
Submitted:
05 September 2024
Posted:
06 September 2024
You are already at the latest version
Abstract
Subcortical beetles interact with a wide range of semiochemicals released from different sources, including trees, fungi, and insect pheromones. While the attraction of bark beetles, their insect predators, and competitors to bark beetle pheromones is well-studied, the attraction of these beetle communities to other sources of semiochemicals remains poorly understood. We tested the attraction of bark and wood-boring beetles and their predators to host stress volatiles, fungal volatiles, and a mountain pine beetle lure in the field. Host stress volatiles were derived from lodgepole pine trees stressed by three fungal symbionts of mountain pine beetle and two common phytopathogens. Our results showed that bark beetles, particularly mountain pine beetles, show a preference for a combination of fungal volatiles, particularly 2-methyl-1-butanol and mountain pine beetle lures. Without the addition of lures, 2-methyl-1-butanol was also identified as a key fungal volatile in the attraction of mountain pine beetle competitors from the Cerambycidae and Buprestidae families. Predators in the Elateridae and Staphylinidae families showed attraction to host stress volatiles and the healthy tree volatile profile. These findings suggest that these chemicals warrant further field testing for potential use in monitoring and management of subcortical beetle populations.
Keywords:
Ophiostoma montium
; Leptographium longiclavatum
; Grosmannia clavigera
; Atropellis piniphila
; Endocronartium harknessii
; Cerambycidae
; Buprestidae
; Elateridae
; Staphylinidae
; VOCs
; forest health
1. Introduction
Subcortical coleopteran beetles such as bark beetles (Coleoptera: Curculionidae, Scolytinae), long-horn (Cerambycidae) and short-horn (Buprestidae) beetles, and predators (Elateridae and Staphylinidae) develop and feed within the subcortical tissues (phloem and sapwood) of host trees. To locate suitable habitats, these beetles interact with a wide range of behaviour-modifying semiochemicals, including volatiles released by host and non-host trees, fungi, and bark beetle pheromones [1,2,3]. However, these interactions are complex as there are multiple sources of semiochemicals, each showing its own unique properties [1,2,4,5,6]. We currently have a little understanding of the semiochemical landscape governing the attraction of this habitat-sharing community of beetles.
Semiochemicals are known to be effective and eco-friendly tools that have been employed for over 50 years to monitor forest pest insect populations and gain a better understanding of insect diversity [6,7,8,9]. Species-specific pheromones are commonly used as monitoring tools for economically important bark beetle species [1,6,10]. Competitors, predators, and other bark beetle species that share the same subcortical habitats can also use bark beetle pheromones as cues to locate prey or host trees [10,11,12,13]. Furthermore, host trees stressed by abiotic and biotic factors release different compositions of volatile chemicals from healthy trees [14,15,16] these volatiles can act as attractants, repellents, or synergistic to bark beetle pheromones [2,17,18]. More recently, studies have explored the volatile compounds emitted by the fungal associates of bark beetles as an additional source of semiochemicals influencing the beetle communities [19,20,21,22]. In contrast to the bark beetle pheromones, our understanding of the attraction of subcortical beetles to host stress volatiles and fungal volatiles is lacking.
Subcortical beetle interactions with their preferred semiochemicals differ between species. Table 1 summarizes the known attractants for various subcortical beetle communities. In general, ethanol and some monoterpenes released by host trees have been found to be attractive to some cerambycid species [23]. Traps combining tree stress volatiles have been found to be the most effective for most buprestid species [24,25]. To our knowledge, no long-range pheromones have been identified for buprestids. Predators of bark and wood-boring beetles, such as species from the Elateridae and Staphylinidae families, are known to use bark beetle pheromones to locate host trees under attack by bark beetles [12]. For example, pheromones produced by several species of Ips; ipsenol and ipsdienol, are known to attract several wood-boring and predatory beetle species [26]. However, most bark beetle predators are habitat specialists– they predate on several species of beetles in the same subcortical habitat– and respond to a wide range of semiochemicals [12,27]. For instance, host volatiles are also known to be sources of attraction of bark beetle predators and can alter predator response to bark beetle pheromones [28,29].
We investigated the subcortical semiochemical landscape in the mountain pine beetle [MPB, Dendroctonus ponderosae (Hopkins)] system. The MPB is an eruptive species that is native to conifer forests in western North America [68,69,70,71]. For MPB, host tree colonization is primarily driven by the release of its aggregation pheromones, trans-verbenol and exo-brevicomin. Host tree volatiles, terpinolene and myrcene, are known to improve MPB attraction to its pheromones [33].
A large diversity of MPB competitors and predators exist throughout MPB ranges. Major insect predators of MPB include species from the families Cleridae, Trogossitidae, Staphylinidae, Histeridae, Elateridae, and Dolichopodidae. In particular, staphylinid and clerid beetles are important predators due to their ability to predate on multiple life stages of MPB [69,72,73]. Competitors include other bark beetle and wood-boring beetle species, such as species from Buprestidae and Cerambycidae families and Scolytinae subfamily [73,74].
In order to successfully survive and reproduce within their selected host, MPBs depend on the presence of symbiotic fungi, primarily Grosmannia clavigera, Ophiostoma montium, and Leptographium longiclavatum. These fungi help beetles overwhelm host defenses during host colonization and also provide essential nutrients such as nitrogen to immature beetles post-host colonization [75,76,77,78,79]. Different concentrations and compositions of fungal volatile organic compounds (FVOCs) are released by each fungal species [80]. These FVOCs could be particularly important in host location by subcortical beetle communities [19,80]. For instance, a blend of volatiles released by ophiostomatoid fungi increased the attraction of Ips typographus to its aggregation pheromones in field tests [21].
In the MPB system, Zaman et al. (2023b) [22] have characterized the chemical volatile profiles of the three main symbiotic fungi of MPB and identified the most abundant FVOCs released by each species. The FVOCs produced by all three fungi were similar and included 2-methyl-2-butanol, acetoin, isobutanol, 3-methyl-1-butanol, and 2-methyl-1-butanol. However, the concentrations of these chemicals varied; i.e., L. longiclavatum had the highest concentrations of isobutanol, 3-methyl-1-butanol, 2-methyl-1-butanol, and acetoin. In olfactometer bioassays, Zaman et al (2023b) [22] also showed MPB attraction to the volatiles emitted by G. clavigera and O. montium. However, the effectiveness of these compounds as attractants for MPB and other subcortical beetles has not yet been tested in the field.
Host stress volatile compounds (SVCs) are another important source of semiochemicals for subcortical beetle communities [30]. These volatile chemicals are released by trees in response to the presence of biotic and abiotic stressors [15]. The two most common groups of SVCs are green leaf volatiles (alcohols, acetates, and aldehydes) and volatile monoterpenoids and sesquiterpenoids [81]. Zaman et al. (2023a) [16] characterized the stress volatile profiles of lodgepole pine (Pinus contorta Dougl. ex Loud. var. latifolia Engelm) trees inoculated with three species of MPB symbiotic fungi and two common phytopathogens, Atropellis piniphila and Endocronartium harknessii. The most abundant monoterpenoids were identified as β-phellandrene, 3-carene, α-pinene, and limonene. Terpene concentrations varied among the three symbiont fungal species; however, the attraction of subcortical beetles, including MPBs to these SVC profiles is not known.
Hence, the primary objective of this study is to examine how SVCs and FVOCs associated with fungal symbionts impact the attraction of beetles colonizing the same subcortical habitat as MPB. Our objectives were to determine whether: (1) individual FVOCs are attractive to subcortical beetle communities, (2) FVOCs synergize the attraction of MPB to its pheromones, and (3) SVCs associated with symbiotic and other phytopathogenic fungi attract subcortical beetle communities. Gaining a better understanding of the interactions between these beetles and their semiochemical landscape will allow managers and researchers to better understand subcortical beetle community dynamics, monitor population trends, and implement timely control measures to mitigate bark beetle populations.
2. Materials and Methods
2.1. Experimental Design
Three experiments were conducted simultaneously. In the first experiment, we tested the effects of FVOCs alone on subcortical beetle attraction. In our second experiment, we tested the potential effects of FVOCs on the MPB pheromones. In the third experiment, we tested the effects of SVCs alone on subcortical beetle attraction. All three experiments were conducted in three blocks in two sites approximately located 20 km south of Cranbrook (British Columbia, Canada) (49°23'42.2"N 115°34'27.6"W and 49°23'33.2"N 115°34'59.7"W) in 2023. All sites were located in open areas surrounded by lodgepole pine forests, which were experiencing low-density MPB attacks during the experiment. A randomized block design was used with a 10 m distance between traps within a block and blocks at least 250 m apart (Figure 1a). Traps at the first site included dispensers containing FVOCs with MPB lure. These traps were placed in a location far away from the other two experiments to avoid possible spillover effects from the use of MPB lure. Traps at our second site tested the effects of FVOCs alone or SVCs alone in two different blocks (Figure 1b).
In all experiments, traps were suspended from 2 m-long conduits (Figure 1d). Flight intercept traps were used for the FVOCs alone or with the MPB lure, and Lindgren traps for the SVC experiment. Insecticide strips were placed inside the collection cups for dry-cup collections. Volatile chemicals were combined with a mineral oil solvent in 15ml, thin-walled, all polyethylene release bottles and hung on the inside of traps (Figure 1c). Mineral oil was used as a solvent since it is not volatile, does not react with the volatile chemicals or the plastic dispensers used, and allows for a controlled release of volatiles. All the volatile chemicals used in this experiment were purchased from commercial vendors (Table 2). The concentrations of FVOCs and SVCs used in our dispensers were based on the concentrations reported by Zaman et al. (2023a & 2023b) [16,22]. Trap collections were completed every week for five weeks during the MPB flight period, starting on July 13th and finishing on August 10th, yielding 15 replications for each treatment. Between the first and second collections, there was a large wind event that knocked down the flight intercept traps; therefore, the second collection was excluded from all data analyses. Liquids were removed from sampling cups in the field using a fine mesh filter. After each collection, the samples were stored in a -20°C freezer before identification.
