
Submitted:
05 September 2024
Posted:
09 September 2024
You are already at the latest version
Abstract
Estrogen exerts its action via estrogen receptors (ERs), including ERα and ERβ, and has effects on immunomodulation during pregnancy. It is known that there is a remodeling in maternal immunology during pregnancy. However, it is not clear if early pregnancy has effects on the expression of ERα and ERβ in the maternal thymus, lymph nodes, spleen, and liver. In this study, these maternal immune organs were harvested at day 16 of the estrous cycle, and at days 13, 16, and 25 of pregnancy. The mRNA and protein expression of ERα and ERβ was analyzed. The results revealed that mRNA and protein expression of both ERα and ERβ was upregulated in the maternal spleen, and expression of ERα and ERβ in the thymus, lymph nodes, and liver was modulated during early pregnancy. In conclusion, early pregnancy modulates the expression of ERα and ERβ in the maternal thymus, lymph nodes, spleen, and liver in a tissue-specific manner, which is related to the regulation of maternal immune function during early pregnancy.
Keywords:
estrogen receptor
; immune organ
; pregnancy
; sheep
1. Introduction
Estrogen not only plays an essential role in the luteinizing hormone peak during the periconceptional period, but is also involved in placentation through remodeling the function of immune cells and the expression of angiogenic factors in the uterus during early pregnancy [1]. Estrogen mediates the process of implantation through regulating the expression of estrogen-modulated paracrine factors in the uterus, and these factors can promote cell growth, or amplify estrogenic effects during early pregnancy [2]. Estrogen can induce vasodilator production via estrogen receptors (ERs), including ERα and ERβ, in the uterine artery, which is necessary in providing nutrient support for fetal growth and survival during pregnancy in humans [3]. The expression of ERα and ERβ in the endometrium is necessary for normal pregnancy, and ERα is related to uterine cell proliferation, but ERβ has a negative effect on ERα action [4]. A high level of estrogen enhances the proportions of CD4+ CD25+ Treg in peripheral blood, which is essential for estrogen-mediated immunomodulation during pregnancy in humans [5].
There is a dynamic adaptation in the maternal immune system during pregnancy, which contributes to a tolerance towards the allogeneic fetus, but not a broad maternal immune inhibition [6]. Estrogens secreted from the ovary and placenta can modulate the immune system development and immune responses via its receptors expressed in the immune cells, which is involved in the adaptation of the organism to the intrinsic physiological changes in females [7]. Estrogen can suppress autophagy of the immune system via ERα and ERβ, which plays a key role in pre-implantation and maintaining embryo survival [8]. In addition, estrogens have effects on innate and adaptive immunity, which modulate the activity of different immune cells in the thymus, spleen, and lymph node through ERα and ERβ [9].
In ruminants, conceptus signal (interferon-tau, IFNT) and high levels of progesterone modulate the maternal innate immune system, which changes immune function both locally and systemically during pregnancy [10]. IFNT has paracrine and endocrine actions in modulating maternal innate immune functions, which is essential for avoiding conceptus rejection by the mother during early pregnancy in ruminants [11]. Our previous studies report that IFNT has effects on bone marrow, thymus, spleen, lymph nodes, and liver through an endocrine manner during early pregnancy in ewes. In sheep, early pregnancy has effects on the expression of melatonin receptors, gonadotropin-releasing hormone and its receptor, and prolactin and its receptor in the ovine thymus, lymph node, spleen, and liver [12], which modulates the function of the maternal immune system via endocrine and/or paracrine/autocrine pathways in a tissue-specific manner. However, it is not clear if early pregnancy has effects on the expression of ERα and ERβ in the maternal thymus, lymph node, spleen, and liver. The aim of the present study was to investigate the expression of ERα and ERβ in these organs from nonpregnant ewes and early pregnant ewes, which will contribute to understanding pregnancy immune tolerance.
2. Materials and Methods
2.1. Material and Methods
The experiments were performed as described previously [12]. Ewes were checked twice daily, and animals that manifested a mating mark (day 0) were assigned to collect thymus, lymph node, spleen, and liver on days 13, 16, and 25 of gestation (P13, P16, and P25), and on day 16 of the estrous cycle (N16). Pregnancy was confirmed by finding a conceptus in the uterus after the ewes were killed.
2.2. RNA Extraction and RT-qPCR Assay
Total RNA was extracted from these samples using TRNzol Universal Reagent (DP424; Tiangen Biotech Co., Ltd., Beijing, China), and RT-qPCR was performed as described previously [12]. Primers (Table 1) of ovine ERα and ERβ genes were designed and synthesized by Shanghai Sangon Biotech Co., Ltd. (Shanghai, China). The 2-ΔΔCt analysis method was applied to analyze the relative levels [13].
2.3. Western Blot Analysis
Western blot analysis was performed as described previously [12], and a rat anti-ERα monoclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA, sc-53490, 1:1000), or mouse anti-ERβ antibody (Abcam, Cambridge, UK, ab187291, 1:1000) was employed to analyze the expression of ERα and ERβ proteins in these tissues in a 1:1000 dilution. The primary antibodies were validated and considered to be species cross-reactive by specific binding to native ovine proteins.
2.4. Immunohistochemistry Analysis
2.5. Statistical Analysis
Statistical analysis was performed using the Proc Mixed models of SAS (Version 9.4; SAS Institute, Cary, NC). The normality of the distribution of data was assessed, and the data of ERα and ERβ mRNA and proteins were in a normal distribution. ANOVA with a post-hoc Tukey test was performed. P-values less than 0.05 were considered significant.
3. Results
3.1. ERα and ERβ in the Thymus
Figure 1 indicated that the expression values of ERα mRNA and protein peaked at P13 (P < 0.05) in the maternal thymus, but declined at P25 compared to N16, P13, and P16. Furthermore, the expression values of ERβ mRNA and protein were the highest at P13 among the four groups (P < 0.05), and ERβ protein was almost undetected at N16 and P25. In addition, the ERα protein was located in the epithelial reticular cells, capillaries, and thymic corpuscles. The staining intensities for ERα protein were 1, 3, 2, and 0.5 for N16, P13, P16 and P25, respectively. The staining intensity was as follows: 0 = negative; 1 = weak; 2 = strong; 3 = stronger.
