
Submitted:
05 September 2024
Posted:
06 September 2024
You are already at the latest version
Abstract
Currently, the NRC amino acids (AA) requirements for pigs published in 2012 are used as a reference in variable swine industries. However, recent results in several articles suggest that the standardized ileal digestible (SID) AA: Lysine (Lys) ratio significantly evolved over the last two decades, while some publications report inconsistent outcomes. Therefore, the present study used a meta-regression analysis to assess the relative ratio to lysine to maximize the feed efficiency of four essential amino acids (tryptophan, valine, isoleucine, and methionine) in pig diets. According to the PRISMA guidelines, articles examining the target AA requirement using a basal diet supplemented with varying levels of crystalline AA (tryptophan, valine, isoleucine, or methionine) were identified across Scopus, PubMed, and Science Direct. As a result, 23, 22, 16, and 9 articles using tryptophan, valine, isoleucine, and methionine were selected and categorized into experiments for inclusion in our meta-analysis. The results suggested that the requirements of tryptophan, valine, isoleucine, and methionine in our meta-regression analysis were superior to NRC recommendations regardless of the regression models and the growth phases with significant RSQ values (RSQ ≈ 1). Also, the QUAD and CLP regression models emphasized higher requirements than the LP model for the great majority of amino acids and growth phases. The results of the QUAD and CLP models were selected as estimations of amino acid requirements for pigs under challenged conditions whereas the LP model was chosen to estimate the amino acid requirements of genetically improved pigs under a modern housing system. The results of this meta-regression analysis might be used to refresh the information on the NRC amino acids (AA) requirements for swine.
Keywords:
young pig
; tryptophan
; valine
; isoleucine
; methionine
; SID
; requirements
; meta-regression
1. Introduction
Nitrogen (N) excessively excreted into natural environments by pigs raised in modern farms has a detrimental impact through its contribution to acidification and eutrophication of sensitive ecosystems and odor emissions [1]. The reduction of crude protein (CP) content in swine diets has proven to decrease these negative effects by improving nitrogen (N) utilization efficiency, with beneficial outputs on both animal performance and health as well as reduced environmental impact [2,3]. However, this feeding strategy should be combined with an appropriate provision of amino acids (AA) in terms of quantity and quality to meet the animals’ genetic potential for protein deposition [4,5]. Up to these days, the NRC [6] has been the reference to provide appropriate amino acid requirements in pig's diet with low levels of CP. However, recent studies suggested that the standardized ileal digestible (SID) AA: Lysine (Lys) ratio significantly evolved in the last two decades, while some publications showed inconsistent outcomes [7,8,9,10]. Moreover, the NRC requirements are the condensed results from only six studies of which pig body weight did not overpass 33 kg [11]. Hence, a more accurate investigation considering every factor influencing the amino acid requirements (especially the limited amino acids) in pig nutrition seems to be necessary. In this context, aggregating the findings across all published papers is crucial to ultimately evaluate the requirements of the target amino acid in swine nutrition. Utilizing meta-regression analysis, a statistical tool to obtain an unbiased evaluation from multiple articles [12], is an appropriate approach to achieve these objectives. Indeed, meta-regression analysis can appropriately combine study findings and summarize the genuine impact of an intervention by drawing plots with statistical models, considering the sources of heterogeneity among studies and increasing the sample size.
Among the essential amino acids, tryptophan (Trp), valine (Val), isoleucine (Ile), and methionine (Met) play important partitions in several biological functions in pig metabolism. Trp is involved in the regulation of the immune response, voluntary feed intake and synthesis of serotonin [13,14,15]. Val and Ile belong to the group of essential branched-chain AA (BCAA), which are interdependent amino acids influencing the inclusion levels of other limiting AA. Furthermore, Val deficiency is known to decrease the feed intake within 1 h after meal ingestion in the pig's growing stage [16]. Concerning Met, it is recognized as a component of the sulphur amino acids (SAA), essential not only for protein accretion but also for producing cytokines, glutathione (GSH), and acute phase proteins (APPs) [17]. In addition to its functions in protein synthesis, Met plays key roles in biological functions (e.g., cellular redox function, cell survival and proliferation) and protection against diseases (e.g., liver cirrhosis, Alzheimer’s disease, and diabetes) [17]. Based on their importance and the fact that they have not been sufficiently investigated, additional studies are needed to estimate their requirements in pig nutrition.
In the process of assessing the requirements of these AA (Trp, Val, Ile, and Met), numerous statistical models have been developed [18] but the choice of method depends essentially on the species of interest, application facilities, and best fit of the data. The linear broken-line model, for instance, assumes a constant efficiency of AA utilization below the breakpoint where the efficiency is a linear function of the nutrient supplementation under the plateau [19]. However, linear broken-line regression supposes that the response to a nutrient dose is linear while it is generally not linear because of the rate of change due to the reduction of nutrient dose as the nutrient dose approaches its requirement [20]. For some data sets, a linear broken-line model will underestimate the requirement in that it represents the average animal in the population [21]. The quadratic model is easy to fit to data (only 3 input levels are needed for quadratic responses) and can easily fit the increases and decreases in performance [22]. However, the great majority of nutritional outcomes are known to have a plateau or “safe” level between the requirement for response levels and levels inducing negative effects, whereas a quadratic model does not characterize the nutrient response data [22]. Considering the diverse impacts of the regression models outlined in the previous statements, a meta-regression analysis was conducted to investigate the requirements of the limited amino acids of interest (Trp, Val, Ile, and Met) in pigs using the linear-plateau (LP), curvilinear-plateau (CLP), and quadratic (QUAD) regression models. The hypothesis tested in this study was that the requirement of the SID Trp, Val, Ile, and Met will be greater than the levels recommended in NRC [6] due to improved genetic potentials and an increased number of studies testing amino acid requirements under variable factors such as disease challenge models and antagonism between amino acids.
2. Materials and Methods
Literature Search and Study Selection
The literature search was conducted according to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) updated guidelines [23]. Research papers were sourced from various databases including Scopus (https://www.scopus.com/), PubMed (https://pubmed.ncbi.nlm.nih.gov/), and ScienceDirect (https://www.sciencedirect.com/), all accessed on April 11, 2023. The keywords used to search for the eligible papers were the name of the target AA, pig, and its synonyms (piglet, hog, barrow, gilt, and swine). The study selection was made through the record of articles containing the keywords in their titles and abstracts. Then, a full-text screening was processed using the same keyword to collect the final list of eligible papers. The articles retained to conduct this meta-regression analysis were peer-reviewed publications between 2000 and 2023.
Inclusion and Exclusion Criteria
The articles identified from the three databases were compiled in EndNote (version 20.6), and duplicates were automatically eliminated. The remaining records were independently screened by two reviewers based on the following criteria: in vivo experiments published in English (1) investigating the target AA requirement using a basal diet supplemented with varying levels of crystalline AA; (2) research providing a standardized ileal digestible (SID) AA profile; (3) studies using pigs weighing less than 50 kg as experimental animals; (4) assessments of the effects of different target AA requirements on growth parameters (average daily gain [ADG], average daily feed intake [ADFI], gain to feed intake ratio [G: F]). Conversely, experiments using unique basal diets were excluded.
Data Extraction and Cleansing
The studies were categorized into experimental groups based on various factors, including the type of experiments conducted, the basal diets used, and the experimental phase. Characteristics of each experimental group, such as authors, publication year, title of the paper, basal diet, crude protein (%DM), target SID AA (%DM), SID Lys (%DM), target SID AA: Lys ratio, initial BW (kg), final BW (kg), ADG (g/day), ADFI (g/day), and G: F ratio were extracted from each study. To further refine our analysis and detect any outliers, we used quantile ranges of the target SID AA: Lys ratio. The classification by BW for the target SID AA: Lys ratio requirements considered the stage of pig growth followed by National Research Council (NRC, 2012) guidelines: Phase 1 (BW <11), Phase 2 (11 ≤ BW <25), Phase 3 (25 ≤ BW <50). During our exploratory data analysis, we observed that in certain treatment groups, the response decreased with an increase in the dose, which was contrary to the expected biological or pharmacological response pattern. This indicated the possible presence of confounding factors or experimental errors, potentially obscuring the true dose-response relationship. To preserve the integrity and reliability of our models, these data points were excluded from the final analysis. This precaution is in alignment with established best practices in data analysis, where data that deviate from expected patterns are thoroughly evaluated and, if deemed necessary, omitted to avoid drawing incorrect conclusions. Focusing on data that display a consistent and interpretable response pattern, we sought to provide a more accurate and robust analysis of the dose-response relationship. This, in turn, enhances the validity of our results and ensures they accurately represent the biological mechanisms at play. We included only the experiments with at least two levels of the target SID AA: Lys ratio to guarantee a valid dose-response analysis.
Statistical Analysis
All statistical analyses were conducted using R software (R Core Team, 2023). The requirements for the target SID AA: Lys ratio to improve G: F in pigs were analyzed. Trials were categorized by the basal diet(s) reported in each experiment. The highest response within each group was set to 100%, and the remaining responses were standardized relative to the highest value observed. The response of pigs to an increase in target AA was modelled using linear-plateau (LP), curvilinear-plateau (CLP), and quadratic (QUAD) regression models:
LP: for
for
Relative response, %
The linear slope
The Intercept
The minimum SID AA: Lys ratio required to reach the plateau value.
CLP: for
for
Relative response, %
The quadratic coefficient
The linear slope
The Intercept
The minimum SID AA: Lys ratio required to reach the plateau value.
QUAD:
Relative response, %
The quadratic coefficient
The linear slope
The Intercept
The LP model identifies the optimal AA level for maximal growth in pigs until a plateau is reached. The CLP model provides deeper insights by analyzing both the growth phase and the plateau, identifying the 95% breakpoint (BP) where additional AA no longer improves performance. The QUAD model highlights the critical 95% peak value, indicating when the effectiveness of AA supplementation begins to decline, thus demonstrating the diminishing returns of excessive supplementation. The adequacy of these models was primarily confirmed by evaluating the Adjusted R-squared value (RSQ), which quantifies the proportion of variance in the dependent variable explained by the independent variables, adjusted for the number of predictors.
3. Results
3.1. Tryptophan
3.1.1. Dataset
The PRISMA flow diagram, presented in Figure 1, summarizes the search strategy employed to investigate Trp requirements. A total of 264 citations were identified across various databases (Scopus: 133, PubMed: 92, ScienceDirect: 37), and 2 additional citations were obtained through reference lists. During the screening process, duplicates and studies that did not meet the specific inclusion criteria were removed. Ultimately, 23 papers were selected for data extraction. Given that, some studies included multiple treatments (involving various experiments, basal diets, and phases), these 23 papers were categorized into several distinct experiments according to the phase (Phase 1: n = 37; Phase 2: n = 23; Phase 3: n = 35). An experiment was defined as the implementation of a basal diet associated with one or more Trp inclusions, expressed as the SID Trp: Lys ratio. The datasets and experimental conditions of the 23 studies, recorded after complete screening, are presented in Table 1. The table lists the authors and year of publication and categorizes each study according to the growth phase of the subjects. It also detailed the range of SID Trp: Lys ratio and the range of SID Lys and provides remarks on specific challenge studies. The compilation of research included in the table spans from 2003 to 2022.
3.1.2. Estimation of the SID Trp: Lys Ratio Requirement
The requirements of SID Trp: Lys ratio by body weight classification (Phase 1, Phase 2, and Phase 3) to maximize G: F are reported in Figure 2. The Trp: Lys ratio according to the LP, CLP and QUAD models were all superior to NRC [6] requirements regardless of the growth phase. In Phase 1, the results presented in the diagram (Figure 2) show that the CLP and QUAD models, with equal values, highlighted higher requirements in terms of SID Trp: Lys ratio than the LP model and NRC requirements (QUAD: 0.229 = CLP: 0.229 > LP: 0.212 > NRC: 0.163). Similarly, the QUAD model induced the highest requirements successively followed by CLP, LP, and NRC outputs in Phase 2 (QUAD: 0.204 > CLP: 0.188 > LP: 0.183 > NRC: 0.163) and Phase 3 (QUAD: 0.207 > CLP: 0.199 > LP: 0.186 > NRC: 0.173). Independently of the phase and regression model, significant RSQ values (RSQ ≈ 1) wavering between 0.670 and 0.757 indicated that the estimation of SID Trp: Lys ratio requirements was explained by the independent variables in the model.
3.2. Valine
3.2.1. Dataset
The PRISMA flow chart depicted in Figure 3, encapsulates the methodology adopted for researching Val requirements. We identified 105 citations from a range of databases: Scopus yielded 51, PubMed 41, and ScienceDirect contributed 13 references. Throughout the vetting process, we excluded duplicate entries and studies that fell outside our predefined selection criteria. Finally, 22 articles were earmarked for in-depth data extraction. Owing to the presence of multifaceted treatment protocols in some studies, encompassing a variety of experiments, basal diets, and growth phases, these 22 articles were organized into multiple unique experimental depending on the phase (Phase 1: n = 30; Phase 2: n = 39; Phase 3: n = 21). We delineated an experiment as any application of a basal diet complemented by one or more valine additions, quantified by the SID Val: Lys ratio. The accumulated data and the specifics of the experimental setups for the 22 scrutinized studies are presented in Table 2. This table highlights the growth phase categorization for the studies, the spectrum of SID Val: Lys ratios, the SID Lys ranges, and annotations regarding interactions with branched-chain amino acids (BCAAs). The compilation of studies spans a publication period from 2009 to 2023.