2.1.1. Testing the Effects of FVOCs on the Attraction of Subcortical Beetles
The most abundant volatiles identified by Zaman et al. (2023b) [22] were used, including isobutanol, 2-methyl-2-butanol, 3-methyl-1-butanol, 2-methyl-1-butanol, and acetoin. Our treatments also included a blend of FVOCs based on the volatile profile of O. montium and a blank control (mineral oil). The O. montium volatile profile was used due to the species being dominant in hotter, drier conditions, which is characteristic of our field site locations in late spring and summer [82]. We tested seven FVOC treatments (five individual volatiles, a blend, and a control) in three blocks for a total of 21 traps. Table 2 contains treatment information and information regarding the individual chemicals used in this experiment.
2.1.2. Testing the Combined Effects of FVOCs and MPB Pheromones on Subcortical Beetle Attraction
Traps that contained FVOCs with MPB lures included the same volatile treatments (the five FVOCs and a blend treatment) with MPB lures added. We used MPB lures alone as the control. The MPB lure blend contained a mix of exo-brevicomin, trans-verbenol, terpinolene, and myrcene. We tested seven treatments in three blocks, for a total of 21 traps.
2.1.3. Testing the Effects of SVCs on the Attraction of Subcortical Beetles
The treatments included the SVC profiles associated with the three main symbiotic fungal species associated with MPB (O. montium, L. longiclavatum, and G. clavigera), the profile of a healthy tree, and the profiles of two phytopathogens infecting lodgepole pine: A. piniphila and E. harknessii. The SVC profiles of these five phytopathogens were composed of geranyl acetate, α-pinene, camphene, β-pinene, 3-carene, β-myrcene, limonene, terpinolene, bornyl acetate, γ-terpinene, camphor, and borneol mixed at different concentrations depending on the stress agent (Table 3, [16]). We tested seven treatments in three blocks, a total of 21 traps.
2.2. Statistical Analysis
All the subcortical beetles collected from our samples belonged to five Coleopteran families and one subfamily, including Staphylinidae, Elateridae, Cerambycidae, Buprestidae, and Curculionidae. Notably, all catches of Curculionidae were species from the Scolytinae subfamily and were almost entirely MPBs. R-studio (version 2023.12.0+369) was used to perform all statistical analyses. To determine the relative attractiveness of the FVOC and SVC treatments, we used the relative catches (% catch) of beetles caught in each treatment throughout collections 1, 3, 4, and 5. Due to a wind event that caused several traps to be knocked down before our second collection, collection 2 was excluded from all analyses. Key treatments for the subcortical beetle attraction were identified by comparing the relative catches from each volatile treatment to the control treatments.
To understand the temporal variation of subcortical beetles caught throughout the field season, we compared the mean number of beetles caught during our collection periods (excluding collection 2). We then conducted a Kruskal Wallis rank-based test to determine whether there were significant differences between catches in collection periods for each subcortical beetle family. The Kruskal-Wallis test was used because the data for each beetle group exhibited non-normal distributions and had similar distributions across groups. For beetle groups that were found to have significant differences between collection periods, we conducted Dunn’s test for pairwise comparisons using Bonferroni’s P-value adjustment. We performed the same analyses using Kruskal-Wallis and Dunn's tests to determine significant differences between catches of subcortical beetles between all three experiments. A non-metric multidimensional scaling (NDMS) ordination was also performed to understand correlations between the mean beetle catches of each subcortical beetle family and volatile treatment groups. Due to the non-normal distribution and zero-inflated nature of the count data, the Bray-Curtis distance measure was used for our ordinations.
3. Results
Throughout the field season, we caught 1,382 subcortical beetles, including 724 scolytids, 436 cerambycids, 96 elaterids, 92 buprestids, and 34 staphylinids (Table 4). Of the 724 scolytids caught in our traps, only 14 were not MPB.
3.1. The Effects of FVOCs, SVCs and MPB Lure on the Attraction of Subcortical Beetles
We found no statistical differences between the volatile treatments used in all three experiments for any of the subcortical beetle groups: FVOCs alone: Cerambycidae (P=0.936, df=6), Buprestidae (P=0.5975, df=6), Elateridae (P=0.855, df=6), and Staphylinidae (P=0.8627, df=6); FVOCs with pheromone lure: Scolytinae (P=0.8959, df=6), Cerambycidae (P=0.6356, df=6), Buprestidae (P=0.9873, df=6), Elateridae (P=0.565, df=6), and Staphylinidae (P=0.6133, df=6); SVCs alone: Scolytinae (P=0.9365, df=6), Cerambycidae (P=0.793, df=6), Buprestidae (P=0.4174, df=6), Elateridae (P=0.2436, df=6), and Staphylinidae (P=0.5332, df=6).
3.1.1. Catches of Subcortical Beetles in FVOC Treatments
For the two families of MPB competitors, cerambycids and buprestids, 2-methyl-1-butanol caught 21% and 36% of beetles, respectively (Figure S1 a & b). Of the total cerambycids caught, 18% were caught in the 3-methyl-1-butanol treatment compared to 11% in the control. Relatively low numbers of bark beetles were caught (<5 individuals). The isobutanol and FVOC blend each caught 22% of staphylinid beetles (Figure S1d). The control treatment caught approximately 11% of staphylinid beetles. We caught very few elaterid beetles (18 total) and the 2-methyl-1-butanol, 2-methyl-2-butanol, acetoin, the blend, and control treatments each caught 17% of the total elaterids (Figure S1e).
3.1.2. Catches of Subcortical Insects in FVOCs with MPB Lure
In traps containing both FVOCs and MPB pheromones, 21% of beetles were caught in the 2-methyl-1-butanol, 17% in the 2-methyl-2-butanol, and 11% in the control (Figure S2a). For cerambycid beetles, the percent catches of each FVOC changed with the addition of MPB lures: the acetoin (32%) and the FVOC blend (25%) accounted for over 50% of all trap catches (Figure S2b). Together, the 2-methyl-1-butanol (10%) and 3-methyl-1-butanol (4%) accounted for less than 15% of cerambycids caught. For buprestids, isobutanol caught 24% and the FVOC blend caught 18% of beetles (Figure S2c). Meanwhile, all the other treatments (including the control) each caught approximately 12% of the buprestid beetles. A quarter of all staphylinids caught were in the acetoin, and 17% were caught in the 3-methyl-1-butanol (Figure S2d). No staphylinids were caught in traps containing the FVOC blend. A quarter of elaterid beetles were caught in the 2-methyl-2-butanol treatment (Figure S2e). The 3-methyl-1-butanol, the blend, and the control treatments each caught 18% of elaterids.
3.1.3. Catches of Subcortical Insects in SVC Treatments
Very few Scolytinae were caught in the SVC treatments. Therefore, key SVC profiles in MPB attraction were difficult to identify (Figure S3a). Overall, the L. longiclavatum profile caught 33% of buprestids and 17% of cerambycids (Figure S3 b & c). Over 20% of the cerambycids caught were in the E. harknessii treatment. Approximately half of the staphylinids caught were found in the G. clavigera (24%) and healthy tree (21%) profile treatments (Figure S3d). The healthy tree treatment also caught 30% of elaterid beetles (Figure S3e).
3.2. Temporal Variation in Subcortical Beetle Catches
With the exception of Buprestidae (χ²4=5.814, P=0.213), we found significant differences between collections for all subcortical beetle families: Scolytinae (χ²4=103.09, P<0.001), Cerambycidae (χ²4=56.585, P<0.001), Elateridae (χ²4=12.981, P=0.0113), and Staphylinidae (χ²4=68.908, P<0.001). Using pooled data from all three experiments, we observed the highest mean catches of Scolytinae at the beginning (July 13) and end (August 10) of our sampling period (Figure 2a), which is likely due to two separate MPB emergence periods occurring during the field season. We caught very low numbers of MPB in our third (July 27) and fourth (August 3) collections. We observed similar trends for the other subcortical beetle families, with the exception of buprestids’ mean catches during the fourth collection being higher than the fifth collection (Figure 2b,c,d, & e). Interestingly, the temporal variation in catches of Elateridae and Scolytinae was found to be very similar. The trends of Cerambycidae and Staphylinidae catches throughout the field season were also very similar, with the lowest mean catches during the fourth collection and the highest in the fifth collection.
3.3. Differences in Subcortical Beetle Catches between Volatile Experiments
An NDMS was performed to identify whether our experiments were correlated with catches of specific subcortical beetle families (Figure 3). We found that Scolytinae catches had a strong positive correlation with the combination of FVOCs and MPB lure in traps. Additionally, Elateridae and Staphylinidae catches were both found to have a strong positive correlation with the SVC experiment. Significant differences in catches between experiments were found for several subcortical beetle families; Scolytinae (χ²2=103.09, P<0.001), Buprestidae (χ²2= 8.1877, P=0.0167) and Staphylinidae (χ²2= 17.764, P=0.0001). Scolytinae were almost entirely caught in the combination of FVOCs and MPB lure (Figure 4a). For Buprestidae, the highest mean catches were observed in the two FVOC experiments, with higher beetle attraction found when MPB lure was added to traps (Figure 4c). Catches of staphylinid beetles were higher in the SVC experiment compared to both FVOC experiments (Figure 4d).
4. Discussion
Our findings show that subcortical beetle communities interact with MPB lure, FVOCs, and SVCs differently. We identified FVOCs and MPB lure as important volatiles in MPB attraction. On the other hand, SVCs played a role in the attraction of predatory beetles. Within each of the volatile groups tested, subcortical beetles also showed differing responses to individual volatile compounds and their responses to those compounds were further altered by the MPB lure. We identified several key volatile compounds and stress volatile profiles for each of the beetle families (Table 5). However, these results require further field testing before being implemented as semiochemical tools for monitoring and management of subcortical beetles.
