Submitted:
04 September 2024
Posted:
09 September 2024
You are already at the latest version
Abstract
This study aimed to explore the genetic information about the yield traits in perennial Chinese rice "Shendao3#" and provide a series of theoretical bases for the innovative utilization of perennial Chinese rice germplasm. The F2 population derived from the cross between perennial Chinese rice "Shendao3#" and one annual rice "XiqingzaoB" was developed to identify QTLs’ underlying yield-related traits. Fifteen yield traits including plant height, panicle plant-1, and thousand-grain weight were measured for the QTL mapping. The software packages Excel 2010, GraphpadPrism 5.0, and QTLIciMapping 4.10 were used to perform the phenotypic data analysis, construction of molecular map, and QTL mapping, respectively. Results indicated 15 yield-related traits displaying normal distribution and belonging to quantitative trait inheritance. A total of 25 QTLs associated with the yield-related traits and 26 epistatic QTLs were detected on the whole genome except for the 6th and 12th chromosomes. This finding explained phenotypic variance with ranging from 4.21% to 27.30% and 1.15% to 19.30%, respectively. Six pleiotropic QTL (chromosomal regions) underlying two or more yield-related traits were detected on the 1st , 4th, 7th, 8th, and 11th chromosomes, respectively. Among 25 QTLs underlying a single yield-related trait, 11 were primarily identified in the perennial Chinese rice "Shendao3#". All QTLs underlying 15 yield-related traits followed the genetic model of additive, dominant, and digenic epistatic of additive by additive, additive by dominant, dominant by additive, and dominant by dominant. This work can serve as a good foundation for yield-related gene mining and its utilization in breeding the perennial Chinese rice "Shendao3#".
Keywords:
perennial Chinese rice"Shendao3#"
; QTLs for yield-related trait
; pleiotropic QTLs
1. Introduction
The eternal goal of rice geneticists and breeders is to pursue superior yield-related traits in rice-breeding projects. However, rice yield-related traits are quantitative traits and thus controlled by polygenes, whose genetic basis is complex and affected by environmental and genetic factors. With the rapid development of molecular quantitative genetics, complex quantitative traits are dissected into a single Gregor Mendel factor through molecular-map-based cloning. Determining its genomic position and analyzing its molecular biological function for the genetic improvement and polymerization breeding of quantitative traits has great significance.
QTLs underlying yield-related traits in rice have been extensively studied using a traditional genetic-mapping population (F2, F2,3, DH, RIL, and BC1F2). A total of 2060 QTLs controlling yield-related traits have been detected on the whole genome of rice. Some QTLs controlling yield-related traits including heading date, effective panicle number, grain number per panicle, grain length, and 1000-grain weight were successfully cloned and functionally analyzed. For the heading date of rice for Hd1[1], Hd6 [2], and Hd3a [3]. In particular, Gn1a is the first QTL control grain number per spike, cloned on the short arm of the first chromosome encoding a cytokinin oxidase dehydrogenase and degraded cytokinin. Meanwhile, Gn1a is a negative regulator that can control the number of spikelets by promoting the number of reproductive organs, thereby achieving the goal of yield control [4]. Ghd7 located on the 7th chromosome is a pleiotropism gene, which affects plant height, flowering period, spikelet number per panicle, and flag leaf area. It plays an important role in rice-yield increase and ecological adaptability [5]. GS3 controlling grain length and grain weight is located on the 3rd chromosome, explaining 80%–90% variation of grain weight and grain length in the near-isogenic lines of Minghui 63 and Chuan7 [6]. GW2 is a major gene on the 2nd chromosome that controls grain width and weight and increases grain-filling rate [7]. IPA1 is a promoter that binds the protein OSSPL14, promotes primary branch differentiation, inhibits tillering, and increases the number and weight of grains per panicle [8,9]. GW8 is located on the 8th chromosome that encodes a promoter-like SPB-box binding protein Osspl16. Osspl16 promotes glume cell division and grain filling, thereby increasing seed width and grain weight [10]. LRK1 is located in the leucine-rich receptor kinase gene cluster. LRK1 overexpression can significantly increase tillers, plant height, grain number, and grain weight per panicle [11]. In summary, the successful isolation and cloning of QTL for yield-related traits in rice have benefited from the innovative utilization of excellent genetic resources. Therefore, a series of unceasing excavation and innovation of rice germplasm resources can promote the transformation and upgrading of the rice-breeding industry.
Perennial Chinese rice can survive the natural cold-winter field environment, sprout from rice tillering node in the following spring, tiller, flower, seed, and be harvested in the following autumn. It is a type of special rice germplasm that can withstand cold and is harvested again at maturity. In agricultural production, if new perennial Chinese rice varieties with "high yield, good quality, and wide adaptability" can be cultivated and commercial released to farmer. Alleviating the negative impact of land wastage on rice production caused by urbanization, industrialization, and even sharp reduction of agricultural population has great practical significance [12]. In 1959, Yan first discovered the overwintering regeneration of rice in the open field in Changsha, Hunan province, and in Yuanjiang and Xiaogan, Hubei Province [13]. Nowadays, rice scientists are focusing on the characteristics of the overwintering germination of perennial rice, especially the yield stability of perennial rice and the genetic expression of yield-related traits such as growth period, tillering, and grain number per panicle in our country. The overwintering male sterile line, overwintering heterosis, combining ability analysis, and breeding of conventional overwintering varieties have also been reported [14,15,16]. Comparative analysis of the previous studies on perennial rice, the study progress on the genetic basis theory perennial rice and breeding application is still sluggish, mostly staying in the field yield traits investigation and breeding research of the preliminary exploration stage. However, little is known about its deep-seated genetic and molecular mechanism. In particular, the understanding of the genetic loci and their modes of action for the yield-related traits of perennial Chinese rice resources is relatively backward. Aggregating different high-yield genes of perennial rice at the molecular level into the same new perennial rice variety remains difficult. In the present study, perennial Chinese rice "Shendao3#" that can withstand cold tolerance to -1 °C of the daily minimum temperature for one day and 0 °C of the daily minimum temperature for three days in January 2021 throughout the cold-winter season of Chongqing Municipality in China was selected as materials [17]. The F2 population derived from the cross between "Shendao3#" and "XieqingzaoB" and its molecular genetic map was constructed to map QTLs underlying the yield-related traits. Our results can lay a foundation for further revealing the genetic mechanism of yield-related traits in perennial Chinese rice and its potential utilization in breeding.
2. Materials and Methods
2.1. Plant Materials
Perennial Chinese rice "Shendao3#" was selected as female parent material. It can withstand cold to -1 °C of the daily minimum temperature for one day, 0 °C of the daily minimum temperature for four days and 1 °C of the daily minimum temperature for four days in January 2021 throughout the cold-winter season of Chongqing Municipality, China (https://lishi.tianqi.com/bishan/202101). It can also sprout from rice tillering node in March 2021, tiller, flower, seed and be harvested again, thereby exhibiting perennial characteristics (Figure 1 a-g). However, XieqingzaoB is an inndica maintainer line and sensitive to low temperature. It cannot survive through the natural cold-winter field environment of Chongqing Municipality, China.