3.2.2. Estimation of the SID Val: Lys Ratio Requirement
Figure 4 showed the optimal SID Val: Lys ratio requirements across different body weight classifications (Phase 1, Phase 2, and Phase 3) to maximize G: F. Irrespective of the growth phase, the Val: Lys ratios determined by the LP, CLP, and QUAD models surpassed the NRC (2012) standards. Notably, in Phase 1 and Phase 3, both the CLP and QUAD models exhibited higher Val: Lys ratio requirements compared to the LP model and NRC recommendations (Phase 1: QUAD: 0.717 = CLP: 0.717 > LP: 0.651 > NRC: 0.637; Phase 3: QUAD: 0.732 = CLP: 0.732 > LP: 0.690 > NRC: 0.653). In Phase 2, the CLP model indicated the highest requirements, followed by QUAD, LP, and NRC (CLP: 0.695 > QUAD: 0.694 > LP: 0.662 > NRC: 0.634). Across all phases and regression models, robust RSQ values (RSQ ≈ 1), ranging from 0.576 to 0.722, affirmed that the estimation of SID Val: Lys ratio requirements was well-explained by the independent variables within the model.
3.3. Isoleucine
3.3.1. Dataset
The PRISMA flow diagram, presented in Figure 5, summarizes the search strategy employed to investigate Ile requirements. A total of 74 citations were identified across various databases (Scopus: 35, PubMed: 30, ScienceDirect: 7), and 2 additional citations were obtained through reference lists. During the screening process, duplicates and studies that did not meet the specific inclusion criteria were removed. Ultimately, 16 papers were selected for data extraction. Given that, some studies included numerous treatments (involving various experiments, basal diets, and phases), these 16 papers were categorized into distinct experiments according to the growth phase (Phase 1: n = 24; Phase 2: n = 34; Phase 3: n = 11). An experiment was defined as the implementation of a basal diet associated with one or more Ile inclusions, expressed as the SID Ile: Lys ratio. The datasets and experimental conditions of the 16 studies, recorded after complete screening, are presented in Table 3. The table lists the authors and year of publication and categorizes each study according to the growth phase of the subjects. It also detailed the range of SID Ile: Lys ratio and the range of SID Lys and provides remarks on specific challenge studies. The compilation of research included in the table spans from 2003 to 2022.
3.3.2. Estimation of the SID Ile: Lys Ratio Requirement
Figure 6 outlines the SID Ile: Lys ratio requirements across different body weight classifications (Phase 1, Phase 2, and Phase 3) to optimize G: F. No matter the stage of growth, the Ile: Lys ratios derived from the LP, CLP, and QUAD models exceeded the NRC (2012) standards. Specifically, in Phase 1, both the CLP and QUAD models indicated higher Ile: Lys ratio requirements compared to the LP model and NRC recommendations (QUAD: 0.648 = CLP: 0.648 > LP: 0.580 > NRC: 0.511). Similarly, the QUAD model demonstrated the highest requirements followed by CLP, LP, and NRC in Phase 2 (QUAD: 0.609 > CLP: 0.558 > LP: 0.548 > NRC: 0.512) and Phase 3 (QUAD: 0.580 > CLP: 0.564 > LP: 0.549 > NRC: 0.520). Moderate RSQ values ranging from 0.517 to 0.588 were observed in Phases 1 and 2, while high RSQ values ranging from 0.935 to 0.968 were noted in Phase 3. However, all results were statistically significant (RSQ ≈ 1), affirming that the estimation of SID Ile: Lys ratio requirements was well-explained by the independent variables in the model.
3.4. Methionine
3.4.1. Dataset
The PRISMA flow diagram, presented in Figure 7, summarizes the search strategy employed to investigate Met requirements. A total of 528 citations were identified across various databases (Scopus: 301, PubMed: 192, ScienceDirect: 35). During the screening process, duplicates and studies that did not meet the specific inclusion criteria were removed. Ultimately, 9 papers were selected for data extraction. Given that, some studies included multiple treatments (involving various experiments, basal diets, and phases), these 9 papers were categorized into numerous distinct experiments according to the phases studied (Phase 1: N = 16; Phase 2: N = 13; Phase 3: N = 21). An experiment was defined as the implementation of a basal diet associated with one or more Met inclusions, expressed as the SID Met: Lys ratio. The datasets and experimental conditions of the 9 studies, recorded after complete screening, are presented in Table 4. The table lists the authors and year of publication and categorizes each study according to the growth phase of the subjects. It also detailed the range of SID Met: Lys ratio and the range of SID Lys and provides remarks on specific challenge studies. The compilation of research included in the table spans from 2003 to 2022.
3.4.2. Estimation of the SID Met: Lys Ratio Requirement
The SID Met: Lys ratio requirements across different body weight classifications (Phase 1, Phase 2, and Phase 3) for maximizing G: F are presented in Figure 8. The Met: Lys ratios determined by the LP, CLP, and QUAD models exceeded the NRC (2012) standards, irrespective of the growth phase. In Phase 1 (QUAD: 0.414 > CLP: 0.413 > LP: 0.317 > NRC: 0.289) and Phase 2 (QUAD: 0.400 > CLP: 0.327 > LP: 0.294 > NRC: 0.293), the results depicted in Figure 8 demonstrated that the QUAD model indicated the highest requirements, followed by CLP, LP, and NRC outputs. For Phase 3, both the CLP and QUAD models, with equal values, emphasized higher requirements in terms of SID Met: Lys ratio compared to the LP model and NRC requirements (QUAD: 0.436 = CLP: 0.436 > LP: 0.351 > NRC: 0.286). In Phase 1, moderate RSQ values ranging between 0.637 and 0.684 were observed, while high RSQ values ranging from 0.735 to 0.908 were noted in Phases 2 and 3. However, all these results were statistically significant (RSQ ≈ 1), confirming that the estimation of SID Met: Lys ratio requirements was accurately explained by the independent variables in the model.
4. Discussion
Several statistical models have been developed to determine the requirements of amino acids and proteins in humans [18] and animals, but the choice of method depends essentially on the species to be worked on, application facilities, and the best fit of the data. This study investigated the amino acid requirements of pigs using the LP, CLP, and QUAD meta-regression models based on numerous outcomes. Indeed, the approximate and controversial results found among individual studies made it difficult to obtain an accurate view of pigs’ amino acid requirements in today’s industry. Therefore, meta-regression analysis, an important instrument to get an unbiased evaluation of the available pieces of evidence, is one of the best methods to reach this target. It can accurately summarize the study findings and detect the real effect of an intervention by considering the sources of heterogeneity between articles and increasing the sample size [89]. The great majority of studies use every growth parameter (ADG, BWG, ADFI, and G: F) to estimate the quantity of amino acids maximizing pig performance [11,37,70]. In the present meta-regression analysis, the feed efficiency (G: F) was selected to assess the amino acid requirements in pig nutrition because it summarized the other growth parameters by expressing the unit of feed consumed per unit of body weight gain.
4.1. Tryptophan
It is commonly asserted that Trp, the second or third limiting AA in cereal-based diets for pigs [90] play an important role in biological functions [13,91] and is involved in the regulation of the immune response, voluntary feed intake and synthesis of serotonin [13,14,15]. Hence, it is necessary to determine the Trp requirement of pigs expressed as the ratio of Trp to Lys, with Lys as the second limiting AA to avoid other variables that are similarly restrictive to Trp, thus leading the Trp to Lys ratio to be underestimated [92].
The SID Trp: Lys ratio according to the LP, CLP and QUAD models were all superior to NRC [6] requirements regardless of the growth phase. The minimum SID Trp: Lys ratio to support optimum G: F in phase 1 (QUAD = CLP = 0.229; LP = 0.212), phase 2 (QUAD = 0.204; CLP = 0.188; LP = 0.183), and phase 3 (QUAD = 0.207; CLP = 0.199; LP = 0.186) of this meta-regression analysis are in average comparable to a previous meta-analysis investigating the Trp requirements [92] and several original experiments on the same topic [27,36,93]. These results might be due to the improvement of pigs’ growth potential by modern breeding and genetic technologies [94]. Indeed, over 70% of the studies used in the present meta-analysis were conducted from 2012 to 2022, after the release of NRC requirements (Under modern housing conditions and with developed lean-type genetics). In addition, the inclusion of challenged studies in this meta-analysis to assess the Trp: Lys requirements could be also a source of greater values for the optimum inclusion levels of Trp, as inflammatory and stress challenges are known to increase partitioning of Trp to the immune system [32,33,45,95]. Indeed, during an immune challenge (natural or induced), Trp utilization is increased due to higher catabolism of Trp via the activation of indoleamine 2, 3 dioxygenases (IDO) [96]. Therefore, during infection, the requirement for Trp is increased. Furthermore, the high Trp: Lys ratio observed in the results of our meta-analysis might be due to the presence of articles in which excess leucine was balanced by higher incorporation of Trp in pigs’ diets [7]. Numerous authors [97,98,99] suggested that the negative effect of excess leucine could be partially improved by increasing dietary Trp. Based on the previous assertions, we considered that the higher Trp requirements highlighted by our meta-regression are partially attributed to factors such as environmental challenges and diets with excess leucine on the one hand and the other hand to modern housing and genetic improvement. Moreover, it has been reported that the regression model could have some influence on the estimation of amino acid requirements. Therefore, we would suggest a category of Trp: Lys ratio under challenged conditions which would be the exact values or range values of QUAD and CLP models and another category of Trp: Lys ratio under modern housing and with genetically enhanced pigs represented by the LP model’s values. Hence, the tryptophan requirements for pigs in challenged conditions and excessive inclusion of leucine would be 0.229, [0188; 0.204], and [0.199; 0.207] in phases 1, 2, and 3 respectively. The Trp: Lys ratio for genetically improved pigs under modern housing could be 0.212, 0.183, and 0.186 in Phases 1, 2, and 3, respectively. However, deeper studies should be conducted to confirm these results.
4.2. Valine
Val belongs to the group of essential branched-chain AA (BCAA), that animals cannot synthesize and therefore must be supplied through feeding. In pig nutrition, Val is considered to be the fifth limiting amino acid, and its deficiency is known to decrease the feed intake within 1 h after meal ingestion in the growing stage [16].
The SID Val: Lys ratios determined by the LP, CLP, and QUAD models surpassed the NRC [6] standards in every growth phase (Phase 1, Phase 2, and Phase 3). Val requirements suggested by the regression models in phase 1 (QUAD = CLP = 0.717; LP = 0.651), phase 2 (CLP = 0.695; QUAD = 0.694; LP = 0.662) and phase 3 (QUAD = CLP = 0.732; LP = 0.690), are comparable to several previous outcomes [50,56,63]. These results might be partially due to the improvement of pigs’ growth potential by modern breeding and genetic technologies [94]. Indeed, over 60% of the studies used in the present meta-analysis were conducted from 2012 to 2022, after the publication of NRC requirements (Under modern housing conditions and with new lean-type genetics). Also, the high number of studies [25,56,62,63] with high leucine levels used in this meta-analysis to assess the SID Val: Lys requirements could also be the source of greater values for the optimum inclusion levels of Val. Indeed, Harper et al. [100] suggested that an oversupply of leucine results in an increase in the requirements of the two other branched-chain amino acids (BCAA), including Val. Moreover, some authors [16,101] asserted that excess leucine in combination with a low level of Val in the diet had a negative effect on feed intake and consequently on the performance of piglets. High levels of leucine can stimulate the BCAA dehydrogenase complex (BCKDH), an enzyme complex that catalyzes the irreversible degradation of all BCAAs, including Val [99]. Therefore, excess leucine increases Val catabolism, which might become deficient and alter insulin-like growth factor-1 (GH-IGF-1) expression [50]. Nevertheless, as previously explained, these deficiencies can be overcome by higher inclusions of Val to balance the amino-acid requirements in terms of BCAA.
In addition, the level of Trp in pig diets is also known to influence the valine requirements [11,59] and could explain the higher requirements obtained in our meta-analysis. Following numerous researchers [7,25,102], Clizer et al. [11] suggested that Val and Trp play an integral role in correcting the negative effects of excess dietary leucine when diets contain high inclusion levels of corn co-products.
Apart from the genetic improvement and the modern housing in the pig industry, several other parameters such as the balance in BCAA and Trp inclusion level might be the cause of higher Val requirements highlighted by our meta-analysis. Also, the regression model was reported to influence the findings of numerous experiments on amino acid ratios in pig nutrition [56]. Thus, we hypothesize that LP results would be the Val requirements for pigs genetically improved and raised in modern housing, while the Val: Lys ratio of CLP and QUAD model would be used for pigs fed with diets containing other BCAA or tryptophan in excess.
Thus, our suggestions for Val requirements in phases 1, 2, and 3 would be 0.717, [0.694; 0.695], and 0.732, respectively in diets with excessive BCAA or Trp inclusion levels. Whereas 0.651, 0.662, and 0.690 could be the Val requirements in phases 1, 2 and 3, respectively, in diets with balanced amino acids, modern housing and genetically improved pigs.
However, further research using the Val requirements emphasized by our meta-analysis should be conducted to confirm our hypothesis.
