
Submitted:
04 September 2024
Posted:
05 September 2024
You are already at the latest version
Abstract
The stress-inducible heat shock protein 70 (Hsp70), which functions as a molecular chaperone and is frequently overexpressed in different cancer cell types, is presented on the cell surface of tumor cells and is actively released into the circulation in free and extracellular lipid vesicle-associated forms. Since the exact pathomechanism of endometriosis has not yet been elucidated but has been associated with the development of endometrial and ovarian cancer, we asked whether extracellular Hsp70 and circulating endometriotic cells (CECs) reflects the presence and development of endometriosis. Therefore, circulating levels of free and lipid microvesicle-associated Hsp70 were measured using the Hsp70-exo ELISA, and the presence of circulating CECs in the peripheral blood of patients with endometriosis was determined using membrane Hsp70 (mHsp70) and EpCAM monoclonal antibody (mAb)-based bead isolation approaches. Isolated CECs were further characterized by immunofluorescence using reagents directed against cytokeratin (epithelial), CD45 (leukocyte), CD105/CD44 (mesenchymal-stemness) and by comparative RNA analysis. Similar to the situation in patients with cancer, levels of circulating Hsp70 were elevated in the blood of patients with histologically proven endometriosis compared to a healthy control cohort, with the highest Hsp70 level in a patient with lymph node endometriosis. Moreover, CECs could be isolated using the cmHsp70.1 mAb- and to a lesser extent EpCAM mAb-based bead approaches in all patients with endometriosis. The longevity in cell culture and the expression of cytokeratin, CD105 and CD44, together with differentially expressed genes related to EMT reveals similarities of mHsp70 expressing CECs with circulating tumor cells (CTCs) and suggest a mesenchymal-stemness origin. These findings may support the involvement of mHsp70 positive, stem cell-like cells in the development of endometriotic lesions. In summary, elevated levels of Hsp70 in the blood and CECs could serve as liquid biopsy markers for endometriosis and help to elucidate the underlying pathomechanism of the disease.
Keywords:
endometriosis
; mesenchymal-stemness biomarker
; heat shock protein 70 (Hsp70)
1. Introduction
Worldwide, 10-15% of all women are affected by endometriosis, an estrogen-dependent benign gynecological disease. The disease is characterized by the presence of functional endometrial cells outside of the endometrium, predominantly in ovarian and other pelvic tissues, but also in more distant organs such as brain, lung and lymph nodes [1,2,3]. Due to the nonspecific symptoms of endometriosis, such as pain during menses, chronic pelvic pain and migraines, the diagnosis of endometriosis is often delayed by up to 10 years. Presently, only invasive laparoscopic methods can diagnose endometriosis in biopsies [4]. Therefore, there is a high unmet clinical need for reliable liquid biopsy-based biomarkers that can improve the early diagnosis of endometriosis which can be repeatedly analyzed by minimally invasive methods.
In addition to retrograde menstruation, immunopathological and endocrine factors as well as stem cells have been suggested to play a role in the development of endometriosis [3,5,6,7], however, the exact pathomechanism has not yet been elucidated. Since endometriotic cells are not locally restricted to the endometrium, but can migrate into extra-genital organs - most likely via the blood stream - we investigated the blood as a source of potential biomarkers of endometriosis.
Members of the HSP70 70kDa heat shock (stress) protein family of molecules reside in nearly all subcellular compartments such as the cytosol, nucleus, lysosomes, ER, mitochondria. The highly conserved, major stress-inducible Hsp70 is responsible for maintaining protein homeostasis, protein transport, antigen processing and prevention of cell death [8,9]. A high intracellular expression of Hsp70 in tumor cells is associated with tumor progression, anti-apoptotic signaling and therapy resistance [10,11]. In tumor cells, but not normal cells, Hsp70 can also be translocated from the cytosol to the plasma membrane by an ER-independent, non-classical, vesicular transport mechanism [9,12,13].Klicken oder tippen Sie hier, um Text einzugeben. It is well established that tumor cells overexpressing Hsp70 in the cytosol [10] and membrane Hsp70 (mHsp70) on the cell surface [14,15] Klicken oder tippen Sie hier, um Text einzugeben.have the capacity to release Hsp70 in extracellular lipid microvesicles with biophysical properties of exosomes [16]. Previous data from our group have demonstrated that elevated levels of Hsp70 in the blood can serve as a biomarker in a large variety of different tumor entities such as breast, lung, head and neck and brain cancers [17,18,19,20,21]. In this study, we analyzed Hsp70 levels in the circulation of patients with endometriosis and endometriotic bladder and lymph nodes to evaluate its role as a potential biomarker of disease.
Circulating tumor cells (CTCs) are most commonly isolated by antibody-based methods targeting the expression of epithelial adhesion molecule (EpCAM, CD326), which is crucial for the malignancy of epithelial tumors [22,23,24,25]. After entering the blood stream, CTCs migrating to different organs in the body extravasate into a different microenvironment. By evading the innate and adaptive immune response in the blood and being able to survive independently of other cells as a cell clone or colony, and by adapting to a new microenvironment, proliferating, and forming vessels, CTCs fulfil certain criteria that are essential for the spread of tumor cells [24], key properties of which might also enable distant organ involvement in endometriosis. CTCs appear in all major carcinoma groups [22,23,24] and the enumeration of CTCs in the blood of cancer patients during therapy is considered as a real-time “liquid biopsy” [26] that can serve as a prognostic marker for tumor diseases and therapeutic responsiveness [27]. It is well documented that the expression of EpCAM is frequently down-regulated after epithelial-to-mesenchymal transition (EMT) [28,29,30,31] and the functional transition of polarized epithelial cells into mobile mesenchymal cells [23,25,30]. Isolation of CTCs relying only on the cell surface expression of EpCAM on CTCs is therefore sub-optimal. We have previously shown that, unlike EpCAM, mHsp70, as detected using the cmHsp70.1 monoclonal antibody (mAb), remains stably expressed on the cell surface of CTCs after EMT [32], and that higher numbers of CTCs could be isolated from the blood of tumor patients using a cmHsp70.1 mAb-based bead approach compared to an equivalent EpCAM mAb-based bead approach [21].
Based on these findings, herein we asked whether free and microvesicle (exosomal)-associated Hsp70 in the blood, as well as circulating endometriotic cells (CECs), can serve as potential diagnostic biomarkers for patients with histologically confirmed endometriosis [33,34]. To address this question, soluble Hsp70 levels were measured using the Hsp70-exo ELISA [35], which is able to quantify free and vesicle-associated Hsp70, and CECs were enumerated after isolation with cmHsp70.1 mAb- and EpCAM mAb-based bead approaches from patients with endometriosis and extra-uterine endometriosis. Furthermore, CECs were characterized by immunofluorescence and RNA analysis with a focus on epithelial, mesenchymal and stem cell markers.
2. Material & Methods
2.1. Sample Collection
This pilot study analyzed EDTA anti-coagulated blood (2x 7.5 ml) from 12 female therapy-naïve patients within an age range of 15-50 with histologically confirmed endometriosis who were treated at the Department of Gynecology and Obstetrics, TUM University Hospital, School of Medicine and Health, Technical University Munich (TUM) and age-matched healthy donors. Blood was collected from the patients with histologically confirmed endometriosis prior to surgery. In 3 patients, blood samples could be collected before and 1 and 6 months after resection of endometriotic lesions by laparoscopy. The study was approved by the Ethical Committee of the TUM University Hospital, School of Medicine and Health (2023-407-S-NP). Before starting the study, written informed consent was obtained from all patients and normal donors. The study was performed in accordance with the guidelines of the Declaration of Helsinki 1975.
2.2. Cell Culture
The human cancer cell line JEG-3 (RRID:CVCL_0363; gestational choriocarcinoma), kindly provided by the group of Prof. Andreas Pichlmair (Institute of Virology, TUM), was cultured in RPMI Medium (Gibco Thermo Fisher Scientific, Waltham MA USA), supplemented with 10% v/v heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich, St. Louis MO USA) and antibiotics (100 IU/ml penicillin and 100 mg/ml streptomycin; Sigma-Aldrich, St. Louis MO USA), and kept in culture at 37°C with 5% v/v CO2 in a humidified atmosphere. The cell line was shown to be positive for mHsp70 expression by flow cytometry and immunofluorescence staining.
2.3. Measurement of Circulating Free and Lipid Vesicle-Associated Hsp70 Levels Using the Hsp70-Exo ELISA [35]
For plasma preparation, EDTA anti-coagulated whole blood (S-Monovette, Sarstedt, Nürmbrecht, Germany) was centrifuged at 1500xg for 15 min at room temperature and aliquots of plasma (300 µl) were stored at -80°C. For the Hsp70-exo ELISA, 96-well MaxiSorp Nunc-Immuno plates (Thermo, Rochester, NY, USA) were coated with the cmHsp70.2 coating mAb (1 µg/ml; multimmune GmbH (Munich, Germany)) in sodium carbonate buffer (0.1 M sodium carbonate, 0.1 M sodium hydrogen carbonate, pH 9.6; Sigma-Aldrich (Darmstadt, Germany)) overnight at room temperature. After rinsing the plates with washing buffer (phosphate-buffered saline (PBS), Life Technologies, Darmstadt, Germany) containing 0.05% v/v Tween-20 (Calbiochem, Merck, Darmstadt, Germany), the plates were blocked with liquid plate sealer (Condor Bioscience GmbH, Wangen i. Allgäu, Germany) for 30 min to prevent non-specific binding. The plates were washed and plasma samples (100 µl) and an 8 point Hsp70 standard (0-100 ng/ml), both diluted 1:5 in StabilZyme Select Stabilizer (Diarect GmbH, Freiburg i. Breisgau, Germany), were added. After 30 minutes incubation and washing, biotinylated cmHsp70.1 detection mAb (multimmune GmbH, Munich, Germany; 200 ng/ml) dissolved in HRP-Protector (Candor Biosciences GmbH, Wangen i. Allgäu, Germany) was added for 30 minutes, after which the plates were rinsed with washing buffer. After another incubation for 30 minutes with 57 ng/ml Streptavidin (Senova GmbH, Weimar, Germany) in HRP-ProtectorTM (Candor Biosciences GmbH), plates were incubated with the substrate reagent (100 µl) (BioFX TMB Super Sensitive One Component HRP Microwell Substrate, Surmodics, Inc., Eden Prairie, MN, USA) for 15 minutes, after which the reaction was stopped with 2N H2SO4 (50 µl) and the absorbances measured at 450 nm in a Microplate Reader (VICTOR X4 Multilabel Plate Reader, PerkinElmer, Waltham, MA, USA). Absorbance readings were corrected with the absorbance at 570 nm.