Catches of MPB were strongly correlated with treatments that combined FVOCs and MPB lures. We also found that FVOCs alone did not attract MPBs; we caught no MPBs in traps containing individual FVOCs or the blend of FVOCs. However, in treatments that did include MPB lures, the 2-methyl-1-butanol treatment caught 21% of MPB; while the MPB lure alone (control) treatment only caught 11% of MPB. Therefore, this compound may be an important FVOC, possibly working synergistically with MPB lure as an attractant for MPB. Similar to the FVOC alone experiment, very few MPBs were caught in our SVC experiment. However, the SVCs were not tested in combination with MPB pheromone lures, therefore, the possible synergistic effects between these two volatile groups on MPB attraction are still unknown.
Catches of predatory beetles from Elateridae and Staphylinidae were highly correlated with SVCs, suggesting that these host volatiles may play a larger role in the attraction of bark beetle predators than previously thought. Although tree volatiles, such as α-pinene, have been known to enhance predatory beetle attraction to bark beetle pheromones, research on bark beetle predator attraction has focused largely on using bark beetle pheromones in combination with other volatiles in traps [83,84]. Therefore, no work has been done investigating the bark beetle predator attraction to SVCs alone.
The attraction of both families of predatory beetles (Elateridae and Staphylinidae) was associated with the chemical profile of healthy trees. Predators are known to utilize bark beetle pheromones and host volatiles as semiochemical cues while foraging, especially host volatiles associated with the presence of their prey [10,29]. Erbilgin et al. (2007) [2] proposed that tree-killing bark beetle species, like MPBs, are attracted to healthy trees since the thicker phloem of such trees increases MPB fitness [74,85]. Therefore, it is possible that volatiles released by healthy trees may act as kairomones for predators by signaling ideal host conditions for prey species, such as MPBs. We speculate that individual monoterpenes like 3-carene within the healthy tree profile may act as an attractant for predators; coincidently, healthy trees have the highest concentrations of this monoterpene relative to the infected trees in this study. Furthermore, 3-carene has been found to be an attractant for several bark beetle species, including red turpentine beetle (Dendroctonus valens) [18,86]. Another monoterpene, α-pinene, could be responsible for the E. harknessii infected tree profile being identified as another key SVC profile for elaterid beetles. Miller (2023) [60] reported that predatory beetles of bark and wood-boring beetles were attracted to α-pinene, which is found in the highest concentration in trees infected by E. harknessii compared to other stress agents tested in the current study. The E. harknessii profile was also identified as a key SVC profile for both families of wood-boring beetles in this study. Therefore, we suspect α-pinene may be acting as an indicator of favourable hosts to wood-boring beetles, signalling their presence to predatory beetles, such as elaterids.
In treatments that did not include the addition of MPB lure, 2-methyl-1-butanol emerged as a key FVOC for the attraction of cerambycid and buprestid species caught in this study. These findings are consistent with previous studies that identified 2-methyl-1-butanol as an attractant for several wood-boring beetle species [47,48,49]. This compound was also identified as a key FVOC for MPB; however, this finding was from our experiment, which combined FVOCs with MPB lure. We observed that the addition of the MPB lure altered cerambycid attraction to 2-methyl-1-butanol, relative to their attraction to the same compound alone since it was not identified as a key FVOC once pheromone lures were included in the treatment. Buprestid and cerambycid beetles were also not attracted to the same FVOCs once MPB pheromones were added; in our experiment combining FVOCs and MPB lure, acetoin was the only key FVOC identified for cerambycids while isobutanol was the only key FVOC identified for buprestids. Interestingly, we caught higher numbers of buprestids in FVOC traps, which included MPB lure, meanwhile, no significant differences between experiments were found for cerambycids. Overall, these results highlight that wood-boring beetles interact differently with FVOCs in the presence of bark beetle pheromones and that these responses further differ depending on the family of wood-boring beetle. These changes in the attractiveness of compounds in the presence of MPB pheromones must be considered when testing semiochemical tools for these species and should be further investigated in the field.
We observed temporal variations in the catches of subcortical beetle communities. The highest catches of Scolytinae and Elateridae beetles occurred during the first collection in July, suggesting these beetles are most abundant early in the field season due to less competition for host trees. Similar trends in trap catches for both families throughout the season indicate closer interactions during host selection. A high catch of MPB during the last collection suggests two emergence periods of MPB in the 2023 field season. Higher catches of Cerambycidae and Staphylinidae beetles in the final collection indicate later summer emergence. Emergence times impact the semiochemical landscape and beetle behaviour, as beetles expend more energy flying in warmer temperatures, affecting dispersion, host colonization, and reproduction [87]. At our field sites, the hottest month was August, corresponding with our last two collection periods (August 3 and August 10), which could mean that cerambycids and staphylinids are more tolerant of hot temperatures during their adult flight period.
This study has potential limitations. First of all, our sites were located in open areas with a lot of wind exposure. Several large wind events occurred during the field season. The high winds at the sites may have also impacted the release rates and dispersal patterns of volatiles and pheromones, potentially altering beetle responses to our traps. Another potential limitation was the duration of our field season. We likely observed two separate emergence events of MPB at our field sites since our highest catches of MPB were in the first and last collections. Thus, it is likely we missed the MBP emergence at the beginning of the field season (before our first collection) and at the end of the second emergence period (after our second collection). In future experiments, it may be more suitable to extend the collection period from 5 weeks to 7 weeks, starting one week earlier and ending one week later, to ensure both populations are represented in the data.
We conclude that groups of subcortical beetle communities interact differently with various sources of volatiles. We identified 2-methyl-1-butanol as a key FVOC in synergizing MPB attraction to its commercially available lure. However, further field testing must be conducted to confirm the role of this volatile in MPB attraction. We also found that the addition of bark beetle pheromones to traps altered subcortical beetle community attraction to FVOCs, especially for wood-boring beetles. Currently, we do not know whether the addition of pheromones to SVCs would similarly alter beetle attraction to traps. Overall, our results suggest that FVOCs and SVCs could be useful semiochemical tools for the monitoring and management of subcortical beetle species but require further field testing before being used as a tool in subcortical insect monitoring. Once developed, these tools will allow us to better understand beetle community composition and interactions between different functional groups of species.
Author Contributions
Conceptualization, Leah Crandall, Rashaduz Zaman and Nadir Erbilgin ; Data curation, Leah Crandall and Rashaduz Zaman ; Formal analysis, Leah Crandall, Rashaduz Zaman and Nadir Erbilgin ; Investigation, Leah Crandall, Marnie Duthie-Holt, Wade Jarvis and Nadir Erbilgin ; Methodology, Leah Crandall, Rashaduz Zaman and Nadir Erbilgin ; Supervision, Rashaduz Zaman and Nadir Erbilgin ; Validation, Leah Crandall, Rashaduz Zaman and Nadir Erbilgin ; Visualization, Leah Crandall, Rashaduz Zaman and Nadir Erbilgin ; Writing – original draft, Leah Crandall, Rashaduz Zaman and Nadir Erbilgin ; Writing – review & editing, Leah Crandall, Rashaduz Zaman , Marnie Duthie-Holt, Wade Jarvis and Nadir Erbilgin.