In mid-July 2022, the hybrid combination of "Shendao3#/XieqingzaoB" was constructed at the Rice Biotechnology Testing Station of Chongqing Normal University (CQNU), Shapingba district, Chongqing (29°32′ N, 106°32′ E), China. In late August of the same year, the seed hybrid F0 was planted at Nanfan breeding bases of the Sichuan Academy of Agricultural Sciences, Yingzhou town, Lingshui County, Hainan Province, China. In late December of the same year, seeds from F1 plants and F2 population were collected to construct the molecular-linkage map and identify the QTLs underlying the yield-related traits in perennial Chinese rice"Shendao3#".

2.2. Phenotyping Experiment
Rice-field experiments were conducted at the Rice Biotechnology Testing Station of CQNU, University Town, Shapingba district, Chongqing (29°32′ N, 106°32′ E), P. R. China. In March 2022, seeds of F2 "Shendao3#/XieqingzaoB" population and biparents (Shendao3# and XieqingzaoB) were sown on 9 March 2022. The 40-day-old seedlings of F2 "Shendao3#/XieqingzaoB" population and biparents (Shendao3# and XieqingzaoB) were transplanted into a single row with 10 plants having a 20 cm gap between plants within each row and a 25 cm gap between rows.
A rice special fertilizer 375 kg hm−2 occupied 70% of the total nutrients, including pure N 150 kg hm−2, 105 kg hm−2 P2O5, and 120 kg hm−2 K2O, was selected as base fertilizer for rice. Rice topdressing fertilizer including pure N 45 kg hm−2, 31.5 kg hm−2 P2O5, and 36 kg hm−2 K2O accounted for only 30% of the total nutrients. Rice topdressing fertilizer and seedling herbicide were applied after two weeks of transplanting. The water-management strategy was adapted in shallow water at the transplanting and tiller stages, and flooding mid-season with drainage–reflooding–moist intermittent irrigation was conducted. Disease, pest, and paddy weed management were performed at different growth stages of rice. At the tiller stage of rice, the leaf of "Shendao3#/XieqingzaoB" F2 individuals and biparents (Shendao3# and XieqingzaoB) were sampled to construct a rice molecular linkage map.
2.3. Trait Measurement
Referring to the method of Shen [18], at maturity stage, we sampled five individual plants of biparent “Shendao3# and XieqingzaoB” and 120 F2 individual plants random for trait measurement. A total of 15 yield-related traits were measured, including plant height (PH, in centimeter), panicle plant-1 (PP), panicle length (PL, in centimeter), fulling grains panicle-1 (FGP), empty grain panicle-1 (EGP), spikelet plant-1 (SP), grain yield major panicle (GYMP, in gram), grain yield plant (GYP, in gram), grain length (GL, in millimeter), grain width (GW, in millimeter), grain thickness (GT, in millimeter), and thousand-grain weight (TGW, in gram). We calculated three derived traits, namely, grain-setting rate (GSR =FGP/SP×100%), grain-setting density (GSD = SP/PL), and length-to-width ratio (LWR=GL/GW). The phenotypic value of single trait in the F2 plant was used as input data to identify QTLs. The measurement of all traits followed the Standard Evaluation System for Rice (http://www.Knowledgebank.irri.org/ses/).
2.4. Data Analysis
All measured trait phenotypic data in the F2 population and its biparents were inputted into Microsoft Excel 2010. With the same software, analysis of standard derivation (SD) and calculation of mean value and coefficient of variation (CV) were conducted. GraphPad Prism 5.0 was used to calculate the T-test value and phenotypic correlation coefficient (PCC), as well as draw the histogram.
2.5. Linkage Map and QTL Analysis
We constructed a linkage map containing 112 SSR markers and covering the whole genomic length of 1950.72cM with an average interval of 17.14 cM between markers by using QTLIcimapping version 4.20 [19]. It was also used to perform QTL analysis for the yield-related traits. Inclusive composite interval mapping (ICIM) for QTL analysis in the F2 population was performed using IciMapping 4.20 software [19]. The PIN of 0.01 and walking speed of 1 cM were set. An LOD threshold was used as 1000 permutations to advocate QTLs. An LOD score of ≥3.0 was used to declare the significant major QTL. The contribution rate (R2,%) was estimated as the percentage of variance explained by each locus in proportion to the total phenotypic variance. QTL nomenclature followed the principle suggested by McCouch [20]. Digenic epistasis QTLs in the F2 population were identified by the ICIM-EPI mapping of IciMapping 4.20 version with mapping parameters of 1 cM steps and 0.001 probabilities in a stepwise regression. Genome-wide threshold F-values (F=0.001) were estimated from 1000 permutions to indicate the presence of epistasis [21].
3. Results
3.1. Phenotypic Variation for Yield-Related Traits in the Biparents and F2 Population
A significant phenotypic difference was observed between Shendao3# and XieqingzaoB for all measured traits, except for EGP, GL, LWR, and TGW (Table 1). It showed a wide T-test value ranging from 0.10 to 13.07. Meanwhile, Shendao3# had significantly higher phenotypic values on PH, FGP, SP, GSR, GSD, GYP, GW, and GT than those of XieqingzaoB. However, XieqingzaoB had significantly higher phenotypic values on PP and PL than those of Shendao3#. All measured traits showed tremendous bidirectional segregation beyond biparents, except for PP, FGP, and GSR, which displayed a wide coefficient of variation ranging from 7.02% for GL to 67.52% for EGP in the F2 population. It continuously demonstrated normal distribution and belonged to quantitative inheritance (Figure 2), which was suited to QTL mapping.
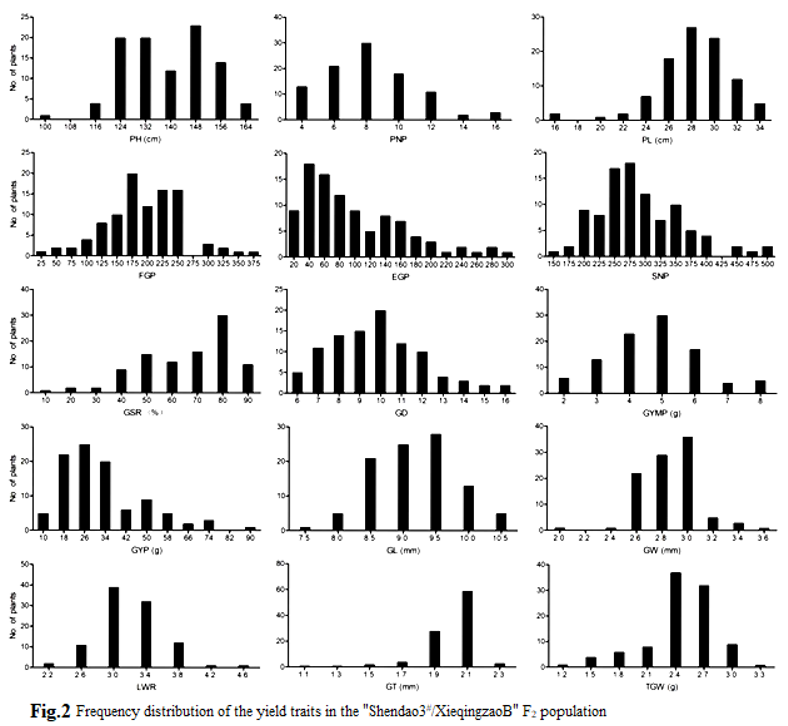
3.2. Trait Correlations
Altogether 61 significant correlation pairs for 15 yield-related traits were estimated in the F2 population (Table 2). In the F2 population, 46 significantly positive correlation pairs and 15 significantly negative correlation pairs exhibiting a wide absolute value ranging from 0 to 1.00 were calculated for all measured traits. PL showed significantly positive correlation with PH, PP, FGP, EGP, SP, GYMP, GYP, GL, and LWR, respectively. Correlation coefficients (r) ranged from 0.19 to 0.44. GYP showed significantly positive correlation with PH, PP, PL, FGP, SP, GSR, GSD, GYMP and TGW, respectively. Correlation coefficients (r) ranged from 0.24 to 0.80. PL and GYP were regarded as some of the most important determinants in the yield-related traits of perennial Chinese rice. GW showed a significantly positive correlation only with PH, EGP, and GSD, with correlation coefficients (r) ranging from 0.16 to 0.24. GW determined the field performance of the three yield-related traits. TGW showed significantly negative correlation with EGP, SP, GSD, and GW, with correlation coefficients (r) ranging from -0.15 to -0.38. It was negatively correlated with the other four measured traits. GW showed a significantly negative correlation with EGP, GSR, GYMP, and GW, with correlation coefficients (r) ranging from -0.15 to -0.27. It was negatively correlated with the other four measured traits. PH showed a significantly negative correlation only with GSR, with a correlation coefficient of -0.21. In summary, the yield-related trait in perennial Chinese rice "Shendao3#" was a very complex character. It showed complex correlations among all measured traits directly or indirectly determined the formation of yield traits.