2.4. Isolation of Circulating Cells with cmHsp70.1 and EpCAM Antibody-Coupled S-pluriBeads
Isolation of circulating CECs from patients with endometriosis and healthy donors was performed according to a method described previously for CTCs [21,32]. Briefly, EDTA anti-coagulated blood (7.5 ml) was incubated with S-pluriBeads (PluriSelect Life Sciences, Leipzig, Germany) which were covalently coupled to the cmHsp70.1 (multimmune GmbH, Munich, Germany) or EpCAM (CD326, clone HEA 125; Origene/Acris GmbH, Herford, Germany) mAbs for 30 minutes under gentle rotation at room temperature. To reduce the number of erythrocytes in the sample, cells bound to the antibody-coupled beads were washed on a 30 µm pluriStrainer® with washing buffer (PluriSelect Life Sciences). CECs were detached from the beads by incubating with detachment buffer (PluriSelect Life Sciences) for 10 min at room temperature. The sample was then filtered through a sterile strainer, washed twice with medium, and incubated overnight at 37°C in a 48-well plate. After 24 h and again after 1 week, cells were counted using a Zeiss Axiovert microscope (Zeiss, Oberkoch, Germany). The isolated cells were kept in culture (DMEM Medium; Gibco Thermo Fisher Scientific, Waltham MA USA) supplemented with 1 nM β-estrogen (Invitrogen Merck, Darmstadt, Germany), 10% v/v heat-inactivated FBS (Sigma-Aldrich, St. Louis, MO, USA) and antibiotics (100 IU/ml penicillin and 100 mg/ml streptomycin; Sigma-Aldrich, St. Louis MO USA) for further analysis.
2.5. Immunohistochemical Staining
Adherent cells were cultivated in an 8-well chamber slide (Invitrogen, Thermo Fisher Scientific, Waltham MA, USA). After three washing steps in flow cytometry buffer (PBS; Life Technologies) containing 10% v/v FBS (Sigma Aldrich, St. Louis, MO, USA) cells were incubated with the following antibodies at room temperature for 30 minutes in the dark: cytokeratin cocktail, CK7 (LP5K, Millipore, Billerica, USA), CK19 (A53-B/A2, Exbio, Vestec, Czech Republic), panCK (C11, Exbio, Vestec, Czech Republic); CD45-Alexa Fluor® 488 (A4-120-C100, Exbio, Vestec, Czech Republic); CD105-PE (42A3, BioLegend, San Diego, CA, USA), CD44-FITC (C44Mab-5, BioLegend, San Diego, Ca, USA) all diluted in PBS (1:50). After another washing nuclei were counter-stained with Hoechst 33342 (1:1000; Invitrogen Thermo Fisher Scientific, Waltham MA, USA) for 5 minutes at room temperature. The cells were imaged on a Zeiss Axio Observer.Z1 microscope (Zeiss, Jena, Germany) at magnifications as indicated and image analysis was performed using AxioVision SE64 Rel.4.9. Multi-color images were produced by merging.
2.6. Flow Cytometry
JEG-3 cells were trypsinized and single cell suspensions washed twice with flow cytometry buffer (PBS; Life Technologies, containing 10% v/v FBS, Sigma Aldrich). Cells were then incubated with the following fluorescently-labeled antibodies: cmHsp70.1-FITC (multimmune GmbH, Munich, Germany); CD44-FITC (C44Mab-5, BioLegend, San Diego, CA, USA); CD9-PE (H19a, BioLegend, San Diego, USA); CD105-PE (43A3, BioLegend, San Diego, CA, USA) and cells (100,000 each) were incubated in the dark for 30 min on ice. After a final washing step, cells were resuspended in flow cytometry buffer and analyzed on a BD FACSCalibur™ flow cytometer (BD Biosciences, New Jersey, USA). Directly before the flow cytometric analysis, propidium iodide (1 µl/ 100 µl; Sigma-Aldrich, St. Louis MO, USA) was added to determine cell viability. Only PI-negative (viable) cells were gated and analyzed. An isotype-matched control antibody (BD Biosciences, New Jersey, USA) was used to determine non-specific staining.
2.7. RNA Preparation [36,37] and Analysis [38]
Library preparation for bulk-sequencing of poly(A)-RNA was performed as described previously [36]. A barcoded cDNA of each sample was generated using a Maxima RT polymerase (Thermo Fisher Scientific, Los Angeles, CA, USA) using oligo-dT primer containing barcodes, unique molecular identifiers (UMIs) and adaptor 5’-ends of the cDNAs were extended by a template switch oligo (TSO) and full-length cDNA was amplified with primers binding to the TSO-site and the adaptor. An NEBNext® Ultra® kit was used to fragment cDNA. After end repair and A-tailing, a TruSeq adaptor was ligated and 3’-end-fragments were finally amplified using primers with Illumina P5 and P7 overhangs. The library was sequenced on a NextSeq 500 (Illumina, Los Angeles, CA, USA) with 65 cycles for the cDNA in read1 and 19 cycles for the barcodes and UMIs in read2. Data were processed using the published Drop-seq pipeline (v1.0) to generate sample- and gene-wise UMI tables [37]. Reference genome (GRCh38) was used for alignment. Transcript and gene definitions were used according to GENCODE v38.
The raw sequencing reads were subjected to quality checks using FastQC (version 0.11.9) and were processed with Trimmomatic to trim low-quality bases and to remove adaptor sequences in FASTQ format. These clean paired-end reads were aligned to the human reference genome (GRCh38) using HISAT2 (version 2.0.5) with default parameters. Gene expression levels were quantified using featureCounts, generating raw count data that were subsequently normalized with DESeq2, which accounts for variations in sequencing depth and other technical factors. The DESeq2 R package in the Bioconductor platform was employed to identify differentially expressed genes (DEGs). DESeq2 filters genes based on their average expression across all samples, excluding those with mean normalized counts below a threshold that is automatically determined to maximize the number of detected genes, depending on user-defined criteria. Statistically significant DEGs were identified by comparing expression levels between cmHsp70.1 mAb- and EpCAM mAb-isolated CEC samples, with significance criteria set at an adjusted p value (padj) < 0.001 and an absolute log2 fold change greater than 1 (> 1).
To compare and analyze the RNA-seq results, we utilized multiple comprehensive and publicly available genomic and proteomic datasets. These resources provided a broad spectrum of gene expression across various cancer types, enabling robust validation and cross-referencing of our findings. Among the identified DEGs which were notably upregulated in mHsp70-positive and EpCAM-positive CECs were genes which are associated with epithelial-to-mesenchymal transition (EMT) pathways, stemness, proliferation, matrix remodelling, cell cycle regulation and adhesion.
2.8. Statistical Evaluations
For the statistical analysis of all dependent samples, the Wilcoxon test (Prism) and for all independent samples, the Mann-Whitney test (Prism) was performed, considering a 5% significance level.
3. Results
3.1. Circulating Free and Vesicle-Associated Hsp70 Levels in Patients with Endometriosis, Pre- and Post-Surgery and Healthy Controls
Free and lipid microvesicle-associated Hsp70 levels in plasma samples from 12 patients with histologically confirmed endometriosis before the start of any therapy were measured using the Hsp70-exo ELISA [35] (Table 1). In a subgroup of 3 patients (n=3) additional blood samples could be collected 1 month (patients #11, #12) and 6 months (patient #10) post-surgery. Although the soluble Hsp70 values in patients with histologically confirmed endometriosis showed a large variation (range, 0 – 1171 ng/ml), the mean Hsp70 levels (mean, 263 ± 345 ng/ml; median, 158 ng/ml) differed from those of a healthy control cohort (n=108, 35 ng/ml; p=0.03). The highest soluble Hsp70 level was detected in a patient with lymph node endometriosis (#10, 1,171 ng/ml). This finding is in line with results obtained from patients with malignant tumors such as lung, brain and breast carcinomas [18,19,20,21,39], who also showed elevated soluble Hsp70 levels.
A comparison of the soluble Hsp70 levels in 3 patients with endometriosis from whom blood samples could be collected before surgery and in the follow-up period (after 1 month in patients #11, #12 and after 6 months in patient #10) shows a large drop from 510 ± 589 ng/ml (median: 313 ng/ml) to 38 ± 45 ng/ml Hsp70 after resection of the endometriotic lesions (Figure 1b). This finding suggests that soluble Hsp70 originates from endometriotic cells expressing mHsp70.
3.2. Circulating Endometriotic Cells (CECs) Can Be Isolated from the Blood of Patients with Confirmed Endometriosis by Bead-Based Approaches Targeting Membrane Hsp70 and EpCAM
After a successful validation of the cmHsp70.1 and EpCAM mAb-based bead approaches for isolating circulating tumor cells (CTCs) from EDTA anti-coagulated blood from patients with different tumor entities [21], we asked whether this approach could isolate circulating endometriotic cells (CECs) from patients with histologically confirmed endometriosis (n=10) and patients with extra-uterine endometriosis (n=5). As a negative control, the EDTA blood of 2 healthy donors (n=2) was used for the isolation approach. As summarized in Table 1, CECs could be isolated from the EDTA blood of all patients (n=12) with endometriosis, whereas no cells were isolated from the blood of the healthy individuals (n=2). The mean number of cells isolated from 7.5 ml EDTA blood using cmHsp70.1 and EpCAM mAb-conjugated beads was 21,442 ± 77,711 (median, 258 cells) and 305 ± 418 (median, 178 cells), respectively. These results indicate a drastically higher CEC yield using cmHsp70.1 mAb-conjugated beads for the isolation. A comparison of the number of CECs in patients pre- and 1-month post-surgery indicated a drop in CECs from 890 to 219 (patient #11), 467 to 123 (patient #12) using the cmHsp70.1 mAb-based bead approach, and from 435 to 384 (patient #11), 384 to 156 (patient #12) using the EpCAM mAb-based bead approach. Unexpectedly, 6 months post-surgery, the number of CECs isolated with the cmHsp70.1 mAb-based bead approach increased from 12,935 to 186,648 in patient #10, but dropped from 379 to 6 when the EpCAM mAb-based bead isolation was used. These data indicate that the origin of CECs isolated via membrane-bound Hsp70 or EpCAM might differ.
