected and non-protected areas during the spawning season [103,104]. Additionally, the Red Hind population recovery rate increased when the Red Hind Bank Marine Conservation District transitioned from seasonal to year-round protection [19]. Thus, extending spatial and temporal MPA closures should be considered to maximize the protection of these and other vulnerable aggregating species [11,104].
4.2. Goliath Grouper
The Atlantic Goliath Grouper E. itajara (Goliath Grouper hereafter) is the largest grouper distributed across the Atlantic basin [105]. Interestingly, it was recently confirmed that Goliath Grouper differs from most other economically relevant groupers in the SEUS by displaying gonochorism (single-sex individuals) with the potential for diandric protogyny, where males can arise from birth or sexual transition [40]. The exploitation of this species in the SEUS can be traced back to the early 1900s (Figure 7), and population declines were evident around the 1980s [106]. In other jurisdictions, it is hypothesized that Goliath Grouper populations are currently experiencing overexploitation and/or apparent population declines [107,108].
In response to the population declines across the SEUS, NMFS implemented a moratorium across its jurisdiction in 1990. Soon following the moratorium, increases in the population were observed through the following decades [109,110,111]. This was followed by stakeholders expressing frustration reporting increases in Goliath Grouper abundance leading to reduced reef fish communities and increased depredation events (consuming fishers’ hooked fish) [112]. These angler perceptions reduced their satisfaction and led management to consider intervention. After years of consideration, the Florida Fish and Wildlife Conservation Commission opened a limited-access fishery for juvenile Goliath Grouper (https://myfwc.com/fishing/saltwater/recreational/goliath/).
While the juvenile Goliath Grouper fishery provides the state with direct revenue ($10 to apply and ≥$150 if selected) and biological samples, the decision to open the fishery remains controversial. Following the upward trend, the juvenile and adult populations have suffered episodic mortality events (e.g., Red Tide Karenia brevis blooms and cold snaps) [110]. In addition, the degradation of Red Mangrove Rhizophora mangle, an essential habitat for juvenile Goliath Grouper, has been reported and may lead to reduced recruitment and hindered population recovery efforts [110]. While assessments have been performed on Goliath Grouper, poor catch records have led to high uncertainty and rejection by peer reviewers [110]. Thus, as the Goliath Grouper population faces other challenges (e.g., episodic mortality events and habitat loss), what effects will the fishery add? Will there be reduced recruitment into an already distressed adult population? The discussion of opening a Goliath Grouper fishery is beyond the scope of this review, but for further dialogue see the following references [110,112,113].
5. Climate Change and Anthropogenically Driven Disturbances
Across the globe, climate change is modifying many mechanisms relating to atmospheric, sea surface, physiochemical, dynamic, seasonal, and regional processes [114]. Investigating the effects of these changes on fish stocks is of primary interest as many have been found to affect fish population dynamics [115,116,117,118,119]. As for groupers, primary processes of concern include oceanographic variables, specifically temperature, and currents. Projected warming is hypothesized to drive changes in maturity and gamete release schedules, as well as transition schedules in sequential hermaphrodites [120,121,122]. Under current climate change projections, grouper spawning dynamics in the southern Gulf of Mexico are expected to be affected in the next 50 years [120]. Changes in current dynamics are also thought to affect larval transport in groupers, which have long pelagic larval duration stages [1,123]. Changes in currents during larval stages may affect where these settle, which can severely affect survival and subsequent recruitment [1]. As many groupers across the SEUS have suffered historic population declines, climate change shall be an important consideration in rebuilding efforts.
Environmental changes have been linked to regime shifts (temporal changes in average recruitment) and recruitment failure (successive poor recruitment events) in fish stocks [115,124]. A recent study found that in many assessed fish stocks, recruitment is more influenced by environmental variables than spawning stock biomass [115]. This same study found that a large proportion of assessed stocks, including GOM Red Grouper, have undergone regime shifts that are often not detected by traditional stock-recruit relationship modeling, and not doing so can lead to biased projections [115]. In the SA, many reef fish stocks are showing signs of recruitment failure [125,126,127]. In addition, there is evidence of a correlation in recruitment failure between these species, suggesting that a common exogenous driver is causing poor recruitment events [127]. Among the species showing signs of recruitment failure in the SA are groupers displaying protogynous hermaphroditism, including Red Grouper, Gag, Scamp, and Snowy Grouper [125,127].
The importance of maintaining healthy habitats for productive fisheries has long been recognized and investigated [128,129,130,131,132]. Many groupers display ontogenetic habitat shifts demonstrated by spending juvenile stages in shallow, inshore habitats followed by deeper, offshore habitats to join the adult stock [133,134,135,136]. Primary habitats associated with early life history stages of groupers are Red Mangrove shorelines and seagrass meadows [135,136,137]. These two coastal habitats are among those facing severe impacts due to climate change (e.g., sea level rise and tropicalization) and anthropogenic activities (e.g., nutrient loading) [138,139]. To provide a relevant example, declines in seagrass beds have been reported in the central west and panhandle regions of Florida [140,141]. These areas are among the core distribution of GOM Gag, which is currently overfished and experiencing overfishing [48]. Therefore, how will declines in seagrass cover impact Gag recruitment? Additionally, juvenile Goliath Grouper exhibit diel movements between seagrass beds and Red Mangroves [142]. How will Goliath Grouper fare against its essential habitats facing climate- and anthropogenically-driven changes?
Anthropogenic activities have been linked to impacting coastal ecosystems and fisheries. Among the most relevant in the SEUS is eutrophication, nutrient loading which can lead to phytoplankton blooms followed by hypoxic events. In the GOM, Red Tide blooms have been linked to episodic fish kills [143]. While these are naturally occurring, their intensity and duration have increased and have been credited to eutrophication [144]. While it’s difficult to quantify the population effects caused by these episodic events, Red Grouper has served as a model species to describe yearly mortality rates induced on a fish stock by Red Tide blooms [145,146].
Other anthropogenic activities across the region that can affect fish stocks are pipelines, seismic air guns, and energy exploration [1]. The Deepwater Horizon oil spill (2010) is attributed with causing damages on a variety of taxa and habitats across the GOM [147,148,149]. An ecosystem model simulating Deepwater Horizon effects found the spill reduced the condition of groupers [147]. Effects caused by the oil spill were considered for the most recent GOM Yellowedge Grouper assessment, but there was no strong evidence to justify including a mortality rate for the event [79]. Lastly, wind energy development in the GOM has recently been announced (https://www.fisheries.noaa.gov/feature-story/noaa-and-bureau-ocean-energy-management-sign-new-interagency-agreement-wind-energy). The effects of these activities on fisheries have been a recent topic of interest across other regions [150,151,152], and shall be a new line of investigation in the GOM.
Climate change and anthropogenic activities can severely impact fish population dynamics and hinder recovery efforts if left unaccounted for. Groupers are at the forefront of species of concern, as many display complex movement ecology and reproductive dynamics. In addition, their high level of exploitation may affect their sensitivity to upcoming environmental changes [120]. Thus, the effects of climate change and anthropogenic activities should be accounted for when monitoring and managing grouper fisheries.
6. Ecosystem Based Considerations in Grouper Assessments
Across the globe, there is a push to abandon single-species management for ecosystem-based fisheries management (EBFM; also known as ecosystem approach to fisheries), a holistic approach that accounts for biotic, abiotic, and human components of ecosystems and their interactions and applies an integrated approach to fisheries within ecologically meaningful boundaries [153,154,155]. In 2016, NMFS released its EBFM policy, in which it declared that EBFM was the preferred way to meet the mandates of sustainably managing the nation’s living marine resources [156].
Across the SEUS, efforts to facilitate moving towards EBFM are well underway [157,158,159,160,161]. Groupers have played an important role as case studies to initiate advancing single-species management towards EBFM. In the GOM, the most recent Red Grouper and Gag assessments included an age-specific Red Tide mortality rate for years where severe mortality was evident [48,52]. These mortality rates were estimated from ecosystem models that included many fishing fleets and functional groups as well as environmental variables [162,163]. Although not included in the assessment, the effects of the Deepwater Horizon oil spill were considered in the latest GOM Yellowedge Grouper assessment [79]. While the majority of fish stocks across the SEUS continue to be managed (and assessed) as single stocks, the inclusion of environmental effects moves them closer to EBFM in the model complexity continuum [155], and gets NMFS closer to reaching its EBFM goals.
7. Using Co-Produced Data to Assess Grouper Stocks
Stakeholder involvement is a necessary component for effective and transparent fisheries management as it provides many benefits [164,165,166,167,168]. In the SEUS, stakeholder involvement can be observed at different stages of the management process for grouper fisheries. The Great Goliath Grouper Count was developed to address the data-poor nature of the Goliath Grouper (lacking reliable landings and age/size compositions) by facilitating cost-effective and timely stakeholder visual surveys that inform trends in abundance and size distribution [169]. These data were incorporated into the most recent Goliath Grouper assessment [170], therefore including stakeholders in multiple stages of the management process.
The Gulf of Mexico Fishery Management Council’s Fishermen Feedback Tool is used to gather stakeholder perceptions on the status of stocks, which has been used to gather information on the status of Gag, Red Grouper, Scamp, and Yellowedge Grouper (https://gulfcouncil.org/blog/2022/fishy-or-not-we-want-your-feedback/). While the data collected are not directly included in stock assessments, this provides the managers with local ecological knowledge, which could provide trends not captured by the assessments. In addition, The South Atlantic Fishery Management Council’s Citizen Science Program provides research priorities (which currently include grouper sampling) for stakeholders to become involved in the data collection process (https://safmc.net/citizen-science/).
Stakeholder involvement is a promising avenue for optimizing cost-effective data collection efforts and facilitating stakeholder sense of inclusion in the management process. It is highly recommended by researchers to improve our understanding of spawning aggregations across the SEUS [171,172]. In the Cayman Islands, stakeholder involvement has been a monumental part of the recovery of the Nassau Grouper population [13]. Thus, lessons from this effort could be applied to improve the monitoring and rebuilding efforts of groupers across the SEUS.
8. Emerging Techniques to Monitor Grouper Populations
In recent years, there’s been many developments in techniques to improve the management and monitoring of groupers across the SEUS. A promising one for management purposes is management strategy evaluation, in which simulations are used to evaluate management strategies, their associated trade-offs, and uncertainty in achieving management goals [173]. This technique was recently employed in two SA fish stocks to evaluate how different management procedures meet the specific objectives of the recreational and commercial sectors [174]. Currently, the South Atlantic Fishery Management Council is using management strategy evaluation to evaluate procedures in the snapper/grouper fishery (https://safmc-mse.netlify.app/). Results from this effort shall provide important information on how to best meet management objectives in complex multispecies reef fish fisheries that could be applied to the GOM and CAB.
There have also been many improvements to monitor grouper populations. Among these are age validation techniques involving bomb radiocarbon, which have been employed to validate the estimated ages and longevities of multiple groupers in the SEUS [59,66,175,176]. Additionally, recent advances in epigenetic aging, which were validated for Red Grouper, could provide accurate, timely, and non-lethal mass aging of fishes [177,178]. These advances shall improve the quantity and quality of assessment inputs, therefore decreasing the assessment uncertainty and providing improved catch advice.
Autonomous systems and passive acoustics have been employed to monitor grouper populations. In the CAB, these techniques are used to investigate grouper spawning-related behaviors [179,180,181,182]. These technologies have potentially identified undiscovered grouper spawning areas [180,182], which can assist in directing future management efforts to improve Nassau Grouper and Red Hind recovery efforts. Acoustic telemetry has been employed to investigate movement ecology, spawning dynamics, and post-release mortality of groupers in the SEUS [26,104,183,184,185]. Geochemistry and isotope studies have also provided important information on grouper ontogenetic changes in diet and habitat use, as well as delineating population structure [85,133,186,187]. The utility of environmental DNA for investigating grouper spawning aggregations in the CAB has been recently explored, and while limited success was reported, this remains a powerful tool that could inform population trends of species with depleted population levels, such as Nassau Grouper [188].