Funding
We acknowledge funding for this research from the Natural Sciences and Engineering Research Council of Canada-Discovery Award to NE. This project received partial funding from SERG-International and the Alberta Conservation Association.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We are grateful to Guncha Ishangulyyeva for the techinical support on this project.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Byers, J. A. (2004). Chemical ecology of bark beetles in a complex olfactory landscape. Bark and wood boring insects in living trees in Europe, a synthesis, 89-134. [CrossRef]
- Erbilgin, N., Christiansen, E., Krokene, P. (2007). A host monoterpene influences Ips typographus responses (Coleoptera: Curculionidae, Scolytinae) to its aggregation pheromone: Implications for host colonization of bark beetles. Agricultural & Forest Entomology. 9: 135-140. [CrossRef]
- Davis, T.S., Crippen, T.L., Hofstetter, R.W., Tomberlin, J.K. (2013). Microbial volatile emissions as insect semiochemicals. Journal of Chemical Ecology, 39: 840–859. [CrossRef]
- Schlyter, F., Byers, J.A. & Löfqvist, J. (1987). Attraction to pheromone sources of different quantity, quality, and spacing: Density-regulation mechanisms in bark beetle Ips typographus. Journal of Chemical Ecology, 13: 1503–1523. [CrossRef]
- Wallin, K. F., & Raffa, K. F. (2000). Influences of host chemicals and internal physiology on the multiple steps of postlanding host acceptance behavior of Ips pini (Coleoptera: Scolytidae). Environmental Entomology, 29(3), 442-453. [CrossRef]
- Seybold, S. J., Bentz, B. J., Fettig, C. J., Lundquist, J. E., Progar, R. A., & Gillette, N. E. (2018). Management of western North American bark beetles with semiochemicals. Annual Review of Entomology, 63(1), 407-432. [CrossRef]
- Gandhi, K.J., Gilmore, D.W., Haack, R.A., Katovich, S.A., Krauth, S.J., Mattson, W.J., Zasada, J.C., Seybold, S.J. (2009). Application of semiochemicals to assess the biodiversity of subcortical insects following an ecosystem disturbance in a sub-boreal forest. Journal of Chemical Ecology, 35(12), 1384-410. [CrossRef] [PubMed]
- Afzal, S., Nahrung, H. F., Lawson, S. A., & Hayes, R. A. (2023). How effective are push–pull semiochemicals as deterrents for bark beetles? a global meta-analysis of thirty years of research. Insects, 14(10), 812. [CrossRef]
- Singh, V.V., Naseer, A., Mogilicherla, K., Trubin, A., Zabihi, K., Roy, A., Jakuš, R., Erbilgin, N. (2024). Understanding bark beetle outbreaks: exploring the impact of changing temperature regimes, droughts, forest structure, and prospects for future forest pest management. Reviews in Environmental Science Biotechnology, 23: 257–290. [CrossRef]
- Erbilgin, N., Powell, J.S., Raffa, K.S. (2003). Effect of varying monoterpene concentrations on the response of Ips pini (Coleoptera: Scolytidae) to its aggregation pheromone: implications for pest management and ecology of bark beetles. Agricultural & Forest Entomology, 5: 269-274. [CrossRef]
- Borden, J. H. (1989). Semiochemicals and bark beetle populations: exploitation of natural phenomena by pest management strategists. Ecography, 12(4), 501-510. [CrossRef]
- Erbilgin, N., Raffa, K.S. (2001a). Kairomonal range of generalist predators in specialized habitats: responses to multiple phloeophagous species emitting pheromones vs. host odors. Entomologia Experimentalis et Applicata, 99: 205-210. [CrossRef]
- Boone, C. K., Six, D. L., & Raffa, K. F. (2008). The enemy of my enemy is still my enemy: Competitors add to predator load of a tree-killing bark beetle. Agricultural and Forest Entomology, 10(4), 411-421. [CrossRef]
- Niinemets, Ü., Kännaste, A., & Copolovici, L. (2013). Quantitative patterns between plant volatile emissions induced by biotic stresses and the degree of damage. Frontiers in Plant Science, 4, 262. [CrossRef]
- Faiola, C., & Taipale, D. (2020). Impact of insect herbivory on plant stress volatile emissions from trees: A synthesis of quantitative measurements and recommendations for future research. Atmospheric Environment: X, 5. [CrossRef]
- Zaman, R., Antonioli, F., Shah, A., Ullah, A., May, C., Klutsch, J.G., Erbilgin, N. (2023a). A pine in distress: how infection by different pathogenic fungi affect lodgepole pine chemical defenses. Microbial Ecology, 86, 2666–2673. [CrossRef]
- Byers, J.A. (1992). Attraction of bark beetles, Tomicus piniperda, Hylurgops palliatus, and Trypodendron domesticum and other insects to short-chain alcohols and monoterpenes. Journal of Chemical Ecology 18: 2385–2402. [CrossRef]
- Erbilgin, N., Raffa, K.F. (2000). Opposing Effects of host monoterpenes on responses by two sympatric species of bark beetles to their aggregation pheromones. Journal of Chemical Ecology, 26: 2527–2548. [CrossRef]
- Kandasamy, D., Gershenzon, J. & Hammerbacher, A. (2016). Volatile organic compounds emitted by fungal associates of conifer bark beetles and their potential in bark beetle control. Journal of Chemical Ecology, 42, 952–969. [CrossRef]
- Kandasamy, D., Gershenzon, J., Andersson, M. N., & Hammerbacher, A. (2019). Volatile organic compounds influence the interaction of the Eurasian spruce bark beetle (Ips typographus) with its fungal symbionts. The ISME journal, 13(7); 1788-1800. [CrossRef]
- Jirošová, A., Modlinger, R., Hradecký, J., Ramakrishnan, R., Beránková, K., Kandasamy, D. (2022). Ophiostomatoid fungi synergize attraction of the Eurasian spruce bark beetle, Ips typographus to its aggregation pheromone in field traps. Frontiers in Microbiology.13:980251. [CrossRef]
- Zaman, R., May, C., Ullah, A., Erbilgin, N. (2023b). Bark beetles utilize ophiostomatoid fungi to circumvent host tree defenses. Metabolites 13(2), 239. [CrossRef]
- Miller, D.R., Asaro, C., Crowe, C.M., Duerr, D.A. (2011). Bark beetle pheromones and pine volatiles: attractant kairomone lure blend for longhorn beetles (Cerambycidae) in pine stands of the southeastern United States. Journal of Economic Entomology, 104(4), 1245–1257. [CrossRef]
- Grant, G. G., Poland, T. M., Ciaramitaro, T., Barry Lyons, D., & Jones, G. C. (2011). Comparison of male and female emerald ash borer (Coleoptera: Buprestidae) responses to phoebe oil and (Z)-3-hexenol lures in light green prism traps. Journal of Economic Entomology, 104(1), 173-179. [CrossRef]
- Domingue, M. J., Baker, T. C., Blanco, J. J., & Fernandes, A. T. (2012). A multi-disciplinary approach for developing tools to monitor invasive buprestid beetle species. Invasive species: threats, ecological impact and control methods. Nova, Hauppauge, 3, 77-100.
- Allison, J.D., McKenney, J.L., Miller, D.R., Gimmel, M.L. (2013). Kairomonal responses of natural enemies and associates of the southern Ips (Coleoptera: Curculionidae: Scolytinae) to ipsdienol, ipsenol and cis-Verbenol. Journal of Insect Behavior 26, 321–335. [CrossRef]
- Miller, D. R. (2023). Coleopteran predators of bark and woodboring beetles attracted to traps baited with ethanol and α-pinene in pine (Pinaceae) forests of the southern United States of America. The Canadian Entomologist, 155, e5. [CrossRef]
- Wood, D. L. (1982). The role of pheromones, kairomones, and allomones in the host selection and colonization behavior of bark beetles.
- Erbilgin, N., Raffa, K.S. (2001b). Modulation of predator attraction to pheromones of two prey species by stereochemistry of plant volatiles. Oecologia, 127: 444-453. [CrossRef]
- Gitau, C. W., Bashford, R., Carnegie, A. J., & Gurr, G. M. (2013). A review of semiochemicals associated with bark beetle (Coleoptera: Curculionidae: Scolytinae) pests of coniferous trees: a focus on beetle interactions with other pests and their associates. Forest Ecology and Management, 297: 1-14. [CrossRef]
- Tóth, M. (2013). Pheromones and attractants of click beetles: an overview. Journal of Pest Science, 86: 3-17. [CrossRef]
- Payne, T.L., Coster, J.E., Richerson, J.V., Edson, L.J., Hart, E.R. (1978). Field response of the southern pine beetle to behavioral chemicals. Environmental Entomology 7(4): 578-582.
- Klutsch, J.G., Cale, J.A., Whitehouse, C., Kanekar, S.S., and Erbilgin, N. (2017). Trap trees: an effective method for monitoring mountain pine beetle activities in novel habitats. Canadian Journal of Forest Research. 47(10): 1432-1437. [CrossRef]
- Borden, J.H., Hunt, D.W.A., Miller, D.R., Slessor, K.N. (1986). Orientation in forest coleoptera: an uncertain outcome to responses by individual beetles to variable stimuli. Clarendon Press, Oxford, UK.
- Payne, T.L., Andryszak, N.A., Weiser, H., Dixon, E.A., Ibrahim, N., Coers, L. (1988). Antennal olfactory and behavioral response of the southern pine beetle, Dendroctonus frontalis, to analogs of its aggregation pheromone frontalin. Journal of Chemical Ecology, 14: 1217–1225. [CrossRef]
- Sullivan, B.T., Shepherd, W.P., Pureswaran, D.S., Tashiro, T., Mori, K. (2007). Evidence that (+)-endo-brevicomin is a male-produced component of the southern pine beetle aggregation pheromone. Journal of Chemical Ecology, 33: 1510–1527. [CrossRef]
- Francke, W., Bartels, J., Meyer, H., Schröder, F., Kohnle, U., Baader, E., Pierre Vité, J. (1995). Semiochemicals from bark beetles: New results, remarks, and reflections. Journal of Chemical Ecology, 21: 1043–1063. [CrossRef]
- Coster, J.E., Vité, J.P. (1972). Effects of feeding and mating on pheromone release in the southern pine beetle. Annals of the Entomological Society of America, 65: 263–266. [CrossRef]
- McCarty, F.A., Billings, P., Richerson, J.V., Payne, T.L., Edson, L.J. (1980). Response of the southern pine beetle to behavioral chemicals in the laboratory. Journal of Entomological Science,15: 307–317.
- Vité, J.P., Bakke, A., Renwick, J.A.A. (1972). Pheromones in Ips (Coleoptera: Scolytidae): occurrence and production. The Canadian Entomologist, 104: 1967–1975. [CrossRef]
- Byers, J.A., Birgersson, G. (1990). Pheromone production in a bark beetle independent of myrcene precursor in host pine species. Naturwissenschaften, 77: 385–387. [CrossRef]
- Vité, J.P., Renwick, J.A.A. (1971). Population aggregating pheromone in the bark beetle, Ips grandicollis. Journal of Insect Physiology, 17: 1699–1704. [CrossRef]
- Byers, J.A., Lanne, B.S., Schlyter, F., Lofqvist, J., Bergstrom, G. (1985). Olfactory recognition host-tree susceptibility by pine shoot beetles. Naturwissenschaften, 72, 324–326. [CrossRef]
- Hofstetter, R. W., Chen, Z., Gaylord, M. L., McMillin, J. D., & Wagner, M. R. (2008). Synergistic effects of α-pinene and exo-brevicomin on pine bark beetles and associated insects in Arizona. Journal of Applied Entomology, 132(5): 387-397.
- Miller, D. R. (2006). Ethanol and (−)-α-pinene: attractant kairomones for some large wood-boring beetles in southeastern USA. Journal of Chemical Ecology, 32: 779-794. [CrossRef]
- Andersson, M. N., Larsson, M. C., & Schlyter, F. (2009). Specificity and redundancy in the olfactory system of the bark beetle Ips typographus: single-cell responses to ecologically relevant odors. Journal of Insect Physiology, 55(6); 556-567.