3.3. Morphological Characterization of Cultured CECs from Patients with Endometriosis without and with Extra-Uterine Involvement
Freshly isolated CECs from all patients using cmHsp70.1 and EpCAM mAb-conjugated beads were cultured in DMEM medium supplemented with estrogen at 37°C in 48-well plates. Interestingly, after 1 week in cell culture, all CECs isolated with EpCAM mAb-based beads lost their viability, whereas those isolated with cmHsp70.1 mAb beads remained viable and became plastic-adherent.
Representative examples of cultured CECs derived from the blood of patient #04 (a) with histologically confirmed endometriosis after 1 week in cell culture, and patients #07 (b) and #10 (c) (both 1 month in cell culture) with endometriosis outside the uterine cavity, which were isolated via mHsp70 targeting (left) and EpCAM (right) targeting, are illustrated in Figure 2. A comparison of the morphology of cmHsp70.1 mAb-bead isolated cells derived from these three patients revealed a larger cell diameter (10-20 µm versus 3-5 µm) in the cells of patients #07 and #10 with endometriosis outside the uterine cavity versus patient #04 without extra-uterine involvement. Moreover, the viability in cell culture was much longer (up to 9 months) in mHsp70 expressing CECs derived from patients #7 and #10 with extra-uterine involvement compared to those derived from patient #4 (less than 1 month) without distant organ involvement.
3.4. Immunofluorescent Characterization of Isolated CEC from a Patient with Bladder Endometriosis
The CECs isolated from the blood of patient #07 with endometriotic lesions in the bladder using the cmHsp70.1 mAb-based bead approach were further characterized by immunofluorescence staining. As illustrated in Figure 3a, cells isolated using cmHsp70.1 mAb-based beads were positively stained for the typical epithelial marker cytokeratin (green), but were negative for the leukocyte marker CD45 (red). This finding shows that circulating cells derived from the blood of endometriosis patients share some typical characteristics with circulating tumor cells (CTCs).
To further characterize the biological nature of the circulating cells derived from patient #07, mesenchymal and stemness (CD105, CD44) markers were analyzed by immunofluorescence. As shown in Figure 3, the cmHsp70.1 mAb-isolated cells positively stained for the mesenchymal-stemness markers CD105 (Figure 3b) and CD44 (Figure 3c). Due to the limited number of cells, it was only possible to undertake immunofluorescence staining of CECs from patient #7.
The presence of the epithelial marker cytokeratin was confirmed by immunofluorescence staining of the gestational choriocarcinoma cell line JEG-3 which shares some characteristics with primary endometriotic cells (Figure 4a). This cell line also expresses mHsp70, the mesenchymal-stemness marker CD105, and the epithelial marker CD9, a tetraspanin, but not CD44 on the cell surface of viable cells (Figure 4b), as determined by flow cytometry.
3.5. RNA Analysis of Isolated CECs from a Patient with Lymph Node Endometriosis
A high-throughput RNA sequencing and data analysis using DESeq2 package in R was performed on CECs isolated from patient #10 with lymph node endometriosis using the cmHsp70.1 mAb- and EpCAM mAb-based bead approaches to determine differentially expressed genes. CECs of patient #10 were chosen because only in this patient the number of isolated cells was sufficient for these analyses. A total number of 430 genes were differentially regulated (Supplementary File S1, Figure 5) in EpCAM and cmHsp70.1 mAb-isolated CECs. Table 2 summarizes the results for the RNA analysis in which an adjusted p value (padj) was < 0.001 and an absolute log2 fold change value was greater 1 in CECs isolated with cmHsp70 mAb- and EpCAM mAb-conjugated beads.
Genes differentially expressed by CECs isolated with the EpCAM mAb- versus cmHsp70.1 mAb-conjugated bead approach were related to the proto-oncogene Src and kinases (SRC, FGR) [40], cell cycle regulators (RGCC, CDK5RAP3, CDKN1A) and matrix metalloproteinases (MMP2-ASI) [41] (Table 2), which regulate progression and invasiveness via cytoskeletal reorganization and extracellular matrix remodeling, cell adhesion and migration (ITGAM, ITGAE) in human melanoma [42].
Genes differentially expressed by CECs isolated using the cmHsp70.1 mAb- versus EpCAM mAb-based approach such as ROCK2 [43], STAT3 [44] and CD82 were related to EMT, stemness and endometrial cell proliferation [45] (Table 2), which is regulated via an up-regulated CD44/STAT3 signaling in tumor cells [46]. Down-regulated STAT3 signaling has been associated with fewer endometriotic lesions in an endometriotic mouse model [47] and in human endometriotic tissues [48]. A potential link between STAT3 and EMT/stemness markers has been shown by a positivity of CECs isolated via mHsp70 with the markers CD44 and CD105 (Figure 3).
4. Discussion
As invasive methods are currently the only option for diagnosing endometriosis [1,2,3,4], there is a high unmet clinical need to establish reliable liquid biopsy-based biomarker approaches, which can improve the diagnosis of the disease. Since endometriosis is a disease with an occurrence in extra-uterine organs such as, for example, the brain, the lung, the bladder or lymph nodes [1,2,3,4] and endometriosis-affected tissues share some pathological characteristics with malignant tumors – inflammation, cell growth, angiogenesis, cell adhesion, cytokine production – [49], this pilot study examined the potential of soluble and mHsp70 as diagnostic targets. We could show that similar to tumor patients, the levels of free and vesicle-associated Hsp70, in the circulation of patients with confirmed endometriosis were higher than those in a healthy control cohort [17,18,19,20,21], however with large variations. With a mean soluble Hsp70 concentration of 263 ng/ml, the results for endometriosis patients were comparable to those in tumor patients [17,18,19,20,21,39], and were higher than in a healthy control group (35 ng/ml). This finding supports the hypothesis that Hsp70 might also play a role in endometriosis. Previous studies have indicated that elevated Hsp70 levels in the blood are associated with an unfavorable tumor prognosis and a higher risk for developing distant metastases in different mammalian species [20,39,50]. In our small cohort, we could demonstrate that the highest soluble Hsp70 levels were found in patients with lymph node and bladder endometriosis indicating that an involvement of extra-uterine organs might be associated with the severity of the disease. Further studies in a larger number of patients are required in order to better understand any relationship(s) between levels of soluble Hsp70 and clinical status.
It has also been demonstrated that only tumor cells expressing mHsp70 actively release Hsp70 in extracellular lipid vesicles with proven biophysical characteristics of exosomes [16], and that high Hsp70 levels in the blood are associated with therapy resistance in patients with cancer [35,51]. Moreover, the density of mHsp70 expression on cancer metastases appears higher than that on the corresponding primary tumor [40]. Since the levels of circulating free and lipid vesicle-associated Hsp70 are comparable in patients with malignant tumors and endometriosis, and that endometriosis also occurs in extra-uterine organs including lymph nodes and bladder, we speculate that soluble Hsp70 levels might originate from the cell membrane of endometriotic cells. This hypothesis is supported by the finding that soluble Hsp70 levels dropped drastically after laparoscopic resection of endometriotic lesions, similar to reduced levels in tumor patients after successful therapies which decrease tumor size [52]. Due to the lack of freshly resected tissue, the final proof for a mHsp70 positivity on primary endometriotic cells could not be provided in this study. However, a strong mHsp70 positivity could be demonstrated on the gestational choriocarcinoma cell line JEG-3, which resembles certain characteristics of endometriotic cells, but is nevertheless a tumor cell. Future studies with larger patient cohorts and access to resected endometriotic tissue are necessary to confirm mHsp70 expression on endometriotic cells.
Since the exact mechanism which results in the occurrence of endometriotic cells in extra-uterine organs has not yet been elucidated, we asked whether CECs might be involved in this process. Therefore, we isolated CECs from EDTA anti-coagulated blood of patients with confirmed endometriosis and an extra-uterine involvement using a bead-based approach which is applied for the isolation of CTCs by targeting mHsp70 or EpCAM and which employs cmHsp70.1 mAb- and EpCAM mAb-conjugated beads, respectively. Interestingly, CECs could be isolated with both mAb-conjugated bead systems from all blood samples of endometriosis patients, with drastically higher cell yields with mHsp70 as a selection target and in patients with extra-uterine endometriosis. Moreover, CECs isolated from the latter patients with cmHsp70.1 mAb beads, in contrast to EpCAM mAb beads, could be cultured, remained viable for several months and were positive for the typical CTC marker cytokeratin, the mesenchymal-stemness markers CD44 and CD105, but were negative for the leukocyte marker CD45. These CTC characteristics might explain the occurrence of endometriotic cells in distant organs, since these cells are essential in the metastatic spread in tumor patients [22,24,53,54].
The morphology of CECs from patients with endometriotic bladder and lymph nodes isolated using cmHsp70.1 mAb beads differed from those of other endometriosis patients as they were rounder in shape and larger in size. Besides the altered morphology, these CECs tended to grow in cell clusters and could be propagated in cell culture for a longer period of time (up to 9 months). Due to these differences [55] and due to their molecular features, we propose that the cells isolated from the patient with endometriotic bladder and lymph nodes can be considered as being “CTC-like”. However, it has yet to be demonstrated whether these characteristics are related/explain the perceived relationship(s) between endometriosis and gynecological cancers [56,57,58].
It is known that EpCAM expression on the surface of CTCs is reduced after EMT, whereas mHsp70 expression remains stably expressed after EMT [25,26,27,28,29,30,32]. Since the cell yield after isolation with cmHsp70.1 mAb-conjugated beads was much higher than after isolation with EpCAM mAb-conjugated beads, we hypothesize a mesenchymal origin of the circulating CECs with mHsp70 as a target structure. To further examine possible mesenchymal characteristics of cells isolated using the cmHsp70.1 mAb-conjugated beads from patients with confirmed endometriosis, isolated CECs were immunofluorescently stained for typical mesenchymal and stem cell markers such as CD105 and CD44 as well as for the epithelial marker cytokeratin. The gestational choriocarcinoma cell line JEG-3 served as a control for the validation of the CTC markers. A positive staining of the isolated CECs for these mesenchymal-stemness markers CD44, a hyaluronic acid receptor, and CD105, a coreceptor for TGF-β, indicate stem cell characteristics of these cells [59]. Together with a positive staining for cytokeratin [60,61] and mHsp70, the cultured endometriotic cells share some characteristics with tumor cells, which supports the hypothesis of certain malignant traits of the CECs isolated from endometriosis patients. This is in line with the finding that CECs isolated based on mHsp70 expression remained viable in cell culture for long time periods, whereas those isolated based on their expression of EpCAM lost viability in cell culture.