3.2. ERα and ERβ in Lymph Nodes
ERα mRNA and protein values were higher at P13 and P16 than at N16 and P25 (P < 0.05; Figure 2), and the ERα mRNA and protein values were the lowest at P25 among the four groups (P < 0.05). In addition, ERβ mRNA and protein peaked at P16 (P < 0.05), and ERβ protein was almost undetected at N16, P13, and P25. The ERα protein was located in the subcapsular sinus and lymph sinus, and the staining intensities for ERα protein were 1, 3, 3, and 0 for N16, P13, P16, and P25, respectively (Figure 2C).
3.3. ERα and ERβ in the Spleen
There were increases in the expression of ERα mRNA and protein at P16 and P25 compared to N16 and P13 (Figure 3; P < 0.05) in the spleen, and ERα protein was almost undetected at N16 and P13. ERβ mRNA and protein peaked at P25 (P < 0.05), and ERβ protein was almost undetected at N16, P13, and P16. In addition, ERα protein was located in the capsule, trabeculae, and splenic cords, and the staining intensities for ERα protein were 0, 0, 1, and 2 for N16, P13, P16, and P25, respectively (Figure 3C).
3.4. ERα and ERβ in the Liver
The mRNA and protein levels of ERα were decreased at P13 and P16 compared with N16 and P25 (P < 0.05), and the level of ERα at N16 was the highest among the four groups (P < 0.05). In addition, expression of ERβ mRNA and protein was inhibited during early gestation (P < 0.05), but there was an increase in expression of ERβ mRNA and protein at P16 and P25 compared to P13 (Figure 4). ERα protein was located in the hepatocyte, endothelial cells of the proper hepatic arteries, and hepatic portal veins (Figure 4C), and the staining intensities for ERα protein were 3, 0, 0, and 1 for N16, P13, P16, and P25, respectively.
4. Discussion
The thymus is responsible for the generation of a T-cell receptor repertoire, which responds to foreign antigens, provides surveillance, and remains tolerant to self [14]. Pregnancy induces a severe reduction in thymus size and thymocyte output, which is associated with the suppression of chemokines in the thymic nonlymphoid cells during pregnancy [15]. Our previous studies report that the immune signaling pathways, including complement pathway, nod-like receptor pathway, nod-like receptor pathway, and toll-like receptor pathway, as well as kappa B subunits and IkappaB protein, are changed in the maternal thymus [16], which is involved in the maternal immune regulation in sheep. Estrogens regulate the expression of Aire via ERs, which modulates the expression of tissue-restricted antigen genes to generate self-peptides to bind to major histocompatibility complex molecules in the medullary thymic epithelial cells (TECs) [17]. Estrogen also can modulate the development and differentiation of T-cells and the immune functions of the TECs, which are involved in autoimmunity via ERs [18].
During the proestrus phase in the mouse thymus, a high level of estradiol decreases the percentage of CD4+CD8+ double-positive T cells via ERα, but in the proestrus phase, CD4+CD8- or CD4-CD8+ single positive T cells are significantly enhanced [19]. In addition, during the first half of pregnancy, estriol is implicated in regulating the processes of myeloid dendritic cell maturation in the thymus and is also related to steroid-induced involution of the thymus [20]. There is an upregulation of estrogen levels during pregnancy, which suppresses thymocyte proliferation and leads to thymic involution during pregnancy in mice [21]. Our data revealed that mRNA and protein levels of ERα and ERβ upregulated at P13, but there was a downregulation of ERα and ERβ levels at P25. Thus, it is supposed that estrogens have effects on thymic function via ERα and ERβ in an endocrine manner during early pregnancy in sheep.
The lymph node consists of antigen-presenting cells and antigen-responsive cells, which are involved in the recruitment of naïve lymphocytes and antigen-presenting cells, the generation of adaptive immune responses, and the suppression of autoreactive cells [22]. The presence of an embryo changes the function of the lymph nodes around the reproductive tract, which is implicated in regulating immune responses in the maternal local immune tissues during early pregnancy in the ovine [23]. It has been reported that prostaglandin synthases, T helper cell cytokines, nod-like receptors, toll-like receptors, complement components, as well as nuclear factor kappa B family are modulated in ovine lymph nodes [24], which is related to the maternal immune regulation in ewes. ERα is expressed by the follicular dendritic cells in lymph nodes, which is related to breast cancer and inflammation in humans [25]. Our data manifested that the expression level of ERα peaked at P13 and P16, and the level of ERβ peaked only at P16. However, the expression levels of ERα and ERβ downregulated at P25. Therefore, ERα and ERβ are involved in the immunoregulation of maternal lymph nodes during early pregnancy, which is in an endocrine manner in ewes.
The spleen comprises the red and white pulp (WP), and the WP is the primary immunologic region, and the red pulp (RP) has an immune function distinct from the WP. The marginal zone (MZ) is between the WP and RP, and there are many macrophages in MZ [26]. Estrogen has effects on the expression of some cytokines in the spleen dendritic cells from the female mice, which is through ERα [27]. 17β-estradiol can regulate the function of mice spleen B cells, which is via toll-like receptor 9 [28]. In addition, 17β-estradiol has beneficial effects on the function of splenic CD4+ T lymphocytes, which is mediated by ERα, but not ERβ, and related to the suppression of endoplasmic reticulum stress [29]. Early pregnancy increases the level of ERα and the percentage of mature macrophages in the spleen of female mice, which plays a cardinal role in regulating immune phenomena during pregnancy [30]. It has been reported that early pregnancy has effects on the expression of T helper cell cytokines, toll-like receptors, and nuclear factor kappa B family in the ovine spleen [24,31], which is implicated in the maternal immune adaptation. Our results revealed that early pregnancy enhanced the expression of ERα and ERβ. Therefore, the upregulation of ERα and ERβ is implicated in modulating maternal splenic function during early pregnancy through an endocrine manner.