3.3. QTLs for Yield-Related Traits
A total of 25 QTLs affecting 15 yield traits were detected on the 1st , 2nd, 3rd, 4th, 5th, 7th, 8th, 9th, 10th, and 11th chromosomes, respectively. LOD values ranged from 3.00 to 6.06, which explained a wide phenotypic variation ranging from 4.21% for qGW2a to 27.30% for qPH1 (Figure 3; Table 3). For PH, only one QTL qPH1 was detected on the 1st chromosome and explained a phenotypic variation of 27.30%. Moreover, the Shendao3# allele increased PH at qPH1 genetic loci. For the number of panicle plant-1, four QTLs (qPP1, qPP3, qPP7 and qPP11) were detected on the 1st, 3rd, 7th, and 11th chromosomes, respectively, which explained the phenotypic variation ranging from 7.33% to 10.82%. Among them, Shendao3# alleles increased the number of panicle plant-1 at qPP1, qPP3, and qPP11 but XieqingzaoB allele increased PP at qPP7. For panicle length, three QTLs (qPL1, qPL2, and qPL11) were detected on the 1st, 2nd, and 11th chromosomes, respectively. It explained the phenotypic variation ranging from 6.07% to 8.23%, XieqingzaoB allele increased PL at qPL1 but Shendao3# alleles increased PL at qPL2, and qPL11. For the grain yield plant-1, four QTLs (qGYP7, qGYP9, qGYP10a, and qGYP10b) were detected on the 7th, 9th, and 10th chromosomes, respectively. It explained the phenotypic variation ranging from 5.16% to 9.07%, XieqingzaoB allele increased GYP at qGYP7, but Shendao3# alleles increased GYP at qGYP9, qGYP10a, and GYP10b, respectively. For spikelet plant-1, grain thickness, and empty grain panicle-1, three QTLs (qSP1, qGT5, and qEGP8) were detected on the 1st, 5th, and 8th chromosomes, respectively. It explained 25.56%, 9.30%, and 14.32% of the phenotypic variation, respectively, XieqingzaoB allele increased EGP at qEGP8 but Shendao3# alleles increased SP at qSP1 and GT at qGT5. For grain-setting rate, two QTLs (qGSR4 and qGSR8) were detected on the 4th and 8th chromosomes, which explained 4.61% and 5.40% of the phenotypic variation, respectively, Shendao3# allele increased GSR at qGSR4 but XieqingzaoB allele increased GSR at qGSR8. For grain-setting density, two QTLs (qGSD1 and qGSD2) were detected on the 1st and 2nd chromosome, which explained 14.11% and 20.65% of the phenotypic variation, respectively, Shendao3# alleles increased GSD at qGSD1 and qGSD2. For thousand-grain weight, two QTLs (qTGW1 and qTGW4) were detected on the 1st and 4th chromosomes, which explained 4.76% and 5.08% of the phenotypic variation, respectively, Shendao3# alleles increased TGW at qTGW1 and qTGW4. For grain width, four QTLs (qGW2a, qGW2b, qGW11a, and qGW11b) were detected on the 2nd and 11th chromosomes, which explained the phenotypic variation ranging from 4.76% to 5.08%. Shendao3# alleles increased GW at qGW2a, qGW2b, qGW11a, and qGW11b, respectively.

3.4. Pleiotropic QTLs for Yield Traits
The chromosomal locations of all QTLs identified in the F2 population revealed six genomic regions on the 1st, 4th, 7th, 8th, and 11th chromosomes where clustered QTLs affected two yield traits (Figure 3; Table 4). For the 1st chromosome, two QTL (qSP1 and qGSD1) genomic regions underlying spikelet panicle-1 and grain-setting density were flanked repeatedly by RM265 and RM3738 on the 1st chromosome. The Shendao3# alleles increased SP at qSP1 and GSD at qGSD1. Two QTL (qPP1 and qPL1) genomic regions affecting panicle plant-1 and panicle length were flanked repeatedly by RM3642 and RM600 on the 1st chromosome. The Shendao3# allele increased PP at qPP1, and XieqingzaoB allele increased PL at qPL1. For the 4th chromosome, two QTL (qGSR4 and qTGW4) genomic regions were flanked repeatedly by RM7051 and RM5633 controlling grain-setting rate and thousand-grain weight. The Shendao3# allele increased GSR at qGSR4 and TGW at qTGW4. For the 7th chromosome, two QTL (qPP7 and qGYP7) genomic regions were flanked repeatedly by RM1135 and RM5793 and repeatedly affected panicle plant-1 and grain yield plant-1, respectively. The XieqingzaoB allele increased PP at qPP7 and GYP at qGYP7. For the 8th chromosome, two QTL (qEGP8 and qGSR8) genomic regions were flanked repeatedly by RM1111 and RM3702, which were affected simultaneously empty grain panicle-1 and grain-setting rate. The Shendao3# allele increased EGP at qEGP8, and XieqingzaoB allele increased GSR at qGSR8. For the 11th chromosome, two QTL (qPP11 and qGW11a) genomic regions were flanked repeatedly by RM7120 and RM6293 and affected simultaneously the number panicle plant-1 and grain width. The XieqingzaoB allele increased PP at qPP11, and the Shendao3# allele increased GW at qGW11a. For the pleiotropic QTLs that controlled two or more yield-related traits, they were located repeatedly at the same marker interval. Thus, pleiotropic QTLs should be given more attention in future rice genetic and breeding projects. It's twice as efficient that multi-yield traits will be synchronous improved through one yield trait was genetically improved in the future rice genetic and breeding project.