Video surveys have become one of the most common methods to monitor reef fish populations across the SEUS [189,190]. These are used to generate fishery-independent relative abundance indices that are directly used in the stock assessments to inform the model on the dynamics of adult populations [48,58]. In addition to providing species composition and abundance, video surveys can also provide fish measurements to inform size composition [191]. A long-term video survey in the GOM has provided descriptions of Marbled Grouper habitat use, a species poorly understood across its range [192]. Additionally, video surveys conducted across the Caribbean reported a depth range expansion of Misty Grouper, another poorly understood deepwater species [193].
9. Conclusions and Future Directions
Groupers have historically supported valuable fisheries across their range, and this continues to be the case. Multiple components make grouper fisheries among the most difficult to manage in the SEUS. First, the dynamics of the multispecies reef fish fisheries which they are a part of make it extremely difficult for traditional management techniques to maintain sustainable fisheries. Being composed of species with varying life histories and temporal harvest closures, multispecies reef fish fisheries are conducive to high levels of regulatory discards, which can induce severe mortality on fish stocks and undermine the purpose of temporal harvest closures and minimum size limits. Thus, recreational effort restriction should be considered by management while accounting for socioeconomic components.
In addition to being part of complicated fisheries, groupers have complex life histories, demonstrated by protogynous hermaphroditism, high longevities, slow growth, maturity and transition, ontogenetic habitat shifts, as well as spawning-related migrations and aggregations. These make groupers especially vulnerable to overexploitation, especially the poorly understood deepwater stocks with high longevities that will presumably face additional fishing pressure as increases in shallow-water reef fish regulations and recreational effort are observed. In the case studies discussed, there is substantial evidence of declines in protogynous hermaphrodites, demonstrated by decreasing male sex ratios and limited recruitment. There was also evidence of rebuilding stocks by using harvest moratoriums and MPAs. In the GOM and SA, the expansion of MPAs could have benefits but may be hindered by the ever-growing recreational fleet. Thus, long-term monitoring will be essential to evaluate MPA effects. As for the CAB, MPAs have shown signs of success, however, MPA effectiveness may be dependent on the surrounding context [194]. Given there is evidence of both self-sustaining and dependent grouper subpopulations across the Gulf of Mexico and the Caribbean regions [92,100,195], an international collaboration to implement effective conservation plans should be considered. Additionally, conservative approaches should be considered when assessing grouper stocks and determining their reference points.
Climate change and anthropogenic impacts are expected to take their toll on grouper stocks. Particularly, changing oceanographic conditions are expected to alter the reproductive phenology and larval survival of groupers. Additionally, declines in nursery habitats are expected to affect juvenile condition and recruitment into adult populations. Some grouper stocks have served as models to transition from single-species management towards EBFM. Efforts shall continue to progress by exploring ecosystem models, as currently done to estimate grouper red tide mortality rates, to identify interactions that can be appropriately modeled under current stock assessment platforms (e.g., environmental effects on recruitment). As regime shifts and poor recruitment for some grouper stocks are evident, assessment techniques should account for exogenous mechanisms driving these.
Many novel tools are available to facilitate the management and monitoring of grouper stocks. Management strategy evaluation could prove to be useful for identifying the most appropriate management procedures across the SEUS and beyond. The progression of epigenetic aging could facilitate cost-effective and timely mass aging of fishes. Acoustic telemetry continues to provide novel insights into movement ecology and post-release behaviors of groupers. Autonomous systems and passive acoustics provide non-invasive methods of monitoring grouper spawning behavior and have been useful for identifying unprotected potential spawning areas. Lastly, video surveys have become one of the most effective ways of describing species composition, abundance, and size composition.
While this review focuses on synthesizing available information from the SEUS, the case studies and future directions covered can be applied beyond this region. In many countries, there are data-limited scenarios that make the use of many management and monitoring techniques covered here unlikely. However, several lessons can be applied almost universally, such as the protection of spawning aggregations and involving stakeholders in the management and monitoring process. Information provided on SEUS groupers can facilitate the production of hypotheses about groupers in other regions that are poorly understood. Given the relatively rich history of scientific studies and management of groupers in the SEUS, this synthesis can assist other jurisdictions in sustainably managing their valuable grouper fisheries for generations to come.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The stock assessments from which Figure 1 was derived are publicly available at https://sedarweb.org/sedar-assessments/. Recreational effort data are publicly available at https://www.fisheries.noaa.gov/data-tools/recreational-fisheries-statistics-queries. Commercial grouper landings are publicly available at https://noaa-sero.s3.amazonaws.com/drop-files/cs/2022_GT_AnnualReport_Final.pdf.
Acknowledgments
I would like to extend my profound gratitude to all the scientists who have pushed forward our understanding of grouper biology, ecology, and fisheries management across the SEUS and beyond.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Coleman, F.C.; Koenig, C.C. The Effects of Fishing, Climate Change, and Other Anthropogenic Disturbances on Red Grouper and Other Reef Fishes in the Gulf of Mexico. Integr Comp Biol 2010, 50, 201–212. [Google Scholar] [CrossRef]
- Ellis, R.D. Red Grouper (Epinephelus Morio) Shape Faunal Communities via Multiple Ecological Pathways. Diversity 2019, 11, 89. [Google Scholar] [CrossRef]
- Sadovy de Mitcheson, Y.; Craig, M.T.; Bertoncini, A.A.; Carpenter, K.E.; Cheung, W.W.L.; Choat, J.H.; Cornish, A.S.; Fennessy, S.T.; Ferreira, B.P.; Heemstra, P.C.; et al. Fishing Groupers towards Extinction: A Global Assessment of Threats and Extinction Risks in a Billion Dollar Fishery. Fish and Fisheries 2013, 14, 119–136. [Google Scholar] [CrossRef]
- Ward, C.H. Habitats and Biota of the Gulf of Mexico: Before the Deepwater Horizon Oil Spill. Habitats and Biota of the Gulf of Mexico: Before the Deepwater Horizon Oil Spill 2017, 2, 1–1757. [Google Scholar] [CrossRef]
- Goodyear, C.P.; Schirripa, M.J. The Red Grouper Fishery of the Gulf of Mexico. Southeast Fisheries Center. Miami Laboratory Contribution No. MIA-92/93-75.
- Chester, A.J.; Huntsman, G.R.; Tester, P.A.; Manooch III, C.S. South Atlantic Bight Reef Fish Communities as Represented in Hook-and-Line Catches. Bull Mar Sci 1984, 34, 267–279. [Google Scholar]
- Simmons, C.M.; Collins, A.B.; Ruzicka, R. Chapter 3: Distribution and Diversity of Coral Habitat, Fishes, and Associated Fisheries in U.S. Waters of the Gulf of Mexico. Interrelationships between corals and fisheries, ed. Bortone, Stephen A. CRC Press, Boca Raton, FL, USA. 2014.
- Sledge, J.H. The Gulf of Mexico: A Maritime History. The University of South Carolina Press. 280pp 2019.
- Saul, S. SEDAR12-DW-11 Quantitative Historical Analysis of the United States and Cuban Gulf of Mexico Red Grouper Commercial Fishery. SEDAR. available at . Available online: https://sedarweb.org/documents/s12dw11-quantitative-historical-analysis-of-the-united-states-and-cuban-gulf-of-mexico-red-grouper-commercial-fishery/2006.
- Sadovy, Y.; Flguerola, M.; Roman, A. Age and Growth of Red Hind Epinephelus Guttatus in Puerto Rico and St. Thomas. Fishery Bulletin 1992, 90, 516–528. [Google Scholar]
- Tuohy, E.; Schärer-Umpierre, M.; Penrod, L.; Appeldoorn, R. Spatial and Temporal Dynamics of a Nassau Grouper Fish Spawning Aggregation Located on an Isolated Seamount in Puerto Rico. Aquat Conserv 2023, 33, 1116–1130. [Google Scholar] [CrossRef]
- Sadovy, Y.; Eklund, A.-M. Synopsis of Biological Data on the Nassau Grouper, Epinephelus Striatus (Bloch, 1792), and the Jewfish, E. Itajara (Lichtenstein, 1822). NOAA Technical Report NMFS 146. Available online: https://repository.library.noaa.gov/view/noaa/30901999.
- Waterhouse, L.; Heppell, S.A.; Pattengill-Semmens, C. V; Mccoy, C.; Bush, P.; Johnson, B.C.; Semmens, B.X. Recovery of Critically Endangered Nassau Grouper (Epinephelus Striatus) in the Cayman Islands Following Targeted Conservation Actions. PNAS 2020, 117, 1587–1595. [CrossRef]
- CFMC Fishery Management Plan, Final Environmental Impact Statement, and Draft Regulatory Impact Review, for the Shallow-Water Reeffish Fishery of Puerto Rico and the U. S. Virgin Islands. Caribbean Fishery Management Council. available at . Available online: https://caribbeanfmc.com/fishery-management/fishery-management-plans1985.
- NOAA Magnuson-Stevens Fishery Conservation and Management Act. NOAA. available at 2007, 1–178. Available online: https://www.fisheries.noaa.gov/topic/laws-policies.
- SAFMC Fishery Management Plan, Regulatory Impact Review, and Final Environmental Impact Statement for the Snapper-Grouper Fishery of the South Atlantic Region. South Atlantic Fishery Management Council. available at https://safmc.net/documents/snapper-grouper-fishery-management-plan/ 1983.
- Caillouet Jr., C. W.; Hart, R.A.; Nance, J.M. Growth Overfishing in the Brown Shrimp Fishery of Texas, Louisiana, and Adjoining Gulf of Mexico EEZ. Fish Res 2008, 92, 289–302. [Google Scholar] [CrossRef]
- GMFMC Environmental Impact Statement and Fishery Management Plan for the Reef Fish Resources of the Gulf of Mexico. Gulf of Mexico Fishery Management Council. available at https://repository.library.noaa.gov/view/noaa/19629 1981.
- Rosemond, R.C.; Nemeth, R.S.; Heppell, S.A. Demographic Recovery of a Reef Fish Population over 30 Years of Spawning Aggregation Site Protection. Front Mar Sci 2022, 9, 931409. [Google Scholar] [CrossRef]
- Whiteman, E.A.; Jennings, C.A.; Nemeth, R.S. Sex Structure and Potential Female Fecundity in a Epinephelus Guttatus Spawning Aggregation: Applying Ultrasonic Imaging. J Fish Biol 2005, 66, 983–995. [Google Scholar] [CrossRef]
- Coggins Jr, L.G.; Catalano, M.J.; Allen, M.S.; Pine III, W.E.; Walters, C.J. Effects of Cryptic Mortality and the Hidden Costs of Using Length Limits in Fishery Management. Fish and Fisheries 2007, 8, 196–210. [Google Scholar] [CrossRef]
- Tetzlaff, J.C.; Pine III, W.E.; Allen, M.S.; Ahrens, R.N.M. Effectiveness of Size Limits and Bag Limits for Managing Recreational Fisheries: A Case Study of the Gulf of Mexico Recreational Gag Fishery. Bull Mar Sci 2013, 89, 483–502. [Google Scholar] [CrossRef]
- Pulver, J.R. Sink or Swim? Factors Affecting Immediate Discard Mortality for the Gulf of Mexico Commercial Reef Fish Fishery. Fish Res 2017, 188, 166–172. [Google Scholar] [CrossRef]
- Runde, B.J.; Buckel, J.A. Descender Devices Are Promising Tools for Increasing Survival in Deepwater Groupers. Marine and Coastal Fisheries 2018, 10, 100–117. [Google Scholar] [CrossRef]
- Stallings, C.D.; Ayala, O.; Cross, T.A.; Sauls, B. Post-Release Survival of Red Snapper (Lutjanus Campechanus) and Red Grouper (Epinephelus Morio) Using Different Barotrauma Mitigation Methods. Fish Res 2023, 264, 106717. [Google Scholar] [CrossRef]
- Runde, B.J.; Michelot, T.; Bacheler, N.M.; Shertzer, K.W.; Buckel, J.A. Assigning Fates in Telemetry Studies Using Hidden Markov Models: An Application to Deepwater Groupers Released with Descender Devices. N Am J Fish Manag 2020, 40, 1417–1434. [Google Scholar] [CrossRef]
- NOAA Fisheries of the Caribbean, Gulf of Mexico, and South Atlantic; Snapper-Grouper Fishery off the Southern Atlantic Region; Regulatory Amendment 29. NOAA. available at https://www.federalregister.gov/documents/2020/06/15/2020-11916/fisheries-of-the-caribbean-gulf-of-mexico-and-south-atlantic-snapper-grouper-fishery-off-the 2020.