As the largest gland of the body, the liver plays cardinal roles in immunologic responses and nutrient metabolism, and there are necessary hepatic adaptations in immunology and nutrition during normal pregnancy [32]. ERα mRNA level in the liver is enhanced from day 7 to 21 of pregnancy, and the expression level of ERβ is variable during pregnancy, which is related to hepatic synthetic functions in rats [33]. Estrogen can change the cytokine/chemokine repertoire in the liver via ERs, which are involved in modulating the liver immunosuppressive microenvironment [34]. 17β-estradiol supplementation can improve mitochondrial function to prevent nonalcoholic fatty liver disease via the ERα pathway in bilateral oophorectomy female rats [35]. The previous reports show that early pregnancy modulates the expression of Th cytokines, toll-like receptor pathway, nuclear factor kappa B signaling, complement components, nod-like receptors, and CD4, which is associated with the hepatic local immunosuppressive milieu in sheep [32]. Our results showed that early pregnancy suppressed the expression of ERα and ERβ, but there were increases in expression levels of ERα and ERβ from P13 to P25. Therefore, the changes in the expression of hepatic ERα and ERβ are involved in regulating maternal hepatic function, including the immune function, during early pregnancy.
It has been reported that pregnancy has effects on estrogen levels [21], which may modulate the expression of ERα and ERβ in the maternal thymus, lymph nodes, spleen, and liver via an endocrine manner. Therefore, estrogen exerts its actions on these maternal immune organs via ERα and ERβ in a tissue-specific manner to regulate maternal immune function during early pregnancy (Figure 5).
5. Conclusions
It is reported for the first time that early pregnancy has tissue-specific effects on the expression of ERα and ERβ in the maternal thymus, lymph nodes, spleen, and liver, which regulates the function of main immune organs via an endocrine manner. Therefore, it is suggested that early pregnancy modulates the expression of ERα and ERβ in these immune organs, which is related to the adaptations in the maternal immune system during early pregnancy in ewes.
Supplementary Materials
Author Contributions
Conceptualization, L.Y.; methodology, Z.Y., Y.Z., Z.C.; validation, Z.L.; investigation, Z.Y., Y.Z., Z.C.; data curation, Z.L.; writing—original draft preparation, L.Y.; writing—review and editing, L.Z.; visualization, L.Z.; supervision, L.Y.; project administration, L.Z.; funding acquisition, L.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the grants from Natural Science Foundation of Hebei Province, China (grant number: C2024402023).
Institutional Review Board Statement
All experiments were approved by the Hebei University of Engineering Animal Care and Use Committee (approval no. 2019-017), and performed following the Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting the findings of this study are available within the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Parisi, F.; Fenizia, C.; Introini, A.; Zavatta, A.; Scaccabarozzi, C.; Biasin, M.; Savasi, V. The pathophysiological role of estrogens in the initial stages of pregnancy: molecular mechanisms and clinical implications for pregnancy outcome from the periconceptional period to end of the first trimester. Hum. Reprod. Update. 2023, 29, 699–720. [Google Scholar] [CrossRef] [PubMed]
- Robertshaw, I.; Bian, F.; Das, S.K. Mechanisms of uterine estrogen signaling during early pregnancy in mice: an update. J. Mol. Endocrinol. 2016, 56, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Qi, Q.R.; Li, Y.; Day, R.; Makhoul, J.; Magness, R.R.; Chen, D.B. Estrogen receptors and estrogen-induced uterine vasodilation in pregnancy. Int. J. Mol. Sci. 2020, 21, 4349. [Google Scholar] [CrossRef]
- Yu, K.; Huang, Z.Y.; Xu, X.L.; Li, J.; Fu, X.W.; Deng, S.L. Estrogen receptor function: impact on the human endometrium. Front. Endocrinol. 2022, 13, 827724. [Google Scholar] [CrossRef]
- Xiong, Y.H.; Yuan, Z.; He, L. Effects of estrogen on CD4(+) CD25(+) regulatory T cell in peripheral blood during pregnancy. Asian. Pac. J. Trop. Med. 2013, 6, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Abu-Raya, B.; Michalski, C.; Sadarangani, M.; Lavoie, P.M. Maternal immunological adaptation during normal pregnancy. Front. Immunol. 2020, 11, 575197. [Google Scholar] [CrossRef]
- Quatrini, L.; Ricci, B.; Ciancaglini, C.; Tumino, N.; Moretta, L. Regulation of the immune system development by glucocorticoids and sex hormones. Front. Immunol. 2021, 12, 672853. [Google Scholar] [CrossRef]
- Komijani, E.; Parhizkar, F.; Abdolmohammadi-Vahid, S.; Ahmadi, H.; Nouri, N.; Yousefi, M.; Aghebati-Maleki, L. Autophagy-mediated immune system regulation in reproductive system and pregnancy-associated complications. J. Reprod. Immunol. 2023, 158, 103973. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, B.; Byemerwa, J.; Krebs, T.; Lim, F.; Chang, C.Y.; McDonnell, D.P. Estrogen receptor signaling in the immune system. Endocr. Rev. 2023, 44, 117–141. [Google Scholar] [CrossRef]
- Ott, T.L. Immunological detection of pregnancy: Evidence for systemic immune modulation during early pregnancy in ruminants. Theriogenology. 2020, 150, 498–503. [Google Scholar] [CrossRef]
- Rocha, C.C.; Da-Silveira, J.C.; Forde, N.; Binelli, M.; Pugliesi, G. Conceptus-modulated innate immune function during early pregnancy in ruminants: a review. Anim. Reprod. 2021, 18, e20200048. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Wu, J.; Ren, Y.; Zhang, L.; Cao, J.; Yang, L. Early pregnancy regulates the expression of prolactin and its receptor in the thymus, the liver, the spleen and lymph nodes in sheep. Domest. Anim. Endocrinol. 2022, 81, 106731. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dinges, S.S.; Amini, K.; Notarangelo, L.D.; Delmonte, O.M. Primary and secondary defects of the thymus. Immunol. Rev. 2024, 322, 178–211. [Google Scholar] [CrossRef]