4.3. Isoleucine
Ile, one of the branched-chain amino acids (BCAA), is the sixth limiting amino acid in pig nutrition [71]. The SID Ile: Lys ratios derived from the LP, CLP, and QUAD models exceeded the NRC [6] standards regardless of the growth phase. These results might be in part due to the improvement of pigs’ lean meat growth potential by modern breeding and genetic technologies [94]. Indeed, over 80% of the studies used in the present meta-analysis were conducted after the release of NRC results in 2012 (Under modern housing conditions and with new pig strains). Also, the presence of studies [72,74] with high leucine or Val levels used in this meta-analysis to assess the SID Ile: Lys requirements could be the source of greater values for the optimum inclusion levels of Ile. Indeed, the Ile requirement increases when other BCAA are in excess, particularly due to an antagonistic effect with an excess leucine [103]. This mechanism occurs when elevated leucine increases levels of the enzyme complex branched-chain keto-acid dehydrogenase, which increases the degradation of all BCAA; thus, catabolizing Ile and increasing the requirement [98,104]. According to the results of our meta-analysis, the optimum SID Ile: Lys ratio of pigs in phase 1 varied from 0.580 to 0.648 independently of the model. These higher requirements compared to NRC recommendation and several articles in the literature review [66,69,70] might be explained by the previous assessment related to genetic improvement and excessive inclusion of leucine or Val. In phases 2 and 3, the Ile requirement oscillated from 0.548 to 0.609 and from 0.549 to 0.580, respectively. These results are also higher than several previous experiments [60,67,71,72] on Ile requirements in pig nutrition.
It is noteworthy to mention that the regression model can significantly influence the estimation of amino acid requirements. In their investigation on Val requirements, Liu et al. [56] emphasized a SID Val: Lys ratio of 0.62 using the linear broken-line model, while it was 0.71 when using the quadratic model. Considering the huge number of factors influencing the Ile requirements, we hypothesized that the requirements of QUAD and CLP models would be the right ones for Ile requirements in diets with excess leucine or Val and the LP model values a suggestion of SID Ile: Lys ratio due to genetic improvements in pig balanced diets. Therefore, our recommendation for Ile requirements in phases 1, 2, and 3 would be 0.648, [0.558; 0.609], and [0.564; 0.580], respectively in diets with excessive BCAA inclusion levels. On the other hand, 0.580, 0.548, and 0.549 could be the Ile requirements in phases 1, 2 and 3, respectively, in diets with balanced BCAA inclusion levels.
However, additional research with these estimations of Ile requirements in pig nutrition is necessary to confirm our results.
4.4. Methionine
According to Grimble [105], Met, a component of the SAA is essential not only for protein accretion but also for the production of cytokines, glutathione (GSH), and acute phase proteins (APPs). In addition to its functions in protein synthesis, Met plays key roles in biological functions (e.g., cellular redox function, cell survival and proliferation) and protection against diseases (e.g., liver cirrhosis, Alzheimer’s disease, and diabetes) [17]. Indeed, a dietary Met deficiency impairs the immune response by affecting the development of lymphoid organs and by reducing the production of antibodies and T-cell proliferation in pigs [106,107] and chickens [108]. Thus, higher SAA supplementation can minimize the negative effects of immunological challenges and help the host maintain health and reduce muscle loss [109]. Met is commonly provided when feeding piglets because it is the second limiting amino acid in typical diets. The Met: Lys ratios determined by the LP, CLP, and QUAD models exceeded the NRC [6] standards, irrespective of the growth phase. These results could be explained by the improvement of pigs’ growth potential by modern breeding and genetic technologies [94]. Indeed, 100% of the studies used in the present meta-analysis were conducted after the release of NRC requirements in 2012 (Under modern housing conditions and with new lean-type genetics). According to the results of our meta-analysis, the optimum Met: Lys ratio of pigs in phase 1 varied from 0.317 to 0.417 regardless of the regression model. On the other hand, Rostagno et al. [110], and Sève [111] recommend a Met: Lys of 0.29–0.30 for maximal growth, similar to the NRC [6] report but lower than the present meta-analysis outcomes. The higher Met requirements in phase 1 observed in our findings compared to previous results might be due to the inclusion of challenged studies [79,80,82] in our meta-analysis. It has been reported that a negative whole-body protein resulting from muscle catabolism to release AA for energy and synthesis of cytokines and APPs may occur when animals are under pathogenic challenge [91,112,113,114]. Therefore, increasing the level of Met in those conditions reduces the adverse effect on pig performance. In phase 2, the Met requirements in our meta-regression analysis oscillated between 0.294 and 0.400 across the regression models. These results are confirmed by several previous experiments [76,80,86] showing the evolution of Met inclusion level in pig nutrition. The Met requirements in phase 3 of the present regression meta-analysis ranged from 0.351 to 0.436. The results of Remus et al. [78] and Zhang et al. [84] emphasizing a linear improvement of G: F with Met: Lys ratio up to 0.50 and 0.38 respectively support on average the findings of our study. The reason behind the enhancement of feed efficiency induced by methionine supplementation might be its ability to increase the villus height in the jejunum [85]. In addition, Tsukahara et al. [115] suggested that Met can improve intestinal digestive capacity by increasing the activity of jejunal enzymes.
However, cysteine (Cys) can be converted from Met as needed [116]. Hence, the amount of Met needed in the diet depends on the amount of Cys present.
Based on the different parameters able to influence the Met requirements and the possibility that the regression models could influence the Met: Lys ratio [56], it has been hypothesized that the required levels of Met inclusion in pig diets should be divided into two categories. The first category represents the Met requirements induced by pig genetic improvement and modern housing expressed by the LP model and the methionine requirements in a challenged environment and unbalanced AA highlighted by the QUAD and CLP models of our meta-regression analysis. Hence, the recommendations of our meta-analysis in terms of Met: Lys ratio are described as follows: 0.317, 0.294, and 0.351 for genetically enhanced pigs under modern housing in phases 1, 2, and 3, respectively and [0.413;0.414]; [0.327;0.400]; 0.436 for pigs raised under challenged conditions in phase 1, 2, and 3, respectively.
Nevertheless, additional original experiments using these estimations of Met requirements in pig nutrition are necessary to confirm our results.
5. Conclusions
This meta-analysis aimed to assess the requirements for maximizing the feed efficiency of four amino acids (Trp, Val, Ile, and Met) in pig growth phases using the QUAD, CLP, and LP regression models. The results suggested that the requirements of Trp, Val, Ile, and Met in our meta-regression analysis were all superior to NRC recommendations regardless of the regression models and the growth phases with significant RSQ values (RSQ ≈ 1). Also, the QUAD and CLP regression models emphasized higher requirements than the LP model for the great majority of amino acids and growth phases. The results of the QUAD and CLP models were selected as estimations of amino acid requirements for pigs under challenged conditions and unbalanced diets, whereas the LP model was chosen to estimate the amino acid requirements of genetically improved pigs under a modern housing system. Since the present meta-analysis used a high number of experiments and considered several factors able to influence amino acid requirements, it could be used as a reference for Trp, Val, Ile, and Met inclusion levels in pig nutrition.
Author Contributions
Conceptualization, B.C., A.-L. L. and J.T.; methodology, B.C., J.I.C.P.D. and I.C.; software, B.C. and I.C.; validation, N.-J.C., A.-L. L. and J.T.; formal analysis, J.I.C.P.D.; investigation, J.I.C.P.D. and B.C.; resources, N.-J.C., A.-L. L. and J.T.; data curation, B.C., J.T. and I.C.; writing—original draft preparation, J.I.C.P.D. and B.C.; writing—review and editing, J.I.C.P.D., J.T. and N.-J.C.; visualization, N.-J.C., A.-L. L. and J.T.; supervision, N.-J.C., J.T; project administration, N.-J.C. and J.T; funding acquisition, N.-J.C. and J.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in this study are included in the article; further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Webb, J., Broomfield, M., Jones, S. and Donovan, B. Ammonia and odour emissions from UK pig farms and nitrogen leaching from outdoor pig production. A review. Sci. Total Environ 2014, 470, pp.865-875.
- Gloaguen, M., Le Floc'H, N., Corrent, E., Primot, Y. and Van Milgen, J. The use of free amino acids allows the formulation of very low crude protein diets for piglets. J. Anim. Sci. 2014, 92(2), pp.637-644.
- Monteiro, A.N.T.R., Bertol, T.M., de Oliveira, P.A.V., Dourmad, J.Y., Coldebella, A. and Kessler, A.M. The impact of feeding growing-finishing pigs with reduced dietary protein levels on performance, carcass traits, meat quality and environmental impacts. Livest. Sci. 2017, 198, pp.162-169.
- Wang, T.C. and Fuller, M.F. The optimum dietary amino acid pattern for growing pigs: 1. Experiments by amino acid deletion. Br. J. Nutr. 1989, 62(1), pp.77-89.
- Gietzen, D. W., and L. J. Magrum. Molecular mechanisms in the brain involved in the anorexia of branched-chain amino acid deficiency. J. Nutr. 2001, 131:851S–855S.
- NRC. Nutrient requirements of swine. 11th rev. ed. Natl. Acad. Press, Washington, DC, USA, 2012.
- Kwon, W.B., Soto, J.A. and Stein, H.H. Effects of dietary leucine and tryptophan on serotonin metabolism and growth performance of growing pigs. J. Anim. Sci. 2022, 100(1), p.skab356.
- Goodarzi, P., Wileman, C.M., Habibi, M., Walsh, K., Sutton, J., Shili, C.N., Chai, J., Zhao, J. and Pezeshki, A. Effect of isoleucine and added valine on performance, nutrients digestibility and gut microbiota composition of pigs fed with very low protein diets. Int. J. Mol. Sci. 2022, 23(23), p.14886.
- Habibi, M., Goodarzi, P., Shili, C.N., Sutton, J., Wileman, C.M., Kim, D.M., Lin, D. and Pezeshki, A. A mixture of valine and isoleucine restores the growth of protein-restricted pigs likely through improved gut development, hepatic IGF-1 pathway, and plasma metabolomic profile. Int. J. Mol. Sci. 2022, 23(6), p.3300.
- Zong, E., Huang, P., Zhang, W., Li, J., Li, Y., Ding, X., Xiong, X., Yin, Y. and Yang, H. The effects of dietary sulfur amino acids on growth performance, intestinal morphology, enzyme activity, and nutrient transporters in weaning piglets. J. Anim. Sci. 2018, 96(3), pp.1130-1139.
- Clizer, D.A., Tostenson, B.J., Tauer, S.K., Samuel, R.S. and Cline, P.M. Impact of increasing standardized ileal digestible valine: lysine in diets containing 30% dried distiller grains with solubles on growing pig performance. J. Anim. Sci. 2022, 100(9), p.skac228.
- Balduzzi, S., Rücker, G. and Schwarzer, G. How to perform a meta-analysis with R: a practical tutorial. BMJ Ment. Health 2019, 22(4), pp.153-160.
- Henry, Y., Seve, B., Colleaux, Y., Ganier, P., Saligaut, C., J´ego, P. Interactive effects of dietary levels of tryptophan and protein on voluntary feed intake and growth performance in pigs, in relation to plasma free amino acids and hypothalamic serotonin. J. Anim. Sci. 1992, 70, 1873–1887.
- Han, Y., Chung, T.K., Baker, D.H. Tryptophan requirement of pigs in the weight category 10 to 20 kilograms. J. Anim. Sci. 1993, 71, 139–143.
- Pastuszewska, B., Tomaszewska-Zaremba, D., Buraczewska, L., Swi ´ ęch, E., Taciak, M. Effects of supplementing pig diets with tryptophan and acidifier on protein digestion and deposition, and on brain serotonin concentration in young pigs. Anim. Feed Sci. Technol. 2007, 132, 49–65.
- Gloaguen, M.; Le Floc’h, N.; Corrent, E.; Primot, Y.; van Milgen, J. Providing a diet deficient in valine but with excess leucine results in a rapid decrease in feed intake and modifies the postprandial plasma amino acid and α-keto acid concentrations in pigs. J. Anim. Sci. 2012, 90, 3135–3142.
- Bauchart-Thevret, C., B. Stoll, S. Chacko, and D.G. Burrin. Sulfur amino acid deficiency upregulates intestinal methionine cycle activity and suppresses epithelial growth in neonatal pigs. Am. J. Physiol. Endocrinol. Metab. 2009, 296:E1239–E1250.
- Elango, R., Ball, R.O. and Pencharz, P.B. Recent advances in determining protein and amino acid requirements in humans. Br. J. Nutr. 2012, 108(S2), pp.S22-S30.
- Gahl, M.J., Crenshaw, T.D. and Benevenga, N.J. Diminishing returns in weight, nitrogen, and lysine gain of pigs fed six levels of lysine from three supplemental sources. J. Anim. Sci. 1994, 72(12), pp.3177-3187.
- Robbins, K.R., Saxton, A.M. and Southern, L.L. Estimation of nutrient requirements using broken-line regression analysis. J. Anim. Sci. 2006, 84(suppl_13), pp.E155-E165.
- Baker, D.H. Problems and pitfalls in animal experiments designed to establish dietary requirements for essential nutrients. J. Nutr. 1986, 116(12), pp.2339-2349.
- Pesti, G.M., Vedenov, D., Cason, J.A. and Billard, L. A comparison of methods to estimate nutritional requirements from experimental data. Br. Poult. Sci. 2009, 50(1), pp.16-32.
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 88, 105906.
- Tolosa, A.F., Tokach, M.D., Goodband, R.D., Woodworth, J.C., DeRouchey, J.M., Gebhardt, J.T. and Wolfe, M.L. Effects of reducing the standardized ileal digestible lysine and tryptophan to lysine ratio to slow growth of finishing pigs. Transl. anim. sci. 2022, 6(2), p.txac057.
- Kerkaert, H.R., Cemin, H.S., Woodworth, J.C., DeRouchey, J.M., Dritz, S.S., Tokach, M.D., Goodband, R.D., Haydon, K.D., Hastad, C.W. and Post, Z.B. Improving performance of finishing pigs with added valine, isoleucine, and tryptophan: validating a meta-analysis model. J. Anim. Sci. 2021, 99(1), p.skab006.