- NOAA Fisheries of the Caribbean, Gulf of Mexico, and South Atlantic; Reef Fish Resources of the Gulf of Mexico; Requirement for a Descending Device or Venting Tool. NOAA. available at https://www.federalregister.gov/documents/2022/01/14/2022-00720/fisheries-of-the-caribbean-gulf-of-mexico-and-south-atlantic-reef-fish-resources-of-the-gulf-of 2022.
- Coleman, F.C.; Figueira, W.F.; Ueland, J.S.; Crowder, L.B. The Impact of United States Recreational Fisheries on Marine Fish Populations. Science (1979) 2004, 305, 1958–1960. [Google Scholar] [CrossRef]
- Shertzer, K.; Crosson, S.; Williams, E.; Cao, J.; DeVictor, R.; Dumas, C.; Nesslage, G. Fishery Management Strategies for Red Snapper in the Southeastern U.S. Atlantic: A Spatial Population Model to Compare Approaches. N Am J Fish Manag 2024, 44, 113–131. [Google Scholar] [CrossRef]
- NOAA Fisheries of the Caribbean, Gulf of Mexico, and South Atlantic; Reef Fish Fishery of the Gulf of Mexico; Amendment 53. NOAA. available at https://www.federalregister.gov/documents/2021/12/09/2021-26504/fisheries-of-the-caribbean-gulf-of-mexico-and-south-atlantic-reef-fish-fishery-of-the-gulf-of-mexico 2021.
- NOAA Fisheries of the Caribbean, Gulf of Mexico, and South Atlantic; Reef Fish Resources of the Gulf of Mexico; Amendment 56. NOAA. available at https://www.federalregister.gov/documents/2024/05/10/2024-10208/fisheries-of-the-caribbean-gulf-of-mexico-and-south-atlantic-reef-fish-resources-of-the-gulf-of 2024.
- Murua, H. Fish Reproduction Assortment: A Wonderful Diversity. Environ Biol Fishes 2014, 97, 329–333. [Google Scholar] [CrossRef]
- Warner, R.R. Sex Change and the Size-Advantage Model. Trends Ecol Evol 1988, 3, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Beverton, R.J.H.; Holt, S.J. On the Dynamics of Exploited Fish Populations. Springer Dordrecht 1983. [Google Scholar] [CrossRef]
- Barneche, D.R.; Robertson, D.R.; White, C.R.; Marshall, D.J. Fish Reproductive-Energy Output Increases Disproportionately with Body Size. Science (1979) 2018, 360, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Hixon, M.A.; Johnson, D.W.; Sogard, S.M. BOFFFFs: On the Importance of Conserving Old-Growth Age Structure in Fishery Populations. ICES Journal of Marine Science 2014, 71, 2171–2185. [Google Scholar] [CrossRef]
- Gamboa-Salazar, K.R.; Wyanski, D.M.; Bubley, W.J.; Klibansky, N. Effects of Age and Size on Spawning and Egg Production in Gag and Scamp Grouper off the Southeastern United States. ICES Journal of Marine Science 2020, 77, 290–299. [Google Scholar] [CrossRef]
- Bullock, L.H.; Murphy, M.D.; Godcharles, M.F.; Mitchell, M.E. Age, Growth, and Reproduction of Jewfish Epinephelus Itajara in the Eastern Gulf of Mexico. Fishery Bulletin 1992, 90, 243–249. [Google Scholar]
- Murie, D.J.; Parkyn, D.C.; Koenig, C.C.; Coleman, F.C.; Malinowski, C.R.; Cusick, J.A.; Ellis, R.D. Age, Growth, and Functional Gonochorism with a Twist of Diandric Protogyny in Goliath Grouper from the Atlantic Coast of Florida. Fishes 2023, 8. [Google Scholar] [CrossRef]
- NOAA Status of Stocks 2023 Annual Report to Congress on the Status of U.S. Fisheries. NOAA. available at https://www.fisheries.noaa.gov/s3/2024-04/2023SOS-final.pdf 2024.
- Crabtree, R.E.; Bullock, L.H. Age, Growth, and Reproduction of Black Grouper, Mycteroperca Bonaci, in Florida Waters. Fishery Bulletin 1998, 96, 735–753. [Google Scholar]
- Renán, X.; Brulé, T.; Galindo-Cortes, G.; Colás-Marrufo, T. Age-Based Life History of Three Groupers in the Southern Gulf of Mexico. J Fish Biol 2022, 101, 857–873. [Google Scholar] [CrossRef]
- Burton, M.L.; Potts, J.C.; Carr, D.R. Age, Growth and Natural Mortality of Coney (Cephalopholis Fulva) from the Southeastern United States. PeerJ 2015, 2015, e825. [Google Scholar] [CrossRef] [PubMed]
- Bortoletto, E.; Filho, G.; Ester, M.; Simoni, R.; Gomes, M.; Rêgo, D.O.; Lúcia, M.; De Araújo, G.; Guilherme, P.; De Oliveira, V.; et al. Reproductive Biology of Cephalopholis Fulva (Linneus, 1758) Caught in the North Coast of Pernambuco. Panam J Aquat Sci 2019, 14, 134–142. [Google Scholar]
- Harris, P.J.; Collins, M.R. Age, Growth and Age at Maturity of Gag, Mycteroperca Microlepis, from the Southeastern United States during 1994-1995. Bull Mar Sci 2000, 66, 105–117. [Google Scholar]
- Lowerre-Barbieri, S.; Menendez, H.; Bickford, J.; Switzer, T.S.; Barbieri, L.; Koenig, C. Testing Assumptions about Sex Change and Spatial Management in the Protogynous Gag Grouper, Mycteroperca Microlepis. Mar Ecol Prog Ser 2020, 639, 199–214. [Google Scholar] [CrossRef]
- SEDAR SEDAR 72 Gulf of Mexico Gag Grouper Final Stock Assessment Report. SEDAR, North Charleston SC. 319 pp. available online at: http://sedarweb.org/sedar-72 2021.
- SEDAR SEDAR 71 South Atlantic Gag Stock Assessment Report. SEDAR, North Charleston, SC. 164 pp. available online at: http://sedarweb.org/sedar-71 2021.
- Burton, M.L.; Potts, J.C.; Ostrowski, A.D.; Shertzer, K.W. Age, Growth, and Natural Mortality of Graysby, Cephalopholis Cruentata, from the Southeastern United States. Fishes 2019, 4, 36. [Google Scholar] [CrossRef]
- Luckhurst, B.E.; Dean, J.M. Age Estimates of Two Large Misty Grouper, Epinephelus Mystacinus (Serranidae) from Bermuda with a Comparison of the Age of Tropical Groupers in the Western Atlantic. Gulf Caribb Res 2009, 21, 73–77. [Google Scholar] [CrossRef]
- SEDAR SEDAR 61 Stock Assessment Report - Gulf of Mexico Red Grouper. SEDAR, North Charleston, SC. available online at: https://sedarweb.org/assessments/sedar-61 2019.
- SEDAR SEDAR 53 - South Atlantic Red Grouper Assessment Report. SEDAR, North Charleston, SC. 159 pp. available online at: http://sedarweb.org/sedar-53 2017.
- Sadovy, Y.; Rosario, A.; Román, A. Reproduction in an Aggregating Grouper, the Red Hind, Epinephelus Guttatus. Environ Biol Fishes 1994, 41, 269–286. [Google Scholar] [CrossRef]
- Burton, M.L.; Potts, J.C.; Carr, D.R. Age, Growth, and Natural Mortality of Rock Hind, Epinephelus Adscensionis, from the Gulf of Mexico. Bull Mar Sci 2012, 88, 903–917. [Google Scholar] [CrossRef]
- Nolan, E.T.; Downes, K.J.; Richardson, A.; Arkhipkin, A.; Brickle, P.; Brown, J.; Mrowicki, R.J.; Shcherbich, Z.; Weber, N.; Weber, S.B. Life-History Strategies of the Rock Hind Grouper Epinephelus Adscensionis at Ascension Island. J Fish Biol 2017, 91, 1549–1568. [Google Scholar] [CrossRef]
- Lombardi-Carlson, L.A.; Cook, M.; Lyon, H.; Barnett, B.; Bullock, L. A Description of Age, Growth, and Reproductive Life History Traits of Scamps from the Northern Gulf of Mexico. Marine and Coastal Fisheries 2012, 4, 129–144. [Google Scholar] [CrossRef]
- SEDAR SEDAR 68 South Atlantic Scamp Stock Assessment Report. SEDAR, North Charleston, SC. 162 pp. available online at: https://sedarweb.org/assessments/sedar-68/ 2022.
- Sanchez, P.J.; Pinsky, J.P.; Rooker, J.R. Bomb Radiocarbon Age Validation of Warsaw Grouper and Snowy Grouper. Fisheries (Bethesda) 2019, 44, 524–533. [Google Scholar] [CrossRef]
- Kolmos, K.J.; Wyanski, D.M.; White, D.B.; Mikell, P.P. Temporal Changes in the Life History of Snowy Grouper (Hyporthodus Niveatus) off North and South Carolina, and Factors That Influence Spawning Dynamics. Fishery Bulletin 2019, 117, 308–321. [Google Scholar] [CrossRef]
- SEDAR SEDAR 36 Update South Atlantic Snowy Grouper Stock Assessment Report. SEDAR, North Charleston, SC. 118 pp. available online at https://sedarweb.org/assessments/sedar-36/ 2021.