3.5. Digenic Epistatic QTLs for Yield-Related Traits
A total of 26 epistatic QTL pairs for 15 yield traits were detected on the whole genome of rice, except for the 6th and 11th chromosomes in the F2 population. It accounted for an extensive phenotypic variation ranging from 1.15% to 19.30%, with wide-ranging LODs from 5.01 to 8.74 (Table 5). Among them, eight epistatic QTL pairs were found for GSR, six for SP, five for GYP, for PL, and one each for PH, EGP, GW, and GT. For all epistatic QTL pairs exhibiting four interaction patterns including additive–additive, additive–dominant, dominant–additive, and dominant–dominant were identified in the F2 population. A total of 11 out of 24 genomic regions of the QTLs interacted repeatedly with two or more marker intervals. For example, the genomic interval RM3642-RM600 underlying PP and PL on the 1st chromosome interacted with the genomic interval RM6519-RM5651 and the genomic interval RM8019-RM6990 on the 8th chromosome. The genomic interval RM250-RM3763 underlying PL and GW on the 2nd chromosome interacted with the interval RM1352-RM3199 on the 3rd chromosome and the genomic interval RM559-RM5979 on the 4th chromosome. The genomic interval RM7051-RM5633 underlying GSR on the 4th chromosome interacted with the genomic interval RM7637-RM5812 on the 2nd chromosome and the genomic interval RM317-RM7051 underlying both GSR and GW on the 4th chromosome, the genomic interval RM24085-RM160 on the 9th chromosome. The genomic interval RM1135-RM5793 underlying PP and GYP on the 7th chromosome interacted with the interval RM3555-RM5481 on the 7th chromosome, the genomic interval RM1111-RM3702 on the 8th chromosome, and the genomic interval RM202-RM7120 on the 11th chromosome. The genomic interval RM1111-RM3702 underlying EGP and GSR on the 8th chromosome interacted with the genomic interval RM5793-RM432 on the 7th chromosome, the genomic interval RM1135-RM5793 underlying PP and GYP on the 7th chromosome, the genomic interval RM257-RM5661 and RM24085-RM160 on the 9th chromosome, and the genomic interval RM5708-RM3882 on the 10th chromosome. the genomic interval RM202-RM7120 and RM6293-RM1341 on the 11th chromosome. The genomic interval RM24085-RM160 underlying GYP on the 9th chromosome interacted with the genomic interval RM7051-RM5633 underlying GSR on the 4th chromosome and the genomic interval RM1111-RM3702 underlying GSR on the 8th chromosome. The genomic interval RM5708-RM3882 underlying GYP on the 10th chromosome interacted with the interval RM85-RM3856 underlying PP on the 3rd chromosome, the genomic interval RM317-RM7051 underlying GSR on the 4th chromosome, and the genomic interval RM1111-RM3702 underlying GSR on the 8th chromosome. the genomic interval RM202-RM7120 on the 8th chromosome. The genomic interval RM202-RM7120 on the 11th chromosome interacted with the interval RM3738-RM8084 underlying PH on the 1st chromosome, the genomic interval RM1135-RM5793 underlying PP and GYP on the 7th chromosome. And the genomic interval RM5708-RM3882 underlying GYP on the 10th chromosome. the genomic interval RM405-RM26 underlying GT on the 5th chromosome. the genomic interval RM7120-RM6293 on the 11th chromosome interacted with the genomic interval RM559-RM5979 underlying GSR on the 4th chromosome and the genomic interval RM24085-RM160 underlying GYP on the 9th chromosome. The remaining 11 epistatic QTL pairs involved only one interaction pair.
4. Discussion
The cardinal goal of any study on the genetics mechanism of quantitatively inherited traits is to identify the major QTLs hidden in different rice germplasm resources and finally mine the interest gene for rice molecular breeding [22]. In this study, perennial Chinese rice "Shendao3#" was selected as plant material to identify QTLs underlying yield-related traits and thus lay a good foundation for the molecular-design breeding of yield-related genes in perennial Chinese rice.
A total of 25 QTLs underlying the 15 yield-related traits were detected on the whole genome except for the 6th and 11th chromosomes. Among them, six major QTLs can explain more than 10% of phenotypic variation. Six pleiotropic QTL genomic regions controlling two or more yield-related traits were detected on the 1st, 4th, 7th, 8th, and 11th chromosomes, respectively. All QTLs followed the genetic model of additive, dominant and epistatic effect of additive by additive, additive by dominant, dominant by additive, and dominant by dominant in the "Shendao3#/XieqingzaoB" combination. However, the remaining 13 QTL genomic regions such as qPP3, qPL2, qPL11, qGYP9, qGYP10a, and qGYP10b were detected for only a single yield trait. Among the 25 QTLs, 14 were located the same genomic region and even cloned. Accordingly, we searched their alignment QTLs identified in the same chromosomal region through the publicly available QTL database (www.gramene.org) published by previous researchers. For qPH1 underlying PH, it has been previously reported in the Yangdao6/Lvhan1# RIL population [23]. For qPP1 related to PP, it has been previously detected on the 1st chromosome in the Zhongjiazao17/D50 F2 population [24]. For qPP3 related to PP, it has been detected on the 3rd chromosome in the Zhaiyeqin8/Jingxi17 DH population [25]. For qPP11 related to PP, it has been detected on the 11th chromosome in the Oryza sativa×Oryza rufipogon BC2F2 population [26]. For qPL1 related to PL, it has been detected on the 1st chromosome in the Zhongjiazao17/D50 DH population [27]. For qPL2 related to PL, it has been detected on the 2rd chromosome in the O.rufipogon/Jefferson BC2F2 population [28]. For qPL11 related to PL, it has been detected on the 11th chromosome in the Lemont/Teqing RIL population [29]. For qGSR4 underlying GSR, it has been previously reported in the two populations of Zhaiyeqing8/Jingxi17 DH [25,30] and Zhongjiazao17 /D50 F2 [24]. For qGSR8 related to GSR, it has been detected on the 8st chromosome in the Nipponbare/Zhongjiazao17 RIL population [31]. For qGW2a and qGW2b related to the GW, it has been detected on the 2nd chromosome in the Nipponbare/Zhongjiazao17 RIL population [31]. For qGW11a related to GW, it has been detected on the 11th chromosome in the O.sativa/O.glaberrima DH population [32]. For qGW11b related to GW, it has been detected on the 11th chromosome in the IR64/Azucena DH population [33]. For qGT5 related to GW, it has been detected on the 11th chromosome in the four populations of Tesanai2/CB F2 and F2,3[34], O.rufipogon/V20A BCF2 [35], and Oryza rufipogon Griff/Guichao2 ILs [36].
However, 11 QTLs associated with the yield-related were primarily mapped in the "Shendao3# /XieqingzaoB" combination. Among them, for qPP7 related to the number of panicle per plant and qGYP7 underlying the grain yield plant-1, XieqingzaoB alleles increased PP at qPP7 and increased GYP at qGYP7. For the remaining nine QTLs related with the yield-related traits, the Shendao3# alleles increased EGP at qEGP8, SP at qSP1, GSD at qGSD1 and qGSD2, GYP at qGYP9, qGYP10a, and qGYP10b, and TGW at qTGW1 and qTGW4, respectively. Nine newly discovered QTLs (qEGP8, qSP1, qGSD1, qGSD2, qGYP9, qGYP10a, qGYP10b, qTGW1, and qTGW4) underlying the yield-related traits might exhibit a unique genetic model of the perennial Chinese rice "Shendao3#". They may also provide a novel genetic resource for perennial Chinese rice varieties’ breeding. Therefore, special efforts should be devoted to fine mapping these newly discovered QTLs and studying their positional cloning for perennial Chinese rice-molecular breeding.
Six QTL chromosomal regions associated with two or more yield-related traits were detected repeatedly in the "Shendao3#/XieqingzaoB" F2 population. For example, the qPP1 and qPL1 were flanked repeatedly at the marker interval of RM1135-RM57930 on the 1st chromosome, and qPP7 and qGYP7 were detected repeatedly at the marker interval of RM1135-RM57930 on the 7th chromosome. Similarly, the clustered distribution of QTL has also been reported in previous studies [37,38,39]. Consequently, paying more attention to pleiotropic QTLs (chromosome regions) in future rice genetic and breeding projects is necessary. It's twice as efficient that multi-yield traits will be synchronous improved through one yield trait was genetically improved in the future rice genetic and breeding project.