- Laan, M.; Haljasorg, U.; Kisand, K.; Salumets, A.; Peterson, P. Pregnancy-induced thymic involution is associated with suppression of chemokines essential for T-lymphoid progenitor homing. Eur. J. Immunol. 2016, 46, 2008–2017. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Yang, Z.; Quan, Y.; Zhao, S.; Zhang, L.; Yang, L. Regulation of IkappaB protein expression by early gestation in the thymus of ewes. Vet. Sci. 2023, 10, 462. [Google Scholar] [CrossRef] [PubMed]
- Taves, M.D.; Ashwell, J.D. Effects of sex steroids on thymic epithelium and thymocyte development. Front. Immunol. 2022, 13, 975858. [Google Scholar] [CrossRef]
- Merrheim, J.; Villegas, J.; Van-Wassenhove, J.; Khansa, R.; Berrih-Aknin, S.; Le-Panse, R.; Dragin, N. Estrogen, estrogen-like molecules and autoimmune diseases. Autoimmun. Rev. 2020, 19, 102468. [Google Scholar] [CrossRef]
- Lee, H.; Kim, H.; Chung, Y.; Kim, J.; Yang, H. Thymocyte differentiation is regulated by a change in estradiol levels during the estrous cycle in mouse. Dev. Reprod. 2013, 17, 441–449. [Google Scholar] [CrossRef]
- Shirshev, S.V.; Orlova, E.G.; Loginova, O.A.; Nekrasova, I.V.; Gorbunova, O.L.; Maslennikova, I.L. Hormonal regulation of dendritic cell differentiation in the thymus. Bull. Exp. Biol. Med. 2018, 165, 230–234. [Google Scholar] [CrossRef]
- Zoller, A.L.; Schnell, F.J.; Kersh, G.J. Murine pregnancy leads to reduced proliferation of maternal thymocytes and decreased thymic emigration. Immunology. 2007, 121, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.M.; Lou, M.; Yao, L.; Germain, R.N.; Radtke, A.J. The lymph node at a glance - how spatial organization optimizes the immune response. J. Cell. Sci. 2020, 133, jcs241828. [Google Scholar] [CrossRef] [PubMed]
- Quirke, L.D.; Maclean, P.H.; Haack, N.A.; Edwards, S.J.; Heiser, A.; Juengel, J.L. Characterization of local and peripheral immune system in pregnant and nonpregnant ewes. J. Anim. Sci. 2021, 99, skab208. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Cai, C.; Bai, Y.; Zhang, L.; Yang, L. Early pregnancy regulates expression of IkappaB family in ovine spleen and lymph nodes. Int. J. Mol. Sci. 2023, 24, 5156. [Google Scholar] [CrossRef]
- Sapino, A.; Cassoni, P.; Ferrero, E.; Bongiovanni, M.; Righi, L.; Fortunati, N.; Crafa, P.; Chiarle, R.; Bussolati, G. Estrogen receptor alpha is a novel marker expressed by follicular dendritic cells in lymph nodes and tumor-associated lymphoid infiltrates. Am. J. Pathol. 2003, 163, 1313–1320. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef]
- Jiang, B.; Sun, L.; Hao, S.; Li, X.; Hou, Y. Estrogen distinctively modulates spleen DC from (NZB x NZW) F1 female mice in various disease development stages. Cell. Immunol. 2007, 248, 95–102. [Google Scholar] [CrossRef]
- Xu, Y.; Fan, H.; Li, X.; Sun, L.; Hou, Y. 17β-Estradiol enhances response of mice spleen B cells elicited by TLR9 agonist. Cell. Immunol. 2012, 278, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Jiang, L.N.; Wang, C.; Li, Y.; Yin, M.; Du, H.B.; Zhang, H.; Fan, Z.H.; Liu, Y.X.; Zhao, M.; Kang, A.L.; Feng, D.Y.; Li, S.G.; Niu, C.Y.; Zhao, Z.G. Estradiol-induced inhibition of endoplasmic reticulum stress normalizes splenic CD4 + T lymphocytes following hemorrhagic shock. Sci. Rep. 2021, 11, 7508. [Google Scholar] [CrossRef]
- Piesta, A.; Maj, T.; Chełmońska-Soyta, A. The influence of mating on estrogen receptor alpha protein level in spleen and uterine macrophages in female mice. Reprod. Biol. 2009, 9, 225–240. [Google Scholar] [CrossRef]
- Hao, S.; Fang, H.; Fang, S.; Zhang, T.; Zhang, L.; Yang, L. Changes in nuclear factor kappa B components expression in the ovine spleen during early pregnancy. J. Anim. Feed. Sci. 2022, 31, 3–11. [Google Scholar] [CrossRef]
- Yang, L.; Meng, Y.; Shi, Y.; Fang, H.; Zhang, L. Maternal hepatic immunology during pregnancy. Front. Immunol. 2023, 14, 1220323. [Google Scholar] [CrossRef] [PubMed]
- Varas, S.M.; Jahn, G.A. The expression of estrogen, prolactin, and progesterone receptors in mammary gland and liver of female rats during pregnancy and early postpartum: regulation by thyroid hormones. Endocr. Res. 2005, 31, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Benslimane, Y.; Amalfi, K.; Lapin, S.; Perrino, S.; Brodt, P. Estrogen receptor blockade potentiates immunotherapy for liver metastases by altering the liver immunosuppressive microenvironment. Cancer. Res. Commun. 2024, 4, 1963–1977. [Google Scholar] [CrossRef]
- Tian, Y.; Hong, X.; Xie, Y.; Guo, Z.; Yu, Q. 17β-Estradiol (E2) Upregulates the ERα/SIRT1/PGC-1α signaling pathway and protects mitochondrial function to prevent bilateral oophorectomy (OVX)-induced nonalcoholic fatty liver disease (NAFLD). Antioxidants. 2023, 12, 2100. [Google Scholar] [CrossRef]
Figure 1.
ERα and ERβ in the thymus. A: Expression values of ERα and ERβ mRNA in the thymus. B: Expression of ERα and ERβ proteins in the thymus. C: Representative immunohistochemical localization of the ERα protein in the thymus. The thymus is divided into the cortex (CO) and the medulla (ME). Note: HE = stained by hematoxylin and eosin; Clt = negative control; T = thymocyte; ER = epithelial reticular cell; CA = capillary; TC = thymic corpuscle; Bar = 20 µm. Different letters within columns indicate significant differences (P < 0.05).
Figure 1.
ERα and ERβ in the thymus. A: Expression values of ERα and ERβ mRNA in the thymus. B: Expression of ERα and ERβ proteins in the thymus. C: Representative immunohistochemical localization of the ERα protein in the thymus. The thymus is divided into the cortex (CO) and the medulla (ME). Note: HE = stained by hematoxylin and eosin; Clt = negative control; T = thymocyte; ER = epithelial reticular cell; CA = capillary; TC = thymic corpuscle; Bar = 20 µm. Different letters within columns indicate significant differences (P < 0.05).