- Andrews, A.H.; Barnett, B.K.; Allman, R.J.; Moyer, R.P.; Trowbridge, H.D. Great Longevity of Speckled Hind (Epinephelus Drummondhayi), a Deep-Water Grouper, with Novel Use of Postbomb Radiocarbon Dating in the Gulf of Mexico. Canadian Journal of Fisheries and Aquatic Sciences 2013, 70, 1131–1140. [Google Scholar] [CrossRef]
- Ziskin, G.L.; Harris, P.J.; Wyanski, D.M.; Reichert, M.J.M. Indications of Continued Overexploitation of Speckled Hind along the Atlantic Coast of the Southeastern United States. Trans Am Fish Soc 2011, 140, 384–398. [Google Scholar] [CrossRef]
- White, D.B.; Wyanski, D.M.; Eleby, B.M.; Lilyestrom, C.G. Tiger Grouper (Mycteroperca Tigris): Profile of a Spawning Aggregation. Bull Mar Sci 2002, 70, 233–240. [Google Scholar]
- Caballero-Arango, D.; Brulé, T.; Nóh-Quiñones, V.; Colás-Marrufo, T.; Pérez-Díaz, E. Reproductive Biology of the Tiger Grouper in the Southern Gulf of Mexico. Trans Am Fish Soc 2013, 142, 282–299. [Google Scholar] [CrossRef]
- Cook, M.; Fitzhugh, G.R.; Franks, J.S. Validation of Yellowedge Grouper, Epinephelus Flavolimbatus, Age Using Nuclear Bomb-Produced Radiocarbon. Environ Biol Fishes 2009, 86, 461–472. [Google Scholar] [CrossRef]
- Bullock, L.H.; Godcharles, M.F.; Crabtree, R.E. Reproduction of Yellowedge Grouper, Epinephelus Flavolimbatus, from the Eastern Gulf of Mexico. Bull Mar Sci 1996, 59, 216–224. [Google Scholar]
- Burton, M.L.; Potts, J.C.; Carr, D.R. Age, Growth, and Natural Mortality of Yellowfin Grouper (Mycteroperca Venenosa) from the Southeastern United States. PeerJ 2015, 3, e1099. [Google Scholar] [CrossRef]
- García-Cagide, A.; García, T. Reproducción de Mycteroperca Bonaci y Mycteroperca Venenosa (Pisces: Serranidae) En La Plataforma Cubana. Rev Biol Trop 1996, 44, 771–780. [Google Scholar]
- Bullock, L.H.; Murphy, M.D. Aspects of the Life History of the Yellowmouth Grouper, Mycteroperca Interstitialis, in the Eastern Gulf of Mexico. Bull Mar Sci 1994, 55, 30–45. [Google Scholar]
- Sanchez, P.J.; Rooker, J.R. Age, Growth, and Mortality of Threatened Warsaw Grouper, Hyporthodus Nigritus, in the Gulf of Mexico. Fish Res 2021, 243, 106097. [Google Scholar] [CrossRef]
- Coleman, F.C.; Koenig, C.C.; Collins, L.A. Reproductive Styles of Shallow-Water Groupers (Pisces: Serranidae) in the Eastern Gulf of Mexico and the Consequences of Fishing Spawning Aggregations. Environ Biol Fishes 1996, 47, 129–141. [Google Scholar] [CrossRef]
- Moffitt, E.A.; White, J.W.; Botsford, L.W. Accurate Assessment of Marine Protected Area Success Depends on Metric and Spatiotemporal Scale of Monitoring. Mar Ecol Prog Ser 2013, 489, 17–28. [Google Scholar] [CrossRef]
- Arias, A.; Pressey, R.L.; Jones, R.E.; Álvarez-Romero, J.G.; Cinner, J.E. Optimizing Enforcement and Compliance in Offshore Marine Protected Areas: A Case Study from Cocos Island, Costa Rica. Oryx 2016, 50, 18–26. [Google Scholar] [CrossRef]
- Harford, W.J.; Sagarese, S.R.; Karnauskas, M. Coping with Information Gaps in Stock Productivity for Rebuilding and Achieving Maximum Sustainable Yield for Grouper–Snapper Fisheries. Fish and Fisheries 2019, 20, 303–321. [Google Scholar] [CrossRef]
- Conn, P.B.; Williams, E.H.; Shertzer, K.W. When Can We Reliably Estimate the Productivity of Fish Stocks? Canadian Journal of Fisheries and Aquatic Sciences 2010, 67, 511–523. [Google Scholar] [CrossRef]
- Shertzer, K.W.; Conn, P.B. Spawner-Recruit Relationships of Demersal Marine Fishes: Prior Distribution of Steepness. Bull Mar Sci 2012, 88, 39–50. [Google Scholar] [CrossRef]
- NOAA Fisheries of the Caribbean, Gulf of Mexico, and South Atlantic; Reef Fish and Red Drum Fisheries of the Gulf of Mexico; Amendments 48/5. NOAA. available at https://www.federalregister.gov/documents/2022/06/08/2022-12339/fisheries-of-the-caribbean-gulf-of-mexico-and-south-atlantic-reef-fish-and-red-drum-fisheries-of-the 2022.
- SEDAR SEDAR 85 Stock Assessment Report - Gulf of Mexico Yellowedge Grouper. SEDAR, North Charleston, SC. 267 pp. available online at: https://sedarweb.org/assessments/sedar-85/ 2023.
- Gulf Fisheries Branch SEDAR 85 Gulf of Mexico Yellowedge Grouper Operational Assessment - Additional Projections. SEDAR. available at https://sedarweb.org/documents/sedar-85-gulf-of-mexico-yellowedge-grouper-operational-assessment-additional-projections/ 2024.
- Brooks, E.N.; Shertzer, K.W.; Gedamke, T.; Vaughan, D.S. Stock Assessment of Protogynous Fish: Evaluating Measures of Spawning Biomass Used to Estimate Biological Reference Points. Fishery Bulletin 2008, 106, 12–23. [Google Scholar]
- Coleman, F.C.; Koenig, C.C.; Huntsman, G.R.; Musick, J.A.; Eklund, A.M.; McGovern, J.C.; Sedberry, G.R.; Chapman, R.W.; Grimes, C.B. Long-Lived Reef Fishes: The Grouper-Snapper Complex. Fisheries (Bethesda) 2000, 25, 14–21. [Google Scholar] [CrossRef]
- Paxton, A.B.; Harter, S.L.; Ross, S.W.; Schobernd, C.M.; Runde, B.J.; Rudershausen, P.J.; Johnson, K.H.; Shertzer, K.W.; Bacheler, N.M.; Buckel, J.A.; et al. Four Decades of Reef Observations Illuminate Deep-Water Grouper Hotspots. Fish and Fisheries 2021, 22, 749–761. [Google Scholar] [CrossRef]
- GMFMC Fisherman Feedback: Yellowedge Grouper Response Summary. Gulf of Mexico Fishery Management Council. available at https://gulfcouncil.org/wp-content/uploads/Fisherman-Feedback_Yellowedge-Grouper_2024_Final.pdf 2024.
- Sanchez, P.J.; Zapp Sluis, M.; Pinsky, J.; Miller, N.R.; Rooker, J.R. Population Structure and Regional Connectivity of Young Snowy Grouper in the Gulf of Mexico and Western Atlantic Ocean. Marine and Coastal Fisheries 2022, 14, e10119. [Google Scholar] [CrossRef]
- NOAA Fisheries of the Caribbean, Gulf of Mexico, and South Atlantic; Snapper-Grouper Fishery off the Southern Atlantic States; Amendment 14. NOAA. available at https://www.federalregister.gov/documents/2009/01/13/E9-497/fisheries-of-the-caribbean-gulf-of-mexico-and-south-atlantic-snapper-grouper-fishery-off-the 2009.
- Bacheler, N.M.; Schobernd, C.M.; Harter, S.L.; David, A.W.; Sedberry, G.R.; Kellison, G.T. No Evidence of Increased Demersal Fish Abundance Six Years after Creation of Marine Protected Areas along the Southeast United States Atlantic Coast. Bull Mar Sci 2016, 92, 447–471. [Google Scholar] [CrossRef]
- Runde, B.J.; Buckel, J.A.; Rudershausen, P.J.; Mitchell, W.A.; Ebert, E.; Cao, J.; Taylor, J.C. Evaluating the Effects of a Deep-Water Marine Protected Area a Decade after Closure: A Multifaceted Approach Reveals Equivocal Benefits to Reef Fish Populations. Front Mar Sci 2021, 8, 775376. [Google Scholar] [CrossRef]
- Sadovy, Y. The Case of the Disappearing Grouper: Epinephelus Striatus, the Nassau Grouper, in the Caribbean and Western Atlantic. Proceedings of the 45th Gulf and Caribbean Fisheries Institute 1999.
- CFMC Amendment Number 1 to the Fishery Management Plan for the Shallow-Water Reeffish Fishery, Preliminary Environmental Assessment and Regulatory Impact Review. Caribbean Fishery Management Council. available at https://caribbeanfmc.com/images/pdf-files/RF%20Amend%201%20ok.pdf 1990.
- NOAA Nassau Grouper Recovery Outline. NOAA. available at. Available online: https://www.fisheries.noaa.gov/s3//dam-migration/nassau-grouper-recovery-outline.pdf2018.
- Jackson, A.M.; Semmens, B.X.; De Mitcheson, Y.S.; Nemeth, R.S.; Heppell, S.A.; Bush, P.G.; Aguilar-Perera, A.; Claydon, J.A.B.; Calosso, M.C.; Sealey, K.S.; et al. Population Structure and Phylogeography in Nassau Grouper (Epinephelus Striatus), a Mass-Aggregating Marine Fish. PLoS One 2014, 9, e97508. [Google Scholar] [CrossRef] [PubMed]
- Harms-Tuohy, C.; Schärer, M.; Ruiz, H.; Tuohy, E.; Figuerola, M. Identifying Critical Habitats of Juvenile Nassau Grouper (Epinephelus Striatus) in Puerto Rico. Final Report. Caribbean Fishery Management Council. 42pp 2022. [CrossRef]
- Legare, B.; Maize, K.; Nemeth, R. Juvenile Nassau Grouper (Epinephelus Striatus) Utilization of Nearshore Habitats with Evidence of Adult Connectivity to a Spawning Aggregation Site. Proceedings of the 63rd Gulf and Caribbean Fisheries Institute 2011.
- Aguilar-Perera, A.; Schärer, M.T.; Nemeth, M. Occurrence of Juvenile Nassau Grouper, Epinephelus Striatus (Teleostei: Serranidae), off Mona Island, Puerto Rico: Considerations of Recruitment Potential. Caribb J Sci 2006, 42, 264–267. [Google Scholar]
- Coffill-Rivera, M.E.; Neal, J.W.; Rodríguez-Ferrer, G.; Lilyestrom, C.G. Using Lessons Learned from a Multidecadal Intercept Survey of Puerto Rico Spear Fishers to Improve Data Collection in the U.S. Caribbean. N Am J Fish Manag 2024, 44, 507–519. [Google Scholar] [CrossRef]
- Hamilton, R.; De Mitcheson, Y.S.; Aguilar-Perera, A. The Role of Local Ecological Knowledge in the Conservation and Management of Reef Fish Spawning Aggregations. Reef Fish Spawning Aggregations: Biology, Research and Management 2012, 331–369. [CrossRef]
- Beets, J.; Friedlander, A. Stock Analysis and Management Strategies for Red Hind, Epinephelus Guttatus in the U.S. Virgin Islands. Proceedings of the 42nd Gulf and Caribbean Fisheries Institute 1992, 66–79.
- Sadovy, Y.; Figuerola, M. The Status of the Red Hind Fishery in Puerto Rico and St. Thomas as Determined by Yield-per-Recruit Analysis. Proceedings of the 42nd Gulf and Caribbean Fisheries Institute 1992, 23–38.
- Sabat, A.M.; Hernández, E.A.; Toledo, C.G. Demographic Analysis of the Effect of Fishing Mortality on the Red Hind (Epinephelus Guttatus) Population in Western Puerto Rico. Proceedings of the 51st Gulf and Caribbean Fisheries Institute 2000, 169–181.
- Eristhee, N.; Kadison, E.; Murray, P.A.; Llewellyn, A. Preliminary Investigations into the Red Hind Fishery in the British Virgin Islands. 57th Gulf and Caribbean Fisheries Institute 2006, 374–384.