5. Conclusion
Perennial Chinese rice "Shendao3#" was selected as parental materia. The F2 population of "Shen dao3#/XieqingzaoB" was constructed to identify QTLs associated with 15 yield-related traits. A total of 25 QTLs and 26 epistatic QTLs were detected on the whole genome, except for the 6th and 12th chromosomes. It explained phenotypic variation ranging from 4.21% to 27.30% and 1.15% to 19.30%. Six pleiotropic QTLs (chromosomal regions) associated with two or more yield-related traits were detected repeatedly in the "Shendao3#/XieqingzaoB" F2 population. Among 24 QTLs, 11 were associated with yield-related traits and were primarily mapped in the "Shendao3#/XieqingzaoB" combination. All QTLs associated with 15 yield-related traits followed the genetic model of additive, dominant, and digenic epistatic of additive by additive, additive by dominant, dominant by additive, and dominant by dominant. This study laid a good foundation for yield-related gene mining and its utilization in breeding perennial Chinese rice "Shendao3#".
Author Contributions
YSL carried out the supervision, data curation, writing-original draft and funding acqusition; JYG provided perennial rice seeds; YXY carried out the agronomic trait investigation and QTL analysis; TSP carried out the agronomic trait investigation and the DNA extraction; JYL carried out the agronomic trait investigation and PCR amplification; LT carried out the agronomic trait investigation; ML and WBW carried out data curation; XJQ carried out the field experiments. All authors have reviewed and approved the final manuscript.
Funding
This work was supported by the State Key Laboratory of Crop Gene Exploration and Utilization in Southwest China (SKL-KF202226) and the Chongqing Natural Science Foundation of China (cstc2021jcyj-msxmX0007), and the Open Project Program of State Key Laboratory of Rice Biology (20190202).
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationship that may appear to influence the work reported in this paper.
Abbreviations
QTL: quantitative trait locus; SSR: single sequence repeat; PH: plant height; PP: panicle plant-1; PL: panicle length; FGP: fulling grains panicle-1; EGP: empty grain panicle-1; SP: spikelet plant-1; GYMP: grain yield major panicle; GYP: grain yield plant; GL: grain length; GW: grain width; GT: grain thickness; TGW: thousand-grain weight; GSR: grain-setting rate; GSD: grain-setting density; LWR: length-to-width ratio; SD: standard derivation; CV: coefficient of variation; PCC: phenotypic correlation coefficient; ICIM: Inclusive composite interval mapping.
References
- Yano M, Katayose Y, Ashikari M, Yamanouchi U. Hd-1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene constans. Plant Cell. 2000, 12, 2473–2483. [CrossRef]
- Takahashi Y, Shomura A, Sasaki T, Yano M. Hd6, a rice quantitative trait locus involved in photoperiod sensitivity, encodes the a subunit of protein kinase CK2. Proceedings of the National Academy of Sciences. 2001, 98, 7922–7927. [CrossRef]
- Kojima S, Takahashi Y, Kobayashi Y. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowing downstream of Hd1 under short-day conditions. Plant Cell Physiology. 2002, 43, 1096–1105.
- Ashikari M, Sakakibara H, Lin SY, Yamamoto T, Takashi T, Nishimura A, et al. Cytokinin oxidase regulates rice grain production. Science. 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Xue WY, Xing YZ, Weng XY, Zhao Y, Tang WJ, Wang L, et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nature Genetics. 2008, 40, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Fan CC, Xing YZ, Mao HL, Lu TT, Han B, Xu CG, et al. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transmembrane protein. Theoretical Applied Genetics. 2006, 112, 1164–1171.
- Song XJ, Huang W, Shi M, Zhu MZ, Lin HX.A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nature Genetics. 2007, 39, 623–630. [CrossRef]
- Jiao YQ, Wang YH, Xue DX, Wang J, Yan MX, Liu GF, et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nature Genetics. 2010, 42, 541–544. [Google Scholar] [CrossRef]
- Miura K, Ikeda M, Matsubara A, Song XJ, Ito M, Asano K, et al. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nature Genetics. 2010, 42, 545–549. [CrossRef]
- Wang SK, Wu K, Yun,QB, Liu XY, Liu ZB, Lin XY, et al. Control of grain size, shape and quality by OsSPL16 in rice. Nature Genetics. 2012, 44, 950–954. [Google Scholar] [CrossRef]
- He GM, Luo XJ, Tian F, Li KG, Zhu ZF, Su W, et al. Haplotype variation in structure and expression of a gene cluster associated with a quantitative trait locus for improved yield in rice. Genome Research. 2006, 16, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Zhang SL, Huang GF, Zhang YJ, Lv XT, Wan KJ, Liang J, et al. Sustained productivity and agronomic potential of perennial rice. Nature Sustainability. 2022, 6, 28–38. [Google Scholar] [CrossRef]
- Yan, F. Research status and prospect of overwintering ratooning rice. Crop Research. 2012, 26, 79–84. [Google Scholar]
- He HH, Hu DL, Fu JR, Liu YB, Li HB, Zou XS, et al. Breeding of surviving in winter lines and surviving in winter sterile lines in rice. Acta Agriculturae universityaties JiangXiensis. 2005, 27, 659–661. [Google Scholar]
- Fu JR, Li HB, Hu DL, Liu YB, Zou XY, He HH. Analysis on heterosis and combining ability of surviving in winter (SW) lines in rice. Acta Agriculturae universityaties JiangXiensis. 2007, 29, 323–330. [Google Scholar]
- Zhang SL, Huang GF, Zhang YJ, Lian XP, Zhang J, Chen R, Hu FY. Breeding progress and prospect of perennial rice. China Rice. 2022, 28, 39–43. [Google Scholar]
- Liang YS, Gong, JY, Yan YX, Wang BB, Gong WA, Wen H, et al. Survey of overwintering trait in Chinese rice cultivars (Oryza sativa L). Euphytica. 2022, 218, 94. [Google Scholar] [CrossRef]
- Shen, ZD. Crop Breeding Experiment. Beijing: Chinese Agricultural Press. 1995, 112-114.
