
Submitted:
03 September 2024
Posted:
04 September 2024
You are already at the latest version
Abstract
Urbanization degrades natural habitats and creates new urban ecosystems, like domestic gardens. The plant composition of these gardens varies with socio-economic factors and urban planning levels. However, the diversity and impact of introduced species are often poorly assessed, causing potential ecological imbalances, particularly in Lubumbashi (DR Congo). The objective was to analyze the spatial structure, plant diversity, propagation strategies, and ecological functions of domestic gardens. Three distinct neighborhoods were selected: planned, unplanned and residential neighborhood. 20 avenues (with 5 plots per avenue) were chosen to represent the diversity within each neighborhood, and stratified random sampling of plots was conducted to analyze gardening practices. Gardens were classified into types, and their vegetation was evaluated based on species origin and ecological impact. The analysis of domestic gardens in Lubumbashi reveals significant variations across different neighborhood types. Residential neighborhoods exhibit larger average garden sizes (315.1 m²), higher species richness (22 species), and larger plot sizes (1032 m²) compared to unplanned and planned neighborhoods, where garden areas and species richness are notably lower. Rectangular gardens dominate in unplanned areas, while planned neighborhoods feature more intentional landscaping elements, such as flowerbeds and hedges. The use of gardens for food production is prominent in planned areas (40.7%), whereas residential neighborhoods prioritize ornamentation (51.4%). The study identifies 232 species across 68 families, with a predominance of exotic species (80%) in all neighborhoods, particularly in unplanned areas (82.25%). The data reveals that Mangifera indica and Persea americana are abundant in all neighborhoods, illustrating their adaptability to different urban contexts. Herbaceous species are most common, followed by woody plants, with vines being sparse. Species dispersal is primarily driven by human activities (anthropochory), accounting for over 85% in all neighborhoods. These findings highlight the strong human influence on the composition and structure of domestic gardens in Lubumbashi, emphasizing the dominance of exotic species and the importance of anthropogenic factors in shaping urban green spaces. Urban policies should incorporate strategies to minimize the negative impacts of exotic species on native flora.
Keywords:
urbanization
; domestic gardens
; plant diversity
; socio-economic impact
; ecological imbalance
; exotic species
1. Introduction
Urbanization refers to the process by which urban areas expand and densify in response to demographic and economic growth [1]. From 1950 to 2022, the global urbanization rate increased from approximately 29% to 57%, reflecting ongoing expansion driven by industrialization and internal migration [1,2]. However, urbanization patterns vary significantly across regions. In Europe and North America, where urbanization is more advanced, rates exceed 80% and are either stabilizing or growing slowly [3]. Conversely, Asia and Africa experience rapid urbanization due to high population growth and significant rural-to-urban migration [2,4,5,6]. For instance, in Asia, countries like China have seen urbanization rates rise from 20% in 1950 to around 60% by 2022 [1]. In Africa, urbanization is accelerating, though rates remain lower at about 45% in 2022, with substantial infrastructure and urban planning challenges [5,6].
Urbanization significantly impacts vegetation, leading to deforestation, habitat fragmentation, and biodiversity loss. The expansion of urban infrastructure, such as roads and buildings, often replaces natural green spaces, reducing vegetation areas [7,8]. Despite this, urban vegetation remains crucial for providing various ecosystem services [9]. For illustration, trees and green spaces contribute to thermal regulation by providing shade and reducing urban heat islands [10]. Additionally, urban vegetation foster biodiversity by providing habitats for birds and insects, playing a role in stormwater management by absorbing and filtering water, thus reducing flood risks [11]. Moreover, urban vegetation enhances human well-being by offering recreational spaces and strengthening connections with nature [12].
In urban environments, vegetation includes both natural and newly introduced ecosystems [13]. Natural urban vegetation comprises native trees and plants that persist despite urban expansion, while urbanization also leads to artificial plant ecosystems [14], and domestic gardens exemplify these newly introduced ecosystems [15]. The plant diversity in domestic gardens is influenced by several factors: residents’ socio-economic context, the level of urban planning, and local environmental conditions [16,17]. In wealthier neighborhoods, diversity is often higher and oriented towards ornamental plants, whereas in less affluent areas, gardens tend to include practical food species [15].
The Democratic Republic of the Congo (DRC) stands out in Africa for its rich phytogeography, particularly in the Southeast around Lubumbashi, a region where diverse ecosystems such as dense riparian forest, woodland, savannas, and copper-rich steppe savanna intersect [18]. Studying domestic gardens in Lubumbashi is crucial due to this unique diversity and the impacts of rapid urbanization on local ecosystems [19]. Since independence, Lubumbashi, driven by its mining heritage and commercial role, has experienced a demographic explosion due to migration and natural growth, with six out of ten people in Haut-Katanga living in the city in 2021 [20]. The rapid urbanization resulting has led to uncontrolled densification and suburbanization, threatening local ecosystems such as woodlands and unique flora areas [21,22,23]. Industrial activities and waste pollution further exacerbate the situation, while the lack of an urban master plan leads to the destruction of green spaces in favor of new constructions [24].
Consequently, domestic gardens, mostly developed in residential plots, have become centers of both accidental and intentional botanical introduction, playing a key role in the spread of exotic plants [14]. Lubumbashi, a major economic center in Haut-Katanga, features various types of neighborhoods: planned, residential, and unplanned [25]. Residential neighborhoods, once reserved for Europeans, are relatively well-preserved despite social transitions. Planned neighborhoods, originally well-organized, are now degraded. Unplanned neighborhoods, often established illegally, suffer from a lack of planning and adequate public services. This unplanned urban expansion leads to the loss of local species and the dominance of exotic plants in residential plots, reflecting local preferences and socio-economic influences [26]. This dynamic underscore the need to understand the impact of urbanization policies on urban biodiversity to better manage vegetation in a context of rapid growth and phytogeographic diversification.
The scientific literature on the flora of domestic gardens highlights their significant plant diversity, including ornamental, medicinal, and food plants, influenced by climate, cultural preferences, and management practices [27]. These gardens play a crucial role in urban biodiversity by providing refuge for wildlife, contributing to microclimatic regulation, and participating in nutrient cycling [17]. Socio-economically, they offer food and medicinal products and help maintain local knowledge [28]. However, gaps remain, notably the lack of longitudinal studies on the evolution of flora in response to climate and urban changes [29]. Additionally, the ecosystem services provided by these gardens, such as carbon sequestration and air purification, are not sufficiently quantified [30]. The relationship between urban policies and the conservation of these green spaces is also underexplored [15,31,32]. In the DRC, research on urban domestic gardens is limited [33]. While some studies have focused on plant diversity in rural areas or fruit-trees in urbanized areas, urban gardens, particularly in Lubumbashi, are under-researched [26,34]. There is a lack of systematic floristic inventories, analyses of the socio-economic interactions influencing gardening practices, and studies on the impact of urban dynamics, such as plot fragmentation and densification, on the preservation of these gardens. Thus, Lubumbashi represents a promising field for further research in this area.
Studies on the flora of domestic gardens in urban settings can rely on reports from specialized state agencies, such as environmental management agencies and urban agriculture departments [35]. These reports provide information on plant species, gardening practices, and green space planning policies. While useful for identifying general trends and types of vegetation, these reports may have limitations [36]. They can lack detail or be outdated, and may omit specific local variations, non-compliant practices, or illegally introduced species [37]. For a more comprehensive assessment, combining socio-economic surveys with floristic inventories is relevant. Socio-economic surveys provide insights into residents’ motivations, resources, and biodiversity knowledge, revealing the impact of economic and social factors on garden composition [15]. Floristic inventories offer precise data on species, their abundance, and distribution [28]. Together, these approaches enable a thorough understanding of domestic gardens, integrating botanical and socio-economic aspects to enhance urban biodiversity and quality of life [15].
The objective of this study is to comprehensively characterize domestic gardens in selected neighborhoods of Lubumbashi by analyzing their spatial structure, plant diversity, propagation strategies, and the ecological and social functions they fulfill. It is expected that (i) the spatial structure of domestic gardens will vary significantly by neighborhood, with more organized layouts in planned areas and more heterogeneous configurations in unplanned areas; (ii) in unplanned neighborhoods, greater plant diversity is likely due to informal urban expansion and the presence of exotic species introduced by residents, with plants often dispersed through human activities such as trade and informal gardening; and (iii) in unplanned neighborhoods, gardens may serve a broader range of functions, including food, medicinal, and cultural uses, due to the lack of urban planning and less favorable socio-economic conditions.
2. Materials and Methods
2.1. Study Area
Lubumbashi (Figure 1), the capital of Haut-Katanga in the southeastern part of DRC, spans seven municipalities (Kampemba, Kenya, Kamalondo, Lubumbashi, Katuba, Ruashi, and Annexe), covering approximately 747 km2 [25]. Situated at an altitude of around 1200 meters on a plateau, Lubumbashi is in the upper basin of the Kafubu River, between 11°20’ and 12°00’ South latitude and 27°10’ and 27°43’ East longitude. The dominant soils in the region are ferrallitic, predominantly yellow and red in color [18]. The climate is classified as Cw according to Köppen’s classification, with a dry season from May to September, a rainy season from November to March, and transitional months in April and October [18]. Recent studies indicate a delay in the onset of rains and a reduction in annual precipitation, which is estimated to be around 1047 mm between 1970 and 2005 [38,39]. The average annual temperature was 20.1°C in the latter half of the 20th century, with a trend toward warming [38,39]. At the beginning of the 20th century, miombo woodland covered 90% of the region, but it has significantly receded, requiring a 35 km journey outside the city to find remnants patches [40]. Deforestation caused by agriculture, wood collection, and mining activities has transformed the miombo woodland into savannah, and subsequently into barren land in mining areas [21]. Termite mounds, once common, are now less abundant, and their soil is used for brick production [41]. Previously well-maintained green spaces are now neglected or abandoned, especially in unplanned neighborhoods [19]. Urban expansion also promotes the proliferation of invasive exotic plants, such as Mexican sunflower [42]. The city still includes a few large recreational spaces, such as a zoological garden and a golf course, but green spaces are increasingly scarce in newly developed areas [19]. Lubumbashi is a major economic center due to its mining activities, particularly copper and cobalt extraction, which drive the regional economy [25]. However, this mining prosperity contrasts with challenges in urban governance [25]. Managing a city with over 2.5 million inhabitants is often characterized by a lack of coherent planning and institutional instability, exacerbating infrastructure and urban organization issues [43]. Environmentally, rapid urban growth and industrial expansion have led to significant degradation of local ecosystems and increased pollution, threatening vital ecosystem services and exacerbating negative environmental impacts [25].
2.2. Selection of Neighborhoods, Avenues, and Residential Plots
Three neighborhoods were chosen for the study based on their distinct typologies. Mampala, located within Lubumbashi municipality (~5km2 and 55,000 inhabitants), was selected to represent a planned neighborhood. Despite its structured layout, it faces challenges of degradation and has only one green space [26]. Bel-Air, situated in Kampemba municipality (~5km2 and 65,000 inhabitants), exemplifies a residential area formerly reserved for Europeans. It retains relatively good conditions reflecting characteristics of older urban development. The neighborhood features some paved roads, moderately sized plots, moderate sanitation conditions, reliable electricity, minimal informal economic activities, easy accessibility, and several public green spaces, with an average to high standard of living among residents [43]. Lastly, Gambela III (~4km2 and 50,000 inhabitants), located in Lubumbashi municipality, was chosen to represent an unplanned neighborhood. It illustrates the challenges associated with informal urban expansion and lack of planning, characterized by low-income residents, significant informal economic activities, chaotic and unplanned constructions, inadequate public infrastructure, difficult accessibility, dusty conditions in dry seasons, and a lack of public green spaces. The living conditions in this area are often precarious due to anarchic urbanization [26].
For the selection of avenues within each neighborhood, a systematic approach was adopted to ensure a representative coverage of the city’s varied characteristics. Avenues were chosen based on their representativeness and accessibility [26]. The selected avenues were intended to reflect typical variations within residential, planned, and unplanned neighborhoods. Once the avenues were selected, specific plots within each avenue were chosen using stratified random sampling [16]. Selection criteria for the plots included diversity of garden types and accessibility for observations [15].
Data collection on each plot primarily involved interactions with the plot owner. If the owner was unavailable, the longest-residing tenant was consulted. This process ensured that the collected information was as accurate and representative as possible, providing a comprehensive view of gardening practices and floral characteristics in each studied area [15]. Field data collection was conducted over a period of three and a half months, from May 10, 2022, to August 28, 2022, using a pre-established field data sheet. Data were collected directly from the plots and households, preferably on Sunday afternoons to ensure the availability of household heads. This approach aimed to obtain precise information on the structure and function of domestic gardens.
For each neighborhood, a total of 20 avenues were randomly selected to ensure diverse coverage. Within each avenue, 5 plots were strategically chosen to represent different points along the avenue. Two plots were selected at the ends of the avenue, one at each end, while one plot was chosen at the center of the avenue, alternating between the left and right sides. This method captured variability in gardening practices and floral characteristics along the avenues [26]. Thus, for each neighborhood, 100 plots were explored, offering a comprehensive and detailed view of domestic gardens in various geographic and urban contexts. This approach ensured a balanced representation of different configurations and types of gardens within each neighborhood, allowing for an in-depth analysis of the collected data [15].
2.3. Data Collection
Domestic gardens were classified into five distinct types [16,44,45]: (i) flowerbeds (areas planted on the ground, often decorative), (ii) lawns (grass areas primarily used for leisure), (iii) pergolas (structures covered with climbing plants providing shade), (iv) hedges (vegetative barriers often used to delineate properties), and (v) Shruberry (clusters of shrubs and flowering plants, generally for ornamentation). This typology of domestic gardens provides insights into the functional and aesthetic roles these elements play within urban environments. Understanding these different types helps to assess how gardens contribute to biodiversity, microclimate regulation, and social well-being in urban areas [45]. The geometric shape of the gardens was assessed based on their configuration, such as rectangular, circular, or irregular, to better understand their spatial arrangement. Understanding the configuration helps in assessing how space is utilized within gardens, which can affect factors like the accessibility of green spaces for residents. Additionally, the shape and layout of gardens can reflect broader urban planning patterns and the socio-economic conditions of neighborhoods, as these factors often dictate how much space is available for gardening and how it is organized.