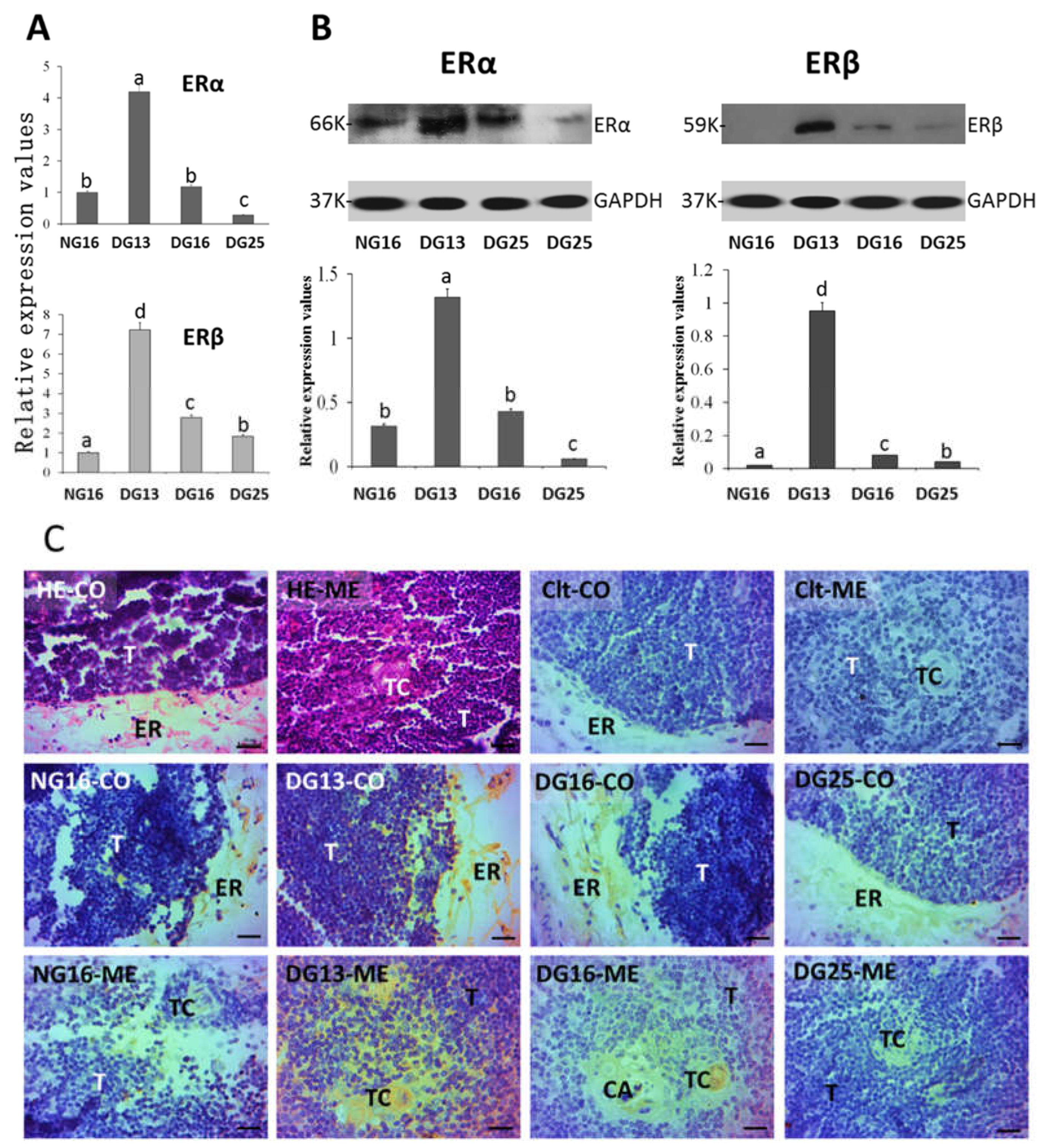
Figure 2.
ERα and ERβ in lymph nodes. A: Expression levels of ERα and ERβ mRNA in the lymph node. B: Expression of ERα and ERβ proteins in the lymph node. C: Representative immunohistochemical localization of the ERα protein in the lymph node. A lymph node is divided into an outer cortex (CO) and an inner medulla (ME). Lymph enters the convex through the subcapsular sinus (SS) and trabeculae (TR) around the lymphoid nodules (LN) and flows into the medulla through the lymph sinus (LS) around the medullary cord (MC). Note: Bar = 20 µm. Different letters within columns indicate significant differences (P < 0.05).
Figure 2.
ERα and ERβ in lymph nodes. A: Expression levels of ERα and ERβ mRNA in the lymph node. B: Expression of ERα and ERβ proteins in the lymph node. C: Representative immunohistochemical localization of the ERα protein in the lymph node. A lymph node is divided into an outer cortex (CO) and an inner medulla (ME). Lymph enters the convex through the subcapsular sinus (SS) and trabeculae (TR) around the lymphoid nodules (LN) and flows into the medulla through the lymph sinus (LS) around the medullary cord (MC). Note: Bar = 20 µm. Different letters within columns indicate significant differences (P < 0.05).