- Li HH, Ribaut JM, Li ZH, Wang JK. Inclusive composite interval mapping (ICIM) for digenic epistasis of quantitative traits in biparental populations. Theoretical Applied Genetics. 2008, 116, 243–260. [Google Scholar] [CrossRef]
- McCouch SR, Cho YG, Yang M. Report on QTL nomenclature. Rice Genet Newsl. 1997, 14, 11–13. [Google Scholar]
- MengL, Li HH, Zhang LY, Wang JK. QTL IciMapping: integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. The Crop Journal. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- Wang JK, Li HH, ZhangXC, Yin CB, Li Y, Ma YZ, et al. Molecular design breeding in crops in China. Acta Agronomical Sinica. 2011, 37, 191–201. [Google Scholar] [CrossRef]
- Han LF, Zhou R, Zhou T, Lin CX, Gan Q, Ni DH, Shi YY, Song FH. Correlation analysis and QTLs mapping of lodging resistance and yield traits in rice. Journal of Biology. 2023, 40, 65–70. [Google Scholar]
- Sheng ZH, Zhu ZL, Ma N, LiW, He JW, Wei XJ, et al. QTL mapping of yield related traits in super rice variety Zhongjiazao 17. Chinese Journal Rice Science. 2016, 30, 35–43. [Google Scholar]
- Tan ZB, Shen LS, Yuan ZL, Lu CF, Chen Y, Zhou KD, Zhu LH. Identification of QTLs for ratooning ability and grain yield traits of rice and analysis of their genetic effects. Acta Agronomica Sinica. 1997, 23, 289–295. [Google Scholar]
- Moncada P, Martinez CP, Borrero J, Chatel M, Gauch H, Guimaraes E, Tohme J, McCouch SR. Quantitative trait loci for yield and yield components in an Oryza sativa×Oryza rufipogon BC2F2 population evaluated in an upland environment. Theoretical Applied Genetics. 2001, 102, 41–42. [Google Scholar] [CrossRef]
- Hu DW, Sheng ZH, Chen W, Li QL, Wei XJ, Shao GN, et al. Identfication of QTLs associated with high yield of super rice variety Zhongjiazao 17. Acta Agronomica Sinica. 2017, 43, 1434–1447. [Google Scholar] [CrossRef]
- Thomson MJ, Tai TH, McClung AM, Lai XH, Hinga ME, Lobos KB, et al. Mapping quantitative trait loci for yield, yield components and morphological traits in an advanced backcross population between Oryza rufipogon and the Oryza sativa cultivar Jefferson. Theoretical Applied Genetics. 2003, 107, 479–493. [Google Scholar] [CrossRef]
- Xu JL, Xue QZ, Luo LJ, Li ZK. QTL dissection of panicle number per plant and spikelet number per panicle in rice (Oryza sativa L). Acta Genetica Sinica. 2001, 28, 752–759. [Google Scholar]
- Lu CF, Shen LH, Tan ZB, Xu YB, He P, Chen Y, Zhu LH. Comparative mapping of QTLs for agronomic traits of rice across environments by using a doubled-haploid population. Theoretical Applied Genetics. 1997, 94, 145–150. [Google Scholar] [CrossRef]
- Zhang YZ, Luo RJ, Sheng ZH, Jiao GA, Tang SQ, Hu PS, Wei XJ. QTL mapping of yield associated traits of Nipponbare/Zhongjiazao17 RIL population. Scientia Agricultura Sinica. 2017, 50, 3640–3651. [Google Scholar]
- Aluko G, Martinez C, Tohme J, Castano C, Bergman C, Oard JH. QTL mapping of grain quality traits from the interspecific cross Oryza sativa×O. glaberrima. Theoretical Applied Genetics. 2004, 109, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Huang N, Parco A, Mew T, Magpantay G, McCouch SR, Guiderdoni E, et al. RFLP mapping of isozymes, RAPD and QTLs for grain shape, brown planthopper resistance in a doubled haploid rice population. Molecular Breeding. 1997, 3, 105–113. [Google Scholar] [CrossRef]
- Zhuang JY, Lin HX, Lu J, Qian HR, Hittalmani S, Huang N, Zheng KL. Analysis of QTL x environment interaction for yield components and plant height in rice. Theoretical Applied Genetics. 1997, 95, 799–808. [Google Scholar] [CrossRef]
- Xiao JH, Li J, Grandillo S, Ahn SN, Yuan L, Tanksley SD, McCouch SR. Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genetics. 1998, 150, 899–909. [Google Scholar] [CrossRef]
- Tian F, Li DJ, Fu Q, Zhu ZF, Fu YC, Wang XK, Sun CQ. Construction of introgression lines carrying wild rice (Oryza rufipogon Griff.) segments in cultivated rice (Oryza sativa L) background and characterization of introgressed segments associated with yield-related traits. Theoretical Applied Genetics. 2005, 1–11.
- Yu SB, Li JX, Xu CG, Tan YF, Li XH, Zhang QF. Identification of quantitative trait loci and epistatic interactions for plant height and heading date in rice. Theoretical Applied Genetics. 2002, 104, 19–35. [Google Scholar]
- You AQ, Lu XG, Jin HJ, Ren X, Liu K, Yang GC, et al. Identification of quantitative trait loci across recombinant inbred lines and testcross populations for traits of agronomi importance in rice. Genetics. 2006, 105, 1287–1300. [Google Scholar]
- Liang YS, Zhan XD, Wang HM, Gao ZQ, Lin ZC, Chen DB, et al. Locating QTLs controlling several adult root traits in an elite Chinese hybrid rice. Gene. 2013, 526, 331–335. [Google Scholar] [CrossRef]

Table 1.
Field performance of yield traits in biparents and "ShenDao3#/XieqingzaoB" F2 population.
| Traits | Parents | F2 population | ||||
|---|---|---|---|---|---|---|
| Shendao3# | XieqingzaoB | T-test value | Means±SD | Range | CV (%) | |
| PH (cm) | 102.80 | 82.40 | 13.07** | 137.96±16.36 | 100.00-167.00 | 9.78 |
| PP | 9.80 | 13.20 | 3.30* | 7.73±2.86 | 3.00-17.00 | 37.80 |
| PL (cm) | 17.60 | 22.04 | 7.89** | 28.33±3.15 | 16.20-34.00 | 11.78 |
| FGP | 157.20 | 96.60 | 6.13** | 193.29±62.46 | 27.00-379.00 | 32.12 |
| EGP | 6.20 | 10.00 | 1.26 | 99.02±68.02 | 12.00-296.00 | 67.52 |
| SP | 163.40 | 106.60 | 4.56** | 292.32±72.86 | 143.00-502.00 | 24.17 |
| GSR (%) | 96.25 | 90.88 | 2.74* | 67.00±0.19 | 8.36-94.59 | 27.74 |
| GSD | 9.28 | 4.83 | 7.85** | 10.32±2.25 | 6.27-17.16 | 22.07 |
| GYMP (g) | 3.99 | 2.71 | 6.29** | 5.26±1.47 | 2.32-8.94 | 27.88 |
| GYP (g) | 33.04 | 24.27 | 4.43* | 34.38±15.84 | 5.57-88.84 | 47.17 |
| GL (mm) | 10.38 | 9.80 | 0.56 | 10.16±9.49 | 7.60-10.72 | 7.02 |
| GW (mm) | 3.43 | 2.58 | 11.63** | 2.87±0.23 | 2.17-3.71 | 7.81 |
| LWR | 4.01 | 3.81 | 0.10 | 3.56±3.35 | 2.18-4.54 | 11.78 |
| GT (mm) | 2.31 | 2.07 | 6.61** | 1.99±0.18 | 1.17-2.21 | 8.70 |
| TGW (g) | 24.60 | 26.80 | 2.42 | 24.70±0.37 | 13.50-33.90 | 15.15 |
*,** represent the significant difference at level of 5% and 1% (α0.05=2.57; α0.01=4.03).
Table 2.
Correlation coefficients (r) among fifteen yield-related traits in the "Shendao3#/XieqingzaoB"F2 population.
Table 2.