Regarding the floristic study, two types of data were collected: (1) plant species present in the domestic gardens, determined through careful observations and identification of collected specimens; and (2) residents’ practices and perceptions regarding this flora. For plant species nomenclature, available floras and specialized literature were used [46,47,48]. The origin status of species was determined by classifying them as exotic if they are not native to a specific geographic region, and as indigenous if they originate from the region (here Africa). The Afro-Asian species were considered as indigenous [47]. This approach distinguished species based on their origin and ecological impact, providing an overview of the floristic diversity of domestic gardens and their influence on the urban environment [49].
Species were also categorized into biological types based on Raunkiaer’s classification (1934), which has been adapted for tropical zones with a pronounced dry season [50]. This classification separates species into Phanerophytes (These are tall, woody plants, such as trees and large shrubs, with buds located high above the ground to survive unfavorable seasons), Chamaephytes (These are small, woody plants or dwarf shrubs, with buds situated close to the ground, allowing them to endure dry conditions), Hemicryptophytes (These plants have their perennating buds at the soil surface, often protected by soil or leaf litter, which is common in herbaceous species), Geophytes (These species survive adverse seasons with underground storage organs like bulbs, tubers, or rhizomes), Therophytes (Annual plants that complete their life cycle quickly and survive unfavorable periods as seeds), and Hydrophytes (Aquatic plants that grow in or near water, adapted to survive submerged or floating environments). This classification helps in understanding the ecological strategies of different plant species within the studied area [14]. Morphological types were distinguished according to Grime’s strategy (1975) into three main forms: herbaceous (non-woody stemmed plants), liana (climbing plants), and woody (including trees, shrubs, and bushes). The morphological types of plants, classified by Grime’s strategy, reflect how urban planning, socio-economic conditions, and the presence or absence of green spaces influence garden composition and biodiversity across different urban settings [51]. Species dispersal strategies—such as anthropochory (human-mediated), zoochory (animal-mediated), autochory (self-dispersal), hydrochory (water-mediated), and anemochory (wind-mediated)—was evaluated for understanding how plants spread and establish in domestic gardens [52]. This understanding helps identify the influence of human activity and environmental factors on plant diversity, aiding in conservation, sustainable garden management, and the resilience of urban green spaces [53,54].
Finally, the uses and functions of domestic gardens were analyzed by collecting information from residents about the goods and ecosystem services provided by the vegetation and the observed floristic composition. Domestic gardens are distinguished by four main uses: ornamental gardens, where plants are grown for aesthetic beauty, contributing to the enhancement of outdoor spaces and creating pleasant environments; food gardens, which provide edible products such as fruits, vegetables, and herbs, essential for the residents’ daily diet; cultural gardens, where certain species have specific cultural importance, used in rituals, celebrations, or local traditions; and medicinal gardens, where plants are cultivated for their medicinal properties, used in the preparation of traditional remedies for various ailments [47]. This approach highlighted the diversity of functions of domestic gardens and their multifunctional role in urban environments [55].
2.3. Data Analysis
Analysis of variance (ANOVA) were performed using R software (4.2), to compare the neighborhood types [26]. When significant differences between means were identified, a post-hoc Tukey test was applied to compare the means of each pair and determine significantly distinct groups. It should be noted that avenues were treated as repetitions in this analysis. The relative frequency of species was calculated to assess their prevalence within the gardens [16]. Additionally, species classification according to their frequency index was conducted using Caratini’s method (1985) [56], as detailed in Table 1. This method characterizes species based on their relative occurrence in the collected samples, providing a precise measure of their abundance and distribution across the different studied neighborhoods [57]. These statistical analyses offer insights into the variations in the floristic composition of domestic gardens and identify significant trends in the data.
3. Results
3.1. Plot Characteristics
The analysis of domestic gardens characteristics in Lubumbashi reveals significant differences between neighborhoods (Table 2). The residential neighborhood shows a higher average number of gardens (2.84 ± 0.6) and a larger average garden area (315.1 ± 336.2 m2) compared to the unplanned (2.4 ± 0.7 and 154.3 ± 106.6 m2) and planned neighborhoods (2.04 ± 1.31 and 42.8 ± 57.3 m2). This trend is also reflected in species richness, which is highest in the residential neighborhood (22 ± 8.272 species) compared to the unplanned (15.96 ± 5.852) and planned neighborhoods (13.355 ± 12.811). The average plot size is also larger in the residential neighborhood (1032.0 ± 205.5 m2), with a higher proportion of vegetation (31.10 ± 11.6%), contrasting with the unplanned (17.80 ± 10.8%) and planned neighborhoods (14.87 ± 19.0%). These results highlight that domestic gardens in the residential neighborhood exhibit larger sizes, greater floral richness, and higher vegetation proportions compared to those in the unplanned and planned neighborhoods, indicating a more extensive and diverse management of green spaces in this area.
3.2. Distribution of Domestic Garden Shapes, Typologies, and Utilizations across Different Neighborhood Types
Square-shaped gardens are relatively rare across all neighborhood types, with the highest occurrence in planned neighborhoods (2.2%) and the lowest in residential neighborhoods (0.7%). This scarcity indicates that square gardens are uncommon, especially in unplanned and residential areas (Table 3). In contrast, rectangular plots are more prevalent, particularly in unplanned neighborhoods (22.0%), followed by residential areas (19.0%). This suggests that rectangular shapes are better suited to the often irregular layouts of unplanned environments. Irregularly shaped plots dominate across all neighborhood types, with the highest percentage found in residential neighborhoods (80.3%). This predominance reflects a preference or necessity for irregular shapes, indicating flexibility in land use.
Pergolas are sparse across all neighborhoods, with planned neighborhoods showing the highest occurrence (2.9%). This rarity suggests that pergolas, likely due to their decorative and space-consuming nature, are not a common feature. Hedges are more frequently found in unplanned neighborhoods (16.5%), indicating their role as natural boundaries or privacy screens in less structured environments. Flowerbeds are a prominent feature in planned neighborhoods (43.1%), reflecting more intentional landscaping efforts, whereas their presence decreases in residential neighborhoods (31.7%), where other vegetation or landscaping types may be prioritized. Lawns are most common in residential neighborhoods (19.0%), likely due to cultural preferences for green open spaces around homes. Conversely, they are less common in planned (6.1%) and unplanned neighborhoods (11.0%). Shrubberies are fairly consistent across all neighborhood types, with a slight dominance in planned neighborhoods (38.0%), suggesting their popularity as a versatile landscaping option (Table 3).
The use of domestic gardens for food production (alimentation) is significant, particularly in planned neighborhoods (40.7%), highlighting a strong emphasis on food security or self-sufficiency. Ornamentation is most prevalent in residential neighborhoods (51.4%), suggesting that aesthetic considerations are more emphasized in these areas. Cultural uses of gardens are relatively low across all neighborhood types, with a slight increase in residential neighborhoods (4.9%), reflecting the incorporation of cultural elements in private spaces. Medicinal plants are most common in unplanned neighborhoods (23.6%), possibly due to limited access to formal healthcare, leading to a reliance on traditional remedies (Table 3).
Overall, the data reveal that planned neighborhoods exhibit a more structured approach, with a focus on food production and ornamentation, while unplanned neighborhoods tend to prioritize practical features like hedges and medicinal plants. Residential areas, on the other hand, place greater emphasis on lawns and ornamentation, reflecting a focus on aesthetics and recreational spaces.
3.2. Flora Analysis
The inventory of domestic gardens in Lubumbashi reveals significant floral diversity varying by neighborhood type. The study identified 232 species across 68 families, with Araceae, Euphorbiaceae, Asteraceae, and Solanaceae being the most prominent. Notable differences among neighborhoods include 169 species in the unplanned area, 181 in the residential area, and 209 in the planned area. Frequency indices show that rare or incidental species dominate all neighborhoods: 94.47% in the planned area, 89.35% in the unplanned area, and 80.66% in the residential area. Accessory species are also significant, representing 10.06% in the unplanned area, 4.31% in the planned area, and 16.57% in the residential area. Frequent and constant species are less represented, with no constant species found in the unplanned area. The data reveals that Mangifera indica and Persea americana are abundant in all neighborhoods, illustrating their adaptability to different urban contexts. The distribution of other species varies by neighborhood: the planned area shows a high presence of woody and rare species, while the unplanned area is characterized by a high proportion of rare or incidental species. The residential area stands out for its richness in herbaceous plants and greater diversity of frequent species. These results illustrate how garden management practices and environmental conditions influence floral diversity. The prevalence of species such as Mangifera indica and Persea americana, along with the diversity of morphological types, provides valuable insights into plant preferences and ecological dynamics within Lubumbashi (See Appendix).
The analysis of domestic gardens in the studied neighborhoods reveals a marked predominance of exotic species over native ones, with at least 80% of the species being exotic. This dominance is paradoxical given the potential ecological benefits of native species for local biodiversity. The distribution of exotic species is remarkably consistent across neighborhoods, with minor variations. The unplanned neighborhood has the highest proportion of exotic species at 82.25%, which may be attributed to gardening practices that favor the introduction of non-native species. In contrast, the residential area has a slightly higher proportion of native species (~20%), which may be related to the larger plot sizes that facilitate the spontaneous establishment and survival of native species (Figure 2). The dominance of exotic species in Lubumbashi’s domestic gardens, despite their homogeneous distribution, raises questions about the long-term impact of these species on local biodiversity, especially in areas where native species have a better chance of establishing themselves.
3.3. Morphological, Biological, and Propagation Characteristics of Domestic Gardens Species across Different Neighborhood Types
The analysis of the morphological types of flora in domestic gardens in the study area shows a clear predominance of herbaceous species across all neighborhoods. These species make up more than half of the floral composition: 57.40% in the unplanned neighborhood, 61.72% in the planned neighborhood, and 60.22% in the residential neighborhood. Woody species are the second most common, with proportions of 38.87% in the unplanned area, 33.97% in the planned area, and 35.36% in the residential area. In contrast, vines are sparsely represented in these gardens, not exceeding 4% in any neighborhood (Table 4). These results highlight the dominance of herbaceous species, likely due to their adaptability and their role in the aesthetics and functionality of gardens. The modest presence of vines may be explained by garden management practices that favor other types of vegetation.
The floral composition of domestic gardens in the three studied neighborhoods is predominantly characterized by therophytes and phanerophytes, reflecting the presence of annual plants and woody species adapted to varying conditions. Geophytes, while secondary, also play a significant role in contributing to the diversity of species. In contrast, hydrophytes and epiphytes are nearly absent, their rarity suggesting environmental conditions or gardening practices that are not conducive to their development (Table 4). These observations highlight a planting strategy focused on more robust species suited to urban environments, leaving little space for species that require more specific conditions.
The analysis results indicate a clear predominance of diaspore dispersal by humans (anthropochory) in the three studied neighborhoods. This phenomenon is particularly pronounced in the planned neighborhood, where 87% of species are dispersed this way, followed closely by the unplanned neighborhood with 86% and the residential neighborhood with 85.05% (Table 4). Anemochory, or wind dispersal, ranks second but at significantly lower levels. Other dispersal modes, such as zoochory or hydrochory, are virtually absent, reflecting a strong human influence on the composition and floral dynamics of domestic gardens in these neighborhoods. This finding underscores the considerable impact of human activities on urban flora, favoring species that can adapt to gardening practices and urban conditions.
4. Discussion
4.1. Methodological Limitations
Selecting only one neighborhood per neighborhood type among more than 40 neighborhoods in the city presents several potential limitations. Specifically, these limitations relate to representativeness, intra-typological variability, and result generalization. However, the chosen neighborhoods were selected based on rigorous criteria to maximize their representativeness for each type [58]. This included preliminary analyses to ensure that the selected neighborhoods are close to the average characteristics of the type. Subsequently, secondary data were used to verify whether the trends observed in the selected neighborhood correspond to those in other neighborhoods of the same type [43]. By following these precautions, selecting a single neighborhood per type does not compromise the validity of the results, provided that the neighborhood is truly representative [59]. Comparative analysis with other studies and cross-validation help reinforce the reliability of the conclusions. Moreover, this approach allows for a more detailed and in-depth analysis of domestic garden characteristics, providing specific insights while limiting biases [31,60].
Collecting data only during the dry season presents certain limitations for a complete understanding of the ecological dynamics of domestic gardens. Floristic diversity and garden structure can vary significantly between seasons. Some species may be less visible or dormant during the dry season, which could lead to an underestimation of species diversity and plant cover. By collecting data only during the dry season, there is a risk of missing important observations regarding plant reproduction and dispersal. However, in domestic gardens, plants are often regularly watered, which mitigates the effects of the dry season [61]. Irrigation supports the growth and blooming of many species, making the data representative of plant composition and garden management practices. Plants in domestic gardens are often selected for their drought resistance or ability to thrive in low-humidity conditions. Also, many domestic garden plants, particularly trees and shrubs, are perennial species whose presence and dominance do not vary considerably between seasons [32]. Therefore, the collected data remains reliable for assessing garden structure and composition.
4.2. Shape and Types of Domestic Garden in Relation to the Land Use Planning
The analysis of data regarding the presence or absence of domestic gardens on plots in the studied neighborhoods reveals a marked trend: the vast majority of plots, approximately 90%, feature domestic gardens. However, this proportion is particularly high in residential neighborhoods compared to non-planned and planned neighborhoods. This observation can be attributed to the larger plot sizes in residential areas, which offer more space for garden establishment. This hypothesis is supported by the Groupe Huit [43] report on the urban planning reference for Lubumbashi, which notes that, in such neighborhoods, the building coverage rarely exceeds half of the total plot area.