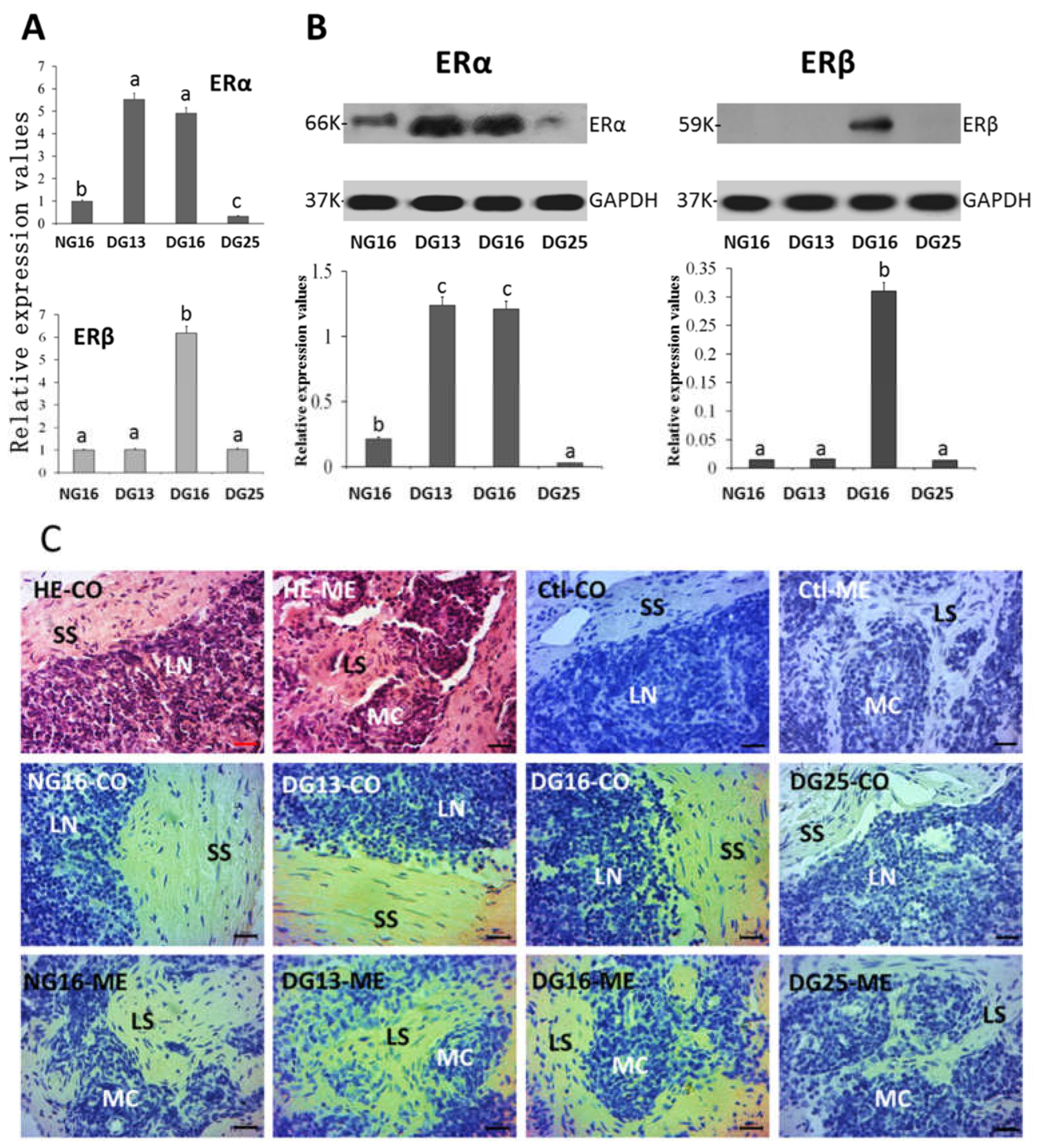
Figure 3.
ERα and ERβ in the spleen. A: Expression values of ERα and ERβ mRNA in the spleen. B: Expression of ERα and ERβ proteins in the spleen. C: Representative immunohistochemical localization of the ERα protein in the spleen. Spleen is divided into red pulp (R) and white pulp (W), and surrounded by a thickened capsule (CP). A capsule (CP) with several trabeculae (TR) projects into the substance of the spleen. Note: HE = stained by hematoxylin and eosin; Clt = negative control; SS = splenic sinuses; SC = splenic cords; MZ = marginal zone; LN = lymphoid nodule; CA = central arteriole; Bar = 50 µm. Different letters within columns indicate significant differences (P < 0.05).
Figure 3.
ERα and ERβ in the spleen. A: Expression values of ERα and ERβ mRNA in the spleen. B: Expression of ERα and ERβ proteins in the spleen. C: Representative immunohistochemical localization of the ERα protein in the spleen. Spleen is divided into red pulp (R) and white pulp (W), and surrounded by a thickened capsule (CP). A capsule (CP) with several trabeculae (TR) projects into the substance of the spleen. Note: HE = stained by hematoxylin and eosin; Clt = negative control; SS = splenic sinuses; SC = splenic cords; MZ = marginal zone; LN = lymphoid nodule; CA = central arteriole; Bar = 50 µm. Different letters within columns indicate significant differences (P < 0.05).