Correlation coefficients (r) among fifteen yield-related traits in the "Shendao3#/XieqingzaoB"F2 population.
| Traits | PH | PP | PL | FGP | EGP | SP | GSR | GSD | GYMP | GYP | GL | GW | LWR | GT | TGW |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PH | 1.00 | ||||||||||||||
| PP | 0.33** | 1.00 | |||||||||||||
| PL | 0.37** | 0.25** | 1.00 | ||||||||||||
| FGP | 0.04 | 0.12 | 0.20** | 1.00 | |||||||||||
| EGP | 0.28** | 0.10 | 0.29** | -0.37** | 1.00 | ||||||||||
| SP | 0.30** | 0.20** | 0.44** | 0.52** | 0.60** | 1.00 | |||||||||
| GSR | -0.21** | -0.02 | -0.13 | 0.68** | -0.90** | -0.24** | 1.00 | ||||||||
| GSD | 0.14 | 0.11 | -0.03 | 0.47** | 0.52** | 0.88** | -0.20* | 1.00 | |||||||
| GYMP | 0.15* | 0.20** | 0.30** | 0.78** | -0.23* | 0.45** | 0.47** | 0.34** | 1.00 | ||||||
| GYP | 0.33** | 0.80** | 0.35** | 0.51** | -0.09 | 0.36** | 0.26** | 0.21** | 0.60** | 1.00 | |||||
| GL | 0.08 | 0.15* | 0.19* | -0.06 | 0.23** | 0.16* | -0.15* | 0.07 | 0.02 | 0.09 | 1.00 | ||||
| GW | 0.16* | 0.09 | -0.14 | -0.20** | 0.24** | 0.09 | -0.27** | 0.18* | -0.17* | -0.06 | -0.03 | 1.00 | |||
| LWR | 0.07 | 0.14 | 0.20* | -0.04 | 0.20** | 0.15* | -0.13 | 0.05 | 0.03 | 0.09 | 1.00** | -0.11 | 1.00 | ||
| GT | -0.01 | 0.02 | 0.07 | -0.11 | -0.10 | -0.19* | -0.00 | -0.25** | 0.14 | 0.07 | 0.01 | 0.12 | 0.00 | 1.00 | |
| TGW | -0.02 | 0.17* | 0.13 | -0.05 | -0.27** | -0.29** | 0.15* | -0.38** | 0.25** | 0.24** | -0.05 | -0.15* | -0.03 | 0.47** | 1.00 |
*,** represent the significant difference at level of 5% and 1% (α0.05=0.146;α0.01=0.191).
Table 3.
QTLs underlying the yield-related traits in perennial Chinese rice "Shendao3#".
| Trait | QTL | Chromosome | Genomic Position | Marker interval | LOD | Additive | Dominant | R2 (%) | Derived |
|---|---|---|---|---|---|---|---|---|---|
| PH | qPH1 | 1 | 34902085-37261443 | RM3738-RM8084 | 6.06 | 9.93 | 7.21 | 27.30 | Shendao3# |
| PP | qPP1 | 1 | 9463544-24866202 | RM3642-RM600 | 3.06 | -3.02 | -4.80 | 8.06 | XieqingzaoB |
| qPP3 | 3 | 4333680-13933574 | RM489-RM6080 | 3.02 | 3.84 | -5.56 | 7.33 | Shendao3# | |
| qPP7 | 7 | 16932001-17489638 | RM1135-RM5793 | 5.23 | -3.58 | -6.52 | 10.82 | XieqingzaoB | |
| qPP11 | 11 | 11763775-2888052 | RM7120-RM6293 | 4.28 | -3.04 | -5.09 | 8.53 | XieqingzaoB | |
| PL | qPL1 | 1 | 27925715-32774365 | RM3642-RM600 | 3.00 | 6.05 | 4.19 | 6.07 | Shendao3# |
| qPL2 | 2 | 13481661-19677083 | RM250-RM3763 | 4.67 | -6.67 | 3.94 | 8.05 | XieqingzaoB | |
| qPL11 | 11 | 1124242-4773752 | RM1341-RM3428 | 3.96 | -3.11 | 5.53 | 8.23 | XieqingzaoB | |
| EGP | qEGP8 | 8 | 35196573-37261443 | RM1111-RM3702 | 3.10 | 49.48 | -132.62 | 14.32 | Shendao3# |
| SP | qSP1 | 1 | 13059580-24116775 | RM265-RM3738 | 5.52 | 48.12 | 33.14 | 26.56 | Shendao3# |
| GSR | qGSR4 | 4 | 11389704-20800963 | RM7051-RM5633 | 3.88 | 0.02 | -0.32 | 4.61 | Shendao3# |
| qGSR8 | 8 | 16932001-17489638 | RM1111-RM3702 | 5.90 | -0.17 | 0.46 | 5.40 | XieqingzaoB | |
| GSD | qGSD1 | 1 | 10811135-19788247 | RM265-RM3738 | 4.53 | 0.47 | 1.82 | 14.11 | Shendao3# |
| qGSD2 | 2 | 2722348-14527760 | RM324-RM262 | 5.25 | 1.49 | -3.80 | 20.65 | Shendao3# | |
| GYP | qGYP7 | 7 | 2722348-13761888 | RM1135-RM5793 | 3.32 | -14.32 | -23.54 | 7.25 | XieqingzaoB |
| qGYP9 | 9 | 4407860-23568212 | RM24085-RM160 | 3.41 | 21.40 | -14.19 | 5.79 | Shendao3# | |
| qGYP10a | 10 | 11389704-15894177 | RM5708-RM3882 | 4.24 | 17.86 | -13.64 | 9.07 | Shendao3# | |
| qGYP10b | 10 | 19677083-28788052 | RM3882-RM8201 | 3.74 | 19.60 | -23.56 | 5.16 | Shendao3# | |
| GW | qGW2a | 2 | 3073406-27342022 | RM5651-RM3732 | 3.04 | 0.34 | -0.37 | 4.21 | Shendao3# |
| qGW2b | 2 | 11389704-15894177 | RM5812-RM324 | 4.07 | 0.28 | -0.26 | 5.38 | Shendao3# | |
| qGW11a | 11 | 11763775-28788053 | RM7120-RM6293 | 3.82 | 0.27 | -0.35 | 5.34 | Shendao3# | |
| qGW11b | 11 | 19677083-28788052 | RM6293-RM1341 | 3.46 | 0.28 | -0.35 | 5.03 | Shendao3# | |
| GT | qGT5 | 5 | 3073406-27342022 | RM405-RM26 | 3.98 | -0.16 | 0.28 | 9.30 | XieqingzaoB |
| TGW | qTGW1 | 1 | 9463544-24866202 | RM3642-RM600 | 4.15 | 0.44 | 0.32 | 4.76 | Shendao3# |
| qTGW4 | 4 | 13059580-24116775 | RM7051-RM5633 | 4.43 | 0.38 | 0.57 | 5.08 | Shendao3# |
Table 4.
Pleiotropic QTLs underlying the yield-related traits in perennial Chinese rice "Shendao3#".
Table 4.