- Nemeth, R.S. Population Characteristics of a Recovering US Virgin Islands Red Hind Spawning Aggregation Following Protection. Mar Ecol Prog Ser 2005, 286, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, R.S.; Blondeau, J.; Herzlieb, S.; Kadison, E. Spatial and Temporal Patterns of Movement and Migration at Spawning Aggregations of Red Hind, Epinephelus Guttatus, in the U.S. Virgin Islands. Environ Biol Fishes 2007, 78, 365–381. [Google Scholar] [CrossRef]
- Nemeth, R.S.; Kadison, E.; Jossart, J.; Shivji, M.; Wetherbee, B.M.; Matley, J.K. Acoustic Telemetry Provides Insights for Improving Conservation and Management at a Spawning Aggregation Site of the Endangered Nassau Grouper (Epinephelus Striatus). Front Mar Sci 2023, 10, 1154689. [Google Scholar] [CrossRef]
- Robins, C.R.; Ray, G.C. A Field Guide to Atlantic Coast Fishes: North America. Houghton Mifflin Harcourt 1986. [Google Scholar]
- McClenachan, L. Historical Declines of Goliath Grouper Populations in South Florida, USA. Endanger Species Res 2009, 7, 175–181. [Google Scholar] [CrossRef]
- Gerhardinger, L.C.; Carvalho Marenzi, R.; Andrade Bertoncini, Á.; Pereira Medeiros, R.; Hostim-Silva, M. Local Ecological Knowledge on the Goliath Grouper Epinephelus Itajara (Teleostei: Serranidae) in Southern Brazil. Neotropical Ichthyology 2006, 4, 441–450. [Google Scholar] [CrossRef]
- Barreiros, J.P.; Coleman, F.C. West African Goliath Grouper: Where Are They between Senegal and Angola? Fishes 2023, 8. [Google Scholar] [CrossRef]
- Koenig, C.C.; Coleman, F.C.; Kingon, K. Pattern of Recovery of the Goliath Grouper Epinephelus Itajara Population in the Southeastern US. Bull Mar Sci 2011, 87, 891–911. [Google Scholar] [CrossRef]
- Koenig, C.C.; Coleman, F.C.; Malinowski, C.R. Atlantic Goliath Grouper of Florida: To Fish or Not to Fish. Fisheries (Bethesda) 2020, 45, 20–32. [Google Scholar] [CrossRef]
- Malinowski, C.; Coleman, F.; Koenig, C.; Locascio, J.; Murie, D. Are Atlantic Goliath Grouper, Epinephelus Itajara, Establishing More Northerly Spawning Sites? Evidence from the Northeast Gulf of Mexico. Bull Mar Sci 2019, 95, 371–391. [Google Scholar] [CrossRef]
- Shideler, G.S.; Carter, D.W.; Liese, C.; Serafy, J.E. Lifting the Goliath Grouper Harvest Ban: Angler Perspectives and Willingness to Pay. Fish Res 2015, 161, 156–165. [Google Scholar] [CrossRef]
- Coleman, F.C.; Nunes, J.A.C.C.; Bertoncini, Á.A.; Bueno, L.S.; Freitas, M.O.; Borgonha, M.; Leite, J.R.; Lima-Júnior, M.J.C.A.; Ferreira, B.; Bentes, B.; et al. Controversial Opening of a Limited Fishery for Atlantic Goliath Grouper in the United States: Implications for Population Recovery. Mar Policy 2023, 155, 105752. [Google Scholar] [CrossRef]
- Lindegren, M.; Brander, K. Adapting Fisheries and Their Management To Climate Change: A Review of Concepts, Tools, Frameworks, and Current Progress Toward Implementation. Reviews in Fisheries Science and Aquaculture 2018, 26, 400–415. [Google Scholar] [CrossRef]
- Sellinger, E.L.; Szuwalski, C.; Punt, A.E. The Robustness of Our Assumptions about Recruitment: A Re-Examination of Marine Recruitment Dynamics with Additional Data and Novel Methods. Fish Res 2024, 269. [Google Scholar] [CrossRef]
- Szuwalski, C.S.; Vert-Pre, K.A.; Punt, A.E.; Branch, T.A.; Hilborn, R. Examining Common Assumptions about Recruitment: A Meta-Analysis of Recruitment Dynamics for Worldwide Marine Fisheries. Fish and Fisheries 2015, 16, 633–648. [Google Scholar] [CrossRef]
- Akimova, A.; Núñez-Riboni, I.; Kempf, A.; Taylor, M.H. Spatially-Resolved Influence of Temperature and Salinity on Stock and Recruitment Variability of Commercially Important Fishes in the North Sea. PLoS One 2016, 11, e0161917. [Google Scholar] [CrossRef]
- Bogstad, B.; Dingsør, G.E.; Ingvaldsen, R.B.; Gjøsæter, H. Changes in the Relationship between Sea Temperature and Recruitment of Cod, Haddock and Herring in the Barents Sea. Marine Biology Research 2013, 9, 895–907. [Google Scholar] [CrossRef]
- Gross, J.M.; Sadler, P.; Hoenig, J.M. Evaluating a Possible New Paradigm for Recruitment Dynamics: Predicting Poor Recruitment for Striped Bass (Morone Saxatilis) from an Environmental Variable. Fish Res 2022, 252, 106329. [Google Scholar] [CrossRef]
- Brulé, T.; Renán, X.; Colás-Marrufo, T. Potential Impact of Climate Change on Fish Reproductive Phenology: A Case Study in Gonochoric and Hermaphrodite Commercially Important Species from the Southern Gulf of Mexico. Fishes 2022, 7, 156. [Google Scholar] [CrossRef]
- Lema, S.C.; Luckenbach, J.A.; Yamamoto, Y.; Housh, M.J. Fish Reproduction in a Warming World: Vulnerable Points in Hormone Regulation from Sex Determination to Spawning. Philosophical Transactions of the Royal Society B: Biological Sciences 2024, 379. [Google Scholar] [CrossRef] [PubMed]
- Miranda, L.A.; Chalde, T.; Elisio, M.; Strüssmann, C.A. Effects of Global Warming on Fish Reproductive Endocrine Axis, with Special Emphasis in Pejerrey Odontesthes Bonariensis. Gen Comp Endocrinol 2013, 192, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Fitzhugh, G.R.; Koenig, C.C.; Coleman, F.C.; Grimes, C.B.; Sturges III, W. Spatial and Temporal Patterns in Fertilization of Young Gag (Mycteroperca Microlepis) along the West Florida Shelf. Bull Mar Sci 2005, 77, 377–396. [Google Scholar]
- Payne, M.R.; C Hatfield, E.M.; Dickey-Collas, M.; Falkenhaug, T.; Gallego, A.; Gröger, J.; Licandro, P.; Llope, M.; Munk, P.; Röckmann, C.; et al. Recruitment in a Changing Environment: The 2000s North Sea Herring Recruitment Failure. ICES Journal of Marine Science 2009, 66, 272–277. [Google Scholar] [CrossRef]
- Bacheler, N.M.; Ballenger, J.C. Decadal-Scale Decline of Scamp (Mycteroperca Phenax) Abundance along the Southeast United States Atlantic Coast. Fish Res 2018, 204, 74–87. [Google Scholar] [CrossRef]
- Bacheler, N.M.; Klibansky, N.; Bubley, W.J.; Smart, T.I. Low Recruitment Drives the Decline of Red Porgy (Pagrus Pagrus) along the Southeast USA Atlantic Coast: Inferences from Fishery-Independent Trap and Video Monitoring. PLoS One 2023, 18, e0286078. [Google Scholar] [CrossRef]
- Wade, K.J.; Shertzer, K.W.; Craig, J.K.; Williams, E.H. Correlations in Recruitment Patterns of Atlantic Reef Fishes off the Southeastern United States Based on Multi-Decadal Estimates from Stock Assessments. Reg Stud Mar Sci 2023, 57, 102736. [Google Scholar] [CrossRef]
- Armstrong, C.W.; Falk-Petersen, J. Habitat-Fisheries Interactions: A Missing Link? ICES Journal of Marine Science 2008, 65, 817–821. [Google Scholar] [CrossRef]
- Minello, T.J.; Able, K.W.; Weinstein, M.P.; Hays, C.G. Salt Marshes as Nurseries for Nekton: Testing Hypotheses on Density, Growth and Survival through Meta-Analysis. Mar Ecol Prog Ser 2003, 246, 39–59. [Google Scholar] [CrossRef]
- Beck, M.B.; Heck, K.L.; Able, K.W.; Childers, D.L.; Eggleston, D.B.; Gillanders, B.M.; Halpern, B.; Hays, C.G.; Hoshino, K.; Minello, T.J.; et al. The Identification, Conservation, and Management of Estuarine and Marine Nurseries for Fish and Invertebrates. Bioscience 2001, 51, 633–641. [Google Scholar] [CrossRef]
- Aarts, B.G.W.; Van Den Brink, F.W.B.; Nienhuis, P.H. Habitat Loss as the Main Cause of the Slow Recovery of Fish Faunas of Regulated Large Rivers in Europe: The Transversal Floodplain Gradient. River Res Appl 2004, 20, 3–23. [Google Scholar] [CrossRef]
- Stamp, T.; West, E.; Robbins, T.; Plenty, S.; Sheehan, E. Large-Scale Historic Habitat Loss in Estuaries and Its Implications for Commercial and Recreational Fin Fisheries. ICES Journal of Marine Science 2022, 79, 1981–1991. [Google Scholar] [CrossRef]
- Vecchio, J.L.; Peebles, E.B. Lifetime-Scale Ontogenetic Movement and Diets of Red Grouper Inferred Using a Combination of Instantaneous and Archival Methods. Environ Biol Fishes 2022, 105, 1887–1906. [Google Scholar] [CrossRef]
- Weisberg, R.H.; Zheng, L.; Peebles, E. Gag Grouper Larvae Pathways on the West Florida Shelf. Cont Shelf Res 2014, 88, 11–23. [Google Scholar] [CrossRef]
- Koenig, C.C.; Coleman, F.C.; Eklund, A.-M.; Schull, J.; Ueland, J. Mangroves Are Essential Nursery Habitat for Goliath Grouper (Epinephelus Itajara). Bull Mar Sci 2007, 80, 567–586. [Google Scholar]
- Casey, J.P.; Poulakis, G.R.; Stevens, P.W. Habitat Use by Juvenile Gag, Mycteroperca Microlepis (Pisces: Serranidae), in Subtropical Charlotte Harbor, Florida (USA). Gulf Caribb Res 2007, 19. [Google Scholar] [CrossRef]
- Switzer, T.S.; Keenan, S.F.; Stevens, P.W.; McMichael, R.H.; MacDonald, T.C. Incorporating Ecology into Survey Design: Monitoring the Recruitment of Age-0 Gags in the Eastern Gulf of Mexico. N Am J Fish Manag 2015, 35, 1132–1143. [Google Scholar] [CrossRef]
- Bardou, R.; Osland, M.J.; Scyphers, S.; Shepard, C.; Aerni, K.E.; Alemu I, J.B.; Crimian, R.; Day, R.H.; Enwright, N.M.; Feher, L.C.; et al. Rapidly Changing Range Limits in a Warming World: Critical Data Limitations and Knowledge Gaps for Advancing Understanding of Mangrove Range Dynamics in the Southeastern USA. Estuaries and Coasts 2023 46:5 2023, 46, 1123–1140. [Google Scholar] [CrossRef]
- Zimmerman, R.C. Scaling up: Predicting the Impacts of Climate Change on Seagrass Ecosystems. Estuaries and Coasts 2021, 44, 558–576. [Google Scholar] [CrossRef]
- Beck, M.W.; Flaherty-Walia, K.; Scolaro, S.; Burke, M.C.; Furman, B.T.; Karlen, D.J.; Pratt, C.; Anastasiou, C.J.; Sherwood, E.T. Hot and Fresh: Evidence of Climate-Related Suboptimal Water Conditions for Seagrass in a Large Gulf Coast Estuary. Estuaries and Coasts 2024. [Google Scholar] [CrossRef]
- Yarbro, L.A.; Carlson, P.R.; Johnsey, E. Extensive and Continuing Loss of Seagrasses in Florida’s Big Bend (USA). Environ Manage 2024, 73, 876–894. [Google Scholar] [CrossRef] [PubMed]
- Rodemann, J.R.; James, W.R.; Rehage, J.S.; Baktoft, H.; Costa, S. V.; Ellis, R.D.; Gonzalez, L.; Santos, R.O. Residency and Fine-Scale Habitat Use of Juvenile Goliath Grouper (Epinephelus Itajara) in a Mangrove Nursery. Bull Mar Sci 2023, 99, 111–117. [Google Scholar] [CrossRef]
- Gannon, D.P.; Berens McCabe, E.J.; Camilleri, S.A.; Gannon, J.G.; Brueggen, M.K.; Barleycorn, A.A.; Palubok, V.I.; Kirkpatrick, G.J.; Wells, R.S. Effects of Karenia Brevis Harmful Algal Blooms on Nearshore Fish Communities in Southwest Florida. Mar Ecol Prog Ser 2009, 378, 171–186. [Google Scholar] [CrossRef]
- Medina, M.; Kaplan, D.; Milbrandt, E.C.; Tomasko, D.; Huffaker, R.; Angelini, C. Nitrogen-Enriched Discharges from a Highly Managed Watershed Intensify Red Tide (Karenia Brevis) Blooms in Southwest Florida. Science of The Total Environment 2022, 827, 154149. [Google Scholar] [CrossRef]
- Sagarese, S.R.; Gray, A.M.; Ainsworth, C.H.; Chagaris, D.D.; Mahmoudi, B. SEDAR42-AW-01 Red Tide Mortality on Red Grouper (Epinephelus Morio) between 1980 and 2009 on the West Florida Shelf. SEDAR, North Charleston, SC. 12 pp. available at https://sedarweb.org/documents/sedar-42-aw-01-red-tide-mortality-on-red-grouper-epinephelus-morio-between-1980-and-2009-on-the-west-florida-shelf/ 2015.