CECs isolated from patient #10 with endometriotic lymph nodes based on EpCAM and mHsp70 expression were profiled for the expression of differentially regulated genes. It appeared that differentially expressed genes of CECs isolated with EpCAM mAb-conjugated beads were found to be related to proto-oncogenes, cell cycle regulation [40], cytoskeletal reorganization, extracellular matrix remodeling [41] and cell adhesion [49]. In cmHsp70.1 mAb-isolated CECs differentially expressed genes were related to markers for EMT [62,63,64], stemness [65] and endometrial cell proliferation, which is regulated via an up-regulation of CD44 and STAT3 signaling [45]. These data are in line with a positive immunofluorescence staining of CD44 in mHsp70 targeted CECs (Figure 3). Although not significantly different, the expression of genes encoding for NCAM1 (CD56), a neuronal cellular adhesion molecule which is described as a signal transducer for the regulation of cell growth, migration, proliferation, apoptosis and differentiation [66] which reflect pathological features of tumor cells [49], as well as additional EMT markers such as ALCAM, ITGB1, and ITGA6 [66,67,68,69] were also differentially expressed in cmHsp70.1 bead-isolated CECs. These findings support the assumption of pathological characteristics in circulating cells that were isolated using cmHsp70.1 mAb-conjugated beads.
5. Conclusion
The occurrence of endometriotic cells in extra-uterine organs stimulated us to determine extracellular Hsp70 concentrations and the presence and prevalence of CECs in the plasma of endometriosis patients as potential biomarkers for a minimal invasive diagnosis of the disease. Similar to tumor patients, we could demonstrate elevated soluble Hsp70 levels and the presence of CECs with characteristics of CTCs in the blood of a small cohort of endometriosis patients. The data warrant further studies in larger patient cohorts and the access to resected endometriotic tissue.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary file S1. Raw data file of DEGs.
Author Contributions
Conceptualization, G.M. and V.S.-K.; methodology, C.G., S.H., L.B., A.P., A.B.D.; software, V.R., R.Ö.; validation, G.M., A.G.P; formal analysis, M-N.T., B.W.; investigation, C.G.; resources, G.M.; data curation, G.M., A.G.P.; writing—original draft preparation, C.G.; writing—review and editing, C.G.; visualization, G.M., V.S.-K.; supervision, G.M.; project administration, V.S.-K.; funding acquisition, G.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by BMBF, grant number 02NUK064A and BAYCELLator, grant number AZ-1568-22.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of the Klinikum rechts der Isar, Technical University Munich (protocol code 2023-407-S-NP, 24.07.203).
Informed Consent Statement
Written informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4413017/), https://www.biocompare.com/Editorial-Articles/597087-A-Guide-to-Mesenchymal-Stem-MSC-markers/).
Acknowledgments
The authors want to thank Anett Lange for editorial help.
Conflicts of Interest
The authors declare no conflicts of interest. The author G.M. is CSO of multimmune GmbH, Munich which provided the patented Hsp70-exo ELISA. The author A.G.P. is CEO of multimmune GmbH.
References
- Tennfjord, M.K.; Gabrielsen, R.; Tellum, T. Effect of physical activity and exercise on endometriosis-associated symptoms: A systematic review. BMC Womens Health. 2021, 21, 355. [Google Scholar] [CrossRef]
- Bulun, S.E.; Yilmaz, B.D.; Sison, C.; Miyazaki, K.; Bernardi, L.; Liu, S.; Kohlmeier, A.; Yin, P.; Milad, M.; Wei, J. Endometriosis. Endocr Rev. 2019, 40, 1048–1079. [Google Scholar] [CrossRef] [PubMed]
- Mehedintu, C.; Plotogea, M.N.; Ionescu, S.; Antonovici, M. Endometriosis still a challenge. J Med Life. 2014, 7, 349–357. [Google Scholar] [PubMed]
- Mounsey, A.L.; Wilgus, A.; Slawson, D.C. Diagnosis and management of endometriosis. Am Fam Physician. 2006, 74, 594–600. [Google Scholar] [PubMed]
- Chan, R.W.; Schwab, K.E.; Gargett, C.E. Clonogenicity of human endometrial epithelial and stromal cells. Biol Reprod. 2004, 70, 1738–1750. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Z.; Yang, F.; Wang, H.; Liang, S.; Wang, H.; Yang, J.; Lin, J. The role of endometrial stem cells in the pathogenesis of endometriosis and their application to its early diagnosisdagger. Biol Reprod. 2020, 102, 1153–1159. [Google Scholar] [CrossRef]
- Sampson, J.A. Metastatic or embolic endometriosis, due to the menstrual dissemination of endometrial tissue into the venous circulation. Am J Pathol. 1927, 3, 93-110 143.
- Saibil, H. Chaperone machines for protein folding, unfolding and disaggregation. Nat Rev Mol Cell Biol. 2013, 14, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Radons, J.; Multhoff, G. Immunostimulatory functions of membrane-bound and exported heat shock protein 70. Exerc.Immunol.Rev. 2005, 11, 17–33. [Google Scholar]
- Albakova, Z.; Armeev, G.A.; Kanevskiy, L.M.; Kovalenko, E.I.; Sapozhnikov, A.M. Hsp70 multi-functionality in cancer. Cells. 2020, 9. [Google Scholar] [CrossRef]
- Vostakolaei, M.A.; Hatami-Baroogh, L.; Babaei, G.; Molavi, O.; Kordi, S.; Abdolalizadeh, J. Hsp70 in cancer: A double agent in the battle between survival and death. J Cell Physiol. 2021, 236, 3420–3444. [Google Scholar] [CrossRef]
- Vega, V.L.; Rodriguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; De Maio, A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. J.Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef]
- Gehrmann, M.; Liebisch, G.; Schmitz, G.; Anderson, R.; Steinem, C.; De, M.A.; Pockley, G.; Multhoff, G. Tumor-specific Hsp70 plasma membrane localization is enabled by the glycosphingolipid Gb3. PLoS.ONE. 2008, 3, e1925. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G.; Botzler, C.; Wiesnet, M.; Muller, E.; Meier, T.; Wilmanns, W.; Issels, R.D. A stress-inducible 72-kda heat-shock protein (hsp72) is expressed on the surface of human tumor cells, but not on normal cells. Int.J.Cancer. 1995, 61, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G.; Botzler, C.; Jennen, L.; Schmidt, J.; Ellwart, J.; Issels, R. Heat shock protein 72 on tumor cells: A recognition structure for natural killer cells. J.Immunol. 1997, 158, 4341–4350. [Google Scholar] [CrossRef] [PubMed]
- Gastpar, R.; Gehrmann, M.; Bausero, M.A.; Asea, A.; Gross, C.; Schroeder, J.A.; Multhoff, G. Heat shock protein 70 surface-positive tumor exosomes stimulate migratory and cytolytic activity of natural killer cells. Cancer Res. 2005, 65, 5238–5247. [Google Scholar] [CrossRef] [PubMed]
- Seier, S.; Bashiri Dezfouli, A.; Lennartz, P.; Pockley, A.G.; Klein, H.; Multhoff, G. Elevated levels of circulating Hsp70 and an increased prevalence of CD94+/CD69+ NK cells is predictive for advanced stage non-small cell lung cancer. Cancers (Basel). 2022, 14. [Google Scholar] [CrossRef]
- Safi, S.; Messner, L.; Kliebisch, M.; Eggert, L.; Ceylangil, C.; Lennartz, P.; Jefferies, B.; Klein, H.; Schirren, M.; Dommasch, M.; et al. Circulating Hsp70 levels and the immunophenotype of peripheral blood lymphocytes as potential biomarkers for advanced lung cancer and therapy failure after surgery. Biomolecules. 2023, 13. [Google Scholar] [CrossRef]
- Lobinger, D.; Gempt, J.; Sievert, W.; Barz, M.; Schmitt, S.; Nguyen, H.T.; Stangl, S.; Werner, C.; Wang, F.; Wu, Z.; et al. Potential role of Hsp70 and activated NK cells for prediction of prognosis in glioblastoma patients. Front Mol Biosci. 2021, 8, 669366. [Google Scholar] [CrossRef]
- Rothammer, A.; Sage, E.K.; Werner, C.; Combs, S.E.; Multhoff, G. Increased heat shock protein 70 (Hsp70) serum levels and low NK cell counts after radiotherapy - potential markers for predicting breast cancer recurrence? Radiat Oncol. 2019, 14, 78. [Google Scholar] [CrossRef]
- Xanthopoulos, A.; Samt, A.K.; Guder, C.; Taylor, N.; Roberts, E.; Herf, H.; Messner, V.; Trill, A.; Holzmann, K.L.K.; Kiechle, M.; et al. Hsp70-a universal biomarker for predicting therapeutic failure in human female cancers and a target for CTC isolation in advanced cancers. Biomedicines. 2023, 11. [Google Scholar] [CrossRef]
- Scully, O.J.; Bay, B.H.; Yip, G.; Yu, Y. Breast cancer metastasis. Cancer Genomics Proteomics. 2012, 9, 311–320. [Google Scholar] [PubMed]
- Allard, W.J.; Matera, J.; Miller, M.C.; Repollet, M.; Connelly, M.C.; Rao, C.; Tibbe, A.G.; Uhr, J.W.; Terstappen, L.W. Tumor cells circulate in the peripheral blood of all major carcinomas but not in healthy subjects or patients with nonmalignant diseases. Clin Cancer Res. 2004, 10, 6897–6904. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science. 2011, 331, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P. Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer. 2002, 2, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Alix-Panabieres, C.; Pantel, K. Circulating tumor cells: Liquid biopsy of cancer. Clin Chem. 2013, 59, 110–118. [Google Scholar] [CrossRef]
- Cristofanilli, M. Circulating tumor cells, disease progression, and survival in metastatic breast cancer. Semin Oncol. 2006, 33, S9–14. [Google Scholar] [CrossRef]
- Nieto, M.A.; Huang, R.Y.; Jackson, R.A.; Thiery, J.P. Emt: 2016. Cell. 2016, 166, 21–45. [Google Scholar] [CrossRef]
- Cano, A.; Perez-Moreno, M.A.; Rodrigo, I.; Locascio, A.; Blanco, M.J.; Del Barrio, M.G.; Portillo, F.; Nieto, M.A. The transcription factor snail controls epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat Cell Biol. 2000, 2, 76–83. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat Rev Mol Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- He, P.; Dai, Q.; Wu, X. New insight in urological cancer therapy: From epithelial-mesenchymal transition (EMT) to application of nano-biomaterials. Environ Res. 2023, 229, 115672. [Google Scholar] [CrossRef]
- Breuninger, S.; Stangl, S.; Werner, C.; Sievert, W.; Lobinger, D.; Foulds, G.A.; Wagner, S.; Pickhard, A.; Piontek, G.; Kokowski, K.; et al. Membrane Hsp70-a novel target for the isolation of circulating tumor cells after epithelial-to-mesenchymal transition. Front Oncol. 2018, 8, 497. [Google Scholar] [CrossRef]
- Annemiek, W.N. Pathogenesis of endometriosis. Best Practice & Research Clinical Obstetrics & Gynaecology. 2004, 18, 233-244.