- Hanks, L.M., Millar, J.G., Moreira, J.A., Barbour, J.D., Lacey, E.S., McElfresh, S., Reuter, F.R., Ray, A.M. (2007). Using generic pheromone lures to expedite identification of aggregation pheromones for the cerambycid beetles Xylotrechus nauticus, Phymatodes lecontei, and Neoclytus modestus modestus. Journal of Chemical Ecology, 33, 889–907. [CrossRef]
- Mitchell, R. F., Hughes, D. T., Luetje, C. W., Millar, J. G., Soriano-Agatón, F., Hanks, L. M., & Robertson, H. M. (2012). Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect biochemistry and molecular biology, 42(7), 499-505. [CrossRef]
- Molander, M.A., Winde, I.B., Burman, J., Nyabuga, F.N., Lindblom, T.U.T., Hanks, L.M., Millar, J.G., Larsson, M.C. (2019). Common cerambycid pheromone components as attractants for longhorn beetles (Cerambycidae) breeding in ephemeral oak substrates in northern Europe. Journal of Chemical Ecology, 45, 537–548. [CrossRef]
- Pajares, J. A., Álvarez, G., Ibeas, F., Gallego, D., Hall, D. R., & Farman, D. I. (2010). Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. Journal of Chemical Ecology, 36: 570-583. [CrossRef]
- Teale, S. A., Wickham, J. D., Zhang, F., Su, J., Chen, Y., Xiao, W., Hanks, L.M. & Millar, J. G. (2011). A male-produced aggregation pheromone of Monochamus alternatus (Coleoptera: Cerambycidae), a major vector of pine wood nematode. Journal of Economic Entomology, 104(5): 1592-1598. [CrossRef]
- Miller, D.R., Allison, J.D., Crowe, C.M., Dickinson, D.M., Eglitis, A., Hofstetter, R.W., Munson, A.S., Poland, T.M., Reid, L.S., Steed, B.E., Sweeney, J.D. (2016). Pine sawyers (Coleoptera: Cerambycidae) attracted to α-Pinene, monochamol, and ipsenol in North America. Journal of Economic Entomology, 109(3): 1205–1214. [CrossRef]
- Schröder, F., Fettköther, R., Noldt, U., Dettner, K., König, W.A., Francke, W. (1994). Synthesis of (3R)-3-hydroxy-2-hexanone, (2R,3R)-2,3-hexanediol and (2S,3R)-2,3-hexanediol, the male sex pheromone of Hylotrupes bajulus and Pyrrhidium sanguineum (Cerambycidae). Liebigs Annalen der Chemie: 1211–1218. [CrossRef]
- Miller, D. R., Dodds, K. J., Eglitis, A., Fettig, C. J., Hofstetter, R. W., Langor, D. W., Mayfield, A. E., Munson, A. S., Poland, T. M., Raffa, K. F. (2013). Trap lure blend of pine volatiles and bark beetle pheromones for Monochamus spp. (Coleoptera: Cerambycidae) in pine forests of Canada and the United States. Journal of Economic Entomology, 106 : 1684 – 1692.
- Silk, P. J., Sweeney, J., Wu, J., Price, J., Gutowski, J. M., & Kettela, E. G. (2007). Evidence for a male-produced pheromone in Tetropium fuscum (F.) and Tetropium cinnamopterum (Kirby)(Coleoptera: Cerambycidae). Naturwissenschaften, 94:697-701. [CrossRef]
- Sweeney, J. D., Silk, P. J., Gutowski, J. M., Wu, J., Lemay, M. A., Mayo, P. D., & Magee, D. I. (2010). Effect of chirality, release rate, and host volatiles on response of Tetropium fuscum (F.), Tetropium cinnamopterum Kirby, and Tetropium castaneum (L.) to the aggregation pheromone, fuscumol. Journal of Chemical Ecology, 36: 1309-1321. [CrossRef]
- Lacey, E. S., Ginzel, M. D., Millar, J. G., & Hanks, L. M. (2004). Male-produced aggregation pheromone of the cerambycid beetle Neoclytus acuminatus acuminatus. Journal of Chemical Ecology, 30: 1493-1507.
- Allison, J. D., Borden, J. H., McIntosh, R. L., De Groot, P., & Gries, R. (2001). Kairomonal response by four Monochamus species (Coleoptera: Cerambycidae) to bark beetle pheromones. Journal of Chemical Ecology, 27: 633-646. [CrossRef]
- Morewood, W. D., Simmonds, K. E., Wilson, I. M., Borden, J. H., & McIntosh, R. L. (2002). α-Pinene and ethanol: key host volatiles for Xylotrechus longitarsis (Coleoptera: Cerambycidae). Journal of the Entomological Society of British Columbia, 99: 117-122.
- Miller, D. R., & Asaro, C. (2023). Predators attracted to combination of bark beetle pheromones and host kairomones in pine forests of southeastern United States. Environmental Entomology, 52(5): 787-794. [CrossRef]
- Peschke, K., Friedrich, P., Kaiser, U., Franke, S., & Francke, W. (1999). Isopropyl (Z9)-hexadecenoate as a male attractant pheromone from the sternal gland of the rove beetle Aleochara curtala (Coleoptera: Staphylinidae). Chemoecology, 9: 47-54. [CrossRef]
- Avşar, İ., & Turan, Y. (2024). An overview of the pheromones of staphylinidae (Coleoptera). Transactions of the American Entomological Society, 150(2): 205-215.
- Faly, L., Brygadyrenko, V., & Paulauskas, A. (2024). Repellent and attractant activities of organic compounds on female and male Philonthus decorus (Coleoptera, Staphylinidae). Biology, 13(5), 294. [CrossRef]
- Tolasch, T., Von Fragstein, M., & Steidle, J. L. (2010). Sex pheromone of Agriotes acuminatus (Stephens, 1830)(Coleoptera: Elateridae). Journal of Chemical Ecology, 36: 314-318. [CrossRef]
- Tóth, M., Furlan, L., Yatsynin, V. G., Ujváry, I., Szarukán, I., Imrei, Z., Subchev, M., Tolasch, T. & Francke, W. (2002). Identification of sex pheromone composition of click beetle Agriotes brevis candeze. Journal of chemical ecology, 28: 1641-1652. [CrossRef]
- Toth, M., Furlan, L., Yatsynin, V. G., Ujvary, I., Szarukan, I., Imrei, Z., Tolasch, T., Francke, W. & Jossi, W. (2003). Identification of pheromones and optimization of bait composition for click beetle pests (Coleoptera: Elateridae) in Central and Western Europe. Pest Management Science, 59(4): 417-425. [CrossRef]
- Tolasch, T., von Fragstein, M., & Steidle, J. L. (2007). Sex pheromone of Elater ferrugineus L.(Coleoptera: Elateridae). Journal of chemical ecology, 33: 2156-2166. [CrossRef]
- Amman, G. D. (1977). The role of the mountain pine beetle in lodgepole pine ecosystems: impact on succession. In The role of arthropods in forest ecosystems (pp. 3-18). Berlin, Heidelberg: Springer Berlin Heidelberg.
- Safranyik, L. and Carroll, A.L. (2006). The biology and epidemiology of the mountain pine beetle in lodgepole pine forests. The Mountain Pine Beetle: A Synthesis of its Biology, Management and Impacts on Lodgepole Pine. 3-66. Natural Resources Canada.
- Cudmore, T. J., Björklund, N., Carroll, A. L., & Staffan Lindgren, B. (2010). Climate change and range expansion of an aggressive bark beetle: evidence of higher beetle reproduction in naïve host tree populations. Journal of Applied Ecology, 47(5), 1036-1043. [CrossRef]
- Sambaraju, K. R., & Goodsman, D. W. (2021). Mountain pine beetle: an example of a climate-driven eruptive insect impacting conifer forest ecosystems. CABI Reviews, (2021). [CrossRef]
- Amman, G. D. (1970). Prey consumption and variations in larval biology of Enoclerus spegeus (Coleoptera: Cleridae). The Canadian Entomologist, 102(11), 1374–1379. [CrossRef]
- Krause, A.M., Townsend, P.A., Lee, Y., Raffa, K.F. (2018). Predators and competitors of the mountain pine beetle Dendroctonus ponderosae (Coleoptera: Curculionidae) in stands of changing forest composition associated with elevation. Agricultural and Forest Entomology, 20.3, 402-413. [CrossRef]
- Carroll, A. L., Aukema, B. H., Raffa, K. F., Linton, D. A., Smith, G. D., & Lindgren, B. S. (2006). Mountain pine beetle outbreak development: the endemic—incipient epidemic transition. Canadian Forest Service, Mountain Pine Beetle Initiative Project, 1, 22.
- Bleiker, K.P., Six, D.L. (2007). Dietary benefits of fungal associates to an eruptive herbivore: potential implications of multiple associates on host population dynamics. Environmental Entomology, 36(6):1384-96. [CrossRef] [PubMed]
- Goodsman, D.W., Erbilgin, N., Lieffers, V.J. (2012). The impact of phloem nutrients on overwintering mountain pine beetles and their fungal symbionts. Environmental Entomology, 41(3), 478–486. [CrossRef]
- Therrien, J., Mason, C.J., Cale, J.A. et al. (2015). Bacteria influence mountain pine beetle brood development through interactions with symbiotic and antagonistic fungi: implications for climate-driven host range expansion. Oecologia 179, 467–485. [CrossRef]
- Cale, J.A., Muskens, M., Najar, A., Ishangulyyeva, G., Hussain, A., Kanekar, S.S., Klutsch, J.G., Taft, S., Erbilgin, N. (2017). Rapid monoterpene induction promotes the susceptibility of a novel host pine to mountain pine beetle colonization but not to beetle-vectored fungi, Tree Physiology 37(12), 1597–1610. [CrossRef]
- Guevara-Rozo, S., Hussain, A., Cale, J.A., Klutsch, J.G., Rajabzadeh, R., Erbilgin, N. (2020). Nitrogen and ergosterol concentration varied in live jack pine phloem following inoculations with fungal associates of mountain pine beetle. Frontiers in Microbiology 11(1703). [CrossRef]
- Cale, J.A., Collignon, R.M., Klutsch, J.G., Kanekar, S.S., Hussain, A., Erbilgin, N. (2016) Fungal volatiles can act as carbon sources and semiochemicals to mediate interspecific interactions among bark beetle-associated fungal symbionts. PLoS ONE 11(9): e0162197. [CrossRef]
- Chiu, C. & Bohlmann, J. (2022). Mountain pine beetle epidemic: an interplay of terpenoids in host defense and insect pheromones. Annual Review of Plant Biology, 73:1, 475-494. [CrossRef]
- Moore, M.L., Six, D.L. (2015). Effects of Temperature on growth, sporulation, and competition of mountain pine beetle fungal symbionts. Microbial Ecology 70, 336–347. [CrossRef]
- Aukema, B.H., Dahlsten, D.L., Raffa, K.F. (2000). Improved population monitoring of bark beetles and predators by incorporating disparate behavioral responses to semiochemicals. Environmental Entomology, 29(3), 618–629. [CrossRef]
- Hofstetter, R. W., Gaylord, M. L., Martinson, S., & Wagner, M. R. (2012). Attraction to monoterpenes and beetle-produced compounds by syntopic Ips and Dendroctonus bark beetles and their predators. Agricultural and Forest Entomology, 14(2), 207-215. [CrossRef]
- Amman, G. D. (1972). Mountain pine beetle brood production in relation to thickness of lodgepole pine phloem. Journal of Economic Entomology, 65(1), 138-140.