- Ma, W., Mao, P., Zhu, Y., Guo, L., Zhang, S., Wang, Z. and Zhao, F. Standardized ileal digestible tryptophan to lysine ratios affect performance and regulate intestinal mRNA expression of amino acid transporters in weaning pigs fed a low crude protein diet. Anim. Feed Sci. Technol. 2021, 275, p.114857.
- Capozzalo, M.M., Kim, J.C., Htoo, J.K., de Lange, C.F.M., Mullan, B.P., Resink, J.W., Hansen, C.F., Stumbles, P.A., Hampson, D.J., Ferguson, N. and Pluske, J.R. Estimating the standardised ileal digestible tryptophan requirement of pigs kept under commercial conditions in the immediate post-weaning period. Anim. Feed Sci. Technol. 2020, 259, p.114342.
- Liu, J.B., Yan, H.L., Cao, S.C., Liu, J., Li, Z.X. and Zhang, H.F. The response of performance in grower and finisher pigs to diets formulated to different tryptophan to lysine ratios. Livest. Sci. 2019, 222, pp.25-30.
- Wensley, M.R., Woodworth, J.C., DeRouchey, J.M., Dritz, S.S., Tokach, M.D., Goodband, R.D., Walters, H.G., Leopold, B.A., Coufal, C.D., Haydon, K.D. and Lee, J.T. Effects of amino acid biomass or feed-grade amino acids on growth performance of growing swine and poultry. Transl. anim. sci. 2020, 4(1), pp.49-58.
- Gonçalves, M.A.D., Tokach, M.D., Bello, N.M., Touchette, K.J., Goodband, R.D., DeRouchey, J.M., Woodworth, J.C. and Dritz, S.S. Dose–response evaluation of the standardized ileal digestible tryptophan: lysine ratio to maximize growth performance of growing-finishing gilts under commercial conditions. Animal 2018, 12(7), pp.1380-1387.
- Capozzalo, M.M., Kim, J.C., Htoo, J.K., De Lange, C.F.M., Mullan, B.P., Hansen, C.F., Resink, J.W. and Pluske, J.R. Pigs experimentally infected with an enterotoxigenic strain of Escherichia coli have improved feed efficiency and indicators of inflammation with dietary supplementation of tryptophan and methionine in the immediate post-weaning period. Anim. Prod. Sci. 2016, 57(5), pp.935-947.
- Jayaraman, B., Htoo, J.K. and Nyachoti, C.M. Effects of different dietary tryptophan: lysine ratios and sanitary conditions on growth performance, plasma urea nitrogen, serum haptoglobin and ileal histomorphology of weaned pigs. Anim. Sci. J 2017, 88(5), pp.763-771.
- Jayaraman, B., Regassa, A., Htoo, J.K. and Nyachoti, C.M. Effects of dietary standardized ileal digestible tryptophan: lysine ratio on performance, plasma urea nitrogen, ileal histomorphology and immune responses in weaned pigs challenged with Escherichia coli K88. Livest. Sci. 2017, 203, pp.114-119.
- Yu, D.Y., Lindemann, M.D., Quant, A.D., Jang, Y.D., Payne, R.L. and Kerr, B.J. Antibiotic inclusion in the diet did not alter the standardized ileal digestible tryptophan to lysine ratio for growing pigs. J. Anim. Sci. 2017, 95(12), pp.5516-5523.
- Capozzalo, M.M., Kim, J.C., Htoo, J.K., de Lange, C.F., Mullan, B.P., Hansen, C.F., Resink, J.W., Stumbles, P.A., Hampson, D.J. and Pluske, J.R. Effect of increasing the dietary tryptophan to lysine ratio on plasma levels of tryptophan, kynurenine and urea and on production traits in weaner pigs experimentally infected with an enterotoxigenic strain of Escherichia coli. Arch. Anim. Nutr. 2015, 69(1), pp.17-29.
- Gonçalves, M.A.D., Nitikanchana, S., Tokach, M.D., Dritz, S.S., Bello, N.M., Goodband, R.D., Touchette, K.J., Usry, J.L., DeRouchey, J.M. and Woodworth, J.C. Effects of standardized ileal digestible tryptophan: lysine ratio on growth performance of nursery pigs. J. Anim. Sci. 2015, 93(8), pp.3909-3918.
- Nørgaard, J.V., Pedersen, T.F., Soumeh, E.A., Blaabjerg, K., Canibe, N., Jensen, B.B. and Poulsen, H.D. Optimum standardized ileal digestible tryptophan to lysine ratio for pigs weighing 7–14 kg. Livest. Sci. 2015, 175, pp.90-95.
- Shen, Y.B., Coffey, M.T. and Kim, S.W.,. Effects of short-term supplementation of L-tryptophan and reducing large neutral amino acid along with L-tryptophan supplementation on growth and stress response in pigs. Anim. Feed Sci. Technol. 2015, 207, pp.245-252.
- Naatjes, M., Htoo, J.K., Walter, K., Tölle, K.H. and Susenbeth, A. Effect of dietary tryptophan to lysine ratio on growth of young pigs fed wheat–barley or corn based diets. Livest. Sci. 2014, 163, pp.102-109.
- Borgesa, G., Woyengo, T.A. and Nyachoti, C.M. Optimum ratio of standardized ileal digestible tryptophan to lysine for piglets. J. Anim. Feed Sci. 2013, 22, 323–328.
- Capozzalo, M.M., Kim, J.C., Htoo, J.K., de Lange, C.F.M., Mullan, B.P., Hansen, C.F., Resink, J.W., Stumbles, P.A., Hampson, D.J. and Pluske, J.R. An increased ratio of dietary tryptophan to lysine improves feed efficiency and elevates plasma tryptophan and kynurenine in the absence of antimicrobials and regardless of infection with enterotoxigenic Escherichia coli in weaned pigs. J. Anim. Sci. 2012, 90(suppl_4), pp.191-193.
- Quant, A.D., Lindemann, M.D., Kerr, B.J., Payne, R.L. and Cromwell, G.L. Standardized ileal digestible tryptophan-to-lysine ratios in growing pigs fed corn-based and non-corn-based diets. J. Anim. Sci. 2012, 90(4), pp.1270-1279.
- Shen, Y.B., Voilqué, G., Kim, J.D., Odle, J. and Kim, S.W. Effects of increasing tryptophan intake on growth and physiological changes in nursery pigs. J. Anim. Sci. 2012, 90(7), pp.2264-2275.
- Zhang, G.J., Song, Q.L., Xie, C.Y., Chu, L.C., Thacker, P.A., Htoo, J.K. and Qiao, S.Y. Estimation of the ideal standardized ileal digestible tryptophan to lysine ratio for growing pigs fed low crude protein diets supplemented with crystalline amino acids. Livest. Sci. 2012, 149(3), pp.260-266.
- Le Floc’h, N., Matte, J.J., Melchior, D., Van Milgen, J. and Sève, B. A moderate inflammation caused by the deterioration of housing conditions modifies Trp metabolism but not Trp requirement for growth of post-weaned piglets. Animal 2010, 4(11), pp.1891-1898.
- Trevisi, P., Corrent, E., Messori, S., Casini, L. and Bosi, P. Healthy newly weaned pigs require more tryptophan to maximize feed intake if they are susceptible to Escherichia coli K88. Livest. Sci. 2010, 134(1-3), pp.236-238.
- Eder, K., Nonn, H., Kluge, H. and Peganova, S. Tryptophan requirement of growing pigs at various body weights. J. Anim. Physiol. Anim. Nutr. 2003, 87(9-10), pp.336-346.
- Goodarzi, P., Habibi, M., Gorton, M.W., Walsh, K., Tarkesh, F., Fuhrig, M. and Pezeshki, A. Dietary Isoleucine and Valine: Effects on Lipid Metabolism and Ureagenesis in Pigs Fed with Protein Restricted Diets. Metabolites 2023, 13(1), p.89.
- Siebert, D., Khan, D.R. and Torrallardona, D. The optimal valine to lysine ratio for performance parameters in weaned piglets. Animals 2021, 11(5), p.1255.
- Millet, S., Minussi, I., Lambert, W., Aluwé, M., Ampe, B., De Sutter, J. and De Campeneere, S. Standardized ileal digestible lysine and valine-to-lysine requirements for optimal performance of 4 to 9-week-old Piétrain cross piglets. Livest. Sci 2020, 241, p.104263.
- Oliveira, M.S., Htoo, J.K., González-Vega, J.C. and Stein, H.H. Bioavailability of valine in spray-dried L-valine biomass is not different from that in crystalline L-valine when fed to weanling pigs. J. Anim. Sci. 2019, 97(10), pp.4227-4234.
- Gonçalves, M.A. , Tokach, M.D., Dritz, S.S., Bello, N.M., Touchette, K.J., Goodband, R.D., DeRouchey, J.M. and Woodworth, J.C. Standardized ileal digestible valine: lysine dose response effects in 25-to 45-kg pigs under commercial conditions. J. Anim. Sci. 2018, 96(2), 591–599. [Google Scholar]
- Xu, Y.T., Ma, X.K., Wang, C.L., Yuan, M.F. and Piao, X.S. Effects of dietary valine: lysine ratio on the performance, amino acid composition of tissues and mRNA expression of genes involved in branched-chain amino acid metabolism of weaned piglets. Asian-Australas. j. anim. sci. 2018, 31(1), p.106.
- Zhang, X., Liu, X., Jia, H., He, P., Mao, X., Qiao, S. and Zeng, X. Valine supplementation in a reduced protein diet regulates growth performance partially through modulation of plasma amino acids profile, metabolic responses, endocrine, and neural factors in piglets. J. Agric. Food Chem. 2018, 66(12), pp.3161-3168.
- Clark, A.B., Tokach, M.D., DeRouchey, J.M., Dritz, S.S., Goodband, R.D., Woodworth, J.C., Touchette, K.J. and Bello, N.M. Modeling the effects of standardized ileal digestible valine to lysine ratio on growth performance of nursery pigs. Transl. anim. sci. 2017, 1(4), pp.448-457.
- Liu, X.T., Ma, W.F., Zeng, X.F., Xie, C.Y., Thacker, P.A., Htoo, J.K. and Qiao, S.Y. Estimation of the standardized ileal digestible valine to lysine ratio required for 25-to 120-kilogram pigs fed low crude protein diets supplemented with crystalline amino acids. J. Anim. Sci. 2015, 93(10), pp.4761-4773.
- Soumeh, E.A., van Milgen, J., Sloth, N.M., Corrent, E., Poulsen, H.D. and Nørgaard, J.V. Requirement of standardized ileal digestible valine to lysine ratio for 8-to 14-kg pigs. Animal 2015, 9(8), pp.1312-1318.
- Nemechek, J.E., Tokach, M.D., Dritz, S.S., Goodband, R.D. and DeRouchey, J.M. Evaluation of standardized ileal digestible valine: lysine, total lysine: crude protein, and replacing fish meal, meat and bone meal, and poultry byproduct meal with crystalline amino acids on growth performance of nursery pigs from seven to twelve kilograms. J. Anim. Sci. 2014, 92(4), pp.1548-1561.
- Millet, S. The interaction between dietary valine and tryptophan content and their effect on the performance of piglets. Animals 2012, 2(1), pp.76-84.
- Waguespack, A.M., Bidner, T.D., Payne, R.L. and Southern, L.L. Valine and isoleucine requirement of 20-to 45-kilogram pigs. J. Anim. Sci. 2012, 90(7), pp.2276-2284.
- Gaines, A.M., Kendall, D.C., Allee, G.L., Usry, J.L. and Kerr, B.J.,. Estimation of the standardized ileal digestible valine-to-lysine ratio in 13-to 32-kilogram pigs. J. Anim. Sci. 2011, 89(3), pp.736-742.
- Gloaguen, M., Le Floc'H, N., Brossard, L., Barea, R., Primot, Y., Corrent, E. and Van Milgen, J. Response of piglets to the valine content in diet in combination with the supply of other branched-chain amino acids. Animal 2011, 5(11), pp.1734-1742.
- Barea, R., Brossard, L., Le Floc'H, N., Primot, Y., Melchior, D. and Van Milgen, J. The standardized ileal digestible valine-to-lysine requirement ratio is at least seventy percent in postweaned piglets. J. Anim. Sci. 2009, 87(3), pp.935-947.
- Nørgaard, J.V. and Fernández, J.A. Isoleucine and valine supplementation of crude protein-reduced diets for pigs aged 5–8 weeks. Anim. Feed Sci. Technol. 2009, 154(3-4), pp.248-253.
- Wiltafsky, M.K., Schmidtlein, B. and Roth, F.X. Estimates of the optimum dietary ratio of standardized ileal digestible valine to lysine for eight to twenty-five kilograms of body weight pigs. J. Anim. Sci. 2009, 87(8), pp.2544-2553.
- Clark, A.B., Tokach, M.D., DeRouchey, J.M., Dritz, S.S., Goodband, R.D., Woodworth, J.C., Touchette, K.J. and Bello, N.M. Modeling the effects of standardized ileal digestible isoleucine to lysine ratio on growth performance of nursery pigs. Transl. anim. sci. 2017, 1(4), pp.437-447.
- Lazzeri, D.B., Castilha, L.D., Costa, P.B., Nunes, R.V., dos Santos Pozza, M.S. and Pozza, P.C. Standardized ileal digestible (SID) isoleucine requirement of barrows (15-to 30-kg) fed low crude protein diets. Semin. Cienc. Agrar. 2017, 38(5), pp.3283-3294.
- Clark, A.B., Tokach, M.D., DeRouchey, J.M., Dritz, S.S., Touchette, K., Goodband, R.D. and Woodworth, J.C. Effects of Dietary Standardized Ileal Digestible Isoleucine: Lysine Ratio on Nursery Pig Performance. KAES Research Reports 2016, 2(8), p.12.