The observation that domestic gardens in residential neighborhoods exhibit larger sizes, greater floral richness, and higher vegetation proportions compared to those in unplanned and planned neighborhoods can be attributed also to socio-economic status [15]. Higher socio-economic status in residential areas often correlates with larger land areas and increased disposable income, enabling residents to invest more in their gardens [62]. For instance, affluent neighborhoods in Limpompo province, South Africa, benefit from extensive properties that allow for diverse and extensive domestic gardens [32]. Cultural and aesthetic preferences also play a significant role [63,64]. In cities like Akure, Nigeria, residents in higher socio-economic neighborhoods place a strong emphasis on aesthetic and recreational spaces, leading to more elaborate and varied garden designs [65]. Similar trends were observed in Kinshasa, DR Congo [34]. Urban planning and development constraints further explain these trends [44]. Planned neighborhoods often face stricter zoning regulations that limit garden size and diversity [66]. Conversely, unplanned areas may have bigger plots, resulting in more extensive green spaces. Economic investment in green spaces is another critical factor. In Limpompo and Nort West Provinces in South Africa, residential areas with significant economic resources see enhanced investment in landscaping and garden maintenance, contributing to larger and more diverse gardens [67]. Lastly, community engagement and social practices influence garden management [68]. In Kinshasa, Lubumbashi and Kolwezi, DR Congo, active community involvement often leads to better vegetation care and investment, promoting larger and more diverse green spaces [26,34].
Plot owners often utilize the vacant space to create domestic gardens for various purposes. This correlation between plot size and the presence of domestic gardens is also observed in other studies. For instance, Lubbe et al. [31] demonstrated that plot size directly influences plant diversity and the number of gardens a plot can accommodate. Larger plots are better able to support a variety of plants and multiple types of gardens. This relationship aligns with the findings of Muratet [69] on the vegetation of abandoned lands in the Hauts-de-Seine department, which showed that species richness is largely determined by the area of the land. Thus, plot size affects not only the presence of gardens but also their typology and configuration.
Regarding domestic garden typology, our results indicate a specific trend: in smaller plots, typical of planned neighborhoods, owners tend to create more compact gardens, such as flowerbeds, hedges, and pergolas. This tendency can be explained by the need to optimize the limited available space [70]. In contrast, in residential neighborhoods, where plots are larger, lawns are more common [71]. This is because larger areas allow for the installation of extensive lawns, which require more space [72]. Flowerbeds, on the other hand, are evenly distributed across all sampled neighborhoods, regardless of plot size. Flowerbeds primarily consist of trees, which, due to their height, can coexist with other activities on the plot without disrupting the overall layout. This observation is supported by Ngur-Ikone [73], who noted that flowerbeds are often preferred for their ability to provide shade, serve as windbreaks, and produce fruit, making them particularly attractive to homeowners.
Most domestic gardens have irregular forms, which can be attributed to their placement in non-conventional interstitial spaces. This observation aligns with the findings of Ngur-Ikone [73] and Cameron et al. [74], who also noted a predominance of irregular shapes in urban gardens, often due to the need to adapt gardens to residual spaces or constraints imposed by urban planning. The results show that larger spaces in residential neighborhoods not only support a higher density of gardens but also a greater diversity of garden types, whereas smaller spaces in planned neighborhoods lead to more compact and optimized configurations [75]. Despite the predominance of irregular shapes, reflecting spatial constraints, domestic gardens play a central role in plot design, demonstrating an effective adaptation to urban realities and residents’ needs [76].
4.3. Flora and Uses of Domestic Gardens
The analysis of the flora in domestic gardens within the studied neighborhoods highlights a complex relationship between the level of urban planning and plant species richness. The results indicate significant variation in floral diversity among the gardens, suggesting that socio-economic and historical factors strongly influence the observed plant composition [14]. Firstly, the species richness of domestic gardens appears to be correlated with the degree of neighborhood planning. This observation is supported by the work of Lubbe et al. [31], who indicate that the socio-economic characteristics of residents, as well as their standard of living, play a crucial role in garden plant diversity. For instance, older or historically established neighborhoods, such as those dating back to the colonial era, exhibit greater species diversity compared to more recently developed areas. This trend is corroborated by Makumbelo et al. [77], who observed similar variations in gardens in Kinshasa.
Another factor influencing plant richness is the historical impact on vegetation. In planned neighborhood, although also affected by initial deforestation, has recorded greater species diversity. This can be attributed to the active efforts of residents to reintroduce diverse species into their gardens after deforestation and the toxic atmospheric fallout from mining activities. Research by Shutcha et al. [78] emphasizes that harsh environmental conditions have driven residents of planned neighborhood to seek a diverse vegetation that can survive in altered conditions. The abundance of the herbaceous layer in domestic gardens is also significant. This predominance is related to the major use of annual species, as shown by Bernholt et al. [28]. The high proportion of therophytes observed in all studied neighborhoods can be explained by the high density of buildings and the limited space available for gardens. This observation is reinforced by Marco et al. [79] in high-density built-up areas, such as Lauris, where a similar trend was identified.
Finally, specific species such as Mangifera indica (mango) and Persea americana (avocado) are particularly common in domestic gardens in the study area. These trees provide various urban services, including shading, fruit production, and economic benefits for households. The results are consistent with studies by Makumbelo et al. [77,80] in Kinshasa, which observed similar trends in the use of plant species in urban environments. The floral diversity of domestic gardens in the studied neighborhoods is the result of a complex interplay between urban planning, historical landscaping practices, and residents’ socio-economic needs.
However, it is noteworthy that planned neighborhood, a newly developed neighborhood, shows lower floral diversity compared to residential and unplanned neighborhoods, despite the latter being established earlier. This difference might be attributed to urban planning practices that often involve the removal of existing vegetation to be replaced by exotic species. This practice aligns with findings on species origin status, where exotic species dominate all studied gardens, with proportions ranging from 80.11% to 82.25% depending on the neighborhood. This dominance of exotic species is also observed in other African cities, as highlighted by Bernholt et al. [28], Marco et al. [79], and Bigirimana et al. [47]. The reasons for this dominance include the intentional introduction of plants for ornamental, cultural, and economic purposes [48].
The dominance of exotic plants can lead to a reduction in local biodiversity, as these species often outcompete native plants that are crucial for supporting local wildlife and maintaining ecosystem functions [13]. For example, in Boston, USA, the widespread planting of exotic species has resulted in a decline in native flora, negatively impacting local fauna that rely on native plants [81]. Additionally, exotic species may disrupt essential ecosystem services provided by native plants, such as soil stabilization, water filtration, and pollination [82]. In Paris, France, the introduction of exotic plants in urban green spaces has disrupted local pollinator networks, as native pollinators depend on native plants for food and habitat [83]. Moreover, exotic plants can alter soil chemistry and habitat conditions, making them less suitable for native species [84]. Lastly, the dominance of exotic species can erode cultural and ecological heritage associated with native plants [85]. In central Europe, the replacement of native plants with exotic species in home gardens has led to the loss of traditional knowledge and cultural practices associated to native flora [86].
Cameron et al. [74] emphasize that domestic gardens play a crucial role in urban vegetation, often designed to meet specific needs of their owners. This perspective is reinforced by Loram et al. [87], which asserts that the presence of plant species in a domestic garden is significantly influenced by socio-economic aspects and personal habits, including nutritional, aesthetic, medicinal, and cultural needs. Our results align with these observations. In the studied neighborhoods, the functions of domestic gardens vary according to the socio-economic context. Planned neighborhoods are often designed with a comprehensive master plan that includes designated areas for various purposes. In 2022, the Human Development Index (HDI) of the DRC was 0.479, placing the country in the "low human development" category and ranking it 180th out of 189 countries and territories. Additionally, a large portion of the population lives on less than $1.25 per day and typically holds less stable jobs [88]. To meet health needs and support human and livestock nutrition, various socio-economically important plants are cultivated in domestic gardens. In the city of Lubumbashi, the species richness in neighborhoods is high, including many native species crucial for local biodiversity conservation. These native species also provide several ecosystem services to residents and surrounding populations. The high frequency of fruit tree species such as Mangifera indica (confirmed by previous study, i.e. Useni et al. [26]) and Carica papaya may result from human preference, due to the ecosystem services these species offer, including shade, edible fruits, and ornamental flowers. Additionally, their leaves and bark are used in traditional medicine, which enhances their popularity despite being introduced species [89]. Although alien weeds like Tithonia diversifolia can negatively impact native biodiversity and agricultural productivity, they also offer significant socio-economic benefits [90]. These gardens also serve for cultivating vegetable species and even cereals such as maize for human consumption. Herbaceous plants like Nicotiana tabacum are grown not only for human consumption but also as pesticides for other crops and for use in livestock management [91]. The cultivation of such diverse species underscores the multifunctional value of these gardens, contributing to food security and sustainable agricultural practices within the community. For example, in Bujumbura, Burundi, urban planning promotes structured use of garden space for both food production and ornamentation [15]. This approach reflects the intentional design of these areas to balance aesthetic and practical functions.
Unfortunately, the consumption of vegetables from urban gardens poses a serious health risk due to high concentrations of trace metals found in these products, in the mining context of Lubumbashi. Studies have shown that vegetables grown in contaminated soils can contain dangerous levels of Copper, Cobalt, and Cadmium, adversely affecting consumer health [92,93]. These metals can cause various health issues, including neurological disorders and kidney diseases [94]. To mitigate this risk, bioponics, a soilless cultivation method, is currently being tested as an alternative [95]. By using nutrient substrates and hydroponic solutions, bioponics allows for better control of growing conditions and reduces the risk of contamination. In contrast, unplanned neighborhoods typically lack formal urban planning, leading to a focus on immediate practical needs [96]. For instance, in informal settlements in Niamey, Niger, gardens often feature practical elements such as hedges for privacy and medicinal plants [28]. Residents prioritize these features due to the absence of structured planning and the necessity to address everyday needs with available resources [97]. In residential areas, especially in more affluent neighborhoods, there is a greater emphasis on aesthetics and recreational spaces. For example, in upscale neighborhoods like those in Niamey (Niger) and Cape Town (South Africa), residents invest in well-maintained lawns and ornamental gardens [98,99]. This trend is driven by higher socio-economic status and a lifestyle that values visual appeal and functional recreational spaces [100].
The vegetation in Lubumbashi’s domestic gardens includes some native species; however, these species face a high risk of local extinction due to various factors. Phanerophytes are notably scarce and less frequent in the city’s vegetation. Their long-term survival is jeopardized by the combined effects of isolation and insufficient population size [101]. Most therophytes, chamaephytes, and ruderal species are found in gardens that are frequently repurposed for new construction. As vegetated lands are progressively destroyed, even short-lived or ruderal species, whether native or alien weeds, face a significant risk of local extinction. Ornamental species also pose a potential threat to biodiversity. Despite comprising a small proportion of naturalized flora, escaped ornamental plants are among the most invasive in the city, with many others posing high invasion risks [42]. The increasing density of properties, driven by population growth and reduced availability of new land for development, further exacerbates the decline in plant species diversity. Additionally, the allocation of new properties often results in the destruction of public green spaces that previously supported diverse cultivated plants.
4.4. Socio-Ecological Implications
Gardens are dominated by herbaceous plants, with high proportions (57.40% in unplanned neighborhood, 61.72% in planned neighborhood, and 60.22% in residential neighborhood). These fast-growing, short-lived plants can influence soil structure and nutrient cycling differently from woody species, which play crucial roles in carbon sequestration and soil stabilization [102]. Their low presence in gardens may limit long-term ecological benefits such as erosion reduction and carbon storage. The predominant anthropogenic dissemination (87% in planned neighborhood) indicates that human practices strongly influence the floristic composition of gardens, potentially reducing ecological resilience and altering local ecosystem functions [48]. To reverse this trend, promoting native plant species, enhancing public awareness of ecological gardening practices, and encouraging natural dispersal methods (zoochory, anemochory) are essential [103]. Implementing community-led garden projects and providing incentives for sustainable landscaping can also help restore biodiversity and strengthen the ecological resilience of urban gardens [104].
Domestic gardens, particularly in unplanned neighborhood, play a critical role in food security for residents. The food and medicinal gardens in this neighborhood provide essential sources of food and medicine, especially in the context of limited incomes. For example, urban vegetable gardens like those observed in residential neighborhood allow low-income families to supplement their diets and diversify their nutritional sources. In residential neighborhood, gardens are more oriented towards environmental and aesthetic functions. The presence of lawns and flowerbeds contributes to improving residents’ quality of life by providing attractive and functional green spaces. These green areas support recreational and social activities, enhance community cohesion, and can increase the real estate value of neighboring properties. To improve this situation, integrating food production with environmental and aesthetic functions in all neighborhoods is key [105]. Encouraging the cultivation of both edible and ornamental plants, supporting community garden initiatives, and providing resources for sustainable gardening can enhance food security, improve quality of life, and strengthen community bonds across urban areas [106].
The challenges faced by both (un)planned and residential neighborhoods in Lubumbashi have significant implications for the persistence and functionality of domestic gardens. In unplanned neighborhoods, where plots are often fragmented and sold to different owners, there is a notable risk of reducing the surface area available for gardens or even the complete disappearance of these spaces. This fragmentation can lead to a lack of continuity and coherence in the urban green space, affecting the ecological and aesthetic value of the area. On the other hand, in planned neighborhoods, the increasing density of construction poses a long-term threat to the sustainability of gardens. As the demand for more housing and commercial spaces grows, the pressure to convert green spaces into built environments intensifies, potentially leading to the loss of existing gardens. Additionally, the sale of plots to external investors who frequently demolish existing structures and vegetation for new developments—such as apartment buildings, commercial centers, or service stations—further exacerbates this issue. Such practices undermine the role of gardens in urban sustainability and highlight the need for integrated urban planning strategies that balance development with the preservation of green spaces. To reverse this trend, implementing urban planning policies that protect and promote domestic gardens is essential [107]. Encouraging green space preservation, limiting plot fragmentation, and integrating gardens into new developments can help maintain urban biodiversity, enhance ecological resilience, and ensure the long-term sustainability of green spaces in Lubumbashi [108].