- Erbilgin, N., Ma, C., Whitehouse, C., Shan, B., Najar, A., & Evenden, M. (2014). Chemical similarity between historical and novel host plants promotes range and host expansion of the mountain pine beetle in a naïve host ecosystem. The New Phytologist, 201(3), 940–950. [CrossRef]
- Wijerathna, A., Evenden, M. (2020). Effect of environmental conditions on flight capacity in mountain pine beetle (Coleoptera: Curculionidae: Scolytinae). Journal of Insect Behaviour, 33; 201–215. [CrossRef]
Figure 1.
Experimental field site and trap setup. B1, B2, and B3 represent the three blocks of traps for each experiment, and arrows indicate the direction of trap rotation after each collection. (a) Trap locations at field site 1; fungal volatiles (FVOCs) combined with pheromone experiment. (b) Location of sites within British Columbia. (c) Trap locations at field site 2; FVOC alone treatments (stars) and host stress volatiles (SVCs) alone treatments (circles) were placed at this site. (d) Beetle traps used in our experiments: a flight intercept trap, used in our FVOC experiments (left) and a Lindgren funnel trap, used for the SVC experiment (right).
Figure 1.
Experimental field site and trap setup. B1, B2, and B3 represent the three blocks of traps for each experiment, and arrows indicate the direction of trap rotation after each collection. (a) Trap locations at field site 1; fungal volatiles (FVOCs) combined with pheromone experiment. (b) Location of sites within British Columbia. (c) Trap locations at field site 2; FVOC alone treatments (stars) and host stress volatiles (SVCs) alone treatments (circles) were placed at this site. (d) Beetle traps used in our experiments: a flight intercept trap, used in our FVOC experiments (left) and a Lindgren funnel trap, used for the SVC experiment (right).

Figure 2.
Mean catches of each subcortical beetle group throughout the collection period. Mean catches were calculated using data from all three experiments. Traps were collected every week for 5 weeks from July 13 (Collection 1) to August 10 (Collection 5). Error bars represent standard error. Kruskal-Wallis tests were run to test for significant differences between collections at P<0.05. Catches from collection 2 are excluded due to the loss of traps during an extreme wind event between collections. Collections with the same letter within each beetle group are not significantly different; Dunn’s test with Bonferroni’s adjustment at P<0.05).
Figure 2.
Mean catches of each subcortical beetle group throughout the collection period. Mean catches were calculated using data from all three experiments. Traps were collected every week for 5 weeks from July 13 (Collection 1) to August 10 (Collection 5). Error bars represent standard error. Kruskal-Wallis tests were run to test for significant differences between collections at P<0.05. Catches from collection 2 are excluded due to the loss of traps during an extreme wind event between collections. Collections with the same letter within each beetle group are not significantly different; Dunn’s test with Bonferroni’s adjustment at P<0.05).
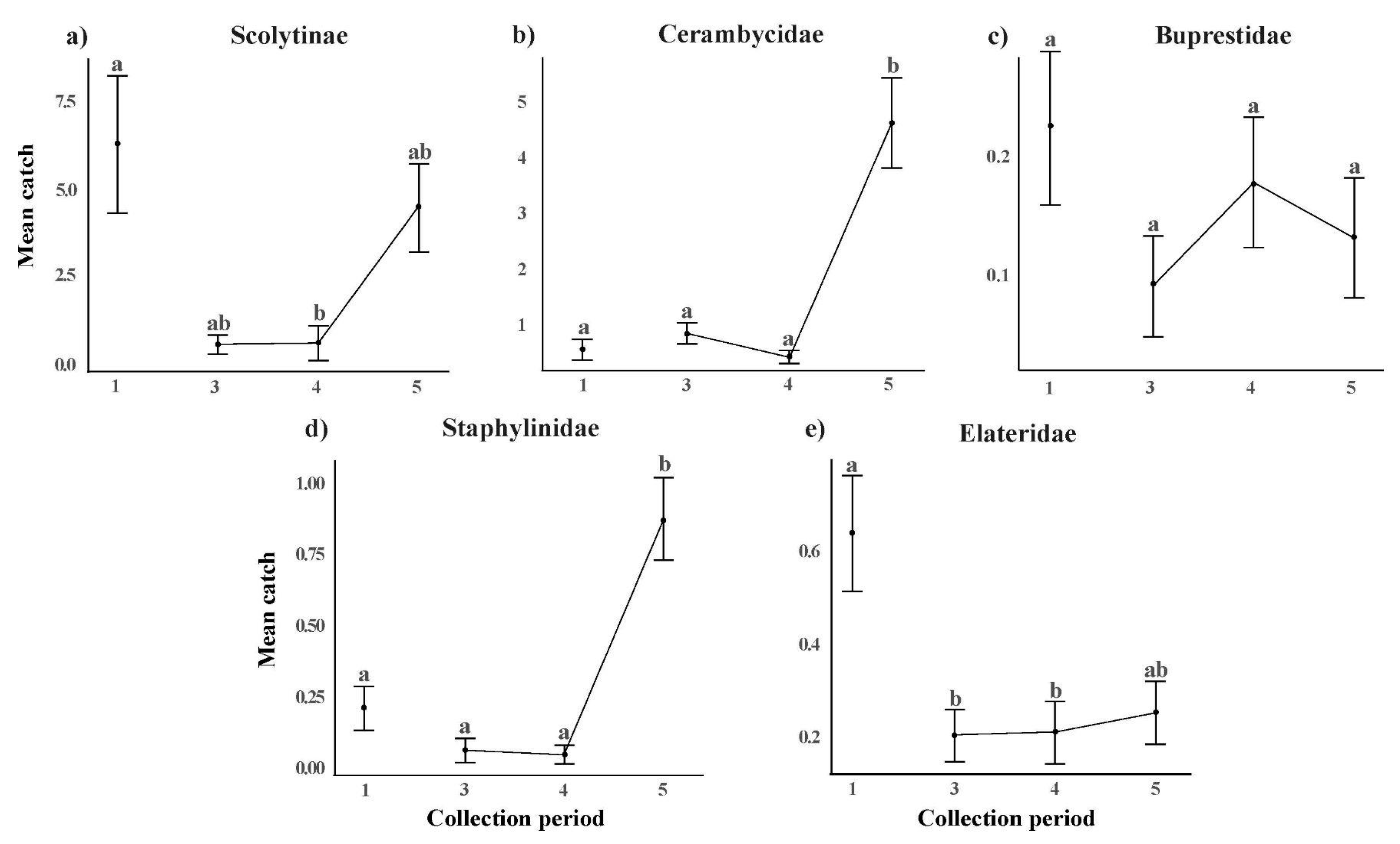
Figure 3.
Effects of different volatile treatment types on catches of subcortical beetle families. Mean beetle catches over the entire field season were analyzed using an NMDS gradient analysis. Bray-Curtis was used as the distance metric. The vectors (black arrows) correspond to our three volatile experiments: (1) Fungal volatiles alone (FVOC), (2) Fungal volatiles combined with MPB pheromones (FVOC+ Pheromone), and (3) Host stress volatiles alone (SVC).
Figure 3.
Effects of different volatile treatment types on catches of subcortical beetle families. Mean beetle catches over the entire field season were analyzed using an NMDS gradient analysis. Bray-Curtis was used as the distance metric. The vectors (black arrows) correspond to our three volatile experiments: (1) Fungal volatiles alone (FVOC), (2) Fungal volatiles combined with MPB pheromones (FVOC+ Pheromone), and (3) Host stress volatiles alone (SVC).