- Sotnikova, N.Y.; Antsiferova, Y.S.; Posiseeva, L.V.; Shishkov, D.N.; Posiseev, D.V.; Filippova, E.S. Mechanisms regulating invasiveness and growth of endometriosis lesions in rat experimental model and in humans. Fertil Steril. 2010, 93, 2701–2705. [Google Scholar] [CrossRef]
- Werner, C.; Stangl, S.; Salvermoser, L.; Schwab, M.; Shevtsov, M.; Xanthopoulos, A.; Wang, F.; Dezfouli, A.B.; Tholke, D.; Ostheimer, C.; et al. Hsp70 in liquid biopsies - a tumor-specific biomarker for detection and response monitoring in cancer. Cancers (Basel). 2021, 13, 3706. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Lersch, R.; De Andrade Kratzig, N.; Strong, A.; Friedrich, M.J.; Weber, J.; Engleitner, T.; Ollinger, R.; Yen, H.Y.; Kohlhofer, U.; et al. In vivo interrogation of regulatory genomes reveals extensive quasi-insufficiency in cancer evolution. Cell Genom. 2023, 3, 100276. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell. 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. Clusterprofiler 4.0: A universal enrichment tool for interpreting omics data. Innovation (Camb). 2021, 2, 100141. https://doi.org/10.1016/j.xinn.2021.100141. [CrossRef]
- Gunther, S.; Ostheimer, C.; Stangl, S.; Specht, H.M.; Mozes, P.; Jesinghaus, M.; Vordermark, D.; Combs, S.E.; Peltz, F.; Jung, M.P.; et al. Correlation of Hsp70 serum levels with gross tumor volume and composition of lymphocyte subpopulations in patients with squamous cell and adeno non-small cell lung cancer. Front Immunol. 2015, 6, 556. [Google Scholar] [CrossRef] [PubMed]
- Manek, R.; Pakzamir, E.; Mhawech-Fauceglia, P.; Pejovic, T.; Sowter, H.; Gayther, S.A.; Lawrenson, K. Targeting src in endometriosis-associated ovarian cancer. Oncogenesis. 2016, 5, e251. [Google Scholar] [CrossRef]
- Kleimenova, T.; Polyakova, V.; Linkova, N.; Drobintseva, A.; Medvedev, D.; Krasichkov, A. The expression of kisspeptins and matrix metalloproteinases in extragenital endometriosis. Biomedicines. 2024, 12. [Google Scholar] [CrossRef]
- Lenci, R.E.; Rachakonda, P.S.; Kubarenko, A.V.; Weber, A.N.; Brandt, A.; Gast, A.; Sucker, A.; Hemminki, K.; Schadendorf, D.; Kumar, R. Integrin genes and susceptibility to human melanoma. Mutagenesis. 2012, 27, 367–373. [Google Scholar] [CrossRef]
- Huang, Z.X.; Mao, X.M.; Wu, R.F.; Huang, S.M.; Ding, X.Y.; Chen, Q.H.; Chen, Q.X. Rhoa/rock pathway mediates the effect of oestrogen on regulating epithelial-mesenchymal transition and proliferation in endometriosis. J Cell Mol Med. 2020, 24, 10693–10704. [Google Scholar] [CrossRef]
- Zhang, G.; Hou, S.; Li, S.; Wang, Y.; Cui, W. Role of STAT3 in cancer cell epithelial-mesenchymal transition (review). Int J Oncol. 2024, 64. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wu, F.; Qin, C.; Lin, Y. Paradoxical role of phosphorylated STAT3 in normal fertility and the pathogenesis of adenomyosis and endometriosisdagger. Biol Reprod. 2024, 110, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Keyoumu, K.; Yu, R.; Wen, D.; Jiang, H.; Liu, X.; Di, X.; Zhang, S. Extracellular matrix marker LAMC2 targets ZEB1 to promote tnbc malignancy via up-regulating CD44/STAT3 signaling pathway. Mol Med. 2024, 30, 61. [Google Scholar] [CrossRef] [PubMed]
- Inui, H.; Kawakita, T.; Murayama, M.; Nakagawa, T.; Sasada, H.; Shinohara, A.; Aragaki, R.; Kagawa, T.; Kadota, Y.; Kato, T.; et al. Effects of STAT inhibitors in mouse models of endometriosis. Reprod Sci. 2023, 30, 2449–2456. [Google Scholar] [CrossRef]
- Choi, J.; Jo, M.; Lee, E.; Kim, S.E.; Lee, D.Y.; Choi, D. Dienogest attenuates stat3 activation in ovarian endometriotic cysts. Eur J Obstet Gynecol Reprod Biol. 2024, 294, 217–221. [Google Scholar] [CrossRef]
- Chopyak, V.V.; Koval, H.D.; Havrylyuk, A.M.; Lishchuk-Yakymovych, K.A.; Potomkina, H.A.; Kurpisz, M.K. Immunopathogenesis of endometriosis - a novel look at an old problem. Cent Eur J Immunol. 2022, 47, 109–116. [Google Scholar] [CrossRef]
- Salvermoser, L.; Flisikowski, K.; Dressel-Bohm, S.; Nytko, K.J.; Rohrer Bley, C.; Schnieke, A.; Samt, A.K.; Tholke, D.; Lennartz, P.; Schwab, M.; et al. Elevated circulating Hsp70 levels are correlative for malignancies in different mammalian species. Cell Stress Chaperones. 2022. [CrossRef]
- Lee, H.W.; Lee, E.H.; Kim, S.H.; Roh, M.S.; Jung, S.B.; Choi, Y.C. Heat shock protein 70 (Hsp70) expression is associated with poor prognosis in intestinal type gastric cancer. Virchows Arch. 2013, 463, 489–495. [Google Scholar] [CrossRef]
- Botzler, C.; Schmidt, J.; Luz, A.; Jennen, L.; Issels, R.; Multhoff, G. Differential Hsp70 plasma-membrane expression on primary human tumors and metastases in mice with severe combined immunodeficiency. Int.J.Cancer. 1998, 77, 942–948. [Google Scholar] [CrossRef]
- Chanteloup, G.; Cordonnier, M.; Isambert, N.; Bertaut, A.; Hervieu, A.; Hennequin, A.; Luu, M.; Zanetta, S.; Coudert, B.; Bengrine, L.; et al. Monitoring Hsp70 exosomes in cancer patients' follow up: A clinical prospective pilot study. J Extracell Vesicles. 2020, 9, 1766192. [Google Scholar] [CrossRef]
- Yu, M.; Stott, S.; Toner, M.; Maheswaran, S.; Haber, D.A. Circulating tumor cells: Approaches to isolation and characterization. J Cell Biol. 2011, 192, 373–382. [Google Scholar] [CrossRef]
- Riethdorf, S.; Fritsche, H.; Muller, V.; Rau, T.; Schindlbeck, C.; Rack, B.; Janni, W.; Coith, C.; Beck, K.; Janicke, F.; et al. Detection of circulating tumor cells in peripheral blood of patients with metastatic breast cancer: A validation study of the cellsearch system. Clin Cancer Res. 2007, 13, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Kralickova, M.; Lagana, A.S.; Ghezzi, F.; Vetvicka, V. Endometriosis and risk of ovarian cancer: What do we know? Arch Gynecol Obstet. 2020, 301, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brilhante, A.V.; Augusto, K.L.; Portela, M.C.; Sucupira, L.C.; Oliveira, L.A.; Pouchaim, A.J.; Nobrega, L.R.; Magalhaes, T.F.; Sobreira, L.R. Endometriosis and ovarian cancer: An integrative review (endometriosis and ovarian cancer). Asian Pac J Cancer Prev. 2017, 18, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Barnard, M.E.; Farland, L.V.; Yan, B.; Wang, J.; Trabert, B.; Doherty, J.A.; Meeks, H.D.; Madsen, M.; Guinto, E.; Collin, L.J.; et al. Endometriosis typology and ovarian cancer risk. JAMA. 2024. [CrossRef]
- Kim, W.T.; Ryu, C.J. Cancer stem cell surface markers on normal stem cells. BMB Rep. 2017, 50, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Barak, V.; Goike, H.; Panaretakis, K.W.; Einarsson, R. Clinical utility of cytokeratins as tumor markers. Clin Biochem. 2004, 37, 529–540. [Google Scholar] [CrossRef]
- Lane, E.B.; Alexander, C.M. Use of keratin antibodies in tumor diagnosis. Semin Cancer Biol. 1990, 1, 165–179. [Google Scholar]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells - current trends and future prospective. Biosci Rep. 2015, 35. [Google Scholar] [CrossRef]
- Miller-Rhodes, P. A guide to mesenchymal stem cell (msc) markers. 2023,.
- Wu, L.; Amjad, S.; Yun, H.; Mani, S.; De Perrot, M. A panel of emerging EMT genes identified in malignant mesothelioma. Sci Rep. 2022, 12, 1007. [Google Scholar] [CrossRef]
- Nielsen, J.S.; Mcnagny, K.M. Novel functions of the CD34 family. J Cell Sci. 2008, 121, 3683–3692. [Google Scholar] [CrossRef]
- Li, J.; Yang, R.; Yang, H.; Chen, S.; Wang, L.; Li, M.; Yang, S.; Feng, Z.; Bi, J. NCAM regulates the proliferation, apoptosis, autophagy, EMT, and migration of human melanoma cells via the Src/Akt/mTOR/Cofilin signaling pathway. J Cell Biochem. 2020, 121, 1192–1204. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.; Wei, F.; Zhang, J.; Li, B. Mir-134 suppresses the migration and invasion of non-small cell lung cancer by targeting ITGB1. Oncol Rep. 2017, 37, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Zeng, J.J.; Yang, Y.; Ruge, F.; Lane, J.; Hargest, R.; Jiang, W.G. Expression of ALCAM in clinical colon cancer and relationship with patients' treatment responses. In Vivo. 2023, 37, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wan, Y.; Su, Z.; Li, J.; Han, M.; Zhou, C. Mesenchymal stem cell-derived exosomal microrna-3940-5p inhibits colorectal cancer metastasis by targeting integrin alpha6. Dig Dis Sci. 2021, 66, 1916–1927. [Google Scholar] [CrossRef]
Figure 1.