5. Conclusion
The objective of this study was to assess the spatial structure, plant diversity, propagation strategies, and functions of domestic gardens in three Lubumbashi neighborhoods, chosen based on their land use planning levels. The findings reveal significant differences in garden structure across neighborhoods, with planned areas having more organized layouts, while unplanned neighborhoods exhibit varied configurations. Residential neighborhoods, however, stand out with larger average garden sizes (315.1 m2), higher species richness (22 species), and bigger plot sizes (1032 m2), compared to both unplanned and planned areas where gardens are smaller and less diverse. In unplanned neighborhoods, rectangular gardens dominate, whereas planned neighborhoods feature more deliberate landscaping elements such as flowerbeds and hedges. Notably, plant diversity peaks in unplanned areas, where 232 species across 68 families were identified, with exotic species comprising 80% of the flora, particularly high in these areas (82.25%). Common species like Mangifera indica and Persea americana are found across all neighborhoods, demonstrating adaptability to urban environments. Unplanned neighborhoods show gardens with more diverse functions, including food, medicinal, and cultural uses, reflecting the lack of urban planning and more challenging socio-economic conditions. Planned areas primarily use gardens for food production (40.7%), while residential neighborhoods emphasize ornamentation (51.4%). Herbaceous species are most prevalent, followed by woody plants, with vines being rare. Human activities (anthropochory) heavily influence species dispersal, accounting for over 85% in all neighborhoods. Despite the study being limited to the dry season and focusing on one neighborhood per type, the results highlight the importance of domestic gardens for urban biodiversity and food security. The findings underscore the predominance of exotic species and anthropogenic influence on plant dispersal. The data suggest that urban management policies should promote local plant diversity and sustainable gardening practices. Additionally, the predominance of gardens for food and ornamentation calls for supporting gardeners to enhance ecological resilience and sustain urban green spaces.
Author Contributions
Y.U.S.: conceptualisation, methodology and writing—original draft preparation; BKK & M.M.M.: methodology, writing—original draft preparation and data curation; J.Y.M, A.M.K & F.M.: writing—review and editing; J.B.: supervision, writing—original draft and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by Academy for Research and Higher Education, Commission for Development Cooperation (Belgium).
Data Availability Statement
The authors confirm that all data supporting the findings of this study are available within the article.
Acknowledgments
The authors would like to thank the development research project "Capacity building for the sustainable management of the miombo clear forest through the assessment of the environmental impact of charcoal production and the improvement of forest resource management practices (CHARLU)" and the administrative services of Lubumbashi city.
Conflicts of Interest
The authors have no conflicts of interest to declare. All co-authors have reviewed and approved the contents of the manuscript, and there are no financial interests to report. We confirm that the submission represents original work and is not currently under review by any other publication.
Appendix A
List of species recorded in domestic gardens of three neighborhoods in Lubumbashi with indications of their morphological type (MT: Herb = Herbaceous, Lign = Woody), propagation strategy (PS: Anthr = Anthropochory, Anem = Anemochory, Zooc = Zoochory, Hydr = Hydrochory, Auto = Autochory), origin status (OS: N = Native, E = Exotic), and absolute frequency (the dash (-) indicates absence of the species in the neighborhood). UN= Unplanned neighborhood; RN=Residential neighborhood; PN=Planned Neighborhood.
| Family | Species | MT | PS | OS | Absolute frequency | ||
| UN | RN | PN | |||||
| Acanthaceae | Justicia brandegeeana Wassh. & L.B.Sm. | Herb | Anthr | E | - | - | 0,01 |
| Justicia secunda Vahl | Herb | Anthr | E | 0,10 | 0,06 | 0,10 | |
| Megaskepasma erythrochlamys Lindau | Herb | Anthr | E | - | - | 0,01 | |
| Odontonema strictum (Nees) Kuntze | Herb | Anthr | E | 0,14 | 0,16 | 0,11 | |
| Pseuderanthemum atropurpureum (W.Bull) Radlk. | Lign | Anthr | E | 0,02 | - | 0,01 | |
| Ruellia simplex C.Wright | Herb | Anthr | E | 0,08 | 0,20 | 0,13 | |
| Sanchezia speciosa Leonard | Lign | Anthr | E | 0,04 | 0,20 | 0,05 | |
| Agavaceae | Agave americana L. | Herb | Anthr | E | 0,02 | - | 0,03 |
| Agave sisalana Perrine | Herb | Anthr | E | 0,04 | 0,02 | 0,04 | |
| Yucca acuminata Sweet | Herb | Anthr | E | - | 0,04 | 0,01 | |
| Amaranthaceae | Alternanthera bettzickiana (Regel) G.Nicholson | Herb | Anthr | E | 0,12 | 0,22 | 0,15 |
| Alternanthera brasiliana (L.) Kuntze | Herb | Anthr | E | 0,12 | 0,14 | 0,10 | |
| Amaranthus hybridus L. | Herb | Anthr | E | 0,14 | 0,18 | 0,13 | |
| Amaranthus spinosus L. | Herb | Aném | N | 0,14 | 0,24 | 0,14 | |
| Celosia argentea L. | Herb | Anthr | E | 0,04 | - | - | |
| Celosia trigyna L. | Herb | Aném | N | 0,04 | - | 0,16 | |
| Iresine herbstii Hook. | Herb | Anthr | E | 0,06 | 0,08 | 0,03 | |
| Amaryllidaceae | Hymenocallis littoralis (Jacq.) Salisb. | Herb | Anthr | E | 0,08 | 0,20 | 0,14 |
| Zephyranthes candida (Lindl.) Herb. | Herb | Anthr | E | - | 0,06 | 0,03 | |
| Anacardiaceae | Mangifera indica L. | Lign | Anthr | E | 0,52 | 0,68 | 0,69 |
| Annonaceae | Annona muricata L. | Lign | Anthr | E | 0,04 | 0,06 | 0,05 |
| Apocynaceae | Catharanthus roseus (L.) G.Don | Herb | Anthr | E | 0,16 | 0,14 | 0,09 |
| Nerium oleander L. | Lign | Anthr | E | 0,02 | 0,06 | 0,01 | |
| Plumeria alba L. | Lign | Anthr | E | 0,04 | - | 0,01 | |
| Plumeria rubra L. | Lign | Anthr | E | 0,02 | 0,02 | 0,01 | |
| Thevetia peruviana (Pers.) K.Schum. | Lign | Anthr | E | 0,02 | - | 0,04 | |
| Araceae | Aglaonema commutatum Schott | Herb | Anthr | E | - | 0,02 | 0,05 |
| Alocasia macrorrhizos (L.) G.Don | Herb | Anthr | E | 0,18 | 0,08 | 0,06 | |
| Alocasia sp | Herb | Anthr | E | 0,08 | 0,12 | 0,09 | |
| Caladium bicolor (Aiton) Vent. | Herb | Anthr | E | 0,10 | 0,16 | 0,05 | |
| Caladium lowii Lem. | Herb | Anthr | E | 0,06 | 0,10 | 0,01 | |
| Caladium sp | Liane | Anthr | E | 0,22 | 0,34 | 0,29 | |
| Caladium lindenii (André) Madison | Herb | Anthr | E | 0,06 | 0,04 | 0,02 | |
| Colocasia esculenta (L.) Schott | Herb | Anthr | E | 0,04 | - | 0,07 | |
| Dieffenbachia amoena Bull. | Herb | Anthr | E | 0,20 | 0,24 | 0,12 | |
| Epipremnum pinnatum (L.) Engl. | Liane | Anthr | E | 0,08 | 0,12 | 0,05 | |
| Eucharis × grandiflora Planch. & Linden | Herb | Anthr | E | 0,08 | 0,16 | 0,04 | |
| Monstera deliciosa Liebm. | Herb | Anthr | E | 0,12 | 0,12 | 0,05 | |
| Philodendron speciosum Schott ex Endl. | Herb | Anthr | E | 0,04 | 0,06 | 0,03 | |
| Philodendron giganteum Schott | Herb | Anthr | E | 0,12 | 0,10 | 0,10 | |
| Philodendron lacerum (Jacq.) Schott | Herb | Anthr | E | - | 0,04 | 0,06 | |
| Philodendron xanadu Croat, Mayo & J.Boos | Herb | Anthr | E | 0,02 | - | 0,04 | |
| Spathiphyllum sp | Herb | Anthr | E | 0,02 | 0,08 | 0,03 | |
| Syngonium auritum (L.) Schott | Herb | Anthr | E | 0,06 | 0,06 | 0,07 | |
| Xanthosoma sagittifolium (L.) Schott | Herb | Anthr | E | 0,02 | 0,18 | 0,07 | |
| Araliaceae | Polyscias balfouriana (André) L.H.Bailey | Lign | Anthr | E | - | 0,08 | 0,05 |
| Arecaceae | Archontophoenix alexandrae (F.Muell.) H.Wendl. & Drude | Lign | Anthr | E | - | 0,02 | - |
| Borassus aethiopum Mart. | Lign | Anthr | E | 0,02 | 0,04 | 0,01 | |
| Chrysalidocarpus lutescens H.Wendl. | Lign | Anthr | E | 0,01 | 0,02 | 0,01 | |
| Cycas revoluta Thunb. | Lign | Anthr | E | - | 0,02 | 0,01 | |
| Elaeis guineensis Jacq. | Lign | Anthr | E | 0,22 | 0,30 | 0,14 | |
| Phoenix dactylifera L. | Lign | Anthr | E | 0,06 | 0,08 | 0,01 | |
| Asparagaceae | Asparagus setaceus (Kunth) Jessop | Herb | Anthr | E | 0,04 | 0,02 | - |
| Dracaena reflexa Lam. | Lign | Anthr | N | - | 0,08 | 0,05 | |
| Ledebouria apertiflora (Baker) Jessop | Herb | Anthr | E | - | 0,02 | - | |
| Sansevieria hyacinthoides (L.) Druce | Herb | Anthr | E | 0,18 | 0,12 | 0,17 | |
| Asphodelaceae | Aloe striata Haw. | Herb | Anthr | E | 0,02 | 0,06 | - |
| Aloe vera (L.) Burm.f. | Herb | Anthr | E | 0,34 | 0,46 | 0,33 | |
| Asteraceae | Ageratum albidum (DC.) Hemsl. | Herb | Anem | N | 0,16 | 0,14 | 0,04 |
| Ageratum conyzoides (L.) L. | Herb | Anem | N | 0,20 | 0,22 | 0,05 | |
| Bidens pilosa L. | Herb | Anthr | N | 0,04 | 0,12 | 0,04 | |
| Calea urticifolia (Mill.) DC. | Lign | Anthr | E | 0,04 | 0,04 | 0,01 | |
| Conyza pyrrhopappa Sch.Bip. ex A.Rich. | Herb | Anem | N | 0,06 | 0,04 | 0,07 | |
| Coreopsis lanceolata L. | Herb | Anthr | E | - | 0,04 | 0,02 | |
| Cosmos langlassei (Sherff) Sherff | Herb | Anthr | E | - | - | 0,03 | |
| Galinsoga parviflora Cav. | Herb | Anem | N | 0,06 | 0,20 | 0,02 | |
| Lactuca serriola L. | Herb | Anem | N | 0,08 | 0,04 | 0,06 | |
| Leucanthemum vulgare (Vaill.) Lam. | Herb | Anthr | E | - | - | 0,02 | |
| Sonchus arvensis L. | Herb | Aném | N | - | 0,06 | - | |
| Tagetes patula L. | Herb | Anthr | E | - | - | 0,05 | |
| Taraxacum sp | Herb | Anem | N | 0,04 | 0,04 | 0,06 | |
| Tithonia diversifolia (Hemsl.) A.Gray | Lign | Anthr | E | 0,02 | - | 0,01 | |
| Wedelia trilobata A.St.-Hil. | Herb | Anthr | E | 0,08 | 0,06 | 0,08 | |
| Zinnia elegans L. | Herb | Anthr | E | - | - | 0,01 | |
| Basellaceae | Basella alba L. | Herb | Anthr | E | 0,02 | 0,14 | 0,05 |
| Begoniaceae | Begonia rex Putz. | Herb | Anthr | E | - | 0,12 | 0,02 |
| Begonia sp | Herb | Anthr | E | 0,02 | 0,14 | 0,04 | |
| Bignoniaceae | Jacaranda mimosifolia D.Don | Lign | Anthr | N | - | 0,02 | - |