- Htoo, J.K., Zhu, C.L., Huber, L., de Lange, C.F.M., Quant, A.D., Kerr, B.J., Cromwell, G.L. and Lindemann, M.D. Determining the optimal isoleucine: lysine ratio for ten-to twenty-two-kilogram and twenty-four-to thirty-nine-kilogram pigs fed diets containing nonexcess levels of leucine. J. Anim. Sci. 2014, 92(8), pp.3482-3490.
- Soumeh, E.A., Van Milgen, J., Sloth, N.M., Corrent, E., Poulsen, H.D. and Nørgaard, J.V. The optimum ratio of standardized ileal digestible isoleucine to lysine for 8–15 kg pigs. Anim. Feed Sci. Technol. 2014, 198, pp.158-165.
- Gloaguen, M. , Le Floc'H, N., Primot, Y., Corrent, E. and Van Milgen, J. Response of piglets to the standardized ileal digestible isoleucine, histidine and leucine supply in cereal–soybean meal-based diets. Animal 2013, 7(6), 901–908. [Google Scholar] [CrossRef] [PubMed]
- Nørgaard, J.V. , Shrestha, A., Krogh, U., Sloth, N.M., Blaabjerg, K., Poulsen, H.D., Tybirk, P. and Corrent, E. Isoleucine requirement of pigs weighing 8 to 18 kg fed blood cell–free diets. J. Anim. Sci. 2013, 91(8), 3759–3765. [Google Scholar] [CrossRef] [PubMed]
- Barea, R. , Brossard, L., Le Floc'H, N., Primot, Y. and Van Milgen, J. The standardized ileal digestible isoleucine-to-lysine requirement ratio may be less than fifty percent in eleven-to twenty-three-kilogram piglets1. J. Anim. Sci. 2009, 87(12), 4022–4031. [Google Scholar] [CrossRef] [PubMed]
- Wiltafsky, M.K. , Bartelt, J., Relandeau, C. and Roth, F.X. Estimation of the optimum ratio of standardized ileal digestible isoleucine to lysine for eight-to twenty-five-kilogram pigs in diets containing spray-dried blood cells or corn gluten feed as a protein source. J. Anim. Sci. 2009, 87(8), 2554–2564. [Google Scholar] [CrossRef]
- da Silva, C.A. , Dias, C.P., Callegari, M.A., de Souza, K.L., Barbi, J.H., Fagundes, N.S., Batonon-Alavo, D.I. and Foppa, L. Increased Sulphur Amino Acids Consumption as OH-Methionine or DL-Methionine Improves Growth Performance and Carcass Traits of Growing-Finishing Pigs Fed under Hot Conditions. Animals 2022, 12(17), 2159. [Google Scholar]
- Yang, Z. , Hasan, M.S., Humphrey, R.M., Htoo, J.K. and Liao, S.F. Changes in growth performance, plasma metabolite concentrations, and myogenic gene expression in growing pigs fed a methionine-restricted diet. Front. Biosci. (Landmark Ed). 2021, 26(9), 413–422. [Google Scholar]
- Ho, T.T. , Htoo, J.K.K., Dao, T.B.A., Carpena, M.E., Le, N.A.T., Vu, C.C. and Nguyen, Q.L. Estimation of the standardized ileal digestible lysine requirement and optimal sulphur amino acids to lysine ratio for 30–50 kg pigs. J. Anim. Physiol. Anim. Nutr. 2019, 103(1), 258–268. [Google Scholar] [CrossRef]
- Remus, A. , Pomar, C., Perondi, D., Gobi, J.P., da Silva, W.C., de Souza, L.J. and Hauschild, L. Response to dietary methionine supply of growing pigs fed daily tailored diets or fed according to a conventional phase feeding system. Livest. Sci. 2019, 222, 7–13. [Google Scholar] [CrossRef]
- Kahindi, R.K. , Regassa, A., Htoo, J.K. and Nyachoti, C.M. Growth performance and expression of genes encoding enzymes involved in methionine and cysteine metabolism in piglets fed increasing sulphur amino acid to lysine ratio during enterotoxigenic Escherichia coli challenge. Can. J. Anim. Sci. 2018, 98(2), 333–340. [Google Scholar]
- Capozzalo, M.M. , Resink, J.W., Htoo, J.K., Kim, J.C., de Lange, F.M., Mullan, B.P., Hansen, C.F. and Pluske, J.R. Determination of the optimum standardised ileal digestible sulphur amino acids to lysine ratio in weaned pigs challenged with enterotoxigenic Escherichia coli. Anim. Feed Sci. Technol. 2017, 227, 118–130. [Google Scholar]
- Capozzalo, M.M. , Kim, J.C., Htoo, J.K., De Lange, C.F.M., Mullan, B.P., Hansen, C.F., Resink, J.W. and Pluske, J.R. Pigs experimentally infected with an enterotoxigenic strain of Escherichia coli have improved feed efficiency and indicators of inflammation with dietary supplementation of tryptophan and methionine in the immediate post-weaning period. Anim. Prod. Sci. 2016, 57(5), 935–947. [Google Scholar]
- Kahindi, R. , Regassa, A., Htoo, J. and Nyachoti, M. Optimal sulfur amino acid to lysine ratio for post weaning piglets reared under clean or unclean sanitary conditions. Anim. Nutr. 2017, 3(4), 380–385. [Google Scholar] [CrossRef] [PubMed]
- Kaewtapee, C. , Krutthai, N. and Bunchasak, C. Effects of supplemental liquid DL-methionine hydroxy analog free acid in diet on growth performance and gastrointestinal functions of piglets. Asian-Australas. J. Anim. Sci. 2016, 29(8), 1166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.J., Thacker, P.A., Htoo, J.K. and Qiao, S.Y. Optimum proportion of standardized ileal digestible sulfur amino acid to lysine to maximize the performance of 25-50 kg growing pigs fed reduced crude protein diets fortified with amino acids. Czech J. Anim. Sci. 2015, 60, (7): 302–310.
- Chen, Y. , Li, D., Dai, Z., Piao, X., Wu, Z., Wang, B., Zhu, Y. and Zeng, Z. L-methionine supplementation maintains the integrity and barrier function of the small-intestinal mucosa in post-weaning piglets. Amino Acids 2014, 46, 1131–1142. [Google Scholar] [CrossRef]
- Conde-Aguilera, J.A. , Cobo-Ortega, C., Mercier, Y., Tesseraud, S. and Van Milgen, J. The amino acid composition of tissue protein is affected by the total sulfur amino acid supply in growing pigs. Animal 2014, 8(3), 401–409. [Google Scholar] [CrossRef]
- Conde-Aguilera, J.A. , Barea, R., Le Floc’h, N., Lefaucheur, L. and Van Milgen, J. A sulfur amino acid deficiency changes the amino acid composition of body protein in piglets. Animal 2010, 4(8), 1349–1358. [Google Scholar] [CrossRef]
- Frantz, N.Z. , Tokach, M.D., Goodband, R.D., Dritz, S.S., DeRouchey, J.M., Nelssen, J.L. and Jones, C.L. The optimal standardized ileal digestible lysine and total sulfur amino acid requirement for finishing pigs fed ractopamine hydrochloride. The professional animal scientist 2009, 25(2), 161–168. [Google Scholar] [CrossRef]
- Ogbuewu, I.; Mokolopi, B.; Mbajiorgu, C. Meta-analysis of growth performance indices of broiler chickens in response to turmeric (Curcuma longa L.) supplementation. Anim. Feed Sci. Technol. 2022, 283, 115155. [Google Scholar] [CrossRef]
- Guzik, A.C. , Pettitt, M.J., Beltranena, E., Southern, L.L., Kerr, J. Threonine and tryptophan ratios fed to nursery pigs. J. Anim. Physiol. Anim. Nutr. 2005, 89, 297–302. [Google Scholar] [CrossRef]
- Le Floc’h, N., Seve, B. Biological roles of tryptophan and its metabolism: potential implications for pig feeding. Livest. Sci. 2007,112, 23–32.
- Simongiovanni, A., Corrent, E., Le Floc’h, N., Van Milgen, J. Estimation of the tryptophan requirement in piglets by meta-analysis. Animal 2012, 6, 594–602.
- Oliveira, G.M. , Ferreira, A.S., Campos, P.F., Rodrigues, V.V., Silva, F.C.O., Santos, W.G., Lima, A.L., Rodrigues, P.G. and Lopes, C.C. Digestible tryptophan to lysine ratios for weaned piglets at 26 days of age. Anim. Prod. Sci. 2016, 57(10), 2027–2032. [Google Scholar] [CrossRef]
- Liu, J.B. , Yan, H.L., Cao, S.C., Liu, J., Li, Z.X. and Zhang, H.F. The response of performance in grower and finisher pigs to diets formulated to different tryptophan to lysine ratios. Livest. Sci. 2019, 222, 25–30. [Google Scholar] [CrossRef]
- Trevisi, P. , Corrent, E., Messori, S., Casini, L. and Bosi, P. Healthy newly weaned pigs require more tryptophan to maximize feed intake if they are susceptible to Escherichia coli K88. Livest. Sci. 2010, 134(1-3), 236–238. [Google Scholar] [CrossRef]
- Le Floc'h, N. , Lebellego, L., Matte, J.J., Melchior, D. and Sève, B. The effect of sanitary status degradation and dietary tryptophan content on growth rate and tryptophan metabolism in weaning pigs. J. Anim. Sci. 2009, 87(5), 1686–1694. [Google Scholar] [CrossRef] [PubMed]
- Gatnau, R. , Zimmerman, D.R., Nissen, S.L., Wannemuehler, M. and Ewan, R.C. Effects of excess dietary leucine and leucine catabolites on growth and immune responses in weanling pigs. J. Anim. Sci. 1995, 73(1), 159–165. [Google Scholar] [CrossRef] [PubMed]
- Wiltafsky, M.K. , Pfaffl, M.W. and Roth, F.X. The effects of branched-chain amino acid interactions on growth performance, blood metabolites, enzyme kinetics and transcriptomics in weaned pigs. Br. J. Nutr. 2010, 103(7), 964–976. [Google Scholar] [CrossRef]
- Wessels, A.G. , Kluge, H., Hirche, F., Kiowski, A., Schutkowski, A., Corrent, E., Bartelt, J., König, B. and Stangl, G.I. High leucine diets stimulate cerebral branched-chain amino acid degradation and modify serotonin and ketone body concentrations in a pig model. PloS one 2016, 11(11), e0150376. [Google Scholar] [CrossRef]
- Harper, A.E. , Miller, R. and Block, K.P. Branched-chain amino acid metabolism. Annu. Rev. Nutr. 1984, 4(1), 409–454. [Google Scholar] [CrossRef]
- Meyer, F. , Van Rensburg, C.J. and Gous, R.M. The response of weaned piglets to dietary valine and leucine. Animal 2017, 11(8), 1279–1286. [Google Scholar] [CrossRef]
- Cemin, H.S. , Tokach, M.D., Dritz, S.S., Woodworth, J.C., DeRouchey, J.M. and Goodband, R.D. Meta-regression analysis to predict the influence of branched-chain and large neutral amino acids on growth performance of pigs. J. Anim. Sci. 2019, 97(6), 2505–2514. [Google Scholar] [CrossRef]
- Morales, A. , Arce, N., Cota, M., Buenabad, L., Avelar, E., Htoo, J.K. and Cervantes, M. Effect of dietary excess of branched-chain amino acids on performance and serum concentrations of amino acids in growing pigs. J. Anim. Physiol. Anim. Nutr. 2016, 100(1), 39–45. [Google Scholar] [CrossRef]
- Langer, S. , Scislowski, P.W., Brown, D.S., Dewey, P. and Fuller, M.F. Interactions among the branched-chain amino acids and their effects on methionine utilization in growing pigs: Effects on plasma amino–and keto–acid concentrations and branched-chain keto-acid dehydrogenase activity. Br. J. Nutr. 2000, 83(1), 49–58. [Google Scholar]
- Grimble, R.F. The effects of sulfur amino acid intake on immune function in humans. J. Nutr. 2006, 136(6), 1660S–1665S. [Google Scholar] [CrossRef] [PubMed]
- Jahoor, F., Wykes, L.J., Reeds, P.J., Henry, J.F., Del Rosario, M.P. and Frazer, M.E. Protein-deficient pigs cannot maintain reduced glutathione homeostasis when subjected to the stress of inflammation. J. Nutr 1995, 125(6), pp.1462-1472.