Soluble Hsp70 levels in the blood of patients with endometriosis and healthy controls. Free and lipid vesicle-associated Hsp70 levels [ng/ml] in the plasma of patients with histologically confirmed endometriosis (n=12) and a healthy cohort (n=108) (a), and selected patients with endometriosis pre-and post-surgery (n=3) (b) as determined using the Hsp70-exo ELISA.
Figure 1.
Soluble Hsp70 levels in the blood of patients with endometriosis and healthy controls. Free and lipid vesicle-associated Hsp70 levels [ng/ml] in the plasma of patients with histologically confirmed endometriosis (n=12) and a healthy cohort (n=108) (a), and selected patients with endometriosis pre-and post-surgery (n=3) (b) as determined using the Hsp70-exo ELISA.
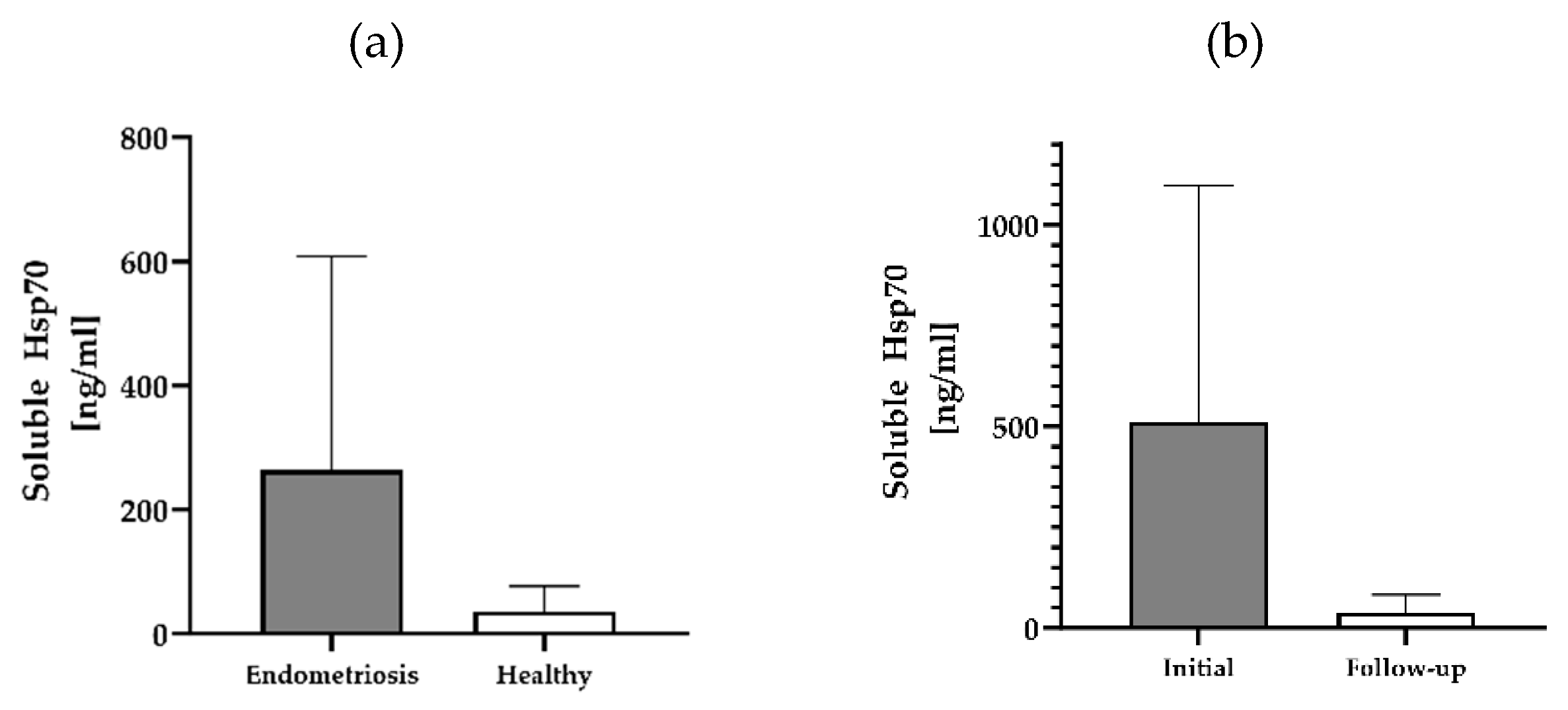
Figure 2.
Morphology of cultured CECs. Morphology of cultured CECs isolated from the EDTA anti-coagulated blood (7.5 ml) of patient #04 (a) with histologically confirmed endometriosis (1 week in cell culture) and patients #07 (b) and #10 (c) with confirmed endometriosis and distant organ involvement (1 month in cell culture) using cmHsp70.1 (left) and EpCAM (right) mAb-conjugated beads. Magnification 20x; scale bar: 100 µm.
Figure 2.
Morphology of cultured CECs. Morphology of cultured CECs isolated from the EDTA anti-coagulated blood (7.5 ml) of patient #04 (a) with histologically confirmed endometriosis (1 week in cell culture) and patients #07 (b) and #10 (c) with confirmed endometriosis and distant organ involvement (1 month in cell culture) using cmHsp70.1 (left) and EpCAM (right) mAb-conjugated beads. Magnification 20x; scale bar: 100 µm.
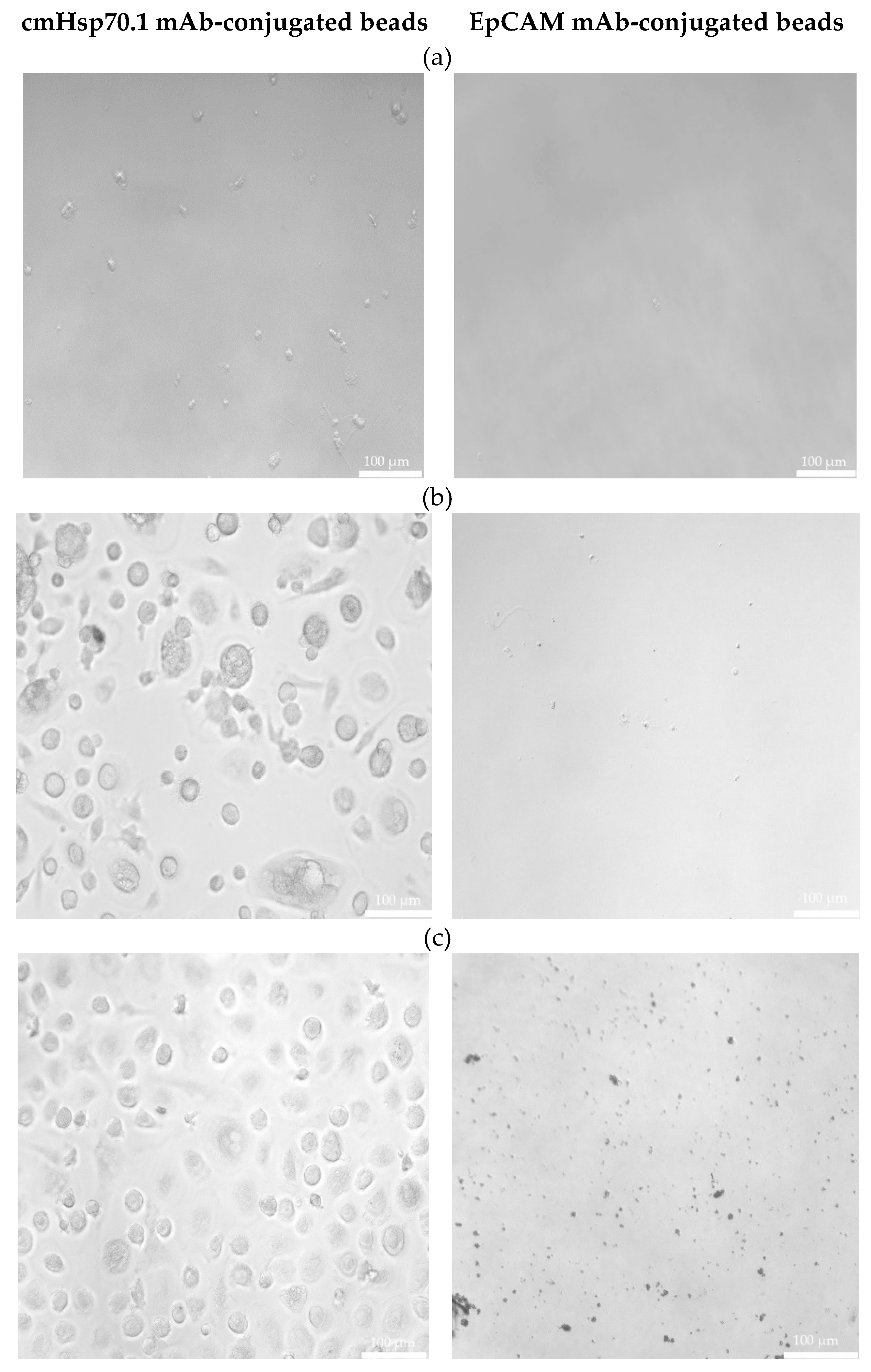
Figure 3.
Representative immunofluorescence photomicrographs of cultured CECs. Immunofluorescent images of isolated cells from endometriosis patient #07 using cmHsp70 mAb-conjugated beads cultured for 7 days and stained with Hoechst 33342 (1:1000; a-c), FITC-labeled cytokeratin cocktail, PE-labeled CD45 (a), CD105-APC (b) and CD44-FITC (c), all diluted 1:50. Magnification: 10x (a), 20x (b, c); scale bar: 200 µm (a), 100 µm (b, c).
Figure 3.
Representative immunofluorescence photomicrographs of cultured CECs. Immunofluorescent images of isolated cells from endometriosis patient #07 using cmHsp70 mAb-conjugated beads cultured for 7 days and stained with Hoechst 33342 (1:1000; a-c), FITC-labeled cytokeratin cocktail, PE-labeled CD45 (a), CD105-APC (b) and CD44-FITC (c), all diluted 1:50. Magnification: 10x (a), 20x (b, c); scale bar: 200 µm (a), 100 µm (b, c).