| Brassicaceae | Brassica chinensis L. | Herb | Anthr | E | 0,06 | 0,04 | 0,03 |
| Brassica juncea (L.) Czern. | Herb | Anthr | E | 0,01 | 0,02 | 0,01 | |
| Bromeliaceae | Ananas comosus (L.) Merr. | Herb | Anthr | E | - | - | 0,05 |
| Cactaceae | Opuntia ficus-indica (L.) Mill. | Lign | Anthr | E | - | - | 0,02 |
| Opuntia sp | Lign | Anthr | E | 0,02 | 0,06 | 0,01 | |
| Cannabaceae | Cannabis sativa L. | Herb | Anthr | E | 0,02 | 0,02 | 0,01 |
| Cannaceae | Canna indica L. | Herb | Anthr | E | 0,14 | 0,26 | 0,14 |
| Caricaceae | Carica papaya L. | Lign | Anthr | E | 0,26 | 0,38 | 0,28 |
| Caryophyllaceae | Dianthus carthusianorum L. | Herb | Anthr | E | 0,02 | 0,06 | 0,01 |
| Chenopodiaceae | Chenopodium ambrosioides L. | Herb | Anem | N | 0,08 | 0,12 | 0,07 |
| Cleomaceae | Gynandropsis gynandra (L.) Briq. | Herb | Anem | N | 0,02 | - | 0,02 |
| Clusiaceae | Garcinia huillensis Welw. | Lign | Anthr | N | - | 0,02 | - |
| Combretaceae | Quisqualis indica L. | Lign | Anthr | E | - | - | 0,01 |
| Terminalia mantaly H.Perrier | Lign | Anthr | E | 0,02 | 0,06 | 0,01 | |
| Commelinaceae | Callisia fragrans (Lindl.) Woodson | Herb | Anthr | E | - | 0,02 | 0,06 |
| Callisia repens (Jacq.) L. | Herb | Anthr | E | 0,10 | 0,12 | 0,08 | |
| Commelina diffusa Burm.f. | Herb | Anthr | N | 0,06 | 0,16 | 0,03 | |
| Tradescantia pallida (Rose) D.R.Hunt | Herb | Anthr | E | 0,10 | 0,20 | 0,05 | |
| Tradescantia spathacea Sw. | Herb | Anthr | E | 0,06 | 0,16 | 0,08 | |
| Tradescantia zebrina Bosse | Herb | Anthr | E | 0,14 | 0,24 | 0,18 | |
| Convolvulaceae | Ipomoea batatas (L.) Lam. | Liane | Anthr | E | 0,38 | 0,40 | 0,31 |
| Ipomoea cairica (L.) Sweet | Liane | Anthr | N | 0,02 | 0,10 | 0,06 | |
| Ipomoea fistulosa Mart. ex Choisy | Lign | Anthr | E | 0,04 | - | 0,02 | |
| Costaceae | Costus sp | Herb | Anthr | E | - | 0,02 | 0,06 |
| Crassulaceae | Bryophyllum daigremontianum (Raym.-Hamet & Perrier) A.Berger | Herb | Anthr | E | 0,12 | 0,18 | 0,14 |
| Bryophyllum pinnatum (Lam.) Oken | Herb | Anthr | E | 0,14 | 0,24 | 0,16 | |
| Cucurbitaceae | Citrullus lanatus (Thunb.) Matsum. & Nakai | Liane | Anthr | E | - | - | 0,01 |
| Cucurbita moschata Duchesne | Liane | Anthr | E | 0,14 | 0,12 | 0,14 | |
| Luffa acutangula (L.) Roxb. | Liane | Anthr | E | - | 0,02 | 0,02 | |
| Cyperaceae | Cyperus involucratus Rottb. | Herb | Auto | N | - | 0,08 | 0,04 |
| Dioscoreaceae | Dioscorea alata L. | Herb | Anthr | N | 0,02 | - | 0,01 |
| Dioscorea bulbifera L. | Herb | Anthr | N | - | - | 0,01 | |
| Dracaenaceae | Cordyline terminalis (L.) Kunth | Lign | Anthr | E | 0,16 | 0,28 | 0,16 |
| Euphorbiaceae | Acalypha godseffiana Mast. | Lign | Anthr | E | 0,08 | 0,10 | 0,13 |
| Acalypha wilkesiana Müll.Arg. | Lign | Anthr | E | 0,22 | 0,32 | 0,24 | |
| Breynia disticha J.R.Forst. & G.Forst. | Lign | Anthr | E | 0,06 | 0,04 | 0,03 | |
| Codiaeum variegatum (L.) Rumph. ex A.Juss. | Lign | Anthr | E | 0,04 | 0,10 | 0,03 | |
| Euphorbia characias L. | Herb | Anthr | E | - | 0,06 | 0,01 | |
| Euphorbia cotinifolia L. | Lign | Anthr | E | 0,04 | 0,02 | 0,03 | |
| Euphorbia heterophylla L. | Herb | Aném | N | 0,06 | - | 0,01 | |
| Euphorbia hirta L. | Herb | Aném | N | 0,06 | 0,08 | 0,04 | |
| Euphorbia milii Des Moul. | Herb | Anthr | E | 0,06 | 0,06 | 0,04 | |
| Euphorbia pulcherrima Willd. ex Klotzsch | Lign | Anthr | E | - | 0,02 | - | |
| Euphorbia sp | Lign | Anthr | E | 0,12 | 0,08 | 0,17 | |
| Euphorbia tirucalli L. | Lign | Anthr | E | 0,06 | - | 0,02 | |
| Euphorbia trigona Mill. | Lign | Anthr | E | - | 0,06 | 0,01 | |
| Jatropha curcas L. | Lign | Anthr | E | 0,06 | - | 0,02 | |
| Manihot esculenta Crantz | Lign | Anthr | E | 0,24 | 0,16 | 0,13 | |
| Manihot glaziovii Müll.Arg. | Lign | Anthr | E | 0,32 | 0,36 | 0,15 | |
| Phyllanthus muellerianus (Kuntze) Exell | Lign | Anthr | N | 0,08 | 0,14 | 0,04 | |
| Ricinus communis L. | Lign | Anthr | E | 0,04 | 0,04 | 0,02 | |
| Fabaceae | Acacia auriculiformis Benth. | Lign | Anthr | E | 0,08 | 0,02 | 0,01 |
| Arachis hypogaea L. | Herb | Anthr | E | - | - | 0,03 | |
| Bauhinia sp | Lign | Anthr | E | - | 0,02 | - | |
| Cajanus cajan (L.) Millsp. | Lign | Anthr | E | 0,04 | - | 0,01 | |
| Leucaena leucocephala (Lam.) de Wit | Lign | Anthr | E | 0,06 | 0,14 | 0,04 | |
| Mimosa pudica L. | Herb | Aném | N | 0,02 | - | - | |
| Phaseolus lunatus L. | Herb | Anthr | N | - | 0,02 | 0,01 | |
| Phaseolus vulgaris L. | Herb | Anthr | E | 0,02 | 0,04 | 0,09 | |
| Senna occidentalis (L.) Link | Lign | Aném | E | 0,02 | 0,04 | 0,02 | |
| Senna siamea (Lam.) H.S.Irwin & Barneby | Lign | Anthr | E | 0,04 | - | - | |
| Glycine max (L.) Merr. | Herb | Anthr | E | - | - | 0,01 | |
| Tephrosia vogelii Hook.f. | Lign | Anthr | E | 0,04 | - | - | |
| Iridaceae | Iris domestica (L.) Goldblatt & Mabb. | Herb | Anthr | E | 0,02 | 0,04 | 0,04 |
| Labiaceae | Ocimum basilicum L. | Herb | Anthr | E | 0,06 | 0,10 | 0,07 |
| Lamiaceae | Ajuga reptans L. | Herb | Anthr | E | - | 0,04 | 0,02 |
| Ocimum gratissimum L. | Herb | Anthr | E | 0,22 | 0,22 | 0,08 | |
| Plectranthus amboinicus (Lour.) Spreng. | Herb | Anthr | N | 0,10 | 0,22 | 0,12 | |
| Plectranthus sp | Lign | Anthr | E | 0,12 | 0,18 | 0,11 | |
| Prunella vulgaris L. | Herb | Anem | N | 0,18 | 0,30 | 0,08 | |
| Rosmarinus officinalis L. | Herb | Anthr | E | 0,06 | 0,06 | - | |
| Salvia officinalis L. | Herb | Anthr | E | - | 0,02 | 0,01 | |
| Solenostemon scutellarioides (L.) Codd | Herb | Anthr | E | 0,04 | 0,08 | 0,08 | |
| Lauraceae | Persea americana Mill. | Lign | Anthr | E | 0,48 | 0,68 | 0,48 |
| Liliaceae | Allium fistulosum L. | Herb | Anthr | E | 0,04 | 0,12 | 0,05 |
| Allium sativum L. | Herb | Anthr | E | - | 0,02 | 0,01 | |
| Chlorophytum comosum (Thunb.) Jacques | Herb | Anthr | E | 0,02 | 0,10 | 0,01 | |
| Tulbaghia violacea Harv. | Herb | Anthr | E | 0,02 | 0,02 | 0,04 | |
| Lythraceae | Cuphea hyssopifolia Kunth | Lign | Anthr | E | 0,04 | 0,04 | 0,05 |
| Punica granatum L. | Lign | Anthr | E | 0,04 | 0,02 | 0,01 | |
| Malvaceae | Abelmoschus esculentus (L.) Moench | Herb | Anthr | E | 0,12 | 0,06 | 0,21 |
| Gossypium hirsutum L. | Lign | Anthr | E | - | - | 0,01 | |
| Hibiscus acetosella Welw. ex Hiern | Herb | Anthr | E | 0,14 | 0,08 | 0,10 | |
| Hibiscus rosa-sinensis L. | Lign | Anthr | E | 0,06 | 0,10 | 0,09 | |
| Hibiscus sabdariffa L. | Herb | Anthr | E | 0,06 | 0,03 | 0,08 | |
| Hibiscus tiliaceus var. abutiloides (Willd.) Hochr. | Lign | Anthr | E | 0,02 | 0,01 | 0,01 | |
| Malva arborea (L.) Webb & Berthel. | Herb | Anthr | E | - | - | 0,07 | |
| Malvaviscus arboreus Cav. | Lign | Anthr | E | 0,07 | 0,10 | 0,15 | |
| Marantaceae | Maranta arundinacea L. | Herb | Anthr | E | - | - | 0,06 |
| Marsileaceae | Marsilea hirsuta R. Br. | Herb | Hydr | N | - | 0,18 | - |
| Meliaceae | Melia azedarach L. | Lign | Anthr | E | - | 0,02 | - |
| Moraceae | Ficus benjamina L. | Lign | Anthr | E | - | 0,04 | 0,01 |
| Ficus pumila L. | Lign | Anthr | N | - | 0,02 | 0,01 | |
| Ficus sp | Lign | Zooc | N | 0,01 | 0,04 | 0,01 | |
| Ficus thonningii Blume | Lign | Anthr | N | - | - | 0,01 | |
| Morus alba L. | Lign | Anthr | E | 0,10 | 0,12 | 0,01 | |
| Moringaceae | Moringa oleifera Lam. | Lign | Anthr | E | 0,06 | 0,04 | 0,07 |
| Musaceeae | Musa sp | Herb | Anthr | E | 0,26 | 0,10 | 0,10 |
| Myrtaceae | Callistemon speciosus (Sims) Sweet | Lign | Anthr | E | 0,04 | 0,06 | 0,03 |
| Eucalyptus sp | Lign | Anthr | E | 0,06 | 0,04 | - | |
| Psidium guajava L. | Lign | Anthr | E | 0,36 | 0,50 | 0,23 | |
| Syzygium guineense (Willd.) DC. | Lign | Anthr | E | 0,10 | 0,14 | 0,11 | |
| Nyctaginaceae | Bougainvillea sp | Lign | Anthr | E | 0,02 | 0,04 | 0,05 |
| Mirabilis jalapa L. | Herb | Anem | N | 0,02 | 0,10 | 0,16 | |
| Oxalidaceae | Oxalis triangularis A. St.-Hil. | Herb | Anem | N | 0,08 | 0,10 | 0,04 |
| Pandanaceae | Pandanus butayei De Wild. | Herb | Anthr | E | 0,02 | 0,08 | 0,09 |
| Passifloraceae | Adenia lobata (Jacq.) Engl. | Liane | Anthr | E | - | - | 0,01 |
| Passiflora edulis Sims | Liane | Anthr | E | 0,06 | 0,10 | 0,02 | |
| Pinaceae | Pinus sp | Lign | Anthr | E | 0,10 | 0,08 | 0,08 |
| Poaceae | Arundo donax L. | Hebarcée | Anthr | E | - | - | 0,03 |
| Cymbopogon citratus (DC.) Stapf | Herb | Anthr | E | 0,26 | 0,30 | 0,15 | |
| Cymbopogon densiflorus (Steud.) Stapf | Herb | Anthr | E | 0,02 | - | - | |
| Cynodon dactylon (L.) Pers. | Herb | Anthr | N | 0,24 | 0,20 | 0,10 | |
| Eleusine indica (L.) Gaertn. | Herb | Aném | N | 0,04 | 0,20 | 0,03 | |
| Imperata cylindrica (L.) Raeusch. | Herb | Aném | N | 0,06 | 0,06 | 0,01 | |
| Panicum maximum Jacq. | Herb | Aném | N | - | 0,02 | - | |
| Paspalum notatum Flüggé | Herb | Anthr | E | 0,16 | 0,32 | 0,01 | |
| Pennisetum purpureum Schumach. | Herb | Anem | N | - | 0,06 | - | |
| Phragmites australis (Cav.) Trin. ex Steud. | Herb | Anthr | N | - | 0,02 | - | |
| Saccharum officinarum L. | Herb | Anthr | E | 0,14 | 0,22 | 0,09 | |
| Setaria pallide-fusca (Schumach.) Stapf & C.E. Hubb. | Herb | Aném | N | - | - | 0,03 | |
| Zea mays L. | Herb | Anthr | E | - | 0,06 | 0,04 | |
| Polygonaceae | Rumex usambarensis (Dammer) Dammer | Herb | Anthr | E | 0,02 | - | 0,04 |
| Polypodiacdeae | Drynaria laurentii (Christ) Hieron. | Herb | Anthr | N | 0,04 | 0,08 | 0,02 |
| Pontedieraceae | Eichhornia crassipes (Mart.) Solms | Herb | Anthr | E | 0,02 | - | 0,01 |
| Portulacaceae | Portulaca grandiflora Hook. | Herb | Anthr | N | - | - | 0,02 |
| Portulaca oleracea L. | Herb | Aném | N | 0,06 | 0,14 | 0,06 | |
| Rosaceae | Fragaria sp | Herb | Anthr | E | - | 0,02 | 0,01 |
| Malus domestica Baumg. | Lign | Anthr | E | 0,02 | - | 0,01 | |
| Rosa sp | Lign | Anthr | E | 0,10 | 0,12 | 0,05 | |
| Rubiaceae | Coffea sp | Lign | Anthr | E | 0,04 | - | - |
| Rutaceae | Casimiroa edulis La Llave | Lign | Anthr | E | 0,06 | - | 0,02 |
| Citrus limon (L.) Osbeck | Lign | Anthr | E | 0,34 | 0,38 | 0,13 | |
| Citrus sinensis (L.) Osbeck | Lign | Anthr | E | 0,14 | 0,06 | 0,09 | |