- Kim, J.C. , Mullan, B.P., Frey, B., Payne, H.G. and Pluske, J.R. Whole body protein deposition and plasma amino acid profiles in growing and/or finishing pigs fed increasing levels of sulfur amino acids with and without Escherichia coli lipopolysaccharide challenge. J. Anim. Sci. 2012, 90 (suppl_4), 362–365. [Google Scholar] [CrossRef] [PubMed]
- Maroufyan, E. , Kasim, A., Hashemi, S.R., Loh, T.C., Bejo, M.H. and Davoodi, H. The effect of methionine and threonine supplementations on immune responses of broiler chickens challenged with infectious bursal disease. Am. J. Appl. Sci. 2010, 7(1), 44. [Google Scholar] [CrossRef]
- Rakhshandeh, A. , Htoo, J.K., Karrow, N., Miller, S.P. and de Lange, C.F. Impact of immune system stimulation on the ileal nutrient digestibility and utilisation of methionine plus cysteine intake for whole-body protein deposition in growing pigs. Br. J. Nutr. 2014, 111(1), 101–110. [Google Scholar] [CrossRef]
- Rostagno, H.S. , Albino, L.F.T., Donzele, J.L., Gomes, P.C., Oliveira, R.D., Lopes, D.C., Ferreira, A.S., Barreto, S.D.T. and Euclides, R.F. Composição de alimentos e exigências nutricionais. Tabelas brasileiras para aves e suínos, 2005, 2. [Google Scholar]
- Sève, B. Alimentation du porc en croissance: intégration des concepts de protéine idéale, de disponibilité digestive des acides aminés et d'énergie nette. Prod. Anim. 1994, 7(4), 275–291. [Google Scholar] [CrossRef]
- Grimble, R.F. and Grimble, G.K. Immunonutrition: role of sulfur amino acids, related amino acids, and polyamines. Nutrition 1998, 14(7-8), 605–610. [Google Scholar] [CrossRef]
- Le Floc'h, N. , Melchior, D. and Obled, C. Modifications of protein and amino acid metabolism during inflammation and immune system activation. Livest. Prod. Sci. 2004, 87(1), 37–45. [Google Scholar] [CrossRef]
- Melchior, D. , Sève, B. and Le Floc'H, N. Chronic lung inflammation affects plasma amino acid concentrations in pigs. J. Anim. Sci. 2004, 82(4), 1091–1099. [Google Scholar] [CrossRef]
- Tsukahara, T. , Inoue, R., Nakatani, M., Fukuta, K., Kishino, E., Ito, T. and Ushida, K. Influence of weaning age on the villous height and disaccharidase activities in the porcine small intestine. Anim. Sci. J. 2016, 87(1), 67–75. [Google Scholar] [CrossRef]
- Lu, S.C. Regulation of glutathione synthesis. Mol. Asp. Med. 2009, 30(1-2), 42–59. [Google Scholar] [CrossRef]
Figure 1.
Systematic literature search selection process. The PRISMA diagram details the applied search and selection process for Trp.
Figure 1.
Systematic literature search selection process. The PRISMA diagram details the applied search and selection process for Trp.
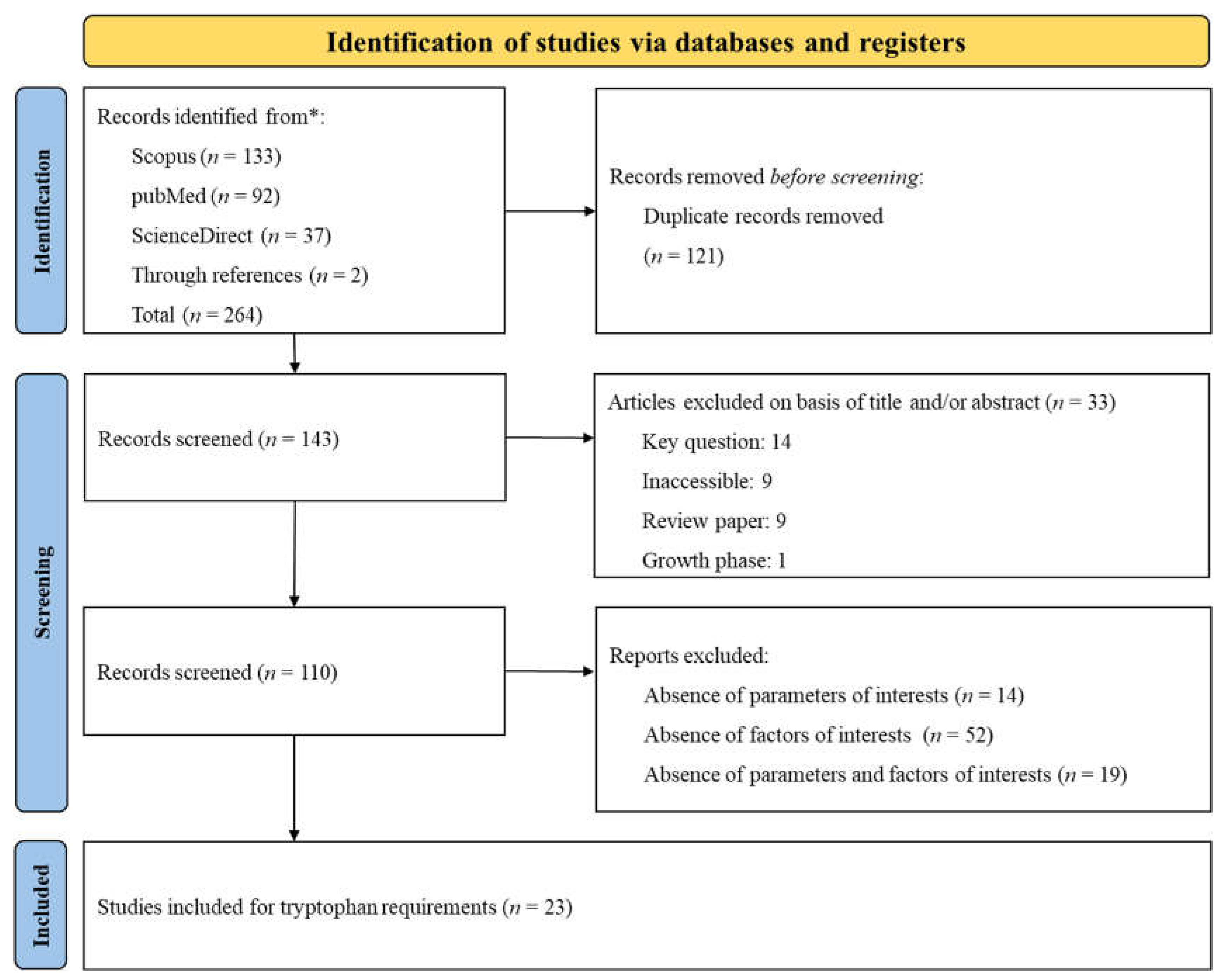
Figure 2.
SID Trp: Lys ratio requirements by body weight classification to maximize GF in pigs.
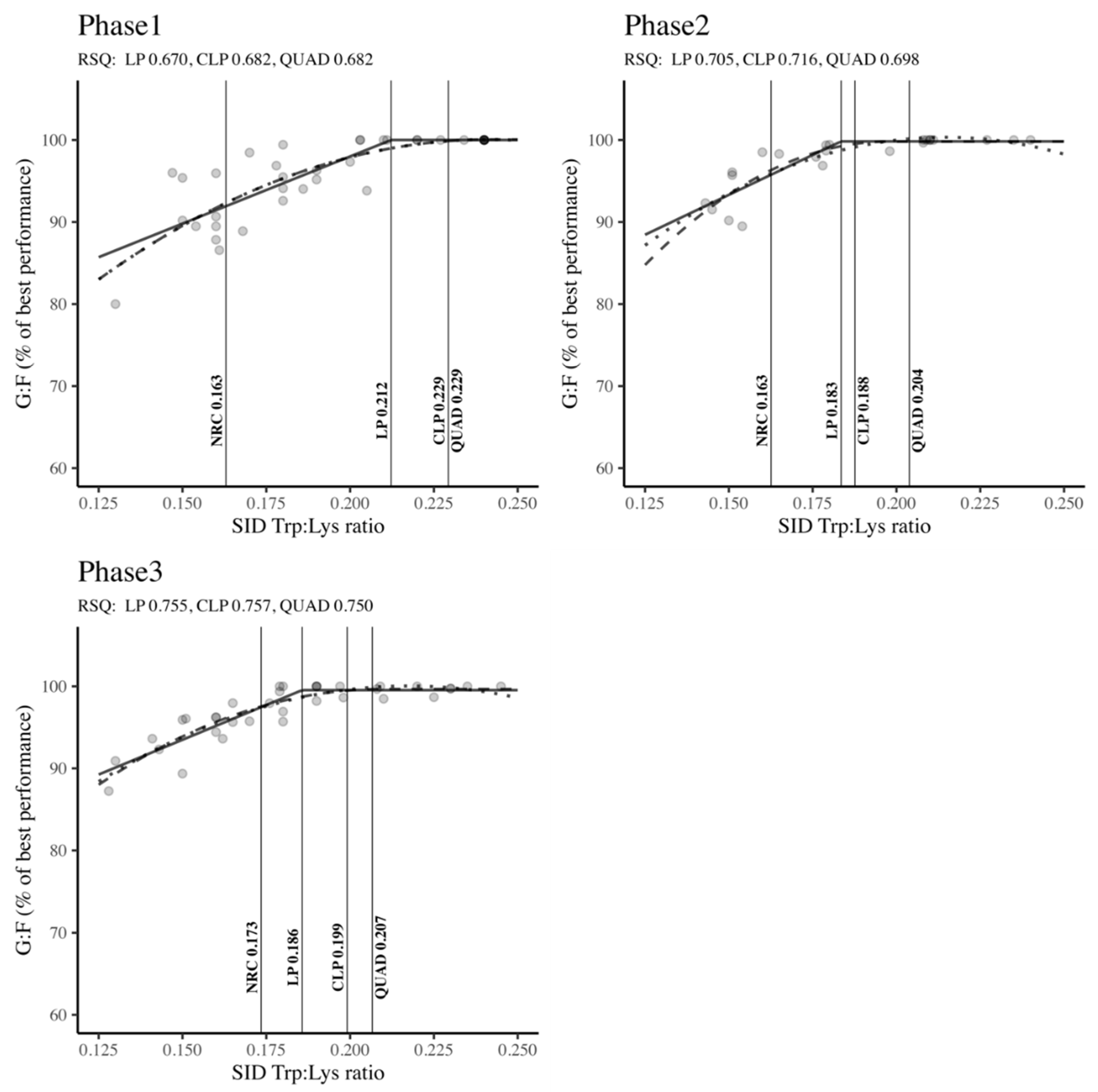
Figure 3.
Systematic literature search selection process. The PRISMA diagram details the applied search and selection process for Val.
Figure 3.
Systematic literature search selection process. The PRISMA diagram details the applied search and selection process for Val.
Figure 4.
SID Val: Lys ratio requirements by body weight classification to maximize GF in pigs.
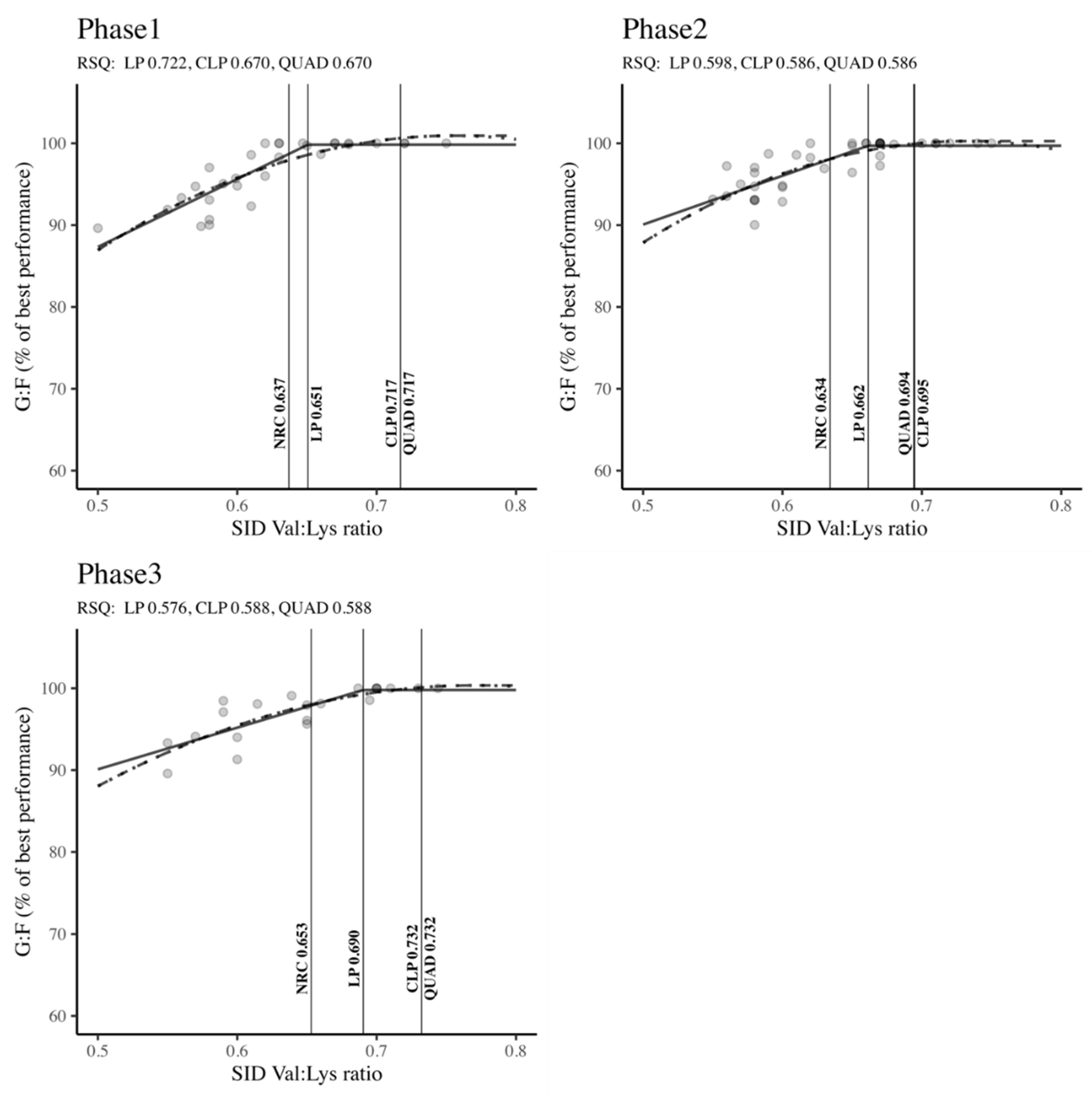
Figure 5.