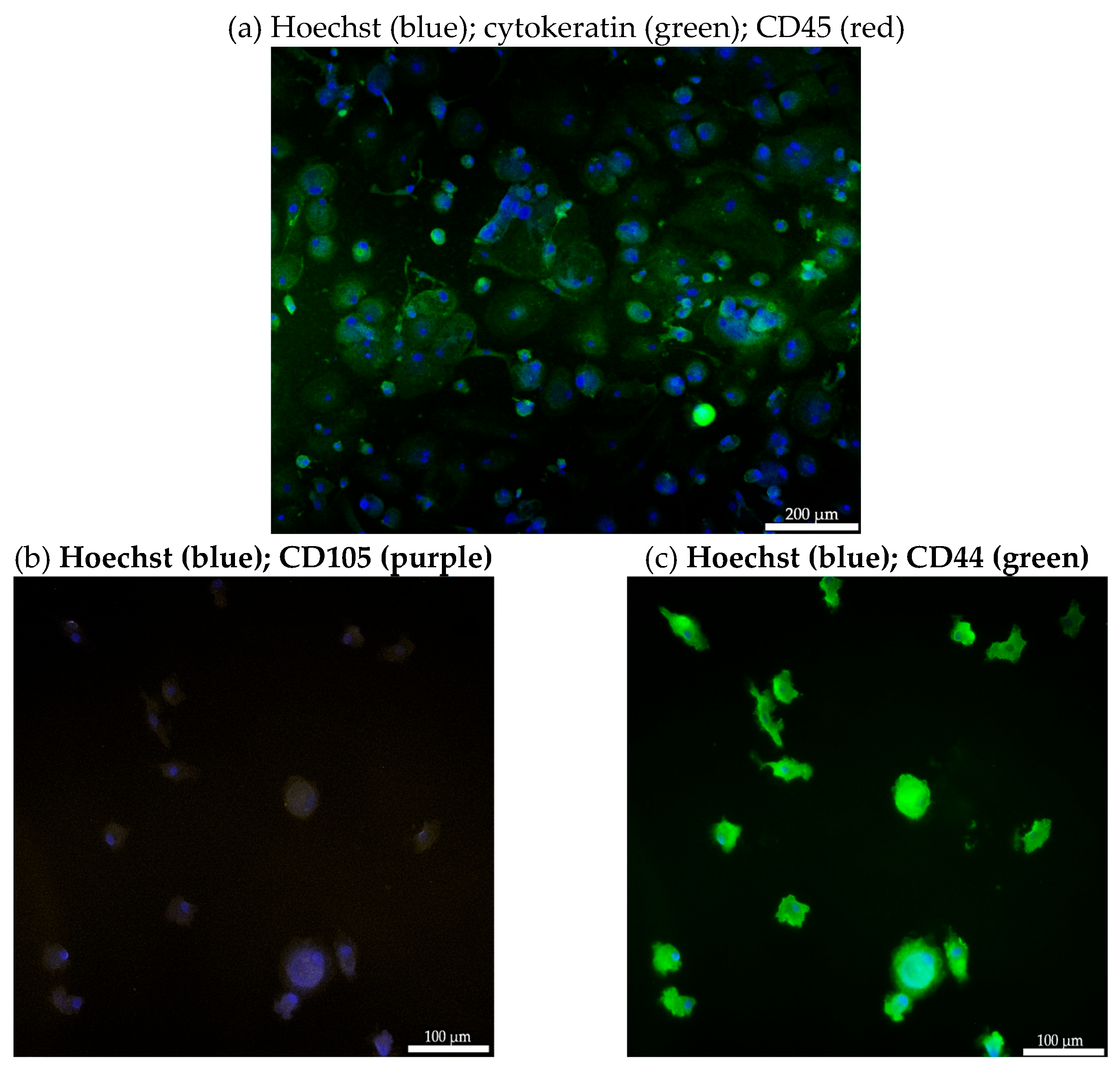
Figure 4.
Immunofluorescence and flow cytometric analysis of JEG-3 cells. The gestational choriocarcinoma cell line JEG-3 was stained with Hoechst 33342 (1:1000) and FITC-labeled cytokeratin cocktail (1:50). Magnification: 20x, scale bar: 50 µm (a). Representative histograms of mHsp70, CD105, CD44 and CD9 expression on viable JEG-3 cells after staining of viable JEG-3 cells with cmHsp70.1-FITC, CD105-PE, CD44-FITC and CD9-PE mAbs (b). The percentages of positively stained cells from 3 independent experiments ± standard deviation (SD) are shown in each histogram.
Figure 4.
Immunofluorescence and flow cytometric analysis of JEG-3 cells. The gestational choriocarcinoma cell line JEG-3 was stained with Hoechst 33342 (1:1000) and FITC-labeled cytokeratin cocktail (1:50). Magnification: 20x, scale bar: 50 µm (a). Representative histograms of mHsp70, CD105, CD44 and CD9 expression on viable JEG-3 cells after staining of viable JEG-3 cells with cmHsp70.1-FITC, CD105-PE, CD44-FITC and CD9-PE mAbs (b). The percentages of positively stained cells from 3 independent experiments ± standard deviation (SD) are shown in each histogram.
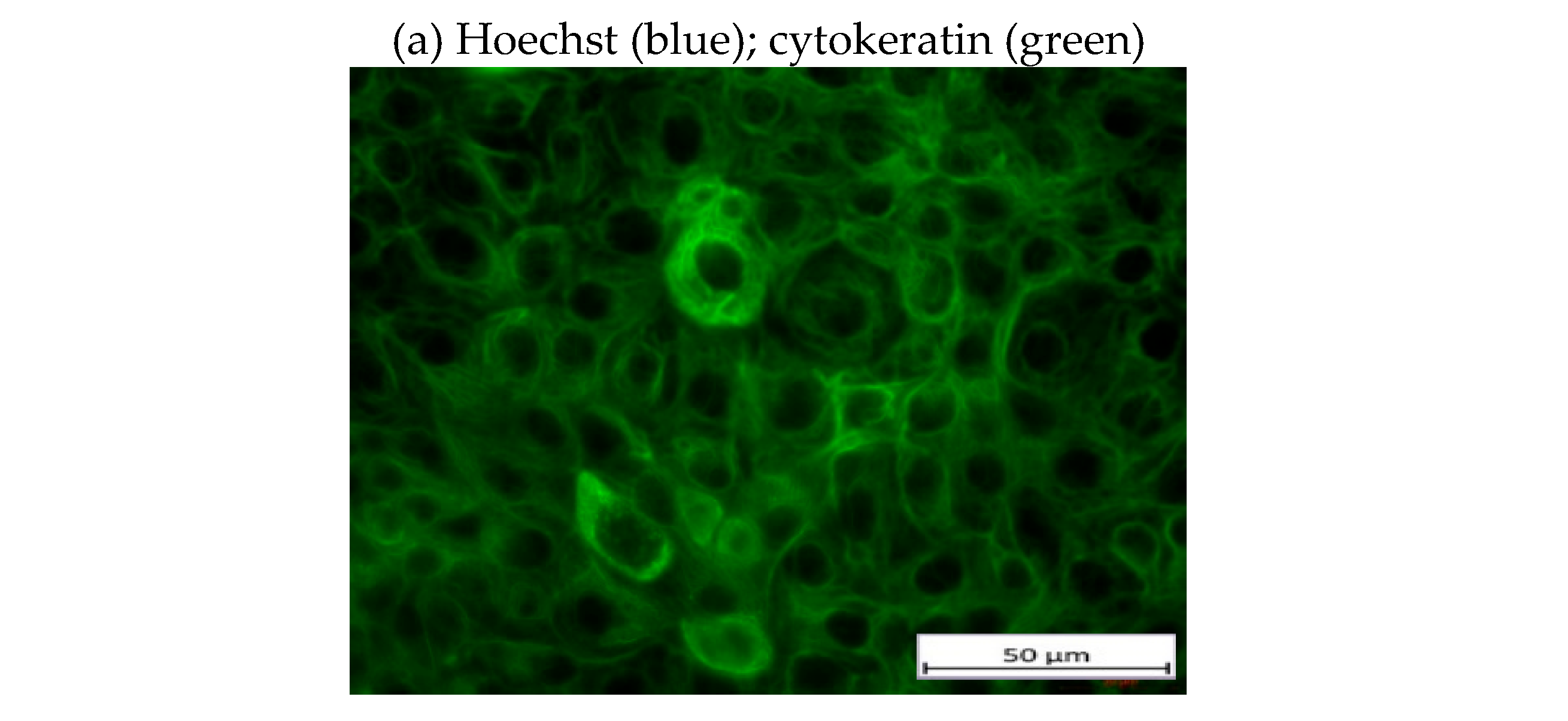
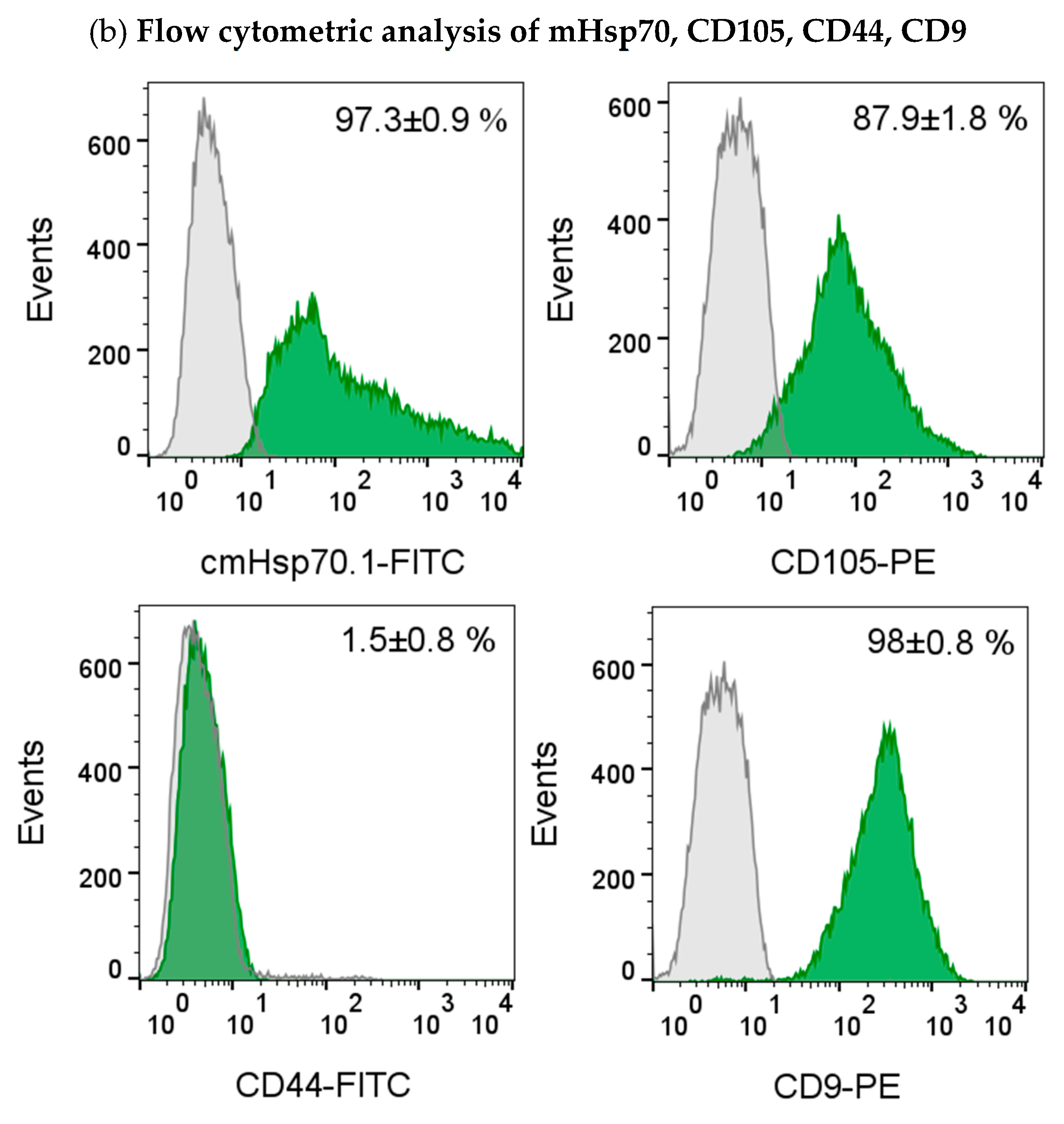
Figure 5.