| Solanaceae | Brugmansia candida Pers. | Lign | Anthr | E | 0,02 | 0,08 | 0,01 |
| Capsicum annuum L. | Herb | Anthr | E | 0,06 | 0,02 | 0,03 | |
| Capsicum chinense Jacq. | Herb | Anthr | E | - | 0,02 | 0,01 | |
| Capsicum frutescens L. | Herb | Anthr | E | 0,02 | 0,06 | 0,07 | |
| Cestrum nocturnum L. | Lign | Anthr | E | 0,06 | 0,12 | 0,03 | |
| Lycopersicon esculentum Mill. | Herb | Anthr | E | 0,12 | 0,26 | 0,23 | |
| Nicandra indica Roem. & Schult. | Herb | Aném | N | 0,02 | 0,04 | 0,02 | |
| Nicotiana tabacum L. | Herb | Anthr | E | 0,04 | - | - | |
| Physalis peruviana L. | Herb | Anthr | N | - | - | 0,01 | |
| Solanum aethiopicum L. | Herb | Anthr | E | 0,08 | 0,04 | 0,02 | |
| Solanum anguivi Lam. | Herb | Anthr | E | - | 0,02 | 0,01 | |
| Solanum melongena L. | Herb | Anthr | E | 0,02 | 0,04 | 0,05 | |
| Solanum torvum Sw. | Herb | Anthr | N | 0,02 | - | 0,01 | |
| Typhaceae | Typha sp | Herb | Anem | N | - | 0,04 | 0,01 |
| Urticaceae | Pilea cadierei Gagnep. & Guillaumin | Herb | Anthr | E | 0,02 | - | 0,03 |
| Verbenaceae | Duranta erecta L. | Lign | Anthr | E | 0,18 | 0,24 | 0,16 |
| Duranta repens L. | Lign | Anthr | E | - | 0,02 | 0,01 | |
| Lantana camara L. | Lign | Anthr | E | 0,10 | 0,18 | 0,02 | |
| Stachytarpheta indica (L.) Vahl | Herb | Anthr | N | - | - | 0,01 | |
| Vitex trifolia L. | Lign | Anthr | E | 0,22 | 0,32 | 0,20 | |
References
- Angel, S. Urban Expansion: Theory, Evidence and Practice. Buildings & Cities 2023, 4, 124–138. [Google Scholar] [CrossRef]
- Yamashita, A. Rapid Urbanization in Developing Asia and Africa. In Urban Development in Asia and Africa: Geospatial Analysis of Metropolises; Springer: Dordrecht, Netherlands, 2017; pp. 47–61. [Google Scholar] [CrossRef]
- Henderson, J.V.; Turner, M.A. Urbanization in the Developing World: Too Early or Too Slow? J. Econ. Perspect. 2020, 34, 150–173. [Google Scholar] [CrossRef]
- Gu, D.; Andreev, K.; Dupre, M.E. Major Trends in Population Growth Around the World. China CDC Wkly. 2021, 3, 604–613. [Google Scholar] [CrossRef]
- Cividino, S.; Halbac-Cotoara-Zamfir, R.; Salvati, L. Revisiting the “City Life Cycle”: Global urbanization and implications for regional development. Sustainability 2020, 12, 1151. [Google Scholar] [CrossRef]
- UN Department of Economic and Social Affairs. World Urbanization Prospects. Demogr. Res. 2018, 12. Available online: https://population.un.org/wup/Publications/Files/WUP2018-Report.pdf (accessed on 14 April 2024).
- Bai, L.; Xiu, C.; Feng, X.; Liu, D. Influence of Urbanization on Regional Habitat Quality: A Case Study of Changchun City. Habitat Int. 2019, 93, 102042. [Google Scholar] [CrossRef]
- Li, G.; Fang, C.; Li, Y.; Wang, Z.; Sun, S.; He, S.; Liu, X. Global Impacts of Future Urban Expansion on Terrestrial Vertebrate Diversity. Nat. Commun. 2022, 13, 1628. [Google Scholar] [CrossRef]
- Useni, S.Y.; Malaisse, F.; Kaleba, S.C.; Mwanke, A.K.; Yamba, A.M.; Khonde, C.N.; Kankumbi, F.M. Tree Diversity and Structure on Green Space of Urban and Peri-Urban Zones: The Case of Lubumbashi City in the Democratic Republic of Congo. Urban For. Urban Green. 2019, 41, 67–74. [Google Scholar] [CrossRef]
- Kraemer, R.; Kabisch, N. Parks Under Stress: Air Temperature Regulation of Urban Green Spaces Under Conditions of Drought and Summer Heat. Front. Environ. Sci. 2022, 10, 849965. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Mata, L.; Mackie, J.A.; Hahs, A.K.; Stork, N.E.; Williams, N.S.; Livesley, S.J. Increasing Biodiversity in Urban Green Spaces Through Simple Vegetation Interventions. J. Appl. Ecol. 2017, 54, 1874–1883. [Google Scholar] [CrossRef]
- Richards, D.R.; Belcher, R.N.; Carrasco, L.R.; Edwards, P.J.; Fatichi, S.; Hamel, P.; Stanley, M.C. Global Variation in Contributions to Human Well-Being from Urban Vegetation Ecosystem Services. One Earth 2022, 5, 522–533. [Google Scholar] [CrossRef]
- Kowarik, I. Novel Urban Ecosystems, Biodiversity, and Conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef]
- Useni Sikuzani, Y.; Sambiéni Kouagou, R.; Maréchal, J.; Ilunga wa Ilunga, E.; Malaisse, F.; Bogaert, J.; Munyemba Kankumbi, F. Changes in the Spatial Pattern and Ecological Functionalities of Green Spaces in Lubumbashi (the Democratic Republic of Congo) in Relation with the Degree of Urbanization. Trop. Conserv. Sci. 2018, 11, 1940082918771325. [Google Scholar] [CrossRef]
- Bigirimana, J.; Bogaert, J.; De Cannière, C.; Bigendako, M.J.; Parmentier, I. Domestic Garden Plant Diversity in Bujumbura, Burundi: Role of the Socio-Economical Status of the Neighborhood and Alien Species Invasion Risk. Landsc. Urban Plan. 2012, 107, 118–126. [Google Scholar] [CrossRef]
- Al-Kofahi, S.D.; Gharaibeh, A.A.; Bsoul, E.Y.; Othman, Y.A.; St Hilaire, R. Investigating Domestic Gardens’ Densities, Spatial Distribution and Types Among City Districts. Urban Ecosyst. 2019, 22, 567–581. [Google Scholar] [CrossRef]
- Al-Kofahi, S.D.; Al-Kafawin, A.M.; Al-Gharaibeh, M.M. Investigating Domestic Gardens Landscape Plant Diversity, Implications for Valuable Plant Species Conservation. Environ. Dev. Sustain. 2024, 26, 21259–21279. [Google Scholar] [CrossRef]
- Malaisse, F. Se Nourrir en Forêt Claire Africaine: Approche Écologique et Nutritionnelle; Les Presses Agronomiques de Gembloux: Belgium, 1997; 384p. [Google Scholar]
- Useni, S.Y.; Kaleba, S.C.; Khonde, C.N.; Mwana, Y.A.; Malaisse, F.; Bogaert, J.; Kankumbi, F.M. Vingt-cinq ans de monitoring de la dynamique spatiale des espaces verts en réponse á urbanisation dans les communes de la ville de Lubumbashi (Haut-Katanga, RD Congo). Tropicultura 2017, 35, 300–311. [Google Scholar]
- INS (Institut National de la Statistique). Anuaire Statistique RDC 2022; Ministère National du Plan: Kinshasa, Democratic Republic of the Congo, 2022; 433p. [Google Scholar]
- Khoji, M.H.; Nghonda, D.D.N.; Malaisse, F.; Waselin, S.; Sambiéni, K.R.; Kaleba, S.C.; Kankumbi, F.M.; Bastin, J.F.; Bogaert, J.; Sikuzani, Y.U. Quantification and Simulation of Landscape Anthropization around the Mining Agglomerations of Southeastern Katanga (DR Congo) between 1979 and 2090. Land 2022, 11, 850. [Google Scholar] [CrossRef]
- Mwitwa, J.; German, L.; Muimba-Kankolongo, A.; Puntodewo, A. Governance and sustainability challenges in landscapes shaped by mining: Mining forestry linkages and impacts in the Copper Belt of Zambia and the DR Congo. For. Policy Econ. 2012, 25, 19–30. [Google Scholar] [CrossRef]
- Useni, S.; Cabala, K.; Halleux, J.M.; Bogaert, J.; Munyemba, K. Caractérisation de la Croissance Spatiale Urbaine de la Ville de Lubumbashi (Haut-Katanga, RD Congo) entre 1989 et 2014. Tropicultura 2018, 38, 98–108. [Google Scholar]
- Useni Sikuzani, Y.; Boisson, S.; Cabala Kaleba, S.; Nkuku Khonde, C.; Malaisse, F.; Halleux, J.M.; Munyemba Kankumbi, F. Dynamique de l’Occupation du Sol autour des Sites Miniers le Long du Gradient Urbain-Rural de la Ville de Lubumbashi, RD Congo. Biotechnol. Agron. Soc. Environ. 2020, 24, 14p. [Google Scholar] [CrossRef]
- Useni, S.Y.; Malaisse, F.; Mwembu, D.D.D.; Bogaert, J. Lubumbashi (DR Congo): Navigating the Socio-Ecological Complexities of a Vital Mining Hub. Cities 2024, 154, 105341. [Google Scholar] [CrossRef]
- Useni, Y.S.; Malaisse, F.; Yona, J.M.; Mwamba, T.M.; Bogaert, J. Diversity, Use and Management of Household-Located Fruit Trees in Two Rapidly Developing Towns in Southeastern DR Congo. Urban For. Urban Green. 2021, 63, 127220. [Google Scholar] [CrossRef]
- Galluzzi, G.; Eyzaguirre, P.; Negri, V. Home Gardens: Neglected Hotspots of Agro-Biodiversity and Cultural Diversity. Biodivers. Conserv. 2010, 19, 3635–3654. [Google Scholar] [CrossRef]
- Bernholt, H.; Kehlenbeck, K.; Gebauer, J.; Buerkert, A. Plant Species Richness and Diversity in Urban and Peri-Urban Gardens of Niamey, Niger. Agrofor. Syst. 2009, 77, 159–179. [Google Scholar] [CrossRef]
- Freeman, C.; Dickinson, K.J.; Porter, S.; Van Heezik, Y. "My Garden Is an Expression of Me": Exploring Householders’ Relationships with Their Gardens. J. Environ. Psychol. 2012, 32, 135–143. [Google Scholar] [CrossRef]
- Kumar, B.M. Carbon Sequestration Potential of Tropical Homegardens. In Tropical Homegardens: A Time-Tested Example of Sustainable Agroforestry; Springer: Dordrecht, Netherlands, 2006; pp. 185–204. [Google Scholar] [CrossRef]
- Lubbe, C.S.; Siebert, S.J.; Cilliers, S.S. Political Legacy of South Africa Affects the Plant Diversity Patterns of Urban Domestic Gardens Along a Socio-Economic Gradient. Sci. Res. Essays 2010, 5, 2900–2910. [Google Scholar]
- Mosina, G.K.; Maroyi, A.; Potgieter, M.J. Comparative Analysis of Plant Use in Peri-Urban Domestic Gardens of the Limpopo Province, South Africa. J. Ethnobiol. Ethnomed. 2014, 10, 1–8. [Google Scholar] [CrossRef]
- Karhagomba, I.B.; Mushagalusa, T.B.; Nabino, V.B.; Koh, K.; Kim, H.S. The Cultivation of Wild Food and Medicinal Plants for Improving Community Livelihood: The Case of the Buhozi Site, DR Congo. Nutr. Res. Pract. 2013, 7, 510–518. [Google Scholar] [CrossRef]
- Etshekape, P.G.; Atangana, A.R.; Khasa, D.P. Tree Planting in Urban and Peri-Urban of Kinshasa: Survey of Factors Facilitating Agroforestry Adoption. Urban For. Urban Green. 2018, 30, 12–23. [Google Scholar] [CrossRef]
- Napawan, N.C. Complexity in Urban Agriculture: The Role of Landscape Typologies in Promoting Urban Agriculture’s Growth. J. Urban. 2016, 9, 19–38. [Google Scholar] [CrossRef]
- Scarlett, L.; Boyd, J. Ecosystem Services and Resource Management: Institutional Issues, Challenges, and Opportunities in the Public Sector. Ecol. Econ. 2015, 115, 3–10. [Google Scholar] [CrossRef]
- Maréchal, J.; Useni, S.Y.; Bogaert, J.; Munyemba, K.F.; Mahy, G. La Perception par des Experts Locaux des Espaces Verts et de Leurs Services Écosystémiques dans une Ville Tropicale en Expansion: Le Cas de Lubumbashi. In Anthropisation des Paysages Katangais; Bogaert, J., Colinet, G., Mahy, G., Eds.; 2018; pp. 59–69.