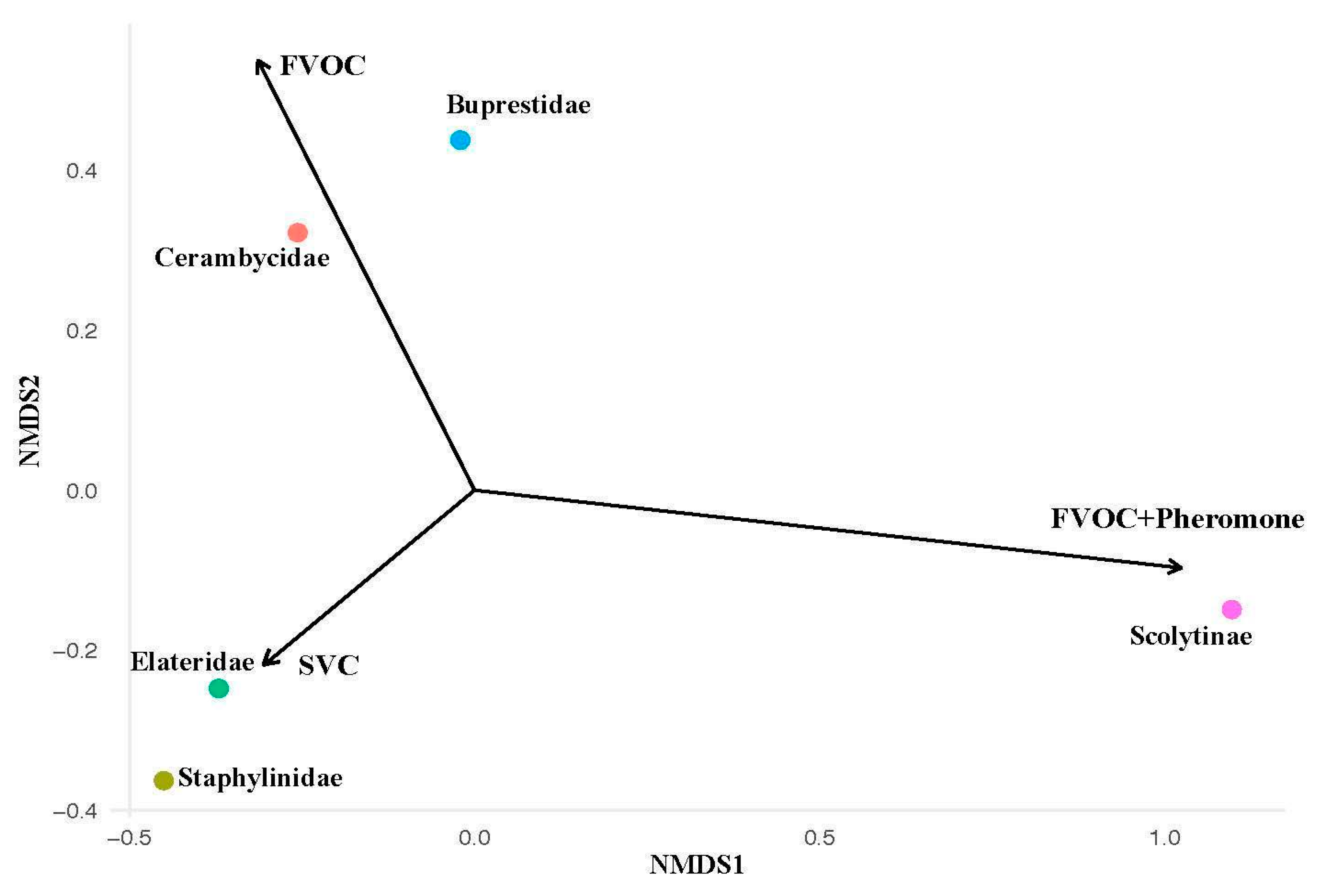
Figure 4.
Effects of fungal volatile (FVOC) and host stress volatile (SVC) experiments on the mean beetle catch of four subcortical beetle families and one subfamily. Experiments include (1) FVOCs alone (FVOC), (2) FVOCs with MPB pheromone lures (FVOCP), and (3) SVCs alone (SVC). Error bars represent standard error. Experiments with the same letter within each beetle group are not significantly different; Dunn’s test with Bonferroni’s adjustment at P<0.05).
Figure 4.
Effects of fungal volatile (FVOC) and host stress volatile (SVC) experiments on the mean beetle catch of four subcortical beetle families and one subfamily. Experiments include (1) FVOCs alone (FVOC), (2) FVOCs with MPB pheromone lures (FVOCP), and (3) SVCs alone (SVC). Error bars represent standard error. Experiments with the same letter within each beetle group are not significantly different; Dunn’s test with Bonferroni’s adjustment at P<0.05).
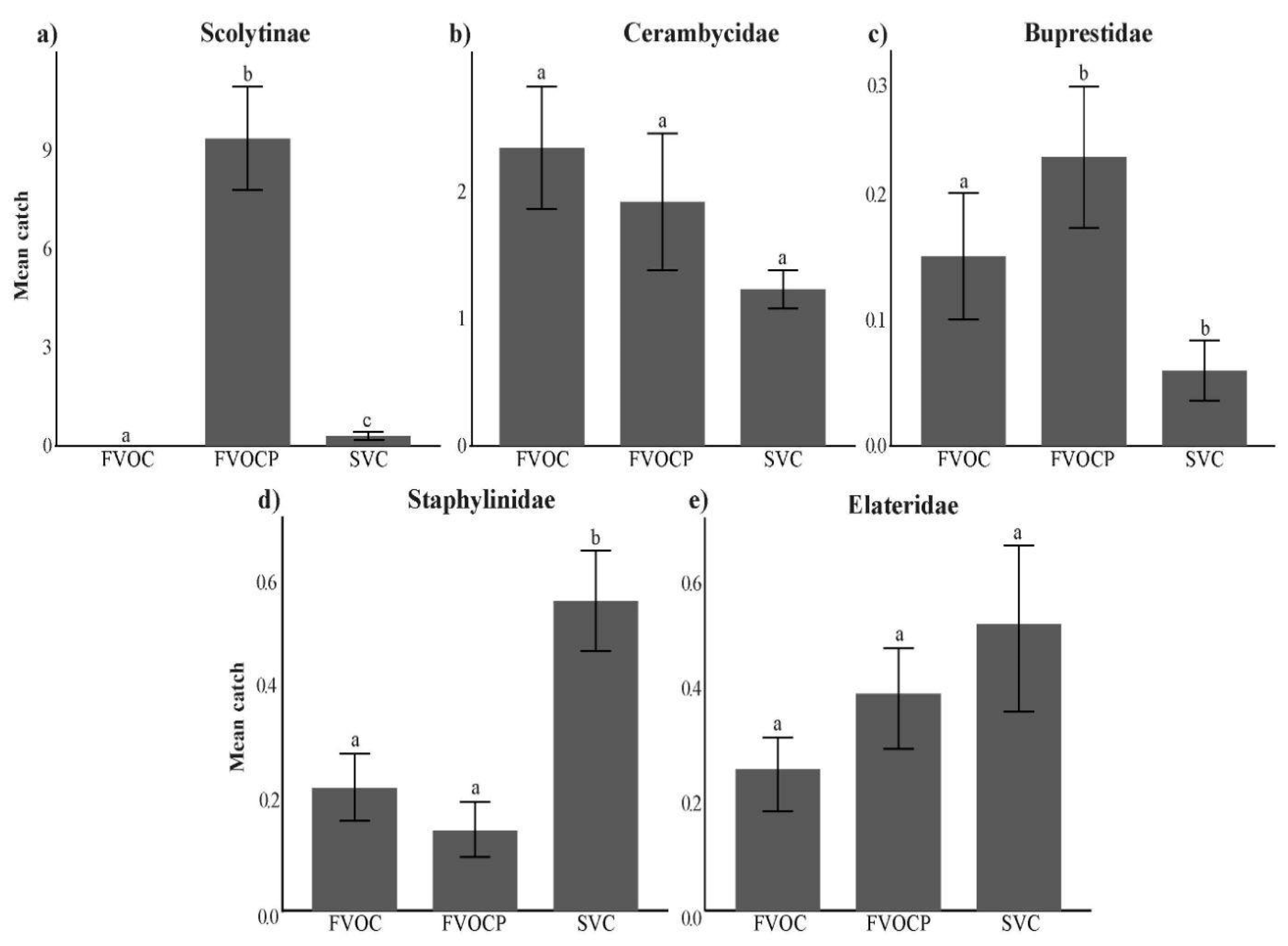

Table 1.
Summary of known attractant compounds (including pheromone components and host volatiles) for coleopteran species in the subcortical beetle families identified in our traps. Synthesized using Gitau et al. (2013) [30] and Tóth (2013) [31].