Systematic literature search selection process. The PRISMA diagram details the applied search and selection process for Ile.
Figure 5.
Systematic literature search selection process. The PRISMA diagram details the applied search and selection process for Ile.
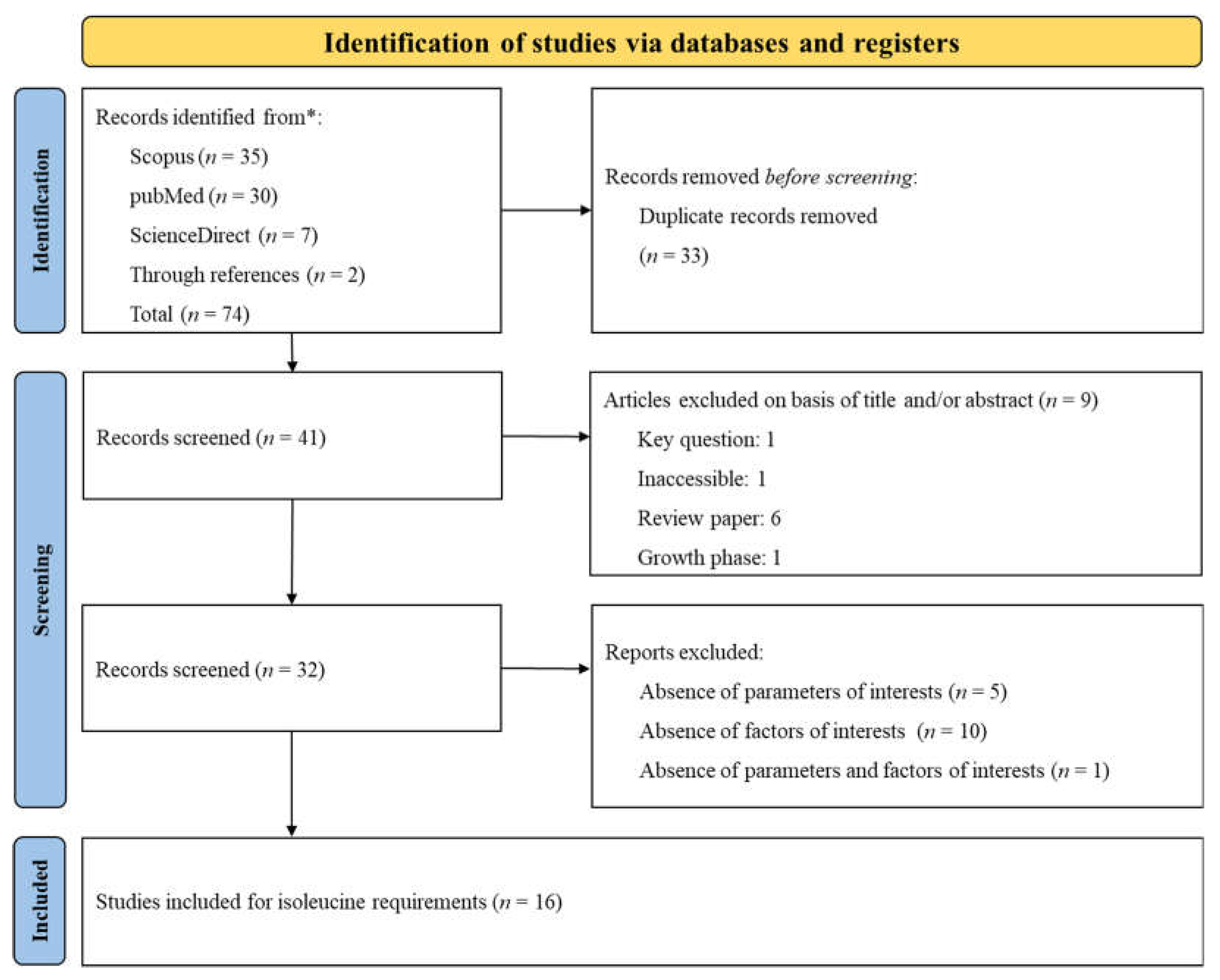
Figure 6.
SID Ile: Lys ratio requirements by body weight classification to maximize GF in pigs.
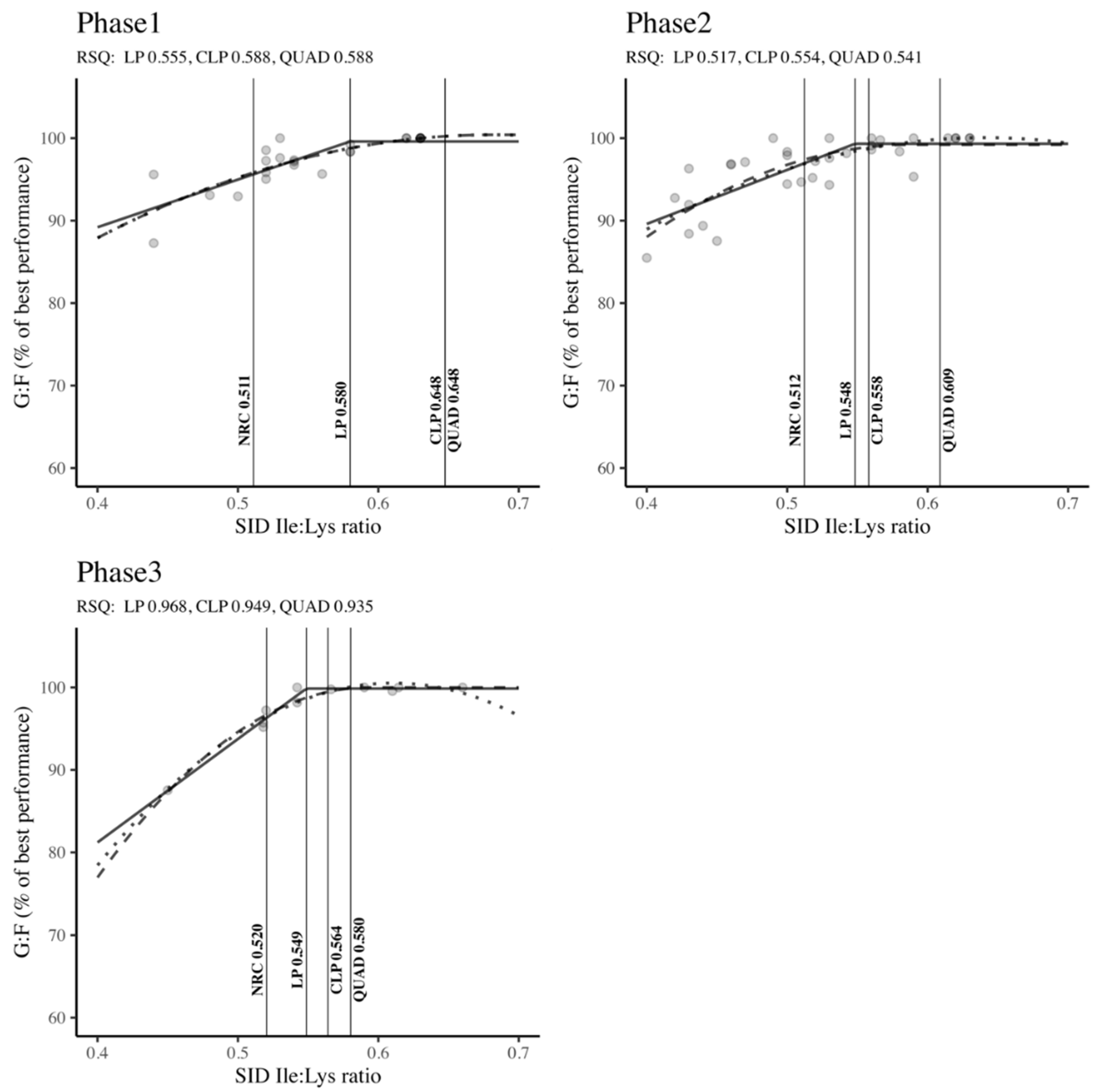
Figure 7.
Systematic literature search selection process. The PRISMA diagram details the applied search and selection process for Met.
Figure 7.
Systematic literature search selection process. The PRISMA diagram details the applied search and selection process for Met.
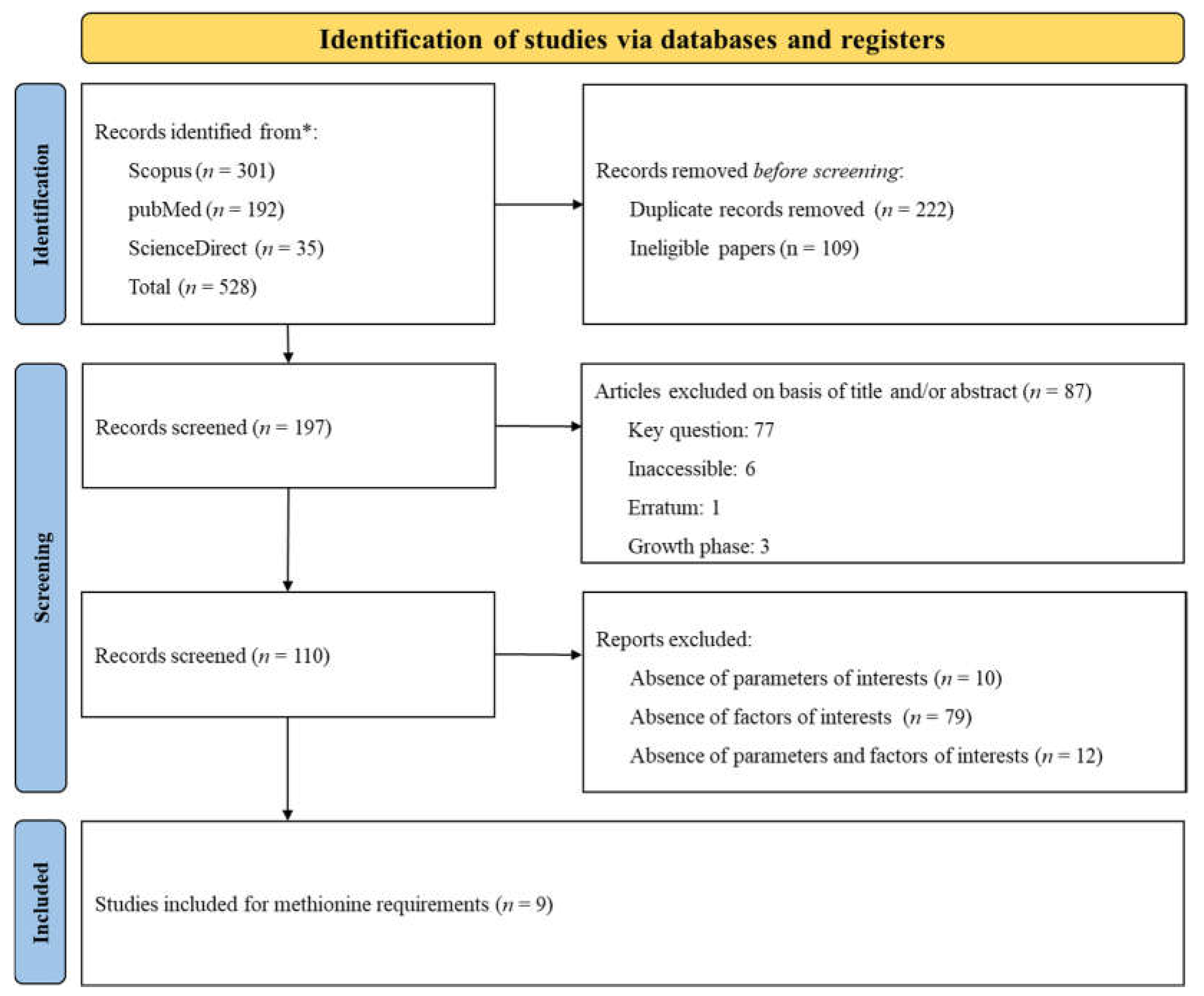
Figure 8.
SID Met: Lys ratio requirements by body weight classification to maximize GF in pigs.