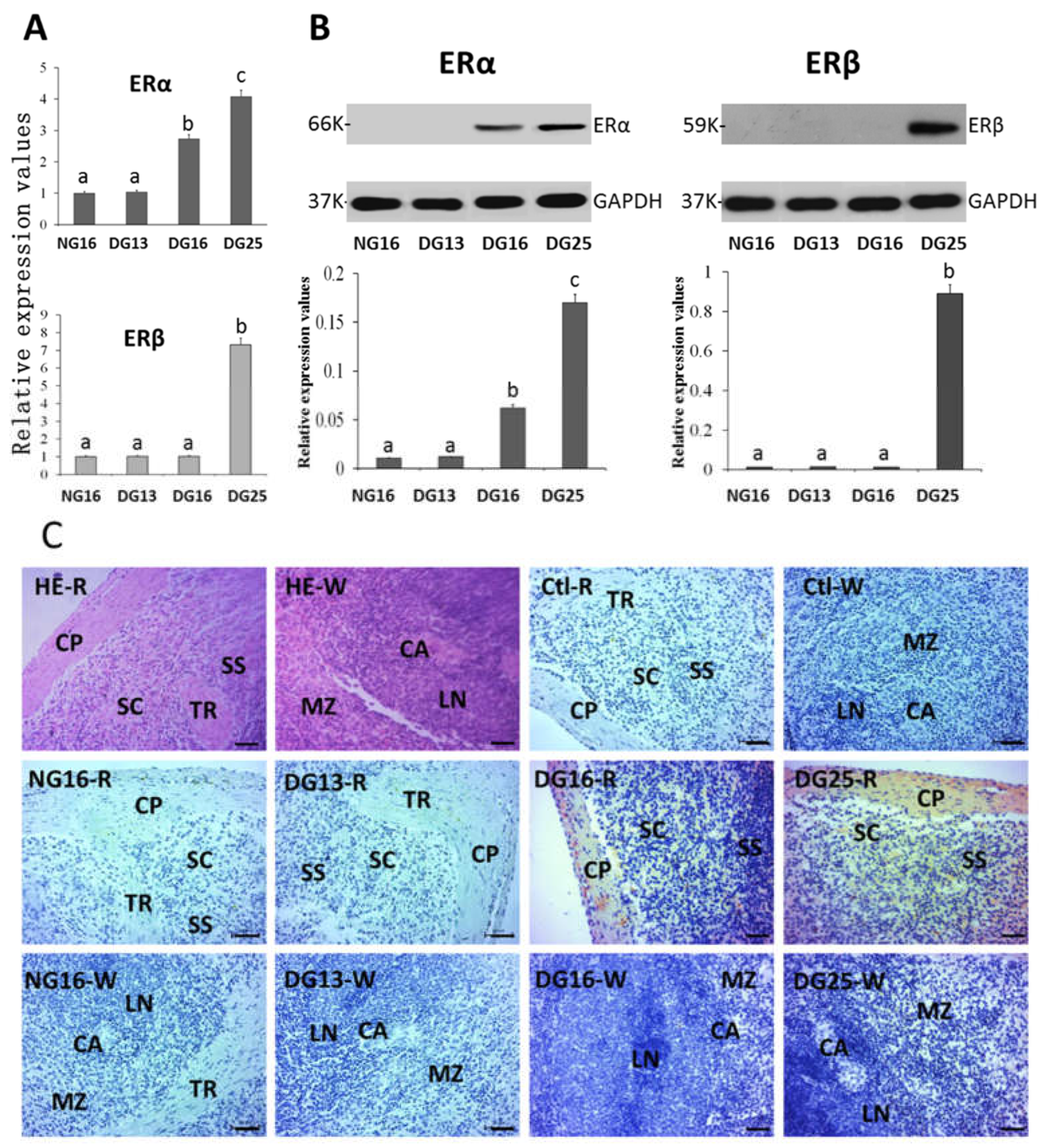
Figure 4.
ERα and ERβ in the liver. A: Expression values of ERα and ERβ mRNA in the liver. B: Expression of ERα and ERβ proteins in the liver. C: Representative immunohistochemical localization of the ERα protein in the liver. A portal triad is a component of the hepatic lobule, and consists of proper hepatic artery (HA), hepatic portal vein (PV), and small bile ductile (BD). Note: HE = stained by hematoxylin and eosin; H = hepatocyte; Bar = 50 µm. Different letters within columns indicate significant differences (P < 0.05).
Figure 4.
ERα and ERβ in the liver. A: Expression values of ERα and ERβ mRNA in the liver. B: Expression of ERα and ERβ proteins in the liver. C: Representative immunohistochemical localization of the ERα protein in the liver. A portal triad is a component of the hepatic lobule, and consists of proper hepatic artery (HA), hepatic portal vein (PV), and small bile ductile (BD). Note: HE = stained by hematoxylin and eosin; H = hepatocyte; Bar = 50 µm. Different letters within columns indicate significant differences (P < 0.05).
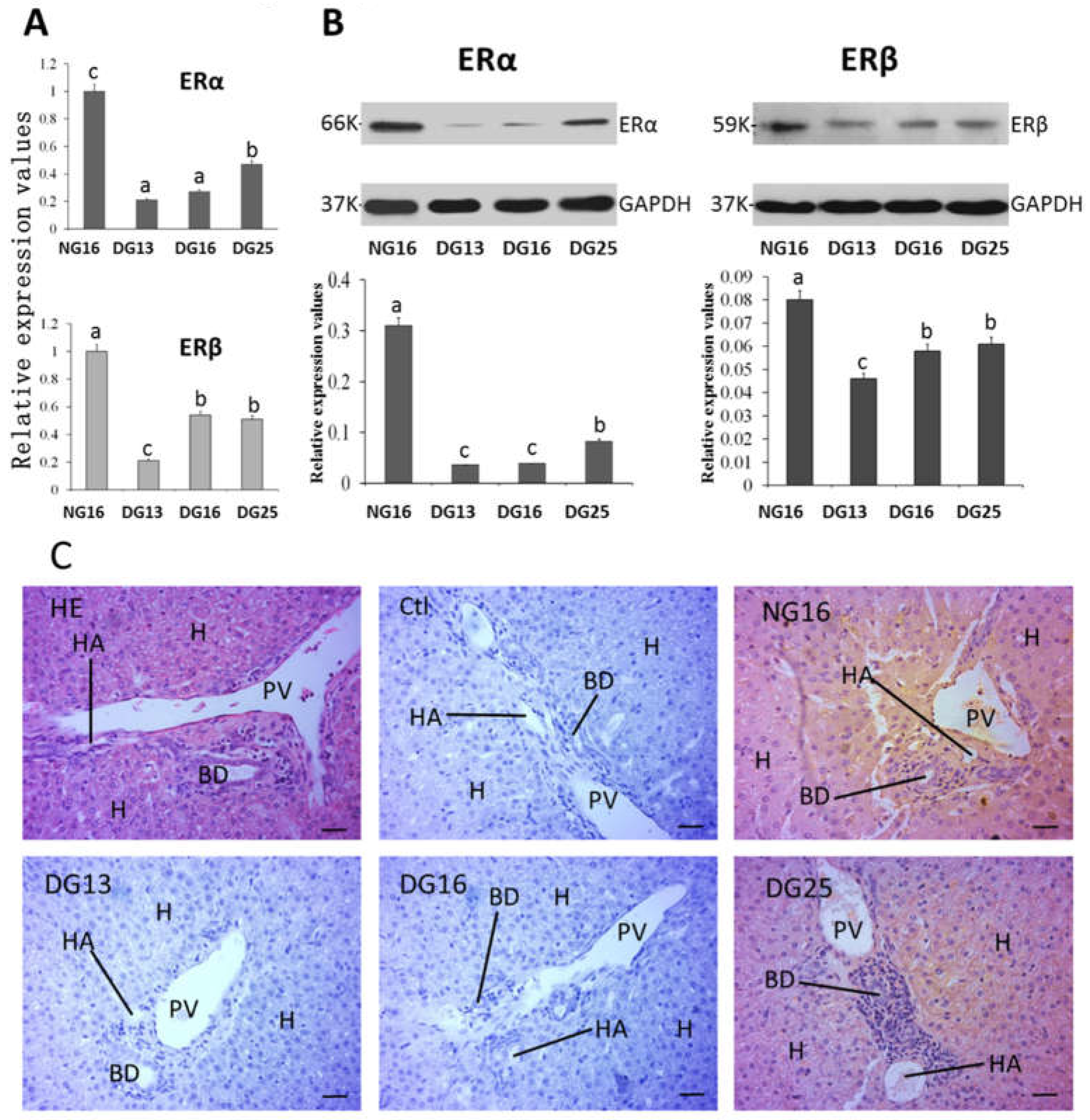
Figure 5.