- Walter III, J.F.; Sagarese, S.R.; Harford, W.J.; Grüss, A.; Stumpf, R.P.; Christman, M.C. SEDAR42-RW-02 Assessing the Impact of the 2014 Red Tide Event on Red Grouper (Epinephelus Morio) in the Northeastern Gulf of Mexico. SEDAR, North Charleston, SC. 13 pp. available at https://sedarweb.org/documents/sedar-42-rw-02-assessing-the-impact-of-the-2014-red-tide-event-on-red-grouper-epinephelus-morio-in-the-northeastern-gulf-of-mexico/ 2015.
- Ainsworth, C.H.; Paris, C.B.; Perlin, N.; Dornberger, L.N.; Patterson III, W.F.; Chancellor, E.; Murawski, S.; Hollander, D.; Daly, K.; Romero, I.C.; et al. Impacts of the Deepwater Horizon Oil Spill Evaluated Using an End-to-End Ecosystem Model. PLoS One 2018, 13, e0190840. [Google Scholar] [CrossRef]
- Murawski, S.A.; Schwing, P.T.; Patterson III, W.F.; Sutton, T.T.; Montagna, P.A.; Milligan, R.J.; Joye, S.B.; Thomas, L.; Kilborn, J.P.; Paris, C.B.; et al. Vulnerability and Resilience of Living Marine Resources to the Deepwater Horizon Oil Spill: An Overview. Front Mar Sci 2023, 10, 1202250. [Google Scholar] [CrossRef]
- Patterson III, W.F.; Robinson, K.L.; Barnett, B.K.; Campbell, M.D.; Chagaris, D.C.; Chanton, J.P.; Daly, K.L.; Hanisko, D.S.; Hernandez, F.J.; Murawski, S.A.; et al. Evidence of Population-Level Impacts and Resiliency for Gulf of Mexico Shelf Taxa Following the Deepwater Horizon Oil Spill. Front Mar Sci 2023, 10, 1198163. [Google Scholar] [CrossRef]
- Stokesbury, K.D.E.; Cassidy, K.; Lowery, T.M. Constructing a Baseline Groundfish Trawl Survey for an Offshore Windfarm Development Area. Marine and Coastal Fisheries 2023, 15, e10267. [Google Scholar] [CrossRef]
- Jech, J.M.; Lipsky, A.; Moran, P.; Matte, G.; Diaz, G. Fish Distribution in Three Dimensions around the Block Island Wind Farm as Observed with Conventional and Volumetric Echosounders. Marine and Coastal Fisheries 2023, 15, e10265. [Google Scholar] [CrossRef]
- Van Hoeck, R. V.; Rowell, T.J.; Dean, M.J.; Rice, A.N.; Van Parijs, S.M. Comparing Atlantic Cod Temporal Spawning Dynamics across a Biogeographic Boundary: Insights from Passive Acoustic Monitoring. Marine and Coastal Fisheries 2023, 15, e10226. [Google Scholar] [CrossRef]
- FAO The Ecosystem Approach to Fisheries. FAO Technical Guidelines for Responsible Fisheries. No. 4, suppl. 2. Rome, FAO. 112 p. available at https://www.fao.org/in-action/globefish/publications/details-publication/en/c/346126/ 2003, 112.
- Howell, D.; Schueller, A.M.; Bentley, J.W.; Buchheister, A.; Chagaris, D.; Cieri, M.; Drew, K.; Lundy, M.G.; Pedreschi, D.; Reid, D.G.; et al. Combining Ecosystem and Single-Species Modeling to Provide Ecosystem-Based Fisheries Management Advice within Current Management Systems. Front Mar Sci 2021, 7, 607831. [Google Scholar] [CrossRef]
- Karp, M.A.; Link, J.S.; Grezlik, M.; Cadrin, S.; Fay, G.; Lynch, P.; Townsend, H.; Methot, R.D.; Adams, G.D.; Blackhart, K.; et al. Increasing the Uptake of Multispecies Models in Fisheries Management. ICES Journal of Marine Science 2023, 80, 243–257. [Google Scholar] [CrossRef]
- NOAA NOAA Fisheries Policy 01-120: Ecosystem-Based Fisheries Management Policy. NOAA. available at https://www.fisheries.noaa.gov/resource/document/ecosystem-based-fisheries-management-policy 2016.
- Anstead, K.A.; Drew, K.; Chagaris, D.; Schueller, A.M.; McNamee, J.E.; Buchheister, A.; Nesslage, G.; Uphoff, J.H.; Wilberg, M.J.; Sharov, A.; et al. The Path to an Ecosystem Approach for Forage Fish Management: A Case Study of Atlantic Menhaden. Front Mar Sci 2021, 8, 607657. [Google Scholar] [CrossRef]
- Grüss, A.; Rose, K.A.; Simons, J.; Ainsworth, C.H.; Babcock, E.A.; Chagaris, D.D.; De Mutsert, K.; Froeschke, J.; Himchak, P.; Kaplan, I.C.; et al. Recommendations on the Use of Ecosystem Modeling for Informing Ecosystem-Based Fisheries Management and Restoration Outcomes in the Gulf of Mexico. Marine and Coastal Fisheries 2017, 9, 281–295. [Google Scholar] [CrossRef]
- Seara, T.; Williams, S.M.; Acevedo, K.; Garcia-Molliner, G.; Tzadik, O.; Duval, M.; Cruz-Motta, J.J. Development and Analyses of Stakeholder Driven Conceptual Models to Support the Implementation of Ecosystem-Based Fisheries Management in the U.S. Caribbean. PLoS One 2024, 19, e0304101. [Google Scholar] [CrossRef]
- Chagaris, D.; Sagarese, S.; Farmer, N.; Mahmoudi, B.; de Mutsert, K.; VanderKooy, S.; Patterson, W.F.; Kilgour, M.; Schueller, A.; Ahrens, R.; et al. Management Challenges Are Opportunities for Fisheries Ecosystem Models in the Gulf of Mexico. Mar Policy 2019, 101, 1–7. [Google Scholar] [CrossRef]
- Chagaris, D.; Drew, K.; Schueller, A.; Cieri, M.; Brito, J.; Buchheister, A. Ecological Reference Points for Atlantic Menhaden Established Using an Ecosystem Model of Intermediate Complexity. Front Mar Sci 2020, 7, 606417. [Google Scholar] [CrossRef]
- Chagaris, D.; Sinnickson, D. SEDAR61-WP-06 An Index of Red Tide Mortality on Red Grouper in the Eastern Gulf of Mexico. SEDAR, North Charleston, SC. 16 pp. available at https://sedarweb.org/documents/sedar-61-wp-06-an-index-of-red-tide-mortality-on-red-grouper-in-the-eastern-gulf-of-mexico/ 2018.
- Vilas, D.; Chagaris, D.; Buczkowski, J. SEDAR72-WP-01 Red Tide Mortality on Gag Grouper from 2002-2018 Generated by an Ecospace Model of the West Florida Shelf. SEDAR, North Charleston, SC. 17 pp. available at https://sedarweb.org/documents/sedar-72-wp-01-red-tide-mortality-on-gag-grouper-from-2002-2018-generated-by-an-ecospace-model-of-the-west-florida-shelf/ 2020.
- Pita, C.; Pierce, G.J.; Theodossiou, I. Stakeholders’ Participation in the Fisheries Management Decision-Making Process: Fishers’ Perceptions of Participation. Mar Policy 2010, 34, 1093–1102. [Google Scholar] [CrossRef]
- Mackinson, S.; Wilson, D.C.; Galiay, P.; Deas, B. Engaging Stakeholders in Fisheries and Marine Research. Mar Policy 2011, 35, 18–24. [Google Scholar] [CrossRef]
- Bentley, J.W.; Hines, D.E.; Borrett, S.R.; Serpetti, N.; Hernandez-Milian, G.; Fox, C.; Heymans, J.J.; Reid, D.G. Combining Scientific and Fishers’ Knowledge to Co-Create Indicators of Food Web Structure and Function. ICES Journal of Marine Science 2019, 76, 2218–2234. [Google Scholar] [CrossRef]
- Cooke, S.J.; Nguyen, V.M.; Chapman, J.M.; Reid, A.J.; Landsman, S.J.; Young, N.; Hinch, S.G.; Schott, S.; Mandrak, N.E.; Semeniuk, C.A.D. Knowledge Co-Production: A Pathway to Effective Fisheries Management, Conservation, and Governance. Fisheries (Bethesda) 2021, 46, 89–97. [Google Scholar] [CrossRef]
- Mills, K.E.; Armitage, D.; Eurich, J.G.; Kleisner, K.M.; Pecl, G.T.; Tokunaga, K. Co-Production of Knowledge and Strategies to Support Climate Resilient Fisheries. ICES Journal of Marine Science 2023, 80, 358–361. [Google Scholar] [CrossRef]
- Collins, A.B.; Fluech, B.; Siders, Z.A.; Sipos, M.; Zangroniz, A. Diving for Data: Florida Sea Grant’s Great Goliath Grouper Count. Oceanography 2024, 37, 102–103. [Google Scholar] [CrossRef]
- SEDAR SEDAR 47 Stock Assessment Report - Southeastern U.S. Goliath Grouper. SEDAR, North Charleston, SC. available at https://sedarweb.org/assessments/sedar-47/ 2016.