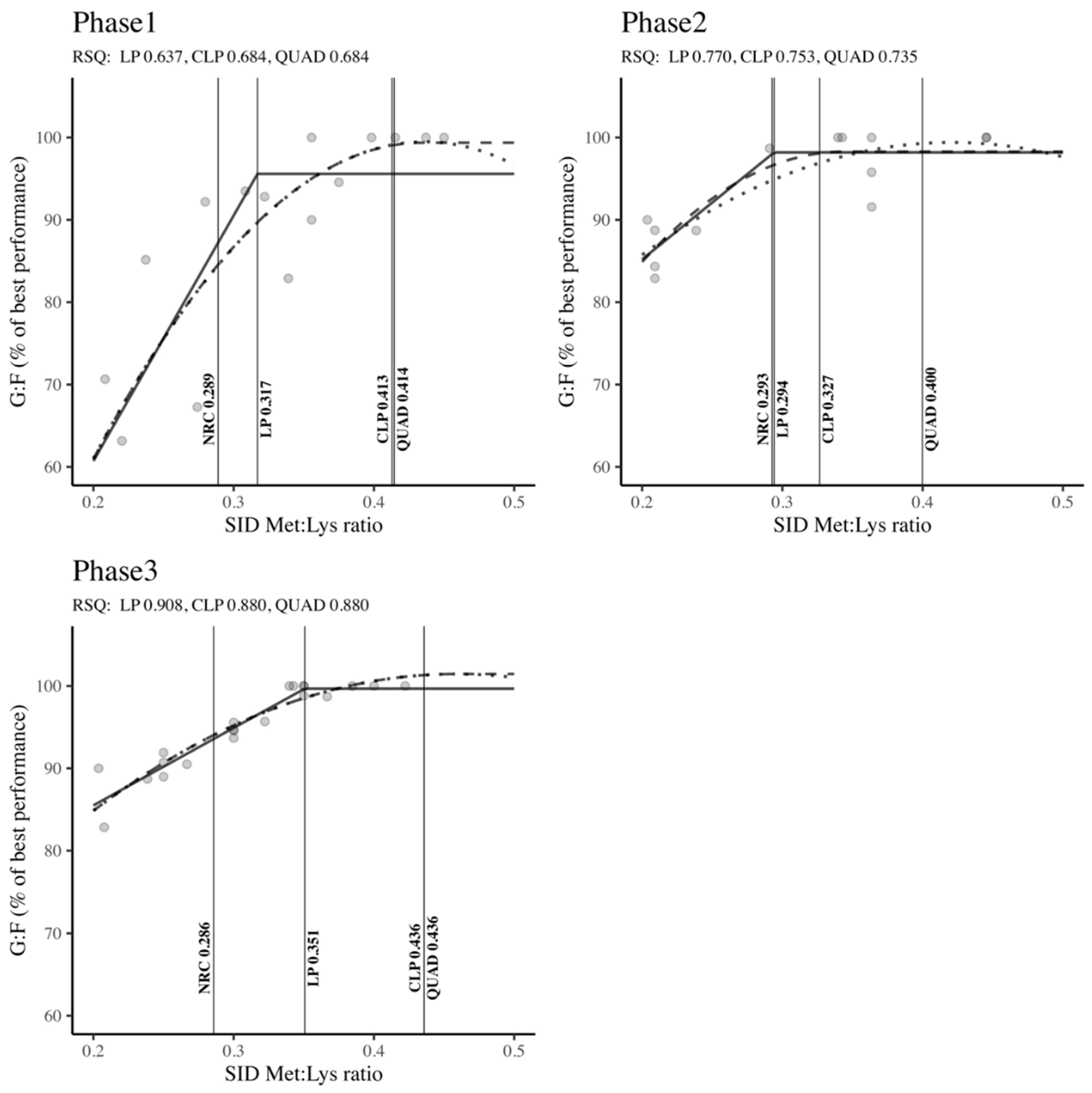

Table 1.
Information on studies used in meta-regression analysis for estimating tryptophan requirements of pigs.
Table 1.
Information on studies used in meta-regression analysis for estimating tryptophan requirements of pigs.
| Source | Phase1 | Range of SID Trp: Lys ratio |
Range of SID Lys, % |
Remarks |
| Kwon et al. [7] | 3 | 0.18-0.28 | 1.00 | |
| Tolosa et al. [24] | 3 | 0.16-0.19 | 0.81-0.89 | |
| Kerkaert et al. [25] | 3 | 0.19-0.21 | 0.98 | |
| Ma et al. [26] | 1 | 0.15-0.24 | 1.10 | |
| Capozzalo et al. [27] | 1 | 0.17-0.25 | 1.23-1.27 | |
| Liu et al. [28] | 2-3 | 0.15-0.23 | 0.90 | |
| Wensley et al. [29] | 2 | 0.16-0.21 | 1.25 | |
| Gonçalves et al. [30] | 3 | 0.15-0.25 | 0.90 | |
| Capozzalo et al. [31] | 1 | 0.16-0.24 | 1.25 | E. coli infection |
| Jayaraman et al. [32] | 1 | 0.16-0.25 | 1.18 | E. coli infection |
| Jayaraman et al. [33] | 1 | 0.19-0.24 | 1.23-1.33 | Unclean condition |
| Yu et al. [34] | 3 | 0.13-0.21 | 0.66 | |
| Capozzalo et al. [35] | 1 | 0.16-0.24 | 1.24 | E. coli infection |
| Gonçalves et al. [36] | 1-2 | 0.15-0.25 | 0.97-1.30 | |
| Nørgaard et al. [37] | 1 | 0.13-0.23 | 1.09 | |
| Shen et al. [38] | 2-3 | 0.16-1.10 | 0.90-1.17 | Stress |
| Naatjes et al. [39] | 2-3 | 0.13-0.21 | 1.05 | |
| Borgesa et al. [40] | 1 | 0.18-0.24 | 0.92 | |
| Capozzalo et al. [41] | 1 | 0.17-0.26 | 1.24 | E. coli infection |
| Quant et al. [42] | 3 | 0.13-0.18 | 0.66 | |
| Shen et al. [43] | 2 | 0.21-1.31 | 0.90 | |
| Zhang et al. [44] | 3 | 0.13-0.25 | 0.90 | |
| Le Floc’h et al. [45] | 1-3 | 0.15-0.24 | 1.05-1.22 | |
| Trevisi et al. [46] | 1 | 0.18-0.22 | ND2 | E. coli infection |
| Eder et al. [47] | 3 | 0.09-0.23 | 0.87 |
1Phase1 (BW < 11), Phase2 (11 ≤ BW < 25), Phase3 (25 ≤ BW< 50). 2No description.
Table 2.
Information on studies used in meta-regression analysis for estimating valine requirements for pigs.
Table 2.
Information on studies used in meta-regression analysis for estimating valine requirements for pigs.
| Source | Phase1 | Range of SID Val: Lys ratio |
Range of SID Lys, % |
Range of SID Leu: Lys ratio |
Range of SID Ile: Lys ratio |
Remarks |
| Goodarzi et al. [48] | 1-2 | 0.39-0.75 | 1.29 | 0.82 | 0.29; 0.60 | BCAA interaction |
| Clizer et al. [11] | 3 | 0.60-0.80 | 0.98 | 1.34 | 0.60 | |
| Goodarzi et al. [8] | 1-2 | 0.39-0.75 | 1.29 | 0.81 | 0.30; 0.55 | BCAA interaction |
| Habibi et al. [9] | 1-2 | 0.37-0.62 | 1.29 | 0.77 | 0.31; 0.55 | BCAA interaction |
| Kerkaert et al. [25] | 3 | 0.70-0.80 | 0.98 | 1.45 | 0.61 | |
| Siebert et al. [49] | 1-3 | 0.70-0.76 | 1.15-1.25 | 1.03-1.06 | 0.58 | |
| Millet et al. [50] | 1-2 | 0.58-0.82 | 1.05 | 1.05 | 0.54 | |
| Oliveira et al. [51] | 1 | 0.71-0.87 | 1.41-1.43 | ND2 | ND2 | |
| Gonçalves et al. [52] | 3 | 0.57-0.78 | 0.85 | 1.54-1.58 | 0.61-0.62 | |
| Xu et al. [53] | 1-2 | 0.50-0.80 | 1.17 | 0.99 | 0.53 | |
| Zhang et al. [54] | 2-3 | 0.45-0.65 | 1.15 | ND2 | ND2 | |
| Clark et al. [55] | 1-2 | 0.50-0.85 | 1.24 | 1.10 | 0.57 | |
| Liu et al. [56] | 3 | 0.55-0.75 | 0.90 | 1.13 | 0.51 | |
| Soumeh et al. [57] | 1-2 | 0.58-0.78 | 1.10 | ND2 | ND2 | |
| Nemechek et al. [58] | 1-2 | 0.57-0.70 | 1.30 | ND2 | ND2 | |
| Millet [59] | 1-2 | 0.58-0.67 | 1.06 | 0.96 | 0.52 | |
| Waguespack et al. [60] | 2-3 | 0.61-0.74 | 0.83 | 1.30 | 0.60 | |
| Gaines et al. [61] | 2-3 | 0.55-0.80 | 1.10 | ND2 | ND2 | |
| Gloaguen et al. [62] | 2 | 0.60-0.80 | 0.95-1.02 | 1.01-0.69 | 0.47-0.64 | |
| Barea et al. [63] | 1-2 | 0.57-0.80 | 0.92-1.00 | ND2 | 0.50-0.60 | |
| Nørgaard and Fernández [64] | 1-2 | 0.60-0.72 | 1.00 | 1.02 | 0.53; 0.62 | BCAA interaction |
| Wiltafsky et al. [65] | 1-2 | 0.49-0.84 | 0.93-1.02 | 0.98-1.06 | 0.59-0.64 |
1Phase1 (BW < 11), Phase2 (11 ≤ BW < 25), Phase3 (25 ≤ BW< 50). 2No description.
Table 3.
Information on studies used in meta-regression analysis for estimating isoleucine requirements for pigs.
Table 3.
Information on studies used in meta-regression analysis for estimating isoleucine requirements for pigs.
| Source | Phase1 | Range of SID Ile: Lys ratio |
Range of SID Lys, % |
Range of SID Leu: Lys ratio |
Range of SID Val: Lys ratio |
Remarks |
| Goodarzi et al. [48] | 1-2 | 0.29-0.60 | 1.29 | 0.82 | 0.39-0.75 | BCAA interaction |
| Clizer et al. [11] | 3 | 0.55-0.75 | 0.73 | 1.61 | 0.78 | |
| Goodarzi et al. [8] | 1-2 | 0.30-0.55 | 1.29 | 0.81 | 0.39-0.75 | BCAA interaction |
| Habibi et al. [9] | 1-2 | 0.31-0.55 | 1.29 | 0.77 | 0.37-0.62 | BCAA interaction |
| Kerkaert et al. [25] | 3 | 0.61-0.66 | 0.98 | 1.45 | 0.70 | |
| Clark et al. [66] | 1 | 0.40-0.63 | 1.24-1.28 | 1.07-1.09 | 0.71 | |
| Lazzeri et al. [67] | 2-3 | 0.45-0.73 | 1.06 | 0.99 | 0.65 | |
| Clark et al. [68] | 1-2 | 0.40-0.63 | 1.24-1.28 | 1.07-1.09 | 0.71 | |
| Htoo et al. [69] | 1-3 | 0.33-0.70 | 0.95 | 1.03-1.33 | 0.69-0.89 | |
| Soumeh et al. [70] | 1-2 | 0.42-0.62 | 1.14 | ND2 | 0.70 | |
| Gloaguen et al. [71] | 2 | 0.40-0.55 | 0.94-0.98 | 1.01-1.09 | ND2 | |
| Nørgaard et al. [72] | 1-2 | 0.42-0.62 | 1.12 | ND2 | 0.70 | |
| Waguespack et al. [60] | 2-3 | 0.52-0.61 | 0.83 | 1.30 | 0.73 | |
| Barea et al. [73] | 2 | 0.46-0.65 | 1.00 | ND2 | ND2 | |
| Nørgaard and Fernández [64] | 1-2 | 0.53-0.62 | 1.00 | 1.02 | 0.60-0.72 | BCAA interaction |
| Wiltafsky et al. [74] | 1-2 | 0.36-0.72 | 0.92-1.02 | 1.08-1.62 | 0.68-1.02 |
1Phase1 (BW < 11), Phase2 (11 ≤ BW < 25), Phase3 (25 ≤ BW< 50). 2No description.
Table 4.
Information on studies used in meta-regression analysis for estimating methionine requirements for pigs.
Table 4.
Information on studies used in meta-regression analysis for estimating methionine requirements for pigs.
| Source | Phase1 | Range of SID SAA: Lys ratio |
Range of SID Met: Lys ratio |
Range of SID Lys, % |
Supplemental amino acid2 |
Remarks |
| da Silva et al. [75] | 2-3 | 0.59-0.74 | ND3 | 0.96-1.07 | SAA | |
| Yang et al. [76] | 2-3 | 0.48-0.62 | 0.20-0.34 | 1.08 | SAA+Met | |
| Ho et al. [77] | 3 | 0.50-0.70 | 0.25-0.45 | 1.00 | SAA+Met | |
| Remus et al. [78] | 3 | 0.37-0.57 | 0.21-0.39 | 1.15-1.30 | SAA+Met | |
| Kahindi et al. [79] | 1 | 0.48-0.71 | 0.22-0.46 | 1.18 | SAA+Met | E. coli infection |
| Zong et al. [10] | 1 | 0.39-0.71 | 0.20-0.52 | 1.35 | SAA+Met | |
| Capozzalo et al. [80] | 1-2 | 0.45-0.78 | 0.21-0.53 | 1.10-1.20 | SAA+Met | E. coli infection |
| Capozzalo et al. [81] | 1 | 0.55-0.66 | 0.28-0.39 | 1.25 | SAA | E. coli infection |
| Kahindi et al. [82] | 1 | 0.52-0.68 | 0.24-0.40 | 1.18 | SAA+Met | Unclean condition |
| Kaewtapee et al. [83] | 2-3 | 0.49-0.69 | ND3 | 1.05 | SAA | |
| Zhang et al. [84] | 3 | 0.50-0.70 | 0.27-0.47 | 0.90 | SAA+Met | |
| Chen et al. [85] | 1 | 0.39-0.48 | 0.20-0.29 | 1.20 | SAA+Met | |
| Conde-Aguilera et al. [86] | 2-3 | 0.50-0.60 | 0.24-0.34 | 1.06-1.09 | SAA+Met | |
| Conde-Aguilera et al. [87] | 2 | 0.38-0.60 | 0.17-0.40 | 1.16 | SAA+Met | |
| Frantz et al. [88] | 3 | 0.47-0.63 | ND3 | 1.05 | SAA |
1Phase1 (BW < 11), Phase2 (11 ≤ BW < 25), Phase3 (25 ≤ BW< 50). 2SAA, supplementation with sulfur amino acids; SAA+MET, supplementation with methionine without concurrent increase in cysteine. 3No description.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