| Subcortical beetle community | Known attractants | Source | Species attracted | References |
|---|---|---|---|---|
| Curculionidae, Scolytinae | Trans-verbenol | Female pheromone | Dendroctonus ponderosae | [32,33] |
| Exo-brevicomin | Male pheromone | Dendroctonus ponderosae; Dendroctonus terebrans; Dendroctonus brevicomis | [34] | |
| (-)-Endo-brevicomin | Male pheromone | Dendroctonus frontalis | [35,36] | |
| (+)-Sulcatol | Female pheromone |
Gnathotrichus sulcatus; Ips sexdentatus |
[37] | |
| 4,6,6- Lineatin | Female pheromone | Trypodendron lineatum | [37] | |
| Frontalin | Female pheromone | Dendroctonus rufipennis; Dendroctonus brevicomis; Dendroctonus pseudotsugae; Dendroctonus terebrans | [35,38,39] | |
| Ipsdienol | Male pheromone | Ip scalligraphus; Ips pini; Ips duplicatus; Ips avulses; Ips paraconfusus; Ips grandicollis; Ips perturbatus; Ips grandicollis | [26,40,41] | |
| Ipsenol | Male pheromone | Ips grandicollis; Ips paraconfusus; Ips duplicatus | [40,42] | |
| Terpinolene | Host tree volatile | Dendroctonus ponderosae | [17,33,43] | |
| Myrcene | Host tree volatile | Dendroctonus ponderosae | [17,33,44] | |
| Alpha-pinene | Phloem tissue of host trees |
Dendroctonus frontalis; Dendroctonus brevicomis; Ips grandicollis; Hylastes porculus; Hylobius pales; Pachylobius picivorus |
[18,32,44,45] | |
| p-Cymene | Host tree volatile | Ips typographus | [46] | |
| (+)-3-carene | Host tree volatile | Trypodendron domesticum; Hylurgops palliatus | [17,18] | |
| Cerambycidae | 2-Methyl-1-butanol | Pheromone components of several species |
Neoclytus acuminatus; Neoclytus mucronatus; Phymatodes lengi; Xylotrechus colonus; Aegomorphus modestus; Astyleiopus variegatus; Lepturges angulatus; Monochamus carolinensis; Megacyllene caryae; Pyrrhidium sanguineum; Phymatodes alni ssp. alni; Phymatodes testaceus |
[47,48,49] |
| Ethanol | Similar compound to the male pheromone 2-(undecyloxy)-ethanol | Monochamus; Acanthocinus nodosus; Acanthocinus obsoletus; Arhopalus r. nubilus; Xylotrechus s. sagittatus | [45,50,51] | |
| Monochamol | Male-produced aggregation pheromone (from Monochamus galloprovincialis; Monochamus alternatus; Monochamus scutellatus) | Monochamus | [52] | |
| 3-Hydroxy-2-hexanone | Male-produced aggregation pheromone (from Pyrrhidium sanguineum; Phymatodes alni spp. alni; Phymatodes testaceus) | Several species from the subfamily Cerambycinae | [49,53] |
|
| Ipsenol | Bark beetle pheromone (Ips species) | Monochamus carolinensis; Monochamus clamator, Monochamus mutator; Monochamus obtusus; Monochamus scutellatus; Monochamus titillator | [52,54] |
|
| 3-Hydroxy-2-octanone | Male-produced aggregation pheromone | Plagionotus arcuatus | [49] | |
| Fuscumol acetate | Male-produced aggregation pheromone (from | Tetropium | [55,56] | |
| C6 (anti) diols (2S,3R;2R,3S) hexanediol | Male-produced aggregation pheromone (from Neoclytus acuminatus acuminatus; Curius dentatus) | Neoclytus acuminatus acuminatus; Curius dentatus | [57] | |
| α-pinene | Monoterpene produced by many coniferous trees, including Pinus and Picea species | Acanthocinus nodosus, A. obsoletus, Arhopalus rusticus nubilus, Asemum striatum, Monochamus titillator, Prionus pocularis, Xylotrechus integer, and X. sagittatus sagittatus; Xylotrechus longitarsis; Monochamus scutellatus | [45,52,58,59] | |
| Buprestidae | α-pinene | Monoterpene produced by many coniferous trees, including Pinus and Picea species | Buprestis lineata | [45,60] |
| Staphylinidae | Isopropyl (Z9)-hexadecenoate | Female-produced aggregation pheromone | Aleochara curtula | [61,62] |
| Methyl alcohol | Tree derived volatile | Female Philonthus decorus | [63] | |
| Elateridae | Ethanol | Released by trees, especially stressed individuals | Alaus myops | [45] |
| Neryl butanoate | Female-produced aggregation pheromone | Agriotes acuminatus | [64] |
|
| Geranyl butanoate | Female-produced aggregation pheromone | Agriotes brevis; Agriotes lineatus; Agriotes sputator | [65,66] | |
| (E,E)- Farnesyl butanoate | Female-produced aggregation pheromone | Agriotes brevis | [65] |
|
| Geranyl octanoate | Female-produced aggregation pheromone | Agriotes lineatus; Agriotes obscurus | [66] |
|
| Geranyl isovalerate | Female-produced aggregation pheromone | Agriotes litigiosus fen. typicus & Agriotes litigosus var. laichartingi | [66] |
|
| Geranyl hexanoate | Female-produced aggregation pheromone | Agriotes obscurus; Agriotes sordidus | [66] |
|
| (E,E)-Farnesyl acetate | Female-produced aggregation pheromone | Agriotes ustulatus | [66] |
|
| 7-Methyloctyl-5-methylhexanoate | Female-produced aggregation pheromone | Elater ferrugineus | [67] |
|
| 7-Methyloctyl octanoate | Female-produced aggregation pheromone | Elater ferrugineus | [67] |
|
| 7-Methyloctyl-7-methyloctanoate | Female-produced aggregation pheromone | Elater ferrugineus | [67] |
|
| 7-Methyloctyl (Z)-4-decenoate | Female-produced aggregation pheromone | Elater ferrugineus | [67] |
Table 2.
Chemical concentrations and release rates of fungal volatile compounds (FVOCs) used in volatile dispensers. All chemicals were purchased from Sigma-Aldrich.
Table 2.
Chemical concentrations and release rates of fungal volatile compounds (FVOCs) used in volatile dispensers. All chemicals were purchased from Sigma-Aldrich.
| Treatments/ Chemical | Chemical purity (%) | Concentrations (uL mL-1) | Release rate* (mg day-1) |
|---|---|---|---|
| Acetoin | ≥96 | 61.87 | 1.48 |
| 3-Methyl-1-butanol | 98 | 39.87 | 0.98 |
| 2-Methyl-1-butanol | ≥99 | 20.92 | 0.56 |
| Isobutanol | ≥99 | 30.33 | 0.62 |
| 2-Methyl-2-butanol | 99 | 33.27 | 0.84 |
| FVOC mixture | 48.39 | 0.96 | |
| * Release rates determined by weight lost in the laboratory at 22°C | |||
Table 3.
Chemical purity of volatile compounds included in dispensers of host stress volatile blends. The release rate was determined by weight lost in the laboratory at 22°C (22.06 mg/day).
Table 3.
Chemical purity of volatile compounds included in dispensers of host stress volatile blends. The release rate was determined by weight lost in the laboratory at 22°C (22.06 mg/day).
| Compounds | Enantiomeric ratios | Chemical purity (%) | Source |
|---|---|---|---|
| Geranyl acetate | ≥97 | Sigma-Aldrich | |
| α-pinene | (-) | 98 | Sigma-Aldrich |
| Camphene | (-) | 90 | SAFC |
| β-Pinene | (+) | ≥94 | TCI Chemicals |
| 3-Carene | 90 | Sigma-Aldrich | |
| β-Myrcene | 90 | Sigma-Aldrich | |
| Limonene | (S)-(-) | 96 | Fluka Analytical |
| Terpinolene | ≥90 | SAFC | |
| Bornyl acetate | ≥99 | SAFC | |
| γ-Terpinene | 97 | Fluka Analytical | |
| Camphor | ≥95 | Fluka Analytical | |
| Borneol | (-) | 97 | Sigma-Aldrich |
Table 4.
Total catches of subcortical beetle families in field traps over the whole collection period. Host stress volatile (SVC) treatments include: Atropellis piniphila (AP), Grosmannia clavigera (GC), Healthy tree (Healthy), Leptographium longiclavatum (LL), Ophiostoma montium (OM), and Endocronartium harknessii (EH).
Table 4.
Total catches of subcortical beetle families in field traps over the whole collection period. Host stress volatile (SVC) treatments include: Atropellis piniphila (AP), Grosmannia clavigera (GC), Healthy tree (Healthy), Leptographium longiclavatum (LL), Ophiostoma montium (OM), and Endocronartium harknessii (EH).
| Group | Treatments | Cerambycidae | Scolytinae | Buprestidae | Elateridae | Staphylinidae |
|---|---|---|---|---|---|---|
| FVOC | 2-Methyl-1-butanol | 36 | 0 | 1 | 3 | 4 |
| 2-Methyl-2-butanol | 27 | 0 | 3 | 3 | 1 | |
| 3-Methyl-1-butanol | 31 | 0 | 1 | 1 | 2 | |
| Acetoin | 17 | 0 | 3 | 3 | 1 | |
| Isobutanol | 20 | 0 | 4 | 2 | 1 | |
| Control | 19 | 0 | 2 | 3 | 2 | |
| Mixture | 21 | 0 | 4 | 3 | 0 | |
| FVOC with pheromones | 2-Methyl-1-butanol | 15 | 148 | 4 | 3 | 2 |
| 2-Methyl-2-butanol | 8 | 118 | 1 | 7 | 2 | |
| 3-Methyl-1-butanol | 6 | 83 | 2 | 5 | 2 | |
| Acetoin | 45 | 95 | 3 | 2 | 2 | |
| Isobutanol | 15 | 85 | 1 | 1 | 4 | |
| Pheromone alone | 18 | 74 | 1 | 5 | 2 | |
| Mixture | 35 | 91 | 0 | 5 | 3 | |
| SVC | AP | 17 | 5 | 9 | 7 | 1 |
| Control | 14 | 6 | 9 | 6 | 2 | |
| GC | 16 | 4 | 15 | 2 | 0 | |
| Healthy | 16 | 3 | 13 | 15 | 0 | |
| LL | 21 | 5 | 5 | 0 | 2 | |
| OM | 13 | 4 | 6 | 7 | 1 | |
| EH | 26 | 3 | 5 | 13 | 0 | |
| Total | 436 | 724 | 92 | 96 | 34 |
Table 5.
Summary of known attractants for subcortical beetles, key fungal volatiles (FVOCs), and host stress volatile (SVC) profiles identified in each of the experiments in our study. The blend refers to a synthetic blend of FVOCs that were based on the Ophiostoma montium profile identified by Zaman et al. (2023b) [22].
Table 5.
Summary of known attractants for subcortical beetles, key fungal volatiles (FVOCs), and host stress volatile (SVC) profiles identified in each of the experiments in our study. The blend refers to a synthetic blend of FVOCs that were based on the Ophiostoma montium profile identified by Zaman et al. (2023b) [22].
| Subcortical beetle family | Known attractants | Key FVOCs | Key FVOCs when pheromones are added | Key SVCs |
|---|---|---|---|---|
| Curculionidae Sub-family: Scolytinae | Trans-verbenol, Exo-brevicomin, Terpinolene, Myrcene | N/A | 2-Methyl-1-butanol | N/A |
| Cerambycidae | 2-Methyl-1-butanol ⍺-Pinene | 2-Methyl-1-butanol | Acetoin, blend | Endocronartium harknessii |
| Buprestidae | Tree stress volatiles primarily | 2-Methyl-1-butanol | Isobutanol |
Leptographium longiclavatum, Endocronartium harknessii |
| Staphylinidae | Bark beetle pheromones, some monoterpenes | Isobutanol, blend | Acetoin | Grosmannia clavigera, Healthy tree |
| Elateridae | Bark beetle pheromones, some monoterpenes | N/A | 2-Methyl-2-butanol | Healthy tree, Endocronartium harknessii |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