- Farmer, N.A.; Heyman, W.D.; Karnauskas, M.; Kobara, S.; Smart, T.I.; Ballenger, J.C.; Reichert, M.J.M.; Wyanski, D.M.; Tishler, M.S.; Lindeman, K.C.; et al. Timing and Locations of Reef Fish Spawning off the Southeastern United States. PLoS One 2017, 12, e0172968. [Google Scholar] [CrossRef] [PubMed]
- Heyman, W.D.; Grüss, A.; Biggs, C.R.; Kobara, S.; Farmer, N.A.; Karnauskas, M.; Lowerre-Barbieri, S.; Erisman, B. Cooperative Monitoring, Assessment, and Management of Fish Spawning Aggregations and Associated Fisheries in the U.S. Gulf of Mexico. Mar Policy 2019, 109, 103689. [Google Scholar] [CrossRef]
- Punt, A.E.; Butterworth, D.S.; de Moor, C.L.; De Oliveira, J.A.A.; Haddon, M. Management Strategy Evaluation: Best Practices. Fish and Fisheries 2016, 17, 303–334. [Google Scholar] [CrossRef]
- Damiano, M.D.; Shertzer, K.W.; Cao, J. Exploring Tradeoffs in Southeast United States Marine Fisheries Management Using Management Strategy Evaluation. Fish Res 2024, 275, 107028. [Google Scholar] [CrossRef]
- Patterson III, W.F.; Barnett, B.K.; TinHan, T.C.; Lowerre-Barbieri, S.K. Eye Lens Δ14C Validates Otolith-Derived Age Estimates of Gulf of Mexico Reef Fishes. Canadian Journal of Fisheries and Aquatic Sciences 2021, 78, 13–17. [Google Scholar] [CrossRef]
- Patterson III, W.F.; Chamberlin, D.W. Application of the Bomb Radiocarbon Chronometer with Eye Lens Core Δ14C for Age Validation in Deepwater Reef Fishes. Canadian Journal of Fisheries and Aquatic Sciences 2023, 80, 1047–1052. [Google Scholar] [CrossRef]
- Weber, D.N.; Fields, A.T.; Patterson, W.F.; Barnett, B.K.; Hollenbeck, C.M.; Portnoy, D.S. Novel Epigenetic Age Estimation in Wild-Caught Gulf of Mexico Reef Fishes. Canadian Journal of Fisheries and Aquatic Sciences 2022, 79, 1–5. [Google Scholar] [CrossRef]
- Weber, D.N.; Fields, A.T.; Chamberlin, D.W.; Patterson, W.F.; Portnoy, D.S. Epigenetic Age Estimation in a Long-Lived, Deepwater Scorpionfish: Insights into Epigenetic Clock Development. Canadian Journal of Fisheries and Aquatic Sciences 2024, 81, 620–631. [Google Scholar] [CrossRef]
- Appeldoorn-Sanders, E.; Schärer-Umpierre, M.T.; Cruz-Motta, J.J. Passive Acoustics as a Tool to Quantify/Characterize Vessel Activity at Fish Spawning Aggregation Sites. Ocean Coast Manag 2022, 226, 106270. [Google Scholar] [CrossRef]
- Woodward, C.; Schärer-Umpierre, M.; Nemeth, R.S.; Appeldoorn, R.; Chérubin, L.M. Spatial Distribution of Spawning Groupers on a Caribbean Reef from an Autonomous Surface Platform. Fish Res 2023, 266, 106794. [Google Scholar] [CrossRef]
- Appeldoorn-Sanders, E.; Zayas-Santiago, C.; Schärer-Umpierre, M. Characterization and Temporal Patterns of Red Hind Grouper, Epinephelus Guttatus, Choruses at a Single Aggregation Site over a 10-Year Period. Environ Biol Fishes 2023, 106, 1953–1969. [Google Scholar] [CrossRef]
- Chérubin, L.M.; Dalgleish, F.; Ibrahim, A.K.; Schärer-Umpierre, M.; Nemeth, R.S.; Matthews, A.; Appeldoorn, R. Fish Spawning Aggregations Dynamics as Inferred from a Novel, Persistent Presence Robotic Approach. Front Mar Sci 2020, 6, 779. [Google Scholar] [CrossRef]
- Keller, J.A.; Herbig, J.L.; Morley, D.; Wile, A.; Barbera, P.; Acosta, A. Grouper Tales: Use of Acoustic Telemetry to Evaluate Grouper Movements at Western Dry Rocks in the Florida Keys. Marine and Coastal Fisheries 2020, 12, 290–307. [Google Scholar] [CrossRef]
- Farmer, N.A.; Ault, J.S. Grouper and Snapper Movements and Habitat Use in Dry Tortugas, Florida. Mar Ecol Prog Ser 2011, 433, 169–184. [Google Scholar] [CrossRef]
- Tharp, R.M.; Hostetter, N.J.; Paxton, A.B.; Taylor, J.C.; Buckel, J.A. Artificial Structure Selection by Economically Important Reef Fishes at North Carolina Artificial Reefs. Front Mar Sci 2024, 11, 1373494. [Google Scholar] [CrossRef]
- Vecchio, J.L.; Ostroff, J.L.; Peebles, E.B. Isotopic Characterization of Lifetime Movement by Two Demersal Fishes from the Northeastern Gulf of Mexico. Mar Ecol Prog Ser 2021, 657, 161–172. [Google Scholar] [CrossRef]
- Hanson, P.J.; Koenig, C.C.; Zdanowicz, V.S. Elemental Composition of Otoliths Used to Trace Estuarine Habitats of Juvenile Gag Mycteroperca Microlepis along the West Coast of Florida. Mar Ecol Prog Ser 2004, 267, 253–265. [Google Scholar] [CrossRef]
- Gonzalez Colmenares, G.M.; Gonzalez Montes, A.J.; Harms-Tuohy, C.A.; Schizas, N. V. Using EDNA Sampling for Species-Specific Fish Detection in Tropical Oceanic Samples: Limitations and Recommendations for Future Use. PeerJ 2023, 11, e14810. [Google Scholar] [CrossRef]
- Bacheler, N.M.; Gillum, Z.D.; Gregalis, K.C.; Pickett, E.P.; Schobernd, C.M.; Schobernd, Z.H.; Teer, B.Z.; Smart, T.I.; Bubley, W.J. Comparison of Video and Traps for Detecting Reef Fishes and Quantifying Species Richness in the Continental Shelf Waters of the Southeast USA. Mar Ecol Prog Ser 2022, 698, 111–123. [Google Scholar] [CrossRef]
- Switzer, T.S.; Keenan, S.F.; Thompson, K.A.; Shea, C.P.; Knapp, A.R.; Campbell, M.D.; Noble, B.; Gardner, C.; Christman, M.C. Integrating Assemblage Structure and Habitat Mapping Data into the Design of a Multispecies Reef Fish Survey. Marine and Coastal Fisheries 2023, 15, e10245. [Google Scholar] [CrossRef]
- Powers, S.P.; Drymon, J.M.; Hightower, C.L.; Spearman, T.; Bosarge, G.S.; Jefferson, A. Distribution and Age Composition of Red Snapper across the Inner Continental Shelf of the North-Central Gulf of Mexico. Trans Am Fish Soc 2018, 147, 791–805. [Google Scholar] [CrossRef]
- Campbell, M.D.; Rademacher, K.R.; Noble, B.; Salisbury, J.; Felts, P.; Moser, J.; Caillouet, R.; Hendon, M.; Driggers, W.B. Status and Trends of Marbled Grouper in the North-Central Gulf of Mexico. Marine and Coastal Fisheries 2019, 11, 114–124. [Google Scholar] [CrossRef]
- Aldridge, S.E.; Dixon, O.F.L.; de Silva, C.; Kohler, J.K.; Shipley, O.N.; Phillips, B.T.; Fernandes, T.F.; Austin, T.; Ormond, R.F.; Gore, M.A.; et al. Depth Range Extension for the Misty Grouper Hyporthodus Mystacinus Documented via Deep-Sea Landers throughout the Greater Caribbean. Fishes 2024, 9, 114. [Google Scholar] [CrossRef]
- Olson, J.C.; Appeldoorn, R.S.; Schärer-Umpierre, M.T.; Cruz-Motta, J.J. Recovery When You Are on Your Own: Slow Population Responses in an Isolated Marine Reserve. PLoS One 2019, 14, e0223102. [Google Scholar] [CrossRef]
- Johnston, M.W.; Bernard, A.M. A Bank Divided: Quantifying a Spatial and Temporal Connectivity Break between the Campeche Bank and the Northeastern Gulf of Mexico. Mar Biol 2017, 164, 12. [Google Scholar] [CrossRef]
Figure 1.
An elusive Marbled Grouper Dermatolepis inermis landed off the coast of Alabama, USA. Photo credit: M. E. Coffill-Rivera.
Figure 1.
An elusive Marbled Grouper Dermatolepis inermis landed off the coast of Alabama, USA. Photo credit: M. E. Coffill-Rivera.

Figure 2.
Number of recreational saltwater angler trips across the U.S. Gulf of Mexico and Atlantic coasts. Points denote the yearly estimate while lines bound the 95% confidence intervals. Data are publicly available at https://www.fisheries.noaa.gov/data-tools/recreational-fisheries-statistics-queries.
Figure 2.
Number of recreational saltwater angler trips across the U.S. Gulf of Mexico and Atlantic coasts. Points denote the yearly estimate while lines bound the 95% confidence intervals. Data are publicly available at https://www.fisheries.noaa.gov/data-tools/recreational-fisheries-statistics-queries.
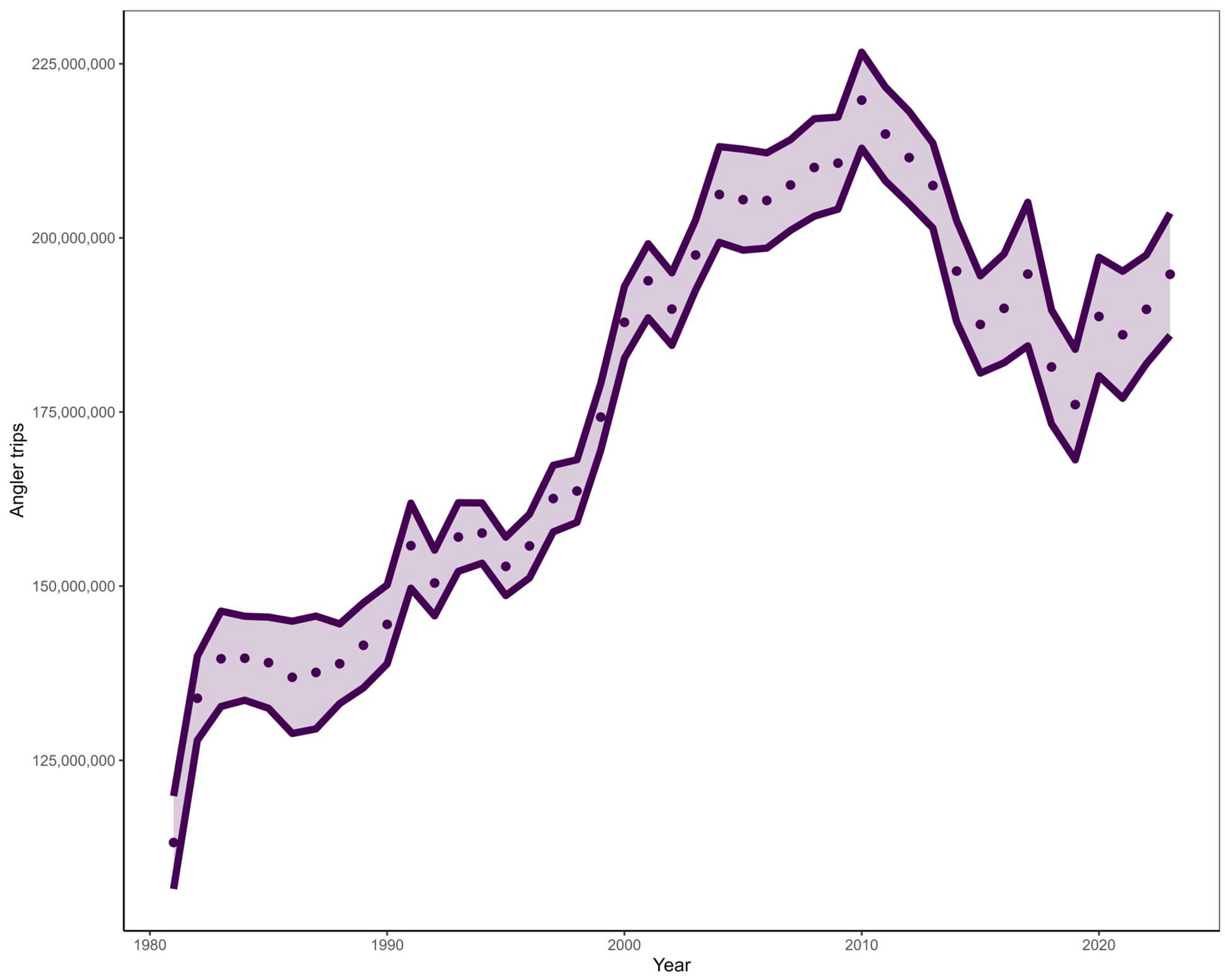
Figure 3.
Examples of groupers distributed across the southeastern U.S. that display monandric protogynous hermaphroditism. (A) Juvenile Red Grouper Epinephelus morio. (B) Juvenile Gag Mycteroperca microlepis. (C) Juvenile Scamp M. phenax. (D) Juvenile Graysby Cephalopholis cruentata. Photo credits: M. E. Coffill-Rivera.
Figure 3.
Examples of groupers distributed across the southeastern U.S. that display monandric protogynous hermaphroditism. (A) Juvenile Red Grouper Epinephelus morio. (B) Juvenile Gag Mycteroperca microlepis. (C) Juvenile Scamp M. phenax. (D) Juvenile Graysby Cephalopholis cruentata. Photo credits: M. E. Coffill-Rivera.

Figure 4.
Population trends represented as fraction unfished for U.S. Gulf of Mexico (GOM) and South Atlantic (SA) grouper stocks that display monandric protogynous hermaphroditism assessed with integrated models. Fraction unfished for GOM stocks is measured using spawning stock biomass while SA stocks are measured using total biomass. Stock assessments are publicly available at https://sedarweb.org/sedar-assessments/.
Figure 4.
Population trends represented as fraction unfished for U.S. Gulf of Mexico (GOM) and South Atlantic (SA) grouper stocks that display monandric protogynous hermaphroditism assessed with integrated models. Fraction unfished for GOM stocks is measured using spawning stock biomass while SA stocks are measured using total biomass. Stock assessments are publicly available at https://sedarweb.org/sedar-assessments/.