Pleiotropic QTLs underlying the yield-related traits in perennial Chinese rice "Shendao3#".
| Traits | QTL | Chromosome | Marker interval | LOD value | Additive | Dominant | R2 (%) |
|---|---|---|---|---|---|---|---|
| SP | qSP1 | 1 | RM265-RM3738 | 5.52 | 48.12 | 33.14 | 26.56 |
| GSD | qGSD1 | 1 | RM265-RM3738 | 4.53 | 0.47 | 1.82 | 14.11 |
| PP | qPP1 | 1 | RM3642-RM600 | 3.06 | -3.02 | -4.80 | 8.06 |
| PL | qPL1 | 1 | RM3642-RM600 | 3.00 | 6.05 | 4.19 | 6.07 |
| GSR | qGSR4 | 4 | RM7051-RM5633 | 3.88 | 0.02 | -0.32 | 4.61 |
| TGW | qTGW4 | 4 | RM7051-RM5633 | 4.43 | 0.38 | 0.57 | 5.08 |
| PP | qPP7 | 7 | RM1135-RM5793 | 5.23 | -3.58 | -6.52 | 10.82 |
| GYP | qGYP7 | 7 | RM1135-RM5793 | 3.32 | -14.32 | -23.54 | 7.25 |
| EGP | qEGP8 | 8 | RM1111-RM3702 | 3.10 | 49.48 | -132.62 | 14.32 |
| GSR | qGSR8 | 8 | RM1111-RM3702 | 5.90 | -0.17 | 0.46 | 5.40 |
| PP | qPP11 | 11 | RM7120-RM6293 | 4.28 | -3.04 | -5.09 | 8.53 |
| GW | qGW11a | 11 | RM7120-RM6293 | 3.82 | 0.27 | -0.35 | 5.34 |
Table 5.
Epistatic QTL underlying the yield-related traits in perennial Chinese rice "Shendao3#".
| Traits | Chr a | Marker interval | Chr a | Marker interval | LOD value | Add b | Add | Dom c | Dom | Add×Add | Add×Dom | Dom×Add | Dom×Dom | R2 (%) d |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PH | 1 | RM3738-RM8084 | 11 | RM202-RM7120 | 5.01 | 4.82 | -8.37 | 0.49 | -28.13 | 8.08 | -21.17 | -5.63 | 30.51 | 19.3 |
| PNP | 1 | RM3642-RM600 | 2 | RM6519-RM5651 | 7.08 | -1.97 | 1.90 | -3.21 | 1.56 | -3.38 | 3.30 | 0.42 | -0.59 | 2.21 |
| 3 | RM3513-RM1352 | 5 | RM405-RM26 | 5.03 | 0.91 | -1.03 | 3.50 | 1.48 | -0.7 | -2.76 | 5.63 | -7.37 | 1.49 | |
| 3 | RM85-RM3856 | 10 | RM5708-RM3882 | 6.40 | -0.34 | 1.58 | 4.55 | 0.98 | -1.78 | 2.01 | 2.20 | -8.64 | 1.98 | |
| 7 | RM3555-RM5481 | 7 | RM1135-RM5793 | 6.13 | -3.07 | -1.71 | -2.01 | -6.46 | 0.69 | 2.89 | 1.55 | 4.18 | 2.19 | |
| 7 | RM5793-RM432 | 8 | RM1111-RM3702 | 5.08 | 0.41 | 0.36 | 0.67 | 1.55 | 0.39 | -5.50 | 4.95 | -6.19 | 1.61 | |
| 7 | RM1135-RM5793 | 11 | RM202-RM7120 | 6.84 | 0.55 | -2.71 | -1.06 | 3.88 | 0.89 | -4.19 | 3.52 | -7.08 | 2.2 | |
| PL | 1 | RM3642-RM600 | 8 | RM8019-RM6990 | 5.48 | -4.58 | -3.57 | -2.01 | -2.00 | -2.56 | 6.12 | 1.03 | 7.14 | 6.28 |
| 2 | RM250-RM3763 | 3 | RM1352-RM3199 | 5.10 | -5.30 | 0.77 | 4.32 | 6.97 | 1.58 | 5.93 | -1.27 | -9.50 | 4.94 | |
| 2 | RM7637-RM5812 | 4 | RM7051-RM5633 | 5.49 | 0.91 | 0.34 | 3.40 | 3.33 | -2.59 | -3.69 | -0.77 | -5.13 | 5.13 | |
| EGP | 8 | RM1111-RM3702 | 9 | RM257-RM5661 | 5.11 | 7.29 | -9.84 | -88.52 | -27.85 | -52.25 | -51.59 | 45.48 | -29.79 | 1.74 |
| GSR | 4 | RM317-RM7051 | 4 | RM7051-RM5633 | 5.94 | -0.01 | 0.07 | 0.07 | -0.10 | 0.04 | -0.36 | 0.09 | -0.14 | 1.53 |
| 4 | RM5633-RM401 | 8 | RM1111-RM3702 | 7.42 | 0.07 | 0.00 | -0.33 | 0.21 | -0.006 | -0.04 | -0.15 | 0.34 | 1.45 | |
| 4 | RM7051-RM5633 | 9 | RM24085-RM160 | 5.69 | 0.00 | 0.02 | -0.41 | -0.13 | 0.02 | 0.13 | -0.10 | 0.56 | 1.24 | |
| 4 | RM317-RM7051 | 10 | RM5708-RM3882 | 7.22 | 0.12 | 0.15 | 0.09 | 0.12 | -0.22 | -0.03 | 0.02 | -0.38 | 1.49 | |
| 4 | RM559-RM5979 | 11 | RM7120-RM6293 | 6.72 | -0.03 | 0.02 | 0.00 | -0.24 | 0.24 | -0.22 | -0.12 | 0.47 | 1.54 | |
| 8 | RM1111-RM3702 | 9 | RM24085-RM160 | 5.32 | -0.04 | 0.16 | -0.12 | -0.09 | 0.04 | 0.16 | -0.43 | 0.34 | 1.30 | |
| 8 | RM1111-RM3702 | 10 | RM5708-RM3882 | 7.95 | -0.06 | 0.20 | 0.28 | -0.01 | 0.18 | 0.18 | -0.17 | -0.01 | 1.50 | |
| 8 | RM1111-RM3702 | 11 | RM6293-RM1341 | 6.80 | -0.05 | 0.16 | 0.26 | -0.06 | 0.19 | 0.12 | -0.13 | 0.07 | 1.52 | |
| GYP | 7 | RM3555-RM5481 | 7 | RM1135-RM5793 | 7.01 | 14.80 | -1.74 | 17.34 | -5.76 | 0.78 | -25.40 | 21.76 | -22.25 | 2.28 |
| 7 | RM1135-RM5793 | 8 | RM1111-RM3702 | 5.68 | 1.06 | -18.79 | -18.91 | -8.21 | -8.94 | 8.78 | 8.10 | 6.61 | 2.52 | |
| 7 | RM1135-RM5793 | 11 | RM202-RM7120 | 5.87 | -2.54 | -3.60 | -3.50 | 12.75 | -12.47 | 24.77 | 7.82 | -26.83 | 2.28 | |
| 9 | RM24085-RM160 | 11 | RM7120-RM6293 | 5.31 | 9.01 | -13.95 | 11.16 | -14.50 | -12.05 | -4.57 | 10.83 | -2.13 | 2.36 | |
| 10 | RM5708-RM3882 | 11 | RM202-RM7120 | 5.40 | 7.41 | -13.27 | 1.72 | 15.35 | -2.44 | 18.95 | 20.25 | -31.15 | 2.94 | |
| GW | 2 | RM250-RM3763 | 4 | RM559-RM5979 | 6.25 | 0.12 | -0.01 | 0.46 | 0.28 | 0.18 | -0.003 | 0.34 | -0.79 | 2.49 |
| GT | 5 | RM405-RM26 | 11 | RM202-RM7120 | 8.74 | -0.04 | 0.11 | 0.17 | -0.25 | 0.15 | 0.18 | -0.10 | 0.26 | 2.65 |
a Chromosome. b Additive effect. c Dominant effect. d R2 the total phenotypic variation explained by single epistatic QTL pairs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