- Kalombo, K.D. Caractérisation de la Répartition Temporelle des Précipitations à Lubumbashi (Sud-Est de la RDC) sur la Période de 1970–2014. In Proceedings of the XXIII Colloque de l’Association Internationale de Climatologie, Liège, Belgium; 2015; pp. 531–536. [Google Scholar]
- Kalombo, K.D. Évaluation des Éléments du Climat en RDC; Editions Universitaires Européennes: Saarbrücken, Germany, 2016. [Google Scholar]
- Useni, S.Y.; Malaisse, F.; Kaleba, S.C.; Kankumbi, F.M.; Bogaert, J. Le Rayon de Déforestation autour de la Ville de Lubumbashi (Haut-Katanga, RD Congo): Synthèse. Tropicultura 2017, 35. [Google Scholar]
- Vranken, I.; Adam, M.; Mujinya, B.B.; Munyemba, F.K.; Baert, G.; Van Ranst, E.; Bogaert, J. Termite Mounds Identification through Aerial Photographic Interpretation in Lubumbashi: Methodology Evaluation. Trop. Conserv. Sci. 2014, 7, 733–746. [Google Scholar] [CrossRef]
- Useni Sikuzani, Y.; Andre, M.; Mahy, G.; Cabala Kaleba, S.; Malaisse, F.; Munyemba Kankumbi, F.; Bogaert, J. Interprétation Paysagère du Processus d’Urbanisation à Lubumbashi (RD Congo): Dynamique de la Structure Spatiale et Suivi des Indicateurs Écologiques entre 2002 et 2008. Tropicultura 2018. [Google Scholar]
- Groupe Huit. Élaboration du Plan Urbain de Référence de Lubumbashi [Development of the Urban Reference Plan of Lubumbashi]; Beau, Ministère des Infrastructures et Travaux Publics, Lubumbashi, Democratic Republic of the Congo, 2009.
- Loram, A.; Warren, P.H.; Gaston, K.J. Urban Domestic Gardens (XIV): The Characteristics of Gardens in Five Cities. Environ. Manag. 2008, 42, 361–376. [Google Scholar] [CrossRef]
- Loram, A.; Warren, P.; Thompson, K.; Gaston, K. Urban Domestic Gardens: The Effects of Human Interventions on Garden Composition. Environ. Manag. 2011, 48, 808–824. [Google Scholar] [CrossRef]
- Lebrun, J.-P.; Stork, A.L. Énumération des Plantes à Fleurs d’Afrique Tropicale et Tropical African Flowering Plants: Ecology and Distribution; Conservatoire et Jardin Botaniques de la Ville de Genève: Geneva, Switzerland, 1991–2015. [Google Scholar]
- Bigirimana, J.; Bogaert, J.; De Cannière, C.; Lejoly, J.; Parmentier, I. Alien Plant Species Dominate the Vegetation in a City of Sub-Saharan Africa. Landsc. Urban Plan. 2011, 100, 251–267. [Google Scholar] [CrossRef]
- Rija, A.A.; Said, A.; Mwamende, K.A.; Hassan, S.H.; Madoffe, S.S. Urban Sprawl and Species Movement May Decimate Natural Plant Diversity in an Afro-Tropical City. Biodivers. Conserv. 2014, 23, 963–978. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a Major Cause of Biotic Homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Al Shaye, N.A.; Masrahi, Y.S.; Thomas, J. Ecological Significance of Floristic Composition and Life Forms of Riyadh Region, Central Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Trait Convergence and Trait Divergence in Herbaceous Plant Communities: Mechanisms and Consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Vittoz, P.; Engler, R. Seed Dispersal Distances: A Typology Based on Dispersal Modes and Plant Traits. Bot. Helv. 2007, 117, 109–124. [Google Scholar] [CrossRef]
- Qvenild, M.; Setten, G.; Skår, M. Politicising Plants: Dwelling and Invasive Alien Species in Domestic Gardens in Norway. Norsk Geogr. Tidsskr.-Nor. J. Geogr. 2014, 68, 22–33. [Google Scholar] [CrossRef]
- Guo, W.Y.; van Kleunen, M.; Pierce, S.; Dawson, W.; Essl, F.; Kreft, H.; Pyšek, P. Domestic Gardens Play a Dominant Role in Selecting Alien Species with Adaptive Strategies that Facilitate Naturalization. Glob. Ecol. Biogeogr. 2019, 28, 628–639. [Google Scholar] [CrossRef]
- Goenster, S.; Wiehle, M.; Kehlenbeck, K.; Jamnadass, R.; Gebauer, J.; Buerkert, A. Indigenous Fruit Trees in Homegardens of the Nuba Mountains, Central Sudan: Tree Diversity and Potential for Improving the Nutrition and Income of Rural Communities. In Proceedings of I All Africa Horticultural Congress 911; 2009, pp. 355–364.
- Caratini, R. Botanique 4; Bordas: Paris, France, 1985. [Google Scholar]
- Michael, G.C.; Tijani-Eniola, H.; Nwagwu, F.A.; Olaniyi, O.W. Weed Flora Dynamics in Maize (Zea mays L.) Field as Influenced by Cover Crop Types and Planting Densities at Jalingo, Northeast, Nigeria. Int. J. Plant Soil Sci. 2021, 33, 1–16. [Google Scholar] [CrossRef]
- Nkuku, K.C.; Rémon, M. Stratégies de Survie à Lubumbashi (R-D Congo). Enquête sur 14000 Ménages Urbains; Archive Congolaise, l’Harmattan: Paris, France, 2006. [Google Scholar]
- Dzhambov, A.M.; Lercher, P.; Browning, M.H.; Stoyanov, D.; Petrova, N.; Novakov, S.; Dimitrova, D.D. Does Greenery Experienced Indoors and Outdoors Provide an Escape and Support Mental Health during the COVID-19 Quarantine? Environ. Res. 2021, 196, 110420. [Google Scholar] [CrossRef]
- Lubbe, C.S.; Siebert, S.J.; Cilliers, S.S. Floristic Analysis of Domestic Gardens in the Tlokwe City Municipality, South Africa. Bothalia 2011, 41, 351–361. [Google Scholar] [CrossRef]
- Ngcaba, P.; Maroyi, A. Home Gardens in the Eastern Cape Province, South Africa: A Promising Approach to Enhance Household Food Security and Well-Being. Biodiversitas 2021, 22, 3969–3978. [Google Scholar] [CrossRef]
- Sierra-Guerrero, M.C.; Amarillo-Suárez, A.R. Socioecological Features of Plant Diversity in Domestic Gardens in the City of Bogotá, Colombia. Urban For. Urban Green. 2017, 28, 54–62. [Google Scholar] [CrossRef]
- Kurz, T.; Baudains, C. Biodiversity in the Front Yard: An Investigation of Landscape Preference in a Domestic Urban Context. Environ. Behav. 2012, 44, 166–196. [Google Scholar] [CrossRef]
- Kendal, D.; Williams, K.J.; Williams, N.S. Plant Traits Link People’s Plant Preferences to the Composition of Their Gardens. Landsc. Urban Plan. 2012, 105, 34–42. [Google Scholar] [CrossRef]
- Fadamiro, J.A.; Adedeji, J.A. Home Gardens and Quality of Life in Akure, Nigeria. In Proceedings of Environmental Design and Management International Conference, 2015, pp. 11–20.
- Taylor, J.R.; Lovell, S.T.; Wortman, S.E.; Chan, M. Ecosystem Services and Tradeoffs in the Home Food Gardens of African American, Chinese-Origin and Mexican-Origin Households in Chicago, IL. Renew. Agric. Food Syst. 2017, 32, 69–86. [Google Scholar] [CrossRef]
- Shackleton, C.M.; Hebinck, P.; Kaoma, H.; Chishaleshale, M.; Chinyimba, A.; Shackleton, S.E.; Gumbo, D. Low-Cost Housing Developments in South Africa Miss the Opportunities for Household Level Urban Greening. Land Use Policy 2014, 36, 500–509. [Google Scholar] [CrossRef]
- Ghose, R.; Pettygrove, M. Actors and Networks in Urban Community Garden Development. Geoforum 2014, 53, 93–103. [Google Scholar] [CrossRef]
- Muratet, A. Diversité Végétale en Milieu Urbain: L’Exemple des Hauts-de-Seine; Doctoral Dissertation, Paris 6, 2006.
- Petschek, P.; Gass, S. (Eds.) Constructing Shadows: Pergolas, Pavilions, Tents, Cables, and Plants; Walter de Gruyter: Berlin, Germany, 2012. [Google Scholar]
- Hayward, G. Small Buildings, Small Gardens: Creating Gardens around Structures; Gibbs Smith: Layton, UT, USA, 2007. [Google Scholar]
- Young, C.; Hofmann, M.; Frey, D.; Moretti, M.; Bauer, N. Psychological Restoration in Urban Gardens Related to Garden Type, Biodiversity and Garden-Related Stress. Landsc. Urban Plan. 2020, 198, 103777. [Google Scholar] [CrossRef]
- Ngur-Ikone, J.K. La Politique Publique de la Gestion des Espaces Verts par l’Hôtel de Ville de Kinshasa. Afr. Dev. 2010, 35, 13–46. [Google Scholar]
- Cameron, R.W.F.; Blanusa, T.; Taylor, J.E.; Salisbury, A.; Halstead, A.J.; Henricot, B.; Thompson, K. The Domestic Garden—Its Contribution to Urban Green Infrastructure. Urban For. Urban Green. 2012, 11, 129–137. [Google Scholar] [CrossRef]
- Wolseley, V.F.G. Gardens; Applewood Books: Carlisle, MA, USA, 2008. [Google Scholar]
- Smith, M.R. The Front Garden: New Approaches to Landscape Design; Houghton Mifflin Harcourt: Boston, MA, USA, 2001. [Google Scholar]
- Makumbelo, E.; Paulus, J.J.; Luyindula, N.; Lukoli, L. Apport des Arbres Fruitiers à la Sécurité Alimentaire en Milieu Urbain Tropical: Cas de la Commune de Limete à Kinshasa, R.D. Congo. Tropicultura 2005, 23, 245–252. [Google Scholar]
- Shutcha, M.N.; Mubemba, M.M.; Faucon, M.P.; Luhembwe, M.N.; Visser, M.; Colinet, G.; Meerts, P. Phytostabilisation of Copper-Contaminated Soil in Katanga: An Experiment with Three Native Grasses and Two Amendments. Int. J. Phytoremediation 2010, 12, 616–632. [Google Scholar] [CrossRef]
- Briggiler Marcó, M.; Reinheimer, J.A.; Quiberoni, A. Phage Adsorption to Lactobacillus plantarum: Influence of Physiological and Environmental Factors. Int. J. Food Microbiol. 2010, 138, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Makumbelo, E.; Paulus, J.J.; Luyindula, N.; Lukoli, L. Apport des Arbres Fruitiers à la Sécurité Alimentaire en Milieu Urbain Tropical: Cas de la Commune de Limete à Kinshasa, R.D. Congo. Tropicultura 2005, 23, 245–252. [Google Scholar]
- Drayton, B.; Primack, R.B. Plant Species Lost in an Isolated Conservation Area in Metropolitan Boston from 1894 to 1993. Conserv. Biol. 1996, 10, 30–39. [Google Scholar] [CrossRef]
- Vilà, M.; Hulme, P.E. Non-Native Species, Ecosystem Services, and Human Well-Being. In Impact of Biological Invasions on Ecosystem Services; Springer: Cham, Switzerland, 2017; pp. 1–14. [Google Scholar] [CrossRef]
- Zaninotto, V.; Thebault, E.; Dajoz, I. Native and Exotic Plants Play Different Roles in Urban Pollination Networks across Seasons. Oecologia 2023, 201, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Weidenhamer, J.D.; Callaway, R.M. Direct and Indirect Effects of Invasive Plants on Soil Chemistry and Ecosystem Function. J. Chem. Ecol. 2010, 36, 59–69. [Google Scholar] [CrossRef]
- Charles, H.; Dukes, J.S. Impacts of Invasive Species on Ecosystem Services. In Biological Invasions; Springer: Berlin, Germany, 2007; pp. 217–237. [Google Scholar] [CrossRef]
- Biró, É.; Babai, D.; Bódis, J.; Molnár, Z. Lack of Knowledge or Loss of Knowledge? Traditional Ecological Knowledge of Population Dynamics of Threatened Plant Species in East-Central Europe. J. Nat. Conserv. 2014, 22, 318–325. [Google Scholar] [CrossRef]
- Loram, A.; Warren, P.; Thompson, K.; Gaston, K. Urban Domestic Gardens: The Effects of Human Interventions on Garden Composition. Environ. Manag. 2011, 48, 808–824. [Google Scholar] [CrossRef]
- UNDP. Rapport sur le Développement Humain 2023-2024 Aperçu: Sortir de l’Impasse Aperçu du Rapport sur le Développement Humain 2023-2024; UNDP, 2024. Available online: https://hdr.undp.org/system/files/documents/global-report-document/hdr2023-24snapshotfr.pdf (accessed on 29 August 2024).