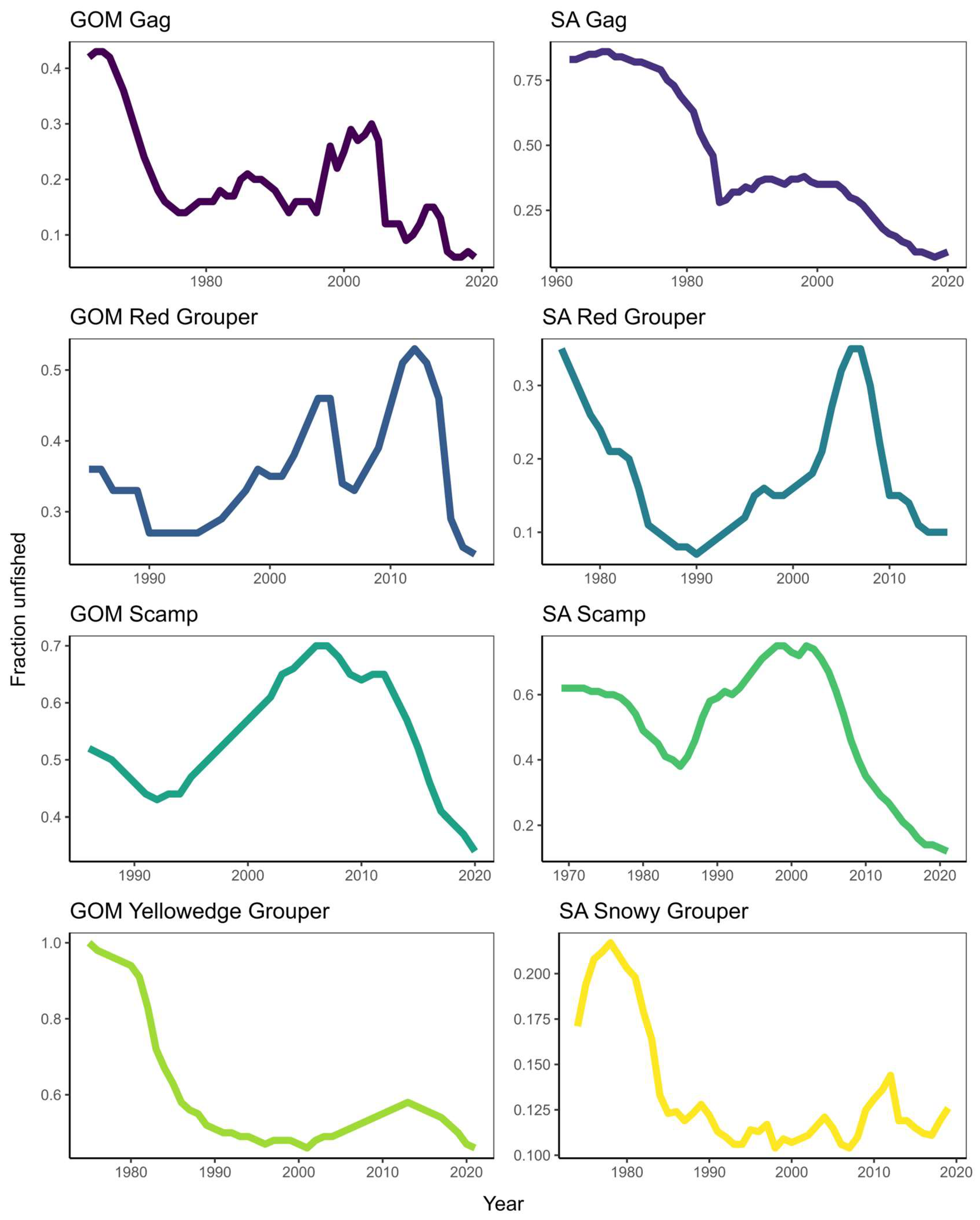
Figure 5.
(A) Yellowedge Grouper Hyporthodus flavolimbatus and (B) Snowy Grouper H. niveatus, the two most landed groupers in the U.S. Gulf of Mexico and South Atlantic’s deepwater grouper complex. Both species display monandric protogynous hermaphroditism. Photo credits: M. E. Coffill-Rivera.
Figure 5.
(A) Yellowedge Grouper Hyporthodus flavolimbatus and (B) Snowy Grouper H. niveatus, the two most landed groupers in the U.S. Gulf of Mexico and South Atlantic’s deepwater grouper complex. Both species display monandric protogynous hermaphroditism. Photo credits: M. E. Coffill-Rivera.

Figure 6.
U.S. Gulf of Mexico commercial landings from groupers managed under the deepwater grouper complex during the Individual Fishing Quota era. Landings are publicly available at https://noaa-sero.s3.amazonaws.com/drop-files/cs/2022_GT_AnnualReport_Final.pdf.
Figure 6.
U.S. Gulf of Mexico commercial landings from groupers managed under the deepwater grouper complex during the Individual Fishing Quota era. Landings are publicly available at https://noaa-sero.s3.amazonaws.com/drop-files/cs/2022_GT_AnnualReport_Final.pdf.
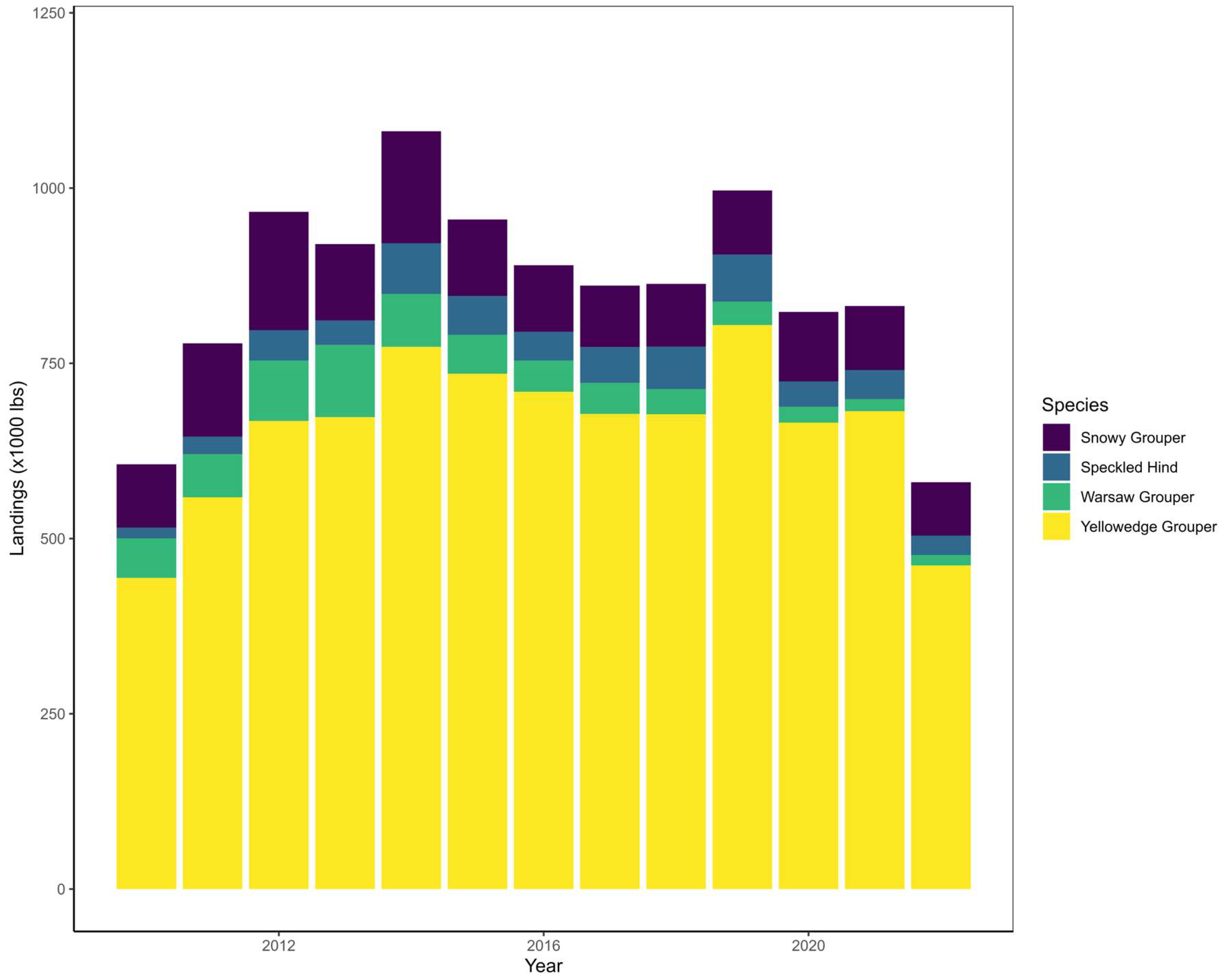
Figure 7.
Atlantic Goliath Grouper Epinephelus itajara displayed at Gene Johnson’s Tackle Shop, Daytona Beach, Florida, circa 1920. Public domain photograph courtesy of State Archives of Florida, Florida Memory. Available at https://www.floridamemory.com/items/show/140117.
Figure 7.
Atlantic Goliath Grouper Epinephelus itajara displayed at Gene Johnson’s Tackle Shop, Daytona Beach, Florida, circa 1920. Public domain photograph courtesy of State Archives of Florida, Florida Memory. Available at https://www.floridamemory.com/items/show/140117.


Table 1.
Grouper species distributed across the southeastern and Caribbean U.S. “DPH” denotes diandric protogynous hermaphroditism, “G” denotes gonochorism, and “MPH” denotes monandric protogynous hermaphroditism. “CAB” denotes the Caribbean, “GOM” denotes the Gulf of Mexico, and “SA” denotes the South Atlantic. Brackets denote the reference number.
Table 1.
Grouper species distributed across the southeastern and Caribbean U.S. “DPH” denotes diandric protogynous hermaphroditism, “G” denotes gonochorism, and “MPH” denotes monandric protogynous hermaphroditism. “CAB” denotes the Caribbean, “GOM” denotes the Gulf of Mexico, and “SA” denotes the South Atlantic. Brackets denote the reference number.
| Common name | Scientific name | Longevity (y) | Reproductive strategy | Managed? | Overfished and/or overfishing? |
|---|---|---|---|---|---|
| Atlantic Goliath Grouper | Epinephelus itajara | >30 [39] | G, DPH [40] | CAB, GOM, SA | CAB [41] |
| Black Grouper | Mycteroperca bonaci | >20 [42] | MPH [43] | CAB,GOM, SA | - |
| Coney | Cephalopholis fulva | >10 [44] | MPH [45] | CAB, SA | - |
| Gag | Mycteroperca microlepis | >20 [46] | MPH [47] | GOM, SA | GOM [48], SA [49] |
| Graysby | Cephalopholis cruentata | >20 [50] | MPH [50] | CAB, SA | - |
| Marbled Grouper | Dermatolepis inermis | - | - | - | - |
| Misty Grouper | Hyporthodus mystacinus | >100 [51] | - | SA | - |
| Nassau Grouper | Epinephelus striatus | >20 [12] | G [12] | CAB, GOM, SA | - |
| Red Grouper | Epinephelus morio | >20 [52] | MPH [43] | CAB, GOM, SA | SA [53] |
| Red Hind | Epinephelus guttatus | >10 [10] | MPH [54] | CAB, SA | - |
| Rock Hind | Epinephelus adscensionis | >30 [55] | MPH [56] | CAB, SA | - |
| Scamp | Mycteroperca phenax | >30 [57] | MPH [57] | GOM, SA | SA [58] |
| Snowy Grouper | Hyporthodus niveatus | >50 [59] | MPH [60] | GOM, SA | SA [61] |
| Speckled Hind | Epinephelus drummondhayi | >40 [62] | MPH [63] | GOM, SA | - |
| Tiger Grouper | Mycteroperca tigris | >10 [64] | MPH [65] | CAB | - |
| Yellowedge Grouper | Hyporthodus flavolimbatus | >80 [66] | MPH [67] | CAB, GOM, SA | - |
| Yellowfin Grouper | Mycteroperca venenosa | >30 [68] | MPH [69] | CAB | - |
| Yellowmouth Grouper | Mycteroperca interstitialis | >20 [70] | MPH [70] | GOM, SA | - |
| Warsaw Grouper | Hyporthodus nigritus | >90 [71] | - | GOM, SA | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