Sketch of expression of estrogen receptors (ERs) in the ovine thymus, lymph nodes, spleen, and liver during early pregnancy. Early pregnancy signal (IFNT) exerts effects on the ovary, which modulates the secretion of estradiol (E2), leads to the changes in the expression of ERα and ERβ in the maternal thymus, lymph nodes, spleen, and liver, and contributes to the adaptations in the maternal immune system.
Figure 5.
Sketch of expression of estrogen receptors (ERs) in the ovine thymus, lymph nodes, spleen, and liver during early pregnancy. Early pregnancy signal (IFNT) exerts effects on the ovary, which modulates the secretion of estradiol (E2), leads to the changes in the expression of ERα and ERβ in the maternal thymus, lymph nodes, spleen, and liver, and contributes to the adaptations in the maternal immune system.
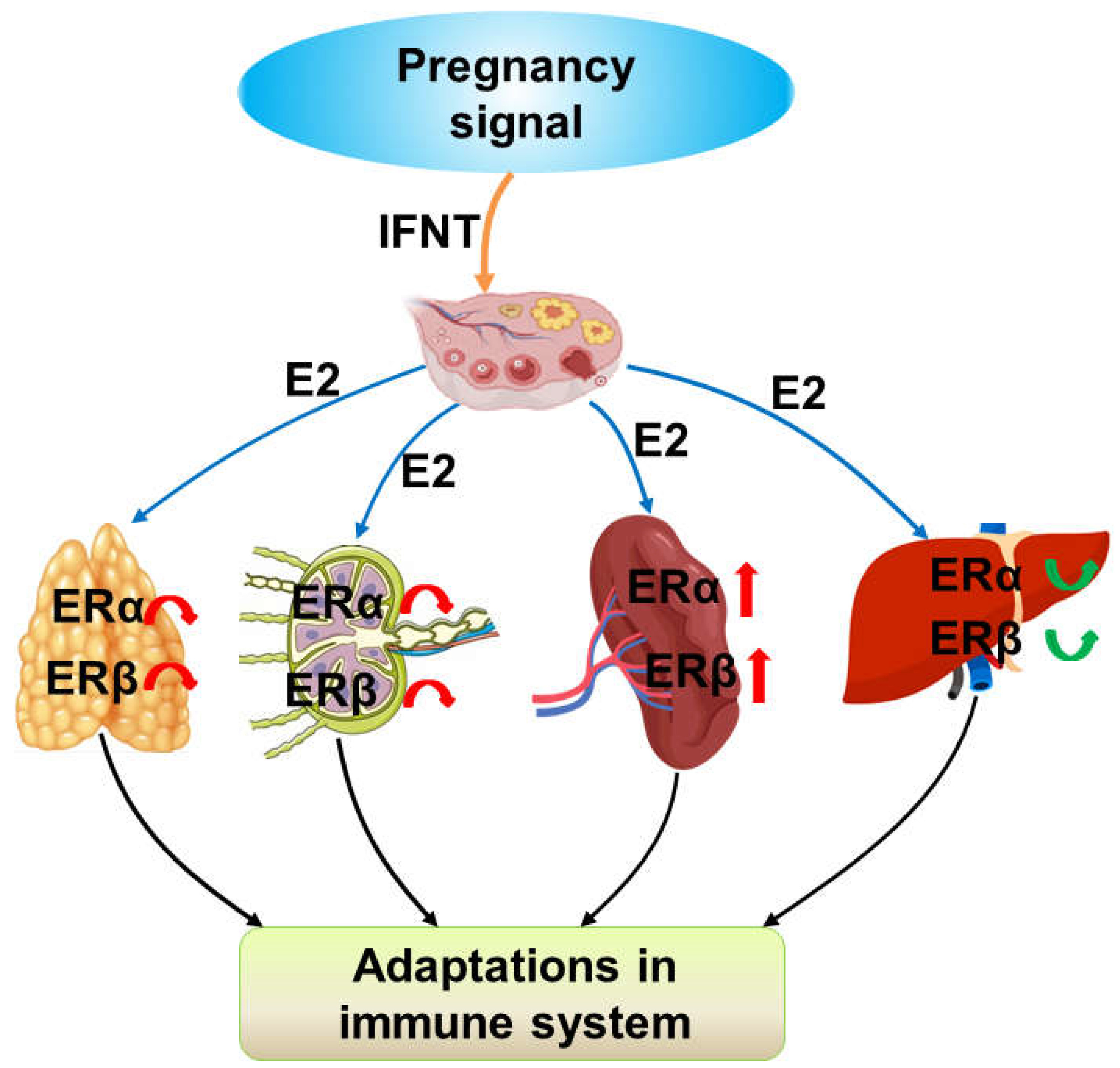

Table 1.
Primers used for RT-qPCR.
| Gene | Primer | Sequence | Size (bp) | Accession numbers |
|---|---|---|---|---|
| ERα | Forward | CTGCTGCTGGAGATGCTGGATG | 88 | XM_042253635.1 |
| Reverse | GCTGGCTCTGATTCACGTCTTCC | |||
| ERβ | Forward | TGCTGCTGGAGATGCTGAATGC | 112 | NM_001009737.1 |
| Reverse | GGTTTCTGGGAGCCCTCTTTGC | |||
| GAPDH | Forward | GGGTCATCATCTCTGCACCT | 176 | NM_001190390.1 |
| Reverse | GGTCATAAGTCCCTCCACGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