- Wilberforce, J.O.; Olivia, E.I.N. Phytochemical Screening and Antimicrobial Activity of Leaves Extracts of Mangifera indica and Carica papaya. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3253–3259. [Google Scholar] [CrossRef]
- Kasongo, L.M.E.; Mwamba, M.T.; Tshipoya, M.P.; Mukalay, M.J.; Useni, S.Y.; Mazinga, K.M.; Nyembo, K.L. Réponse de la Culture de Soja (Glycine max L. [Merril]) à l’Apport des Biomasses Vertes de Tithonia diversifolia (Hemsley) A. Gray comme Fumure Organique sur un Ferralsol à Lubumbashi, RD Congo. J. Appl. Biosci. 2013, 63, 4727–4735. [Google Scholar] [CrossRef]
- Paul, A.; Khan, Z.; Bhattacharyya, A.; Majumder, S.; Banerjee, K. Multiclass Pesticide Residue Analysis in Tobacco (Nicotiana tabacum) Using High Performance Liquid Chromatography-High Resolution (Orbitrap) Mass Spectrometry: A Simultaneous Screening and Quantitative Method. J. Chromatogr. A 2021, 1648, 462208. [Google Scholar] [CrossRef]
- Michel, M.M.M.; Yannick, U.S.; François, N.N.; Emmanuel, M.M.; Prisca, K.K.; Muyembe, M.; Luciens, N.K. Évaluation des Teneurs en Éléments Traces Métalliques dans les Légumes Feuilles Vendus dans les Différents Marchés de la Zone Minière de Lubumbashi. J. Appl. Biosci. 2013, 66, 5106–5113. [Google Scholar] [CrossRef]
- Mpundu Mubemba, M.; Useni, Y.; Nyembo, L.; Colinet, G. Effets d’Amendements Carbonatés et Organiques sur la Culture de Deux Légumes sur Sol Contaminé à Lubumbashi (RD Congo). Biotechnol. Agron. Soc. Environ. 2014, 18. [Google Scholar]
- Banza, C.L.N.; Nawrot, T.S.; Haufroid, V.; Decrée, S.; De Putter, T.; Smolders, E.; Nemery, B. High Human Exposure to Cobalt and Other Metals in Katanga, a Mining Area of the Democratic Republic of Congo. Environ. Res. 2009, 109, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Mununga Katebe, F.; Szekely, I.; Mpundu Mubemba, M.; Burgeon, C.; Jijakli, M.H. Bioponic Cultivation Using Chicken Droppings to Produce Lettuce Plants (Lactuca sativa rz) Uncontaminated by Trace Metals. Horticulturae 2024, 10, 605. [Google Scholar] [CrossRef]
- Cilliers, S.S.; Siebert, S.J.; Du Toit, M.J.; Barthel, S.; Mishra, S.; Cornelius, S.F.; Davoren, E. Garden Ecosystem Services of Sub-Saharan Africa and the Role of Health Clinic Gardens as Social-Ecological Systems. Landsc. Urban Plan. 2018, 180, 294–307. [Google Scholar] [CrossRef]
- Useni Sikuzani, Y.; Mpanda Mukenza, M.; Mwenya, I.K.; Muteya, H.K.; Nghonda, D.D.; Mukendi, N.K.; Bogaert, J. Quantifying Forest Cover Loss during the COVID-19 Pandemic in the Lubumbashi Charcoal Production Basin (DR Congo) through Remote Sensing and Landscape Analysis. Resources 2024, 13, 7. [Google Scholar] [CrossRef]
- Hungerford, H.; Moussa, Y. Seeing the (Urban) Forest through the Trees: Governance and Household Trees in Niamey, Niger. Afr. Geogr. Rev. 2017, 36, 286–304. [Google Scholar] [CrossRef]
- Mistry, A.; Spocter, M. Production of Edibles and Use of Garden Waste in Domestic Gardens of a Middle-Class Suburb in Cape Town, South Africa. J. Urban. 2020, 13, 1–14. [Google Scholar] [CrossRef]
- Jensen, J.L.; Sørensen, E.B. Recreation, Cultivation and Environmental Concerns: Exploring the Materiality and Leisure Experience of Contemporary Allotment Gardening. Leis. Stud. 2020, 39, 322–340. [Google Scholar] [CrossRef]
- Von Thaden, J.; Badillo-Montaño, R.; Lira-Noriega, A.; García-Ramírez, A.; Benítez, G.; Equihua, M.; Pérez-Maqueo, O. Contributions of Green Spaces and Isolated Trees to Landscape Connectivity in an Urban Landscape. Urban For. Urban Green. 2021, 64, 127277. [Google Scholar] [CrossRef]
- Nowak, D.J.; Greenfield, E.J.; Hoehn, R.E.; Lapoint, E. Carbon Storage and Sequestration by Trees in Urban and Community Areas of the United States. Environ. Pollut. 2013, 178, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.; Butler, E.; Orians, C.M. Native Plant Enthusiasm Reaches New Heights: Perceptions, Evidence, and the Future of Green Roofs. Urban For. Urban Green. 2012, 11, 1–10. [Google Scholar] [CrossRef]
- Gough, M.Z.; Accordino, J. Public Gardens as Sustainable Community Development Partners: Motivations, Perceived Benefits, and Challenges. Urban Aff. Rev. 2013, 49, 851–887. [Google Scholar] [CrossRef]
- Simoncini, R. Developing an Integrated Approach to Enhance the Delivering of Environmental Goods and Services by Agro-Ecosystems. Reg. Environ. Change 2009, 9, 153–167. [Google Scholar] [CrossRef]
- Music, J.; Large, C.; Charlebois, S.; Mayhew, K. Gardening from the Ground Up: A Review of Grassroots Governance and Management of Domestic Gardening in Canada. Local Environ. 2022, 27, 1046–1058. [Google Scholar] [CrossRef]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling Up from Gardens: Biodiversity Conservation in Urban Environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef]
- Yacamán Ochoa, C.; Ferrer Jiménez, D.; Mata Olmo, R. Green Infrastructure Planning in Metropolitan Regions to Improve the Connectivity of Agricultural Landscapes and Food Security. Land 2020, 9, 414. [Google Scholar] [CrossRef]
Figure 1.
Geographical map of the Lubumbashi city in south-eastern DRC. The map also shows the study sites, the Gambela 3, bel air 1 and mampala neighbourhood.
Figure 1.
Geographical map of the Lubumbashi city in south-eastern DRC. The map also shows the study sites, the Gambela 3, bel air 1 and mampala neighbourhood.
Figure 2.
Origin status of plant species inventoried in domestic gardens across neighborhood types.
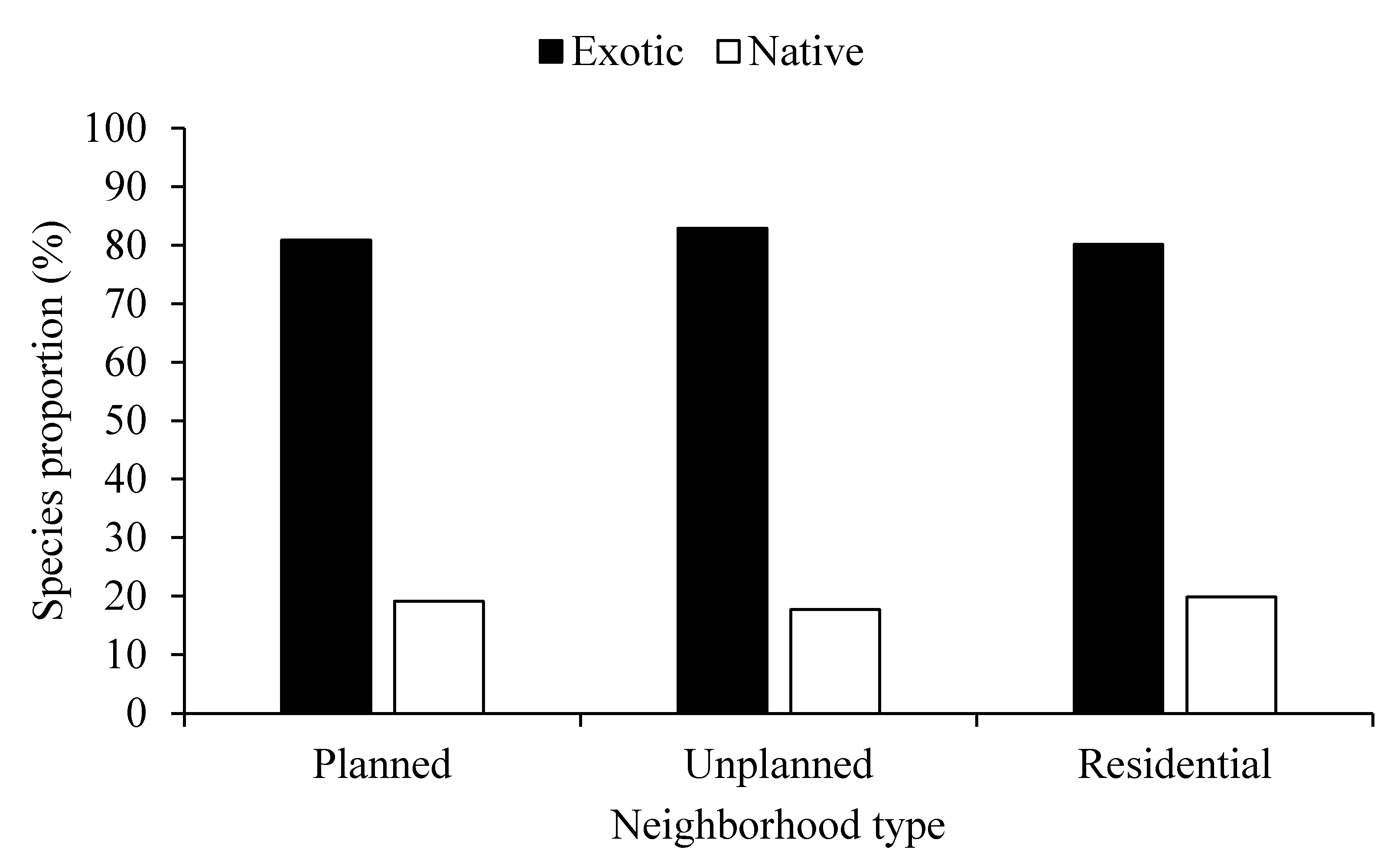

Table 1.
Caratini’s frequency index classification for species abundance and distribution.
| Frequency | Indices | Qualification |
|---|---|---|
| 0.8 to 1.0 | V | Constant |
| 0.6 to 0.8 | IV | Abundant |
| 0.4 to 0.6 | III | Frequent |
| 0.2 to 0.4 | II | Accessory |
| 0.0 to 0.2 | I | Rare or accidental |
Table 2.
Characteristics of domestic gardens (DG) and plots across planned neighborhood (PN), unplanned neighborhood (UN) and residential neighborhood (RN) in the city of Lubumbashi. Mean ± Standard Deviation, letters indicate significant differences between neighborhood means. ***p<.001.
Table 2.
Characteristics of domestic gardens (DG) and plots across planned neighborhood (PN), unplanned neighborhood (UN) and residential neighborhood (RN) in the city of Lubumbashi. Mean ± Standard Deviation, letters indicate significant differences between neighborhood means. ***p<.001.
| Neighborhood types |
Presence of DG (%) | Number of DG | DG area (m2) |
Species richness |
Plot area (m2) |
|---|---|---|---|---|---|
| PN | 89.0 | 2.0±1.3c | 42.8±57.3c | 13,355±12,811c | 526,8 ±192,8c |
| UN | 90.0 | 2.4±0.7b | 154.3±106.6b | 15,96±5,852b | 963,0 ±226,0b |
| RN | 94.0 | 2.8±0.6a | 315.1±336.2a | 22±8,272a | 1032,0 ±205,5a |
| P-value | - | *** | *** | *** | *** |
Table 3.
Distribution of domestic garden shapes, typologies, and utilizations across different neighborhood types. Data are expressed in percentage. n refers to the number of domestic gardens.
Table 3.
Distribution of domestic garden shapes, typologies, and utilizations across different neighborhood types. Data are expressed in percentage. n refers to the number of domestic gardens.
| Parameter | Form | Neighborhood type | ||
|---|---|---|---|---|
| Planned (n=89) | Unplanned (n=90) | Residential (n=94) | ||
| Shape | Square | 2.2 | 0.8 | 0.7 |
| Rectangular | 18.6 | 22.0 | 19.0 | |
| Irregular | 79.2 | 77.2 | 80.3 | |
| Typology | Pergolas | 2.9 | 1.6 | 2.1 |
| Hedge | 9.8 | 16.5 | 13.4 | |
| Flowerbed | 43.1 | 38.6 | 31.7 | |
| Lawn | 6.1 | 11.0 | 19.0 | |
| Shruberry | 38.0 | 32.3 | 33.8 | |
| Utilization | Alimentation | 40.7 | 37.0 | 32.4 |
| Ornementation | 35.3 | 37.0 | 51.4 | |
| Cultural | 4.4 | 2.4 | 4.9 | |
| Medécinal | 19.6 | 23.6 | 11.3 | |
Table 4.
Morphological, biological, and propagation characteristics of domestic gardens species across different neighborhood types. Data are expressed in percentage. n refers to the number of domestic gardens.
Table 4.
Morphological, biological, and propagation characteristics of domestic gardens species across different neighborhood types. Data are expressed in percentage. n refers to the number of domestic gardens.
| Parameter | Form | Neighborhood type | ||
|---|---|---|---|---|
| Planned (n=89) |
Unplanned (n=90) |
Residential (n=94) |
||
| Morphological type | Liana | 2.2 | 0.8 | 0.7 |
| Woody | 18.6 | 22.0 | 19.0 | |
| Herbaceous | 79.2 | 77.2 | 80.3 | |
| Biological type | Phanerophyta | 29.7 | 33.7 | 30.9 |
| Chemephyta | 5.7 | 7.1 | 6.1 | |
| Hemicryptophyta | 3.4 | 4.1 | 4.4 | |
| Therophyta | 36.8 | 33.7 | 32.6 | |
| Epiphyta | 1.0 | 0.6 | 1.1 | |
| Hydrophyta | 0.5 | 0.6 | 0.0 | |
| Geophyta | 23.0 | 20.1 | 24.9 | |
| Propagation mode | Hydrochory | 0.0 | 0.0 | 0.6 |
| Anemochory | 12.0 | 11.0 | 11.0 | |
| Anthropochory | 86.0 | 87.0 | 85.1 | |
| Autochory | 1.0 | 1.0 | 1.1 | |
| Zoochory | 1.0 | 1.0 | 1.1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