DESeq2. Gene expression in CECs isolated with cmHsp70.1 mAb- and EpCAM mAb-conjugated beads from patient #10 with histologically confirmed lymph node endometriosis.
Figure 5.
DESeq2. Gene expression in CECs isolated with cmHsp70.1 mAb- and EpCAM mAb-conjugated beads from patient #10 with histologically confirmed lymph node endometriosis.
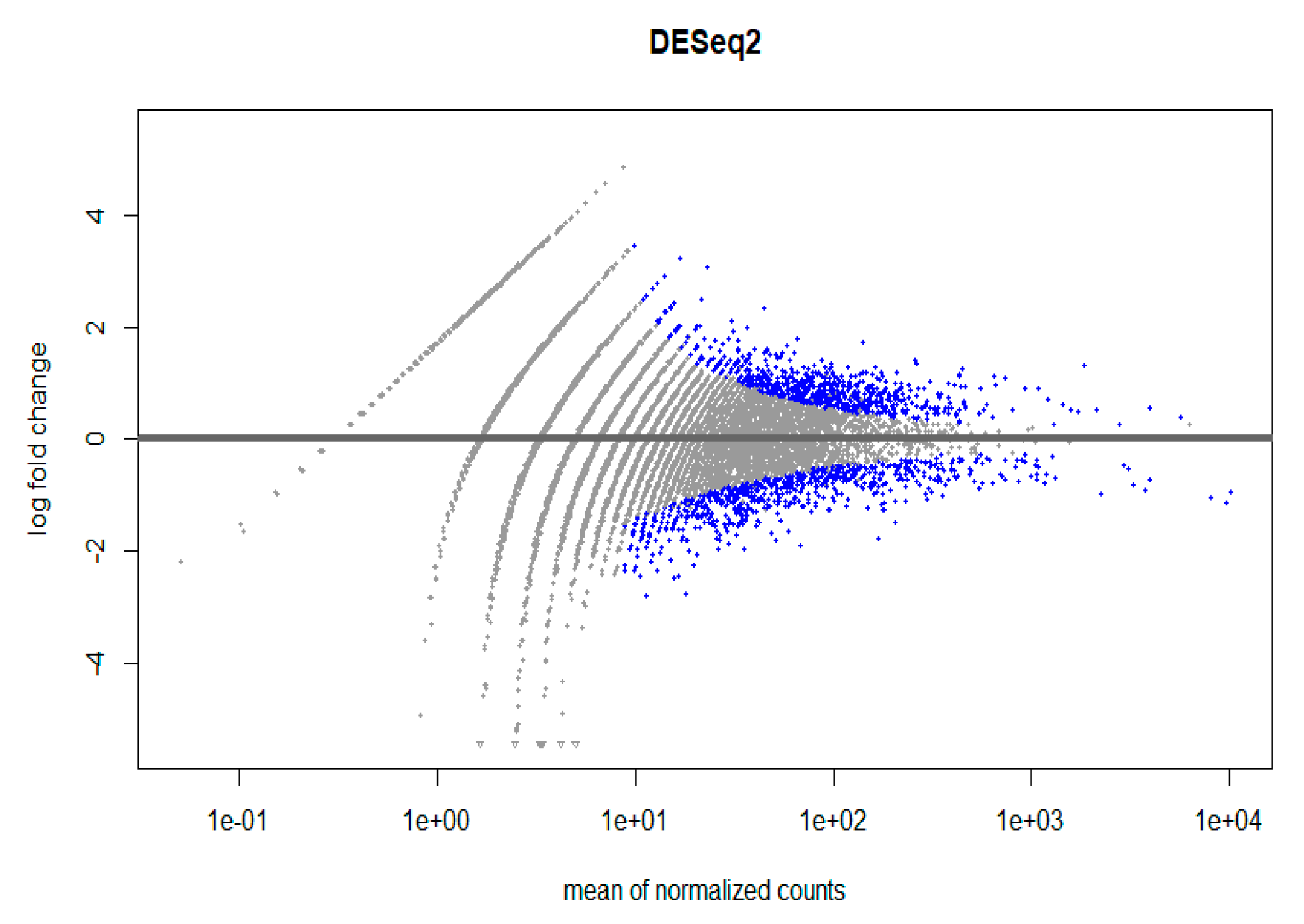

Table 1.
Enumeration of circulating endometriotic cells (CECs). Numbers of isolated CECs using the cmHsp70.1 and EpCAM mAb-based isolation technique derived from EDTA anti-coagulated blood (7.5 ml) of patients with histologically confirmed endometriosis (n=12) and two healthy donors (HD); patients #02, #06, #07, #08 had an involvement of the bladder and #10 had endometriotic lymph nodes. Additional EDTA blood samples were obtained from patients #11 and #12 before and 1-month post-surgery, and from patient #10 before and 6-months post-surgery.
Table 1.
Enumeration of circulating endometriotic cells (CECs). Numbers of isolated CECs using the cmHsp70.1 and EpCAM mAb-based isolation technique derived from EDTA anti-coagulated blood (7.5 ml) of patients with histologically confirmed endometriosis (n=12) and two healthy donors (HD); patients #02, #06, #07, #08 had an involvement of the bladder and #10 had endometriotic lymph nodes. Additional EDTA blood samples were obtained from patients #11 and #12 before and 1-month post-surgery, and from patient #10 before and 6-months post-surgery.
| Endometriosis | Distant organ involvement | Bead functionalization | |
|---|---|---|---|
| Patient ID | cmHsp70.1 mAb | EpCAM mAb | |
| 01 | No | 150 | 589 |
| 02 | Bladder | 1,013 | 1,845 |
| 03 | No | 120 | 98 |
| 04 | No | 552 | 30 |
| 05 | No | 34 | 57 |
| 06 | Bladder, Rectum | 4,963 | 770 |
| 07 | Bladder | 346,896 | 50 |
| 08 | Bladder | 214 | 199 |
| 09 | No | 98 | 139 |
| 10 | Lymph node | 12,935 | 379 |
| 10: follow-up | 186,648 | 6 | |
| 11 | No | 890 | 435 |
| 11: follow up | 219 | 384 | |
| 12 | No | 467 | 384 |
| 12: follow up | 123 | 156 | |
| Healthy donors | |||
| 01 | No | 0 | 0 |
| 02 | No | 0 | 0 |
Table 2.
Comparative RNA analysis in CECs. Genes differentially expressed by CECs isolated using EpCAM mAb- or cmHsp70.1 mAb-conjugated beads from patient #10 with histologically confirmed endometriotic lymph nodes; statistically significant DEGs comparing expression levels between cmHsp70.1 mAb- and EpCAM mAb-isolated CECs at an adjusted p value (padj) < 0.001 and an absolute log2 fold change greater than 1.
Table 2.
Comparative RNA analysis in CECs. Genes differentially expressed by CECs isolated using EpCAM mAb- or cmHsp70.1 mAb-conjugated beads from patient #10 with histologically confirmed endometriotic lymph nodes; statistically significant DEGs comparing expression levels between cmHsp70.1 mAb- and EpCAM mAb-isolated CECs at an adjusted p value (padj) < 0.001 and an absolute log2 fold change greater than 1.
| Genes | Function | Base mean | Log2 fold change | Padj |
|---|---|---|---|---|
| EpCAM mAb | ||||
| SRC | Progression Cytoskeletal reorganization |
44 | 1.1 | 4E−3 |
| FGR | Src kinase | 230.5 | 1.1 | 2E−13 |
| MMP2-AS1 | Extracellular matrix remodeling | 11.8 | 1.7 | 4E−3 |
| CDK5RAP3 | Cell cycle regulation | 10.2 | 2.1 | 7E−4 |
| CDKN1A | Cell cycle regulation | 51.9 | 1.3 | 1E−4 |
| RGCC | Cell cycle regulation | 94.8 | 1.3 | 3E−8 |
| ITGAM | Cell adhesion and migration | 20 | 1 | 1E−4 |
| ITGAE | Cell adhesion and migration | 29.5 | 1.4 | 1E−3 |
| cmHsp70.1mAb | ||||
| ROCK2 | EMT Cytoskeletal organization |
66.2 | -1.8 | 4E−5 |
| STAT3 | EMT Stemness-related marker |
107.8 | -1 | 1E−3 |
| CD82 | Cell adhesion | 134.2 | -1 | 2E−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



